Embed Size (px)
Citation preview

VIROLOGY 144, 373-383 (1985)
Purification and Amino-Terminal Protein Sequence Analysis of the Mumps Virus Fusion Protein
ALFRED C. SERVER,**-l.@ JOHN A. SMITH,*,* M. NEAL WAXHAM,@ JERRY S. WOLINSKY,$ AND HOWARD M. GOODMAN*,?
*Department of Molecular Biology, Massachusetts General Hos@al, Boston, Massachusetts 02114 Departmxnts of tGenetics and $Pathology, Harvard Medical SCM, Boston, Massachusetts 02115; and
ODepa&nmt of Neurology, University of Texas Health .%imce Center, Houston, Texas 77225
Received February 14, 1985; accepted March 25, 1985
The fusion (F) protein of mumps virus was purified by immunoafllnity chromatography using an anti-F monoclonal antibody. The F protein was reduced and alkylated, and the Fi and Fa chains were isolated by high-pressure size exclusion chromatography. Twenty- three amino acid residues from the amino terminus of each chain were identified following automated Edman degradation. The amino-terminal sequence of the Fi chain was homologous to previously reported F1 sequences from three other paramyxoviruses (simian virus 5, Newcastle disease virus, and Sendai virus). Secondary structure predictions suggest an a-helical conformation for the mumps virus F1 amino-terminal sequence. A helical wheel model of the paramyxovirus F1 NH* terminus is presented which defines conserved and variable arcs of the helix and provides a spatial representation of this critical functional domain of the paramyxovirus fusion protein. 0 1985 Academic
Press, Inc.
INTRODUCTION
Mumps virus has two surface glycopro- teins, a hemagglutinin-neuraminidase (HN) protein and a fusion (F) protein (Jensik and Silver, 1976, Orvell, 1978a, b; Server et aL, 1982). The HN protein binds to sialic acid residues on target cells to ini- tiate virus infection. The F protein me- diates the fusion of lipid bilayers that is required for virus penetration and the cell-to-cell spread of infection. The F pro- tein of mumps virus is composed of two disulfide-linked glycopeptides (F, and Fz) that are generated by proteolytic cleavage of a precursor glycopeptide chain (NHz- Fz-F1-COOH) (Rima et aL, 1980; Naruse et a& 1981; Herrler and Compans, 1982; Merz et a,!., 1983). This post-translational processing of the F protein is character- istic of paramyxoviruses and has been best studied with viruses that can be grown to high titer: Newcastle disease
1 Author to whom requests for reprints should be addressed at the Department of Molecular Biology, Massachusetts General Hospital, Boston, Mass. 62114.
virus (NDV) (Hightower and Bratt, 1974; Nagai et aL, 19’76; Nagai and Klenk, 1977), Sendai virus (Homma and Ohuchi, 1973; Scheid and Choppin, 1974, 1977) and sim- ian virus 5 (SV5) (Peluso et aL, 1977). In the case of NDV (Nagai et aL, 1976; Nagai and Klenk, 1977) and Sendai virus (Homma and Ohuchi, 1973; Scheid and Choppin, 1974,1976), it has been demonstrated that cleavage of the F precursor is required for the expression of the biological activity of the fusion protein. The cleavage event results in the generation of a hydrophobic Fr NH2 terminus whose sequence is highly conserved among the three paramyxovi- ruses studied to date (SV5, NDV, and Sendai virus) (Gething et aL, 1978; Scheid et d, 1978; Richardson et aL, 1980). It has been suggested that the hydrophobic NH2 terminus of the F1 chain is the critical functional domain of the fusion protein interacting directly with the membrane lipid bilayer (Gething et aL, 1978; Scheid et al, 1980; Hsu et aL, 1983; Richardson and Choppin, 1983).
Studies of the F, NH, terminus of
373 6642-6822/35 $3.00 Copyright Q 1995 by Academic Press, Inc. All rights of reproduction in any form reserved.

374 SERVER ET AL.
mumps virus have been hampered by the difficulty in obtaining sufficient quantities of purified protein. However, advances in techniques for automated protein micro- sequencing have significantly reduced the amounts of material required for analysis (Hewick et a!., 1981). In this study, sub- nanomole quantities of purified mumps virus F protein were isolated by immu- noaffinity chromatography using an anti- F monoclonal antibody. Sequence analysis identified the first 23 amino acid residues of both the F1 and F2 chains. It was found that the F1 NH2-terminal region of mumps virus is homologous to those of SV5, NDV, and Sendai virus, with 12 of the first 20 amino acids conserved. The secondary structure of the mumps virus F1 NH2 terminus was predicted with the use of Chou and Fasman parameters (Chou and Fasman, 1978) and the sequence was found to have a high potential for a-helical conformation. Assignment of the F1 NHz- terminal sequences of mumps virus, SV5, NDV, and Sendai virus, to an a-helical wheel (Schiffer and Edmundson, 1967) de- fines conserved and variable arcs of the helix and provides a spatial representation of this critical functional domain of the paramyxovirus fusion protein.
MATERIALS AND METHODS
l+u.s. The Kilham strain of mumps virus (Kilham, 1951) was grown in CV-1 cells in the presence of rH]leucine and was isolated as previously described (Server et aL, 1982).
Immunoc&nity chromatography. Mono- clonal antibodies against the glycoproteins of mumps virus were generated and char- acterized as previously described (Server et aL, 1982). The antibodies were precipi- tated from mouse ascites fluids with am- monium sulfate and coupled to an acti- vated affinity support, Affigel-10 (Bio-Bad), according to the procedure of Staehelin et al. (1981). Immunoaffinity chromatography was performed using a modification of the method of Staehelin et al. (1981). Purified mumps virus was disrupted in 0.025M Tris ( hydroxymethyl ) aminomethane hy- drochloride (Tris-HCl), pH 7.5 containing
0.15 M NaCl, 0.1% sodium dodecyl sulfate (SDS), 1.0% (w/v) sodium deoxycholate, 1.0% (w/v) Triton X-100, 0.002 M phenyl- methylsulfonyl fluoride, and 100 Kallikrein inhibitor units/ml Aprotinin (Sigma) (buffer A). The virus lysate was centri- fuged at 95,000 g for 1 hr, and the super- natant was applied to the immunoaffinity column (3 ml bed volume) at a flow rate of 10 ml/hr. Following application of the virus lysate, the column was washed with buffer A (13 bed volumes), buffer B (0.025 M Tris-HCl, pH 7.5 containing 0.5 M NaCl and 0.2% Triton X-100) (26 bed volumes), and buffer C (0.15 M NaCl with 0.1% Tri- ton X-100) (10 bed volumes) at a flow rate of 30 ml/hr. Specifically bound virus gly- coprotein was eluted from the column with 0.2 N acetic acid, pH 2.5 containing 0.15 MNaCl and 0.1% Triton X-100 (buffer D) at a flow rate of 10 ml/hr and was precipitated from buffer D using the method of Horigome and Sugano (1983): 1 vol of protein solution was mixed with 4 vol of methanol:acetone (1:l v/v), heated at 37” for 2 min, and the protein precipi- tate pelleted by centrifugation at 10,000 g for 30 min.
Alkylation and reduction. The mumps virus fusion protein was reduced with 4.7 mM dithiothreitol in 7 M guanidine hy- drochloride/0.5 M Tris-HCl, pH 8.2 con- taining 0.005 M EDTA at 37” for 90 min. Excess iodoacetic acid was added, and the reaction was incubated for 1 hr at 37” and stopped by addition of glacial acetic acid.
High-pressure size exclusion chromatog- raphy. The mixture of the reduced and alkylated chains was lyophilized and re- dissolved in 7.5% trifluoroacetic acid (TFA) (Pierce)/92.5% elution buffer con- taining glacial acetic acid/Bpropanol/wa- ter (20115165, v/v/v). The chains were separated on a Spherogel TSK-SW 3000 analytical column (Beckman) (0.75 x 30 cm) with an SW precolumn (Beckman) (0.75 X 10 cm) using elution buffer at a flow rate of 0.5 ml/min (Bhown et al., 1981). The column outflow was monitored at 280 nm.
Amino acid an&y&s. Amino acid com- positions were determined using a Beck-

MUMPS VIRUS F-PROTEIN NH2 TERMINI 375
man 6300 analyzer after acid hydrolysis of samples at 110” for 24 hr in constant boiling HCl (0.025 ml) (Pierce, Sequanal grade) (Moore, 1972). Ninhydrin and two- channel (440 and 570 nm) integration pro- vided analysis of all amino acids except tryptophan, cysteine, asparagine, and glu- tamine, with l-7% confidence values at 100 pmol/amino acid and with a lower limit of detection of 25 pmol.
Protein sequence analyses. Sequence analyses of the mumps virus fusion pro-
A II 1
- ii
zo- I
lo" I ‘0 IS-
; ;
z z IO-
L3 3 2 5-
f :
oc ’ ,L. I\ 0 IO 20 30 40 50 60 70 so
FRACTION NUMBER
tein (F,-,) and the isolated F1 and F2 chains were performed with an Applied Biosystems 470A protein sequencer. The sequence program (Hewick et al, 1981) contains one coupling step (44”, 21 min), a double-cleavage (44O, 13.4 min), and automated conversion of the 2-anilino-5- thiazolinone derivatives into the corre- sponding 3-phenyl-2-thiohydantoin deriv- atives (Pth-amino acids) using 25% TFA (50°, 33 min). Polybrene (Tarr et al, 19’78; Klapper et al, 1978) was added to the
- 25.7
- 18.4 - 12.3
FIG. 1. Immunoaflinity purification of the mumps virus F protein. Mumps virus was metabolically labeled with [8H’Jeucine and disrupted in buffer A (see Materials and Methods for buffer compositions). The I-IN protein was separated from the other mumps virus proteins by immu- noaffinity chromatography using an anti-HN monoclonal antibody as described under Materials and Methods. A solution with the remaining mumps virus proteins (70 ml) was then applied to a second immunoaffinity column (3 ml bed volume) containing an anti-F monoclonal antibody; the resulting chromatogram is shown in panel A. Following sample application (point 0), the column was washed with 13 bed vol of buffer A (point a), 26 bed vol of buffer B (point b), and 10 bed vol of buffer C (point c). The specifically bound F protein was eluted from the immunoaffinity matrix with buffer D (point d). The flow rate during sample application and acid elution with buffer D was 10 ml/hr and during the wash steps with buffers A, B, and C was 30 ml/hr. Fractions of 1.5 ml were collected and 10 ~1 from each were subjected to liquid scintillation counting. The appropriate fractions were pooled based on radioactive counts, and an aliquot from each was saved for SDS-PAGE. (B) The fluorogram from the SDS-PAGE performed under reducing conditions. The proteins of mumps virus are shown in lane V. An aliquot of the sample applied to the anti-F immunoaflinity column is shown in lane 0 and contained a mixture of the mumps virus proteins following the removal of the HN protein. Aliquots from peaks I and II, in the chromatogram in pane1 A, are shown in lanes I and II, respectively. The designations to the left of lane V, used to label the polypeptide bands, are those of Scheid and Choppin (197’4): HN, hemagglutinin-neuraminidase; NP, nucleocapsid protein; F,, Fi polypeptide chain of the F protein; P, polymerase; M, membrane protein; Fz, Fz polypeptide chain of the F protein. The electrophoretic mobilities of protein standards of known molecular weights are indicated to the right of lane II. The standards used were myosin-H chain (200.0 kDa); phosphorylase B (92.5 kDa); bovine serum albumin (68.0 kDa); ovalbumin (43.0 kDa); a-chymotrypsinogen (25.7 kDa); a-lactoglobin (18.4 kDa), and cytochrome c (12.3 kDa).

376 SERVER ET AL.
glass filter disc, and five sequence cycles were run to reduce Polybrene-related con- taminants. The Pth-amino acids were identified by high-pressure liquid chro- matography on cyano columns (IBM), us- ing a 15 mM sodium acetate buffer (pH 5.8) and a complex gradient of acetoni- trile/methanol (92.5Y.5, v/v) (Hunkapiller and Hood, 1983).
SDS-polyacrylamide gel ekctrophoresis (SDS-PAGE). SDS-PAGE was performed under reducing conditions, according to the procedure of Laemmli (1970). After electrophoresis, the gels were processed for fluorography by the method of Bonner and Laskey (1974). Prestained protein standards (Bio-Rad) were used for molec- ular weight determinations.
RESULTS
ImmunoaJZnity pu@fication of mumps virus F protein. Radiolabeled mumps virus was disrupted with detergent in buffer A and used for the serial extraction of the virus glycoproteins (Fig. 1). The HN gly- coprotein was removed quantitatively by immunoaffinity chromatography using an anti-HN monoclonal antibody (Fig. lB, lanes V, 0). The effluent from the first column with the remaining mumps virus proteins was applied to a second immu- noaffinity column containing an anti-F monoclonal antibody (Figs. 1A and B). As shown in Fig. lB, removal of the F protein from the virus protein mixture was in- complete (lanes 0, I), although residual F protein could be removed by reapplication of the effluent to the column (data not
1 5
shown). The F protein, eluted from the column at acid pH (Fig. lA, peak II), contained no detectable contamination by other radiolabeled virus proteins (Fig. lB, lane II). Moreover, amino acid analysis of the eluent from the column prior to the application of virus proteins indicated that no appreciable antibody was lost from the immunoaffinity matrix during acid elution. The purified F protein was precipitated from the detergent-containing eluent with methanol/acetone prior to reduction and alkylation or sequence analysis.
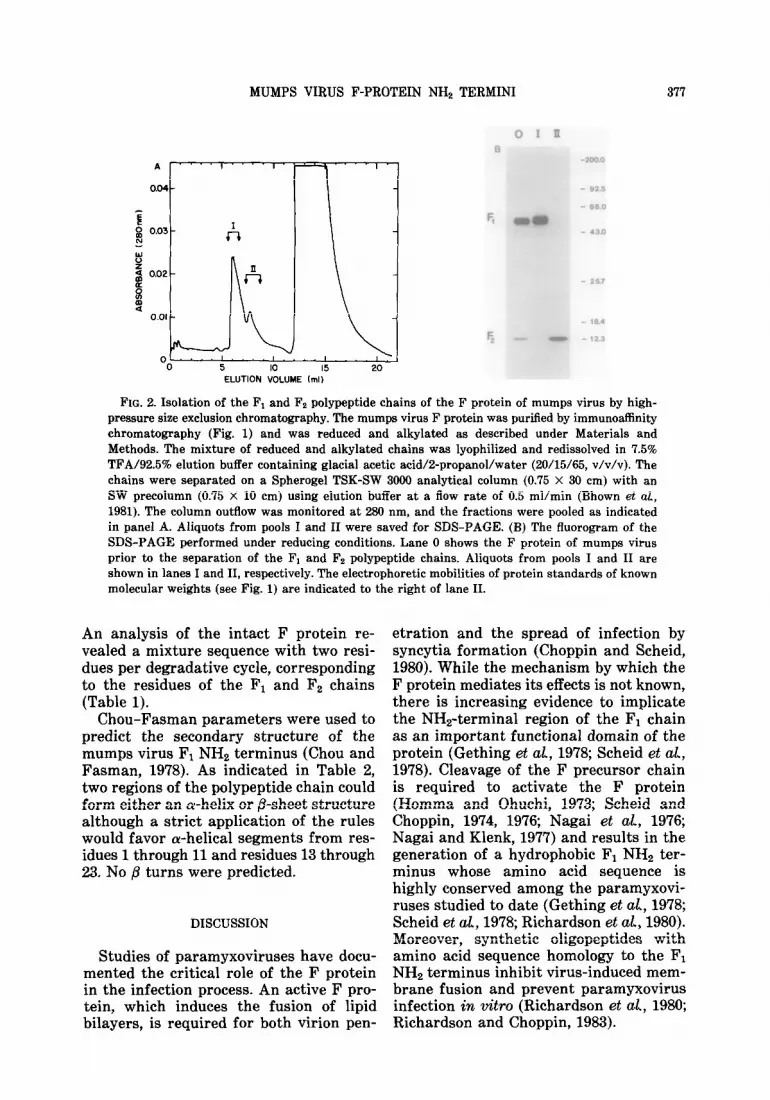
Purification of the Fl and Fe chains. Purified F protein was reduced and alkyl- ated under denaturing conditions. The sample was applied to a high-pressure size exclusion chromatography column equilibrated with acetic acid/2-propanol/ water (20/15/65, v/v/v). Three peaks were detected by uv absorbance at 280 nm (Fig. 2A). The broad peak eluting in the inclu- sion volume (12-19 ml) was shown to contain guanidine-HCl by an analysis of the reduction and alkylation buffer in the absence of protein. Fractions from the other two peaks were pooled, and an ali- quot from each pool was analyzed by SDS-PAGE. As illustrated in Fig. 2B, the F1 and F2 chains were separated from one another: pool I contained F1 (lane I) and pool II contained Fz (lane II).
Protein sequence analysis of the Fl and F2 chains. The purified F1 and Fz peptide chains were sequenced by automated Ed- man degradation with a gas-phase se- quenator (Table 1). Twenty-three amino acid residues were identified for each chain in the following sequences:
10 F1 NH~-Phe-Ala-Gly-Ile-Ala-Ile-Gly-Ile-Ala-Ala-Leu-Gly-Val-
15 20 Ala-Thr-Ala-Ala-Gln-Val-Thr-Ala-Ala-Val-
1 5 10 F2 NH~-Val-Asn-Ile-Asn-Ile-Leu-Gln-Gln-Ile-Gly-~r-Ile-Lys-
15 20 Gln-Gln-Val-Arg-Gln-Leu-Ser-Tyr-Tyr-Ser-
MUMPS VIRUS F-PROTEIN NH2 TERMINI 377
0 5 IO 15 20 ELUTION VOLUME (ml)
FIG. 2. Isolation of the F1 and Fz polypeptide chains of the F protein of mumps virus by high- pressure size exclusion chromatography. The mumps virus F protein was purified by immunoaffinity chromatography (Fig. 1) and was reduced and alkylated as described under Materials and Methods. The mixture of reduced and alkylated chains was lyophilized and redissolved in 7.5% TFA/92.5% elution buffer containing glacial acetic acid/2-propanol/water (20/15/65, v/v/v). The chains were separated on a Spherogel TSK-SW 3000 analytical column (0.75 X 30 cm) with an SW precolumn (0.75 X 10 cm) using elution buffer at a flow rate of 0.5 ml/min (Bhown et al, 1981). The column outflow was monitored at 280 nm, and the fractions were pooled as indicated in panel A. Aliquots from pools I and II were saved for SDS-PAGE. (B) The fluorogram of the SDS-PAGE performed under reducing conditions. Lane 0 shows the F protein of mumps virus prior to the separation of the Fi and Fz polypeptide chains. Aliquots from pools I and II are shown in lanes I and II, respectively. The electrophoretic mobilities of protein standards of known molecular weights (see Fig. 1) are indicated to the right of lane II.
An analysis of the intact F protein re- vealed a mixture sequence with two resi- dues per degradative cycle, corresponding to the residues of the F, and Fz chains (Table 1).
Chou-Fasman parameters were used to predict the secondary structure of the mumps virus F1 NH2 terminus (Chou and Fasman, 1978). As indicated in Table 2, two regions of the polypeptide chain could form either an a-helix or #I-sheet structure although a strict application of the rules would favor a-helical segments from res- idues 1 through 11 and residues 13 through 23. No @ turns were predicted.
DISCUSSION
Studies of paramyxoviruses have docu- mented the critical role of the F protein in the infection process. An active F pro- tein, which induces the fusion of lipid bilayers, is required for both virion pen-
etration and the spread of infection by syncytia formation (Choppin and Scheid, 1980). While the mechanism by which the F protein mediates its effects is not known, there is increasing evidence to implicate the NHz-terminal region of the F1 chain as an important functional domain of the protein (Gething et al., 1978; Scheid et al, 1978). Cleavage of the F precursor chain is required to activate the F protein (Homma and Ohuchi, 1973; Scheid and Choppin, 1974, 1976; Nagai et cd, 1976; Nagai and Klenk, 1977) and results in the generation of a hydrophobic F1 NH2 ter- minus whose amino acid sequence is highly conserved among the paramyxovi- ruses studied to date (Gething et cd, 1978; Scheid et aL, 1978; Richardson et cd, 1980). Moreover, synthetic oligopeptides with amino acid sequence homology to the FI NH2 terminus inhibit virus-induced mem- brane fusion and prevent paramyxovirus infection in vitro (Richardson et cd., 1980; Richardson and Choppin, 1983).

378 SERVER ET AL.
TABLE 1
SEQUENCE DATA FOR THE F, AND Fz PEPTIDE CHAINS AND FOR THE F PROTEINS
Cycle Fl F2
1 Phe 81 2 Ala 55 3 Gly ‘71 4 Ile 64 5 Ala 55 6 Ile 80 7 Gly 35 8 Ile 48 9 Ala 51
10 Ala 60 11 Leu 36 12 Gly 134 13 Val 86 14 Ala 31 15 Thr 15 16 Ala 22 17 Ala 15 18 Gln 16 19 Val 28 20 Thr 13 21 Ala 13 22 Ala 23 23 Val 9
ALb 213
RY” 95.3%
Val 797 Phe 180 Val 307 Asn 675 Ala 230 Asn 255 Ile 270 Gly 190 Ile 257 Asn 574 Ile 226 Asn 191 Ile 301 Ala 193 Ile 301 Leu 274 Ile 272 Leu 195 Gln 321 Gly 164 Gln 142 Gln 380 Ile 193 Gln 208 Ile 428 Ala 168 Ile 212 Gly 377 Ala 190 Gly 196 Tyr 395 Leu 163 Tyr 171 Ile 203 Gly 140 Ile 57 Lys 75 Val 138 Lys 53 Gln 117 Ala 129 Gln 67 Gln 183 Thr 54 Gln 107 Val 187 Ala 96 Val 56 Arg 277 Ala 118 Arg 63 Gln 74 Gln 109 Leu 128 Val 92 Leu 78 Ser 67 Thr 35 Ser 18 Tyr 91 Ala 71 Tyr 78 Tyr 134 Ala 91 Tyr 107 Ser 25 Val 60 Ser 9
858
94.3% 95.3% 93.2%
F protein Cycle
2 3 4 5 6 7 8 9
10 11 12 13 14 15 16 17 18 19 20 21 22 23
a Yields in picomoles of 3-phenyl-2-thiohydantoin(Pth)-amino acids plus phentylthiocarbamyl(Ptc)-amino acids, when present, minus background amino acids.
b AL-amounts loaded based on amino acid analyses (this work) and molecular weight estimates (Herrler and Compans, 1983).
’ RY-repetitive yields calculated from yields of representative, stable, Pth-amino acids.
The initial studies of the primary struc- ture of the F protein were done with three representative paramyxoviruses: SV5, NDV, and Sendai Virus (Gething et al, 1978; Scheid et al, 1978; Richardson et al, 1980). These viruses could be grown to suffi- ciently high titer for an NH2-terminal sequence analysis of the F protein by conventional solid-phase methods (Rich- ardson et aL, 1980). A similar analysis of mumps virus, an important human patho- gen in the paramyxovirus genus, was not done because of the difficulty in obtaining sufficient material for study. However, with the advent of new methods for au- tomated protein microsequencing (Hewick
et aL, 1981), this virus is now amenable to analysis, and the results are presented in this paper. Subnanomole quantities of the F protein of mumps virus were ob- tained by immunoaffinity chromatography using an anti-F monoclonal antibody. The high degree of specificity afforded by the use of a monoclonal antibody allowed the isolation of purified F protein with no detectable contamination. This material was subjected to amino acid sequence analysis both before and after reduction and alkylation, and 23 amino acids from the NH2 terminus of each of the peptide chains of the F protein were identified.
As anticipated, the NH2 terminus of

MUMPS VIRUS F-PROTEIN NH2 TERMINI 379
TABLE 2
SECONDARY STRUCWRE PARAMETERS FOR THE MUMPS VIRUS F1 NH2 TERMINUS PREDICTED FROM AMINO ACID SEQUENCE a
Position number
(9 Residue P* (Pa> i-i+3 PO Pd i-i+3 pt x 104
1 Phe 1.13 2 Ala 1.42 3 GUY 0.57 4 Ile 1.08 5 Ala 1.42 6 Ile 1.08 7 GUY 0.57 8 Ile 1.08 9 Ala 1.42
10 Ala 1.42 11 Leu 1.21 12 GUY 0.57 13 Val 1.06 14 Ala 1.42 15 Thr 0.83 16 Ala 1.42 17 Ala 1.42 18 Gln 1.11 19 Val 1.06 20 Thr 0.83 21 Ala 1.42 22 Ala 1.42 23 Val 1.06
- I 1.05
1.12 1.04 2 1.04 l-i 1.04 II 1.04
a 5
1.12 1.28
I 1.16 1.07 - 1.07 - 0.97
I 1.18 1.27 1.20 2 1.25 4 1.11 II 1.11
3 a 1.18 1.18
1.38 -
0.83 0.75 I 1.60 z 0.83 2 1.60 II 0.75 2 1.60 v
0.83 0.83 1.30 I _ 0.75 1.70 0.83 - 1.19 0.83 I 0.83 2 1.10 4 1.70 A
1.14 0.48 1.00 0.04 1.20 0.07 1.20 0.07 1.20 0.22 1.20 0.03 1.00 0.07 1.14 0.08 0.93 0.25 1.15 0.15 1.15 0.08 1.12 0.14 1.14 0.18 0.92 0.13 0.99 0.22 1.12 0.09 1.21 0.13 1.21 0.13 1.14 0.14 1.14 0.12
’ P, and P, are the amino acid conformational parameters for o-helix and B-sheet, respectively. (P.) and (P,) are the average a-helix and B-sheet conformational parameters, respectively, for any tetrapeptide extending from residue i to residue i + 3. pt is the product of the bend frequencies for the amino acids of a tetrapeptide extending from residue i to residue i + 3. The arrows indicate the extent of elongation of a potential conformation. Any peptide segment with (Pa) > 1.03 and with (P.) > (P,) is predicated as (Y- helical. Any segment with (P,) > 1.05 and with (P,) > (P.) is predicated as b-sheet. &turns are predicated for tetrapeptides with pt > 0.75 X lo-‘. The parameters listed and the rules for their application are those of Chou and Fasman (1978).
the Fi chain of mumps virus shows a high degree of amino acid sequence homology to the corresponding regions of the F proteins of other members of the para- myxovirus genus. The F1 NHe-terminal sequences of mumps virus, SV5, NDV and three stains of Sendai virus are listed in Fig. 3, including those determined directly and those predicted from nucleotide se- quence data. As indicated, 12 of the first 20 amino acid residues at the F1 NH2 terminus are conserved among the differ- ent viruses (Fig. 3). In contrast, a com- parison of Fz NH2 termini of mumps virus,
SV5, and Sendai virus shows that only 5 of the first 20 amino acids are conserved (Fig. 4). The high degree of sequence conservation at the F1 NH2 terminus is striking and is consistent with the current view that this region of the F1 polypeptide chain has a role in the biological function of the protein.
The primary structure of the fusion protein of respiratory syncytial virus (RSV) has recently been predicted from the nucleotide sequence of its mRNA (Collins et al, 1934). This virus is included in the paramyxovirus family but on the

380 SERVER ET AL.
1 5 10 15 20
@km&6vinrs phe-NbGl~Il~~IleG1~Il~N~~~~l~v~~~~~~~v~~-
sv5 -Na-ml-val--Leu-Na---val--Na--val-
uw - 1le - NbIle--Gly-val-- --al- --a-- Ile-
sen&iVirus1-Phe-Na-Val--IlF-Ile---Pro--
sendai viNsz -Pin?-NE-Val-- 'lb-Ile - - - Val - - Na - - Ile -
smdai Vicy -I&-Na-~al--%r-11e----Cal--per---I&Z-
FIG. 3. Comparison of the NHr-terminal sequence of the F1 polypeptide of mumps virus to those of other members of the paramyxovirus genus. The dashed lines indicate identity with the mumps virus sequence. The amino acid sequence for SV5 was determined directly (Richardson et al, 1980) and an identical sequence was predicated from nucleotide sequence data (Paterson et al, 1984). The amino acid sequence for NDV was determined directly (Richardson et al, 1980). The amino acid sequences for Sendai virus1 and Sendai virus2 were determined directly by Gething et al (1978) and by Richardson et al. (1980), respectively, and the sequence for Sendai virus3 was predicted by both Hsu and Choppin (1984) and Blumberg et al (1985).
basis of its uncharacteristic structure is conservation at this region of the F protein separated from the prototypic paramyxo- among members of the paramyxovirus viruses by inclusion in a separate genus, genus is a reflection of the evolutionary pneumovirus (Kingsbury et cd, 1978). The relatedness of these viruses. There is, hydrophobic F1 NH2 terminus of RSV has however, considerable evidence at variance only limited sequence homology to the with this interpretation. As indicated corresponding regions of the F proteins above, it is unlikely that the sequence of the prototypic paramyxoviruses with homology at the F1 NH2 terminus among only 4 of the first 20 amino acid residues the prototypic paramyxoviruses is simply conserved, at positions 1, 3, 7, and 17 in a measure of their relatedness since there the sequence (Collins et al, 1984). This is considerably less sequence conservation observation has led to the suggestion that of another region of the F protein, the F2 the fusion activity of the F protein is NH2 terminus. Moreover, a comparison of dependent upon the overall hydrophobicity the predicated amino acid sequences of of the F1 NHa terminus rather than on a the F proteins of SV5 (Paterson et al, common amino acid sequence. According 1984) and Sendai virus (Hsu and Choppin, to this view, the high degree of sequence 1934; Blumberg et aL, 1985) indicates that
1 5 lfl 15 -2n
rrrpsw- ~~IleA81-nt~~Il~~-Il~L~~~~~~r-
sv5 -ma-ma--val-- Pro-au--n - lwt-
aerdai vins tan-Il ~-ser~--~--val~u-Gly-Lys-ser-L~
FIG. 4. Comparison of the NHr-terminal sequence of the Fr polypeptide of mumps virus to those of SV5 and Sendai virus. The amino acid sequences for SV5 and Sendai virus were derived from nucleotide sequence data by Paterson et al. (1984) and by Blumberg et al (1985), respectively. The sequences are aligned to maximize the number of conserved amino acids. The dashed lines indicate identity with the mumps virus sequence. The SV5 sequence was predicated to begin as shown (Paterson et o& 1984) while gas-liquid chromatography and mass spectrometry were used to identify the first three amino acids of the Sendai virus sequence (Blumberg et a& 1985).

MUMPS VIRUS F-PROTEIN NH2 TERMINI 381
the degree of sequence conservation of either the hydrophobic signal peptide or the membrane-anchoring region does not exceed 25%. This is in marked contrast to the 60% conservation at the F1 NH2 ter- minus and suggests that there is selective pressure to maintain a substantial portion of the amino acid sequence, as well as the overall hydrophobicity, of this functionally significant region of the paramyxovirus F protein. The considerable variation in the sequence of the Fi NH2 terminus of the fusion protein of RSV may reflect impor- tant differences in the structure and mode of action of this protein.
Chou-Fasman empirical predictions sug- gest an a-helical structure for the mumps virus F1 NHz-terminal sequence. The ac- curacy of this predictive method for hydrophobic segments of membrane-as- sociated proteins has been debated. How- ever, the method has been used success- fully for selected hydrophobic peptides (Rosenblatt et ah, 1980; Briggs and Gier- asch, 1984) and the results presented here suggest that representation of the F1 NH2 terminus of mumps virus as an a-helix is a reasonable first approximation. Figure 5 depicts the first 20 amino acid residues of the F1 NH2 terminus of mumps virus on an a-helical wheel (Schiffer and Ed- mundson, 1967). In this model the a-helix is viewed down its axis with the polypep- tide backbone represented by the perim- eter of the wheel and the amino acid side chains indicated by spokes radiating from the wheel every 100” of arc. As anticipated by the uniformly hydrophobic amino acid sequence, the wheel fails to define an amphipathic helix with a clear separation of hydrophobic and hydrophilic residues. However, of considerable interest is the observation that all of the variable posi- tions in the paramyxovirus F1 NHz-ter- minal sequence (i.e., 2, 4, 5, 8, 9, 13, 16, and 19) fall on one side of the a-helix (Fig. 5). The opposite side of the helix, indicated in bold type in Fig. 5, is com- posed of amino acid residues that are absolutely conserved across four para- myxovirus species. This sequence-depen- dent model provides a spatial represen- tation of the mumps virus F1 NHz termi-
“Thr
VQl
FIG. 5. Alpha-helical wheel representation of the NH*-terminal sequence of the mumps virus F1 poly- peptide. The a-helix is viewed down its axis with the polypeptide chain represented by the perimeter of the wheel and the amino acid side chains repre- sented by the external spokes. The amino acids are separated by 100’ of arc, consistent with the 3.6 residues per turn of the a-helix. The amino acids of the NH2 terminus of the mumps virus F1 polypeptide are shown with their position number in the se- quence. The circled numbers indicate the positions of sequence variation among the different para- myxoviruses shown in Fig. 3.
nus and may offer insight into the action of this critical functional domain of the paramyxovirus F protein.
It has been suggested that the F protein initiates its interaction with membrane by binding to a cell surface receptor (Richardson and Choppin, 1983). It is con- ceivable that the conserved arc of an CY- helix formed by the F1 NH2 terminus mediates this interaction. Alternatively, the conserved arc of the helix may act directly on the membrane to destabilize lipid bilayers as a prerequisite to fusion. Further studies on the action of the F protein are clearly indicated and one sug- gested approach is to utilize synthetic oligopeptides that mimic the F1 NH2 ter- minus and that block the action of the F protein at the cell surface (Richardson et aL, 1980; Richardson and Choppin, 1983). The use of an a-helical wheel to represent the F1 NH2 terminus not only offers a novel method of comparing the corre- sponding sequences of four paramyxovi- ruses but provides a model for the rational design of synthetic peptide analogs for use in future studies of the paramyxovirus F protein.

382 SERVER ET AL.
ACKNOWLEDGMENTS
Technical assistance by Kathleen M. Martell (pro- tein sequence analysis), and Mark A. Malia, III, and James L. Campbell (amino acid analysis) is gratefully acknowledged. We thank Dr. Gerald D. Fasman, Dr. Andrzej Galat, Dr. Jiri Novotny, Dr. Lila M. Gierasch, and Dr. George D. Rose for helpful discussions and Dr. Benjamin M. Blumberg, Dr. Colomba Giorgi, Dr. Keith Rose, and Dr. Daniel Kolakofsky for access to their manuscript in press prior to its publication. This work was supported by grants from Hoechst AG (West Germany) and the National Institutes of Health (NS 20352).
REFERENCES
BHO~N, A. S., BENNETT, J. C., MOLE, J. E., and HUNTER, E. (1981). Purification and characteriza- tion of the gag gene products of avian-type c retroviruses by high-pressure liquid chromatog- raphy. And Biochem 112, 128-134.
BLUMBERG, B. M., GIORGI, C., ROSE, K., and KOLAK- OFSKY, D. (1985). Sequence determination of the Sendai virus fusion protein gene. J. Gen Fir01 66, 317-331.
BONNER, W. M., and LASKEY, R. A. (1974). A film detection method for tritium-labelled proteins and nucleic acids in polyacrylamide gels. Eur. J. Biochem 46,83-88.
BRIGGS, M. S., and GIERASCH, L. M. (1984). Exploring the conformational roles of signal sequences: Syn- thesis and conformational analysis of receptor protein wild-type and mutant signal peptides. Bio- chemist?y 23, 3111-3114.
CHOPPIN, P. W., and SCHEID, A. (1980). The role of viral glycoproteins in adsorption, penetration, and pathogenicity of viruses. Rev. Iv&x% Dis. 2,40-61.
CHOW, P. Y., and FASMAN, G. D. (1978). Empirical predictions of protein conformation. Ann Rev. Bio&em. 47,251-276.
COLLINS, P. L., HUANG, Y. T., and WERTZ, G. W. (1984). Nucleotide sequence of the gene encoding the fusion (F) glycoprotein of human respiratory syncytial virus. Proc. Nat1 Ad Sci. USA 81, 7683-7687.
GETHING, M. J., WHITE, J. M., and WATERFIELD, M. D. (1978). Purification of the fusion protein of Sendai virus: Analysis of the NH,-terminal se- quence generated during precursor activation. Proc Nat1 Acad Sci USA 75,2737-2740.
HERRLER, G., and COMPANS, R. W. (1982). Synthesis of mumps virus polypeptides in infected Vero cells. Vim&g 119, 430-438.
HERRLER, G., and COMPANS, R. W. (1983). Posttrans- lational modification and intracellular transport of mumps virus glycoproteins. J. I&-d 47, 354- 362.
HEWICK, R. M., HUNKAPILLER, M. W., HOOD, L. E., and DREYER, W. J. (1981). A gas-liquid solid phase peptide and protein sequenator. .J. Biol. Chem 256, 7990-7997.
HIGHTOWER, L. E., and BRATT, M. A. (1974). Protein synthesis in Newcastle disease virus-infected chicken embryo cells. J. viral 13, 788-800.
HOMMA, M., and OHUCHI, M. (1973). Trypsin action on the growth of Sendai virus in tissue culture cells. III. Structural difference on Sendai viruses grown eggs and tissue culture cells. J. Viral. 12, 1457-1465.
HORIGOME, T., and SUGANO, H. (1983). A rapid method for removal of detergents from protein solution. And B&hem. 130,393-396.
Hsu, M.-c., and CHOPPIN, P. W. (1984). Analysis of Sendai virus mRNAs with cDNA clones of viral genes and sequences of biologically important re- gions of the fusion protein. Proc. Nat1 Ad Sti USA 81, 7732-7736.
Hsu, M.-c., SCHEID, A., and CHOPPIN, P. W. (1983). Fusion of Sendai virus with liposomes: Dependence on the viral fusion protein (F) and the lipid composition of liposomes. fir- 126, 361-369.
HUNKAPILLER, M. W., and HOOD, L. E. (1983). Anal- ysis of phenylthiohydantoins by ultrasensitive gradient high-performance liquid chromatography. In “Methods in Enzymology” (C. H. W. Hirs and S. N. Timasheff, eds.), Vol. 91, pp. 486-493. Aca- demic Press, New York.
JENSIK, S. C., and SILVER, S. (1976). Polypeptides of mumps virus. J. fir01 17, 363-373.
KILHAM, L. (1951). Mumps virus in human milk and in milk of infected monkey. J. Amer. Med Assoc. 146, X3-1232.
KINGSBURY, D. W., BRATT, M. A., CHOPPIN, P. W., HANSON, R. P., HOSAKA, Y., TER MEULEN, V., NORRBY, E., PLOWRIGHT, W., ROTT, R., and WUNNER, W. H. (1978). Paramyxoviridae. Intervirology 10, 137-152.
KLAPPER, D. G., WILDE, C. E., and CAPRA, J. D. (1978). Automated amino acid sequence of small peptides utilizing polybrene. And Bzbchem 85, 126-131.
LAEMMLI, U. K. (1970). Cleavage of the structural proteins during the assembly of the head of bac- teriphage T,. Nature (Lundm) 227, 680-685.
MERZ, D. C., SERVER, A. C., WAXHAM, M. N., and WOLINSKY, J. S. (1983). Biosynthesis of mumps virus F glycoprotein: Non-fusing strains efficiently cleave the F glycoprotein precursor. J. Gen F&X 64, 1457-1467.
MOORE, S. (1972). The precision and sensitivity of amino acid analysis. In “Chemistry and Biology of Peptides” (J. Meienhofer, ed.), pp. 629-653. Ann Arbor Science, Ann Arbor, Mich.
NAGAI, Y., and KLENK, H.-D. (1977). Activation of precursors to both glycoproteins of Newcastle dis-

MUMPS VIRUS F-PROTEIN NH2 TERMINI
ease virus by proteolytic cleavage. Erology 77, 125-134.
NAGAI, Y., KLENK, H.-D., and Roar, R. (1976). Pro- teolytic cleavage of the viral glycoproteins and its significance for the virulence of Newcastle disease virus. Virolm 72, 494-508.
NARUSE, H., NAGAI, Y., YOSHIDA, T., HAMAGUCHI, M., MATSUMATO, T., ISOMURA, S., and SUZUKI, S. (1981). The polypeptides of mumps virus and their synthesis in infected chick embryo cells. fir- 112, 119-130.
ORVELL, C. (1978a). Immunological properties of purified mumps virus glycoproteins. J. Gen vird 41, 517-526.
ORVELL, C. (197813). Structural polypeptides of mumps virus. J. Gen. viral 41, 527-539.
PATERSON, R. G., HARRIS, T. J., and LAMB, R. A. (1984). Fusion protein of the parmyxovirus Simian virus 5: Nucleotide sequence of mRNA predicts a highly hydrophobic glycoprotein. Proc NatL Ad Sci. USA 81.6706-6710.
PELUSO, R. W., LAMB, R. A., and CHOPPIN, P. W. (1977). Polypeptide synthesis in Simian virus 5- infected cells. J. vird 23, 177-187.
RICHARDSON, C. D., and CHOPPIN, P. W. (1983). Oli- gopeptides that specifically inhibit membrane fu- sion by paramyxoviruses: Studies on the site of action. Irirdosy 131, 518-532.
RICHARDSON, C. D., SCHEID, A., and CHOPPM, P. W. (1980). Specific inhibition of paramyxovirus and myxovirus replication by oligopeptides with amino acid sequences similar to those at the N-termini of the F1 or HAP viral polypeptides. I%-* 105, 205-222.
RIMA, B. K., ROBERTS, M. W., MCADAM, W. D., and MARTIN, S. J. (1980). Polypeptide synthesis in mumps virus-infected cells. J. Gen Erol. 46, 501- 505.
ROSENBLATT, M., BEAUDETTE, N. V., and FASMAN, G. D. (1980). Conformational studies of the syn- thetic precursor-specific region of preproparathy- roid hormone. Proc. Natl. Acad Sci USA 77,3983- 3987.
SCHEID, A., and CHOPPIN, P. W. (1974). Identification of biological activities of paramyxovirus glycopro- teins. Activation of cell fusion, hemolysis, and infectivity by proteolytic cleavage of an inactive precursor protein of Sendai virus. Virology 57, 475-490.
SCHEID, A., and CHOPPIN, P. W. (1976). Protease activation mutants of Sendai virus. Activation of biological properties by specific proteases. virdogy 69, 265-277.
SCHEID, A., and CHOPPIN, P. W. (1977). Two disulfide- linked polypeptide chains constitute the active F protein of paramyxoviruses. Virology 80, 54-66.
SCHEID, A., GRAVES, M. C., SILVER, S. M., and CHOP- PIN, P. W. (1978). Studies on the structure and functions of paramyxovirus glycoproteins. In “Negative Strand Viruses and the Host Cell” (B. W. J. Mahy, and R. D. Barry, eds.), pp. 181- 193. Academic Press, New York/London.
SCHEID, A., Hsu, M.-c., and CHOPPIN, P. W. (1980). Role of paramyxovirus glycoproteins in the inter- actions between viral and cell membranes. In “Membrane-Membrane Interactions” (N. B. Gilu- lay, ed.), pp. 119-130. Raven Press, New York.
SCHIFFER, M., and EDMUNDSON, A. B. (1967). Use of helical wheels to represent the structures of pro- teins and to identify segments with helical poten- tial. Biophys. J. 7, 121-135.
SERVER, A. C., MERZ, D. C., WAXHAM, M. N., and WOLINSKY, J. S. (1982). Differentiation of mumps virus strains with monoclonal antibody to the HN glycoprotein. Zqfect Immun 35,179-186.
STAEHELIN, T., HOBBS, D. S., KUNG, H.-F., LAI, C.-Y., and PESTKA, S. (1981). Purification and character- ization of recombinant human leukocyte interferon (IFLrA) with monoclonal antibodies. J. Bid Chem 256,9750-9754.
TARR, G. E., BEECHER, J. F., BELL, M., and MCKEAN, D. J. (1978). Polyquarternary amines prevent pep- tide loss from sequenators. Ad Biochem. 84, 622-627.





























