Embed Size (px)
Citation preview

This article was downloaded by: [Ingenta Content Distribution (Publishing Technology)]On: 07 October 2014, At: 10:03Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office:Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
GranaPublication details, including instructions for authors and subscriptioninformation:http://www.tandfonline.com/loi/sgra20
Pollen morphology and systematics ofBurseraceaeMadeline M Harley a , Unsook Song b & Hannah I Banks aa Palynology Unit , Herbarium, Royal Botanic Gardens , Kew, Richmond,Surrey, TW9 3AE, UKb Unsook Song, Dept Forest Resources, College of Agriculture , ChonbukNational University , 561‐756, South Koreac Palynology Unit , Herbarium, Royal Botanic Gardens , Kew, Richmond,Surrey, TW9 3AE, UKPublished online: 18 Feb 2007.
To cite this article: Madeline M Harley , Unsook Song & Hannah I Banks (2005) Pollen morphology andsystematics of Burseraceae, Grana, 44:4, 282-299, DOI: 10.1080/00173130500477902
To link to this article: http://dx.doi.org/10.1080/00173130500477902
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”)contained in the publications on our platform. However, Taylor & Francis, our agents, and ourlicensors make no representations or warranties whatsoever as to the accuracy, completeness, orsuitability for any purpose of the Content. Any opinions and views expressed in this publicationare the opinions and views of the authors, and are not the views of or endorsed by Taylor &Francis. The accuracy of the Content should not be relied upon and should be independentlyverified with primary sources of information. Taylor and Francis shall not be liable for anylosses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilitieswhatsoever or howsoever caused arising directly or indirectly in connection with, in relation to orarising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Any substantialor systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, ordistribution in any form to anyone is expressly forbidden. Terms & Conditions of access and usecan be found at http://www.tandfonline.com/page/terms-and-conditions

Pollen morphology and systematics of Burseraceae
MADELINE M. HARLEY, UNSOOK SONG and HANNAH I. BANKS
Harley, M. M., Song, U. & Banks, H. I. 2005. Pollen morphology and systematics of Burseraceae. –Grana 44: 282–299. ISSN 0017-3134.
The Burseraceae are a medium-sized family in which 18 genera are currently recognised. They are thesubject of a long-term project to describe the pollen morphology from light, scanning electron andtransmission electron microscopy. The pollen morphology of tribe Protieae has been published, as wellas an account of the pollen of the African taxa in the family. Pollen data for the other two tribes,Bursereae and Canarieae, are more or less complete. The pollen of all the genera have been examined,with the exception of the recently described Pseudodacryodes Pierlot for which, currently, there is nopollen material available. This paper summarises the results.
There is considerable variation in exine and aperture features between, and occasionally within, thegenera and 14 major pollen types are defined, including two previously undescribed types: ‘Canarium
oleiferum’ and ‘Canarium gracile’. The distribution of pollen characteristics throughout the family iscompared with previously published tribal and subtribal groupings, as well as with current ideas ofgeneric relationships from molecular analyses. Comparisons show notable congruence of pollen datawith molecular data. To some extent pollen morphology is different for each of the subtribes.Nevertheless, there are some notable exceptions, for example, the pollen of Garuga and Boswellia areremarkably similar, although Garuga has been included, somewhat tenuously, in tribe Protieae, andBoswellia is included in tribe Bursereae, subtribe Boswelliinae. In a recent molecular tree Garuga andBoswellia appear to be closely related, and this supports the conclusion, based on severalmacromorphological characters as well as pollen, that Garuga should be transferred to tribe Bursereae.
Madeline M. Harley* & Hannah I. Banks, Palynology Unit, Herbarium, Royal Botanic Gardens, Kew,
Richmond, Surrey, TW9 3AE, UK; Unsook Song, Dept Forest Resources, College of Agriculture,
Chonbuk National University, 561-756, South Korea.
*Corresp. Author
(Manuscript received 19 April 2005; accepted 28 October 2005)
The Burseraceae are a medium-sized pantropical family of
18 genera comprising shrubs and trees with prominent resin
ducts in the bark, as well as in other parts of the plant.
Economically, among the best-known members of Burser-
aceae are: Gaboon Mahogany (Aucoumea klaineana Pierre);
Indian White Mahogany (Canarium euphyllum Kurz),
Myrrh (Commiphora myrrha (Nees) Engl.) and Frankincense
(Boswellia species, notably B. sacra Flueck.). The resin from
Bursera, especially in Mexico, is extracted to make varnish.
The species are widely distributed in the Old and New
Worlds, but centred especially in Malesia, Africa, Meso- and
South America. This distribution is reflected in the widely
used tribal classification of Lam (1932) where genera of tribe
Protieae are predominantly South American, those of tribe
Bursereae predominantly African or Mesoamerican, while
those of tribe Canarieae are predominantly Malesian.
Systematically the family has been placed in various
orders by different authors (see Brummitt 1992): within
the Geraniales sensu Bentham & Hooker (1862), Dalla Torre
& Harms (1901); Rutales sensu Melchior (1964), Thorne
(1983); Dahlgren (1983), Young (1982), Takhtajan (1987);
Sapindales sensu Cronquist (1981). The Angiosperm Phylo-
geny Group (1998, 2003) also includes Burseraceae in their
circumscription of Sapindales. Of the 38 families that have,
at times, been placed in the same order as Burseraceae in
different systematic treatments, only three families have
consistently been placed with Burseraceae – Meliaceae,
Rutaceae and Simaroubaceae. Interestingly, in all recent
systematic treatments, a sister relationship between Burser-
aceae and Anacardiaceae has been recognised. This has been
further confirmed by molecular studies including Soltis et al.
(2000) and Savolainen et al. (2000).
Prior to the studies initiated (MMH) for the World Pollen
and Spore Flora, the pollen morphology of Burseraceae had
not been addressed in relation to the systematics of the
family. A long-term project to describe the pollen morphol-
ogy of each of the three widely accepted tribes, from light,
scanning electron and transmission electron microscopy, for
The World Pollen and Spore Flora (WPSF) is nearing
completion. The pollen morphology of tribe Protieae has
already been published (Harley & Daly 1995), and an
account of the pollen of the African taxa in the family
(Harley & Hall 1999). Pollen data for the other two tribes,
Bursereae and Canarieae, are more or less complete. The
pollen of all the genera have been examined, with the
exception of the recently described Pseudodacryodes Pierlot
(1997) for which, currently, there is no pollen material
available. In Harley & Daly (1995) there is a summary of
previously published accounts of pollen morphology for
some of the taxa included in the family. More recently
comprehensive pollen morphological accounts for the
species of each of the three tribes which occur in South
America have been published by Aguilar-Sierra & Melhem
(1998a, b, c).
Grana 44: 282–299, 2005
Grana 44 (2005) # 2005 Taylor & Francis. ISSN 0017-3134
DOI: 10.1080/00173130500477902
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
10:
03 0
7 O
ctob
er 2
014

The pollen morphology of Anacardiaceae, Meliaceae,
Rutaceae and Simaroubaceae is more or less distinct from
that of Burseraceae. Pollen morphology of Meliaceae, and
references to, is summarised by Van der Ham (1995). Pollen
morphology has been described for about 60% of Rutaceae
genera, representative of all seven subfamilies (Engler 1931),
although the coverage is more comprehensive for some
subfamilies than for others; for example, within subfamily
Rutoideae pollen studies appear to be completely lacking in
tribe Boronieae, while in tribe Xanthoxyleae pollen studies
have been carried out for only four genera. Pollen accounts for
Rutaceae include: Arreguın-Sanchez et al. (1995), Barth 1980,
1982, 1983, 1985; Correa et al. (1992), Erdtman (1952), Grant
et al. (2000), Harley (1986), Lobreau-Callen et al. (1975),
Morton & Kallunki (1993), Mziray (1992), Rasolodimby
(1983), Victor & Van Wyk (1998, 1999a, b, 2001). The pollen of
the small, predominantly pantropical, family Simaroubaceae
has not been extensively studied. Probably the most compre-
hensive account remains that of Basak (1963) for Indian
members of the family. The pollen of Anacardiaceae is also
generally distinguishable from that of Burseraceae (Erdtman
1952, Baksi 1976, Ibe & Leis 1979). Notably, a distinctly striate
exine – which is not particularly common among colporate
edicots – is fairly common in the Sapindales, including:
Anacardiaceae, Rutaceae, Sapindaceae, Simaroubaceae, as
well as in Burseraceae.
MATERIAL AND METHODS
The pollen data presented are based on extensive sampling of all
currently recognised genera of Burseraceae with the exception of the
recently described Pseudodacryodes Pierlot (1987) for which no
pollen material is currently available (Pierlot: pers. comm.). Pollen
data for tribe Protieae (Crepidospermum, Garuga, Protium, Tetra-
gastris) are already published (Harley & Daly 1995), while accounts
of the pollen morphology of tribes Bursereae (Aucoumea, Beiselia,
Boswellia, Bursera, Commiphora, Triomma) and Canarieae (Canar-
ium, Dacryodes, Haplolobus, Pseudodacryodes, Rosselia, Santiria,
Scutinanthe, Trattinnickia) are in progress.
Pollen preparation methods for light microscopy (LM), scanning
electron (SEM) and transmission electron microscopy (TEM) follow
the protocols described in Harley & Daly (1995), including the
methods for calculating and defining endoaperture shape, endoa-
perture width and colpus length relative to overall pollen size.
Endoaperture shape index (ENSI) for lalongate endoapertures
(Table I) – the result of dividing the average polar length of the
endoaperture by the average equatorial width (E) of the pollen
grain. The endoaperture shape is interpreted: 3.6 or more, per-
lalongate; 2.6 – 3.5 average lalongate; 2.5 or less, brevi-lalongate.
Endoaperture width index (ENWI) for lalongate endoapertures
(Table II) – the average equatorial width of the endoaperture in
proportion to the average equatorial width of the pollen grain. The
average equatorial width of the endoaperture 63, is divided by two,
and the result divided into the average equatorial width of the
pollen grain. The endoaperture width index is interpreted: 1.45 or
less, very wide; 1.46 – 2.0, wide; 2.01 – 2.50, average; 2.51 – 3.00,
short; 3.01w, very short [NB. The reasoning for the first part of the
calculation is based on the equatorial width of the pollen grain, in
LM, being a planar dimension, whereas the width occupied by the
endoapertures, is effectively 3-dimensional. Dividing the sum total
of the three endoapertures by two provides a measurement which is
more compatible with the equatorial width of the pollen grain]. The
colpus length index (CLI) see Table III, is the result of dividing the
average colpus length into the average polar length of the pollen
grain (P). The colpus length is interpreted: v1.05 – 1.35, long;
1.36 – 1.69, average; 1.70 – 2.00w, short.
Terminology follows Segaar & Van der Ham (1993), Punt et al.
(1994 & the updated website version at: http://www.bio.uu.nl/palaeo/
glossary/ ), Harley & Daly (1995).
RESULTS
As well as our unpublished pollen data for tribes Bursereae
and Canarieae (MH, US, HB) we have drawn on the
following published sources for our morphological data:
Segaar & Van der Ham (1993), Forman et al. (1989, 1994),
Harley & Daly (1995), Harley & Hall (1999). The pollen
types described here focus on the major differences in pollen
morphology within the family. Some of these major types
include the more finely delimited types from the pollen
account of tribe Protieae (Harley & Daly 1995), where
species and collection data for all species studied for tribe
Protieae are given. Full lists of species and collections
studied will also be provided for tribes Bursereae and
Canarieae when the tribal pollen morphological accounts for
Bursereae (M. Harley: in prep.) and Canarieae (M. Harley:
in prep.) are published.
General pollen morphology (Tables IV & V)
Pollen grains usually isopolar, rarely heteropolar. Aperture
system tricolporate, rarely tricolporate lobed-vestibulate or
triporate projectate; aperture disposition usually fossapertu-
rate, angulaperturate less frequently planaperturate. Pollen
Table I. Endoaperture shape index (ENSI).
(Harley & Daly 1995).
Endoaperture shape Index
PER-LALONGATE 3.6 or moreAVERAGE LALONGATE 3.5 – 2.6BREVI-LALONGATE 2.5 or less
Table II. Endoaperture width index (ENWI).
(Harley & Daly 1995).
Endoaperture Width index
VERY WIDE 1.45 or lessWIDE 1.46 – 2.00AVERAGE 2.01 – 2.50SHORT 2.51 – 3.00VERY SHORT 3.01 or more
Table III. Colpus length index (CLI).
(Harley & Daly 1995).
Colpus Length index
LONG 1.35 – 1.05 or lessAVERAGE 1.36 – 1.69SHORT 1.70 – 2.00 or more
Pollen of Burseraceae 283
Grana 44 (2005)
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
10:
03 0
7 O
ctob
er 2
014
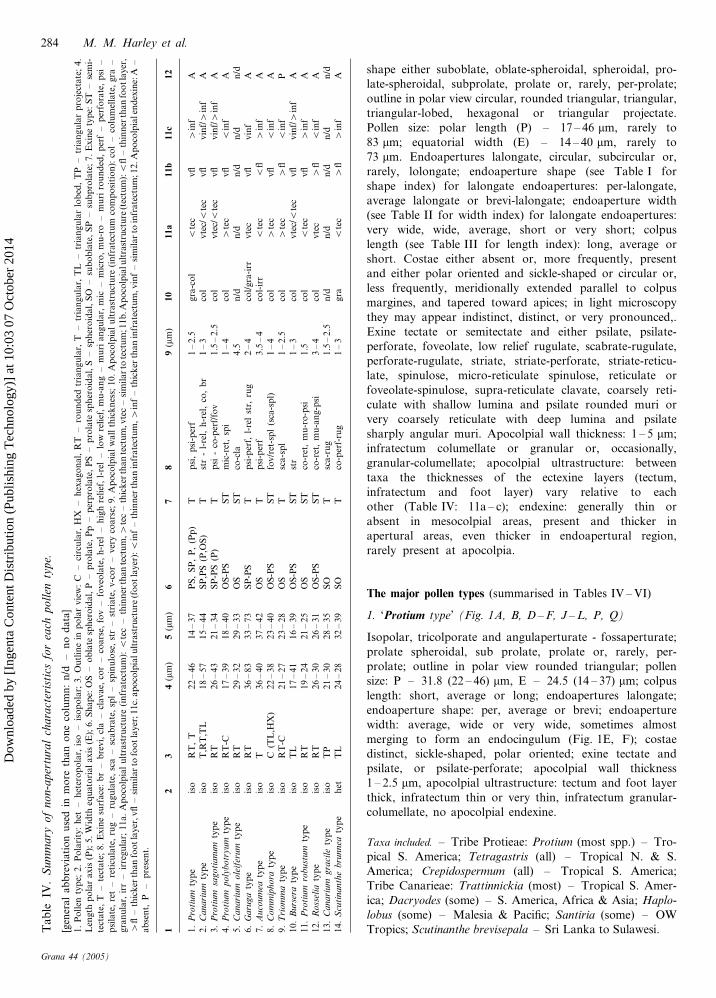
shape either suboblate, oblate-spheroidal, spheroidal, pro-
late-spheroidal, subprolate, prolate or, rarely, per-prolate;
outline in polar view circular, rounded triangular, triangular,
triangular-lobed, hexagonal or triangular projectate.
Pollen size: polar length (P) – 17 – 46 mm, rarely to
83 mm; equatorial width (E) – 14 – 40 mm, rarely to
73 mm. Endoapertures lalongate, circular, subcircular or,
rarely, lolongate; endoaperture shape (see Table I for
shape index) for lalongate endoapertures: per-lalongate,
average lalongate or brevi-lalongate; endoaperture width
(see Table II for width index) for lalongate endoapertures:
very wide, wide, average, short or very short; colpus
length (see Table III for length index): long, average or
short. Costae either absent or, more frequently, present
and either polar oriented and sickle-shaped or circular or,
less frequently, meridionally extended parallel to colpus
margines, and tapered toward apices; in light microscopy
they may appear indistinct, distinct, or very pronounced,.
Exine tectate or semitectate and either psilate, psilate-
perforate, foveolate, low relief rugulate, scabrate-rugulate,
perforate-rugulate, striate, striate-perforate, striate-reticu-
late, spinulose, micro-reticulate spinulose, reticulate or
foveolate-spinulose, supra-reticulate clavate, coarsely reti-
culate with shallow lumina and psilate rounded muri or
very coarsely reticulate with deep lumina and psilate
sharply angular muri. Apocolpial wall thickness: 1 – 5 mm;
infratectum columellate or granular or, occasionally,
granular-columellate; apocolpial ultrastructure: between
taxa the thicknesses of the ectexine layers (tectum,
infratectum and foot layer) vary relative to each
other (Table IV: 11a – c); endexine: generally thin or
absent in mesocolpial areas, present and thicker in
apertural areas, even thicker in endoapertural region,
rarely present at apocolpia.
The major pollen types (summarised in Tables IV – VI)
1. ‘Protium type’ (Fig. 1A, B, D – F, J – L, P, Q)
Isopolar, tricolporate and angulaperturate - fossaperturate;
prolate spheroidal, sub prolate, prolate or, rarely, per-
prolate; outline in polar view rounded triangular; pollen
size: P – 31.8 (22 – 46) mm, E – 24.5 (14 – 37) mm; colpus
length: short, average or long; endoapertures lalongate;
endoaperture shape: per, average or brevi; endoaperture
width: average, wide or very wide, sometimes almost
merging to form an endocingulum (Fig. 1E, F); costae
distinct, sickle-shaped, polar oriented; exine tectate and
psilate, or psilate-perforate; apocolpial wall thickness
1 – 2.5 mm, apocolpial ultrastructure: tectum and foot layer
thick, infratectum thin or very thin, infratectum granular-
columellate, no apocolpial endexine.
Taxa included. – Tribe Protieae: Protium (most spp.) – Tro-
pical S. America; Tetragastris (all) – Tropical N. & S.
America; Crepidospermum (all) – Tropical S. America;
Tribe Canarieae: Trattinnickia (most) – Tropical S. Amer-
ica; Dacryodes (some) – S. America, Africa & Asia; Haplo-
lobus (some) – Malesia & Pacific; Santiria (some) – OW
Tropics; Scutinanthe brevisepala – Sri Lanka to Sulawesi.Ta
ble
IV.
Su
mm
ary
of
no
n-a
per
tura
lch
ara
cter
isti
csfo
rea
chp
oll
enty
pe.
[gen
era
la
bb
rev
iati
on
use
din
mo
reth
an
on
eco
lum
n:
n/d
–n
od
ata
]1.
Po
llen
typ
e;2.
Po
lari
ty:
het
–h
eter
op
ola
r,is
o–
iso
po
lar;
3.
Ou
tlin
ein
po
lar
vie
w:
C–
circ
ula
r,H
X–
hex
ago
nal,
RT
–ro
un
ded
tria
ngu
lar,
T–
tria
ng
ula
r,T
L–
tria
ngu
lar
lob
ed,
TP
–tr
ian
gu
lar
pro
ject
ate
;4.
Len
gth
po
lar
axis
(P);
5.
Wid
theq
uato
rial
axis
(E);
6.
Sh
ap
e:O
S–
ob
late
sph
ero
idal,
P–
pro
late
,P
p–
per
pro
late
,P
S–
pro
late
sph
ero
idal,
S–
sph
ero
idal,
SO
–su
bo
bla
te,
SP
–su
bp
rola
te;
7.
Exin
ety
pe:
ST
–se
mi-
tect
ate
,T
–te
ctate
;8.
Exin
esu
rface
:b
r–
bre
vi,
cla
–cl
avae,
cor
–co
ars
e,fo
v–
foveo
late
,h
-rel
–h
igh
reli
ef,
l-re
l–
low
reli
ef,
mu
-an
g–
mu
rian
gu
lar,
mic
–m
icro
,m
u-r
o–
mu
riro
un
ded
,p
erf
–p
erfo
rate
,p
si–
psi
late
,re
t–
reti
cula
te,
rug
–ru
gu
late
,sc
a–
scab
rate
,sp
l–
spin
ulo
se,
str
–st
riate
,v-c
or
–ver
yco
ars
e;9.
Ap
oco
lpia
lw
all
thic
kn
ess;
10.
Ap
oco
lpia
lu
ltra
stru
ctu
re(i
nfr
ate
ctu
mco
mp
osi
tio
n):
col
–co
lum
ella
te,
gra
–
gra
nu
lar,
irr
–ir
regu
lar;
11a.
Ap
oco
lpia
lu
ltra
stru
ctu
re(i
nfr
ate
ctu
m):
vte
c–
thin
ner
than
tect
um
,wte
c–
thic
ker
than
tect
um
,vte
c–
sim
ilar
tote
ctu
m;1
1b
.Ap
oco
lpia
lult
rast
ruct
ure
(tec
tum
):v
fl–
thin
ner
than
foo
tla
yer
,
wfl
–th
ick
erth
an
foo
tla
yer
,vfl
–si
mil
ar
tofo
ot
layer
;11c.
ap
oco
lpia
lult
rast
ruct
ure
(fo
ot
layer
):v
inf
–th
inn
erth
an
infr
ate
ctu
m,w
inf
–th
ick
erth
an
infr
ate
ctu
m,v
inf
–si
mil
ar
toin
fra
tect
um
;12.A
po
colp
iale
nd
exin
e:A
–
ab
sen
t,P
–p
rese
nt.
12
34
(mm
)5
(mm
)6
78
9(m
m)
10
11
a11
b1
1c
12
1.
Pro
tium
typ
eis
oR
T,
T22
–46
14
–37
PS
,S
P,
P,
(Pp
)T
psi
,p
si-p
erf
1–
2.5
gra
-co
lv
tec
vfl
win
fA
2.
Canari
um
typ
eis
oT
,RT
,TL
18
–57
15
–44
SP
,PS
(P,O
S)
Tst
r-
l-re
l,h
-rel
,co
,b
r1
–3
col
vte
c/v
tec
vfl
vin
f/w
inf
A
3.
Pro
tium
sagoti
anum
typ
eis
oR
T26
–43
21
–34
SP
-PS
(P)
Tp
si-
co-p
erf/
fov
1.5
–2
.5co
lv
tec/
vte
cvfl
vin
f/w
inf
A
4.
Pro
tium
poly
botr
yum
typ
eis
oR
T-C
17
–39
18
–40
OS
-PS
ST
mic
-ret
,sp
i1
–4
col
wte
cvfl
vin
fA
5.
Canari
um
ole
ifer
um
typ
eis
oR
T29
–32
29
–33
OS
ST
co-c
la4.5
n/d
n/d
n/d
n/d
n/d
6.
Garu
ga
typ
eis
oR
T36
–83
33
–73
SP
-PS
Tp
si-p
erf,
l-re
lst
r,ru
g2
–4
col/
gra
-irr
vte
cvfl
vin
fA
7.
Auco
um
eaty
pe
iso
T36
–40
37
–42
OS
Tp
si-p
erf
3.5
–4
col-
irr
vte
cv
flw
inf
A
8.
Co
mm
iph
ora
typ
eis
oC
(TL
,HX
)22
–38
23
–40
OS
-PS
ST
fov/r
et-s
pl
(sca
-sp
l)1
–4
col
wte
cvfl
vin
fA
9.
Tri
om
ma
typ
eis
oR
T-C
21
–27
23
–28
OS
Tsc
a-s
pl
1–
2.5
col
wte
cw
flv
inf
P
10.
Burs
era
typ
eis
oT
L17
–41
16
–39
OS
-PS
ST
str
1–
3co
lv
tec/
vte
cvfl
vin
f/w
inf
A
11.
Pro
tium
robust
um
typ
eis
oR
T19
–24
21
–25
OS
ST
co-r
et,
mu
-ro
-psi
1.5
col
vte
cvfl
win
fA
12.
Ross
elia
typ
eis
oR
T26
–30
26
–31
OS
-PS
ST
co-r
et,
mu
-an
g-p
si3
–4
col
vte
cw
flv
inf
A
13.
Canari
um
gra
cile
typ
eis
oT
P21
–30
28
–35
SO
Tsc
a-r
ug
1.5
–2
.5n
/dn
/dn
/dn
/dn
/d
14.
Scu
tinanth
ebru
nnea
typ
eh
etT
L24
–28
32
–39
SO
Tco
-per
f-ru
g1
–3
gra
vte
cw
flw
inf
A
284 M. M. Harley et al.
Grana 44 (2005)
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
10:
03 0
7 O
ctob
er 2
014
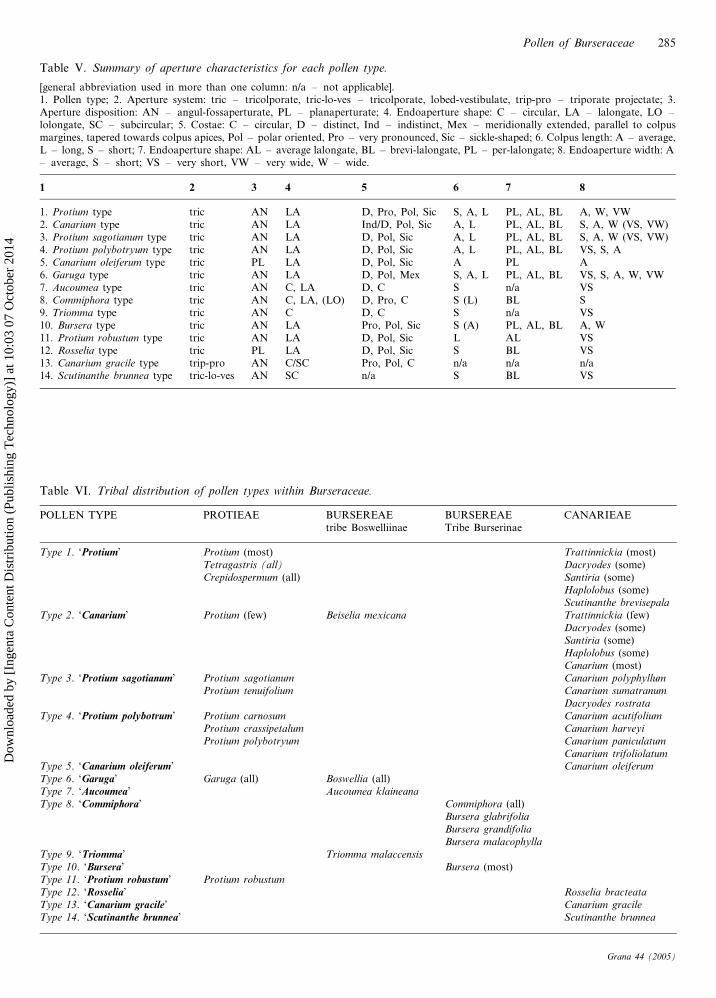
Table V. Summary of aperture characteristics for each pollen type.
[general abbreviation used in more than one column: n/a – not applicable].1. Pollen type; 2. Aperture system: tric – tricolporate, tric-lo-ves – tricolporate, lobed-vestibulate, trip-pro – triporate projectate; 3.Aperture disposition: AN – angul-fossaperturate, PL – planaperturate; 4. Endoaperture shape: C – circular, LA – lalongate, LO –lolongate, SC – subcircular; 5. Costae: C – circular, D – distinct, Ind – indistinct, Mex – meridionally extended, parallel to colpusmargines, tapered towards colpus apices, Pol – polar oriented, Pro – very pronounced, Sic – sickle-shaped; 6. Colpus length: A – average,L – long, S – short; 7. Endoaperture shape: AL – average lalongate, BL – brevi-lalongate, PL – per-lalongate; 8. Endoaperture width: A– average, S – short; VS – very short, VW – very wide, W – wide.
1 2 3 4 5 6 7 8
1. Protium type tric AN LA D, Pro, Pol, Sic S, A, L PL, AL, BL A, W, VW2. Canarium type tric AN LA Ind/D, Pol, Sic A, L PL, AL, BL S, A, W (VS, VW)3. Protium sagotianum type tric AN LA D, Pol, Sic A, L PL, AL, BL S, A, W (VS, VW)4. Protium polybotryum type tric AN LA D, Pol, Sic A, L PL, AL, BL VS, S, A5. Canarium oleiferum type tric PL LA D, Pol, Sic A PL A6. Garuga type tric AN LA D, Pol, Mex S, A, L PL, AL, BL VS, S, A, W, VW7. Aucoumea type tric AN C, LA D, C S n/a VS8. Commiphora type tric AN C, LA, (LO) D, Pro, C S (L) BL S9. Triomma type tric AN C D, C S n/a VS10. Bursera type tric AN LA Pro, Pol, Sic S (A) PL, AL, BL A, W11. Protium robustum type tric AN LA D, Pol, Sic L AL VS12. Rosselia type tric PL LA D, Pol, Sic S BL VS13. Canarium gracile type trip-pro AN C/SC Pro, Pol, C n/a n/a n/a14. Scutinanthe brunnea type tric-lo-ves AN SC n/a S BL VS
Table VI. Tribal distribution of pollen types within Burseraceae.
POLLEN TYPE PROTIEAE BURSEREAE BURSEREAE CANARIEAEtribe Boswelliinae Tribe Burserinae
Type 1. ‘Protium’ Protium (most) Trattinnickia (most)Tetragastris (all) Dacryodes (some)Crepidospermum (all) Santiria (some)
Haplolobus (some)Scutinanthe brevisepala
Type 2. ‘Canarium’ Protium (few) Beiselia mexicana Trattinnickia (few)Dacryodes (some)Santiria (some)Haplolobus (some)Canarium (most)
Type 3. ‘Protium sagotianum’ Protium sagotianum Canarium polyphyllum
Protium tenuifolium Canarium sumatranum
Dacryodes rostrata
Type 4. ‘Protium polybotrum’ Protium carnosum Canarium acutifolium
Protium crassipetalum Canarium harveyi
Protium polybotryum Canarium paniculatum
Canarium trifoliolatumType 5. ‘Canarium oleiferum’ Canarium oleiferum
Type 6. ‘Garuga’ Garuga (all) Boswellia (all)Type 7. ‘Aucoumea’ Aucoumea klaineana
Type 8. ‘Commiphora’ Commiphora (all)Bursera glabrifolia
Bursera grandifolia
Bursera malacophylla
Type 9. ‘Triomma’ Triomma malaccensis
Type 10. ‘Bursera’ Bursera (most)Type 11. ‘Protium robustum’ Protium robustum
Type 12. ‘Rosselia’ Rosselia bracteata
Type 13. ‘Canarium gracile’ Canarium gracile
Type 14. ‘Scutinanthe brunnea’ Scutinanthe brunnea
Pollen of Burseraceae 285
Grana 44 (2005)
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
10:
03 0
7 O
ctob
er 2
014

286 M. M. Harley et al.
Grana 44 (2005)
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
10:
03 0
7 O
ctob
er 2
014

WPSF pollen types included (Harley & Daly 1995). – Protium
cubense-type; Protium macgregorii-type; Protium strumo-
sum-type; Crepidospermum goudotianum-type; Tetragastris
mucronata-type.
2. ‘Canarium type’ (Fig. 1C, G – I, M – O, R)
Isopolar, tricolporate and angulaperturate; usually subpro-
late, sometimes prolate spheroidal, rarely, prolate or oblate
spheroidal; outline in polar view triangular, rounded
triangular or, sometimes, triangular lobate; pollen size: P
– 19.6 – 50.0 (18 – 57) mm, E. – 17.0 – 41.4 (15 – 44) mm;
colpus length: long or average; endoapertures lalongate;
endoaperture shape: brevi, average or per; endoaperture
width: wide, average, short, rarely, very short or very wide;
costae indistinct to distinct , sickle-shaped, polar oriented;
exine tectate with fine low or high relief striate to coarsely
striate, aligned or non aligned, occasionally brevi-striate,
striate-perforate or, rarely, striae transversely ridged;
apocolpial wall thickness 1 – 3 mm, apocolpial ultrastructure:
foot layer, tectum, infratectum columellate and similar in
thickness, or infratectum may be thinner, no apocolpial
endexine.
Taxa included. – Tribe Protieae: Protium (a few mainly OW
Tropics spp.); Tribe Bursereae: Beiselia mexicana – Mexico;
Tribe Canarieae: Dacryodes (some) – Tropical S. America;
Africa; Asia; Santiria (some) – OW Tropics; Haplolobus
(some) – Malesia and Pacific; Canarium (most) – OW Tro-
pics.
WPSF types included (Harley & Daly 1995). – Protium fimbria-
tum-type; Protium apiculatum-type; Protium connarifolium-
type; Protium serratum-type; Protium urophyllidium-type;
Protium javanicum-type; Protium madagascariense-type.
3. ‘Protium sagotianum type’ (Fig. 2A, D, G, J – L)
Isopolar, tricolporate and angulaperturate; prolate spheroidal,
subprolate or, less commonly, prolate; outline in polar view
rounded triangular; pollen size: P – 30.8 (26 – 43) mm, E – 25.8
(21 – 34) mm; colpus length: average or long; endoapertures
lalongate; endoaperture shape: per, average or brevi; endoa-
perture width: short, average or wide; costae distinct, sickle-
shaped, polar oriented; exine tectate and psilate with coarse
perforations or foveolae frequently denser and/or coarser in
apolcolpial area; apocolpial wall thickness 1.75 – 2.5 mm,
apocolpial ultrastructure: tectum, foot layer and columellate
infratectum more or less equal in thickness or, infratectum may
be thinner, no apocolpial endexine.
Taxa included. – Tribe Protieae: Protium sagotianum, P.
tenuifolium – Peru, Trinidad, Costa Rica; Tribe Canar-
ieae: Canarium polyphyllum, C. sumatranum – Sumatra,
Malay Peninsula; Dacryodes rostrata – Malesia.
WPSF type included (Harley & Daly 1995). – Protium sagotia-
num-type.
4. ‘Protium polybotryum type’ (Fig. 2B, E, H, I, M – O)
Isopolar, tricolporate and angulaperturate; oblate spheroi-
dal to prolate spheroidal; outline in polar view rounded
triangular to circular; pollen size: P – 26 (17 – 39) mm, E –
25.4 (18 – 40) mm; colpus length: average or long; endoa-
pertures: lalongate, endoaperture shape: brevi, average or
long; endoaperture width: average, short or very short;
costae distinct, sickle-shaped, polar oriented; exine semi-
tectate and micro-reticulate, spinulose; apocolpial wall
thickness: 1 – 2 mm or 3 – 4 mm; apocolpial ultrastructure:
tectum and foot layer similar in thickness, infratectum
columellate and wider, no apocolpial endexine.
Taxa included. – Tribe Protieae. – Protium carnosum,
P. crassipetalum, P. polybotryum – Brazil, Surinam;
Tribe Canarieae: Canarium acutifolium, C. harveyi, C.
paniculatum, C. trifoliolatum – Mascarenes, Mauritius,
Fiji, New Guinea, New Caledonia.
WPSF type included (Harley & Daly 1995). – Protium polybo-
tryum-type.
5. ‘Canarium oleiferum type’ (Fig. 2C, F, P – R)
Isopolar, tricolporate and planaperturate; oblate spheroidal;
outline in polar view rounded triangular; pollen size: P – 30.7
(29 – 32) mm, E – 30.9 (29 – 33) mm; colpus length: average;
endoapertures lalongate; endoaperture shape: per; endoaper-
ture width: average; costae distinct, sickle-shaped, polar-
oriented; exine semitectate, coarsely and densely clavate;
apocolpial wall thickness c. 4.5 mm, apocolpial ultrastructure
unknown (embedded but not yet thin sectioned), however,
from light microscopy it appears that the clavae project from a
thin, finely reticulate tectum beneath which is a columellate
infratectum and a thin foot layer.
Taxon included. – Tribe Canarieae: Canarium oleiferum –
New Caledonia.
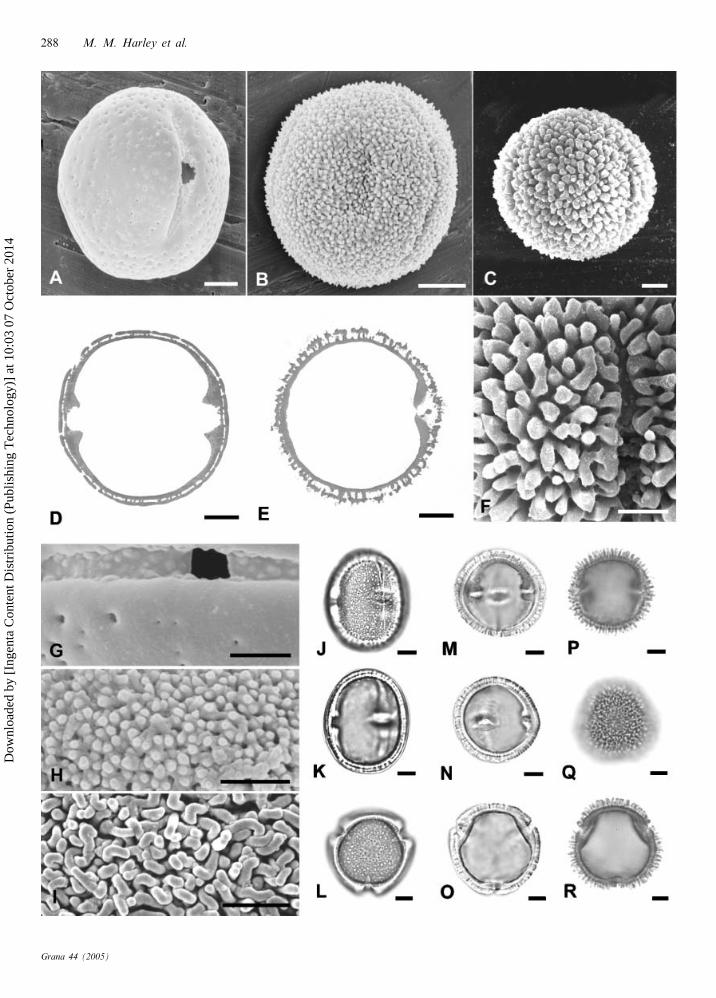
6. ‘Garuga type’ (Fig. 3A – F)
Isopolar, tricolporate and angulaperturate; prolate spher-
oidal or subprolate; outline in polar view rounded
triangular; pollen size: P – 55.5 (36 – 83) mm, E – 43.8
(33 – 73) mm [NB. Boswellia: P – 61.7 (43 – 83) mm, E – 44.9
(40 – 73) mm; Garuga: P – 49.3 (36 – 62) mm, E – 42.8
(33 – 57) mm]; colpus length: short, average or long;
endoapertures lalongate; endoaperture shape: brevi, average,
or per [per-lalongate apparently confined to Boswellia];
Fig. 1. Type 1. ‘Protium’ [A, B, D – F, J – L, P, Q] and Type 2. ‘Canarium’ [C, G – I, M – O, R]. (A, P) Protium copal: (A) equatorialmesocolpial view (SEM), (P) polar plane (TEM). (B, Q) Crepidospermum multijugum: (B) equatorial colporus view (SEM), (Q) polarplane (TEM). (C) Canarium denticulatum, mescolpial view (SEM). (D) Protium ferrugineum close up of aperture (SEM). (E, F, L) Cre-
pidospermum rhoifolium, different grains, equatorial views – note extreme width of endoapertures (LM): (E) high-mid focus, (F) midfocus, (L) low focus. (G) Canarium strictum, close up of exine surface and colporus (SEM). (H) Canarium sylvestre, close up of meso-colpium surface (SEM). (I) Canarium muelleri, close up of mesocolpium surface (SEM). (J, K) Protium opacum, different grains, equa-torial views, both high-mid focus (LM). (M – O) Canarium baileyanum (LM): (M) polar view mid-low focus, (N) equatorial view highfocus, (O) equatorial view mid focus, same grain as ‘N’. (R) Protium javanicum, polar plane (TEM). Scale bar – 2.5 mm (D, G – I);5 mm (A – C & N – R); 7.5 mm (J – M); 10 mm (E, F).
Pollen of Burseraceae 287
Grana 44 (2005)
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
10:
03 0
7 O
ctob
er 2
014
288 M. M. Harley et al.
Grana 44 (2005)
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
10:
03 0
7 O
ctob
er 2
014
endoaperture width: variable: very short, short, average,
wide or very wide [very short and very wide apparently
confined to Garuga]; costae distinct, meridionally extended,
parallel to colpi, broad at equator and tapered to a point at
colpus apices (i.e. a very elongated triangle); exine tectate
and psilate-perforate, less frequently low relief striate or
rugulate mesocolpia [differentiation of mesocolpia more
common in Garuga]; apocolpial wall thickness 2 – 4 mm,
apocolpial ultrastructure: foot layer, tectum and irregularly
columellate infratectum similar in thickness, columellae
sometimes interspersed with granular elements, no apoc-
olpial endexine.
Taxa included. – Tribe Protieae: Garuga (all) – Himalayas,
Indo-Malaya to W. Pacific; Tribe Bursereae: Boswellia
(all) – OW Tropics.
WPSF type included (Harley & Daly 1995). – Garuga flori-
bunda-type.
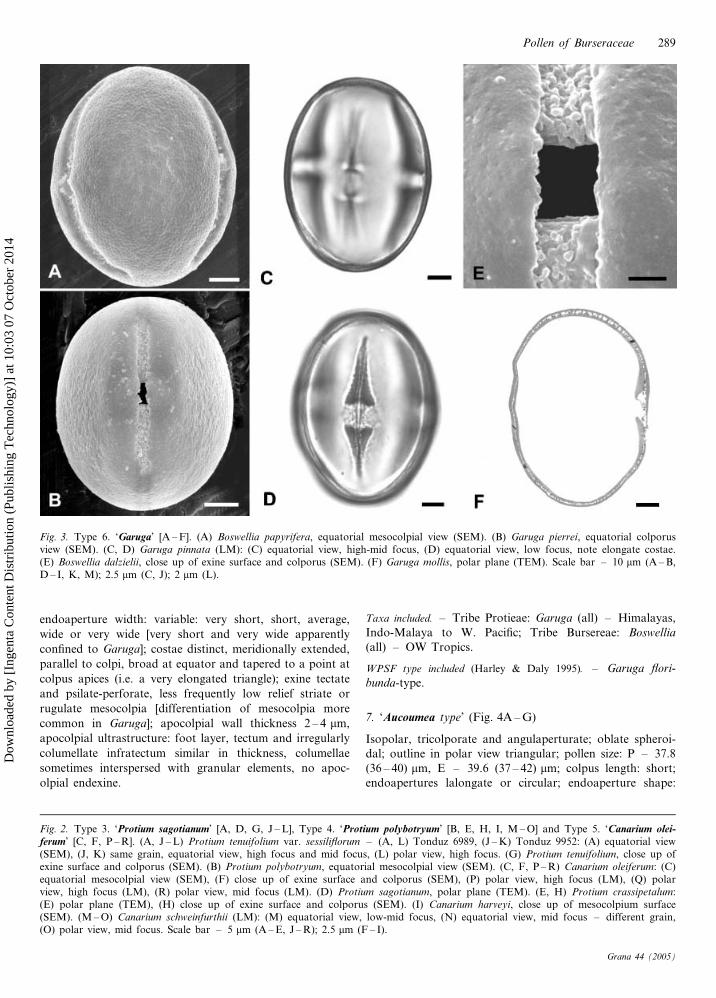
7. ‘Aucoumea type’ (Fig. 4A – G)
Isopolar, tricolporate and angulaperturate; oblate spheroi-
dal; outline in polar view triangular; pollen size: P – 37.8
(36 – 40) mm, E – 39.6 (37 – 42) mm; colpus length: short;
endoapertures lalongate or circular; endoaperture shape:
Fig. 2. Type 3. ‘Protium sagotianum’ [A, D, G, J – L], Type 4. ‘Protium polybotryum’ [B, E, H, I, M – O] and Type 5. ‘Canarium olei-ferum’ [C, F, P – R]. (A, J – L) Protium tenuifolium var. sessiliflorum – (A, L) Tonduz 6989, (J – K) Tonduz 9952: (A) equatorial view(SEM), (J, K) same grain, equatorial view, high focus and mid focus, (L) polar view, high focus. (G) Protium tenuifolium, close up ofexine surface and colporus (SEM). (B) Protium polybotryum, equatorial mesocolpial view (SEM). (C, F, P – R) Canarium oleiferum: (C)equatorial mesocolpial view (SEM), (F) close up of exine surface and colporus (SEM), (P) polar view, high focus (LM), (Q) polarview, high focus (LM), (R) polar view, mid focus (LM). (D) Protium sagotianum, polar plane (TEM). (E, H) Protium crassipetalum:(E) polar plane (TEM), (H) close up of exine surface and colporus (SEM). (I) Canarium harveyi, close up of mesocolpium surface(SEM). (M – O) Canarium schweinfurthii (LM): (M) equatorial view, low-mid focus, (N) equatorial view, mid focus – different grain,(O) polar view, mid focus. Scale bar – 5 mm (A – E, J – R); 2.5 mm (F – I).
Fig. 3. Type 6. ‘Garuga’ [A – F]. (A) Boswellia papyrifera, equatorial mesocolpial view (SEM). (B) Garuga pierrei, equatorial colporusview (SEM). (C, D) Garuga pinnata (LM): (C) equatorial view, high-mid focus, (D) equatorial view, low focus, note elongate costae.(E) Boswellia dalzielii, close up of exine surface and colporus (SEM). (F) Garuga mollis, polar plane (TEM). Scale bar – 10 mm (A – B,D – I, K, M); 2.5 mm (C, J); 2 mm (L).
Pollen of Burseraceae 289
Grana 44 (2005)
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
10:
03 0
7 O
ctob
er 2
014

Fig. 4. Type 7. ‘Aucoumea’ [A – G] and Type 8. ‘Commiphora’ [H – M]. (A – G) Aucoumea klaineana: (A) polar plane (TEM), (B) equa-torial plane (TEM), (C) close up of exine surface and colporus (SEM), (D, E) same grain equatorial view, high and mid foci (LM), (F,G) same grain, polar view, mid and low foci (LM). (H – M) Commiphora: (H, I) C. sarandensis, equatorial view, high and mid foci(LM). (J) C. mossambicencis, close up of mesocolpial exine (SEM). (K) C. africana var. tubak, group of pollen grains in different orien-tations (SEM). (L) C. insisa, close up of exine surface and colporus (SEM). (M) C. mollis, polar plane (TEM). Scale bar – 5 mm (A –E, G – H); 10 mm (F, J – N); 2.5 mm (I).
290 M. M. Harley et al.
Grana 44 (2005)
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
10:
03 0
7 O
ctob
er 2
014

brevi or circular; endoaperture width: very short; costae
distinct, circular; exine tectate and psilate-perforate; apoc-
olpial wall thickness 3.5 – 4.0 mm; apocolpial ultrastructure:
foot layer thick, tectum slightly thinner, infratectum very
thin irregular columellate, no apocolpial endexine.
Taxon included. – Tribe Bursereae: Aucoumea klaineana –
Trop. W. Africa.
8. ‘Commiphora type’ (Fig. 4H – M)
Isopolar, tricolporate and angulaperturate; oblate spheroi-
dal or prolate spheroidal; outline in polar view: circular,
less frequently lobed triangular or hexagonal; pollen size:
P – 26.5 (22 – 38) mm, E – 29.0 (23 – 40) mm; colpus length:
short or, rarely, long; endoapertures circular or lalongate,
less frequently lolongate; endoaperture shape: probably
brevi; endoaperture width: probably short; costae distinct,
or very distinct, more or less circular, less frequently sickle-
shaped, polar oriented; exine semi-tectate, foveolate or
reticulate spinulose, rarely scabrate-spinulose; apocolpial
wall thickness 1 – 4 mm; apocolpial ultrastructure: foot layer
and tectum thin, infratectum wide and columellate, no
apocolpial endexine.
Taxa included. – Tribe Bursereae: Commiphora (all) –
mostly subtropical and tropical Africa, Madagascar, a few
from Mexico, Cuba, Bahamas, Brazil; Bursera glabrifolia
(Mexico), B. grandifolia (Mexico), B. malacophylla (Ecua-
dor).
9. ‘Triomma type’ (Fig. 5A – C, H, J – L)
Isopolar, tricolporate and angulaperturate; oblate spheroi-
dal; outline in polar view rounded triangular to circular;
pollen size: P – 23.9 (21 – 27) mm, E – 25.05 (23 – 28) mm;
colpus length: short; endoapertures circular; endoaperture
width: very short; costae distinct, more or less circular; exine
tectate, exine topology scabrate-spinulose; apocolpial wall
thickness 1 – 2.5 mm; apocolpial ultrastructure: tectum thin,
foot layer very thin, infratectum columellate c. twice the
width of the tectum, distinct apocolpial endexine – thicker
than foot layer.
Taxon included. – Tribe Bursereae: Triomma malaccensis –
W. Malesia.
10. ‘Bursera type’ (Fig. 5D – G, I, M, N)
Isopolar, tricolporate, lobed and angulaperturate; oblate
spheroidal or prolate spheroidal; outline in polar view: lobed
triangular; pollen size: P – 26.3 (17 – 41) mm, E – 25.3
(16 – 39) mm; colpus length: short, less frequently average;
endoapertures lalongate; endoaperture shape: average or
per; endoaperture width: average or wide; costae very
distinct, sickle-shaped, polar oriented; exine semitectate,
distinctly striate – striae non-aligned or ‘woven’ or with
uni-directional alignment or striate-reticulate or striate
perforate; apocolpial wall thickness 1 – 3 mm; apocolpial
ultrastructure foot layer, tectal elements and columellate
infratectum similar in width or infratectum may be thinner,
no apocolpial endexine.
Taxon included. – Tribe Bursereae: Bursera (most) – Meso-
and tropical South America.
11. ‘Protium robustum type’ (Fig. 6A – E, J, K)
Isopolar, tricolporate and angulaperturate; oblate spheroidal;
outline in polar view rounded triangular; pollen size: P – 21.4
(19 – 24) mm, E – 22.5 (21 – 25) mm; colpus length: long;
endoapertures lalongate; endoaperture shape: average; endoa-
perture width: very short; costae distinct, sickle-shaped, polar
oriented; exine semi-tectate, and coarsely reticulate, lumina
shallow and bases sparsely granular, muri psilate and rounded;
apocolpial wall thickness c. 1.5 mm; apocolpial ultrastructure:
thick muri and foot layer, infratectum slightly thinner and
columellate, no apocolpial endexine.
Taxon included. – Tribe Protieae: Protium robustum –
Brazil.
WPSF type included (Harley & Daly 1995). – Protium robus-
tum-type.
12. ‘Rosselia type’ (Fig. 6F – I, L – N)
Isopolar, tricolporate and planaperturate; oblate spheroidal
to prolate spheroidal; outline in polar view rounded
triangular; pollen size: P – 28.2 (26 – 30) mm, E – 29.0
(26 – 31) mm; colpus length: short; endoapertures lalongate;
endoaperture shape: brevi; endoaperture width: very short;
costae distinct, sickle-shaped, polar oriented; exine semi-
tectate and very coarsely reticulate, lumina deep with
granular bases, muri psilate and angular; apocolpial wall
thickness 3 – 4 mm, apocolpial ultrastructure: muri thin and
pointed, supported by columellae of similar height and
width, foot layer thinner than the columellar layer or the
muri, no apocolpial endexine.
Taxon included. – Tribe Canarieae: Rosselia bracteata –
New Guinea (Rossel Island).
13. ‘Canarium gracile type’ (Fig. 7A, D, G, H)
Isopolar or subisopolar, triporate projectate, and angula-
perturate; suboblate; outline in polar view, triangular
projectate; pollen size: P – 25.9 (21 – 30) mm, E – 31.3
(28 – 35) mm; costae very pronounced, polar oriented, slight
meridional elongation; exine tectate, exine topology sca-
brate-rugulate; apocolpial wall thickness 1.5 – 2.5 mm, apoc-
olpial ultrastructure unknown.
Taxon included. – Tribe Canarieae: Canarium gracile Engl.
– Philippines.
14. ‘Scutinanthe brunnea type’ (Fig. 7B, C, E, F, I, J)
Heteropolar, tricolporate lobed vestibulate (rarely dicolpo-
rate lobed vestibulate) and angulaperturate, frequently (c.
75%) with radial asymmetry of polarisation for each
aperture lobe; suboblate; outline in polar view triangular
lobate; pollen size: P – 26.4 (24 – 28) mm, E – 34.5
(32 – 39) mm; colpus length: short; endoapertures subcircu-
lar, endoaperture shape: brevi; endoaperture width: very
short; costae not present; exine tectate, and coarsely
perforate-rugulate, aperture lobes more or less psilate with
Pollen of Burseraceae 291
Grana 44 (2005)
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
10:
03 0
7 O
ctob
er 2
014

Fig. 5. Type 9. ‘Triomma’ [A – C, H, J, K] and Type 10. ‘Bursera’ [D – G, I, M, N]. (A – C, H, J – L) Triomma malaccensis – (A – C, H, J)Yong 94742; (L) Kedah KFN27479: (A) equatorial view (SEM), (B) polar plane, endexine (arrows) underlies a very thin foot layer (TEM),(C) equatorial plane (TEM), (H) polar view (SEM), (J) equatorial view, high focus (LM), (K) equatorial view, mid focus (LM), (L) polarview, high-mid focus (LM). (D – G, I, M, N) Bursera: (D) B. confusa, equatorial view (SEM), (E – G, I) B. kerberi: (E) polar plane (TEM),(F) polar view (SEM), (G) equatorial plane (TEM), (I) close up of mesocolpial exine surface (SEM). (M – N) B. discolor: (M) equatorialview, high-mid focus (LM), (N) polar view, high-mid focus (LM). Scale bar – 5 mm (A – H); 2 mm (I); 10 mm (J – N).
292 M. M. Harley et al.
Grana 44 (2005)
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
10:
03 0
7 O
ctob
er 2
014
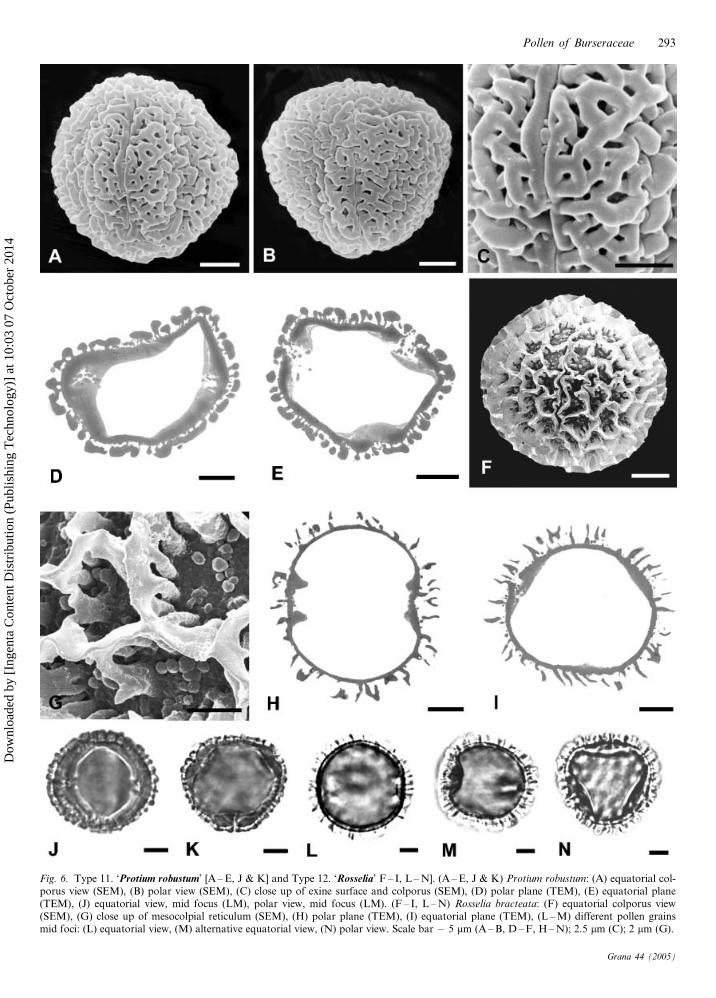
Fig. 6. Type 11. ‘Protium robustum’ [A – E, J & K] and Type 12. ‘Rosselia’ F – I, L – N]. (A – E, J & K) Protium robustum: (A) equatorial col-porus view (SEM), (B) polar view (SEM), (C) close up of exine surface and colporus (SEM), (D) polar plane (TEM), (E) equatorial plane(TEM), (J) equatorial view, mid focus (LM), polar view, mid focus (LM). (F – I, L – N) Rosselia bracteata: (F) equatorial colporus view(SEM), (G) close up of mesocolpial reticulum (SEM), (H) polar plane (TEM), (I) equatorial plane (TEM), (L – M) different pollen grainsmid foci: (L) equatorial view, (M) alternative equatorial view, (N) polar view. Scale bar – 5 mm (A – B, D – F, H – N); 2.5 mm (C); 2 mm (G).
Pollen of Burseraceae 293
Grana 44 (2005)
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
10:
03 0
7 O
ctob
er 2
014
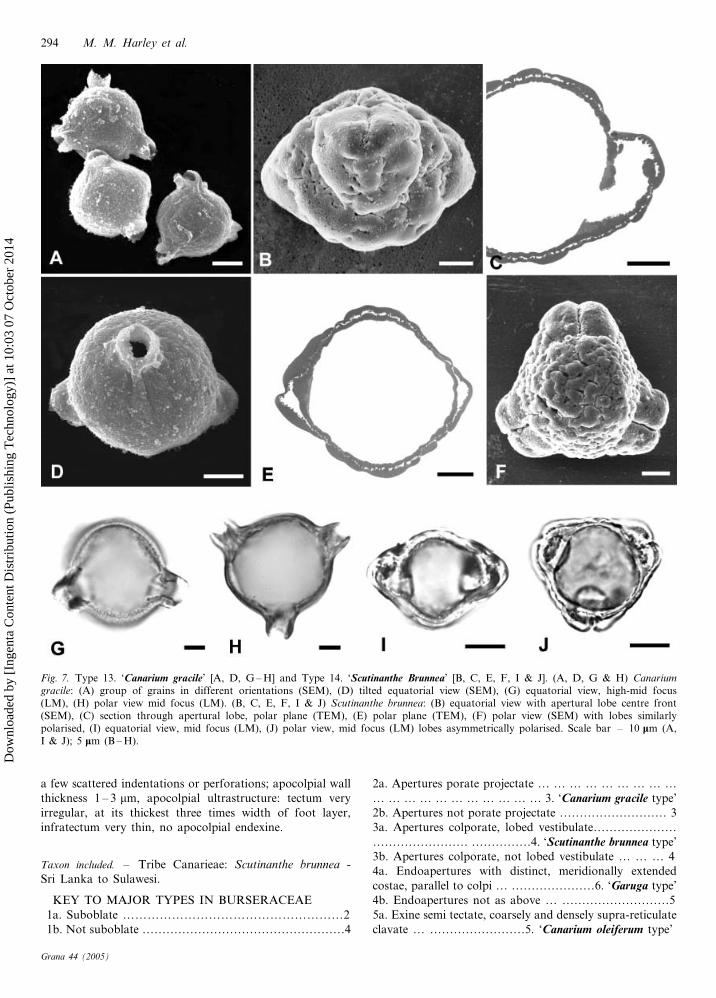
a few scattered indentations or perforations; apocolpial wall
thickness 1 – 3 mm, apocolpial ultrastructure: tectum very
irregular, at its thickest three times width of foot layer,
infratectum very thin, no apocolpial endexine.
Taxon included. – Tribe Canarieae: Scutinanthe brunnea -
Sri Lanka to Sulawesi.
KEY TO MAJOR TYPES IN BURSERACEAE
1a. Suboblate ………………………………………………2
1b. Not suboblate ……………………………………………4
2a. Apertures porate projectate … … … … … … … … …
… … … … … … … … … … … 3. ‘Canarium gracile type’
2b. Apertures not porate projectate ……………………… 3
3a. Apertures colporate, lobed vestibulate…………………
…………………… ……………4. ‘Scutinanthe brunnea type’
3b. Apertures colporate, not lobed vestibulate … … … 4
4a. Endoapertures with distinct, meridionally extended
costae, parallel to colpi … …………………6. ‘Garuga type’
4b. Endoapertures not as above … ………………………5
5a. Exine semi tectate, coarsely and densely supra-reticulate
clavate … ……………………5. ‘Canarium oleiferum type’
Fig. 7. Type 13. ‘Canarium gracile’ [A, D, G – H] and Type 14. ‘Scutinanthe Brunnea’ [B, C, E, F, I & J]. (A, D, G & H) Canarium
gracile: (A) group of grains in different orientations (SEM), (D) tilted equatorial view (SEM), (G) equatorial view, high-mid focus(LM), (H) polar view mid focus (LM). (B, C, E, F, I & J) Scutinanthe brunnea: (B) equatorial view with apertural lobe centre front(SEM), (C) section through apertural lobe, polar plane (TEM), (E) polar plane (TEM), (F) polar view (SEM) with lobes similarlypolarised, (I) equatorial view, mid focus (LM), (J) polar view, mid focus (LM) lobes asymmetrically polarised. Scale bar – 10 mm (A,I & J); 5 mm (B – H).
294 M. M. Harley et al.
Grana 44 (2005)
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
10:
03 0
7 O
ctob
er 2
014

5b. Exine not as above … …………………………………6
6a. Exine coarsely reticulate ……………………………… 7
6b. Exine not as above …………………………………… 2
7a. Lumina very wide, and deep with angular muri
……………………………………8. ‘Rosselia bracteata type’
7b. Lumina wide, and shallow with rounded muri …
……………………………………11. Protium robustum type’
8a. Exine striate …………………………………………… 9
8b. Exine not striate ………………………………………10
9a. Outline in polar view strongly lobed-triangular, shape
spheroidal or prolate spheroidal …………10. ‘Bursera type’
9b. Outline in polar view triangular, or rounded triangular,
shape usually subprolate … ……………2. ‘Canarium type’
10a. Exine tectate, and psilate-perforate or foveolate …11
10b. Exine not as above ……………………………………13
11a. Shape oblate spheroidal, outline in polar view
distinctly triangular … …………………7. ‘Aucoumea type’
11b. Shape subprolate, prolate-spheroidal to prolate,
outline in polar view rounded triangular ………………12
12a. Psilate, usually finely perforate in mesocolpia, psilate
apocolpia …………………………………1. ‘Protium type’
12b. Coarsely perforate or foveolate, frequently more
pronounced in apocolpia ……3. ‘Protium sagotianum type’
13a. Exine spinulose, tectate ……………9. ‘Triomma type’
13b. Exine spinulose, semitectate … ……………………14
14a. Foveolate or reticulate, colpi usually short ……………
…………………………………………8. ‘Commiphora type’
14b. Microreticulate, colpi average to long…………………
………………………………4. ‘Protium polybotryum type’
DISCUSSION
The pollen types described are based on combinations of
varying aperture and exine morphology, as well as shape
and size. Of the 14 pollen types described 13 are
tricolporate, and range from oblate spheroidal to
prolate, characteristics frequently encountered in other
eudicotyledous families. Nevertheless, there are only four
pollen types where the overall combination of characteristics
is not obviously diagnostic to family level, these are,
‘Canarium type’, ‘Protium sagotianum type’, ‘Garuga type’
and ‘Protium robustum type’.
The pollen of Rosselia, originally described in Forman
et al. (1994), is not only unique within Burseraceae, it is also
unusual outside the family. In particular, the appearance of
the reticulum and the granulae in the bases of the lumina are
very similar to the reticulate pollen exine in some species of
Ruellia (Acanthaceae). However, the exine of Rosselia pollen
has a clearly defined foot layer while, in Ruellia, pollen with
a similar exine has no foot layer. Furthermore, in Ruellia
pollen there are 3, rarely 5, pores or, occasionally a short
colporus (Furness & Grant 1996).
The remaining nine pollen types are unique to Burser-
aceae, ‘Protium type’, ‘Protium polybotryum type’, ‘Canar-
ium oleiferum type’, ‘Aucoumea type’, ‘Commiphora type’,
‘Triomma type’, ‘Bursera type’, ‘Canarium gracile type’ and
‘Scutinanthe brunnea type’. Of these seven are represented by
only one species, ‘Canarium oleiferum type’, ‘Aucoumea
type’, ‘Triomma type’, ‘Protium robustum type’ ‘Rosselia
type’, ‘Canarium gracile type’ and ‘Scutinanthe brunnea
type’. The last two types represent extremely unusual
dicotyledonous pollen aperture morphologies. The distribu-
tion of pollen morphological types across the three most
widely accepted tribes of Burseraceae (Table VI) is inter-
esting, as it draws attention to some of the often debated
relationships within the family (Harley & Daly 1995).
Protium (147 species) is the largest genus of tribe Protieae
and Canarium (75 species) is the largest genus of tribe
Canarieae. The ‘Protium type’ (Fig. 1A, B, D – F, J – L, P,
Q) and the ‘Canarium type’ (Fig. 1C, G – I, M – O, R) share
some basic characteristics which are frequently encountered
in eudicot pollen: tricolpory, and a prolate spheroidal to
prolate shape, with a tectate exine. However, in the ‘Protium
type’ the often unusually wide endoapertures (Fig. 1E, F,
J – L), and a thin granular-columellate infratectum (Fig. 1P,
Q) are less commonly encountered than the smaller
endoapertures (Fig. 1N, O) and columellate infratectum
(Fig. 1P) of the ‘Canarium type’. Notably, however, with the
exception of the pollen of Beiselia mexicana, neither of these
pollen types occurs in tribe Bursereae.
In tribe Protieae all species of Tetragastris and Crepidos-
permum, as well as most Protium species have ‘Protium type’
pollen. A few species included in ‘Protium type’, particularly
in Crepidospermum, are distinguished by very wide endoa-
pertures which more or less form an endocingulum around
the equator of the grain (Fig. 1E, F & L); in all other
respects the pollen conforms to the ‘Protium type’. In
Protium there are a few Old World species: P. connarifolium
Merrill, P. javanicum Burm. f., P. madagascariense Engl., P.
obtusifolium Marchand, P. serratum (Wall. ex Colebr.) Engl.
and a few New World species: P. apiculatum Swart, P.
amazonicum (Cuatrec.) Daly (syn. P. fimbriatium Swart), P.
urophyllidium Daly which have pollen with a striate
‘Canarium type’ exine. In tribe Canarieae most species of
Trattinnickia and Scutinanthe brevisepala, have ‘Protium
type’ pollen, while in Dacryodes, Santiria and Haplolobus
some species have ‘Protium type’ pollen, and some species
have ‘Canarium type’ pollen. However, the majority of
Canarium species have striate ‘Canarium type’ pollen. It is
difficult to make clear distinctions between striate Protium
pollen, and the striate pollen in tribe Canarieae. Never-
theless, there are characters which, in a more refined
treatment of the ‘Canarium type, would probably support
the circumscription of one or two subtypes; for example,
shape – 60% of the Protium species included in ‘Canarium
type’ has prolate pollen, while in 97% of the Canarieae
species included the pollen is either prolate-spheroidal or
sub-prolate.
The coarsely perforate ‘Protium sagotianum type’
(Fig. 2A, D, G, J – L) and the microreticulate, spinulose
‘Protium polybotryum type’ (Fig. 2B, E, H, I, M – O) are also
represented only in tribes Protieae and Canarieae, albeit the
number of species in each case is few. The original ‘Protium
polybotryum type’ (Harley & Daly 1995) included three
Protium species, but now four species of Canarium are also
included. However, the exine surface and infratectum of the
New World Protium species are slightly different (cf.
Pollen of Burseraceae 295
Grana 44 (2005)
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
10:
03 0
7 O
ctob
er 2
014

Fig. 2H with Fig. 2I) and probably justify the distinction of
a subtype for the Canarium species included in this type.
The pollen of Canarium oleiferum (Fig. 2C, F & P – R) is
described here for the first time. It is one of the seven pollen
types in Burseraceae represented by only one species. The
clavate exine has not yet been thin-sectioned but, from light
microscopy, it appears that the clavae arise from a thin,
finely reticulate tectum beneath which is a columellate
infratectum and a thin foot layer; endexine also appears to
be present away from aperture areas.
The two genera included in the ‘Garuga type’ (Fig. 3A – F)
are usually placed in separate tribes, Garuga in Protieae, and
Boswellia in Bursereae, although a clearly defined suite of
characters which justify two distinct tribes has never been
fully resolved (Harley & Daly 1995). Both genera are from
the Old World: Garuga – Himalayas, Indomalaya to
western Pacific; Boswellia – Old World tropics, especially
Africa and India. The pollen is strikingly similar in the two
genera, and of a type commonly encountered in the
eudicotyledons, although within Burseraceae it is unique
to these two genera. The pollen type is based partly on size,
but mainly on the configuration of the colporate apertures
which have meridionally extended, tapering costae
(Fig. 3D). Notably, a DNA study of Burseraceae (Clarkson
et al. 2002) resolves Garuga and Boswellia as sister taxa.
The pollen of the monospecific genus Aucoumea
(Fig. 4A – G) is very distinctive both within and outside
the family. However, the characteristics of circular endoa-
pertures and circular costae are shared with two other
genera of Burseraceae: Triomma and Commiphora. All three
genera are in tribe Bursereae, although Triomma is in
subtribe Boswelliinae, and Commiphora is in subtribe
Bursereae (Table VI). Interestingly the ultrastructure of
Aucoumea pollen, thin infratectum, thick tectum and thick
foot layer is very similar to the ultrastructure of ‘Protium
type’ (cf. Fig. 1P, Q with Fig. 4A) but in other respects the
pollen is notably dissimilar.
‘Triomma type’ pollen (Fig. 5A – C, H & J – L) is
apparently unique in the family; in no other species of
Burseraceae is there pollen with an endexine which extends
across the apocolpial region (Fig. 5B). Furthermore, with
one rare exception in Commiphora – C. erosa Vollesen, it is
the only example of tectate spinulose pollen in the family.
Commiphora is the largest genus in the family with almost
200 species. Its pollen is distinctive and characteristic of the
genus (Fig. 4H – M). However, a few tropical American
species of the mainly Mexican genus Bursera, which is
considered to be closely related to the predominantly
African Commiphora, have been transferred to Commiphora:
C. leptophloeus (Mart.) Gillett (Gillett 1980), C. sarcopoda
(Paul G. Wilson) Rzed. & R. Palacios, C. tecomaca (DC.)
Rzed. & R. Palacios (Rzedowski & Palacios-Chavez 1985),
C. angustata (Griseb.) Moncada, C. glauca (Griseb.)
Moncada and C. inaguensis (Britton) Moncada (Moncada-
Ferrera 1989). Apart from the situation in C. inaguensis,
where two of the three collections which have been
examined have Bursera-like pollen, the pollen data strongly
supports the transfer of these species to Commiphora. The
DNA study by Clarkson et al. (2002) also supports a close
relationship between Commiphora and Bursera. In addition
there are other Bursera species which have Commiphora-type
pollen: B. glabrifolia Engl, B. grandifolia Engl. and B.
malacophylla Robinson.
The ‘Bursera type’ (Fig. 5D – G, I, M, N), like the
‘Canarium type’, has a striate exine. However, the striae
are more strongly pronounced in Bursera pollen and often
have a distinctly reticulate underlying structure (cf. Fig. 1H,
I with Fig. 5F, I). Furthermore, the striae, combined with a
triangular, trilobate outline in polar view, short colpi
(usually) and pronounced costae give the pollen of Bursera
a characteristic and unique appearance.
In ‘Protium robustum type’ (Fig. 6A – E, J & K) the colpi
are very long with a small apocolpial area (Fig. 6B). A
coarsely reticulate exine is rare in the family, the only other
example being the pollen of ‘Rosselia bracteata type’.
However, the reticulum and ultrastructure are strikingly
different between the two species. In P. robustum (Fig. 6C)
the lumina are small and shallow, the muri wide and
rounded, and the columellae short, while in R. bracteata
(Fig. 6F – I, M & N) the lumina are large and deep, the muri
narrow and angular and the columellae long.
‘Canarium gracile type’ (Fig. 7A, D, G & H) is the second
of two previously undescribed Burseraceae pollen types,
both are in Canarium. The three sub equatorial, porate,
projectate apertures are probably unique among extant
angiosperms, and it is all the more remarkable for occurring
in a large (c. 75 spp.) genus where all other species have
pollen which is isopolar and tricolporate. The best known
examples of tri-projectate pollen are those produced by
some members of the Loranthaceae (Feuer & Kuijt 1979);
these appear to have some affinity with the fossil pollen
genus Aquilapollenites Rouse (Rouse 1957). However, the
Loranthaceous examples are extreme triprojectate, syncol-
pate, and decidedly unlike the pollen of Canarium gracile.
Pollen of some species of Persoonia (Proteaceae) (Feuer
1986) has greater similarity to the pollen of C. gracile, but
there are two important differences: the porate projections
are strongly oriented towards one pole, and not visible when
viewed from the opposing pole which is not the case with C.
gracile. Secondly, in C. gracile pollen the porate projections
have very distinct costate thickenings (Fig. 7G, H) which are
not present in Persoonia pollen (Feuer 1986).
The pollen morphology of Scutinanthe brunnea (Fig. 7B,
C, E, F, I & J) is also very unusual, and this is amplified by
the simple, isopolar and tricolporate pollen of the only other
species of the genus: S. brevisepala Leenh. A detailed
description and discussion of the curious heteropolar pollen,
frequently with radially asymmetric polarisation of each
aperture lobe, was provided by Segaar & Van der Ham
(1993). There is no comparable pollen in extant angios-
perms. However, Segaar and Van der Ham (1993) discuss
the ‘‘shift’’ of the ecto- and endoaperture to opposite non
equatorial positions, and cite the example of the fossil
Normapolles group, where this unusual irregularity is also
known to occur (see Batten & Christopher 1981). The
Normapolles group was first proposed by Pflug (1953) to
include fossil pollen grains distinctly different from pollen
produced by living angiosperms. The first representatives are
noted from the middle Cenomanian, followed by rapid
diversification throughout the remainder of the Cretaceous
296 M. M. Harley et al.
Grana 44 (2005)
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
10:
03 0
7 O
ctob
er 2
014

and early Tertiary, but by the end of the Eocene they have
become virtually extinct. There a number of examples of
Normapolles pollen bearing notable similarity to ‘Scuti-
nanthe brunnea type’, for example, some species of
Emscheripollis Krutsch, Trudopollis Pflug (Pacltova 1981)
and Plicapollis Pflug (Mikhelis 1981). A great many genera
have been described for dispersed pollen assigned to the
Normapolles group (see for example, Batten & Christopher
1981). So far, however, fossil flowers with in situ
Normapolles type pollen (see Friis et al. 2003 for summary)
have been associated with the Fagales (sensu APG 1998,
2003). The fossil flowers are very small and usually bisexual.
While the inflorescences of Scutinanthe brunnea have small
(c. 3 – 5 mm long) unisexual flowers; the male flowers have
8 – 10 stamens and the female flowers have 2-celled ovaries
and one-seeded fruits. Unisexual fossil flowers with
Normapolles type pollen are rare, they include Bedellia
(Sims et al. (1999) and Endressianthus (Friis et al. 2003). The
suggested affinity for both genera is with Betulaceae (Sims
et al. (1999, Friis et al. 2003).
CONCLUSIONS
The pollen morphology of Burseraceae is extremely varied in
both exine and aperture variants, and includes a number of
pollen types unique to the family. Of 14 major pollen types
seven of the more unusual types are represented by only one
taxon; three of the taxa represent monospecific genera. It is
particularly remarkable that the family has two outstanding
examples of pollen with highly modified aperture systems.
One, Scutinanthe brunnea, shares similarities with some taxa
in the extinct Normapolles group, while the other, Canarium
gracile, occurs in a genus with generally unspecialised
tricolporate pollen. A substantial number of taxa in tribes
Protieae and Canarieae share the characteristics of pollen
types 1 to 4, and these are morphologically the least
specialised pollen types in the family. With the exceptions of
the pollen of Beiselia mexicana and Boswellia spp. the pollen
of tribe Bursereae is highly distinctive, and unique to the
tribe. Nevertheless, tribe Canarieae also includes four very
distinctive, monospecific pollen types. Pollen morphology
and DNA data support the long held opinion of a close
relationship between Bursera and Commiphora (Bursereae)
and, perhaps most interestingly, pollen morphology and
DNA data suggest a relationship between Garuga and
Boswellia which supports Daly’s comment (in Harley &
Daly 1995) that ‘‘Garuga is a morphological outlier in the
Protieae and should probably be transferred to the
Bursereae.’’
ACKNOWLEDGEMENTS
We thank the staff of the Leiden and Utrecht Herbaria for
permitting MMH to remove pollen samples from their Herbarium
collections, especially for tribe Canarieae. We also thank Dr Andrea
Weeks, University of Texas at Austin, for sending us samples of
Bursera pollen; as well as our reviewers Dr Raymond Van der Ham
and Dr Douglas Daly for their helpful comments on the submitted
manuscript.
SPECIMENS ILLUSTRATED
Aucoumea klaineana Pierre, Klaine 100, Gabon (K)
Boswellia dalzielii Hutch., Dalziel 279, Nigeria (K)
Boswellia papyrifera (Del.) Hochst., Eggeling 1238, Uganda (K)
Bursera confusa (Rose) Engl., Hinton 7772, Mexico (K)
Bursera discolor Rzed., Hinton et al. 7736, Mexico (K)
Bursera kerberi Engl., Hinton 10429, Mexico (K)
Canarium baileyanum Leenh., Kew Herbarium No. 227, Australia
(K)
Canarium denticulatum Blume, Parkinson 19, Andaman Isls. (K)
Canarium gracile Engl., Ramos 1130, Philippines (U)
Canarium harveyi Seem., A.C. Smith 1151, Fiji (K)
Canarium muelleri F.M. Bailey, Herbarium Lugd. Bat. 401390,
Australia (L)
Canarium oleiferum Baill., de Alleizette s.n., New Caledonia (L)
Canarium paniculatum Benth. ex Engl., Vaughan 12515, New
Guinea (K)
Canarium strictum Roxb., Venkata Reddi 99026, India (L)
Canarium sylvestre Gaertn., Robinson 878, Philippines (L)
Commiphora africana (A. Rich.) Engl. var. tubuk (Sprague)
Gillett, Carter 886, Somalia (K)
Commiphora incisa Chiov., Carter & Stannard 595, Kenya (K)
Commiphora mollis (Oliv.) Engl., Menezes 888, Angola (K)
Commiphora mossambicencis (Oliv.) Engl., Burtt 4589, Tanzania
(K)
Commiphora sarandensis Burtt, Burtt 3979, Tanganyika (K)
Crepidospermum multijugum Swart, Klug 2284, Peru (K)
Crepidospermum rhoifolium (Benth.) Tr. & Pl., Palacios 3021,
Ecuador (K)
Garuga floribunda Decne., Koorders VIB89, Celebes (K)
Garuga pierrei Guillaumin, Collins 73, Thailand (K)
Garuga pinnata Roxb., Drummond 3112, India (K)
Protium copal (Schltdl. & Cham.) Engl., Borgeau 2058, Mexico (K)
Protium crassipetalum Cuatrec., Campbell et al. P21923, Brazil (K)
Protium ferrugineum (Engl.) Engl., Pipoly et al. 6725 (K)
Protium javanicum Burm., f. Kostermans 18001, Indonesia (K)
Protium opacum Swart, Krukoff 4911, Brazil (K)
Protium polybotryum (Turcz.) Engl., Hostman 1268, Surinam (K)
Type
Protium robustum ( Swart) D.M. Porter, Daly et al. 1166, Brazil (K)
Protium sagotianum Marchand, Daly et al. 5124, Peru (K)
Protium tenuifolium (Engl.) Engl., Rogers 10117, s.l. Trinidad (K)
Protium tenuifolium (Engl.) Engl. var. sessiliflorum (Rose)
D.M. Porter, Tonduz 6989 (K - Type); 9952 Costa Rica (K)
Rosselia bracteata Forman, Katik et al. LAE70973, New
Guinea: Rossel Island (L)
Scutinanthe brunnea Thwaites, Endert 25 E IP 513, Indonesia:
Sumatra (K)
Triomma malaccensis Hook. f., Yong 94742 (K); Kedah Kepong
Field no. 27479, Malysia: Malay Penins. (K)
REFERENCES
Aguilar-Sierra, C. I. & Melhem, T. S. 1998a. Morfologia polınica da
tribo Bursereae (Burseraceae) na America do Sul. – Rev. Bras.
Bot. Reg. (Sao Paulo) 21: 17 – 26.
Aguilar-Sierra, C. I. & Melhem, T. S. 1998b. Morfologia polınica da
tribo Canarieae (Burseraceae) na America do Sul. – Rev. Bras.
Bot. Reg. (Sao Paulo) 21: 27 – 34.
Aguilar-Sierra, C. I. & Melhem, T. S. 1998c. Morfologia polınica da
Pollen of Burseraceae 297
Grana 44 (2005)
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
10:
03 0
7 O
ctob
er 2
014

tribo Protieae (Burseraceae) na America do Sul. – Rev. Bras.
Bot. Reg. (Sao Paulo) 21: 35 – 63.
Angiosperm Phylogeny Group (APG) 1998. An ordinal classifica-
tion for the families of flowering plants. – Ann. Mo. Bot. Gard.
85: 531 – 553.
Angiosperm Phylogeny Group (APG) 2003. An update of the
Angiosperm Phylogeny Group classification for the orders and
families of flowering plants: APG II. – Bot. J. Linn. Soc. 141:
399 – 436.
Arreguın-Sanchez, M. L., Palacios-Chavez, R. & Quiroz-Garcıa, D.
L. 1995. Morfologıa de los granos de polen de las familias
Rutaceae y Zygophyllaceae de la Estacion de Biologıa Chamela,
Jalisco, Mexico. – In: Mems 8o Coloq. Int. Paleobot. Palinol.,
Jalisco 1994, pp. 95 – 106. – Mexico DF.
Baksi, S. K. 1976. Pollen morphology of the genera Gluta Linnaeus
and Melanorrhoea Wallich (Anacardiaceae). – In: The evolu-
tionary significance of the exine (ed. I. K. Ferguson & J. Muller),
pp. 379 – 405. – Linn. Soc. Symp. Ser. I. Acad. Press, London/
New York.
Barth, O. M. 1980. Pollen morphology of a Brazilian Rutaceae:
Zanthoxylum (Fagara). – Pollen Spores 22: 425 – 436.
Barth, O. M. 1982. Variacoes polinicas em especies Brasileiras da
familia Rutaceae. – Bol. Inst. Geosci., Univ. Sao Paulo 13:
129 – 134.
Barth, O. M. 1983. Pollen morphology of Brazilian Rutaceae:
Dictyoloma and Hortia. – Pollen Spores 25: 409 – 420.
Barth, O. M. 1985. Pollen morphology of Brazilian Rutaceae:
Pilocarpus. – Pollen Spores 27: 145 – 153.
Basak, R. K. 1968. Studies on the pollen morphology of
Simaroubaceae. – Bull. Bot. Surv. India (1967) 9: 63 – 67.
Batten, D. J. & Christopher, R. A. 1981. Key to the recognition of
Normapolles and some morphologically similar pollen genera. –
Rev. Palaeobot. Palynol 35: 359 – 383.
Bentham, G. & Hooker, J. D. 1862. Geraniales: 49. Rutaceae, 50.
Simaroubaceae, 52. Burseraceae, 53. Meliaceae. – In: Genera
Plantarum. Vol. 1, P. 1, (ed. G. Bentham & J. D. Hooker),
pp. x – xi, 278 – 316 & 321 – 340. – A. Black, W. Pamplin, L,
Reeve, Williams & Norgate, London.
Brummitt, R. K. 1992. Vascular plant families and genera. – R.
Bot. Gards, Kew.
Clarkson, J. J., Chase, M. W. & Harley, M. M. 2002. Phylogenetic
relationships in Burseraceae based on plastid rps16 intron
sequences. – Kew Bull. 57: 183 – 193.
Correa, A. M. S., Watanabe, H. M. & Melhem, T. S. 1992. Flora
polınica da reserva do Parque estadual das Fontes do Ipiringa
(Sao Paulo, Brasil). Famılias: 44. Tiliaceae e 116. Rutaceae. –
Hoehnea 19: 117 – 124.
Cronquist, A. 1981. An integrated system of classification of
flowering plants. – Columbia Univ. Press, New York.
Dahlgren, R. M. T. 1983. General aspects of angiosperm evolution
and macrosystematics. – Nord. J. Bot. 3: 119 – 149.
Dalla Torre, C. G. de & Harms, H. 1901. 137. Rutaceae, 138.
Simaroubaceae, 139. Burseraceae, 140. Meliaceae. – In: Genera
Siphonogamarum ad systema Englerianum conscripta. 4. (ed. C.
G. Dalla Torre, C. G. de & H. Harms, H.), pp. 250 – 262. – W.
Engelmann, Lipsiae (Leipzig).
Engler, A. 1931. Rutaceae. – In: Die naturlichen Pflanzenf-
Familien. 2 Aufg, 19a. Pandales, Geraniales (ed. A. Engler, A.
& K. Prantl), pp. 405 – 456. – W. Engelmann, Leipzig.
Erdtman, G. 1952. Pollen morphology and plant taxonomy.
Angiosperms. – Almqvist & Wiksell, Stockholm.
Feuer, S. 1986. Pollen morphology and evolution in the Persoo-
nioideae, Sphalmioideae and Carnarvonioideae (Proteaceae). –
Pollen Spores 28: 123 – 156.
Feuer, S. & Kuijt, J. 1979. Fine structure of Mistletoe pollen II.
Pollen morphology and evolution in Psittacanthus (Lorantha-
ceae). – Bot. Not. 132: 295 – 309.
Forman, L. L., Brandham, P. E., Harley, M. M. & Lawrence, T. J.
1989. Beiselia mexicana (Burseraceae) and its affinities. – Kew
Bull. 44: 1 – 31.
Forman, L. L., van der Ham, R. W. J. M., Harley, M. M. & Lawrence,
T. J. 1994. Rosselia, a new genus of Burseraceae from the Louisade
Archipelago, Papua New Guinea. – Kew Bull. 49: 601 – 621.
Friis, E.-M., Pedersen, K. R. & Schoneneberger, J. 2003. End-
ressianthus, a new Normapolles-producing plant genus of
Fagalean affinity from the Late Cretaceous of Portugal. –
Int. J. Pl. Sci. 164(5 Suppl.): 201 – 223.
Furness, C. A. & Grant, M. C. 1996. Pollen morphology of some
Ruellia species (Acanthaceae) from Africa and Madagascar. –
Grana 35: 231 – 239.
Gillett, J. 1980. Commiphora (Burseraceae) in South America and its
relationship to Bursera. – Kew Bull. 34: 569 – 587.
Grant, M., Blackmore, S. & Morton, C. 2000. Pollen morphology
of the subfamily Aurantoideae (Rutaceae). – Grana 39: 8 – 20.
Harley, M. M. 1986. Pollen of Haplophyllum, Ruta and Thamnosma
(Rutaceae). – In: Taxonomic revision of the genus Haplophyllum
(Rutaceae) by C.C. Townsend – Hooker’s Icones Plant. 40: 6 – 11.
Harley, M. M. & Hall, D. 1999. Pollen morphology of the African
Burseraceae and related genera. – Palaeoecol. Africa & Sur. Isls
26: 225 – 242.
Harley, M. M. & Daly, D. 1995. Burseraceae: Protieae. – World
Pollen Spore Flora 20. Scand. Univ. Press, Stockholm/Oslo/
Copenhagen.
Ibe, R. A. & Leis, R. A. 1970. Pollen morphology of the
Anacardiaceae of northeastern North America. – Bull. Torrey
Bot. Club 106: 140 – 144.
Lam, H. J. 1932. The Burseraceae of the Malay Archipelago and
Peninsula. – Bull. Jard. Bot. Buitenzorg 3e Ser 12: 281 – 561.
Lobreau-Callen, D., Oltmann, O. & Straka, H. 1975. Rutaceae. –
Palynologia Madagassica et Mascarenica. Fam. 104. (ed. H.
Straka). Pollen Spores 17: 24 – 27.
Melchior, H. (Ed.) 1964. 27. Rutales. – In: Syllabus der
Pflanzenfamilien. 12 ed. Bd 2 (ed. A. Engler), pp. 262 – 277. –
Gebr. Borntraeger, Berlin.
Mikhelis, A. A. 1981. Normapolles pollen in Cretaceous/Palaeogene
boundary deposits of the Priazov’ye (Azov Sea area). – Rev.
Palaeobot. Palynol. 35: 209 – 229.
Moncada Ferrera, M. 1989. Reporte del genero Commiphora Jacq.
(Burseraceae) para Cuba. – Rev. Jard. Bot. Nacl. 10: 3 – : 10.
Morton, C. M. & Kallunki, J. A. 1993. Pollen morphology of the
subtribe Cuspariinae (Rutaceae). – Brittonia 45: 286 – 314.
Mziray, W. 1992. Taxonomic studies in Toddalieae Hook. f. (Rutaceae)
in Africa. – Symb. Bot. Upsal. 30, Uppsala Univ., Uppsala.
Pacltova, B. 1981. The evolution and distribution of Normapolles
pollen during the Cenophytic. – Rev. Palaeobot. Palynol. 35:
175 – 208.
Pflug, H. D. 1953. Zur Enstehung und Entwicklung des angios-
permiden Pollens in der Erdgeschichte. – Palaeontographica B
95: 60 – 171.
Pierlot, R. 1997. Pseudodacryodes Pierlot genre nouveau de
Burseraceae de l’est de la Rep. dem. du Congo. – Bull. Jard.
Bot. Natl Belg. 66: 175 – 186.
Punt, W., Blackmore, S., Nilsson, S. & Le Thomas, A. 1994.
Glossary of Pollen and Spore Terminology. – LPP Contr. Ser.
1. LPP Found., Utrecht. (updated – http://www.bio.uu.nl/
palaeo/glossary/).
Rasolodimby, J. F. C. 1983. Contribution palynologique a la
connaissance des Rutaceae de Madagascar. – These 3eme cycle.
131pp.
Rouse, G. E. 1957. The application of a new nomenclatural
298 M. M. Harley et al.
Grana 44 (2005)
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
10:
03 0
7 O
ctob
er 2
014

approach to Upper Cretaceous plant microfossils from western
Canada. – Can. J. Bot. 35: 349 – 375.
Rzedowski, J. & Palacios-Chavez, R. 1985. La presencia de
Commiphora (Burseraceae) en Mexico. – Taxon 34: 207 – 210.
Segaar, P. J. & van der Ham, R. W. J. M. 1993. Pollen of
Scutinanthe brunnea compared with other burseraceous pollen
types: a remarkable case of divergence. – Rev. Palaeobot.
Palynol. 79: 297 – 334.
Sims, H. J., Herendeen, P. S., Lupia, R., Christopher, R. A. &
Crane, P. R. 1999. Fossil flowers with Normapolles pollen from
the Upper Cretaceous of southeastern North America. – Rev.
Palaeobot. Palynol. 106: 131 – 151.
Soltis, D. E., Soltis, P. S., Chase, M. W., Mort, M. E., Albach,
D.C., Zanis, M., Savolainen, V., Hahn, W. H., Hoot, S. B., Fay,
M. F., Axtell, M., Swensen, S. M., Prince, L. M., Kress, W. J.,
Nixon, K. C. & Farris, J. A. 2000. Angiosperm phylogeny
inferred from 18S rDNA, rbcL, and atpB sequences. – Bot. J.
Linn. Soc. 133: 381 – 461.
Savolainen, V., Fay, M. F., Albach, D. C., Backlund, A., van der Bank,
M., Cameron, K. M., Johnson, S. A., Lledo, M. D., Pintaud, J-C.,
Powell, M., Sheahan, M. C., Soltis, D. E., Soltis, P. S., Weston, P.,
Whitten, W. N., Wurdack, K. J. & Chase, M. W. 2000. Phylogeny of
the eudicots: a nearly complete familial analysis based on rbcL gene
sequences. – Kew Bull. 55: 257 – 309.
Takhtajan, A. 1987. Systema Magnoliophytorum. – USSR Acad.
Sci. Press, Leningrad (St. Petersburg).
Thorne, R. F. 1983. Proposed new realignments in the angiosperms.
– Nord. J. Bot. 3: 85 – 117.
Van der Ham, R. W. J. M. 1995. Palynology. – In: Flora
Malesiana, Ser. 1. Spermatophyta. Vol. 12 P. 1. Meliaceae
(ed. D. J. Mabberley, C. M. Pannell & A. M. Sing), pp. 6 – 7.
– Found. Flora Males. Herb. Neerl. Natl. Bot. Gard., Leiden.
Victor, J. & Van Wyk, A. E. 1998. Palynology of Acmadenia
(Diosminae: Rutaceae) and its taxonomic implications. – Grana
37: 143 – 154.
Victor, J. & Van Wyk, A. E. 1999a. Palynology of Diosma and
Coleonema (Rutaceae: Diosminae) and its taxonomic implica-
tions. – Grana 38: 12 – 19.
Victor, J. & Van Wyk, A. E. 1999b. Palynology of Adenandra
(Rutaceae: Diosminae) and its taxonomic implications. – Grana
38: 1 – 11.
Victor, J. & Van Wyk, A. E. 2001. Palynology of Euchaetis and
Macrostylis (Diosminae - Rutaceae) and its taxonomic implica-
tions. – Grana 40: 105 – 110.
Young, D. A. 1982. Proposal. – In: Amended outlines and indices
for six recently published systems of angiosperm classification
(ed. H. G. Bedell & J. L. Reveal). – Phytologia 51:
65 – 156.
Pollen of Burseraceae 299
Grana 44 (2005)
Dow
nloa
ded
by [
Inge
nta
Con
tent
Dis
trib
utio
n (P
ublis
hing
Tec
hnol
ogy)
] at
10:
03 0
7 O
ctob
er 2
014





























