Embed Size (px)
Citation preview

Pb
HJCa
b
c
d
e
f
g
h
i
j
k
l
a
ARRAA
KTPEEV
de
Uf
m
0d
Atherosclerosis 221 (2012) 341– 349
Contents lists available at ScienceDirect
Atherosclerosis
jo ur nal homep age : www.elsev ier .com/ locate /a therosc leros is
TEN overexpression attenuates angiogenic processes of endothelial cells bylockade of endothelin-1/endothelin B receptor signaling
siao-Mei Kuoa,b,1, Chun-Yao Linc,1, Hing-Chung Lamd, Pey-Ru Linb,e, Hoi-Hung Chana,f,ui-Cheng Tsengg, Cheuk-Kwan Sunh, Te-Fa Hsui, Chia-Ching Wuj, Chao-Yuh Yangk,hing-Mei Hsua,∗, Ming-Hong Taia,b,l,∗∗
Department of Biological Sciences, National Sun Yat-Sen University, 70 Lien-Hai Rd., Kaohsiung 804, TaiwanDepartment of Medical Education and Research, Kaohsiung Veterans General Hospital, Kaohsiung 813, TaiwanDivision of Cardiosurgery, Kaohsiung Veterans General Hospital, Kaohsiung 813, TaiwanDivision of Metabolism, Kaohsiung Veterans General Hospital, Kaohsiung 813, TaiwanInstitute of Basic Medicine, National Cheng Kung University, Tainan 701, TaiwanDivision of Gastroenterology, Kaohsiung Veterans General Hospital, Kaohsiung 813, TaiwanDivision of Allergy, Immunology, Kaohsiung Veterans General Hospital, Kaohsiung 813, TaiwanDivision of General Surgery, Department of Surgery, Chang Gung Memorial Hospital-Kaohsiung Medical Center, Kaohsiung, TaiwanDepartment of Cosmetic Applications & Management, Tung Fang Institute of Technology, Kaohsiung 829, TaiwanDepartment of Cell Biology and Anatomy, School of Medicine, National Cheng Kung University, Tainan 701, TaiwanDivision of Cardiology, Department of Medicine, China Medical University Hospital, Taichung 40447, TaiwanInstitute of Biomedical Sciences, National Sun Yat-Sen University, Kaohsiung 804, Taiwan
r t i c l e i n f o
rticle history:eceived 12 February 2010eceived in revised form 20 August 2010ccepted 20 August 2010vailable online 31 August 2010
eywords:hrombosisTENndothelin-1ndothelin B receptorascular endothelial growth factor
a b s t r a c t
Arteriovenous (AV) graft is frequently used as vascular access in hemodialysis patients. However, clot-ting or thrombosis of AV grafts often occurs and requires surgical removal. At present, the molecularpathogenesis underlying thrombosis of AV graft is not clear. The PTEN/Akt signaling has been impli-cated in the pathogenesis of vascular diseases. In this study, elevated PTEN expression and concomitantAkt inactivation was observed in endothelium of atherosclerotic brachial arteries from hemodialysispatients. To investigate whether PTEN upregulation affects endothelial function, adenovirus-mediatedPTEN (Ad-PTEN) overexpression was performed in aorta rings and cultured endothelial cells. It was foundthat PTEN overexpression potently inhibited the microvessel sprouting in aorta rings and the angiogenicactivities of endothelial cells including migration and tube formation. On the contrary, PTEN knockdownby RNA interference promoted the endothelial migration and reversed the Ad-PTEN-induced inhibition ofendothelial migration. Expression analysis showed that PTEN overexpression attenuated the expression
of endothelin-1 (ET-1) and endothelin B receptor (ETBR) in endothelial cells at transcriptional levels. How-ever, exogenous ET-1 supply only partially reversed the PTEN-induced inhibition of migration and tubeformation. This was delineated due to that PTEN overexpression also perturbed endothelial nitric oxidesynthase (eNOS) activation and vascular endothelial growth factor (VEGF) release. In summary, PTENupregulation induces endothelial dysfunction by attenuating the availability and signaling of multiplendot
angiogenic pathways in eAbbreviations: AV, arteriovenous; PTEN, phosphatase and tensin homologeleted on chromosome ten; ET-1, endothelin-1; ETBR, endothelin B receptor; eNOS,ndothelial nitric oxide synthase; VEGF, vascular endothelial growth factor.∗ Corresponding author. Tel.: +886 7 252000x3619; fax: +886 7 5253609.
∗∗ Corresponding author at: Institute of Biomedical Sciences, National Sun Yat-Senniversity, 70 Lien-Hai Rd., Kaohsiung 804, Taiwan. Tel.: +886 7 5252000x5816;
ax: +886 7 5250197.E-mail addresses: [email protected] (C.-M. Hsu),
[email protected] (M.-H. Tai).1 These authors contribute equally to this work.
021-9150/$ – see front matter © 2010 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.atherosclerosis.2010.08.067
helial cells, thereby may contribute to thrombosis of AV graft.© 2010 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
When native vasculature dose not permit a fistula, arteriove-nous bridge graft is often created in uremia patients by connectingbrachial artery and vein with a synthetic vessel for vascular access.The graft usually is made of a synthetic material such as poly-tetrafluoroethylene (PTFE) and ready for use within weeks afterformation. However, compared with AV fistula, AV bridge graft is
prone to failure due to atherosclerosis in the arterial anastomo-sis, which leads to decreased hemodialysis flow and even graft orbrachial artery thrombosis. At present, the molecular pathogenesisunderlying thrombosis or atherosclerosis of AV graft is not clear.
3 sclero
toa(gpiHeser(cep
Aopuroaip[phavwb
oAtwhco(ewtoflkwp
2
2
tft(ccC
42 H.-M. Kuo et al. / Athero
Endothelial dysfunction, an early marker of atherosclerosis,riggers the hemodynamic perturbations that lead to vari-us inflammatory and vascular diseases such as thrombosis,therosclerosis and diabetes. An imbalance of de novo endothelin-1ET-1) and nitric oxide (NO) synthesis is involved in the patho-enesis of endothelial dysfunction. ET-1, a potent vasoconstrictoreptide of 21-amino acids mainly secreted by endothelial cell, play
mportant role in vascular physiology and cardiovascular diseases.uman ET-1 is derived from a 212 amino acids precursor, prepro-ndothelin-1 (ppET-1), after the post-translational cleavage bypecific membrane-bound endopeptidases, endothelin-convertingnzymes. ET-1 acts via two main classes of G-protein-coupledeceptors: endothelin A receptor (ETAR) and endothelin B receptorETBR) [1]. ET-1 is a potent angiogenesis activator [2] and enhancesell proliferation by reducing apoptosis in human umbilical veinndothelial cells via the ETB receptor [3], which in turn induce cellroliferation and migration [4,5].
Phosphoinositide 3-kinases (PI3Ks) and its downstream effectorkt are activated by various stimuli and involved in the physiol-gy and diseases of the vascular system [6]. The PI3K/Akt pathwayarticipates in prototypical endothelial functions such as the reg-lation of vascular tone, angiogenesis, control of adhesion, andecruitment of leucocytes to the vessel wall. The positive effectf PI3K on angiogenesis is counteracted by PTEN (phosphatasend tensin homolog deleted on chromosome ten), which plays anmportant role in many cellular processes including tumor sup-ression, embryonic development, cell migration and apoptosis7]. Moreover, PTEN regulates various cellular processes in theathogenesis of vascular diseases including, cardiac hypertrophy,eart failure, preconditioning and hypertension [8,9]. PTEN acts as
negative regulator of angiogenesis both in vitro, where it inhibitsascular sprouting and VEGF-induced tube formation, and in vivo,here PTEN overexpression or administration of PI3K inhibitors
locks tumor angiogenesis.Endothelial cells, the primary cells responsible for production
f ET-1 and NO, play a pivotal important role in angiogenesis.ngiogenesis is a process by which new vessels are formed from
he pre-existing blood vessels via endothelial cells migration asell as proliferation [10]. It has been shown that ET-1 and NOas a direct angiogenic effect on endothelial and peri-vascularells as well as an indirect action through the increased releasef the potent angiogenic factor vascular endothelial growth factorVEGF) [11]. The relationship between PTEN and tumor angiogen-sis has been extensively studied. However, it remains unclearhether the cellular PTEN level modulates the physiological func-
ions of endothelial cells. In this study, we observed a unique profilef endothelial PTEN accumulation in the atherosclerotic arteriesrom hemodialysis patients. Subsequently, we employed variousaboratory approaches to elucidate how PTEN overexpression ornockdown affected endothelial function and the angiogenic path-ays in endothelial cells particularly the ET-1 and its receptorsathway.
. Materials and methods
.1. Cell cultures and reagents
For production and propagation of Ad5 adenovirus, E1a-ransformed human embryonic kidney 293 cells were purchasedrom Microbix Biosystems Inc. (Toronto, Canada) and maintainhem at low-passage. Human umbilical vein endothelial cells
HUVEC; passage 3–6) were isolated from umbilical veins andultured in M199 medium (Life Technologies, Gaithersburg, MD)ontaining 15% fetal calf serum, 20 U/ml porcine heparin (Sigmahemical Co.), and 100 �g/ml endothelial cell growth supplementsis 221 (2012) 341– 349
(Calbiochem; La Jolla, CA) as previously described [12]. Matrigelwas from BD PharMingen (La Jolla, CA). For RNA interference stud-ies, the control double-stranded, small interference RNA (siRNA)against PTEN and control siRNA were purchased from Santa CruzInc. (Santa Cruz; CA). ET-1 was from Peninsula Laboratories (Bel-mont, California).
2.2. Collection of explanted brachial arteries from hemodialysispatients and immunohistochemical analysis
The collection of surgical tissues during graft removal surgerywas approved by the Institutional Reviewer Boards of Kaohsi-ung Veterans General Hospital (Kaohsiung, Taiwan). Due to graftsthrombosis with the brachial artery anastomosis involvement,the brachial arteries were excised from six hemodialysis patientswithin 2 months after grafts implantation. The specimens weresegmented by 1 cm from the distal ends of the excised artery. Byvisual examination, the segments containing the atheroscleroticand non-atherosclerotic segments were used for subsequent analy-sis. Immunohistochemical analysis of PTEN expression in dissectedarteries was performed as previously described [13].
2.3. Adenoviral vectors
Recombinant adenovirus vectors encoding human PTEN cDNAwith hemagglutinin tag (Ad-PTEN) and green fluorescent protein(Ad-GFP) were generated as previously described [14]. The viruswas amplified by two rounds of cesium chloride ultracentrifugationand desalted by G-25 gel-filtration chromatography. After titter-ing by plaque-forming assay in 293 cells, the virus solution wasaliquoted and stored at −80 ◦C before use.
2.4. Rat aortic ring assay
This ex vivo angiogenesis assay was performed as previouslydescribed [15,16]. Briefly, the thoracic aortas were excised fromSprague–Dawley rats (male; 8-week-old) and immediately placedinto DMEM medium containing 10% FBS. The dissected aortas werecut into cross-sectional rings (1–1.5 mm in length) in a 24-wellplate and infected with adenovirus vectors (5 × 109 pfu per ring)for 1 h. After infection, the rings were placed into wells of a 24-wellplate coated with 0.5 ml of Matrigel in MCDB131 media and over-laid with 0.5 ml of MCDB131 media contain 5% fetal calf serum andbFGF (500 ng/ml; R&D, Minneapolis, MN). The rings were main-tained at 37 ◦C for 7–10 days and the vascular sprouting wasexamined daily using microscope equipped with digital images sys-tem (Olympus; Tokyo, Japan). The greatest distance from the aorticring body to the end of the vascular sprouts (sprout length) wasmeasured by NIH Image program at 3 distinct points per ring.
2.5. Quantitative reverse transcription-polymerase chainreaction (qRT-PCR)
RNA was isolated from endothelial cells using RNAzol (TEL-TEST,Inc., Friendswoods, TX). 5 �g of total RNA was used SuperscriptaseIII (Invitrogene; Carlsbad, CA), real-time PCR in Lightcycler (Roche)using a SYBR green I assay. PCR reaction was performed in SYBRGreen PCR Master Mix (Roche) following protocols provided bymanufacturer. The primer sequences for ET-1: forward primer 5′-CTTCTGCCACCTGGACATCA-3′, reverse primer 5′-GGCTTCCAAGT-CCATACGGA-3′. The primer sequences for ETAR: forward 5′-TC-TGATGACCTC GGTCCC-3′, reverse 5′-GTTCATGCTGTCCTTATGGCT-
3′. ETBR: forward 5′-AAT TACGATGGACTACAAAGGAAGTTA-3′, pri-mer 5′-GCAAGCAGAAATAGA AACTGAATAGC-3′. VEGF: forward5′-CCCTGATGAGATCGAGTACA-3′, reverse 5′-AGGAAGCTCATCTCT-CCTAT-3. Expression was normalized to �-actin: forward
sclero
5C
2
HicoaCaistGp
2
[1pAbtcws
2
aCei2
2
k[r1wp
2
cipm1acn(da(
H.-M. Kuo et al. / Athero
′-TCACCCACACTGTGCCCATC TACGA-3′, primer 5′-CAGCGGAA-CGCTC ATTGCCAATGG-3′.
.6. Migration assay
Migration assay was performed as previously described [12].UVEC were infected with adenovirus vectors for 24 h and seeded
n triplicate in the upper compartment of the chamber (1.2 × 105
ells in 400 �l). The lower compartment was filled with 200 �lf the DMEM media containing bFGF (100 ng/ml) as the chemo-ttractant. A polycarbonate filter (8 �m pore size; Nucleopore,ostar, Cambridge, MA) coated with 0.005% gelatin to allow celldhesion separated the compartments. After incubation for 2–4 hn a humidified 5% CO2 atmosphere at 37 ◦C, cells on the upperide of the filter were removed, and those that had migrated tohe lower side were fixed in absolute methanol, stained with 10%iemsa solution (Merck, Germany), and counted as a mean ± SEMer filter under five different high power fields (200×).
.7. Tube formation assay
Tube formation assay was performed as previously described12]. Briefly, Matrigel was diluted with cold serum-free medium to0 mg/ml. The diluted Matrigel solution were added into 24-welllate (200 �l per well) and allowed to form a gel at 37 ◦C for 30 min.fter infection for 24 h, HUVEC (1.5 × 105 cells per ml) were incu-ated with Matrigel for 6–8 h at 37 ◦C in 5% CO2. After incubation,he endothelial tubes were fixed with 3% paraformaldehyde andounted in four different lower power fields (100×). The tube ringas examined daily using microscope equipped with digital images
ystem (Olympus; 400×).
.8. Transfection with siRNA
HUVEC (in 6-well plate) were transfected with control siRNAnd PTEN siRNA at 20 �M using the lipofectamine 2000 (Invitrogen;arlsbad, CA) according to the manufacturer’s instructions. In somexperiments, HUVEC were transfected for 24 h then subsequentlynfected with adenovirus vectors (at a MOI of 200) for an additional4 h before harvest for further analysis.
.9. Enzyme-linked immunosorbent assay (ELISA)
ET-1 was extracted and measured using enzyme immunoassayit (R&D systems, Inc., Minneapolis, MN) as previously described17]. The intra-assay and inter-assay CVs were 4.5% and 5.5%,espectively. The minimum detectable dose is typically less than.0 pg/ml. The VEGF concentrations in cultured media of HUVECere measured by VEGF ELISA kit (R&D Systems Inc.) followingrotocols provided by manufacturer.
.10. Western blot analysis
After infection with adenovirus vectors for 48 h, endothelialells were harvest and extracted using lysis buffer (Cell Signal-ng Technology, Inc.; Danvers, MA). After separation in SDS-PAGE,rotein samples were transferred onto a polyvinylidene fluorideembrane (Millipore) using blotting apparatus. After blocking for
h, the membrane was incubated with primary antibodies for 1 ht room temperature. After incubation with secondary antibodyonjugated with HRP (1:5000 dilutions) for 30 min, the protein sig-als on membrane were detected using ECL-plus luminal solution
Pharmacia; Piscataway, NJ) and expose to X-ray film for autora-iogram. The primary antibodies used in this study were antibodiesgainst PTEN (1:1000 dilutions; Cell Signaling Technology, Inc.), HA1:1000 dilutions; Santa Cruz Inc.; Santa Cruz, CA), pAkt (Ser473sis 221 (2012) 341– 349 343
and Thr308; 1:1000 dilutions; Cell Signaling Technology, Inc.), Akt(1:1000 dilutions; Cell Signaling Technology, Inc.), ETBR (1:1000dilutions; Santa Cruz Inc.), ppET-1 (1:1000 dilutions, Peninsula Lab-oratories), ETAR (1:1000 dilutions; Santa Cruz Inc.), p-eNOS (1:1000dilutions; Cell Signaling Technology, Inc.), eNOS (1:1000 dilutions;Cell Signaling Technology, Inc.), VEGF (1:1000 dilutions; Santa CruzInc.), �-actin (1:5000 dilutions; Sigma)
2.11. Immunofluorescence and flow cytometry analysis
After infection with adenovirus vectors for 48 h, HUVEC werepermeablized using buffer containing 1.5% normal goat serum and0.2% Triton X-100 in PBS, incubated with ETBR antibody (1:100)for 2 h, then incubated with Alexa 488-conjugated secondary anti-body for 30 min at room temperature. After mounting in anti-Fademedia, the slides were visualized under a fluorescence microscope.The surface ETBR expression in HUVEC was determined by flowcytometry analysis. After infection with adenovirus vectors for 48 h,HUVEC were trypsinized and incubated with ETBR antibody (1:200)in Flow Cytometry Staining Buffer (eBioscience; San Diego, CA) at37 ◦C for 1 h. After wash with PBS twice, cells were incubated withrhodamine-conjugated secondary antibody (1:200) at 37 ◦C for 1 h,washed with PBS twice, and resuspended in PBS for analysis in aflow cytometer (BD Biosciences; San Jose, CA).
2.12. NO release assay
The NO release was determined by measuring the nitrite con-tents in the culture media as previously described [18]. Briefly, afterviral gene delivery, 100 �l cultured supernatants of HUVEC wereharvested and mixed with 100 �l Griess reagent (1 part 0.1% naph-thylethylenediamine dihydrochloride to 1 part 1% sulfanilamide in5% phosphoric acid). The absorbance at 550 nm was measured, andthe nitrite concentration was determined by interpolation of a cal-ibration curve of standard sodium nitrite concentrations againstabsorbance.
2.13. Nuclear factor kappa B (NF�B)-driven luciferase reporterassays
The NF�B activities in endothelial cells were investigated byluciferase activities assay as previously described [19]. HUVEC (in6-well plate) at 80% confluence were co-transfected with NF�B-driven luciferase vector and the renilla luciferase reporter vector(Promega; Madison, WI) at a ratio of 1: 1/10 using the lipofectamine2000 (Invitrogen; Carlsbad, CA) according to the manufacturer’sinstructions. After subsequent infection with adenovirus vectors for24 h, the NFkB-driven luciferase activities in cells were determinedusing a Dual-Light kit (Promega; Madison, WI) in luminometer(Microlumat Plus LB96 V; EG& G Berthold, Germany) and normal-ized with that of renilla luciferase.
2.14. Statistical analysis
The results were mean ± SD. Multiple comparison was per-formed by one-way analysis of variance in SPSS 10.0 for Windows(Chicago, IL). A P value less than 0.05 was considered significant.
3. Results
3.1. Elevated PTEN expression and reduced Akt phosphorylationin endothelium of atherosclerotic vessels
To investigate whether atherosclerosis is involved in thrombo-sis of AV bridge graft, the explanted brachial arteries were collectedfrom six hemodialysis patients for histological analysis, which
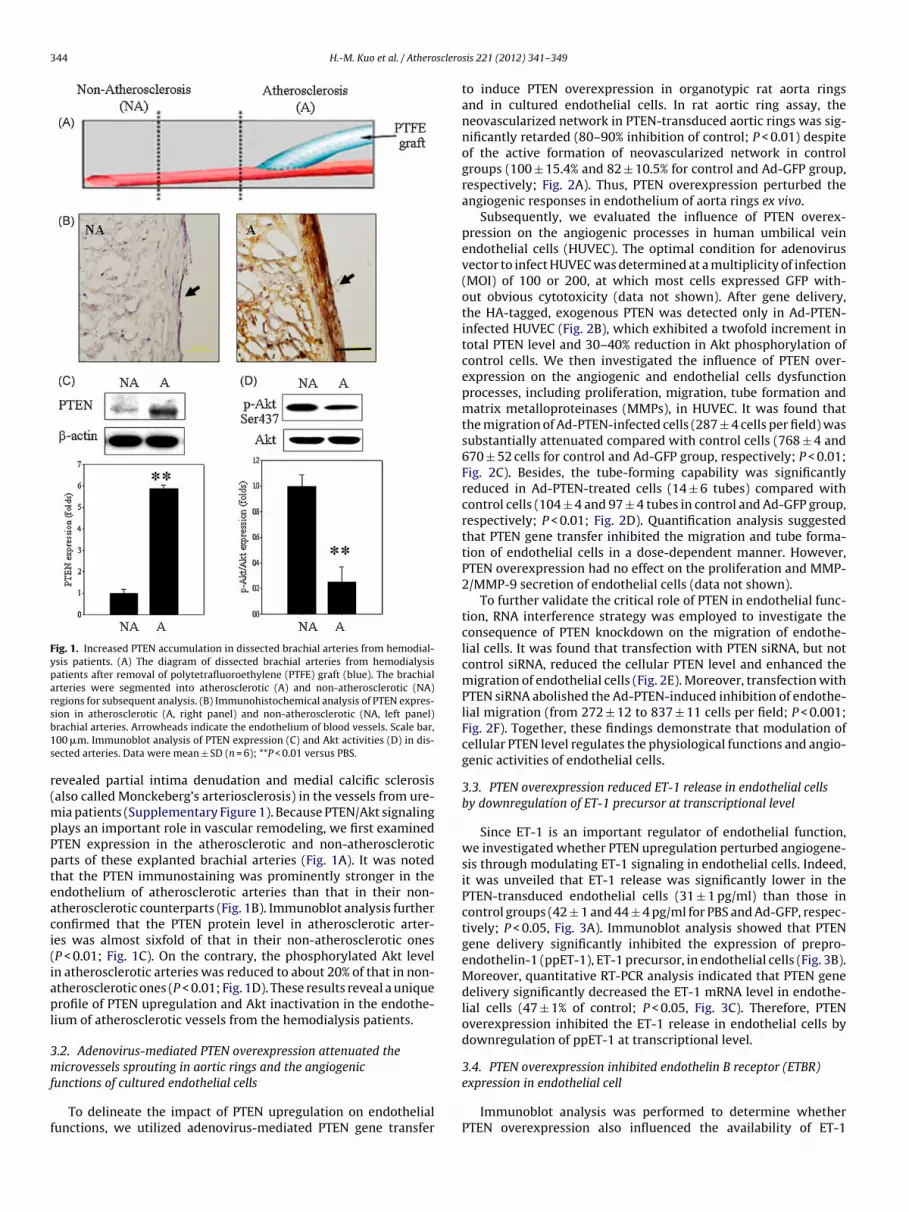
344 H.-M. Kuo et al. / Atherosclero
Fig. 1. Increased PTEN accumulation in dissected brachial arteries from hemodial-ysis patients. (A) The diagram of dissected brachial arteries from hemodialysispatients after removal of polytetrafluoroethylene (PTFE) graft (blue). The brachialarteries were segmented into atherosclerotic (A) and non-atherosclerotic (NA)regions for subsequent analysis. (B) Immunohistochemical analysis of PTEN expres-sb1s
r(mpPpteaci(iapl
3mf
f
ion in atherosclerotic (A, right panel) and non-atherosclerotic (NA, left panel)rachial arteries. Arrowheads indicate the endothelium of blood vessels. Scale bar,00 �m. Immunoblot analysis of PTEN expression (C) and Akt activities (D) in dis-ected arteries. Data were mean ± SD (n = 6); **P < 0.01 versus PBS.
evealed partial intima denudation and medial calcific sclerosisalso called Monckeberg’s arteriosclerosis) in the vessels from ure-
ia patients (Supplementary Figure 1). Because PTEN/Akt signalinglays an important role in vascular remodeling, we first examinedTEN expression in the atherosclerotic and non-atheroscleroticarts of these explanted brachial arteries (Fig. 1A). It was notedhat the PTEN immunostaining was prominently stronger in thendothelium of atherosclerotic arteries than that in their non-therosclerotic counterparts (Fig. 1B). Immunoblot analysis furtheronfirmed that the PTEN protein level in atherosclerotic arter-es was almost sixfold of that in their non-atherosclerotic onesP < 0.01; Fig. 1C). On the contrary, the phosphorylated Akt leveln atherosclerotic arteries was reduced to about 20% of that in non-therosclerotic ones (P < 0.01; Fig. 1D). These results reveal a uniquerofile of PTEN upregulation and Akt inactivation in the endothe-
ium of atherosclerotic vessels from the hemodialysis patients.
.2. Adenovirus-mediated PTEN overexpression attenuated theicrovessels sprouting in aortic rings and the angiogenic
unctions of cultured endothelial cells
To delineate the impact of PTEN upregulation on endothelialunctions, we utilized adenovirus-mediated PTEN gene transfer
sis 221 (2012) 341– 349
to induce PTEN overexpression in organotypic rat aorta ringsand in cultured endothelial cells. In rat aortic ring assay, theneovascularized network in PTEN-transduced aortic rings was sig-nificantly retarded (80–90% inhibition of control; P < 0.01) despiteof the active formation of neovascularized network in controlgroups (100 ± 15.4% and 82 ± 10.5% for control and Ad-GFP group,respectively; Fig. 2A). Thus, PTEN overexpression perturbed theangiogenic responses in endothelium of aorta rings ex vivo.
Subsequently, we evaluated the influence of PTEN overex-pression on the angiogenic processes in human umbilical veinendothelial cells (HUVEC). The optimal condition for adenovirusvector to infect HUVEC was determined at a multiplicity of infection(MOI) of 100 or 200, at which most cells expressed GFP with-out obvious cytotoxicity (data not shown). After gene delivery,the HA-tagged, exogenous PTEN was detected only in Ad-PTEN-infected HUVEC (Fig. 2B), which exhibited a twofold increment intotal PTEN level and 30–40% reduction in Akt phosphorylation ofcontrol cells. We then investigated the influence of PTEN over-expression on the angiogenic and endothelial cells dysfunctionprocesses, including proliferation, migration, tube formation andmatrix metalloproteinases (MMPs), in HUVEC. It was found thatthe migration of Ad-PTEN-infected cells (287 ± 4 cells per field) wassubstantially attenuated compared with control cells (768 ± 4 and670 ± 52 cells for control and Ad-GFP group, respectively; P < 0.01;Fig. 2C). Besides, the tube-forming capability was significantlyreduced in Ad-PTEN-treated cells (14 ± 6 tubes) compared withcontrol cells (104 ± 4 and 97 ± 4 tubes in control and Ad-GFP group,respectively; P < 0.01; Fig. 2D). Quantification analysis suggestedthat PTEN gene transfer inhibited the migration and tube forma-tion of endothelial cells in a dose-dependent manner. However,PTEN overexpression had no effect on the proliferation and MMP-2/MMP-9 secretion of endothelial cells (data not shown).
To further validate the critical role of PTEN in endothelial func-tion, RNA interference strategy was employed to investigate theconsequence of PTEN knockdown on the migration of endothe-lial cells. It was found that transfection with PTEN siRNA, but notcontrol siRNA, reduced the cellular PTEN level and enhanced themigration of endothelial cells (Fig. 2E). Moreover, transfection withPTEN siRNA abolished the Ad-PTEN-induced inhibition of endothe-lial migration (from 272 ± 12 to 837 ± 11 cells per field; P < 0.001;Fig. 2F). Together, these findings demonstrate that modulation ofcellular PTEN level regulates the physiological functions and angio-genic activities of endothelial cells.
3.3. PTEN overexpression reduced ET-1 release in endothelial cellsby downregulation of ET-1 precursor at transcriptional level
Since ET-1 is an important regulator of endothelial function,we investigated whether PTEN upregulation perturbed angiogene-sis through modulating ET-1 signaling in endothelial cells. Indeed,it was unveiled that ET-1 release was significantly lower in thePTEN-transduced endothelial cells (31 ± 1 pg/ml) than those incontrol groups (42 ± 1 and 44 ± 4 pg/ml for PBS and Ad-GFP, respec-tively; P < 0.05, Fig. 3A). Immunoblot analysis showed that PTENgene delivery significantly inhibited the expression of prepro-endothelin-1 (ppET-1), ET-1 precursor, in endothelial cells (Fig. 3B).Moreover, quantitative RT-PCR analysis indicated that PTEN genedelivery significantly decreased the ET-1 mRNA level in endothe-lial cells (47 ± 1% of control; P < 0.05, Fig. 3C). Therefore, PTENoverexpression inhibited the ET-1 release in endothelial cells bydownregulation of ppET-1 at transcriptional level.
3.4. PTEN overexpression inhibited endothelin B receptor (ETBR)
expression in endothelial cellImmunoblot analysis was performed to determine whetherPTEN overexpression also influenced the availability of ET-1

H.-M. Kuo et al. / Atherosclerosis 221 (2012) 341– 349 345
Fig. 2. PTEN gene delivery inhibited the angiogenic processes in aorta rings and in cultured endothelial cells. (A) Effect of PTEN gene transfer on the microvessel sprouting inaorta rings. After infection with various adenovirus vectors (1 × 109 pfu) for 7 days, the representative profile of microvessel outgrowth in different groups of aorta rings wasrecorded under light microscopy (top panel). Scale bar, 2 mm. The vessel length was measured by NIH Image program at three distinct points per ring and expressed mean ± SDpercentages of control (n = 6; bottom panel). **P < 0.01 versus control. (B) Effect of PTEN gene delivery on Akt activities in endothelial cells. After infection with recombinantadenovirus vectors (MOI 100 or 200) for 48 h, the cell extract of endothelial cells were collected for immunoblot analysis. The total PTEN protein level was evaluated byanti-PTEN (60 and 63 kDa, respectively) and the exogenous PTEN was detected using anti-HA (63 kDa). The Akt activity was determined by measuring anti-phospho-Akt(Ser473) and total Akt protein levels and expressed as fold over control. (C) Effect of PTEN overexpression on migration of endothelial cells. The representative photographs ofendothelial migration in different treatment groups were shown (top panel). Quantification analysis of migrated cells was performed by counting cell number from three highpower fields and expressed as mean ± SD of triplicates (bottom panel). (D) Effect of PTEN overexpression on tube formation of endothelial cells. The representative profilesthe tubular structures of HUVEC were recorded under light microscopy (top panel). Quantification analysis of tube formation were performed by counting the numberof rings and expressed as mean ± SD from quadruplicates (bottom panel). **P < 0.01 versus control. (E) Effect of PTEN siRNA on the cellular PTEN levels by immunoblotanalysis and migration in endothelial cells. In immunoblot analysis (top panel), HUVEC were transfected with siRNA for 24 h then harvested for western blot analysis usinganti-PTEN and normalized with that of �-actin level. For migration assay (bottom panel), HUVEC were transfected with siRNA for 24 h then subjected to migration assayusing Boyden chamber. Data were mean ± SD from quadruplicates. *P < 0.05 versus control groups. (F) Effect of PTEN siRNA on the Ad-PTEN-induced inhibition of endothelialmigration. HUVEC were transfected with siRNA for 24 h then infected with adenovirus vectors for another 24 h before harvested for migration assay. Data were mean ± SDfrom quadruplicates. **P < 0.001 versus control groups; #P < 0.001 versus Ad-PTEN group.

346 H.-M. Kuo et al. / Atherosclero
Fig. 3. PTEN overexpression attenuated ET-1 homeostasis in endothelial cells. (A)ELISA analysis of ET-1 protein release by endothelial cells after PTEN gene deliveryat MOI of 200 for 48 h. Data were mean ± SD of triplicates. (B) Western blot analysisof cellular ppET-1 expression in endothelial cells. After infection with recombinantadenovirus vectors (MOI 100 or 200) for 48 h, the expression of ET-1 precursor,prepro-endothelin-1 (ppET-1) in endothelial cells was evaluated by western blotanalysis and normalized with �-actin level. (C) qRT-PCR analysis of ET-1 mRNAlevel in endothelial cells after infection with recombinant adenovirus vectors (MOI200) for 48 h. The relative ET-1 mRNA level was determined by normalization witht*
re(i(atfEcgPlcgee
uremia patients. This seems consistent with previous studies that
hat of �-actin and expressed as mean ± SD percentages of control from triplicates.P < 0.05, **P < 0.01.
eceptors, the endothelin receptor A and B (ETAR and ETBR), inndothelial cells. Though ETAR expression was barely detecteddata not shown), ETBR protein level was significantly decreasedn Ad-PTEN-treated endothelial cells compared with controlsP < 0.05; Fig. 4A). Besides, immunofluorescence analysis revealed
significant reduction in ETBR immunostaining in the PTEN-ransduced endothelial cells (Fig. 4B). Flow cytometry analysisurther confirmed that Ad-PTEN-treated cells exhibited decreasedTBR cell surface expression (16.9 ± 2.6%) compared with that inontrol groups (41.7 ± 2.6 and 42.0 ± 3.2% for control and Ad-GFProup, respectively; P < 0.01; Fig. 4C). Finally, quantitative RT-CR analysis showed that the ETBR mRNA level was significantlyower in Ad-PTEN-infected endothelial cells (29 ± 1%) than that inontrol groups (100 ± 26% and 116 ± 12% for control and Ad-GFP
roup, respectively; P < 0.01; Fig. 4D). Therefore, PTEN gene deliv-ry inhibited the expression of ET-1 as well as its receptor ETBR inndothelial cells.sis 221 (2012) 341– 349
3.5. Exogenous ET-1 partially reversed the PTEN-inducedinhibition of migration and tube formation
Because the PTEN-induced reduction in ET-1 secretion wasmoderate (less than 30% of control groups), we examined whetherexcessive ET-1 supply could rescue the PTEN-mediated angiogen-esis perturbation. It was found that ET-1 supply only partiallyreversed the PTEN-induced inhibition of endothelial migration(from 287 ± 4 to 569 ± 19 cells per field; P < 0.001; SupplementaryFigure 2A) and tube formation (from 14 ± 6 to 29 ± 4 tubes per field;P < 0.05; Supplementary Figure 2B). Therefore, exogenous ET-1 sup-ply was not sufficient to reverse the PTEN-mediated angiogenesisinhibition, implicating that additional pathway(s) contributed toPTEN-mediated inhibition of endothelial functions.
3.6. PTEN overexpression perturbed eNOS and VEGF signaling inendothelial cells
Since Akt regulates eNOS and VEGF in endothelial cells, weexamined whether PTEN overexpression influenced eNOS activityand VEGF expression in endothelial cells. Immunoblot analy-sis showed that PTEN gene delivery effectively inhibited eNOSphosphorylation at Ser1175, which is essential to eNOS activity,without affecting its protein level (Fig. 5A). To confirm such find-ing, we also evaluated eNOS activity by determining the NO releasein endothelial cells after PTEN gene transfer. By measuring thenitrite level, it was found that the nitrite concentration in PTEN-transduced cells (13 ± 0.5 �M) was significantly lower than controlgroups (21 ± 0.4 and 22 ± 0.3 �M for control and Ad-GFP group,respectively; P < 0.01; Fig. 5B). Similarly, PTEN overexpression sig-nificantly attenuated VEGF secretion and decreased VEGF mRNAlevels in HUVEC (P < 0.05, Fig. 5C and D).
Because ET-1/ETBR and VEGF are transcriptionally regulatedby nuclear factor kappa B (NF-�B) pathway [20–22], we eval-uated the NF-�B activity in endothelial cells after PTEN genetransfer using luciferase reporter gene assay. It was shown thatNF-�B activity was significantly reduced in Ad-PTEN-infected cellscompared with controls (P < 0.05, Fig. 5E). Together, PTEN overex-pression elicited simultaneous blockade of multiple pro-angiogenicpathways, ultimately resulting in angiogenesis inhibition andendothelial dysfunction. Moreover, NF-�B inhibition participatedin PTEN-mediated inhibition of ET-1/ETBR and VEGF expression.
4. Discussion
The present study demonstrates for the first time that PTENaccumulation as well as Akt inactivation is found in the endothe-lium of thrombotic arteries from uremia patients. Adenoviral genetransfer studies indicate that PTEN overexpression potently dis-rupts the angiogenic functions of endothelial cells ex vivo andin vitro. Such perturbation of endothelial function may explain theendothelial dysfunction in endothelium with PTEN accumulation.We further delineated that PTEN overexpression inhibits the ET-1release as well as its receptor ETBR expression in endothelial cells,thereby perturbing the ET-1/NO homeostasis and angiogenic pro-cesses. Finally, we unveiled that PTEN upregulation simultaneouslyperturbs the expression of multiple angiogenic pathways, includ-ing ET-1, VEGF and NO, which may ultimately lead to endothelialdysfunction and thrombosis (as summarized in Fig. 5F).
In this study, we identified partial intima denudation as well asMonckeberg’s arteriosclerosis in the thrombotic arteries from the
Monckeberg’s arteriosclerosis is more common in patients withchronic kidney disease than in general population [23,24]. Moncke-berg’s arteriosclerosis, also called medial calcific sclerosis, is a form

H.-M. Kuo et al. / Atherosclerosis 221 (2012) 341– 349 347
Fig. 4. PTEN overexpression reduced endothelin receptor B (ETBR) expression in endothelial cells. (A) Western blot analysis of ETBR level after PTEN gene delivery. Afterinfection with adenovirus vectors for 48 h, the ETBR protein level in endothelial cells was detected by immunoblot analysis and normalized with that of �-actin. (B)Immunofluorescence analysis of ETBR expression after PTEN gene delivery. After infection with adenovirus vectors for 48 h, the ETBR expression in endothelial cells wereanalyzed by immunofluorescence staining using anti-ETBR followed by Alexa Fluor® 488-conjugated secondary antibodies (green). The cell nuclei were visualized using DAPIstaining (blue). Scale bar, 20 �m. (C) Flow cytometry analysis of surface ETBR expression after PTEN gene delivery. The cell surface ETBR expression in endothelial cells wasanalyzed by FACScan using anti-ETBR followed by Alexa Fluor® 488-conjugated secondary antibodies (red). (D) qRT-PCR analysis of ETBR mRNA level in endothelial cellsafter PTEN gene delivery. After infection with adenovirus vectors at MOI of 200 for 48 h, the ETBR mRNA level was determined by qRT-PCR analysis by normalization with� 1. (Fot
ooaIpcsmreup
vpawspiAtnu
nrimE
-actin and expressed as mean ± SD percentages of control from triplicates. **P < 0.0o the web version of the article.)
f arteriosclerosis where calcium deposits form in the middle layerf the walls of medium sized vessel sclerosis. The pathogenic mech-nism for Monckeberg’s arteriosclerosis is largely unknown [25].t is hypothesized that mechanical/chemical injuries during graftrocedure might elevate the cellular PTEN level in the endothelialells to induce endothelial dysfunction or to progress to apopto-is. Subsequently, the vascular smooth muscle cells proliferate andigrate at the endothelial cell denudation site, which consequently
esults in sclerosis. Future studies are warranted to delineate thetiological agent(s) and molecular mechanism underlying PTENpregulation during Monckeberg’s arteriosclerosis in hemodialysisatients.
PTEN upregulation in endothelial cells has been observed inarious cellular models of insulin resistance [26,27]. For exam-le, PTEN upregulation was delineated to participate in free fattycids-induced inhibition of insulin signaling in endothelial cells,hich would likely contribute to the pathogenesis of metabolic
yndrome [27]. Besides, resistin, an insulin resistance-inducingeptide, has been reported to induce PTEN expression by activat-
ng stress signaling p38 pathway and target transcription factorTF-2 expression, thereby inhibiting insulin signaling and leading
o cardiovascular diseases [26]. However, the pathogenic mecha-ism underlying PTEN upregulation in the thrombotic arteries fromremia patients remain to be elucidated.
The present study also shed lights on the therapeutic mecha-ism for local PTEN gene therapy to alleviate the angiogenesis-
elated diseases [14,28]. For example, Huang et al. employedntraarterial delivery of Ad-PTEN to suppress the neointima for-ation in rats after balloon injury [28]. Given the pivotal role ofT-1 and VEGF in restenosis, the simultaneous blockade of ET-1
r interpretation of the references to color in this figure legend, the reader is referred
and VEGF signaling pathways may explain the potent efficacy ofPTEN gene therapy. Likewise, it has been shown that intraarticularPTEN gene transfer ameliorates the collagen-induced rheumatoidarthritis in rats through inhibition of VEGF and interleukin-1 [14].Because ET-1 is increased in patients with rheumatoid arthritis[29–31], the perturbed ET-1 release provides further evidence sup-porting the potential of local PTEN gene transfer for managementof rheumatoid arthritis.
PTEN is an important tumor suppressor gene and well knownfor its potent inhibition of tumor angiogenesis. However, the func-tion of PTEN on physiological processes of vascular endothelial cellshas not been characterized yet. In this study, PTEN gene deliveryattenuated the migration and tube formation of cultured endothe-lial cells in vitro and reduced the microvessel sprouting of the rataorta rings in vivo. On the contrary, PTEN knockdown by RNA inter-ference promotes the migration of endothelial cells and possiblythe angiogenic activities. This observation of PTEN downregulationstimulates angiogenesis is consistent with the enhanced postnatalneovascularization in transgenic mice with endothelial cell-specificPTEN mutation [32,33]. Together, our data strongly support theimportant role of PTEN in regulating the angiogenic processes evenin non-transformed endothelial cells.
One novel finding of this study is that PTEN negatively regulatesangiogenesis via attenuating the ET-1/ETBR signaling pathway.The reduction in ET-1 secretion and precursor ppET-1 mightdirectly contribute to the PTEN-mediated angiogenesis inhibition
in endothelial cells. Indeed, ET-1 supplementation partially revertthe attenuated migration and tube formation by PTEN gene trans-fer, supporting that altered ET-1/NO homeostasis is involved inthe anti-angiogenesis function of PTEN. This finding is also in
348 H.-M. Kuo et al. / Atherosclerosis 221 (2012) 341– 349
Fig. 5. PTEN overexpression perturbed endothelial nitric oxide synthase (eNOS) phosphorylation and vascular endothelial growth factors (VEGF) expression in endothelialcells. (A) Western blot analysis of phosphorylated eNOS (p-eNOS) expression after PTEN gene delivery. After infection with adenovirus vectors for 48 h, the p-eNOS level inendothelial cells was detected by immunoblot analysis and normalized with that of eNOS level. (B) Measurement of NO release in endothelial cells after PTEN gene delivery.After infection with adenovirus vectors for 48 h, the nitrite concentration in cultured media of endothelial cells was measuring by Griess reagent. Data were mean ± SD oftriplicates. *P < 0.05, versus control groups. (C) ELISA analysis of VEGF release in endothelial cells after PTEN gene delivery. After infection with adenovirus vectors for 48 h, theVEGF release in endothelial cells was measuring by ELISA. Data were mean ± SD of triplicates. *P < 0.05, versus control groups. (D) qRT-PCR analysis of VEGF mRNA level afterPTEN gene delivery. After infection with recombinant adenovirus vectors at MOI of 200 for 48 h, the relative VEGF mRNA level in endothelial cells was determined by qRT-PCRa contrN enovia EN-m
atElaEt�bhrEHrPt1
ot
nalysis and normalized with that of �-actin. Data were mean ± SD percentage ofF�B activity in endothelial cells after PTEN gene delivery. After infection with adnalysis and normalized with that of renilla level. (F) The hypothetical scheme for PT
greement with the essential role of ET-1 in chemo-attractant andhe formation of capillaries networks [34]. Given that both ET-1 andTBR transcripts were decreased in the PTEN-transduced endothe-ial cells, it seems probable that PTEN gene transfer regulated ET-1nd ETBR expression at transcriptional levels. Because ET-1 andTBR are regulated by NF-�B pathway [20–22], it seems plausiblehat PTEN overexpression leads to Akt inactivation and reduced NF-B activities. In addition to NF-�B pathway, ET-1 can be regulatedy hypoxia and possibly by its downstream transcription factor,ypoxia-inducible factor 1� (HIF-1�). Because PTEN is known toegulate the stability of HIF-1�, it is likely that PTEN might reduceT-1 expression through inhibition of HIF-1� signaling pathway.owever, we cannot rule out the involvement of post-translational
egulation of ET-1 release by endothelin-converting enzyme inTEN-transduced endothelial cells. Future studies are warrantedo delineate the detailed mechanism underlying PTEN-induced ET-
/ETBR downregulation.In conclusion, the present study demonstrates the involvementf endothelial PTEN accumulation and its correlation with Akt inac-ivation in atherosclerotic arteries from patients underwent graft
ol from triplicates. *P < 0.05, versus control groups. (E) NF�B-luciferase analysis ofrus vectors for 48 h, the NF�B level in endothelial cells was detected by luciferaseediated inhibition of ET-1/ETBR, VEGF, NF�B and NO pathways in endothelial cells.
removal. Besides, we further shed lights on how PTEN upregulationinduces angiogenesis inhibition and possibly endothelial dysfunc-tion. Finally, we show for the first time that PTEN overexpressionattenuated ET-1/ETBR signaling in endothelial cells. Future stud-ies are warranted to elucidate the mechanism through which PTENwas up-regulated in endothelium of atherosclerotic arteries.
Acknowledgments
This work was supported by grants from National ScienceCouncil, Taiwan (NSC-99-2321-B-110 -005 and NSC-100-2325-B-110-002), Kaohsiung Veterans General Hospital (VGHKS98-044 toJYL and VGHKS97G-12 to HCL), Kaohsiung Chang Gung MemorialHospital (CMRPG880531), and National Sun Yat Sen University-Kaohsiung Medical University Joint Research Center (Kaohsiung,Taiwan).
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.atherosclerosis.2010.08.067.

sclero
R
[[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
H.-M. Kuo et al. / Athero
eferences
[1] Davenport AP, Battistini B. Classification of endothelin receptors and antago-nists in clinical development. Clin Sci (Lond) 2002;103(Suppl. 48):1S–3S.
[2] Salani D, Taraboletti G, Rosano L, et al. Endothelin-1 induces an angiogenicphenotype in cultured endothelial cells and stimulates neovascularization invivo. Am J Pathol 2000;157:1703–11.
[3] Dong F, Zhang X, Wold LE, Ren Q, Zhang Z, Ren J. Endothelin-1 enhances oxida-tive stress, cell proliferation and reduces apoptosis in human umbilical veinendothelial cells: role of ETB receptor, NADPH oxidase and caveolin-1. Br JPharmacol 2005;145:323–33.
[4] Noiri E, Hu Y, Bahou WF, Keese CR, Giaever I, Goligorsky MS. Permissive role ofnitric oxide in endothelin-induced migration of endothelial cells. J Biol Chem1997;272:1747–52.
[5] Wren AD, Hiley CR, Fan TP. Endothelin-3 mediated proliferation in woundedhuman umbilical vein endothelial cells. Biochem Biophys Res Commun1993;196:369–75.
[6] Morello F, Perino A, Hirsch E. Phosphoinositide 3-kinase signalling in the vas-cular system. Cardiovasc Res 2008.
[7] Li J, Yen C, Liaw D, et al. PTEN, a putative protein tyrosine phos-phatase gene mutated in human brain, breast, and prostate cancer. Science1997;275:1943–7.
[8] Maehama T, Dixon JE. PTEN: a tumour suppressor that functions as a phospho-lipid phosphatase. Trends Cell Biol 1999;9:125–8.
[9] Oudit GY, Penninger JM. Cardiac regulation by phosphoinositide 3-kinases andPTEN. Cardiovasc Res 2009;82:250–60.
10] Folkman J, Shing Y. Angiogenesis. J Biol Chem 1992;267:10931–4.11] Knowles J, Loizidou M, Taylor I. Endothelin-1 and angiogenesis in cancer. Curr
Vasc Pharmacol 2005;3:309–14.12] Tai MH, Kuo SM, Liang HT, et al. Modulation of angiogenic processes in cultured
endothelial cells by low density lipoproteins subfractions from patients withfamilial hypercholesterolemia. Atherosclerosis 2006;186:448–57.
13] Hu TH, Huang CC, Lin PR, et al. Expression and prognostic role of tumorsuppressor gene PTEN/MMAC1/TEP1 in hepatocellular carcinoma. Cancer2003;97:1929–40.
14] Wang CR, Shiau AL, Chen SY, et al. Amelioration of collagen-induced arthri-tis in rats by adenovirus-mediated PTEN gene transfer. Arthritis Rheum2008;58:1650–6.
15] Srivastava K, Kundumani-Sridharan V, Zhang B, Bajpai AK, Rao GN.15(S)-hydroxyeicosatetraenoic acid-induced angiogenesis requires STAT3-dependent expression of VEGF. Cancer Res 2007;67:4328–36.
16] Bee YS, Sheu SJ, Ma YL, et al. Topical application of recombinant calreticulinpeptide, vasostatin 48, alleviates laser-induced choroidal neovascularizationin rats. Mol Vis 2010;16:756–67.
17] Lam HC, Chu CH, Wei MC, et al. The effects of different doses of atorvastatin on
plasma endothelin-1 levels in type 2 diabetic patients with dyslipidemia. ExpBiol Med (Maywood) 2006;231:1010–5.18] Ikeda U, Kanbe T, Kawahara Y, Yokoyama M, Shimada K. Adrenomedullinaugments inducible nitric oxide synthase expression in cytokine-stimulatedcardiac myocytes. Circulation 1996;94:2560–5.
[
[
sis 221 (2012) 341– 349 349
19] Liu GS, Liu LF, Lin CJ, et al. Gene transfer of pro-opiomelanocortin prohormonesuppressed the growth and metastasis of melanoma: involvement of alpha-melanocyte-stimulating hormone-mediated inhibition of the nuclear factorkappaB/cyclooxygenase-2 pathway. Mol Pharmacol 2006;69:440–51.
20] Ohkita M, Takaoka M, Sugii M, Shiota Y, Nojiri R, Matsumura Y. The role ofnuclear factor-kappa B in the regulation of endothelin-1 production by nitricoxide. Eur J Pharmacol 2003;472:159–64.
21] Sugii M, Ohkita M, Taniguchi M, et al. Xanthoangelol D isolated from the rootsof Angelica keiskei inhibits endothelin-1 production through the suppressionof nuclear factor-kappaB. Biol Pharm Bull 2005;28:607–10.
22] Xu CB, Zheng JP, Zhang W, Zhang Y, Edvinsson L. Lipid-soluble smoke par-ticles upregulate vascular smooth muscle ETB receptors via activation ofmitogen-activating protein kinases and NF-kappaB pathways. Toxicol Sci2008;106:546–55.
23] Shioi A, Taniwaki H, Jono S, et al. Monckeberg’s medial sclerosis and inorganicphosphate in uremia. Am J Kidney Dis 2001;38:S47–49.
24] McCullough PA, Agrawal V, Danielewicz E, Abela GS. Accelerated atheroscle-rotic calcification and Monckeberg’s sclerosis: a continuum of advancedvascular pathology in chronic kidney disease. Clin J Am Soc Nephrol2008;3:1585–98.
25] Doherty TM, Fitzpatrick LA, Inoue D, et al. Molecular, endocrine, and geneticmechanisms of arterial calcification. Endocr Rev 2004;25:629–72.
26] Shen YH, Zhang L, Gan Y, et al. Up-regulation of PTEN (phosphatase andtensin homolog deleted on chromosome ten) mediates p38 MAPK stress signal-induced inhibition of insulin signaling. A cross-talk between stress signalingand insulin signaling in resistin-treated human endothelial cells. J Biol Chem2006;281:7727–36.
27] Wang XL, Zhang L, Youker K, et al. Free fatty acids inhibit insulin signaling-stimulated endothelial nitric oxide synthase activation through upregulatingPTEN or inhibiting Akt kinase. Diabetes 2006;55:2301–10.
28] Huang J, Niu XL, Pippen AM, Annex BH, Kontos CD. Adenovirus-mediatedintraarterial delivery of PTEN inhibits neointimal hyperplasia. ArteriosclerThromb Vasc Biol 2005;25:354–8.
29] Haq A, El-Ramahi K, Al-Dalaan A, Al-Sedairy ST. Serum and synovial fluidconcentrations of endothelin-1 in patients with rheumatoid arthritis. J Med1999;30:51–60.
30] Iwabuchi H, Kasama T, Hanaoka R, et al. Downregulation of intercellular adhe-sion molecule-1 expression on human synovial fibroblasts by endothelin-1. JRheumatol 1999;26:522–31.
31] Kuryliszyn-Moskal A, Klimiuk PA, Sierakowski S, Ciolkiewicz M. A studyon vascular endothelial growth factor and endothelin-1 in patients withextra-articular involvement of rheumatoid arthritis. Clin Rheumatol 2006;25:314–9.
32] Hamada K, Sasaki T, Koni PA, et al. The PTEN/PI3K pathway governs nor-mal vascular development and tumor angiogenesis. Genes Dev 2005;19:
2054–65.33] Suzuki A, Hamada K, Sasaki T, Mak TW, Nakano T. Role of PTEN/PI3K pathwayin endothelial cells. Biochem Soc Trans 2007;35:172–6.
34] Wilasrusmee C, Da Silva M, Siddiqui J, et al. Role of endothelin-1 in microvas-cular dysfunction caused by cyclosporin A. J Am Coll Surg 2003;196:584–91.





























