Embed Size (px)
Citation preview

Bioscience Reports, Vol. 22, No. 1, February 2002 ( 2002)
MINI REVIEW
Rat Hindlimb Unloading: Soleus and Extensor DigitorumLongus Histochemistry, Mitochondrial DNA Contentand Mitochondrial DNA Deletions
V. Pesce,1 A. Cormio, F. Fracasso,1 A. M. S. Lezza,1, P. Cantatore1,2 andM. N. Gadaleta1,2,3
Receiûed Noûember 8, 2001
Mitochondrial phenotypic alterations, mitochondrial DNA content and mitochondrialDNA deletions in a slow, Soleus, and a fast, Extensor Digitorum Longus, skeletal muscleof 3- and 15-month-old hindlimb suspended rats have been studied. Cytochrome c oxidase-negative fibers appeared after unloading in all examined animals and their percentageincreased with increasing unloading time. After 14 days of suspension the mitochondrialDNA content did not change in 3-month-old but decreased significantly in 15-month-oldrats. Soleus was much more affected by unloading than Extensor Digitorum Longus. Themitochondrial DNA deletion of 4834 bp as well as other mtDNA deletions, researched withLong Distance-PCR, were absent in both studied muscles before and after unloading.
KEY WORDS: Mitochondrial DNA; COX-negative fibers; skeletal muscles; hindlimbunloading.
INTRODUCTION
It has been reported that, in both human and rodent, deficits of motor capacity,strength and endurance properties of skeletal muscle are induced by space flight(SF). The reduced muscle strength is associated, in part, with a reduction in musclemass as reflected in smaller cross-sectional areas of both fast- and slow-twitch fibers.It seems that slow-twitch fibers exposed to microgravity are more sensitive to theatrophying process than fast-twitch ones. Accompanying the atrophy is a transform-ation of the fibers phenotype from slow to fast involving myosin heavy chain andsarcoplasmic reticulum protein isoforms [1]. It is known that slow fibers are charac-terized by a higher number and more differentiated mitochondria than fast fibers[2, 3]. Mitochondria are the major source of energy in the cell because they containthe oxidative phosphorylation system (OXPHOS) responsible for the maximum out-put of ATP. Mitochondria contain their own DNA whose genes encode components
1Department of Biochemistry and Molecular Biology, University of Bari, via Orabona, 4, 70125, Bari,Italy.
2Center for the Study of Mitochondria and Energetic Metabolism, Bari, Italy.3To whom correspondence should be addressed. Fax: 39-080-5443403; email: [email protected]
115
0144-8463�02�0200-0115�0 2002 Plenum Publishing Corporation

116 Pesce et al.
of the OXPHOS system. The expression of mitochondrial DNA (mtDNA) is there-fore absolutely critical for the function of those tissues that are highly dependent onaerobic metabolism, such as skeletal muscle and heart [4]. Morphological and func-tional data were reported on rat and human skeletal muscle mitochondria in microg-ravity [3]. Oxidative enzymes were reported to be unaltered or slightly elevated bothin the slow antigravity muscle, Soleus, as in the fast muscle, Extensor DigitorumLongus (EDL) [2, 3]. However, a loss of oxidative enzymes and of mitochondriafrom the subsarcolemmal and intermyofibrillar regions of the fiber with SF in theSoleus was reported [5]. Microgravity significantly reduced (41%) cytochrome c oxi-dase (COX) activity in skeletal muscle triceps brachii although mRNA levels ofnuclear and mitochondrial encoded subunits remained unaltered [6]. A loss of mito-chondrial proteins with SF has been reported in rats, although at a lower degreethan that observed for the contractile proteins and for cell atrophy [3]. No data havebeen reported until now on mitochondrial biogenesis and on mtDNA content inmicrogravity.
In this paper we used histochemical and PCR techniques to follow phenotypicand genotypic alterations of mitochondria as well as mitochondrial DNA content ina slow, Soleus, and a fast, EDL, skeletal muscle of 3- and 15-month-old hindlimbrats suspended up to 14 days.
METHODS
Animal Groups
Rats were suspended by tail harnessing and housed separately in plastic boxcages. The temporal changes during suspension were studied by killing the animalsafter 3, 11 and 14 days of suspension. Suspended rats were fed Purina chow andwater ad libitum, and the control animals were pair fed.
Experimental rats were suspended by their tails using a tail harness that con-sisted of a triangular shaped wire (18 gauge) sandwiched between two layers of vinylcloth glued with Dural contact cement to the dorsal proximal four-fifths of the tail.The tail was washed, dried, and coated with cement before applying the cloth strips.The harness was further strengthened by loosely wrapping the tail with vinyl stripsand elastic tape (Medipore-3M). This method of harnessing distributed the loadalong the length of the tail, avoiding excessive tension on a small area. Rats weresuspended according to the non-invasive procedure of Morey-Holton [7]. Dailyhealth checks confirmed that the exposed tip of the tail remained pink, indicatingadequate blood flow. A fish swivel was attached to the exposed apex of the wiretriangle, and fishing line was tied to the swivel to elevate the rat’s hindquarters,unloading the hindlimbs. The height of suspension was adjusted so that the hindfeed just cleared the grid floor. The forelimbs maintained contact with the floor,thus allowing the animals access to food and water.
Histochemistry
Serial eight-microns-thick transverse sections from frozen muscle biopsies werecut with the cryotome (HM 505 E-Microm), mounted on polylysine-coated glass

MtDNA in Rat Skeletal Muscle 117
slides and stained for COX activity [8], for SDH activity [9] and for both activities.250 to 500 fibers for each muscle sample were analyzed. Fibers with abnormal accu-mulation of mitochondria, the so called ‘‘ragged red fibers’’ (RRF) for their redstaining in the subsarcolemmal region with the modified Gomori trichrome stain,appeared hyperreactive with the modified SDH stain. The modified SDH reactionwas chosen to count the number of RRF because it was more sensitive than themodified Gomori trichrome staining [10]. Sections were viewed on a Zeiss trans-mitted light microscope and photographed using a 10× objective. In each specimenthe total number of fibers in a representative selected field was counted on a photo-micrograph and the percentage of those having no detectable COX activity (COX-negative), normal COX activity (COX-positive), normal SDH activity (SDHn) andhyperreactive SDH activity (RRF) was calculated.
Determination of mtDNA Content in Skeletal Muscle Biopsies
Total DNA was prepared from about 40–50 mg of skeletal muscle as describedby Arnaudo et al. [11] and suspended in 30 µl of water. 5 µg of total DNA weredigested with Pvu II (MBI-Fermentas) and run on a 0.35% agarose gel (SeakemGold, FMC). The gel was blotted onto a Hybond-N membrane (Amersham-Pharm-acia) and simultaneously hybridized with a mitochondrial and a nuclear probe. Themitochondrial probe was a 657 bp fragment obtained by PCR using the primers DN(L 15758–15777) and DR (G 117–98). PCR reaction contained 0.1 µM of eachprimer, 100 ng DNA, 200 µM dNTPs, 2.5 U Taq Polymerase (Roche Mol. Biochem.,Germany), 1.5 mM MgCl2 in a 100 µl volume of 1× reaction buffer. PCR conditionswere: denaturation of 94°C for 1 min, annealing at 51°C for 1 min and extension at72°C for 1 min for 25 cycles. The nuclear probe was a 413 bp fragment containingpart of the 18S rRNA gene and subcloned in the TA vector (Clontech). Both probeswere labeled by random priming (Random Primed DNA Labeling Kit, Roche Mol.Biochem., Germany) and used in a 10:1 ratio of the nuclear DNA (nDNA) probeto the mtDNA probe. Blotting, prehybridization, hybridization and washings werecarried out as described by Sambrook et al. [12]. The filter was exposed to a X-rayfilm at −70°C with an intensifying screen and the hybridization signals were quant-ified by densitometry with a LKB-Pharmacia Ultrascan-XL Laser densitometerequipped with a GelScan-LX-Evaluation software.
Detection of Mitochondrial DNA Deletion of 4834 bp (mtDNA4834)
About 50 mg of skeletal muscle were used to extract nucleic acids enriched formtDNA [13]. The DNA pellet was suspended in 50 µl of water and stored at 4°C.100 ng of DNA were amplified as described elsewhere [14] by using the followingprimers: 7825–For (L 7825-7844) and 13117 Rev (H 13117-13099) for mtDNA4834
(460 bp product, annealing temperature 60°C). The initial PCR reaction contained0.1 µM of each primer, 200 µM dNTPs, 100 ng DNA and 2.5 U Taq polymerase(Roche Mol. Biochem., Germany) in a 100 µl volume of 1Breaction buffer. ThePCR conditions consisted of denaturation at 94°C for 1 min, annealing at 60°C for

118 Pesce et al.
1 min and extension at 72°C for 1 min in a Peltier Thermal Cycler (MJ Research)for 35 cycles.
A secondary seminested PCR round, using one of the primers of the first ampli-fication (13117 Rev) and a new internal primer 7978 For (L 7978–7997) with a 1 µlaliquot of the first amplification mixture diluted 1:100 at an annealing temperatureof 60°C, was performed to obtain a 309 bp product. PCR products were size-frac-tionated on a 1.5% agarose gel (Amresco) 1BTBE buffer and visualized by ethidiumbromide staining. Only those subjects not showing the expected amplification prod-ucts in two successive PCR rounds were considered not containing the searchedmtDNA deletions. Rat mtDNA nucleotide positions are according to Gadaleta etal. [15]. The identity of the amplification products was confirmed by direct DNAsequencing as described elsewhere [16].
Long Distance-PCR
Primers used were 5881-For (L 5881–5906) and 15141-Rev (H 15141–15118).Gene Amp carry-over prevention kit (Perkin Elmer–Norwalk) was used. The PCRmixture contained 100 ng of DNA, 0.1 µM of each primer, 200 µM dNTP (withdUTP instead of dTTP), 2.5 mM MgCl2 and 2.5 U of Long Amplification Taq Poly-merase (Takara—Japan) in a 100 µl reaction. PCR products were separated on a1.5% agarose gel (Amresco—OH, USA) in 1BTBE buffer and visualized by ethid-ium bromide staining. To exclude that the low molecular weight bands were due toPCR artefacts, a secondary seminested LD-PCR round, very similar to the first, wasperformed on an aliquot of the first reaction mixture added after a 2 min at 50°Cand 10 min at 94°C step, employing the 15118-Rev primer and a new internal primer5974-For (L 5974–5996). In all analyzed cases we obtained a pattern consistent withthat observed in the first PCR round (data not shown).
Statistics
Statistical analysis was carried out by using StatView SECversion 1.03 software(Abacus Concepts, Berkeley, CA).
RESULTS
Atrophy
Suspension caused a progressive loss of mass of the two studied muscles (wetwt). In Fig. 1A the loss of mass of the Soleus and EDL in 15-month-old animals isreported. The loss of mass was 39% in Soleus and 27% in EDL. In Fig. 1B the ratioof muscle weight (mg) to total body weight (g) is reported. The values of this ratiowere for the Soleus and the EDL muscles of 15-month-old animals, respectively,0.45J0.02 (mean JSD) and 0.51J0.02 (mean JSD) in control rats and 0.34J0.2(mean JSD) and 0.45J0.02 (mean JSD) after 14 days of unloading. Therefore,the Soleus lost 24% and the EDL 12% of their mass after 14 days of unloading.
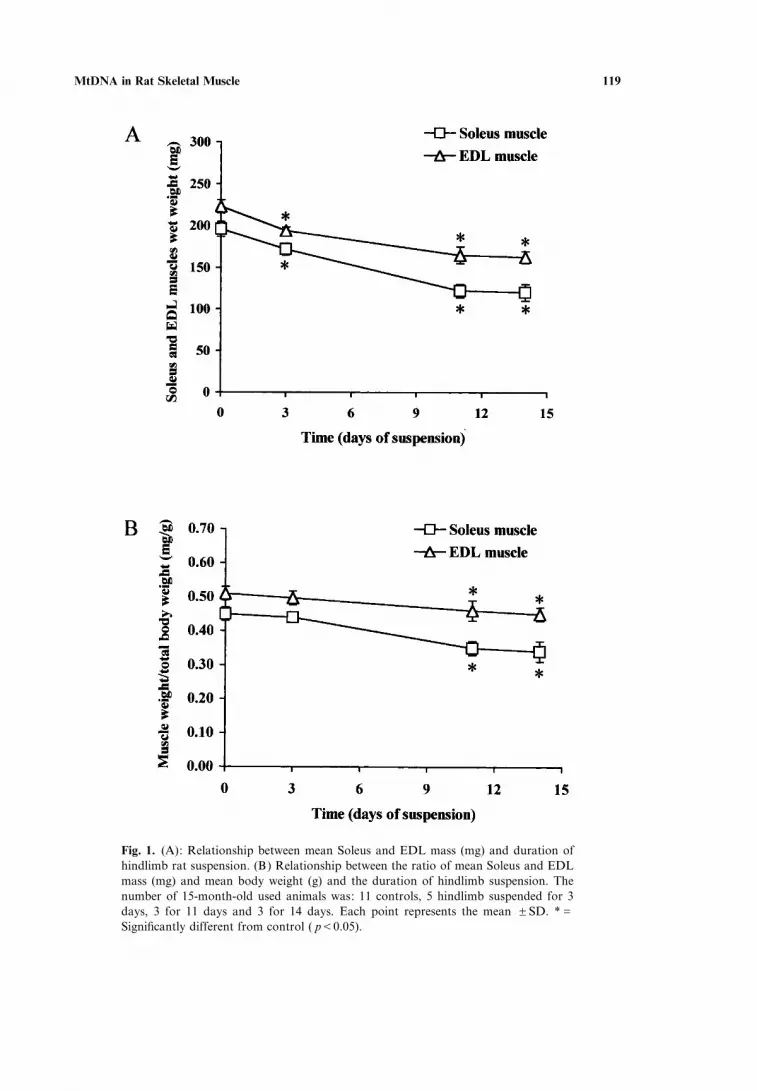
MtDNA in Rat Skeletal Muscle 119
Fig. 1. (A): Relationship between mean Soleus and EDL mass (mg) and duration ofhindlimb rat suspension. (B) Relationship between the ratio of mean Soleus and EDLmass (mg) and mean body weight (g) and the duration of hindlimb suspension. Thenumber of 15-month-old used animals was: 11 controls, 5 hindlimb suspended for 3days, 3 for 11 days and 3 for 14 days. Each point represents the mean JSD. *GSignificantly different from control ( pF0.05).

120 Pesce et al.
Histochemistry
Staining for COX and SDH activities to identify COX-negative fibers and RRFas well as double staining to distinguish two subtypes of RRF, COX-positive RRFand COX-negative RRF, in 3- and 15-month-old rats were performed on Soleus andEDL skeletal muscles. COX-negative fibers were absent in 3-month-old and presentin 50% of 15-month-old control rats whereas, after unloading, they appeared in allexamined animals. In Fig. 2 the mean percentage of COX-negative fibers, respect-ively, in Soleus and EDL skeletal muscles of 3-month-old rats, unloaded for 14 days,is reported. After 14 days hindlimb suspension of 3-month-old rats, 0.4% COX-negative fibers in Soleus muscle and 0.2% in EDL, respectively, have been found. InFig. 3 the mean percentage of COX-negative fibers, respectively, in Soleus and EDLskeletal muscles of 15-month-old rats, unloaded for 3, 11 and 14 days, is reported.COX-negative fibers appeared 3 days after suspension in both muscles of all ana-lyzed rats and their percentage increased after 11 days of suspension. After 14 daysof suspension the Soleus muscle showed fibers disorganization whereas a three-foldincrease of the COX-negative fibers percentage was found in EDL muscle. RRFwere not found in 3- and 15-month-old control and unloaded rats.
Mitochondrial DNA Content
In order to investigate whether a variation of mtDNA copy number occurredin the two rat skeletal muscles after unloading, we measured the mtDNA�nDNAratio in 3-month-old and 15-month-old rats, by hybridizing Pvu II-digested totalDNA extracted from Soleus and from EDL muscles with a mitochondrial andnuclear probe. The mitochondrial probe detected a band of 16.2 kbp correspondingto linearized mtDNA, whereas the nuclear probe detected a 12 kbp band, corre-sponding to a Pvu II fragment of the nuclear 18S rDNA gene (data not shown). Theratio of the intensities of the two bands was used to estimate the relative amount ofmtDNA in each subject. No change of the mtDNA�nDNA ratio was found in 3
Fig. 2. Percentage of COX-negative fibers in the skeletal muscle of 3-month-old rats after 14 days ofunloading. Three controls and 3 unloaded rats were used. Values represent the mean JSE of the percent-ages of COX-negative fibers.

MtDNA in Rat Skeletal Muscle 121
Fig. 3. Percentage of COX-negative fibers in the skeletal muscle of 15-month-old control rats and ratsafter 3, 11 and 14 days of unloading. The number of animals used was the same as in Fig. 1. #GMorphological disorganization of the skeletal muscle. Values represent the mean JSE of the percentagesof COX-negative fibers, calculated by excluding, in the controls, animals without COX-negative fibers.Significant differences in the percentage of COX-negative fibers (Student’s t-test) were found for thecomparisons between the values of control rats and rats after 11 days of unloading for Soleus and betweenthose of control rats and rats after 14 days of unloading for EDL ( pF0.05).
months old rats after 14 days of suspension (data not reported). However, themtDNA content increased after 3 days of unloading in the Soleus muscle of 15-month-old rats and then significantly decreased reaching 40% of the control valueafter 14 days of suspension (Fig. 4A). The mtDNA content of the EDL muscle wasalso reduced after 14 days of unloading although at a lower extent (Fig. 4B).
MtDNA Deletions
Soleus and EDL muscle samples were screened by two PCR rounds with nestedprimers for the presence of the mtDNA4834 in 3- and 15-month-old rats. Althoughit was found in 28-month-old rats (A. Cormio, personal communication), suchdeletion was absent both in 3- and 15-month-old control and unloaded animals herestudied (results not shown). In order to obtain a more comprehensive representationof mtDNA deletions in each animal we used LD-PCR. With this technique, whichrevealed in 28-month-old rats 4 to 8 bands on the gel, corresponding to subgenomicmtDNA particles (A. Cormio, personal communication), no mtDNA deletions atall were found here in 3- and 15-month-old rats, both before and after unloading(results not shown).
DISCUSSION
Mitochondria play a central role in processes such as stress response, metabolicremodeling, apoptosis and aging [17]. After SF some endocrine axes seem to undergochanges resembling those generally described during aging [18, 19]. We reportedrecently the results of an extensive analysis of age-related mitochondrial phenotypicand genotypic alterations in a large number of skeletal muscle biopsies from healthy

122 Pesce et al.
Fig. 4. Relative mitochondrial DNA content in Soleus and EDL skel-etal muscles of 15-month-old control rats and rats after 3, 11 and 14days of hindlimb suspension. The relative mtDNA content wasobtained from the ratio of signals intensities corresponding to mtDNAand nDNA bands and determined in 11 control rats, 5 rats after 3days, 3 after 11 days and 3 after 14 days of hindlimb suspension. Therelative mtDNA content was normalized with respect to the controlrats value. Data are the mean JSD of two experiments. *GStatisti-cally significant decrease of mtDNA content in Soleus muscle after 14days of hindlimb suspension.

MtDNA in Rat Skeletal Muscle 123
human individuals. We were able to show that, in individuals older than 50 years ofage, the appearance of COX-positive RRF and, thereafter, of COX-negative RRF,the increase of the mtDNA4977 and the increase of mtDNA deleted species wereaccompanied by the increase of mtDNA content as an attempt of the cell to counter-act the age-related mitochondrial dysfunction [14]. Consistent with such a compensa-tory response was, after 50 years of age, the increased level of nuclear respiratoryfactor-1 (NRF-1) and mitochondrial transcription factor A (Tfam) [20]. Theseresults prompted us to verify in two rat skeletal muscles, known to have a differentfibers composition, namely Soleus and EDL, and in microgravity conditions, knownto induce atrophy of slow fibers and�or transformation of slow to fast-twitch fibers[1–3], the involvement of mitochondria and of their genetic system.
Here we report, for the first time, that microgravity induces the appearance ofCOX-negative fibers in Soleus and EDL skeletal muscles of young and adult rats anda depletion of mtDNA in adult rats. Moreover, no RRF and no mtDNA deletions inboth groups of rats, before and after unloading, were found.
A 41% reduction of COX activity was already reported in the skeletal muscletriceps brachii after 6 days of microgravity with no change of mRNA for somemtDNA and nDNA-encoded COX subunits. The Authors suggested that proteindegradation was responsible for the reduced COX activity measured [6]. Theseresults were obtained by measuring COX activity in tissue homogenate so that if theCOX deficiency was equally distributed in all fibers was not clear. The histochemicalresults here reported show that the deficiency of COX activity affects in a mosaic-like fashion both the Soleus and EDL muscles of all young and adult examined ratsafter 3, 11 and 14 days of unloading. These data, together with the high degree ofmtDNA depletion after 14 days of unloading in adult rats, might suggest that theCOX deficiency is due to a selective mtDNA depletion in affected fibers. However,the values of relative mtDNA content here reported have been obtained from wholetissues extracts so that it is impossible to specify if unloading affects the relativemtDNA content (a) of all fibers of the tissue, (b) of COX-negative fibers only, (c)of a higher number of fibers than that reported in (b) i.e. fibers with defective Com-plex I that, in our case, might appear as COX-positive. Complex I defects have beenreported in unloading rats [21]. In fact, in the case of mtDNA depletion all com-plexes of the OXPHOS system, except Complex II, which has only nDNA-encodedpolypeptides, should be affected. Therefore, the COX-negative fibers found in theskeletal muscles here analyzed might be fibers defective in Complex IV or fibers inwhich all the OXPHOS system is altered. Reduced mtDNA availability has beenreported to be a limiting factor for mitochondrial gene expression in type II but notin type I fibers [22, 23]. On the other hand, an increased oxidative capacity, followingexercise training or chronic electrical stimulation [22, 23], was primarily associated,at molecular level, with increased mitochondrial DNA content in glycolytic type IIfibers and with increased mitochondrial transcripts in highly oxidative type I musclefibers [24]. Unstimulated fibers, due to hindlimb suspension of rat, might lose signalsinducing mtDNA replication and mitochondrial biogenesis and�or receive apoptoticsignals.
No fibers with hyperproliferation of mitochondria (RRF) have been found inSoleus and EDL muscles before and after unloading. The absence of mtDNA

124 Pesce et al.
deletions in the presence of COX-negative fibers reported in unloading rats suggeststhat mtDNA replication is impaired. This is in agreement with the decrease ofmtDNA content after loading here reported and with the results already reportedby Riley et al. [5] who found that in Soleus, after 14 days of unloading, SDH activity,measured as marker of the respiratory chain activity and presence of mitochondria,was reduced and type I and IIa fibers decreased in area of 63% and 43%, respectively;both subsarcolemmal and intermyofibrillar mitochondria were degraded, with thefirst ones being degraded more rapidly. The same Authors reported a much lesssevere atrophy in EDL [5], in good agreement with the data here reported.
Reduced ATP availability, because of OXPHOS system dysfunction, can inter-fere with nuclear gene expression and cause a decreased synthesis of myofibrillarproteins leading to a decrease in sarcomers number resulting in fiber atrophy[19, 25, 26]. An increased degradation of myonuclei has been reported in Soleus afterunloading suggesting the activation of cellular apoptotic pathways. Apoptosis hasalso been hypothesized as a mechanism contributing to remodeling of skeletal musclein response to hindlimb unweighting [27]. More detailed analyzes at molecular leveland shorter times treatments are necessary to understand which is the mitochondrialrole in triggering the fiber type change and�or the atrophy in skeletal musclesexposed to microgravity [28].
ACKNOWLEDGMENTS
We are grateful to Dr. Anna De Marzo and Adriana Di Benedetto for technicalassistance and Ms. R. Longo for word processing. This work was supported byItalian Space Agency (ASI) grants, Contract No. I�R.168�00 and Sigma Tay—Indu-strie Farmaceutiche Riunite S.p.A.
REFERENCES
1. Baldwin, K. M. (1996) Effect of spaceflight on the functional, biochemical, and metabolic propertiesof skeletal muscle. Med. Sci. Sports Exerc. 28:983–987.
2. Edgerton, V. R. and Roy, R. R. (1996) Neuromuscular adaptations to actual and simulated spa-ceflight. In: Handbook of Physiology, Enûironmental Physiology. Bethesda, MD: Am. Physiol. Soc.,sec. 4, Vol. II, 32:721–763.
3. Fitts, R. H., Riley, D. R., and Widrick, J. J. (2000) Microgravity and skeletal muscle. J. Appl. Physiol.89:823–839.
4. Wallace, D. C. (1999) Mitochondrial diseases in man and mouse. Science 283:1482–1488.5. Riley, D. A., Slocum, G. R., Bain, J. L. W., Sedlak, F. R., Sowa, T. E., and Mellender, J. W.
(1990) Rat hindlimb unloading: soleus histochemistry, ultrastructure, and electromyography. J. Appl.Physiol. 69:58–66.
6. Connor, M. K. and Hood, D. A. (1998) Effect of microgravity on the expression of mitochondrialenzymes in rat cardiac and skeletal muscles. J. Appl. Physiol. 84:593–598.
7. Morey-Holton, E. R. and Globus, R. K. (1998) Hindlimb unloading of growing rats: a model forpredicting skeletal changes during space flight. Bone 22:83S–88S.
8. Seligman, A. M., Karnovsky, M. J., Wasserkrug, H. L., and Hanker, J. W. V. (1968) Non-dropletultrastructural demonstration of cytochrome c oxidase activity with a polymerising osmiophilicreagent diaminobenzidine (DAB). J. Cell. Biol. 38:1–14.
9. Dubowitz, U. K. and Brooke, M. H. eds. (1973) Muscle Biopsy: A Modern Approach. Saunders,W.B.: Philadelphia, PA, USA.

MtDNA in Rat Skeletal Muscle 125
10. Rifai, Z., Welle, S., Kamp, C., and Thornton, C. A. (1995) Ragged red fibers in normal aging andinflammatory myopathy. Ann. Neurol. 37:24–29.
11. Arnaudo, E., Dalakas, M., Shanske, S., Moraes, C. T., DiMauro, S., and Schon, E. A. (1991)Depletion of muscle mitochondrial DNA in AIDS patients with zidovudine-induced myopathy. Lan-cet 337:508–510.
12. Molecular Cloning: A Laboratory Manual, 2nd edn. (1989) (J. Sambrook, E. F. Fritsch, and T. Mani-atis, eds.) Cold Spring Harbor, N.Y., USA: Cold Spring Harbor Laboratory Press.
13. Aspnes, L. E., Lee, C. M., Weindruch, R., Chung, S. S., Roecker, E. R., and Aiken, J. M. (1997)Caloric restriction reduces fiber loss and mitochondrial abnormalities in aged rat muscle. FASEB J.11:573–581.
14. Pesce, V., et al. (2001) Age-related mitochondrial genotypic and phenotypic alterations in humanskeletal muscle. Free Radic. Biol. Med. 30:1223–1233.
15. Gadaleta, G., Pepe, G., De Candia, G., Quagliariello, C., Sbisa, E., and Saccone, C. (1989) Thecomplete nucleotide sequence of the Rattus norvegicus mitochondrial genome: cryptic signalsrevealed by comparative analysis between vertebrates. J. Mol. Eûol. 28:497–516.
16. Gadaleta, M. N., Rainaldi, G., Lezza, A. M. S., Milella, F., Fracasso, F., and Cantatore, P. (1992)Mitochondrial DNA copy number and mitochondrial DNA deletion in adult and senescent rat.Mutat. Res. 275:181–193.
17. Salvioli, S., Bonafe, M., Capri, M., Monti, D., and Franceschi, C. (2001) Mitochondria, aging andlongevity—a new perspective. FEBS Lett. 492:9–13.
18. Grigoriev, A. I. and Egorov, A. D. (1993) Physiological aspects of adaptation of main body systemsduring and after space flight. Adû. Space Biol. Med., Vol. 2 (S. L. Bonting, ed.) 43–82, JAI PressInc., London, UK.
19. Scano, A. and Strollo, F. (1997) Life sciences experiments in space bring benefits on earth. (ESA BR-119), ESTEC Publ. Div. Noordwjik, NL.
20. Lezza, A. M. S. et al. (2001) Increased expression of mitochondrial transcription factor A and nuclearrespiratory factor-1 in skeletal muscle from aged human subjects. FEBS Lett. 501:74–78.
21. Yajid, F., Mercier, J. G., Mercier, B. M., Dubouchaud, H., and Prefaut, C. (1998) Effects of 4 wkof hindlimb suspension on skeletal muscle mitochondrial respiration in rats. J. Appl. Physiol. 84:479–485.
22. Williams, R. S. (1986) Mitochondrial gene expression in mammalian striated muscle. Evidence thatvariation in gene dosage is the major regulatory event. J. Biol. Chem. 261:12390–12394.
23. Williams, R. S., Garcia-Moll, M., Mellor, J., Salmons, S., and Harlan, W. (1987) Adaptation ofskeletal muscle to increased contractile activity. Expression of nuclear genes encoding mitochondrialproteins. J. Biol. Chem. 262:2764–2767.
24. Armstrong, R. B. and Phelps, R. O. (1984) Muscle fiber type composition of the rat hindlimb. Am.J. Anat. 171:259–272.
25. Wanagat, J., Cao, Z., Pathare, P. and Aiken, J. M. (2001) Mitochondrial DNA deletion mutationscolocalize with segmental electron transport system abnormalities, muscle fiber atrophy, fiber split-ting, and oxidative damage in sarcopenia. FASEB J. 15:322–332.
26. Richter, C., Schewizer, M., Cossarizza, A., and Franceschi, C. (1996) Control of apoptosis by thecellular ATP level. FEBS Lett. 378:107–110.
27. Allen, D. et al. (1997) Apoptosis: a mechanism contributing to remodeling of skeletal muscle inresponse to hindlimb unweighting. Am. J. Physiol. 273:C579–C587.
28. Moraes, C. T. (2001) What regulates mitochondrial DNA copy number in animal cells? Trends Genet.17:199–205.





























