Embed Size (px)
Citation preview

Pfliigers Arch (1984) 401:132-142 Pflfigers Archiv
European Journal of Physiology
�9 Springer-Verlag 1984
Pyramidal actions in identified radial motornuclei of the cat
Michael lllert and Ernst Wiedemann Physiologisches Institut der Universit~it Mfinchen, Pettenkoferstrasse 12, D-8000 Miinchen 2, Federal Republic of Germany
Abstract. 1. This study aimed to establish the projection from the corticospinal tract (CST) to the motoneurones in- nervating the deep radial (DR) forelimb muscles. In the anaesthetized cat stimulation of the contralateral pyramid and intracellular recording from identified forelimb motoneurones was used.
2. A train of pyramidal stimuli evoked disynaptic EPSPs in DR motoneurones. The effects were very similar in the different nuclei. Pyramidal IPSPs had a slightly longer latency and occurred in most cases together with disynaptic EPSPs. It is suggested that the inhibitory actions to the distal forelimb are predominantly relayed in a trisynaptic pathway, but that a disynaptic linkage seems possible as well.
3. The disynaptic pyramidal EPSPs remained after CST transection in C5. They were abolished after CST transections in C2. It is concluded that disynaptic corticospinal excitation of distal DR motornuclei is relayed in a short midcervical propriospinal system. Transection experiments at different cervical levels suggest that the majority of the propriospinal neurones is located in C3-C4.
4. The CST facilitated a variety of reflex pathways to motoneurones innervating distal forelimb muscles. Disynap- tic excitatory and inhibitory effects from cutaneous and low threshold group I muscle afferents were common. They were present in all investigated nuclei and powerfully facilitated from the CST. It is suggested that this allows the brain to adapt the reflex mechanisms of the distal forelimb to the synergistic-antagonistic relations between the muscles, which are changing according to the performed movement.
Key words: Forelimb motoneurones - Corticospinal tract - Short propriospinal neurones - Forelimb afferents
Introduction
In the C3-C, segments of the cat Illert et al. (1977, 1978) described a system of excitatory propriospinal neurones (PNs) with monosynaptic projection to forelimb moto- neurones. The PNs receive extensive monosynaptic excita- tory convergence from different higher motor centres and from forelimb afferents; they are also subject to a complex inhibitory control (Lundberg 1979). It has been postulated that this propriospinal relay acts as a premotoneuronal in- tegrative centre, and that the continuous regulation of ongo-
Offprint request to: M. Illert at the above address
ing fast forelimb movements may be one of its main functions (Illert et al. 1977; Lundberg 1979).
Alstermark et al. (1981) investigated the effects of spinal cord lesions on the ability of cats to retrieve food with the forelimb from a test tube. A target-reaching movement (paw is brought in contact with the food) and a food-taking move- ment (toe grasping and paw supination) were differentiated. Both movements were affected by spinal cord lesions at the C5 level: the target-reaching movement was impaired when the ventrolateral quadrant (which includes the axons of the C3-C4 PNs) was sectioned, the food-taking movement when the dorsolateral quadrant (which includes the cortico- and rubrospinal tract axons) was lesioned. When the dorso- lateral quadrant was transected in C2, both movements were affected at the same time. Based on these findings the authors postulate that the C3-C4 PNs may transmit the command for target-reaching; they Suggest that the food-taking movement depends on direkt activation of neuronal networks within the forelimb segments from the cortico- and/or rubrospinal tracts (see also Alstermark 1983).
This hypothesis asks for detailed knowledge on the ac- tion of the corticospinal tract (CST) and of the C3-C~ pro- priospinal system on the motoneurones to the various forelimb muscles. Whereas detailed data are available for muscles acting on the elbow (Illert et al. 1976a, 1977), the information on muscles crossing the distal joints is limited. In the present investigation we have therefore aimed to describe the CST actions on the different deep radial (DR) motornuclei and to analyze if they are transmitted in C3-C4 PNs. Special emphasis was placed on the DR nuclei located in the Cs-Thl segments (= distal nuclei) which innervate the distal limb muscles (Fritz et al. 1981). It will be shown that all DR motornuclei may be disynaptically activated from the CST via a C3-C4 propriospinal relay. We will further summarize findings on the interaction of the CST with pri- mary afferents. Preliminary results have been published (Illert and Wiedemann 1980, 1981).
Methods
The experiments were performed on cats operated under ether and subsequently anaesthetized with ~-chloralose (40-60 mg/kg i.v.). Supplementary doses of nembutal ( 5 - 1 0 mg/kg) were added. During recording the animals were immobilized with gallamintriethiodide and artificially ventilated. The endtidal CO2 concentration was kept within 3.5 - 5.0 vol.%. Blood pressure was continuously monitored and a drop below 80 mm Hg was counteracted by an intrave-

133
nous infusion of dextran and noradrenalin. The rectal temperature was kept within 36-38~
A laminectomy was made from the C5 to the Th2 segments. Corticospinal fibres were stimulated in the contra- lateral pyramid with a tungsten electrode (50 - i00 kO) in- serted from the exposed 4th ventricle at a level 4 mm rostral to the obex and about 0.8 mm lateral to the midline. The histological controls after the experiments showed that the stimulation, sites were in all cases located well within the pyramid. The contralateral pyramid was transected rostral to the stimulation site in most experiments avoid effects caused by antidromic stimulation of corticospinal fibres (Tsukahara et al. 1968).
To position the electrode within the centre of the pyramid we measured the threshold of the descending pyramidal volley with surface recording from the lateral funicle in C5 (Lance 1954). Electrode positions were accepted when the thresholds were below 30 ~tA and when stimulation with 200 laA evoked a maximal pyramidal volley (see Illert et al. 1976a). The descending fibres activated from this site were located in the ipsilateral dorsolateral spinal cord quadrant as was revealed with recordings in C5 from dissected dorsolateral and ventrolateral spinal cord quadrants (Laporte et al. 1956). A lesion of the ipsilateral dorsolateral funicle in C2 completely abolished the descending pyramidal volley (Fig. 5).
Intraceltular recording (potassium acetate electrodes, 2 - 5 M t2) was done from forelimb motoneurones identified by their antidromic invasion from one of the following nerves: biceps (Bi), brachialis (Br), brachioradialis (BRD), extensor carpi radialis longus (ECRL), extensor carpi radialis brevis (ECRB), extensor digitorum communis (EDC), extensor digitorum lateralis (EDL), extensor carpi ulnaris (ECU), extensor indicis proprius (EIP), abductor pollicis longus (APL, Crouch 1969), supinator (Sup), triceps (Tri) with its different heads, medianus (Mn) and ulnaria (U1). In some experiments two nerve branches of the DR were mounted together on a common stimulating electrode and the respective motoneurones (less than 12 % of the sam- ple) identified according to the I a convergence they received (Fritz et ai. 1984). In addition we always dissected the cutaneous superficial radial nerve (SR). Surface potentials recorded from the cord dorsum near the dorsal root entry zone were used for monitoring strengths of stimulation of the peripheral nerves, which are expressed in multiples of threshold (T). Motoneurones with a resting membrane potential of at least 40 mV or an action potential of more than 50 mV were accepted for analysis.
Results
L PSPs evoked from the CST
EPSPs. Figure 1 illustrates the finding that a train of pyra- midal volleys usually evoked short latency ESPSs. In the EDC motoneurone of L-O three stimuli in the pyramid elicited a distinct ESPS with a steep onset (N, O). Since two pyramidal volleys were without such an effect (M), the EPSP is related to the third volley. The segmental latency is 1.2 ms. The other recordings of Fig. I illustrate comparable results in different motoneurones projecting through the DR. The segmental latencies (below 2.2 ms) of the corticospinal EPSPs are given in the histograms of Fig. 2. In A 102 DR motoneurones from cats with an intact CST (hatched area)
are collected. The Iatencies distribute from 1.1 to 2.2 ms, with a peak around 1.5 ms. This distribution is similar to the one obtained by Illert et al. (1976a) in Bi and Br motoneurones. In the histograms B-H (Fig. 2) the neurones are grouped according to the different motornuclei they belong to. Although the material is small for some of them, the latency distribution mirrors in all cases that of the whole population. The values between 1.1 and 1.5 ms are compati- ble with a disynaptic linkage from the CST. We therefore postulate that the different DR motornuclei all receive disynaptic excitation from the CST. This is independent of the segmental location of the motornuclei (for location of the different nuclei see Fritz et al. 1981), of a proximal or distal insertion of the respective muscles and of the joints the muscles are acting at. The recordings of Fig. I exemplify muscles which act at the elbow (BRD, A - D; ECRL, E - G), at the wrist (ECRL, E - G ; ECU, P - S ) , at the wrist and the phalangeal joints (EDC, L - O ) and at the radio-ulnar joints (Sup, H - K ) .
To evoke disynaptic pyramidal EPSPs it was nearly always necessary to use short trains constisting of two to four stimuli, a single stimulus was only exceptionally effec- tive. The thresholds could be as low as 4 0 - 6 0 gA, but were mainly between 100-200 gA. The pyramidal origin of the disynaptic EPSPs was ascertained by withdrawal of the stimulating electrode (Iltert et al. 1976a); after a dorsal displacement of 0 . 5 - 1 . 0 m m the threshold increased drastically and, with unchanged stimulation strength, the amplitude of the EPSPs decreased. With a stimulation strength of 200 jaA the disynaptic effect disappeared when the electrode was moved to a position about 1.5 mm dorsally of the pyramid.
In addition to the disynaptic effects pyramidal stimula- tion often evoked long latency EPSPs (examples are illustrated in Fig. I J, N, R). They occurred in most cases parallel to the disynaptic effects. Sometimes the late EPSPs were the threshold effect of the pyramidal stimulation. They had a rather gradual onset, on which the disynaptic effects became superimposed. Very often the slow EPSPs grew to a considerable size. On withdrawal of the pyramidal electrode the slow EPSPs usually disappeared in parallel with the disynaptic effects, indicating that they were a genuine pyra- midal effect. In rare cases they were also evoked from regions dorsal to the pyramid, and then may have been due to coactivation of other neuronal systems (Illert et al. 1976 a).
IPSPs. Inhibition from the pyramid was often observed in DR motoneurones to distal muscles. The ECU motoneurone of Fig. 3 illustrates a corticospinal IPSP with a segmental latency of 2.1 ms. Always a train of volleys was necessary to evoke inhibition and the number of stimuli required was usually higher for the IPSPs than for the EPSPs (compare Figs. 1 and 3). The histogram D of Fig. 3 (hatched area) shows the segmental latencies of the IPSPs recorded from 41 identified Cs-Thl DR motoneurones. The values distribute from 1.5 to 3.0 ms and are slightly longer than the respective values of the EPSPs (compare Fig. 2 A). This suggests that the inhibitory actions from the CST to motoneurones of distal muscles are predominantly mediated in trisynaptic pathways.
In four motoneurones the IPSPs were evoked without a preceding EPSP, and Fig. 3 presents one example. In most neurones, however, the IPSPs occurred together with disynaptic EPSPs (for measuring of the IPSP latencies in

134
BRD
ECRL
Sup
EDC
ECU
A B Pyr 200 C D
E "1 40mv, A,H,L,P F G j 2 my, B-G, I-K,
M-O, Q-S
_ 2 ms, B,C,F,I. J, L- N.Q,R 1 ms, A,D,E,G,H,O,P,S
0,Sins, K
H L _ ~ ~ l Pyr200 J j ~ ! ~ '~ ~ ' t K / / ~
L M Pyr 200 N 0
P Q Pyr 200 R ~ / . 7 S I ~
Fig. 1A--S Pyramidal short latency EPSPs in different forelimb motoneurones. In this and the following figures the upper traces are intracellular records, the lower traces records from the spinal cord surface at the posi- tion of the respective motoneurones. The voltage calibrations apply for the intra- cellular records. Specimen records from a BRD (A--D), ECRL (E--G), Sup (H--K), EDC (L--O) and ECV (P--S) motoneurone are shown. Antidromic identification of the neurones in the left hand column. B, I, M, Q show the effect of 2 or 3 (Q) stimuli in the pyramid (Pyr, 200 gA), C, F, J, N, R the effect of one additional stimulus. The part of the traces indicated in this column by a bar is displayed at an expanded time scale in the right hand column
these cases refer to Fig. 1 F of Illert and Tanaka 1976). When superimposed on EPSPs, the IPSPs were more easily detected when depolarizing currents were applied. This enlarged the amplitude but not the latency of the IPSPs. Table 1 gives an overview on the occurrence of EPSPs, IPSPs and of mixed effects in the different nuclei. IPSPs were found in virtually all investigated nuclei. Pure IPSPs were seen in ECU and APL motoneurones. Mixed effects were common in the motoneurones to the finger extensors EDC, EDL, EIP and APL. In these neurones the excitatory actions were prominent, but inhibition was often detected when the number of stimuli was increased or when a depolarizing DC- current was used. Presence and frequency of the IPSPs as shown in Table 1 do certainly not mirror the potential inhibi- tory connections to the various motornuclei since depolariz- ing current passage was not done regularly and concealed inhibitory effects have possibly not been detected.
IL Segmental location of relay neurones
Mediation of disynaptic pyramidal EPSPs in a propriospinal relay was investigated with CST lesions at different cervical levels. We used either sampling of motoneurones or record- ing from the same neurone before and after the lesions. The records from the EDC motoneurone of Fig. 4 were obtained with the latter technique. Before the lesion two pyramidal volleys evoked a small EPSP (B), which was facilitated when a third volley was added (C, D). The segmental latency of the EPSP was 1.3 ms and thus of clear disynaptic origin. The microelectrode was withdrawn from the cell and the
ipsilateral dorsolateral quadrant was lesioned between C5 and C~ (E). The lesion virtually abolished the negativity of the early pyramidal volley (F), indicating that the transection was complete for the CST. The same cell was repenetrated ( H - K ) and stimulation of the pyramid evoked EPSPs which were almost identical in shape and amplitude with those recorded before. Similar disynaptic EPSPs were obtained with the sampling technique. It is concluded that the cells mediating this disynaptic excitation from the pyra- mid are located rostral to the CST section.
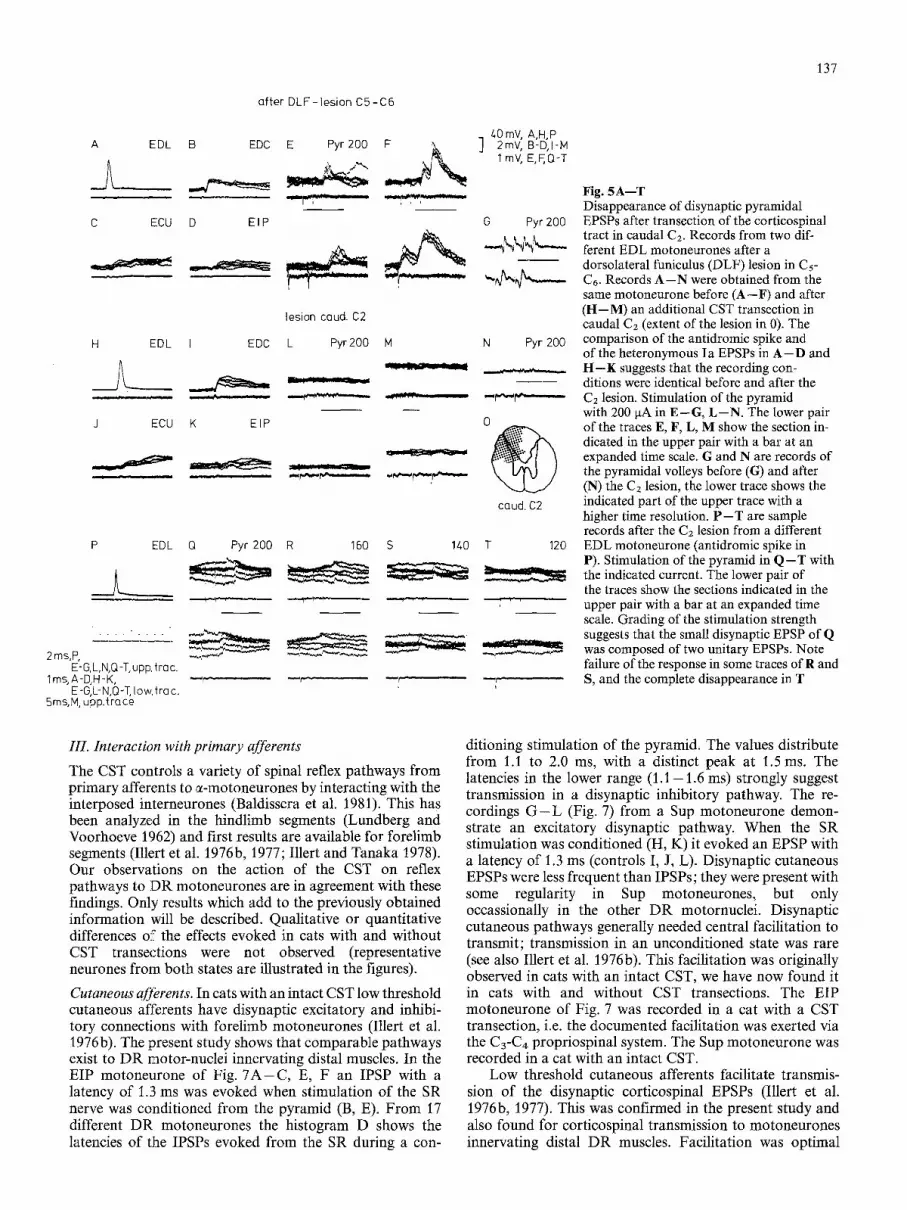
When the CST was transected ipsilaterally in C1 or Cz the disynaptic EPSPs disappeared completely. This is shown in Fig. 5. Records A - F were obtained from an EDL motoneurone after a complete CST transection in C5-C6. Stimulation in the pyramid evoked a disynaptic EPSP with a latency of 1.4 ms (E, F). After CST transection in caudal C2 ( H - M ; pyramidal volleys before and after the lesion in G, N; extent of the lesion in O) the pyramidal EPSPs disappeared completely, while the antidromic spike (A, H) and the heteronymous Ia EPSPs ( B - D , I - K ) were unchanged. These findings suggest that the cells relaying the corticospinal EPSPs are located in the spinal cord and not in the brainstem. Records P - T were obtained later on from another EDL motoneurone, in which a very small disynaptic EPSP had remained (Q). Grading of the stimulation strength suggests that it was composed of two unitary EPSPs (note the failure of the responses in some traces of R and S, and the complete disappearance in T). Apparently the most rostral location of the neurones intercalated in this disynap- tic pathway is in caudal Cz. The differential effect of CST

135
Table 1. Distribution of pyramidal PSPs in forelimb motornuelei before/after CST transections. Segmental tatencies: EPSPs < 2.2 ms, IPSPs < 3.2 ms
Motoneurones Bi Br BRD TLo TLa TM Sup ECR EDC EDL ECU APL EIP
n 29/10 20/13 2/3 10/2 3/0 3/0 9/5 13/6 23/48 19/35 17/17 14/16 7/15 EDSP 23/10 16/13 2/3 7/2 1/0 7/4 12/6 16 /47 12/33 11/11 0/13 3/14 IPSP 1/0 2/0 1/1 3/2 Mixed 6/0 4/0 3/0 1/0 i/0 2/1 1/0 7/1 7/2 5/5 11/1 4/1
6 0 -
n
A CST ~ intact [ D transected
I I i 0 I i i
1.2 2.0 ms
1 5 - EDC D EOL E ECU
I I I I I I I
1•t F SUP G APL H EIP
Fig. 2A--H. Segmental latency of the pyramidal EPSPs in the dif- ferent DR motornuclei. The ordinates of the histograms give the number of ceils (n, note different scales), the abscissae the time interval between arrival of the effective pyramidal volley at the position of the motoneurones and onset of the EPSP. The hatched bars indicate values obtained in animals with an intact corticospinal tract (CST), the open bars give values obtained in animals with a complete CST transection in the C5-C6 segments. (A) All DR motoneurones recorded in the different experiments (without/with CST transection: 102/151). (B) ECR 13/6, (C) EDC 23/48, (D) EDL 19/35, (E) ECU 16/16, (F) Sup 9/5, (G) APL 11/14, (H) gIP 7/15. The numbers in A are larger than the sum of cells given in B - - H since A includes motoneurones which were not identified in the projection to a particular muscte
lesions in the caudal C2 and in the C5-C6 segments suggests that the cell bodies of the relaying PNs are located between the C2 and the C5-C6 segments.
The propriospinally mediated disynaptic excitation reached all DR motornuclei irrespective of the segmental location of the motoneurones, the action of the muscles, or their proxime-distal location within the limb (Table 1). This is shown in the histograms of Fig. 2 (open areas) which collect the segmental latencies of the corticospinal EPSPs (below 2.2 ms) from cats with complete CST transections
(between the C5-C6 segments or, in case of the C6-C7 nuclei, in cranial C5). Histogram A presents the data of 151 DR motoneurones, B - H the values for the different nuclei. In all nuclei the latencies distribute between /.2 or 1.3 ms to 2.0 ms and are thus compatible with a disynaptic linkage (the small number of ECR motoneurones was not enlarged since most of the C 6 - C 7 DR motoneurones analyzed by Illert et al. 1977, belonged to this nucleus). When the amplitudes and the shapes of the propriospinally mediated disynaptic EPSPs were compared, clearcut differences of the potentials relayed to the various motornuclei were not obvious. How- ever, propriospinal excitation was particularly pronounced in the Sup motoneurones, where the onset of the EPSPs was very steep and two or three pyramidal volleys were sufficient to evoke maximal effects.
From different experiments we got the impression that the size of the disynaptic corticospinal EPSPs was reduced by lesions of the dorsolateral funicle in the C3 and C, segments, but not affected by corresponding lesions in C~ and C6. This would indicate that the propriospinal nucleus is restricted to the C3-C4 segments. To verify this impression we performed in one animal consecutive lesions at various spinal cord levels. The main findings are summarized in Fig. 6 with recordings from three different EDC motoneurones. In the control situation with an intact CST, pyramidal stimulation evoked a disynaptic EPSP (B, C). When the CST was transected between the Cs-C6 segments ( D - H ) a disynaptic EPSP with comparable amplitude and latency was recorded in 9 cells (E, F). A further CST transec- tion was performed in caudal C4 ( I - M). In 5 motoneurones recorded afterwards the amplitudes, latencies and shapes of the EPSPs were in the range of the control cells (Fig. 6 J, K), in two neurones they were decreased. CST transections in middle C4 or in caudal C3 drastically decreased the amplitudes of the disynaptic EPSPs (4 cells). This indicates that at least the majority of the PNs with monosynaptic projection to Cs-Tht DR motoneurones is located within C3 and C4.
Transmission of pyramidally evoked IPSPs has not been investigated to a comparable degree. After CST transections in caudal C5 IPSPs were found in C8-Th~ DR motoneurones (Table 1). The segmental latencies are given in the histogram D of Fig. 3 (open area), but the number of cells is too small (n = 14) to state that the latency distribution is identical before and after the lesions. Nevertheless, the linkage seems trisynaptic, although the value of 1.6 ms could indicate a disynaptic connection. IPSPs were no longer observed after CST transections in C1 and C2. In view of the general parallelism with the pyramidally evoked disynaptic EPSPs we postulate that a C3-C4 propriospinal relay may transmit IPSPs from the CST to DR motoneurones innervating distal forelimb muscles.

A ECU B Pyr 200*3 C Pyr 200,, 4
D E F t " ~ i 10 7
0 1.5 3.0 ms
20 mY, A 2 mY, B,C,E,F
2 + 10 ms, B,C 1 + 5 ms, A,E,F
Fig. 3A--F. Corticospinal inhibition in forelimb motoneurones. The specimen records are from an ECU motoneurone of a cat with an intact CST (identification in A). The pyramid was stimulated with 200 gA (3 shocks in B, 4 in C). E and F display at an expanded time scale the part of the records indicated with a bar in B and C. The histogram D presents the segmental latencies between 1.5 and 3.1 ms of corticospinal IPSPs in DR motoneurones. The hatched bars give neurones from cats with an intact CST (n = 41), the open bars neurones from cats with a CST transection in the C5-C6 segments (n = 14)
A E00
before ~ lesion ~_-- ,
] 2mV
E C5-C6 F Pyr G 50- [ 7
after n ~ L~ lesion
r o - r , , 1.2 2.0 m s
H EDC t Pyr 200 J K
after ~ ~ ~ ~ ~ _ ~ lesion :~_. _._: .... ~ " - - - ~ ' ' - . . . . . . . : , . ~ " ~ . r ~ ' . " - . - - -
2ms,B,C,I,J 1 ms,A,D,F,H,K
Fig. 4A--K. Disynaptic pyramidal EPSPs before and after a transection of the corticospinal tract at the C5-C6 level. A--D and H--K are records from the same EDC motoneurone before and after the lesion in E. A comparison of the M-spike and of the homonymous Ia EPSPs in A and H suggests that the recording conditions were similar in both cases. Stimulation of the contralateral pyramid in B, C, I, J with 200 IxA (D and K give the part of the traces indicated in C and J with a bar at an expanded time scale). F shows the surface recordings of the descending pyramidal volleys before and after the lesion. The histogram G gives the segmental latencies (below 2.2 ms) of the pyramidal EPSPs in 140 Cs-Thl DR motoneurones after CST transection in C5
after DLF- les ion C5-C6
A EDL B EDC E Pyr 200 F
C ECU D E I P
lesion coud. C2
H EDL I EDC L Pyr 200 M
J ECU K EIP
P EDL O Pyr 200 R 160 S 140
2 ms, P, E-G, L,N,Q-T, upp. tra c.
1 ms, A-D,H-K, E-G,L-N,Q-T, low.ira c.
5ms, M, upp.trace
Z,O mY, A,H,P ] 2mY, B-D,I-M
1 mV, E,F,Q-T
O Pyr 200
qW'G--
N Pyr 200
o
coud. C2
T 120
137
Fig. 5A--T Disappearance of disynaptic pyramidal EPSPs after transection of the corticospinal tract in caudal C2. Records from two dif- ferent EDL motoneurones after a dorsolateral funiculus (DLF) lesion in C5- C6. Records A--N were obtained from the same motoneurone before (A--F) and after (H--M) an additional CST transection in caudal C2 (extent of the lesion in 0). The comparison of the antidromic spike and of the heteronymous Ia EPSPs in A--D and H--K suggests that the recording con- ditions were identical before and after the C2 lesion. Stimulation of the pyramid with 200 laA in E--G, L--N. The lower pair of the traces E, F, L, M show the section in- dicated in the upper pair with a bar at an expanded time scale. G and N are records of the pyramidal volleys before (G) and after (N) the Cz lesion, the lower trace shows the indicated part of the upper trace with a higher time resolution. P--T are sample records after the C2 lesion from a different EDL motoneurone (antidromic spike in P). Stimulation of the pyramid in Q--T with the indicated current. The lower pair of the traces show the sections indicated in the upper pair with a bar at an expanded time scale. Grading of the stimulation strength suggests that the small disynaptic EPSP of Q was composed of two unitary EPSPs. Note failure of the response in some traces of R and S, and the complete disappearance in T
IlL Interaction with primary afferents
The CST controls a variety of spinal reflex pathways from primary afferents to ~-motoneurones by interacting with the interposed interneurones (Baldissera et al. 1981). This has been analyzed in the hindlimb segments (Lundberg and Voorhoeve 1962) and first results are available for forelimb segments (Illert et al. 1976b, 1977; Illert and Tanaka 1978). Our observations on the action of the CST on reflex pathways to DR motoneurones are in agreement with these findings. Only results which add to the previously obtained information will be described. Qualitative or quantitative differences of the effects evoked in cats with and without CST transections were not observed (representative neurones from both states are illustrated in the figures).
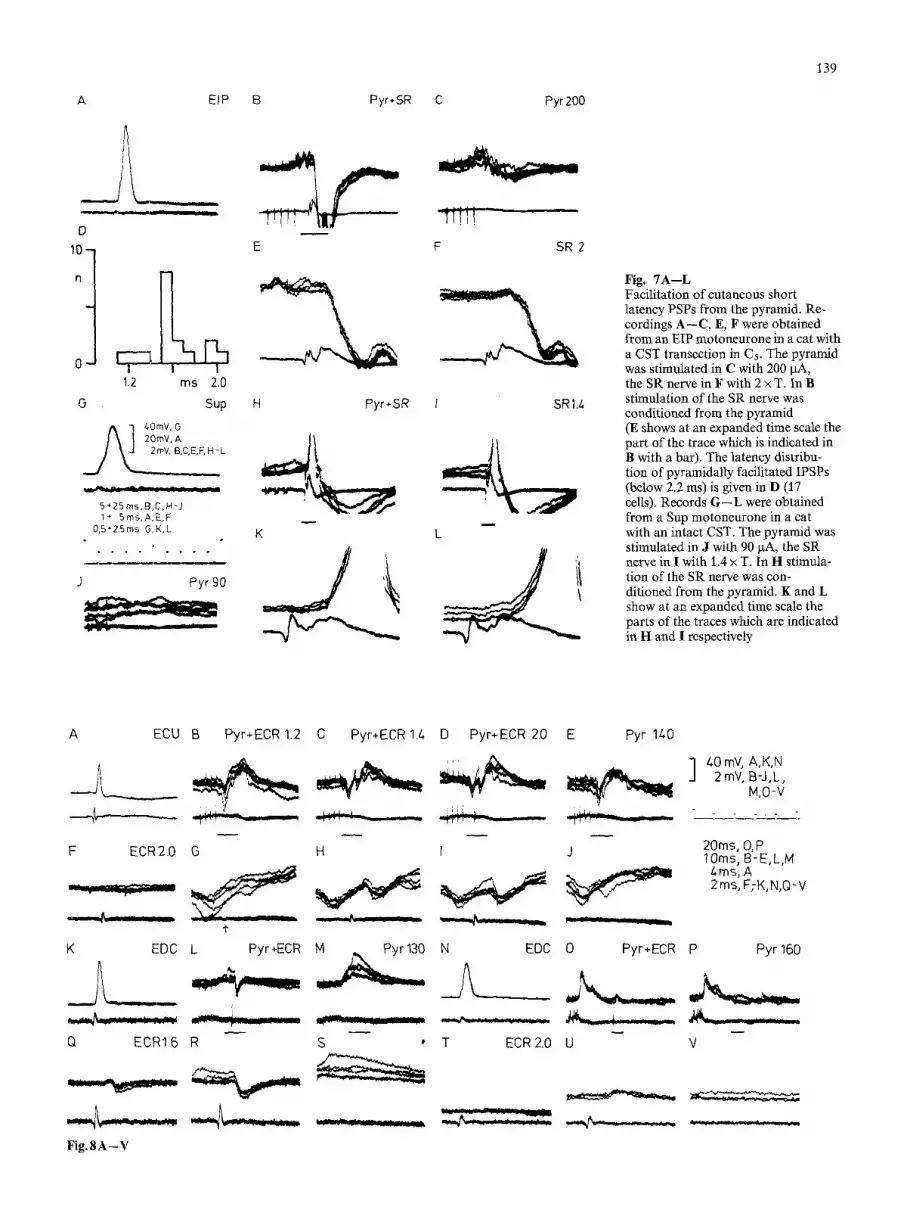
Cutaneous afferents. In cats with an intact CST low threshold cutaneous afferents have disynaptic excitatory and inhibi- tory connections with forelimb motoneurones (Illert et al. 1976 b). The present study shows that comparable pathways exist to DR motor-nuclei innervating distal muscles. In the EIP motoneurone of Fig. 7 A - C , E, F an IPSP with a latency of 1.3 ms was evoked when stimulation of the SR nerve was conditioned from the pyramid (B, E). From 17 different DR motoneurones the histogram D shows the latencies of the IPSPs evoked from the SR during a con-
ditioning stimulation of the pyramid. The values distribute from 1.1 to 2.0 ms, with a distinct peak at 1.5 ms. The latencies in the lower range (1.1-1.6 ms) strongly suggest transmission in a disynaptic inhibitory pathway. The re- cordings G - L (Fig. 7) from a Sup motoneurone demon- strate an excitatory disynaptic pathway. When the SR stimulation was conditioned (H, K) it evoked an EPSP with a latency of 1.3 ms (controls I, J, L). Disynaptic cutaneous EPSPs were less frequent than IPSPs; they were present with some regularity in Sup motoneurones, but only occassionally in the other DR motornuclei. Disynaptic cutaneous pathways generally needed central facilitation to transmit; transmission in an unconditioned state was rare (see also Illert et al. 1976b). This facilitation was originally observed in cats with an intact CST, we have now found it in cats with and without CST transections. The EIP motoneurone of Fig. 7 was recorded in a cat with a CST transection, i.e. the documented facilitation was exerted via the C3-C4 propriospinal system. The Sup motoneurone was recorded in a cat with an intact CST.
Low threshold cutaneous afferents facilitate transmis- sion of the disynaptic corticospinal EPSPs (Illert et al. 1976b, 1977). This was confirmed in the present study and also found for corticospinal transmission to motoneurones innervating distal DR muscles. Facilitation was optimal

138
A EDC
_2 B Pyr 200 C 4-0 mY, !r . . . . - . . . .
20mV, A,D ~ ~ 2rnV, B,C 2ms, A,B,D,E,T,J
' lm V, E,F, J,K 1 ms, C,F,H,K,M
lesion C5-C6
EDC E Pyr200 F G H before
after
lesion caud. CL
EDC J Pyr200 K L M-- I \ , ~ before I
after Fig. 6A--M. Disynaptic pyramidal EPSPs after transections of the corticospinal tract at different cervical levels. Records from three different EDC motoneurones in the same cat (antidromic spikes in A, D, I). A--C: intact CST; D--F: after DLF lesion between Cs-C6 (shown in G); I--K: after DLF lesion in caudal C4 (shown in L); descending pyramidal volleys before and after the lesions in H and M. In B, E, .1 the pyramid was stimulated with 3 shocks at 200 laA; C, F, K display the part of the traces indicated in B, E, ./at an expanded time scale
when the cutaneous volley preceded the pyramidal volley by about 0.5 ms. When the conduction distances are taken into account, this would leave time for one synapse intercalated in the cutaneous-propriospinal projection. Accordingly we suggest a monosynaptic linkage as in the case of the proximal motoneurones (see Discussion in Illert et al. 1977).
Group I muscle afferents. Similarly as described by Illert et al. (1976b) we frequently found pyramidal facilitation of disynaptic group I IPSPs. This was regularly observed in the reciprocal connection between the Bi/Br and Tri muscles which is relayed by Ia inhibitory interneurones (Illert and Tanaka 1978). The situation became complex in the pathways interconnecting distal forelimb muscles. The re- cors A - J of Fig. 8 were obtained from an ECU motoneurone (transection of the CST) and illustrate facilita- tion of group I IPSPs between wrist muscles. In this neurone stimulation of the pyramid elicited a presumed trisynaptic IPSP (E, J). Stimulation of the ECR with a strength supramaximal for group I afferents was ineffective (F), but when it was combined with stimulation of the pyramid a disynaptic IPSP was facilitated. The threshold of the IPSP was low (B, G), and the effect was maximal at 1.4 x T (C, H) when the incoming volley was about half the size of the maximal group I volley.
A comparable effect has been observed from ECU afferents in ECR motoneurones. Conditioning stimulation of the pyramid revealed similar inhibitory effects between a variety of other distal forelimb muscles. In the EDC motoneurone of K - M and Q - S the ECR volley evoked a monosynaptic EPSP (see Fritz et al. 1984). Superimposed on its decay phase is a disynaptic TPSP which was facilitated by conditioning pyramidal stimulation (L, R). Threshold was at 1.1 x T, the effect was maximal at 1.6 x T (see also Illert et al. 1976b).
In most of these cases we do not know the receptor origin of the facilitated group I IPSPs. ECR and EDC are very close synergists, which finds its expression in strong bidirectional monosynaptic Ia connections between them (Fritz et al. 1984). The group I IPSPs of Fig. 8, K - M , Q - S could accordingly correspond to a typical inhibitory I b connection between synergists (Laporte and Lloyd 1952), but we are hesi- tant with this interpretation. The IPSPs were evoked at the same low threshold as were monosynaptic I a EPSPs. This could be due to an absence of threshold separation between Ia and Ib afferents, but it could also reflect convergence of Ia afferents onto inhibitory Ib interneurones (Fetz et al. 1979). Similar explanations could apply for the inhibitory connections between ECR and ECU illustrated in Fig. 8 A - J .
A
L - m i
D
k 0 -
1.2 ms 2.0
O Sup
j~] 40mV, G
, ,C,E,F, N IL
5 + 2 5 m s , B , C , H - J 1 + 5ms, A,E,F
0,5§ Z5 ms G,K,L
J Pyr 90
EIP B Pyr+SR C Pyr 200
E F SR 2
C:& Pyr*SR 1 SR1.4
K
L
L
139
Fig. 7A--L Facilitation of cutaneous short latency PSPs from the pyramid. Re- cordings A--C, E, F were obtained from an EIP motoneurone in a cat with a CST transection in C5. The pyramid was stimulated in C with 200 I.tA, the SR nerve in F with 2 x T. In B stimulation of the SR nerve was conditioned from the pyramid (E shows at an expanded time scale the part of the trace which is indicated in B with a bar). The latency distribu- tion of pyramidally facilitated IPSPs (below 2.2 ms) is given in D (17 cells). Records G - - L were obtained from a Sup motoneurone in a cat with an intact CST. The pyramid was stimulated in J with 90 ~tA, the SR nerve in I with 1.4 x T. In H stimula- tion of the SR nerve was con- ditioned from the pyramid. K and L show at an expanded time scale the parts of the traces which are indicated in H and I respectively
A ECU
F ECR2.0
K EDC
,L__ Q ECR1.6
Fig.8A-V
B Pyr+ECR 1.2 C Pyr+ECR 1.L D Pyr+ECR 2.0 E Pyr 1L0
. , , , . ~ 2mY, B-J,L,
M,O-V
G H I J
L Pyr+ECR M ,,, Pyr130 N EDC
"J'" ~'r " - - 2 :-2 ~
R S ' T ECR 2.0
20ms, (3, P 10rns, B-E,L,M /-,ms, A 2 ms, F;K,N,Q-V
0 Pyr§ P Pyr 160
U V

140
Disynaptic group I EPSPs were frequently encountered between distal forelimb muscles. As in case of the IPSPs the thresholds were often low and the effects maximal at a stimulation strength submaximal for group I afferents. The EDC motoneurone of Fig. 8 N - P , T - V (intact CST) illustrates such an effect between ECR and EDC. The disynaptic linkage of these excitatory effects points to their transmission in I b pathways, but as in case of the disynaptic group I IPSPs we can not differentiate between I a and Ib afferents.
The two EDC motoneurones of Fig. 8 differ in their monosynaptic I a activation from the ECR. In the neurone with monosynaptic Ia excitation (K) pyramidal stimulation facilitated disynaptic group I inhibition from the ECR; in the neurone without monosynaptic Ia excitation from the ECR (N) disynaptic group I excitation from the same nerve was facilitated. The presence of disynaptic group I inhibition was regular in EDC motoneurones with monosynaptic la excitation from the ECR (see also Illert et al. 1976b), but our material is too small to establish that disynaptic group I excitation from ECR is a common finding in EDC motoneurones without monosynaptic Ia excitation from ECR (note that only 10% of the EDC motoneurones are devoid ofmonosynaptic I a excitation from ECR; Fritz et al. 1984). Nevertheless, these EDC motoneurones are certainly interesting in view of the varying functional synergism be- tween ECR and EDC.
Discussion
Short latency excitation from the CST is well documented in c~-motoneurones innervating proximal muscles (Illert et al. 1976a) and has now been demonstrated in the DR motoneurones to the distal forelimb. The distribution from 1.1 to 2.0 ms of the segmental latencies from the CST with a peak at 1.6 ms is in agreement with the distribution re- ported for other forelimb motornuclei (Illert et al. 1976a; Alstermark and Sasaki 1984a). At least in the shorter range it is compatible with a disynaptic linkage from the CST. Disynaptic excitation may have shorter segmental latencies in some other pathways (e.g. Griller and Hongo 1972; Hongo et al. 1969), but in the case of the CST the dispersed conduction velocity in this tract system and in the inter- calated interneurones (Illert et al. 1978) must be considered. Distribution of the segmental latencies was unimodal in the Bi and Br motoneurones investigated by Illert et al. (1976 a), but in the present material it is probably bimodal. This could indicate that the latencies in the longer range are due to one additional synapse.
Our analysis showed that a system of midcervical short PNs may mediate disynaptic excitation to distal forelimb motoneurones, similarly as was described for motoneurones located in the C6-Cv segments (Illert et al. 1977). Alstermark et al. (1984b, see also Alstermark 1983) reached the same conclusion with an independent technique. The location of
the propriospinal relay to the distal limb muscles should be compared with the location of midcervical short pro- priospinal neurones as established by neuroanatomical techniques. Localized HRP injections into the lateral forelimb motornuclei in C6-C7 (Grant et al. 1980; Molenaar 1978) and in C8 (Molenaar 1978) revealed propriospinal neurones from the C3 to the Th2 segments. The rostral border of this nucleus in C2 is identical with the rostral border of the electrophysiologically defined propriospinal system; the caudal border, however, extends further caudally. Illert et al. (1977) discussed if the propriospinal neurones in Cs may represent the caudal extension of the C3- C4 propriospinal system, and if these neurones may transmit propriospinal excitation to motoneurones innervating distal DR muscles. Our results can not exclude this possibility, but they indicate that the majority of the PNs to the distal muscles is located in the C3-C4 segments.
In the action of the C3-C4 PNs onto C8-Thl (DR) o r C 6-
C7 motoneurones (Bi, Br, DR) conspicious differences were not found. In particular, the latency, amplitude and shape of the disynaptic EPSPs seemed identical. It is therefore tempting to speculate that a common system of C3-C~ PNs projects to both, proximal and distal forelimb motornuclei. Previous studies with direct recording from the C3-C4 FNs have pointed in this direction. Illert et al. (1978) established the "tapering" curves of individual PNs and found that the neurones give off collaterals a considerable distance before their final termination. Due to the length of the forelimb motornuclei (Fritz et al. 1981, 1982) one propriospinal axon could contact several of them, thus combining the respective muscles to a common function. However, there are also clear indications for differentiated subpopulations within the C3- C4 PNs. Illert et al. (1978) observed that the PNs differ in the monosynaptic convergence they receive. Furthermore, recent experiments by Alstermark and Sasaki (1984b) indi- cate that specific subpopulations of C3-C4 PNs may distribute their excitation to particular forelimb motor- nuclei. The interesting question of how the C3-C4 pro- priospinal system combines several motornuclei for a move- ment asks for electrophysiological experiments with stimula- tion of the different collaterals of the same PN.
For mediation of inhibition the CST may use several parallel systems. In cats with an intact CST Illert et al. (1976a) described disynaptic latencies for pyramidally evoked IPSPs. They discussed that segmentally located Ib inhibitory interneurones could mediate this effect (Illert et al. 1976b). In cats with CST transections corticospinal in- hibition was mediated in a trisynaptic pathway, and it was postulated that the C3-C4 PNs act as first order, the segmentally located I a inhibitory interneurones as last order interneurones (Illert and Tanaka 1978). Alstermark et al. (1984) found evidence that inhibitory C3-C4 PNs can mediate disynaptic inhibition from the CST to triceps motoneurones. They suggested that this disynaptic pathway is used in parallel to the trisynaptic pathway previously
Fig. 8A--V. Facilitation from the pyramid of disynaptic PSPs evoked in motoneurones from group I muscle afferents. Three different motoneurones (A--J; K - M , Q - S ; N--P, T--V). Antidromic identification of the neurones in A, K, N. The pyramid was stimulated at the indicated strength in B - E , L, M, O, P (G--J, R, S, U, V display from the respective upper records the part of the traces at an expanded time scale which is indicated with a bar). In the ECU neurone of A--J (CST transection) stimulation of the ECR (F with 2 • T) was conditioned from the pyramid (alone in E and J), The a r r o w in G indicates arrival of the ECR volley at the spinal cord. In the EDC neurone of K--M, Q - S (CST transection) stimulation of the ECR (alone in Q with 1.6 • T) was conditioned from the pyramid (alone in M and S). In the EDC neurone of N--P, T--V (intact CST) stimulation of the ECR (alone in T) was conditioned from the pyramid (alone in P and V)

141
described. We have studied inhibition not to the same detail, and thus differences in the inhibitory linkage to particular muscles may have escaped our attention. Nevertheless, our material seems to indicate that also distal DR motornuclei can be disynaptically inhibited from the CST. Short latencies of 1.2 ms which would firmly establish a disynaptic projec- tion are obviously missing, but latencies around 1.5 ms are difficult to explain by a trisynaptic connection. Such values have been found in both, cats with and without CST transections. In the majority of the motoneurones inhibition had latencies longer than 1.8 ms and thus was in all likeli- hood trisynaptically evoked. A dominance of trisynaptic inhibitory connections to distal muscles seems not surprising to us since the functional relations between the muscles acting on the elbow are different from those acting on the wrist and hand. In the former case the synergistic-antagonis- tic relation is fixed, in the latter case it is changing according to the movement to be performed. This should give a segmental control of centrally evoked inhibition an import- ant role.
The pronounced facilitatory effect of conditioning pyra- midal stimulation on transmission in spinal reflex systems is well known (Baldissera et al. 1981). Some of our findings seem to be specific for the mechanisms coordinating the action of the distal forelimb muscles. Disynaptic exitatory and inhibitory cutaneous pathways to distal motornuclei have now been demonstrated. They may be correlated in some way with the versatile manipulatory repertoire of the forelimb. A strong and shaping influence of cutaneous afferents on forelimb motor mechanisms has been demon- strated (Marsden et al. 1972; Garnett and Stephens 1981). The described disynaptic connections could be the substrate for such effects, but at the present state with our only superfi- cial knowledge of these connections a detailed discussion is not possible.
We have not been able to ascribe the electrically evoked di- or polysynaptic group I PSPs to the Ia or Ib receptor system (see also Illert et al. 1976b). In addition, the recently found convergence of I a afferents onto "I b-interneurones" (Fetz et al. 1979) makes it questionable to treat both receptor systems as separate entities (Jankowska 1979; Jankowska and Lundberg 1981). With respect to disynaptic inhibition only effects which are relayed by I a inhibitory interneurones may be regarded with some certainty as being of Ia origin. It would therefore be important to test with recurrent depression from axon collaterals and Renshaw cells (Hultborn 1976) mediation of electrically evoked IPSPs via these interneurones, but we have to accept that this experimental approach is not yet routinely available in the forelimb. Facilitation of disynaptic group I IPSPs was very pronounced in pathways to motoneurones innervating distal muscles (see also Illert et al. 1976b). This became evident in the enhancement of transmission in pathways conducting in the control state and in the opening of additional lines. ParalM excitatory and inhibitory group I pathways to one motornucleus were a regular finding. This organization could reflect the varying functional synergism between the distal forelimb muscles and would, together with the differentiated excitatory monosynaptic Ia pathways (Fritz et al. 1984), give the brain the possibility to combine different muscles into a common functional group of synergists, according to the movement to be performed.
During this investigation we were struck by the similarity of pyramidal effects in cats with and without a segmental
input from the CST. This was observed not only for the effects of pyramidal stimulation in forelimb motoneurones, but also for the influence from the pyramid on transmission in spinal reflex systems. Sofar the only effect that can be ascribed to a direct projection from the CST on the spinal reflex apparatus (i. e. bypassing the C3-C4 propriospinal system) is the monosynaptic projection from the CST onto the "I b-interneurones" (Illert et al. 1976 b). It seems possible that in the electrophysiological experiments the pyramidal effects were largely mediated via the C3-C~ propriospinal system. Behavioural studies show that the direct projection of the CST into the segmental reflex apparatus is important during skilled forelimb movements (Alstermark et al. 1981). It will be one of the main tasks for future experiments to find the electrophysiological correlate of these "segmental" effects. Acknowledgements. We thank Drs. B. Alstermark and A. Lundberg for criticism of the manuscript. Excellent technical assistance was provided by Miss L. Schindler. This study was supported by the Deutsche Forschungsgemeinschaft.
References
Alstermark B (1983) Functional role of propriospinal neurones in the control of forelimb movements. A behavioural and electrophysiological study. Acta Physiol Scand (Sappl) (in press)
Alstermark B, Sasaki S (1984a) Integration in descending motor pathways controlling the forelimb in the cat. 13. Distribution of pyramidal effects in shoulder, elbow, wrist and digit motoneurones in the cat. Exp Brain Res (to be published)
Alstermark B, Sasaki S (1984b) Integration in descending motor pathways controlling the forelimb in the cat. 15. A quantitative comparison of the excitatory projection from C3-C4 pro- priospinal neurones to various forelimb motoneurones in the cat. Exp Brain Res (to be published)
Alstermark B, Lundberg A, Norssell U, Sybirska E (1981) Integra- tion in descending motor pathways controlling the forelimb in the cat. 9. Differential behavioural defects after spinal cord lesions interrupting defined pathways from higher centres to motoneurones. Exp Brain Res 42: 299- 318
Alstermark B, Lundberg A, Sasaki S (1984) Integration in descend- ing motor pathways controlling the forelimb in the cat. 10. Inhibitory pathways to forelimb motoneurones via C3-C4 pro- priospinal neurones. Exp Brain Res (to be published)
Baldissera F, Hultborn H, Illert M (1981) Integration in spinal neuronal systems. In: Brooks VB (ed) Handbook of physiology. The nervous system. Motor control, sect 2, vol 1, pt 1, chapt 12. American Physiological Society, Bethesda, MD, pp 509- 595
Crouch JE (1969) Atlas of cat anatomy. Lea and Febiger, Philadelphia
Fetz EE, Jankowska E, Johannisson T, Lipski J (1979) Autogenetic inhibition of motoneurones evoked by impulses in group Ia muscle spindle afferents. J Physiol 293:173 - 195
Fritz N, Illert M, Saggau P (1981) Location of dorsal interosseus motornuctei in the cat. Neurosci Lett 21:243--248
Fritz N, Illert M, Reeh P (1982) Location of median and ulnar motornuclei in the cat. Neurosci Lett 30:103-108
Fritz N, Illert M, Saggau P (1984) I a-synergism in the cat forelimb. I. Connections with muscles supplied by the deep radial nerve. Exp Brain Res (to be published)
Garnett R, Stephens JA (1981) Changes in the recruitment threshold of motor units produced by cutaneous stimulation in man. J Physiol 311:463-473
Grant G, Illert M, Tanaka R (1980) Integration in descending motor pathways controlling the forelimb in the cat. 6. Anatomical evidence consistent with the existence of C3-C4 propriospinal neurones projecting to forelimb motornuclei. Exp Brain Res 38:87-93

142
Grillner S, Hongo T (1972) Vestibulospinal effects on motoneurones and interneurones in the lumbosacral cord. In: Brodal A, Pompeiano O (eds) Basic aspects of central vestibular mechanisms. Progress in brain research, vol 37. Elsevier, Amsterdam, pp 243- 262
Hongo T, Jankowska E, Lundberg A (1969) The rubrospinal tract. I. Effects on alpha-motoneurones innervating hindlimb muscles in cat. Exp Brain Res 7:344--364
Hultborn H (1976) Transmission in the pathway of reciprocal Ia inhibition to motoneurones and its control during the tonic stretch reflex. In: Homma S (ed) Understanding the stretch reflex. Progress in brain research, vol 44. Elsevier, Amsterdam, pp 235-255
Illert M, Tanaka R (1976) Transmission of corticospinal IPSPs to cat forelimb motoneurones via high cervical propriospinal neurones and Ia inhibitory interneurones. Brain Res 103:143-- 146
Illert M, Tanaka R (1978) Integration in descending motor pathways controlling the forelimb in the cat. 4. Cortieospinal inhibition of forelimb motoneurones mediated by short pro- priospinal neurones. Exp Brain Res 31 : 131 -- 141
Illert M, Wiedemann E (1980) Distribution of pyramidal actions mediated by midcervical propriospinal neurones to identified radial motornuclei. Neurosci Lett (Suppl) 5:S 88
Illert M, Wiedemann E (1981) Pyramidal actions in identified radial motornuclei of the cat. Pfliigers Arch (Suppl) 389:R21
Illert M, Lundberg A, Tanaka R (1976 a) Integration in descending motor pathways controlling the forelimb in the cat. 1. Pyramidal effects on motoneurones. Exp Brain Res 26: 509- 519
IUert M, Lundberg A, Tanaka R (1976b) Integration in descending motor pathways controlling the forelimb in the cat. 2. Conver- gence on neurones mediating disynaptie cortico-motoneuronal excitation. Exp Brain Res 26: 521- 540
Illert M, Lundberg A, Tanaka R (1977) Integration in descending motor pathways controlling the forelimb in the cat. 3. Conver- gence on propriospinal neurones transmitting disynaptic excita- tion from the corticospinal tract and other descending tracts. Exp Brain Res 29:323-346
Illert M, Lundberg A, Padel Y, Tanaka R (1978) Integration in descending motor pathways controlling the forelimb in the cat. 5. Properties of and monosynaptic excitatory convergence on C3-C4 propriospinal neurones. Exp Brain Res 33:101 -- 130
Jankowska E (1979) New observations on neuronal organization of reflexes from tendon organ afferents and their relation to re- flexes evoked from muscle spindle afferents. In: Granit R, Pompeiano O (eds) Reflex control of posture and movement. Progress in brain research, vol 50. Elsevier, Amsterdam, pp 2 9 - 36
Jankowska E, Lundberg A (1981) Interneurones in the spinal cord. TINS 4:230-233
Lance JW (1954) Pyramidal tract in spinal cord of eat. J Neuro- physiol 17:253 -270
Laporte Y, Lloyd DPC (1952) Nature and significance of the reflex connections established by large afferent fibers of muscular origin. Am J Physiol 169:609-621
Laporte Y, Lundberg A, Oscarsson O (1956) Functional organiza- tion of the dorsal spino-eerebellar tract in the cat. I. Recording of mass discharge in dissected Flechsig's fasciculus. Acta Physiol Scand 36:175 -- 187
Lundberg A (1979) Integration in a propriospinal motor centre controlling the forelimb in the cat. In: Asanuma H, Wilson VJ (eds) Integration in the nervous system. Igaku-Shoin, Tokyo New York, pp 47-64
Lundberg A, Voorhoeve P (1962) Effects from the pyramidal tract on spinal reflex arcs. Acta Physiol Scand 56:201 - 219
Marsden CD, Merton PA, Morton HB (1972) Servo action in human voluntary movement. Nature (Lond) 238:140-- 143
Molenaar I (1978) The distribution of propriospinal neurons pro- jecting to different motoneuronal cell groups in the cat's brachial cord. Brain Res 158: 203 - 206
Tsukahara N, Fuller DRG, Brooks VB (1968) Collateral pyramidal influences on the eorticorubrospinal system. J Neurophysiol 31 : 467 - 484
Received December 14, 1983/Accepted February 1, 1984





























