Embed Size (px)
Citation preview

Plant Physiol. (1976) 58, 710-716
Protein Bodies of Castor Bean EndospermISOLATION, FRACTIONATION, AND THE CHARACTERIZATION OF PROTEIN COMPONENTS1
Received for publication June 3, 1976 and in revised form August 24, 1976
RAYMOND E. TULLY AND HARRY BEEVERSDivision of Natural Sciences, University of California, Santa Cruz, California 95064
ABSTRACT
Protein bodies in the endosperm of castor bean seeds (Ricinus com-munis L.) contain phytin globoids and protein crystalloids embedded inan amorphous proteinaceous matrix. The protein bodies are apparendysurrounded by a single membrane. The protein bodies were isolated bygrinding and centrifuging in glycerol. Such isolated protein bodies werealmost identical (after cytological fixation) to those observed in situ,except that the globoids were lost. However, membrane-like structuresappear to have surrounded the globoids. Histochemical analysis of theisolated protein bodies showed that carbohydrates (glycoproteins) arelocalized only in the matrix region.
Addition of water to protein bodies in glycerol caused dissolution ofthe matrix, and release of the globoids and crystalloids. When thecrystalloids were centrifuged on sucrose density gradients, they wererecovered at an equilibrium density of 1.29 to 1.30 g/ml. The crystalloidswere only slighty soluble in most aqueous buffers but were very solublein sodium dodecyl sulfate, urea, or NaOH solutions.
Polyacrybamide gel electrophoresis in the presence of sodium dodecylsulfate and chromatography on ion exchange celiuloses show that theprotein bodies are composed of one major and several minor anodicproteins. The major protein, along with a few of the minor proteins, islocalized in the crystafloids.The major protein (molecular weight 65,000) was converted by mer-
captoethanol into subunits with molecular weights of 32,000 and15,800. It is proposed that the protein is made up of two of the smallersubunits and one of the lager, linked by disulfide bridges. None of thecrystaloid proteins appear to be glycosylated.The water-soluble matrix fraction is composed mainly of two proteins,
with molecular weights of 12,500 and 10,300 on the gels. Neither is aglycoprotein, and neither can be reduced with mercaptoethanol to givesubunits. The soluble fraction also contains other lesser componentsamong which are several glycoprotein lectins. One of these is ricin D,which is the main glycoprotein in the protein bodies.
sucrose density gradients (21), the protein bodies of many oil-seeds must be isolated nonaqueously, since they disintegrate inaqueous media (17). Various such procedures have been devel-oped. Protein bodies have been isolated from cottonseed (16),Cannabis (29), and peanut (6) by grinding in cottonseed oil;from squash (41) and cottonseed (43) by grinding in glycerol;and from Ricinus by grinding in acetone (33). Such isolatedprotein bodies have been fractionated on the basis of the differ-ential solubility of the inclusions. The crystalloids of Cannabiswere prepared by treating protein bodies isolated on cottonseedoil-CCl4 gradients with an aqueous NaCl/SDS solution, whichruptured the membranes and dissolved the matrix proteins (29).The crystalloids were then pelleted, and were dissolved in phos-phate buffer for further study of the proteins. The globoids andcrystalloids of Ricinus protein bodies were prepared by treatingthem with 70% glycerol, which dissolved the matrix proteins(37). The globoids and crystalloids were separated and purifiedby repeated suspensions and centrifugations in acetone-CC14mixtures of increasing densities.
Although much work has been done on the characterization ofthe globoids of Ricinus protein bodies (33, 36, 37) the matrixand crystalloid fractions have not been extensively examined(34). In this study, the protein bodies were isolated from theendosperm of ungerminated castor beans in order to comparethe ultrastructure of the isolated protein bodies with those insitu, and to separate and isolate the matrix and crystalloid frac-tions. The protein constituents of the matrix and crystalloidfractions were separated by column chromatography and gelelectrophoresis, and mol wt estimated by comparison withknown proteins.
Ricin, the highly toxic lectin of castor beans (9), is a potentinhibitor of protein synthesis by cytoplasmic ribosomes (19).Sequestration in the protein bodies, analogous to that of theglycoprotein lectin in the vicilin fraction of Phaseolus (7, 27)might explain why it is nontoxic in the endosperm tissue. Thispossibility was examined.
The reserve proteins of seeds have long been known to bestored in the protein bodies, or aleurone grains. Rost (28),summarizing previous studies on protein bodies, describes threestructural types: those consisting of amorphous protein withoutinclusions (e.g. those of peanut, soybean, and corn), those withphytin globoids in the protein matrix (cotton), and those withboth globoids and protein crystalloids (Cannabis, Cucurbita,Yucca). Rost (28) incorrectly placed Ricinus in the secondgroup; its protein bodies actually have both types of inclusions(24) (Fig. 1).Although the protein bodies of cereals can be isolated on
1 This work was supported by National Science Foundation GrantPCM 75-23566 to H. B., and by a National Science Foundation Gradu-ate Fellowship to R. T.
MATERIALS AND METHODS
Protein Body Isolation and Fractionation. The nonaqueousmethod of Yatsu and Jacks (43) was employed, using 130 mlglycerol/130 castor bean seeds (Ricinus communis var. Hale).The protein body pellet was washed by resuspending in glyceroland recentrifuging, and was stored at 2 C until needed. Thesoluble albumins (SOL fraction) were extracted by the additionof 10 ml of 5 mm tris-HCl buffer (pH 8.5) to the protein bodypellet. The insoluble crystalloids were sedimented, and werewashed in the same buffer (CRY fraction).
Electron Microscopy. Intact endosperm from dry seeds andprotein bodies in glycerol were fixed according to Lott et al. (14)by suspending them in a mixture of 5% glutaraldehyde in 90%glycerol for 2 hr at room temperature. The preparations werethen washed in 0.2 M cacodylate buffer at pH 7.4, and postfixed
710

RICINUS PROTEIN BODIES
in 0.1% OS04 or KMnO4. Protein body crystalloids in aqueoussolutions were fixed in Karnovsky's cacodylate-buffered glutar-aldehyde-formaldehyde at pH 7.4 for 2 hr (13). All material wasthen dehydrated in an acetone series and embedded in hardSpurr's resin (35). Silver-gold thin sections were cut on a SorvallPorter-Blum ultramicrotome with a DuPont diamond knife. Sec-tions were stained for 10 min in 7.5% uranyl magnesium ace-tate, followed by 2 min in 0.4% Reynold's lead citrate. Sectionswere viewed and photographed with an RCA EMU-3H trans-mission electron microscope. Thick sections (1-2 ,m) for lightmicroscopy were sectioned with a glass knife and viewed withphase contrast optics.
Periodic Add-Schiff (PAS)2 Staining of Protein Bodies. Themethod used was essentially that of Chayen et al. (3). Beforefixing in picrate-formalin, however, protein bodies isolated inglycerol were first transferred to 95% ethanol. Controls withoutperiodate treatment were also examined.
Electrophoresis. Polyacrylamide gels (0.5 x 8.5 cm) contain-ing 5.6% acrylamide and 0.1% SDS were prepared according toFairbanks et al. (8). The acrylamide monomer was purified byrecrystallizing from benzene; the methylene-bisacrylamide was
recrystallized from acetone.In initial experiments, samples were prepared by adding an
aliquot of protein solution to an equal volume of treatmentbuffer, which consisted of 2% SDS, 0.2 M tris-HCl, and 2 mmEDTA (pH 8); for the reduction of disulfide bonds, the final mixwas made 1.0% (v/v) in ME.2 The solution was held at 60 C for20 min, and was then dialyzed for several hours against 0.01 M
tris-HCl, 1 mm EDTA, and 0.1% SDS (pH 8). It was laterfound sufficient merely to dissolve the protein directly in thedialysis buffer (with or without ME), treat at 60 C, and electro-phorese immediately. The sample mixture was prepared by com-
bining 200 Al of sample-dialysis buffer, 2 drops of glycerol, 20Ml of 0.1% pyronin Y, and (if needed) 10 Ml of ME. The gelswere overlaid with 100 Ml of this solution, followed by thechamber buffer. Electrophoresis was performed at 4 to 6 mamp/gel in a Bio-Rad model 150 gel electrophoresis cell, equippedwith an ISCO model 490 power supply.
Gels were stained for protein with Coomassie blue accordingto Weber and Osborn (40). Gels were stained for about 30 minat 60 C, and were diffusion-destained in 7% acetic acid at 60 C.Glycoproteins were stained according to Segrest and Jackson(31), omitting periodate from control gels. The excess Schiffreagent was removed by treating the gels at 60 C in severalchanges of 0.1% Na-metabisulfite in 7% acetic acid until thewashings failed to turn pink upon the addition of formaldehyde(8).
Ion Exchange and Gel Chromatography. Crystalloids weredissolved in 5 mm Na-acetate, pH 5, containing 6 M urea. Somepreparations were warmed to 60 C. After centrifuging to clarify,the solution was passed through a CM-cellulose column (1.5 x27 cm) in the acetate-urea buffer. The column was washed with50 ml of buffer, and bound proteins were eluted with a 250-mllinear NaCl gradient (0-0.15 M) in acetate-urea, followed by 1 MNaCl. To prevent crystallization of the urea, chromatographywas performed at room temperature. Protein was estimated byfollowing the absorbance at 280 nm.The soluble matrix albumins extracted in tris buffer were
chromatographed on a DEAE-cellulose column (1.5 x 27 cm)buffered with 5 mm tris-HCl, pH 8.5. After washing the columnwith buffer, bound proteins were eluted with a 300 ml linearNaCl gradient (0-0.3 M) in the tris buffer. The chromatographywas performed at about 6 C. Protein peaks from the columnwere concentrated by ammonium sulfate precipitation and fur-ther fractionated on a Sephadex G-100 column (2.5 x 48 cm) in
Abbreviations: ME: mercaptoethanol; PAS: periodic acid-Schiff re-
agent; PBS: phosphate-buffered saline.
the tris buffer, calibrated with standard proteins.Molecular Weight Estimations. The following proteins of
known mol wt were used to calibrate the SDS gels and theSephadex G-100 column: BSA, 68,000; ovalbumin, 44,000;aldolase, 40,000; glyceraldehyde-3-phosphate dehydrogenase,35,000; chymotrypsin, 25,700; myoglobin, 17,200; and Cyt c,12,400.Afinity Chromatography of Lectins. The method of Nicolson
et al. (18) was followed, with slight modifications. Phosphate-buffered saline (PBS) extracts (or column fractions in tris pH8.5) were applied to a column of Sepharose 6B (2.5 x 9 cm).After washing the column with PBS, the lectins binding to theSepharose were eluted with PBS containing 0.1 or 0.2 M galac-tose.
RESULTS
ULTRASTRUCrURE
Ultrastructural observation of sections from the endosperm ofdry seeds shows protein bodies surrounded by numerous sphero-somes (Fig. 1), as observed by Ory et al. (22). The proteinbodies are 10 to 12 ,um in diameter, and contain crystalloids andgloboids as inclusions. Some crystalloids tend to be polygonal incross-section, as has been noted by others (2, 22, 25), althoughmany are irregular. The crystalloids are dense and brittle, andoften give rippled sections. Note also the shattering of thegloboid by sectioning, which is also commonly observed (11, 14,26). The globoid is situated in the center of a cavity, termed thesoft globoid by Lott et al. (14) (Fig. 1). This cavity appears hereto be empty (i.e. filled with embedding resin).
PROTEIN BODY ISOLATION AND FRACTIONATION
The protein bodies isolated in glycerol were only slightlycontaminated by an occasional cell wall fragment (Fig. 2). Ultra-structural examination of such protein bodies showed them to belike those in situ. The globoids, however, dissolved during fixa-tion. A membrane appears to have surrounded the globoid (Fig.3). However, since it lacks the fine structure of a membrane, itmay be some type of interface or boundary layer. These struc-tures appear identical to the so-called membrane circles of Sim-ola (32).
Isolated protein bodies gave a positive stain with PAS reagentin the matrix region only, with no staining in the crystalloids.Controls without periodate showed no staining. Thus, glycopro-teins appear to be localized in the matrix region.
If protein bodies in glycerol are treated with water or aqueousbuffers, the matrix proteins (17% of the total) dissolve, leavingbehind the insoluble crystalloids (Fig. 4). Such crystalloids arealways fractured, perhaps due to imbibitional stresses set up bythe sudden contact with water. Under the light microscope, suchfractured crystalloids appear lobed, as observed by Wiley (41) insquash crystalloids. Around the crystalloids in Figure 4, whichwere prepared by adding gutaraldehyde-formaldehyde fixativeto protein bodies in glycerol, is what appears to be the mem-brane which surrounded the intact protein body.
Isolated crystalloids are insoluble or very slightly soluble inwater or aqueous buffers (Table I) or Triton X-100 (data notshown). However, they are soluble in NaOH, urea; and SDSsolutions. Thus, all further experiments on crystalloid proteins.were done in the presence of 0.1 to 1.0% SDS, or 4 to 6 M urea,as solubilizing agent. The crystalloids were also soluble in con-centrated NaCl solutions.When isolated crystalloids were layered onto sucrose density
gradients and centrifuged to equilibrium, they formed a sharpband at a density of 1.29 to 1.30 g/ml (Fig. 5). This compareswith densities of 1.26 to 1.28 for Phaseolus aureus protein
711Plant Physiol. Vol. 58, 1976
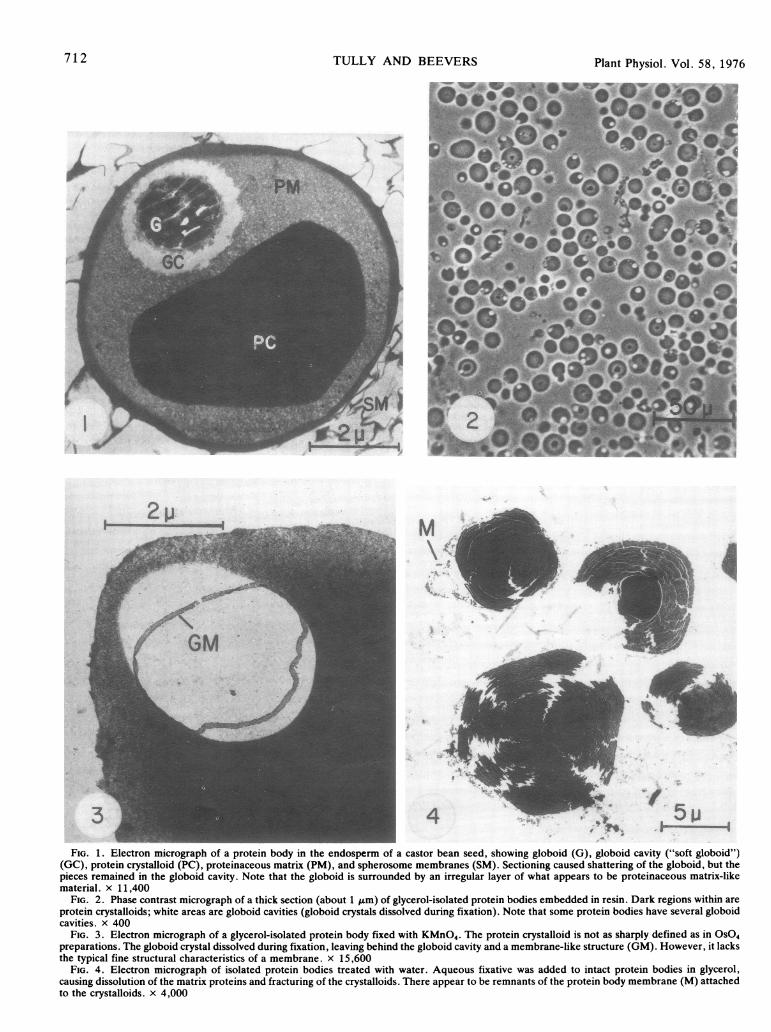
Plant Physiol. Vol. 58, 1976
MV :
3 4 - 5FIG. 1. Electron micrograph of a protein body in the endosperm of a castor bean seed, showing globoid (G), globoid cavity ("soft globoid")
(GC), protein crystalloid (PC), proteinaceous matrix (PM), and spherosome membranes (SM). Sectioning caused shattering of the globoid, but thepieces remained in the globoid cavity. Note that the globoid is surrounded by an irregular layer of what appears to be proteinaceous matrix-likematerial. x 11,400
FIG. 2. Phase contrast micrograph of a thick section (about 1 tum) of glycerol-isolated protein bodies embedded in resin. Dark regions within areprotein crystalloids; white areas are globoid cavities (globoid crystals dissolved during fixation). Note that some protein bodies have several globoidcavities. x 400
FIG. 3. Electron micrograph of a glycerol-isolated protein body fixed with KMnO4. The protein crystalloid is not as sharply defined as in OSO4preparations. The globoid crystal dissolved during fixation, leaving behind the globoid cavity and a membrane-like structure (GM). However, it lacksthe typical fine structural characteristics of a membrane. x 15,600
FIG. 4. Electron micrograph of isolated protein bodies treated with water. Aqueous fixative was added to intact protein bodies in glycerol,causing dissolution of the matrix proteins and fracturing of the crystalloids. There appear to be remnants of the protein body membrane (M) attachedto the crystalloids. x 4,000
712 TULLY AND BEEVERS
''a

Plant Physiol. Vol. 58, 1976 RICINUS PROTEIN BODIES
Table I: Solubility of Protein Body Crystalloids
Two ml of a crystalloid suspension containing about 14 pro-tein per ml (estimated in NaOH extract) were mixed with 2 mlof the solution to be tested. After removing the insolubleprotein by centrifuging, the rm-ining protein was precipitatedwith trichloroacetic acid, redissolved in NaOH, and assayedby the Lowry procedure.
Treatment mg/ml Dissolved
H20 0.1350.1 M Na-borate pH 9.5 0.830
n K-glycine " 0.850KK-EDTA " 0.535K-tris pH 8.0 0.425K-phosphate pH 7.0 0.485K-acetate pH 4.5 0.200
0.1 N HC1 1.020.01 N NaOH 13.70.1 N NaOH 14.00.1 M K-glycine +
4M urea pH 9.5 12.40.1 M Na-borate +
0.57 SDS pH 9.5 12.7
N
at
c0
0
5 10 20 25 30FRACTION
FIG. 5. Sucrose density gradient centrifugation of isolated crystal-loids. Crystalloids in 19 ml of 30% sucrose were layered on a lineargradient of 45 to 65% sucrose (40 ml). After centrifuging at 78,000g for2 hr, the gradient was fractionated. The first fraction consists of thepooled supernatant; remaining fractions are 1.2 ml each. Crystalloidsbanded at 1.29 to 1.30 g/ml.
cC W S
FIG. 6. SDS electropherograms of whole protein body (W), proteincrystalloid (C), and soluble matrix (S) fractions. Most of the protein inthe whole protein body preparation is in one heavy band, which islocalized in the crystalloids. Gel of soluble extracts was loaded so thatweak bands would show up. Arrow indicates direction of migration,which in all figures is downward (toward the anode).
bodies (7), 1.28 to 1.32 for soybean (39), and 1.26 for sun-flower (30). Suvorov et al. (37) found the density of Ricinuscrystalloids in acetone-CCl4 to be between 1.37 and 1.39.
SDS ELECTROPHORESIS
Figure 6 shows typical anodic SDS electropherograms of awhole protein body preparation (W), a crystalloid preparation(C), and the soluble matrix fraction (S). The whole protein bodypreparation yielded one major band, along with several minorones. The major band, and several of the others, are crystalloidproteins (gel C). The remaining proteins are found in the solublefraction (gel S).The apparent mol wt of the proteins on gels are shown in
Figure 7, A and C, numbering the bands as in Figure 7B. Themain crystalloid protein (CRY band 1) has a mol wt of about65,000; the remaining crystalloid proteins are smaller. The mainproteins in the soluble preparations are two low mol wt bands,SOL bands 6 and 7. There are also several bands with higher molwt.When a crystalloid preparation was treated with ME, bands 1
and 2 disappeared, and bands at one-half (CRY band 7) andone-quarter (CRY band 6) of their mol wt appeared or intensi-fied (Fig. 7B, CRY gels). CRY band 3 also disappeared,whereas 4 and 5 remained unchanged. Thus, CRY bands 6 and 7are subunits, which, in the untreated samples, are joined by S-Slinks to give the higher mol wt proteins.Some of the larger proteins from the soluble fraction were also
reduced by ME. The higher mol wt SOL bands 1 through 4yielded upon reduction bands 8 and 9 (band 8 being heteroge-neous). Reduction had little or no effect on SOL bands 5, 6, and7.PAS staining of gels of soluble preparations showed that SOL
bands 1 through 4 are glycoproteins. Their subunits, bands 8 and9, are also PAS-positive. SOL bands 6 and 7, which comprisemost of the soluble protein, showed no PAS staining. None ofthe crystalloid proteins were PAS-positive. These results confirm
ME
3 -
4 ,
- . . .w * ww _= w * *
C C_ < A ;x, _ _, _ _ , - , _c _, w _
A
ME
2~~~~~~~b^
7 57IS_ w
6 d6C
CRY SOL
FIG. 7. B: SDS electropherograms of crystalloid (CRY) and soluble(SOL) preparations in the presence and absence of ME, showing num-bering system used throughout text; A and C: apparent mol wt of thevarious soluble and crystalloid protein bands on the gels.
the histochemical finding that glycoprotein is confined to thematrix region of the protein bodies.
SEPARATION AND PURIFICATION OF THE PROTEIN COMPONENTS
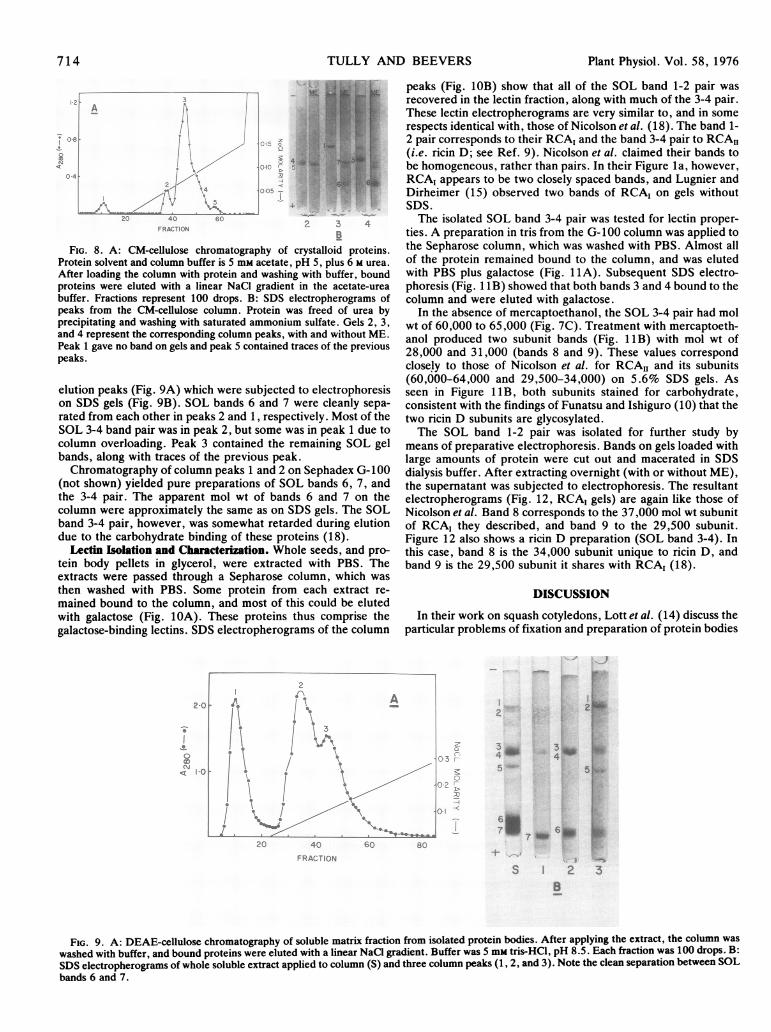
Crystalloid Components. The main crystalloid componentswere separated by chromatography on CM-cellulose (Fig. 8A),and were then electrophoresed on SDS gels with or without ME(Fig. 8B). The first column peak yielded no stainable bands ongels. The second peak was the CRY band 4-5 pair, which wasunaffected by ME. The third peak was an almost pure CRYband 1, which gave bands 6 and 7 upon ME treatment. Thefourth peak (shoulder) consisted largely of CRY band 3, whichwas converted to band 6 upon ME treatment. Peak 5 (gel notshown) consisted of minor components and traces of the otherpeaks.
Soluble Matrix Albumins. Chromatography of a water-solublematrix preparation on a DEAE-cellulose column yielded three
713
TULLY AND BEEVERS
4L
4 c
4~ ~ ~ ~~~~~~~~~A.C' 0N
... 4. |
FIG. 8. A: CM-cellulose chromatography of crystalloid proteins.Protein solvent and column buffer is 5 mm acetate, pH 5, plus 6 M urea.After loading the column with protein and washing with buffer, boundproteins were eluted with a linear NaCI gradient in the acetate-ureabuffer. Fractions represent 100 drops. B: SDS electropherograms ofpeaks from the CM-cellulose column. Protein was freed of urea byprecipitating and washing with saturated ammonium sulfate. Gels 2, 3,and 4 represent the corresponding column peaks, with and without ME.Peak 1 gave no band on gels and peak 5 contained traces of the previouspeaks.
elution peaks (Fig. 9A) which were subjected to electrophoresison SDS gels (Fig. 9B). SOL bands 6 and 7 were cleanly sepa-rated from each other in peaks 2 and 1, respectively. Most of theSOL 3-4 band pair was in peak 2, but some was in peak 1 due tocolumn overloading. Peak 3 contained the remaining SOL gelbands, along with traces of the previous peak.Chromatography of column peaks 1 and 2 on Sephadex G-100
(not shown) yielded pure preparations of SOL bands 6, 7, andthe 3-4 pair. The apparent mol wt of bands 6 and 7 on thecolumn were approximately the same as on SDS gels. The SOLband 3-4 pair, however, was somewhat retarded during elutiondue to the carbohydrate binding of these proteins (18).
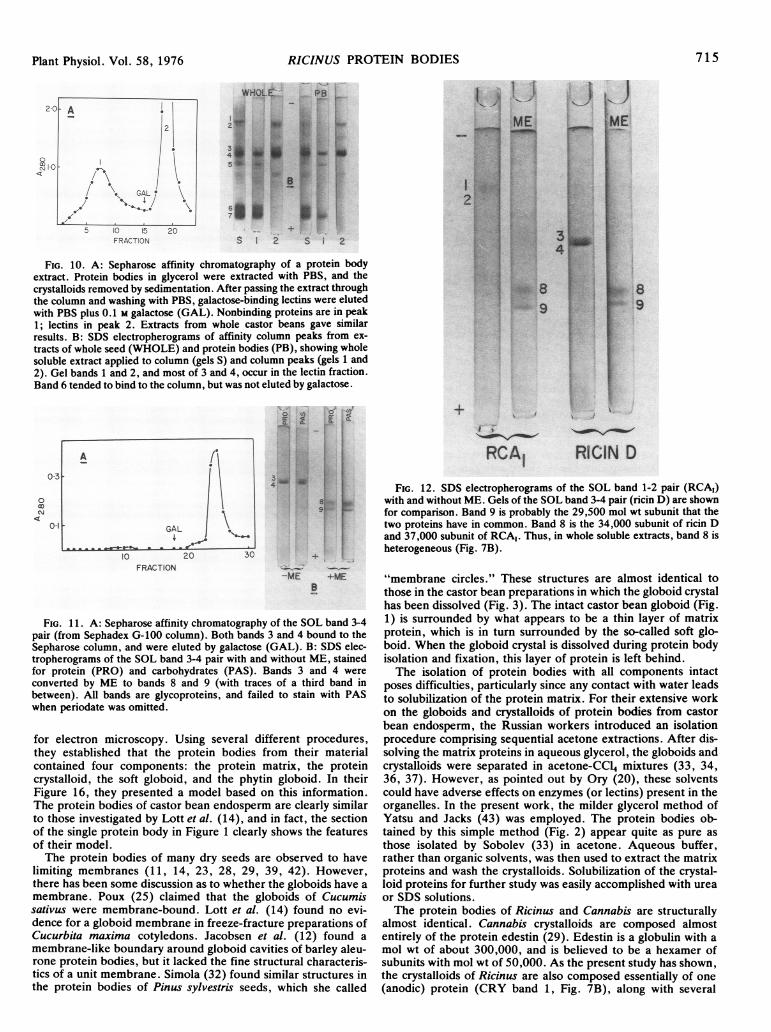
Lectin Isolation and Characterization. Whole seeds, and pro-tein body pellets in glycerol, were extracted with PBS. Theextracts were passed through a Sepharose column, which wasthen washed with PBS. Some protein from each extract re-mained bound to the column, and most of this could be elutedwith galactose (Fig. 10A). These proteins thus comprise thegalactose-binding lectins. SDS electropherograms of the column
IO * A2 I0
* 5' 5
e,
peaks (Fig. 10B) show that all of the SOL band 1-2 pair wasrecovered in the lectin fraction, along with much of the 3-4 pair.These lectin electropherograms are very similar to, and in somerespects identical with, those of Nicolson et al. (18). The band 1-2 pair corresponds to their RCA, and the band 3-4 pair to RCAII(i.e. ricin D; see Ref. 9). Nicolson et al. claimed their bands tobe homogeneous, rather than pairs. In their Figure la, however,RCA, appears to be two closely spaced bands, and Lugnier andDirheimer (15) observed two bands of RCA, on gels withoutSDS.The isolated SOL band 3-4 pair was tested for lectin proper-
ties. A preparation in tris from the G-100 column was applied tothe Sepharose column, which was washed with PBS. Almost allof the protein remained bound to the column, and was elutedwith PBS plus galactose (Fig. 1 A). Subsequent SDS electro-phoresis (Fig. 1 1B) showed that both bands 3 and 4 bound to thecolumn and were eluted with galactose.
In the absence of mercaptoethanol, the SOL 3-4 pair had molwt of 60,000 to 65,000 (Fig. 7C). Treatment with mercaptoeth-anol produced two subunit bands (Fig. 1 1B) with mol wt of28,000 and 31,000 (bands 8 and 9). These values correspondclosely to those of Nicolson et al. for RCA1I and its subunits(60,000-64,000 and 29,500-34,000) on 5.6% SDS gels. Asseen in Figure 1lB, both subunits stained for carbohydrate,consistent with the findings of Funatsu and Ishiguro (10) that thetwo ricin D subunits are glycosylated.The SOL band 1-2 pair was isolated for further study by
means of preparative electrophoresis. Bands on gels loaded withlarge amounts of protein were cut out and macerated in SDSdialysis buffer. After extracting overnight (with or without ME),the supematant was subjected to electrophoresis. The resultantelectropherograms (Fig. 12, RCA1 gels) are again like those ofNicolson et al. Band 8 corresponds to the 37,000 mol wt subunitof RCA1 they described, and band 9 to the 29,500 subunit.Figure 12 also shows a ricin D preparation (SOL band 3-4). Inthis case, band 8 is the 34,000 subunit unique to ricin D, andband 9 is the 29,500 subunit it shares with RCA1 (18).
DISCUSSION
In their work on squash cotyledons, Lott et al. (14) discuss theparticular problems of fixation and preparation of protein bodies
2 2
-- 3_4
.;
3 _4
5
5-
5. *
2C 4f.
FRAOT ON60 B
-t-_
S 2 3B
FIG. 9. A: DEAE-cellulose chromatography of soluble matrix fraction from isolated protein bodies. After applying the extract, the column waswashed with buffer, and bound proteins were eluted with a linear NaCl gradient. Buffer was 5 mM tris-HC1, pH 8.5. Each fraction was 100 drops. B:SDS electropherograms of whole soluble extract applied to column (S) and three column peaks (1, 2, and 3). Note the clean separation between SOLbands 6 and 7.
714 Plant Physiol. Vol. 58, 1976
C.
-.-A
; ....
17 6 owI'm 7 OM
RICINUS PROTEIN BODIES
5 10 15 20FRACTION
WHOL.C PB
si1 2 s 2
FIG. 10. A: Sepharose affinity chromatography of a protein bodyextract. Protein bodies in glycerol were extracted with PBS, and thecrystalloids removed by sedimentation. After passing the extract throughthe column and washing with PBS, galactose-binding lectins were elutedwith PBS plus 0.1 M galactose (GAL). Nonbinding proteins are in peak1; lectins in peak 2. Extracts from whole castor beans gave similarresults. B: SDS electropherograms of affinity column peaks from ex-tracts of whole seed (WHOLE) and protein bodies (PB), showing wholesoluble extract applied to column (gels S) and column peaks (gels 1 and2). Gel bands 1 and 2, and most of 3 and 4, occur in the lectin fraction.Band 6 tended to bind to the column, but was not eluted by galactose.
0-a
A
0-3
~~~~GALI
g.
10 20 30 +
FRACTION-ME -ME
FIG. 11. A: Sepharose affinity chromatography of the SOL band 3-4pair (from Sephadex G-100 column). Both bands 3 and 4 bound to theSepharose column, and were eluted by galactose (GAL). B: SDS elec-tropherograms of the SOL band 3-4 pair with and without ME, stainedfor protein (PRO) and carbohydrates (PAS). Bands 3 and 4 were
converted by ME to bands 8 and 9 (with traces of a third band inbetween). All bands are glycoproteins, and failed to stain with PASwhen periodate was omitted.
for electron microscopy. Using several different procedures,they established that the protein bodies from their materialcontained four components: the protein matrix, the proteincrystalloid, the soft globoid, and the phytin globoid. In theirFigure 16, they presented a model based on this information.The protein bodies of castor bean endosperm are clearly similarto those investigated by Lott et al. (14), and in fact, the sectionof the single protein body in Figure 1 clearly shows the featuresof their model.The protein bodies of many dry seeds are observed to have
limiting membranes (11, 14, 23, 28, 29, 39, 42). However,there has been some discussion as to whether the globoids have a
membrane. Poux (25) claimed that the globoids of Cucumissativus were membrane-bound. Lott et al. (14) found no evi-dence for a globoid membrane in freeze-fracture preparations ofCucurbita maxima cotyledons. Jacobsen et al. (12) found a
membrane-like boundary around globoid cavities of barley aleu-rone protein bodies, but it lacked the fine structural characteris-tics of a unit membrane. Simola (32) found similar structures inthe protein bodies of Pinus sylvestris seeds, which she called
I ;.
RCAI RICIN D
FIG. 12. SDS electropherograms of the SOL band 1-2 pair (RCAI)with and without ME. Gels of the SOL band 3-4 pair (ricin D) are shownfor comparison. Band 9 is probably the 29,500 mol wt subunit that thetwo proteins have in common. Band 8 is the 34,000 subunit of ricin Dand 37,000 subunit of RCA,. Thus, in whole soluble extracts, band 8 isheterogeneous (Fig. 7B).
"membrane circles." These structures are almost identical tothose in the castor bean preparations in which the globoid crystalhas been dissolved (Fig. 3). The intact castor bean globoid (Fig.1) is surrounded by what appears to be a thin layer of matrixprotein, which is in turn surrounded by the so-called soft glo-boid. When the globoid crystal is dissolved during protein bodyisolation and fixation, this layer of protein is left behind.The isolation of protein bodies with all components intact
poses difficulties, particularly since any contact with water leadsto solubilization of the protein matrix. For their extensive workon the globoids and crystalloids of protein bodies from castorbean endosperm, the Russian workers introduced an isolationprocedure comprising sequential acetone extractions. After dis-solving the matrix proteins in aqueous glycerol, the globoids andcrystalloids were separated in acetone-CC14 mixtures (33, 34,36, 37). However, as pointed out by Ory (20), these solventscould have adverse effects on enzymes (or lectins) present in theorganelles. In the present work, the milder glycerol method ofYatsu and Jacks (43) was employed. The protein bodies ob-tained by this simple method (Fig. 2) appear quite as pure as
those isolated by Sobolev (33) in acetone. Aqueous buffer,rather than organic solvents, was then used to extract the matrixproteins and wash the crystalloids. Solubilization of the crystal-loid proteins for further study was easily accomplished with ureaor SDS solutions.The protein bodies of Ricinus and Cannabis are structurally
almost identical. Cannabis crystalloids are composed almostentirely of the protein edestin (29). Edestin is a globulin with a
mol wt of about 300,000, and is believed to be a hexamer ofsubunits with mol wt of 50,000. As the present study has shown,the crystalloids of Ricinus are also composed essentially of one
(anodic) protein (CRY band 1, Fig. 7B), along with several
20 A_ ~~~I
d
. i
i II /.
x Jx
._ME
I .wME
s~
3.4
715Plant Physiol. Vol. 58, 1976

TULLY AND BEEVERS
minor components. The mol wt of this protein on SDS gels isabout 65,000. Treatment with mercaptoethanol (Fig. 8, gel 3)yields subunits with mol wt of 15,800 (band 6) and 32,000 (band7). The protein is presumably a band 6-6-7 combination, heldtogether by S-S bridges. As summarized by Derbyshire et al. (5)in their Table 1, many dicotyledenous seeds contain largeamounts of protein of mol wt 300,000 to 400,000. The tableshows that such a protein has been isolated from Ricinus com-
munis seeds, with a mol wt of about 332,000. If this protein isanalogous to edestin from Cannabis seeds, then perhaps it is a
hexamer of the crystalloid protein of mol wt 65,000.The water-soluble proteins of the protein bodies, which make
up th amorphous matrix, also present a simple picture (Fig.7B). Most of the (anodic) protein exists as two low mol wt
components, SOL bands 6 and 7. These proteins, along withband 5, yield no subunits upon mercaptoethanol treatment, andthey are apparently not glycosylated. The remaining solubleproteins, the band 1-2 and band 3-4 pairs, are all glycoproteins,and can be broken into subunits by reduction. The two bandswithin each pair are close together, and often appear as one bandon gels. The SOL band 1-2 and 3-4 pairs appear to be RCA, andRCAII, respectively, as defined by Nicolson et al. (18). RCAII,however, is a misnomer, since the pure protein has no agglutin-ating activity (9). In the present discussion, this fraction will becalled ricin D (9). That these gel bands are identical to those ofNicolson et al. (18) is verified by several pieces of evidence: theyhave about the same mol wt on SDS gels (Fig. 7C); they are
reduced by mercaptoethanol to subunits of mol wt comparableto those reported by Nicolson et al.; both band pairs bind toagarose, and subsequently can be eluted with galactose (Figs. 10and 11). Also, the band 3-4 pair was slightly retarded duringelution from a Sephadex column, giving it a lower apparent molwt than that exhibited on SDS gels, as also observed by Nicolsonetal.
It is thus clear that these two lectins are localized in the proteinbodies. As germination proceeds, the protein body membranesfuse, forming the tonoplast (1). The contents of the proteinbodies remain in this vacuole and are hydrolyzed during germi-nation. Thus, the toxic ricin is probably kept out of contact withribosomes in the cytoplasm.
It is noteworthy that the two lectins are the main glycoproteinsin the protein bodies and the crystalloid proteins show no PASstaining on gels. This is in contrast with protein bodies fromPhaseolus aureus, in which vicilin and legumin, the major stor-age proteins, are glycosylated (7). The five subunits of 7S soy-
bean globulin are also glycosylated (38).In the course of this study, we observed that a portion of the
protein in a soluble extract precipitates at 0 C and rapidly redis-solves on warming. The precipitate is heterogeneous, containingSOL bands 3, 4, 5, and 7. It was possible to separate some of theband 7 fraction cleanly from band 6 using this method. Otherseed storage proteins exhibit cryoprecipitability, as reviewed byDaussant et al. (4).
LITERATURE CITED
1. ASHTON, F. M. 1976. Mobilization of storage proteins of seeds. Annu. Rev. Plant Physiol.27: 95-117.
2. BuTrROSE, M.S. 1971. Ultrastructure of barley aleurone cells as shown by freeze-etching.Planta 96: 13-26.
3. CHAYEN, J., L. BrrENSKY, AND R. G. BUTCHER. 1973. Practical Histochemistry. JohnWiley & Sons, New York. pp. 72-73.
4. DAUSSANT, J., N.J. NEUCERE, AND L. Y. YATSU. 1969. Immunochemical studies on
Arachis hypogaea proteins with particular reference to the reserve proteins. I. Characteri-zation, distribution, and properties of a-arachin and a-conarachin. Plant Physiol. 44: 471-479.
5. DERBYSHIRE, E., D.J.WRIGHT, AND D. BOULTER. 1976. Legumin and vicilin, storageproteins of legume seeds. Phytochemistry 15: 3-24.
6. DIECKERT,J.W., J. E. SNOWDEN, A. T. MOORE, D. C. HEINGELMAN, AND A. M.
ALTSCHUL. 1962. Composition of some subcellular fractions from seeds of Arachishypogaea.J .Food. Sci. 27: 321-325.
7. ERICSON, M. C. AND M. J. CHRISPEELS. 1973. Isolation and characterization of glucosa-mine-containing storage glycoproteins from the cotyledons of Phaseolus aureus. PlantPhysiol. 52: 98-104.
8. FAIRBANKS, G., T. L. STECK, AND D. F. H. WALLACH. 1971. Electrophoretic analysis of themajor polypeptides of the human erythrocyte membrane. Biochemistry 10: 2606-2617.
9. FUNATSU, M. 1972. The structure and toxic function of ricin. In: M. Funatsu, K. Hiromi, K.Imahori, T. Murachi, and K. Narita, eds., Proteins: Structure and Function, Vol. 2. JohnWiley & Sons, New York. pp. 103-140.
10. FUNATSU, M. AND M. ISHIGURO. 1971. Chemical structure and toxicities of ricins. Proteinsisolated from Ricinus communis. Kaguku To Seibutsu 9: 490-497 (Chem. Abstr. 76:1 10323j).
11. HORNER, H. T. AND H. J. ARNOT-. 1965. A histochemical and ultrastructural study ofYucca seed proteins. Am. J. Bot. 52: 1027-1038.
12. JACOBSEN, J. V., R. B. KNOX, AND N. A. PYLioTns. 1971. The structure and composition ofaleurone grains in the barley aleurone layer. Planta 101: 189-209.
13. KARNOVSKY, M. J. 1965. A formaldehyde-glutaraldehyde fixative of high osmolarity for usein electron microscopy. J. Cell Biol. 27: 137-138A.
14. LoTr, J. N. A., P. L. LARSEN, AND J. J. DARLEY. 1971. Protein bodies from the cotyledonsof Cucurbita maxima. Can. J. Bot. 49: 1777-1782.
15. LUGNIER, A. AND G. DIRHEIMER. 1973. Differences between ricin and phytohemaggluti-nins from Ricinus communis seeds. FEBS Lett. 35: 117-120.
16. LuI, N. S. T. AND A. M. ALTSCHUL. 1967. Isolation of globoids from cottonseed aleuronegrains. Arch. Biochem. Biophys. 121: 678-684.
17. MIKoLA, J., L. Y. YATSU, T. J. JACKS, AND J. J. HEBERT. 1975. Disruption of certainaleurone grains by various homogenizing agents. Plant Cell Physiol. 16: 933-937.
18. NICOLSON, G. L., J. BLAUSTEI-., AND M. E. ETZLER. 1974. Characterization of two plantlectins from Ricinus communis and their quantitative interaction with a murine lymphoma.Biochemistry 13: 196-204.
19. OLSNES, S., K. REFSNES, AND A. PIHL. 1974. Mechanism of action of the toxic lectins abrinand ricin. Nature 249: 627-631.
20. ORY, R. L. 1972. Enzyme activities associated with protein bodies of seeds. In: G. E.Inglett, ed., Symposium: Seed Proteins. Avi Publishing Co.,Westport, Conn. pp. 86-98.
21. ORY, R. L. AND K. W. HENNINGSEN. 1969. Enzymes associated with protein bodiesisolated from ungerminated barley seeds. Plant Physiol. 44: 1488-1498.
22. ORY, R. L., L. Y. YATSU, AND H.W. KIRCHER. 1968. Association of lipase activity with thespherosomes of Ricinus communis. Arch. Biochem. Biophys. 1)3: 255-264.
23. PALEG, L. AND B. HYDE. 1964. Physiological effects of gibberellic acid. VII. Electronmicroscopy of barley aleurone cells. Plant Physiol. 39: 673-680.
24. PFEFFER,W. 1872. Untersuchungenuber die Proteinkorner und die Bedeutung des Aspara-gins beim Keimen der Samen. Jahrb.Wiss. Bot. 8: 429-571.
25. Poux, N. 1965. Localisation del'activite phosphatasique acide et des phosphates dans lesgrains d'aleurone. I. Grains d'aleurone renfermont Sla fois globoides et crystalloides. J.Microscopie 4: 771-782.
26. RICKSON, F. R. 1968. Anatomy, histochemistry and fine structure of the germinatingembryo of Paulowniatomentosa. Am. J. Bot. 55: 280-290.
27. ROMERO, J., S-M. M. SUN, R. C. MCLEESTER, F. A. BLISS, AND T. C. HALL. 1975.Heritable variation in a polypeptide subunit of the major storage protein of the bean,Phaseolus vulgaris L. Plant Physiol. 56: 776-779.
28. ROST, T. L. 1972. The ultrastructure and physiology of protein bodies and lipids fromhydrated dormant and nondormant embryos of Setaria lutescens (Gramineae). Am. J. Bot.59: 607-616.
29. ST. ANGELO, A. J., L. Y. YATSU, AND A. M. ALTSCHUL. 1968. Isolation of edestin fromaleurone grains of Cannabis sativa. Arch. Biochem. Biophys. 124: 199-205.
30. SCHNARRENBERGER, C., A. OESER, AND N. E. TOLBERT. 1972. Isolation of protein bodieson sucrose gradients. Planta 104: 185-194.
31. SEGREST, J. P. AND R. L. JACKSON. 1972. Molecular weight determination of glycoproteinsby polyacrylamide gel electrophoresis in sodium dodecyl sulfate. Methods Enzymol. 28:54-63.
32. SIMOLA, L. K. 1974. The ultrastructure of dry and germinating seeds of Pinus sylvestris L.Acta Bot. Fennica 103: 1-31.
33. SOBOLEV, A. M. 1966. On the state of phytin in the aleurone grains of mature andgerminating seeds. Soviet Plant Physiol. 13: 177-183.
34. SOBOLEV, A. M., V. I. SUVOROV, M. P. SAFONOVA, AND A. A. PROKOF'EV. 1972. Synthesisof storage proteins in endosperm of ripening castor seeds. Soviet Plant Physiol. 19: 894-899.
35. SPURR, A. R. 1969. A low-viscosity epoxy resin embedding medium for electron micros-copy. J. Ultrastruct. Res. 26: 31-43.
36. SUVOROV, V. 1. AND A. M.SOBOLEV. 1972. Cationic composition of aleurone grains ofseeds. Soviet Plant. Physiol. 19: 486-489.
37.SUVOROV, V. I., N. P. BUZULUKOVA, A. M. SOBOLEV, AND1. N.SVESHNIKOVA. 1970.Structure and chemical composition of globoids from aleurone grains of castor seeds.Soviet Plant Physiol. 17: 1020-1027.
38. THANH, V. H., K. OKUBO, AND K. SHIBASAKI. 1975. Isolation and characterization of themultiple 7S globulins of soybean proteins. Plant Physiol. 56: 19-22.
39.TOMBS, M. P. 1967. Protein bodies of the soybean. Plant Physiol. 42: 797-813.40.WERER, K. AND M. OSBORN. 1969. The reliability of molecular weight determinations by
dodecyl sulfate-polyacrylamide gel electrophoresis. J. Biol. Chem. 244: 4406-4412.41. WILEY, L. 1971. The fate of protein bodies in germinating seeds of Cucurbita maxima Duch.
Ph.D. dissertation. Univ. of Calif., Davis.42. YATSU, L. Y. 1965. The ultrastructure of cotyledonary tissue from Gossypium hirsutum L.
seeds. J. Cell Biol. 25: 193-199.43. YATSU, L. Y. AND T. J. JACKS. 1968. Association of lysosomal activity with aleurone grains
in plant tissue. Arch. Biochem. Biophys. 124: 466-471.
716 Plant Physiol. Vol. 58, 1976






![of [3H]PGF1x. - NCBI](https://img.pdfslide.net/doc/110x75/634d8ccba3f58bc38f088adc/of-3hpgf1x-ncbi.jpg)






















