Embed Size (px)
Citation preview

Mitochondrion 12 (2012) 363–369
Contents lists available at SciVerse ScienceDirect
Mitochondrion
j ourna l homepage: www.e lsev ie r .com/ locate /mi to
Short communication
Preliminary evidences on mitochondrial injury and impaired oxidative metabolism inbreast cancer
Lorenza Putignani a,⁎, Salvatore Raffa b, Roberta Pescosolido c, Teresa Rizza d, Federica Del Chierico a,Laura Leone b, Laura Aimati b, Fabrizio Signore e, Rosalba Carrozzo d, Francesco Callea f,Maria Rosaria Torrisi b, Paola Grammatico c,1
a Parasitology Unit, Bambino Gesù Children's Hospital, IRCCS, Piazza Sant'Onofrio 4, 00165, Rome, Italyb Department of Clinical and Molecular Medicine, “Sapienza” University of Rome Sant'Andrea Hospital, Via di Grottarossa, 1035-00189, Rome, Italyc Medical Genetics, Molecular Medicine Department, “Sapienza” University of Rome, San Camillo-Forlanini Hospital, Circonvallazione Gianicolense 87, 00152, Rome, Italyd Unit of Molecular Medicine for Neuromuscular and Neurodegenerative Diseases, Bambino Gesù Children's Hospital and Research Institute, Piazza Sant'Onofrio 4, 00165, Rome, Italye Department of Obstetrics and Gynaecology, San Camillo-Forlanini Hospital, Circonvallazione Gianicolense 87, 00152, Rome, Italyf Department of Pathology, “Bambino Gesù” Children's Hospital and Research Institute, Piazza Sant'Onofrio 4, 00165, Rome, Italy
Abbreviations: CIEM, cryo-immunoelectron microscocomplex IV subunit IV; EM, electron microscopy; F1β ATinfiltrating ductal carcinoma; IEM, immuno electron micMAb203, monoclonal antibody against human complexbody against human complex IV subunit I; MAb408, moF1β; MAb14C10, monoclonal antibody against human GAtein 3; OMM, outer mitochondrial membrane; OXPHOS,complex II, subunit A flavoprotein (Fp); SDH-B, succinattegral membrane protein; SDH-D, succinate–dehydrogechrome c reductase complex III, subunit core protein 2;⁎ Corresponding author. Tel.: +39 06 68592598; fax:
E-mail addresses: [email protected] (L. Pu1 Tel.: +39 06 58704355; fax: +39 06 58704647.
1567-7249/$ – see front matter © 2012 Elsevier B.V. andoi:10.1016/j.mito.2012.02.003
a b s t r a c t
a r t i c l e i n f oArticle history:Received 18 May 2011Received in revised form 6 February 2012Accepted 13 February 2012Available online 18 February 2012
Keywords:MitochondriopathyBreast cancerMitochondrial damageOxidative phosphorylation
Mitochondriopathy is emerging as a new cancer theory; however, the relevance of mitochondrial pathobiologyin breast cancer has not yet been completely explored. Herein we report on altered expression levels of the ox-idative phosphorylation system (OXPHOS) subunits, mitochondrial structural injury and impaired ATP contentfrom a breast-infiltrating ductal carcinoma (IDC). With this purpose, a human mammary carcinoma (HMC-1)cell, referred to a humanmammary epithelial cell (HMEC) line, was assayed for: a) OXPHOS levels by quantita-tive cryo-immunoelectron microscopy (CIEM) labeling; b) morphological characterization by a newly intro-duced damage grading (scale Mt-g1–3), calculated on the % of intact cristae carrying mitochondria;c) bioenergetic impairment by luminometric determinations of cellular ATP content and cytochemical visualiza-tion of COX activity. Drastic OXPHOS reduction was observed in HMC-1 cells for the succinate-dehydrogenasecomplex II SDH-B protein, while decreasing was reported for the NADH–ubiquinone oxidoreductase complex INDUFS3 and the ubiquinol cytochrome c reductase complex III UQCRC2 subunits. A significant dropping wasdetected for the ATP-synthase complex V F1β protein. For the COX complex near-depletion of themitochondrial-encoded COXI and no apparent variation of the COXIV subunits were observed. Injury gradingwas categorized assigning three levels of morphological damage in HMC-1 mitochondria: i) severe (4.6%),ii) moderate (23.1%), iii) slight (44.6%), corresponding to 0%, 1–50% and 51–75% of area occupied by intact cris-tae. ATP generation andCOXactivity appeared significantly reduced inHMC-1 cells. The structural damage gradinghere described could provide new insight on IDC mitochondrial impairment and represent hallmark in the breastcancer mitochondriopathy.
© 2012 Elsevier B.V. and Mitochondria Research Society. All rights reserved.
py; COX, cytochrome c oxidase complex IV; COXI, cytochrome c oxidase complex IV subunit I; COXIV, cytochrome c oxidaseP, synthase complex V F1β subunit; HMC-1, human mammary carcinoma-1; HMEC, human mammary epithelial cells; IDC,roscopy; IMM, inner mitochondrial membrane; MAb112, monoclonal antibody against human complex I NDUFS3 subunit;II SDH-B subunit; MAb304, monoclonal antibody against human complex III UQCRC2 subunit; MAb404, monoclonal anti-noclonal antibody against human complex IV subunit IV; MAb503, monoclonal antibody against human complex V subunitPDH, Mt-g(1–3)-grading scale of mitochondrial damage; NDUFS3, NADH–ubiquinone oxidoreductase complex I Fe–S pro-oxidative phosphorylation system; ROS, reactive oxygen species; SD, standard deviation; SDH-A, succinate–dehydrogenasee–dehydrogenase complex II, subunit B iron–sulfur protein (Ip); SDH-C, succinate–dehydrogenase complex II, subunit C in-nase complex II, subunit D integral membrane protein; TEM, transmission electron microscopy; UQCRC2, ubiquinol–cyto-WB, western blot.+39 06 68592218.tignani), [email protected] (P. Grammatico).
d Mitochondria Research Society. All rights reserved.

364 L. Putignani et al. / Mitochondrion 12 (2012) 363–369
1. Introduction
The classical paradigm of mitochondrial structure–function inti-mately relates the two aspects of this organelle biology in a highly dy-namic way. Cellular events that alter mitochondrial bioenergeticinvolve internal changes and structural aberrations of the organellearchitecture compromising, in turn, the biochemical activities mainlyassociated with its internal membrane and matrix. For this reason, in-dices of structural change and impairment of the mitochondrial archi-tecture may reveal functional aberration and progressive cellularpathological shifts. The structure modification is mainly associatedwith loss of cristae and internal convolution, morphological alterationsof size and shape (Hackenbrock, 1966, 1968).
Therefore, it affects the functional activity, often associated withmodification of the terminal cytochrome c oxidase (COX, complexIV) of the oxidative phosphorylation system (OXPHOS) (Oka et al.,2008), change of the OXPHOS electron transfer and redox signalingby altering the reactive oxygen species (ROS) interaction, and decou-pling the ATP synthase complex V (Campello and Scorrano, 2010). Inhuman cells, in condition of severe mitochondrial damage, atrophiedmitochondria appear round-shaped and deprived of convoluted in-ternal membranes, more similar to “ancient” protist mitochondriarather than to typical Eukaryota mitochondria, similar to those de-scribed for the Barth or other syndromes (Acehan et al., 2006;Duong Van Huyen et al., 2006).
In the last decade, a huge work of research has provided new in-sights in the role of mitochondria in cancer cell (Brandon et al.,2006; Owens et al., 2011; Polyak et al., 1998; Singh, 1998) suggestingthat the compromised mitochondria bioenergetic function, combinedwith altered phenotype, is a hallmark of carcinogenesis (Isidoro et al.,2004). In the last few years, these results have been confirmed forbreast cancer (Owens et al., 2011), fully responding to Warburg'searly hypothesis according to which, during tumorigenesis, there isa significant reduction of OXPHOS and an increased ATP productionfrom cytosolic glycolysis (Isidoro et al., 2005; Warburg, 1929, 1956).However, despite decades of research based on this assumption, na-ture and origin of respiratory metabolic alterations in cancer cellsstill remain largely unknown.
In order to study the modified respiratory metabolism of breastcancer cells, cryo-immunoelectron microscopy (CIEM) was employedto assay the expression profiling of the OXPHOS I–V complexes in abreast cancer primary cell culture, which was compared with datapreviously described (Putignani et al., 2008). The altered mitochon-drial phenotype in cancer cells associated to impaired OXPHOS wasclassified by introducing new indexes of morphological degenera-tions. Altered cristae and reduced convolution of inner mitochondrialwere associated to low cellular ATP content.
2. Materials and methods
2.1. Cell line and culture conditions
The local Ethics Committee of S. Camillo-Forlanini Hospital (Rome,Italy) approved tissue collection and patient's consent. A tissue speci-men from a breast-infiltrating ductal carcinoma (IDC) was processedto generate a breast cancer primary cell culture (HumanMammary Car-cinoma, HMC-1), maintained in 20% RPMI 1640 medium (Euroclone,Milan, Italy), 20% heat-inactivated fetal calf serum, 2 mM L-glutamine(Euroclone), 100 μg/ml streptomycin and penicillin each (Eurobio,France) as previously described (Putignani et al., 2008). HMC-1 was re-ferred to a reference Human Mammary Epithelial Cell line (HMEC, Clo-netics, Walkersville, MD, USA), cultured in specific optimized, fullysupplementedmedium(Clonetics™MEGM), according tomanufacturerinstructions.
2.2. Ultrastructural evaluation of mitochondrial damage
Cell samples were fixed in 0.1 M phosphate buffer, 2% paraformal-dehyde, 0.2% glutaraldehyde for 2 h at 25 °C, washed and embeddedin 0.1 M phosphate buffer, 12% gelatin that was solidified in ice. Gelatinblocks were infused with 2.3 M sucrose ON at 4 °C, frozen in liquid ni-trogen, and cryosectioned with an Ultracut EMFCS cryoultramicrotome(Leica Microsystems, Wetzlar, Germany).
2.2.1. Definition of grading and IMM/OMM index of mitochondrial damageA grading scale for the evaluation of mitochondrial damage
(Cadwell et al., 2008) was generated and categorized on the basis ofthe percentage of mitochondrial area carrying intact cristae: i) 51–75%(Mt-g1 degree, slight damage); ii) 1–50% (Mt-g2 degree, moderatedamage); iii) 0% (Mt-g3 degree, severe damage). A total of 20 differentultrathin cryosections, randomly taken and recorded at 28,000× forboth HMC-1 and HMEC cell lines, were analyzed and all mitochondrialstructures were classified and graded. An additional parameter of themitochondrial injury was the convolution degeneration related to theinner mitochondrial membrane (IMM) length. All mitochondria ob-served in 30 ultrathin cryosections, randomly observed at 56,000× forboth HMC-1 and HMEC cell lines, were measured for both lengths ofIMM and outer mitochondrial membrane (OMM); hence, the IMM/OMM index was calculated for each mitochondrion. Mitochondrialdamagewas correlated to a lowmean value of IMM/OMM index, corre-sponding to partial or total loss of IMM cristae.
2.3. Cryo-immunoelectron microscopy (CIEM)
Ultrathin cryosections, obtained as described above, were collect-ed with sucrose and methyl cellulose and incubated with primarymouse monoclonal antibodies against OXPHOS subunits (NDUFS3,SDH-B, UQCRC2, COI, COIV and F1β) (Mitosciences, Eugene, OR,USA) diluted 1:50 in PBS-1% BSA, followed by either 10-nm- or 18-nm-diameter colloidal gold particles conjugated with protein A(Pharmacia, Uppsala, Sweden) diluted 1:10 in PBS-1% BSA. Finally,ultrathin cryosections were stained with a solution of 2% methyl cel-lulose and 0.4% uranyl acetate and examined under a Morgagni 268Dtransmission electron microscopy (FEI Company, Hillsboro, OR, USA).
2.3.1. Quantification of OXPHOS subunits by immunogold labelingTotal mitochondrial area, total colloidal gold particles and immuno-
labeling density (colloidal gold particles/μm2 mitochondrial area) wereanalyzed in at least 20 different ultrathin cryosections (28.000×) ran-domly taken (Lucocq, 1994). Specificity of gold immunolabeling wasaddressed by achieving as indicator of specific/background labelingratio in the HMEC reference line a value ranging from 7 to 100 times.
For each OXPHOS subunit, the normalized labeling density HMC-1/HMEC (%) was measured and correlated with previous densitomet-ric data of WB expression profiles (Putignani et al., 2008). For theimmunogold background evaluation, the density of colloidal goldparticles/μm2 extramitochondrial area was measured.
2.4. Imaging analyses and statistics
The digital images of ultrathin sections were recorded with aMegaView II charge-coupled device camera (Soft Imaging SystemGmbH, Münster, Germany) and analyzed with AnalySIS software.The analysis was performed by two observers in a blinded fashionto avoid operator-dependent bias. Results were expressed asmean±standard deviation (SD). Statistical analysis was based onChi-square and Student's t tests. p values b0.05 were considered toindicate the statistical significance. Through Pearson correlation coef-ficient (r), the regression analysis was used to test the relationship ofimmunogold labeling data with densitometric data of WB expressionprofiles of OXPHOS subunits.

365L. Putignani et al. / Mitochondrion 12 (2012) 363–369
2.5. Measurement of cellular ATP content
The content of cellular ATP was assayed luminometrically usingthe ATPLITE 1 STEP (Perkin Elmer, Boston, MA) according to the pro-cedure recommended by the manufacturer, using 1×104 cells foreach determination and expressed as counts per second (CPS)/cells.HMC-1 and HMEC cells were grown either in RPMI-based and specificMEGM regular media, respectively, as well as in galactose supple-mented media for 24 h. The experiments were carried out twiceboth in RM and GAL-supplemented media to assess inter-assay repro-ducibility. For either assays, each ATP content experiment was per-formed 3 times at the same moment to verify intra-assayreproducibility (Ferreira et al., 2011).
2.6. Cytochemical reaction to COX
COX activity was visualized cytochemically, as described elsewhere(Tiranti et al., 1998). Briefly, both HMEC and HMC-1 cells were plated
Mt-g1 Mt-g2 Mt-g3
HMC-1 44.6% 23.1% 4.6%
HMEC 13.9% 4.2% 1.4%
A
C
IMM/OMM TYPE
HMC-1 1.47±0.08 A,B,C
HMEC 2.38±0.31 D
200 nm
Intact cristae (%)
Fig. 1. Graphical representation of the morphological damage in HMC-1 and HMEC cells. Thintact cristae (Panels A, B) and convolution degree (Panel C). Blue and yellow contours reprconvolution in the mitochondria population, compared to normal mitochondria (D).
on sterile coverslips, air dried, and incubated for 15 min at RT in 10 mlof pre-incubation medium (Tris–HCl 50 mM pH 7.6, 10 g sucrose,28 mg cobalt chloride). The reactionwas performed in 10 ml of incuba-tion medium (10% sucrose in 0.1 M sodium phosphate pH 7.6, 10 mgcytochrome c, 10 mg 3,3′-Diaminobenzidine-tetrahydrochloride-dihydrate (DAB), 2.0 mg catalase, pH 7.4) at 37 °C for 5 h. Coverslipswere mounted on warm gelatine. COX activity was visualized as abrown DAB reaction product. Images were acquired with a micro-scope (ZEISS AXIOSKOP-HBO50) equipped with a 20× objectiveand a RT Color Spot camera system (Diagnostic Instrument) associatedwith the IAS 2000 acquisition software.
3. Results
3.1. Morphological damage of mitochondria
In order to describe the morphological alterations in the mito-chondrial population of HMC-1 cells, two parameters of structural
B
e injury was quantified on the basis of the percentage of mitochondrial area displayingesent OMM and IMM, respectively. Letters A, B and C represent the different degree of

366 L. Putignani et al. / Mitochondrion 12 (2012) 363–369
degeneration were considered: i) the percentage of mitochondrialarea carrying intact cristae; and ii) the loss of convolutions in the mi-tochondria indirectly assessed as the ratio IMM/OMM. Therefore, itwas introduced a “grading” nomenclature of themorphological damagecategorized as i) slight; ii) moderate; and iii) severe, according to the
Fig. 2. Expression profiles of I–V subunits of the OXPHOS system assayed by CIEM on HMClabeling for complex II, IV and V subunits in HMC-1 and HMEC cells, respectively (Panel(Panel B). Symbols *, ** refer to pb0.05 and b0.01, respectively (see Table 1).
three levels so-called Mt-g1–3. The Mt-g1 category was associated to44.6% and 13.9% (ratio 3.2) of the mitochondrial population of HMC-1and HMEC cells, respectively. The Mt-g2 category was observed in23.1% and 4.2% (ratio 5.5) of the mitochondrial population of theHMC-1 and HMEC cells, respectively. The Mt-g3 category was observed
-1 and HMEC intact cells. Selected CIEM images are reported for showing comparativeA). Histogram representation of immunolabeling density for each OXPHOS complex

Table 1Quantization of OXPHOS expression profiles by quantitative CIEM in mitochondria from HMC-1 and HMEC cells.
Mitochondrial labeling Extra-mitochondrial labeling
Specific immunogold Background immunogold
OXPHOS complexsubunit
Cellline
Mitochondrialarea (μm2)
Colloidalgold particles(N°)
Labelingdensity(colloidal goldparticles/μm2)
HMC-1/HMEC (%) Extra-mitochondrialarea (μm2)
Colloidalgoldparticles(N°)
Backgrounddensity(colloidalgoldparticles/μm2)
Labeling density WB-based densitometric analysisa
Complex I NDUFS3 HMC-1 3.32 3 0.90±0.42 48.09* 32.78 7.91 2 0.25±0.08HMEC 7.46 14 1.87±0.91 11.71 3 0.26±0.05
Complex II SDH-B HMC-1 4.34 27 6.21±3.37 30.39** 3.38 10.49 2 0.19±0.03HMEC 3.81 78 20.4±3.71 11.17 3 0.27±0.06
Complex III UQCRC2 HMC-1 7.55 18 2.34±0.84 60.26* 50.34 10.68 2 0.19±0.04HMEC 4.11 16 3.88±2.33 14.02 1 0.07±0.02
Complex IV COI HMC-1 5.10 7 1.37±0.49 18.27** 4.37 9.79 2 0.20±0.04HMEC 8.26 62 7.50±0.65 10.21 2 0.19±0.08
Complex IV COIV HMC-1 4.47 28 6.25±2.28 75.56° 97.26 9.25 2 0.21±0.08HMEC 3.62 30 8.27±1.71 11.35 2 0.17±0.02
Complex V F1β HMC-1 4.02 6 1.49±1.01 39.01** 18.07 8.17 1 0.12±0.04HMEC 5.75 22 3.82±0.76 8.61 2 0.23±0.04
p Value (by Student's t test): °not significant; *pb0.05, **pb0.01a Data from Putignani et al. (2008), Breast Cancer Res. Treat. 110, 439–452; Pearson correlation coefficient (r) +0.95; pb0.01.
367L. Putignani et al. / Mitochondrion 12 (2012) 363–369
in 4.6% and 1.4% (ratio 3.3) of the mitochondrial population of HMC-1and HMEC cells, respectively. The overall damage (Mt-g1,3) was ob-served in 72.3% of HMC-1 compared to 19.5% of HEMC cells, with atotal ratio of 3.7 (Fig. 1A, B). The observations were supported statisti-cally by a Chi-square test probability pb0.001. The inferred fourth cate-gory, corresponding to the absence of morphological damage andassociated with 76–100% range of intact cristae, was observed in27.7% of HMC-1 and 80.5% of HMEC cells.
The IMM/OMM index, associated with loss of convolutions insidethe mitochondria, was characterized by the values 1.47±0.08 and2.38±0.31 for HMC-1 cells and HMEC, respectively, demonstratingthe presence of a length of IMM rather large for HMEC cells and al-most halved for HMC-1 cells, as expected for mitochondria collapsedand deprived of internal invaginations. Observations were supportedby a pb0.001 of Student's t test (Fig. 1C).
0
100000
200000
300000
400000
500000
600000
700000
800000
900000
1000000
HMECHMC-1RM GALRM GAL
CP
S/1
x104
cells
Fig. 3. Cellular ATP content. Luminometric measurement of total ATP in cultured HMC-1 and HMEC cells growth either in regular medium (RM) or in medium supplementedfor 24 h with galactose (GAL).
3.2. Quantification of OXPHOS subunits immunogold labeling
The values of expression profiles of subunits I–V of the OXPHOSsystem, previously obtained by densitometry approach (Putignaniet al., 2008), were compared with the values obtained with acompletely different approach such as the CIEM. The NDUFS3, SDH-B, UQCRC2, COXI, COXIV, and F1β subunits of OXPHOS complexes I–V were mapped by gold particle immunolabeling within sectionsfrom HCM-1 whole cells (Fig. 2A). The number of particles was quan-tified and reported to the mitochondrial area. This procedure providedimmunolabeling density data for all OXPHOS complexes of HMC-1cells, which were compared to the reference values obtained forHMEC cells. By using this approach, the background noise was consid-ered as nonspecific labeling density of the non mitochondrial area andwas removed from the real signal in the mitochondrial marking. Afterbackground removal, the following values of the labeling density, nor-malized to the HMEC cells, were obtained: 48.09% with pb0.05 (com-plex I); 30.39% with pb0.01 (complex II); 60.26% with pb0.05(complex III); 18.27% with pb0.01; (complex IV COI); 75.56% with pnot significant (complex IV COIV); 39.01% with pb0.01 (complex V)(Fig. 2B, Table 1). The values showed full consistency with the data pre-viously obtained (Putignani et al., 2008) by quantitative densitometricanalysis of Western Blot profiles (Table 1). The two sets of valuesshowed a linear relationship characterized by a high Pearson correla-tion coefficient (r: +0.95) with a significant probability value pb0.01.
The lowest labeling valueswere observed for complexes II, IV andVpro-tein targets.
3.3. Measurement of cellular ATP content
HMC-1 and HEMC cells, were assayed for ATP content. The totalamount of ATP content in HMC-1 cells was lower (44%) than in theHEMC cells (Fig. 3). In order to disclose the overall metabolism ofthe HMC-1 cells and verify the integrity of OXPHOS system, cellswere treated with galactose for 24 h, an approach that force thecells to get ATP through the OXPHOS; also in this case, we detectedfor the HMC-1 cells an ATP content lower (47%) than in the HEMCcells, growth in similar conditions (Fig. 3). No significant differencein ATP content for HEMC after addition of galactose was detected(95.5%), while for HMC-1 cells the reduction was 20% (Fig. 3).
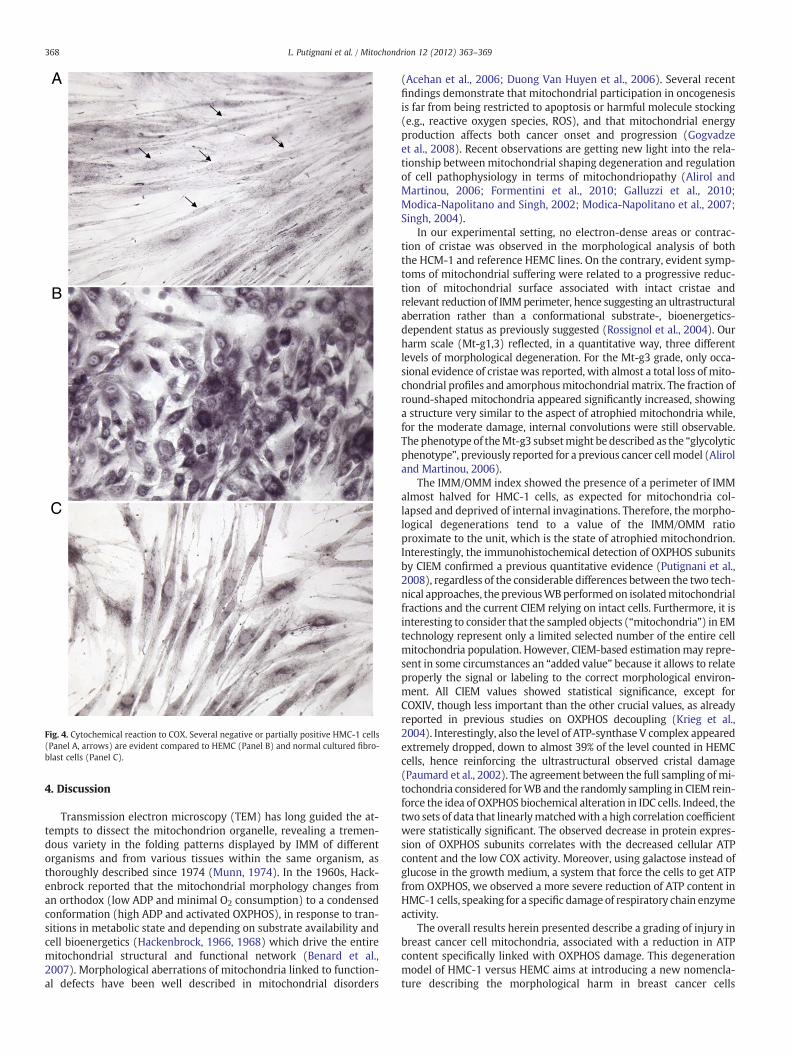
3.4. COX cytochemistry
Cytochemistry staining for COX in HMC-1 cells showed a verycomplicated scenario, with normal, partially positive or negativecells (Fig. 4A) compared with normal activity in HEMC (Fig. 4B) andnormal cultured fibroblasts (Fig. 4C).
A
B
C
Fig. 4. Cytochemical reaction to COX. Several negative or partially positive HMC-1 cells(Panel A, arrows) are evident compared to HEMC (Panel B) and normal cultured fibro-blast cells (Panel C).
368 L. Putignani et al. / Mitochondrion 12 (2012) 363–369
4. Discussion
Transmission electron microscopy (TEM) has long guided the at-tempts to dissect the mitochondrion organelle, revealing a tremen-dous variety in the folding patterns displayed by IMM of differentorganisms and from various tissues within the same organism, asthoroughly described since 1974 (Munn, 1974). In the 1960s, Hack-enbrock reported that the mitochondrial morphology changes froman orthodox (low ADP and minimal O2 consumption) to a condensedconformation (high ADP and activated OXPHOS), in response to tran-sitions in metabolic state and depending on substrate availability andcell bioenergetics (Hackenbrock, 1966, 1968) which drive the entiremitochondrial structural and functional network (Benard et al.,2007). Morphological aberrations of mitochondria linked to function-al defects have been well described in mitochondrial disorders
(Acehan et al., 2006; Duong Van Huyen et al., 2006). Several recentfindings demonstrate that mitochondrial participation in oncogenesisis far from being restricted to apoptosis or harmful molecule stocking(e.g., reactive oxygen species, ROS), and that mitochondrial energyproduction affects both cancer onset and progression (Gogvadzeet al., 2008). Recent observations are getting new light into the rela-tionship betweenmitochondrial shaping degeneration and regulationof cell pathophysiology in terms of mitochondriopathy (Alirol andMartinou, 2006; Formentini et al., 2010; Galluzzi et al., 2010;Modica-Napolitano and Singh, 2002; Modica-Napolitano et al., 2007;Singh, 2004).
In our experimental setting, no electron-dense areas or contrac-tion of cristae was observed in the morphological analysis of boththe HCM-1 and reference HEMC lines. On the contrary, evident symp-toms of mitochondrial suffering were related to a progressive reduc-tion of mitochondrial surface associated with intact cristae andrelevant reduction of IMMperimeter, hence suggesting an ultrastructuralaberration rather than a conformational substrate-, bioenergetics-dependent status as previously suggested (Rossignol et al., 2004). Ourharm scale (Mt-g1,3) reflected, in a quantitative way, three differentlevels of morphological degeneration. For the Mt-g3 grade, only occa-sional evidence of cristaewas reported, with almost a total loss of mito-chondrial profiles and amorphousmitochondrial matrix. The fraction ofround-shaped mitochondria appeared significantly increased, showinga structure very similar to the aspect of atrophied mitochondria while,for the moderate damage, internal convolutions were still observable.The phenotype of theMt-g3 subsetmight be described as the “glycolyticphenotype”, previously reported for a previous cancer cell model (Aliroland Martinou, 2006).
The IMM/OMM index showed the presence of a perimeter of IMMalmost halved for HMC-1 cells, as expected for mitochondria col-lapsed and deprived of internal invaginations. Therefore, the morpho-logical degenerations tend to a value of the IMM/OMM ratioproximate to the unit, which is the state of atrophied mitochondrion.Interestingly, the immunohistochemical detection of OXPHOS subunitsby CIEM confirmed a previous quantitative evidence (Putignani et al.,2008), regardless of the considerable differences between the two tech-nical approaches, the previousWBperformed on isolatedmitochondrialfractions and the current CIEM relying on intact cells. Furthermore, it isinteresting to consider that the sampled objects (“mitochondria”) in EMtechnology represent only a limited selected number of the entire cellmitochondria population. However, CIEM-based estimationmay repre-sent in some circumstances an “added value” because it allows to relateproperly the signal or labeling to the correct morphological environ-ment. All CIEM values showed statistical significance, except forCOXIV, though less important than the other crucial values, as alreadyreported in previous studies on OXPHOS decoupling (Krieg et al.,2004). Interestingly, also the level of ATP-synthase V complex appearedextremely dropped, down to almost 39% of the level counted in HEMCcells, hence reinforcing the ultrastructural observed cristal damage(Paumard et al., 2002). The agreement between the full sampling of mi-tochondria considered forWB and the randomly sampling in CIEM rein-force the idea of OXPHOS biochemical alteration in IDC cells. Indeed, thetwo sets of data that linearlymatchedwith a high correlation coefficientwere statistically significant. The observed decrease in protein expres-sion of OXPHOS subunits correlates with the decreased cellular ATPcontent and the low COX activity. Moreover, using galactose instead ofglucose in the growth medium, a system that force the cells to get ATPfrom OXPHOS, we observed a more severe reduction of ATP content inHMC-1 cells, speaking for a specific damage of respiratory chain enzymeactivity.
The overall results herein presented describe a grading of injury inbreast cancer cell mitochondria, associated with a reduction in ATPcontent specifically linked with OXPHOS damage. This degenerationmodel of HMC-1 versus HEMC aims at introducing a new nomencla-ture describing the morphological harm in breast cancer cells

369L. Putignani et al. / Mitochondrion 12 (2012) 363–369
depleted of fully working OXPHOS activity. The “model” could beexploited in pre-clinical investigations to gain new insights into mito-chondrial pathophysiology of breast cancer.
Acknowledgments
This studywas supported by theMinistero dell'Istruzione, Universitàe Ricerca, (MIUR); Associazione Italiana per la Ricerca sul Cancro (AIRC);Ricerca Corrente (RC) “Bambino Gesù” Children's Hospital, IRCCS; andFondazione Monte dei Paschi di Siena (MPS). The authors thankAndreina Santoro for her careful English revision of the manuscript.
References
Acehan, D., Xu, Y., Stokes, D.L., Schlame, M., 2006. Comparison of lymphoblast mitochondriafrom normal subjects and patients with Barth syndrome using electron microscopictomography. Lab. Invest. 87, 40–48.
Alirol, E., Martinou, J.C., 2006. Mitochondria and cancer: is there a morphologicalconnection? Oncogene 25, 4706–4716.
Benard, G., Bellance, N., James, D., Parrone, P., Fernandez, H., Letellier, T., Rossignol, R.,2007. Mitochondrial bioenergetics and structural network organization. J. Cell Sci.120, 838–848.
Brandon, M., Baldi, P., Wallace, D.C., 2006. Mitochondrial mutations in cancer. Oncogene25, 4647–4662.
Cadwell, K., Liu, J.Y., Brown, S.L., Miyoshi, H., Loh, J., Lennerz, J.K., Kishi, C., Kc, W.,Carrero, J.A., Hunt, S., Stone, C.D., Brunt, E.M., Xavier, R.J., Sleckman, B.P., Li, E.,Mizushima, N., Stappenbeck, T.S., Virgin, H.W., 2008. A key role for autophagyand the autophagy gene Atg16l1 in mouse and human intestinal Paneth cells. Nature456, 259–263.
Campello, S., Scorrano, L., 2010. Mitochondrial shape changes: orchestrating cellpathophysiology. EMBO Rep. 11, 678–684.
Duong Van Huyen, J.P., Batisse, D., Heudes, D., Belair, M.F., Piketty, C., Gonzalez-Canali, G.,Weiss, L., Kazatchkine, M.D., Bruneval, P., 2006. Alteration of cytochrome oxidase sub-unit I labeling is associated with severe mitochondriopathy in NRTI-related hepato-toxicity in HIV patients. Mod. Pathol. 19, 1277–1288.
Ferreira, M., Torraco, A., Rizza, T., Fattori, F., Meschini, M.C., Castana, C., Go, N.E., Nargang, F.E.,Duarte, M., Piemonte, F., Dionisi-Vici, C., Videira, A., Vilarinho, L., Santorelli, F.M.,Carrozzo, R., Bertini, E., 2011. Progressive cavitating leukoencephalopathy associatedwith respiratory chain complex I deficiency and a novel mutation in NDUFS1. Neuroge-netics 12, 9–17.
Formentini, L., Martínez-Reyes, I., Cuezva, J.M., 2010. The mitochondrial bioenergeticcapacity of carcinomas. IUBMB Life 62, 554–560.
Galluzzi, L., Morselli, E., Kepp, O., Vitale, I., Rigoni, A., Vacchelli, E., Michaud, M., Zischka,H., Castedo, M., Kroemer, G., 2010. Mitochondrial gateways to cancer. Mol. AspectsMed. 31, 1–20.
Gogvadze, V., Orrenius, S., Zhivotovsky, B., 2008. Mitochondria in cancer cells: what isso special about them? Trends Cell Biol. 18, 165–173.
Hackenbrock, C.R., 1966. Ultrastructural bases for metabolically linked mechanical activityin mitochondria. I. Reversible ultrastructural changes with change in metabolic steadystate in isolated liver mitochondria. J. Cell Biol. 30, 269–297.
Hackenbrock, C.R., 1968. Ultrastructural bases for metabolically linked mechanical activityin mitochondria. II. Electron transport-linked ultrastructural transformations in mito-chondria. J. Cell Biol. 37, 345–369.
Isidoro, A., Martínez, M., Fernández, P.L., Ortega, A.D., Santamaría, G., Chamorro, M.,Reed, J.C., Cuezva, J.M., 2004. Alteration of the bioenergetic phenotype of mitochondriais a hallmark of breast, gastric, lung and oesophageal cancer. Biochem. J. 378, 17–20.
Isidoro, A., Casado, E., Redondo, A., Acebo, P., Espinosa, E., Alonso, A.M., Cejas, P.,Hardisson, D., Fresno Vara, J.A., Belda-Iniesta, C., González-Barón, M., Cuezva, J.M.,2005. Breast carcinomas fulfill the Warburg hypothesis and provide metabolicmarkers of cancer prognosis. Carcinogenesis 26, 2095–2104.
Krieg, R.C., Knuechel, R., Schiffmann, E., Liotta, L.A., Petricoin III, E.F., Herrmann, P.C.,2004. Mitochondrial proteome: cancer-alteredmetabolism associatedwith cytochromec oxidase subunit level variation. Proteomics 4, 2789–2795.
Lucocq, J., 1994. Quantitation of gold labelling and antigens in immunolabelled ultrathinsections. J. Anat. 184, 1–13.
Modica-Napolitano, J.S., Singh, K.K., 2002. Mitochondria as targets for detection andtreatment of cancer. Expert Rev. Mol. Med. 11, 1–19.
Modica-Napolitano, J.S., Kulawiec, M., Singh, K.K., 2007. Mitochondria and human cancer.Curr. Mol. Med. 7, 121–131.
Munn, E.A., 1974. The Structure of Mitochondria. Academic Press, London.Oka, T., Sayano, T., Tamai, S., Yokota, S., Kato, H., Fujii, G., Mihara, K., 2008. Identification
of a novel proteinMICS1 that is involved inmaintenance of mitochondrial morphologyand apoptotic release of cytochrome c. Mol. Biol. Cell 19, 2597–2608.
Owens, K.M., Kulawiec, M., Desouki, M.M., Vanniarajan, A., Singh, K.K., 2011. ImpairedOXPHOS complex III in breast cancer. PLoS One 6, e23846.
Paumard, P., Vaillier, J., Coulary, B., Schaeffer, J., Soubannier, V., Mueller, D.M., Brèthes, D., diRago, J.P., Velours, J., 2002. The ATP synthase is involved in generating mitochondrialcristae morphology. EMBO J. 21, 221–230.
Polyak, K., Li, Y., Zhu, H., Lengauer, C., Willson, J.K., Markowitz, S.D., Trush, M.A., Kinzler,K.W., Vogelstein, B., 1998. Somatic mutations of the mitochondrial genome inhuman colorectal tumours. Nat. Genet. 20, 291–293.
Putignani, L., Raffa, S., Pescosolido, R., Aimati, L., Signore, F., Torrisi, M.R., Grammatico,P., 2008. Alteration of expression levels of the oxidative phosphorylation system(OXPHOS) in breast cancer cell mitochondria. Breast Cancer Res. Treat. 110,439–452.
Rossignol, R., Gilkerson, R., Aggeler, R., Yamagata, K., Remington, S.J., Capaldi, R.A.,2004. Energy substrate modulates mitochondrial structure and oxidative capacityin cancer cells. Cancer Res. 64, 985–993.
Singh, K.K., 1998. Mitochondrial DNA Mutations in Aging, Disease, and Cancer. Springer,New York.
Singh, K.K., 2004. Mitochondrial dysfunction is a common phenotype in aging and cancer.Ann. N. Y. Acad. Sci. 1019, 260–264.
Tiranti, V., Hoertnagel, K., Carrozzo, R., Galimberti, C., Munaro, M., Granatiero, M.,Zelante, L., Gasparini, P., Marzella, R., Rocchi, M., Bayona-Bafaluy, M.P., Enriquez,J.A., Uziel, G., Bertini, E., Dionisi-Vici, C., Franco, B., Meitinger, T., Zeviani, M.,1998. Mutations of SURF-1 in Leigh disease associated with cytochrome c oxidasedeficiency. Am. J. Hum. Genet. 63, 1609–1621.
Warburg, O., 1929. Ist die aerobe Glykolyse spezifish fur die Tumoren? Biochem. Z. 204,482–483.
Warburg, O., 1956. On the origin of cancer cells. Science 123, 309–314.





























