Embed Size (px)
Citation preview

Dermatol Clin 25 (2007) 577–588
Receptors of Eccrine, Apocrine, and HolocrineSkin Glands
Uwe Wollina, MDa,*, Mohamed Badawy Abdel-Naser, MDb,Ruta Ganceviciene, MDc,d, Christos C. Zouboulis, MDc,e
aDepartment of Dermatology and Allergology, Academic Teaching Hospital Dresden-Friedrichstadt,
Friedrichstrasse 41, 01067 Dresden, GermanybFaculty of Medicine, Ain Shams University Hospital, 4 Al Rahman Tower 11281, El Sawah Square, Cairo, Egypt
cLaboratory for Biogerontology, Dermato-Pharmacology and Dermato-Endocrinology,
Institute of Clinical Pharmacology and Toxicology, Charite Universitaetsmedizin Berlin,
Campus Benjamin Franklin, 12200 Berlin, GermanydDepartment of Dermatology, Vilnius University Hospital, Santariskiu Klinikos, Vilnius, Lithuania
eDepartment of Dermatology, Venereology, and Allergology, Dessau Medical Center, 06847 Dessau, Germany
Eccrine, apocrine, and holocrine glands belongto the group of specialized miniorgans of skinknown as skin appendages. Eccrine glands de-
velop from the sweat gland anlagen in thepalmoplantar skin between 14 to 16 weeks’estimated gestational age (EGA). Canalization
of the dermal component (ie, coils and dermalduct) is complete by 16 weeks’ EGA. Canalizationof the epidermal duct or acrosyringium is notcomplete until 22 weeks’ EGA. The specific end
product of eccrine glands is sweat. They do notfunction in utero but start functioning immedi-ately after birth.
Apocrine glands usually arise from the upperportion of hair follicles not before the fifth monthof EGA. They function transiently during the
third trimester and become quiescent in theneonate. Their function reappears during puberty.Their secretions play a role in sexuality. Apocrine
glands, such as Moll’s glands of the eyelid,contribute to ocular host defense. The largestapocrine gland, however, is the breast gland,which is not discussed in this article.
The development of holocrine sebaceous glandsis associated closely with the formation of hairfollicles during embryonic life. They rest until
puberty. Their specific end product is sebum [1,2].
* Corresponding author.
E-mail address: [email protected] (U. Wollina).
0733-8635/07/$ - see front matter � 2007 Elsevier Inc. All
doi:10.1016/j.det.2007.06.001
Sex steroid receptors
Sex-steroid receptors are distributed widelyamong human tissues. As an example of their ac-
tion, the estrogen receptors (ERs) are discussed inmore detail.
Estrogens effects are genomic or nongenomic.
Estrogens mediate their genomic activity by in-teraction with and activation of specific nuclearERs. The ER-ligand complex binds to specificDNA sequences within the regulatory region of
the target genes. By interaction with other cellularcomponents, these target genes are activated orsuppressed. This is known as the classical path-
way. The nonclassical pathway of genomic actionalso can be mediated by interactions with ERs, inthis case with membrane-bound ERs localized in
the caveolae. Another option is the binding toG-protein–coupled receptor. This interaction ac-tivates kinases that act on nuclear transcription
factors (ie, nongenomic) [3,4].Two isoforms of ERs are known (ie, ER-a and
ER-b). Whereas a large number of natural andsynthetic estrogens show a similar binding affinity
to both receptors, other ligands are more selective,such as genistein or daidzein, which activateER-b much more strongly. ER immunostaining
usually is seen in a nuclear pattern [5].ER-a is expressed exclusively in basal and
partially differentiated cells of normal sebaceous
glands but not in sebaceous tumors. ER-a is seen in
rights reserved.
derm.theclinics.com

578 WOLLINA et al
only a small portion of the eccrine gland epitheliawith a weak cytoplasmic staining. ER-b is ex-pressed by sebaceous glands and cultured human
sebocytes in the same distribution as ER-a [6]. Hu-man eccrine sweat glands express ER-b. In mousesebaceous glands there is a gender difference inthe expression of ERs and androgen receptors
(ARs) [7] but in human skin there is no gender dif-ference in the pattern or distribution of ERs [8].Treatment of gonadectomizedmalemicewith pros-
taglandin E2 leads to expression of ER-a [7].ER-b expression is diminished in many human
eccrine neoplasms [9,10]. A case of a 45-year-old
woman who had a signet cell carcinoma of the ec-crine sweat gland of the eyelid, however, showedpositivity for ER and progesterone receptor(PR) [11]. Cases of ER-positive malignant eccrine
spiradenoma also are described [12]. In the axil-lary apocrine glands, a strong nuclear expressionof ER-a is seen in the secretory epithelium [13].
Sebaceous glands belong to the peripheralintracrine tissues in which estrogens are formedby transformation of prohoromone, dehydroe-
piandrosterone (DHEA), locally. There is noDHEA receptor in human tissue but DHEA isa major source of estrogen in women. Oral DHEA
supplementation in postmenopausal women whohave mostly sebostatic skin is capable of increas-ing sebum production by almost 80% after 12months. Apart from this, other beneficial cutane-
ous effects are documented, such as improved skinhydration, reduced facial pigmentation, and in-creased skin thickness [14,15].
PRs a and b have been identified in humanskin. In normal sebaceous glands, PRa and PRbare expressed by basal and differentiated sebo-
cytes [9]. PR expression has been observed in sy-ringoma, suggesting the possibility of hormonalcontrol of this benign eccrine tumor [16,17].
ARs are expressed strongly by all sebocytes, in
eccrine and apocrine sweat glands [18]. In seba-ceous glands, undifferentiated basal sebocytes ex-press AR in higher quantity than differentiated
sebocytes [19]. In eccrine and apocrine sweatglands, luminal cells of the secretory portion arevariably AR reactive [20]. In male mice, but not
in female animals, gonadectomy leads to a diminu-tion of the AR expression by sebaceous glands [7].
In benign eccrine tumors, the expression of AR
is diminished [9]. In apocrine carcinoma, AR andAR messenger RNA (mRNA) may be absent [21].ARs are expressed by sebaceous and sweat glandswithin nevus sebaceous of Jadassohn [22]. Papil-
lary hidradenomas may overexpress AR [23].
In addition, AR is expressed by the secretoryepithelium of axillary apocrine glands in correla-tion to their secretory activity [13]. An exception
to this rule is the apocrine gland of the eyelid (ie,Moll’s gland). Not all primates express ARs inthis gland [24]. In patients who have nonautoim-mune dry eyes with meibomian gland dysfunc-
tion, the level of bioavailable testosterone isreduced significantly [25].
The available data suggest that all adnexal
glands of human skin interact with sex steroids.This can be of developmental importance, such asfor sebaceous glands during puberty, or of regu-
latory importance, such as in apocrine glands,where phermones are produced under the influ-ence of androgens [2,13].
Extramammary Paget’s disease is an uncom-
mon neoplasm of the skin with apocrine differen-tiation. Eighty percent of these tumors and manyof the lymph node metastases are AR positive
[26]. These tumor cells are characterized, however,by a consistent lack of ERs and PRs [27].
In the case of acne, the adenarche (ie, the
starting point of endogenous androgen produc-tion) sebaceous glands increase their volume andtheir sebum production [2,28]. A major stimulus
for this is the DHEA, later on testosterone playsa major role. There is no acne without androgens.In women who have acne, the androgen effectscan be balanced with lower doses of estrogen com-
bined with progestins, such as levonogestrel, deso-gestrel, norgestinate, gestodene, cyproteroneacetate, chlormadinone acetate, and drospirenone
[29]. Studies of oral contraceptives show them tobe efficient and safe in women, reducing inflam-matory and noninflammatory lesions [29,30].
In therapeutically resistant cystic acne of menand women, an androgen excess occurs. DHEAsulfate and 17-0H-progesterone levels are higherand sex-hormone–binding globulin is lower in
affected subjects compared with controls.Therefore, low-dose dexamethasone can be usedfor 6 months to decrease DHEA sulfate [31]. In
hyperandrogenism, with the triade of acne, alope-cia, and hypertrichosis, contraceptives and antian-drogens are the cornerstone of treatment [32].
In ER-positive sweat gland adenocarcinoma,the use of hormone-blocking agents, such astamoxifen, has been discussed for the treatment
of metastasizing disease [12,33,34].Luteinizing hormone/human chorionic gonad-
otrophin receptor (LH/hCG-R) was detected inskin by immunohistochemistry and reverse tran-
scription polymerase chain reaction. In eccrine

579RECEPTORS OF SKIN GLANDS
sweat glands, LH/hCG-R is localized in clearcells that are involved in fluid and ion transport.Changes in sweating during puberty andmenopause might be related to changes in LH
levels. In the secretory coils, cells adjacent to thebasement membrane were stained with mono-clonal antibody against LH/hCG-R. In seba-
ceous glands, thin cells with round nuclei closeto the duct were stained intensively, whereaspolyhedral sebocytes with fat droplets showed
a faint staining [35].
Mineralocorticoid and glucocorticoid receptors
Mineralocorticoid receptor (MCR) mediates
aldosterone actions on salt and water balance inrestricted target cells. MCR is a member of thenuclear receptor superfamily and acts as a ligand-induced transcription factor modulating the
expression of specific proteins involved in physi-ologic responses to aldosterone [36]. Two humanMCR isoforms are identified (ie, a and b). Bothisoforms are coexpressed in sweat gland ducts.In patients who have abnormal mineralocorticoidstatus, the expression may be compromised. In
primary hyperaldosteronism and Liddle syn-drome, MCRb but not MCRa expression in sweatglands is reduced markedly [37].
The enzyme, 11-b-hydroxysteroid dehydroge-nase type II, is shown to confer specificity onMCR by inactivating glucocorticoids. Colocaliza-tion of the enzyme and the MCR is seen in
a majority of sodium-transporting epithelia. Inskin, MCR is expressed in basal cell of the sweatducts but not in apocrine or sebaceous glands.
The dehydrogenase is localized in luminal epithe-lia of the eccrine duct [38].
Like ARs or PRs, unliganded glucocorticoid
receptors (GCRs) are located in the cytoplasmattached to heat shock proteins (hsp 90, 70, and56). When hormones bind to GCRs, hsps are
released and, through an energy-dependent process,the hormone receptor complexes form homo- orheterodimers. The zinc fingers of their DNAdomains slot into the glucocorticoid response
elements in the DNA helix where they exertactivation of suppression of protein synthesis. Inaddition, nongenomic effects of glucocorticoids
are mediated by interaction with surface proteinsand lipids [39].
GCRs immunoreactivity is found in sweat
gland and sebaceous gland epithelia [40]. Thisprobably explains the induction of steroid-induced acne.
Orphan nuclear hormone receptors
Nuclear hormone receptors are important inthe regulation of epithelial cell differentiation andlipid metabolism. Different receptors are identified
in human skin. Some of them play a crucial role inlipid metabolism of sebaceous glands. In seba-ceous glands maintained ex vivo and in human
sebocyte cultures, peroxisome proliferation-activated receptors (PPARs) (types a, d, and g),retinoid X receptor (RXR)-a, liver X receptor-a,and pregnane X receptor but not farnesoid Xreceptor could be detected (Fig. 1) [41–43]. AllPPARs subtypes also are expressed in sebaceous
glands in vivo. Furthermore, DAX1 and SF1 areexpressed by sebaceous glands and cultured hu-man sebocytes and eccrine sweat glands in humanskin [44,45].
PPARa and PPARg ligands inhibited the lipidsynthesis in sebaceous glands maintained ex vivo[42] but stimulated lipogenesis in diabetic patients
in vivo [46]. In agreement with the in vivodata, natural and some synthetic PPARa, g, anda/d agonists increased lipid synthesis in human
sebocytes cultures (SZ95 and SEB-1 cell lines)[43,46,47].
Investigations on human SZ95 sebocytes in
vitro suggest that PPARg activation by ultravioletlight B irradiation and PPARg-dependent cyclo-oxygenase-2 activation are important regulatorsof cellular responses to oxidative stress [48]. Inhi-
bition of the synthesis of the natural PPARa ago-nist leukotriene B4 in vivo led to significantimprovement of inflammatory acne lesions and
seborrhoea [49–51].In sebocyte development, there is a limited
cooperation between PPAR and RXR agonists.
In vitro, a submaximal dose of rexinoidda RXRagonistdaugmented the stimulation of sebocytedifferentiation by PPAR agonists [52].
The development of new RXR agonists might
lead to cancer prevention for diseases, such asMuir-Torre syndrome, where sebaceous tumorsarise on skin [53].
Neurotrophic factor receptor
Ret is the specific receptor for glial cell line–derived neurotrophic factor (GDNF), a factor
initially identified as a neurotrophic factor fordoaminergic neurons. Ret was found in rats in theepithelial cells of the secretory duct and the coiled
secretory terminal of eccrine sweat glands. Electronmicroscopy showed thatRet immunoreactivity waslocalized in the basal area of secretory cells adjacent
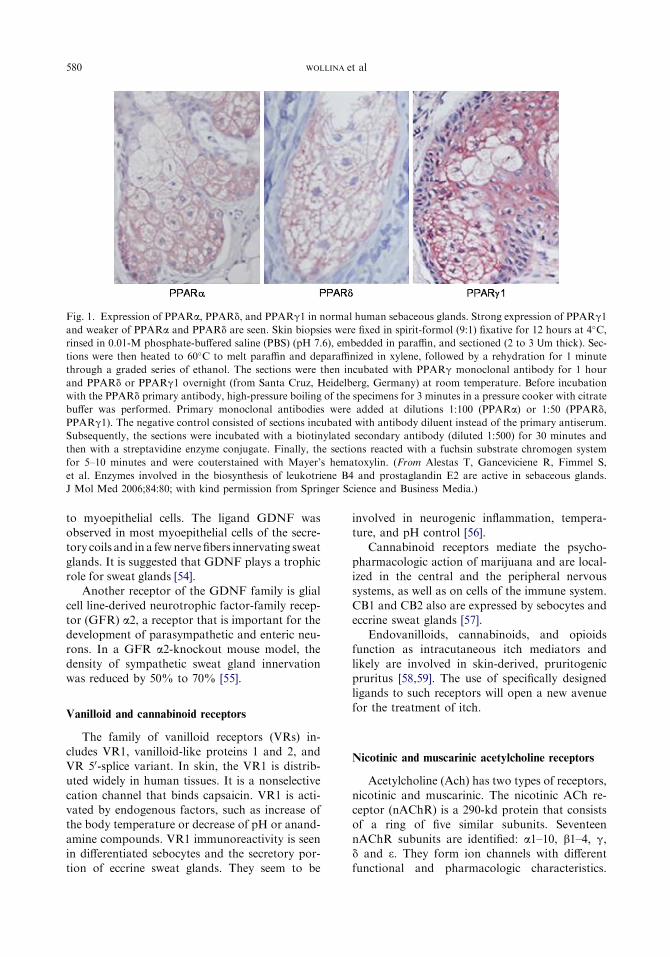
580 WOLLINA et al
Fig. 1. Expression of PPARa, PPARd, and PPARg1 in normal human sebaceous glands. Strong expression of PPARg1
and weaker of PPARa and PPARd are seen. Skin biopsies were fixed in spirit-formol (9:1) fixative for 12 hours at 4�C,rinsed in 0.01-M phosphate-buffered saline (PBS) (pH 7.6), embedded in paraffin, and sectioned (2 to 3 Um thick). Sec-
tions were then heated to 60�C to melt paraffin and deparaffinized in xylene, followed by a rehydration for 1 minute
through a graded series of ethanol. The sections were then incubated with PPARg monoclonal antibody for 1 hour
and PPARd or PPARg1 overnight (from Santa Cruz, Heidelberg, Germany) at room temperature. Before incubation
with the PPARd primary antibody, high-pressure boiling of the specimens for 3 minutes in a pressure cooker with citrate
buffer was performed. Primary monoclonal antibodies were added at dilutions 1:100 (PPARa) or 1:50 (PPARd,
PPARg1). The negative control consisted of sections incubated with antibody diluent instead of the primary antiserum.
Subsequently, the sections were incubated with a biotinylated secondary antibody (diluted 1:500) for 30 minutes and
then with a streptavidine enzyme conjugate. Finally, the sections reacted with a fuchsin substrate chromogen system
for 5–10 minutes and were couterstained with Mayer’s hematoxylin. (From Alestas T, Ganceviciene R, Fimmel S,
et al. Enzymes involved in the biosynthesis of leukotriene B4 and prostaglandin E2 are active in sebaceous glands.
J Mol Med 2006;84:80; with kind permission from Springer Science and Business Media.)
to myoepithelial cells. The ligand GDNF wasobserved in most myoepithelial cells of the secre-tory coils and in a fewnervefibers innervating sweat
glands. It is suggested that GDNF plays a trophicrole for sweat glands [54].
Another receptor of the GDNF family is glial
cell line-derived neurotrophic factor-family recep-tor (GFR) a2, a receptor that is important for thedevelopment of parasympathetic and enteric neu-rons. In a GFR a2-knockout mouse model, the
density of sympathetic sweat gland innervationwas reduced by 50% to 70% [55].
Vanilloid and cannabinoid receptors
The family of vanilloid receptors (VRs) in-
cludes VR1, vanilloid-like proteins 1 and 2, andVR 50-splice variant. In skin, the VR1 is distrib-uted widely in human tissues. It is a nonselective
cation channel that binds capsaicin. VR1 is acti-vated by endogenous factors, such as increase ofthe body temperature or decrease of pH or anand-
amine compounds. VR1 immunoreactivity is seenin differentiated sebocytes and the secretory por-tion of eccrine sweat glands. They seem to be
involved in neurogenic inflammation, tempera-ture, and pH control [56].
Cannabinoid receptors mediate the psycho-
pharmacologic action of marijuana and are local-ized in the central and the peripheral nervoussystems, as well as on cells of the immune system.
CB1 and CB2 also are expressed by sebocytes andeccrine sweat glands [57].
Endovanilloids, cannabinoids, and opioidsfunction as intracutaneous itch mediators and
likely are involved in skin-derived, pruritogenicpruritus [58,59]. The use of specifically designedligands to such receptors will open a new avenue
for the treatment of itch.
Nicotinic and muscarinic acetylcholine receptors
Acetylcholine (Ach) has two types of receptors,
nicotinic and muscarinic. The nicotinic ACh re-ceptor (nAChR) is a 290-kd protein that consistsof a ring of five similar subunits. Seventeen
nAChR subunits are identified: a1–10, b1–4, g,d and 3. They form ion channels with differentfunctional and pharmacologic characteristics.

581RECEPTORS OF SKIN GLANDS
Nicotin is an ACh antagonist and interacts withall AChR types except a9 subunits [51].
Five subtypes of muscarinic AChR (mAChR)are known. The mAChRs are expressed by
keratinocytes in human epidermis and skinglands. The mAChRs are involved in the earlysteps of embryologic differentiation of eccrine
sweat glands. During development, the innerva-tion of eccrine sweat glands undergoes a strikingchange from noradrenergic to cholinergic func-
tion. In rat skin it was observed that mAChRs aredetectable as early as postnatal day 4 when glandsbegin to form. Within the next 2 weeks, m3 AChR
protein and mRNA increase. m3 AChR expres-sion does not depend on intact gland innervation.The m3 AChR is coupled to phospholipase C [60].
Muscarinic blockage during development or in
adult animals results in the loss of secretoryresponsiveness, which is regulated by innervation[61]. In postnatal life, ACh is the major transmit-
ter for sudomotoric activity and seems to be ofimportance for sebaceous gland activity. In ratskin, m2 AChR is expressed by nerve fibers that
innervate eccrine sweat glands [62] but m3AChR seems to be the major receptor subtype inthe regulation of sweat gland secretion [63].
Scopolamin is a substance with similar bindingaffinities to all of the five known muscarinicreceptor subtypes. By 0.6-mg scopolamine givenorally, a significant reduction of sweat gland
activity occurs as measured by skin conductancein normal adult volunteers [64].
In undifferentiated sebocytes, the nAChR sub-
units a3, a9, b4, and mAChR subtypes m2–m5are found. Mature sebocytes express a7, b2, b4,m2, and m4 AChR. In eccrine sweat glands, the
a3, a7, and m2–m5 AChR are most prominent inmyoepithelial cells whereas a9, b2, m1, m3, andm4 AChR are present in acinar epithelia [65].
The expression of a7 AChR is abolished in the
epidermis adjacent to the acrosyringium in pal-moplantar pustulosis, a disease associated withnicotine abuse [66]. Dysfunction of mAChR is
a possible cause of idiopathic sudomotor failure(a disease with sudden onset, concomitant sharppain, or cholinergic urticaria over the entire
body), elevated serum IgE, lack of autonomic dys-function other than generalized anhidrosis, andmarked response to steroids. Although thermo-
regulatory sweating is absent, emotional sweatingis preserved [67].
Botulinum toxin is the most potent pharma-cologic inhibitor of focal hyperhidrosis. It
blocks specific intracellular transport proteins
of ACh but itself does not interact with anyAChR [68].
Goldstein and colleagues [69] reported a patientwho had frequent episodes of pain, sweating, and
flushing bilaterally in the hairless skin of the oph-thalmic and maxillary distributions of the trigem-inal nerve. The patient reported benefit from
anticholinergic quarternary amine trospium chlo-ride whereas intravenous edrophonium evokedpain and sweating. The authors concluded that
facial pain and sweating can result from by occu-pation of muscarinic cholinergic receptors afterACh release from local nerves.
Angiotensin II receptors
The tissue response to angiotensin IIdanoctapeptide that regulates blood pressuredis
mediated by specific cell surface receptors, AT1and AT2. AT1 is predominant in the vascularsystem. AT1 is expressed by eccrine and apocrine
sweat glands. In apocrine glands, luminal andperiluminal cells of the acrosyringium are AT1positive. The luminal cells of the dermal ductexpress AT1 whereas the basal cells, secretory
cells, and myoepithelial cells are negative. Thewhole gland is AT2 negative. In eccrine sweatglands, the complete acrosyringium and the lumi-
nal cells of the dermal duct are AT1 positive andthe remaining cellular elements negative. AT2 isnot expressed by eccrine sweat glands [70]. AT1
also is found in sebaceous glands [71].In eccrine poroma, some of the tumor cells in
cell strands and cells surrounding luminal struc-
tures are AT1 positive; AT2 is not expressed [70].
Melanocortin receptors
Melanocortins regulate immune functions,
pigmentation, lipid metabolism, and adrenalhormone secretion. Melanocortins include adre-nocorticotrophic hormone, a-melanocyte-stimu-
lating hormone (aMSH), b-lipotrophin, andendorphins. Melanocortins bind to specificG-protein coupled receptors. Five different mela-nocortin receptor (MR) subtypes are known [72].
MR-1 transcripts and immunoreactivity areseen in sebocytes in vitro and in vivo. Sebumproduction can be increased by aMSH. By in-
teraction of aMSH with MR-1, interleukin (IL)-8expression of sebocytes is modulated. Immunoe-lectronmicroscopy demonstrates MR-1 along the
cell surface and intracytoplasmatically withintubular endosomes, suggesting the internalizationof MR-1 [73].

582 WOLLINA et al
MR-1 is expressed by a variety of sebaceousmalformations, hamartomas, and neoplasms,such as sebaceoma, cylindroma, and extramam-
mary Paget’s disease. MR-1 is found in eccrinesweat glands in dermal ducts and secretoryepithelia. Eccrine tumors, such as syringoma orhidroacanthoma simplex, also express this type of
MR [74].MR-5 is expressed by differentiating, lipid-
laden sebaceous cells but not in basal sebocytes.
Therefore, MR-5 seems to represent a sebocytedifferentiation marker [75]. In transgenic mice,a targeted disruption ofMR-5 resulted in amarked
decrease in sebum production by sebaceousglands. MR-5 also is expressed by eccrine sweatglands [76]. Others could not identify MR-5immunoreactivity in sebaceous glands [73].
Because melanocortins can suppress in-flammation by prevention of nuclear factor kBactivation, MR ligands are of potential creating
new anti-inflammatory acne drugs [77].
Further neuropeptide receptors
Vasoactive intestinal peptide receptor (VIP-R)has been localized by immunohistochemistry in
acinar cells of the secretory coil and in luminalcells of the duct in eccrine sweat glands. It hasbeen suggested that VIP is involved in regulation
of chloride reabsorption from primary sweat atthe ductal segment [78]. Further analysis revealedthat two functional receptor types are expressed,namely a high-affinity VIP-R (kd ¼ 0.58–2.4 kd)
and a low-affinity type (kd ¼ 175–288 kd)(Fig. 2) [79]. VIP-R immunoreactivity is seen in
cystic structures of steatocystoma multiplex [80]but is absent in normal sebaceous glands [81].
Corticotropin-releasing hormone (CRH) is the
most proximal element of the hypothalamic-pitu-itary-adrenal axis, and it acts as central coordina-tor for neuroendocrine and behavioral responsesto stress. In human skin, the CRH receptor
(CRH-R) 1 is expressed in all major cellularpopulations of epidermis, dermis, and subcutiswith CRH-R1a the most prevalent isoform. The
CRH-R2 gene is expressed solely in hair folliclekeratinocytes and papilla fibroblasts, whereasCRH-R2 antigen is localized predominantly in
hair follicles, sebaceous and eccrine glands, andmuscle and blood vessels [82,83].
Investigations on the expression of CRH,CRH-binding protein (CRH-BP), and CRHRs
were performed in SZ95 sebocytes in vitro andnormal and acne-involved sebaceous glands invivo [83,84]. CRH, CRH-BP, CRH-R1, and
CRH-R2 were detectable in SZ95 sebocytes atthe mRNA and protein levels with CRH-R1the predominant type (CRH-R1/CRH-R2 ¼ 2)
(Fig. 3). CRH was biologically active on humansebocytes. It induced a biphasic increase of syn-thesis of sebaceous lipids with a maximum stimu-
lation at 10�7 Mol (M) and upregulated mRNAlevels of 3b-hydroxysteroid dehydrogenase/D(5-4)isomerase, although it did not affect cell viabil-ity, cell proliferation, or IL-1 beta-induced IL-8
release. Testosterone at 10�7 M downregulatedCRH-R1 and CRH-R2 mRNA expression at 6to 24 hours, and growth hormone switched
CRH-R1 mRNA expression to CRH-R2 at 24hours. Based on these findings, CRH may be anautocrine hormone for human sebocytes that
Fig. 2. Expression of VIP-R. Immunoperoxidase staining with VIP-R specific monoclonal antibody 109.10 (Immuno-
tech, Marseille, France) diluted 1:50. (A) Periglandular cells and cutaneous nerve fibers around the sebaceous gland
were stained whereas the sebocytes remained negative. (B) VIP-R expression in the most mature layer of epithelial cells
of steatocystoma multiplex.
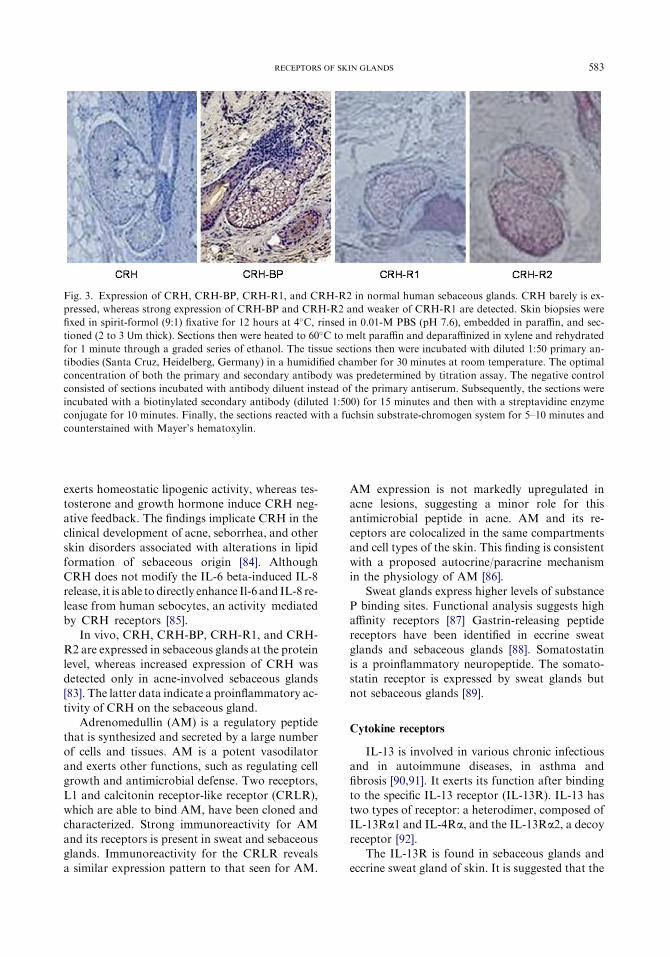
583RECEPTORS OF SKIN GLANDS
Fig. 3. Expression of CRH, CRH-BP, CRH-R1, and CRH-R2 in normal human sebaceous glands. CRH barely is ex-
pressed, whereas strong expression of CRH-BP and CRH-R2 and weaker of CRH-R1 are detected. Skin biopsies were
fixed in spirit-formol (9:1) fixative for 12 hours at 4�C, rinsed in 0.01-M PBS (pH 7.6), embedded in paraffin, and sec-
tioned (2 to 3 Um thick). Sections then were heated to 60�C to melt paraffin and deparaffinized in xylene and rehydrated
for 1 minute through a graded series of ethanol. The tissue sections then were incubated with diluted 1:50 primary an-
tibodies (Santa Cruz, Heidelberg, Germany) in a humidified chamber for 30 minutes at room temperature. The optimal
concentration of both the primary and secondary antibody was predetermined by titration assay. The negative control
consisted of sections incubated with antibody diluent instead of the primary antiserum. Subsequently, the sections were
incubated with a biotinylated secondary antibody (diluted 1:500) for 15 minutes and then with a streptavidine enzyme
conjugate for 10 minutes. Finally, the sections reacted with a fuchsin substrate-chromogen system for 5–10 minutes and
counterstained with Mayer’s hematoxylin.
exerts homeostatic lipogenic activity, whereas tes-
tosterone and growth hormone induce CRH neg-ative feedback. The findings implicate CRH in theclinical development of acne, seborrhea, and otherskin disorders associated with alterations in lipid
formation of sebaceous origin [84]. AlthoughCRH does not modify the IL-6 beta-induced IL-8release, it is able todirectly enhance Il-6 and IL-8 re-
lease from human sebocytes, an activity mediatedby CRH receptors [85].
In vivo, CRH, CRH-BP, CRH-R1, and CRH-
R2 are expressed in sebaceous glands at the proteinlevel, whereas increased expression of CRH wasdetected only in acne-involved sebaceous glands[83]. The latter data indicate a proinflammatory ac-
tivity of CRH on the sebaceous gland.Adrenomedullin (AM) is a regulatory peptide
that is synthesized and secreted by a large number
of cells and tissues. AM is a potent vasodilatorand exerts other functions, such as regulating cellgrowth and antimicrobial defense. Two receptors,
L1 and calcitonin receptor-like receptor (CRLR),which are able to bind AM, have been cloned andcharacterized. Strong immunoreactivity for AM
and its receptors is present in sweat and sebaceousglands. Immunoreactivity for the CRLR revealsa similar expression pattern to that seen for AM.
AM expression is not markedly upregulated in
acne lesions, suggesting a minor role for thisantimicrobial peptide in acne. AM and its re-ceptors are colocalized in the same compartmentsand cell types of the skin. This finding is consistent
with a proposed autocrine/paracrine mechanismin the physiology of AM [86].
Sweat glands express higher levels of substance
P binding sites. Functional analysis suggests highaffinity receptors [87] Gastrin-releasing peptidereceptors have been identified in eccrine sweat
glands and sebaceous glands [88]. Somatostatinis a proinflammatory neuropeptide. The somato-statin receptor is expressed by sweat glands butnot sebaceous glands [89].
Cytokine receptors
IL-13 is involved in various chronic infectiousand in autoimmune diseases, in asthma andfibrosis [90,91]. It exerts its function after binding
to the specific IL-13 receptor (IL-13R). IL-13 hastwo types of receptor: a heterodimer, composed ofIL-13Ra1 and IL-4Ra, and the IL-13Ra2, a decoyreceptor [92].
The IL-13R is found in sebaceous glands andeccrine sweat gland of skin. It is suggested that the

584 WOLLINA et al
receptor might play a functional role in theinflammatory cascade of atopic dermatitis [93].
IL-13 and IL-13R are increased in lesional skin
of bleomycin-induced scleroderma in mice [94].Spontaneous scleroderma in humans shows a re-markable impact of skin gland architecture. It isyet not clear, however, whether or not IL-13 and
IL-13R of these glands contribute to dermal fibro-sis and sclerosis.
Cytokine receptors have been identified in
clear cells and duct epithelia of eccrine sweatglands, such as IL-1aR, IL-1bR, IL-6R, andtumor necrosis factor a-R. Inflammatory skin,
as in cutaneous leishmaniasis, shows minor mod-ifications of the staining intensity in particular forIL-6R [95].
Transforming growth factor-beta
and morphogenetic protein receptors
The transforming growth factor-beta (TGF-b)superfamily is involved in growth and differenti-ation. In mammals, TGF-b is found in threehighly homologous isoforms that exert their
effects via heteromeric complexes of type-I andtype-II receptors (TGF-b receptor proteins type Iand type II [TbRI and TbRII]). TGF-b regulates
the growth and metabolism of various cell types,
including keratinocytes. These growth factors sig-nal through transmembrane serine/threoninekinase receptors. Activation of type I receptor
kinase phosphorylates intracellular signaling pro-teins named Smads.
In normal human skin, immunoreactive TGF-b2, but not TGF-b1, is detected predominantly in
the epidermis, hair follicles, and sebaceous glands.The epidermal expression of TbRI and TbRII isvery weak in the majority of normal skin samples.
In addition, no expression of TGF-b2 is detectedin the eccrine secretory portion or in eccrinespiradenoma, but it is detected in the upper
eccrine ducts and in eccrine poroma [96].In an immunohistochemical study of human
eccrine sweat glands, expression of TGFb-RI wasrestricted to myoepithelial cells, whereas bone
morphogenetic protein-receptor (BMPR) type IAwas expressed selectively by dermal duct andacrosyringium. BMPR type IB monoclonal anti-
body gave a faint staining of secretory epitheliumand myoepithelial cells. It is suggested thatmyoepithelial cells and duct cells are major targets
of the TGF-b signaling pathway [97].The authors investigated the expression of
TGF-b1, -b2, and -b3; latent TGF-b–bindingprotein (LTBP); and TbRI and TbRII during in-duced hair growth in C57 BL-6 mice. They havefound a positive reactivity for LTBP and TbRI
Table 1
Some implications of ligand-receptor interactions of skin glands in clinical dermatology
Receptor type Pathogenic and diagnostic relevance Therapeutic relevance
Estrogen Y in eccrine neoplasms Hormone replacement therapy for menopausal
women; hormone therapy for hyperandrogenism
and female acne
Progesteron [ in syringoma ?
Androgen [ in cystic acne Dexamethasone treatment of cystic acne;
antiandrogens for female acne
Mineralocorticoid ? ?
Glucocorticoid Steroid acne ?
Orphan nuclear Acne, seborrhoea Chemoprevention of cancer
Neurotrophic factor ? ?
Vanilloid, cannabinoid,
opioid
Itch, pain Potential for new itch and pain therapy
Nicotinic/muscarinic Hyperhidrosis, pain Pain and antihydrotic therapy
Melanocortin [ in hamartomas and tumors Potential for anti-inflammatory drugs
VIP Variable expression in eccrine
tumors
?
Corticotropin-releasing
hormone
? Potential new acne treatments
Adrenomedullin ? Antimicrobial substances
IL-13 Fibrosis, sclerosis ?
TGF-b Promotes growth of sebaceous glands ?

585RECEPTORS OF SKIN GLANDS
in sebocytes. TbRII was found in sebaceousglands without significant variations during thehair cycle. A bidirectional interaction of sebocytesand hair follicle epithelium in the TGF-b/LTBPseems possible. Sebocytes can be considered tobe a target for TGFs, because they express bothTbRI and TbRII. The general properties of
TGF-b as a growth inhibitor of epithelial cellsmay suggest involvement in either the abrogationof extensive growth at the end of anagen or the
initiation of catagen for the follicle epitheliumand growth control for sebaceous glands [98].
Clinical implications and therapeutic relevance
The knowledge of specific ligand-receptor in-teractions during development and function ofskin glands not only is of academic interest but
also this knowledge has implications for diagnos-tics and treatments. The expression of ligands andreceptors is a tool in dermatopathology of adnexal
tumors. Available treatments of female acne,hyperandrogensism, focal hyperhidrosis, itch, andpain are in part based on ligand-receptor effects.Further substances and indications are on the
horizon. A summary is provided in Table 1.
Summary
The skin is the largest independent peripheralendocrine organ and exerts a great complexity ofinteraction with hormones, neuropeptides, PPARligands, growth factors, and cytokines [99,100].
Knowledge of receptor expression and receptor-ligand function offers new insight into pathologicconditions and opens new opportunities in drug
development.
References
[1] Loomis CA, BirgeMB. Fetal skin development. In:
Eichenfield LF, Frieden IJ, Esterly NB, editors.
Textbook of neonatal dermatology. Philadelphia:
WB Saunders; 2001. p. 1–17.
[2] Zouboulis CC, Fimmel S, Ortmann J, et al.
Sebaceous glands. In: Hoath SB, Maibach HI, edi-
tors. Neonatal skindstructure and function. 2nd
edition. NewYork:Marcel Dekker; 2003. p. 59–88.
[3] Thornton MJ. The biological actions of estrogens
on skin. Exp Dermatol 2002;11:487–502.
[4] Vevrier-Sevrain S, Bonte F, Gilchrest B. Biology of
estrogens in skin: implications for aging skin. Exp
Dermatol 2006;15:83–94.
[5] De Angelis M, Stossi F, Waibel M, et al. Isocou-
marins as estrogen receptor beta selective li-
gands: isomers from isoflavone phytoestrogens
and their metabolites. Bioorg Med Chem 2005;
23:6529–42.
[6] Makrantonaki E, Adjaye J, Herwig R, et al. Age-
specific hormonal decline is accompanied by
transcriptional changes in human sebocytes in
vitro. Aging Cell 2006;5:331–44.
[7] Azzi L, El-Alfy M, Labrie F. Gender differences
and effects of sex steroids and dehydroepiandros-
terone on androgen and oestrogen a receptors in
mouse sebaceous glands. Br J Dermatol 2006;154:
21–7.
[8] Thornton MJ, Taylor AH, Mulligan K, et al. Oes-
trogen receptor beta is the predominant oestrogen
receptor in human scalp skin. Exp Dermatol
2003;12:181–90.
[9] Kariya Y, Moriya T, Suzuki T, et al. Sex steroid
hormone receptors in human skin appendage and
its neoplasms. Endocr J 2005;52:317–25.
[10] Hu S, Bakshandeh H, Kerdel FA, et al. Eccrine
syringofibroadenoma of clear cell variant: an
immunohistochemical study. Am J Dermatopathol
2005;27:228–31.
[11] Betis F, Hofman V, Lagier J, et al. Adenocarci-
nome sudoral primitive de la paupiere de type
eccrine a cellules en bague a chaton. J FrOphtalmol
2002;25:547–51.
[12] Mirza I, Kloss R, Sieber SC. Malignant eccrine
spiradenoma. Arch Pathol Lab Med 2002;126:
591–4.
[13] Beier K, Ginez I, Schaller H. Localization of ste-
roid hormone receptors in the apocrine sweat
glands of the human axilla. Histochem Cell Biol
2005;123:61–5.
[14] Baulien E-E, Thomas G, Legrain S, et al. Dehy-
droepiandrosterone (DHEA), DEHEA sulphate,
and aging: contribution of the DHEAge study to
a sociabiological issue. Proc Natl Acad Sci USA
2000;97:4279–84.
[15] Labrie F, Lun-The V, Belanger A, et al. Is dehy-
droepiandrosterone a hormone? J Endocrinol 2005;
187:169–96.
[16] Wallace ML, Smoller BR. Progesterone receptor
positivity supports hormonal control of syringo-
mas. J Cutan Pathol 1995;22:442–5.
[17] Timpanidis PC, Lakhani SR, Groves RW. Proges-
terone receptor-positive syringoma associated with
diabetes. J Am Acad Dermatol 2003;48(Suppl 5):
S103–4.
[18] Fimmel S, Saborowski A, Terouanne B, et al. Inhi-
bition of the androgen receptor by antisense
oligonucleotides regulates the biological activity
of androgens in SZ95 sebocytes. Horm Metab
Res 2007;39:149–56.
[19] Choudhry R, Hodgins MB, Van der Kwast TH,
et al. Localization of androgen receptors in human
skin by immunohistochemistry: implications for
the hormonal regulation of hair growth, sebaceous
glands and sweat glands. J Endocrinol 1992;133:
467–75.

586 WOLLINA et al
[20] Shikata N, Kurokawa I, Andachi H, et al. Expres-
sion of androgen receptors in skin appendage
tumors: an immunohistochemical study. J Cutan
Pathol 1995;22:149–53.
[21] Bratthauer GL, Lininger RA, Man YG, et al. An-
drogen and estrogen receptor mRNA status in
apocrine carcinomas. Diagn Mol Pathol 2002;11:
113–8.
[22] Hamilton KS, Johnson S, Smoller BR. The role of
androgen receptors in the clinical course of nevus
sebaceous of Jadassohn. Mod Pathol 2001;14:
539–42.
[23] Offidani A, Campanati A. Papillary hidradenoma:
immunohistochemical analysis of steroid receptor
profile with a focus on apocrine differentiation.
J Clin Pathol 1999;52:829–32.
[24] Stoeckelhuber M, Stoeckelhuber BM, Welsch U.
Apocrine glands in the eyelid of primates contrib-
ute to the ocular host defense. Cells Tissues Organs
2004;176:187–94.
[25] Tamer C, Oksuz H, Sohut S. Androgen status of
the nonautoimmune dry eye. Ophthalmic Res
2006;38:280–6.
[26] Fujimoto A, Takata M, Hatta N, et al. Expression
of structurally unaltered androgen receptor in
extramammary Paget’s disease. Lab Invest 2000;
80:1465–71.
[27] Diaz de Leon E, Carcangiu ML, Prieto VG, et al.
Extramammary Paget disease is characterized by
the consistent lack of estrogen and progesterone re-
ceptors but frequently expresses androgen receptor.
Am J Clin Pathol 2000;113:572–5.
[28] Zouboulis CC. Acne: sebaceous gland action. Clin
Dermatol 2004;22:360–6.
[29] Huber J, Walch K. Treating acne with oral contra-
ceptives: use of lower doses. Contraception 2006;
73:23–9.
[30] Katsabas AD, Stefanaki C, Cunliffe WJ. Guidelines
for treating acne. Clin Dermatol 2004;22:439–44.
[31] Marynick SP, Chakmakjian ZH, McAffree DL,
et al. Androgen excess in cystic acne. N Engl
J Med 1983;308:981–6.
[32] Shaw JC. Acne: effect of hormones on pathogen-
esis and treatment. Am J Clin Dermatol 2002;3:
571–8.
[33] SchroderU,Dries V,Klussmann JP, et al. Successful
adjuvant tamoxifen therapy for estrogen receptor-
positive metastasizing sweat gland adenocarcinoma:
need for a clinical trial? Ann Otol Rhinol Laryngol
2004;113:242–4.
[34] Daniel SJ, Nader R, Kost K, et al. Facial sweat
gland carcinoma metastasizing to neck nodes: a di-
agnostic and therapeutic challenge. Arch Otolar-
yngol Head Neck Surg 2001;127:1495–8.
[35] Venencie PY, Meduri G, Pissard S, et al. Luteiniz-
ing hormone/human chorionic gonadotrophin
receptors in various epidermal structures. Br J Der-
matol 1999;141:438–46.
[36] Verrey F. Transcriptional control of sodium trans-
port in tight epithelia by adrenal steroids. J Membr
Biol 1995;144:93–110.
[37] Zennaro M-C, Farman N, Bonvalet J-P, et al. Tis-
sue-specific expression ofa andbmessenger ribonu-
cleic acid isoforms of the human mineralocorticoid
receptor in normal and pathological states. J Clin
Endocrinol Metab 1997;82:1345–52.
[38] Hirasawa G, Sasano H, Takahasi K, et al. Colocal-
ization of 11 beta-hydroxysteroid dehydrogenase
type II andmineralocorticoid receptor in human ep-
ithelia. J Clin Endocrinol Metab 1997;82:3859–63.
[39] Nussey S, Whitehead S. Endocrinology. An inte-
grated approach. Oxford (UK): BIOS Scientific
Publisher; 2001.
[40] Karstila T, Rechardt L, Honkaniemi J, et al. Im-
munocytochemical localization of glucocorticoid
receptor in rat skin. Histochemistry 1994;102:
305–9.
[41] ChenW,Yang C-C, SheuH-M, et al. Expression of
peroxisome proliferator-activated receptor and
CCAAT/enhancer binding protein transcription
factors in cultured human sebocytes. J Invest Der-
matol 2003;121:441–7.
[42] DownieMM, Sanders DA,Maier LM, et al. Perox-
isome proliferation-activated receptor and farne-
soid X receptor ligands differentially regulate
sebaceous differentiation in human sebaceous
glands cultures in vitro. Br J Dermatol 2004;151:
766–75.
[43] Alestas T, Ganceviciene R, Fimmel S, et al. En-
zymes involved in the biosynthesis of leukotriene
B4 and prostaglandin E2 are active in sebaceous
glands. J Mol Med 2006;84:75–87.
[44] Patel MV, McKay IA, Burrin JM. Transcriptional
regulators of steroidogenesis, DAX-1 and SF-1, are
expressed in human skin. J Invest Dermatol 2001;
117:1559–65.
[45] ChenW,YangC, Liao CY, et al. Expression of sex-
determining genes in human sebaceous glands and
their possible roles in pathogenesis of acne. J Eur
Acad Dermatol Venereol 2006;20:846–52.
[46] Trivedi NR, Cong Z, Nelson AM, et al. Peroxi-
some proliferator-activated receptors increase hu-
man sebum production. J Invest Dermatol 2006;
126:2002–9.
[47] Wrobel A, Seltmann H, Fimmel S, et al. Differenti-
ation and apoptosis in human immortalized sebo-
cytes. J Invest Dermatol 2003;120:175–81.
[48] Zhang Q, Seltmann H, Zouboulis CC, et al. In-
volvement of PPARgamma in oxidative stress-
mediated prostaglandin E(2) production in SZ95
human sebaceous gland cells. J Invest Dermatol
2006;126:42–8.
[49] Zouboulis CC, Nestoris S, Adler YD, et al. A new
concept for acne therapy: a pilot study with zileu-
ton, an oral 5-lipoxygenase inhibitor. Arch Derma-
tol 2003;139:668–70.

587RECEPTORS OF SKIN GLANDS
[50] Zouboulis CC, Saborowski A, Boschnakow A.
Zileuton, an oral 5-lipoxygenase inhibitor, directly
reduces sebum production. Dermatology 2005;210:
36–8.
[51] Misery L. Nicotine effects on skin: are they positive
or negative? Exp Dermatol 2004;13:665–70.
[52] Kim MJ, Deplewski D, Ciletti N, et al. Limited
cooperation between peroxisome proliferators-
activated receptors and retinoid X receptor antago-
nists in sebocyte growth and development. Mol
Genet Metab 2001;74:362–9.
[53] Lippman SM, Lotan R. Advances in the develop-
ment of retinoids as chemopreventive agents.
J Nutr 2000;130:479S–82S.
[54] Yoshida T. Immunohistochemical localization of
glial cell line-derived neurotrophic factor and its
receptor Ret in the rat sweat gland. Kurume Med
J 2004;51:193–202.
[55] Hiltunen PH, Airaksinen MS. Sympathetic cholin-
ergic target innervation requires GDNF family
receptor GFR alpha 2. Mol Cell Neurosci 2004;
26:450–7.
[56] Stander S, Moormann C, SchumacherM, et al. Ex-
pression of vanilloid receptor subtype 1 in cutane-
ous sensory nerve fibers, mast cells, and epithelial
cells of appendage structures. Exp Dermatol 2004;
13:129–39.
[57] Stander S, SchmelzM,MetzeD, et al. Distrubution
of cannabinoid receptor 1 (CB1) and 2 (CB2) on
sensory nerve fibers and adnexal structures in
human skin. J Dermatol Sci 2005;38:177–88.
[58] Paus R, Schmelz M, Biro T, et al. Frontiers in pru-
ritus research: scratching the brain. J Clin Invest
2006;116:1174–86.
[59] Ikoma A, SteinhoffM, Stander S, et al. The neuro-
biology of itch. Nat Rev Neurosci 2006;7:535–47.
[60] Grant MP, Landis SC. Developmental expression
of muscarinic cholinergic receptors and coupling
to phospholipase C in rat sweat glands are indepen-
dent of innervation. J Neurosci 1991;11:3772–82.
[61] Landis SC. Development of muscarinic receptors
and regulation of secretory responsiveness in ro-
dent sweat glands. Life Sci 1999;64:381–5.
[62] Haberberger RV, Bodenbenner M. Immunohisto-
chemical localization of muscarinic receptors (M2)
in the rat skin. Cell Tissue Res 2000;300:389–96.
[63] Schiavone A, Brambilla A. Muscarinic M3 recep-
tors mediate secretion from sweat glands in the
rat. Pharmacol Res 1991;23:233–9.
[64] Golding JF, Stott JR. Comparison of the effects of
a selective muscarinic receptor antagonist and
hyoscine (scopolamine) on motion thickness, skin
conductance and heart rate. Br J Clin Pharmacol
1997;43:633–7.
[65] Kurzen H, Berger H, Jager C, et al. Phenotypical
and molecular profiling of the extraneuronal cho-
linergic system of the skin. J Invest Dermatol
2004;123:937–49.
[66] Hagforsen E, Edvinsson M, Nordlind K, et al. Ex-
pression of nicotinic receptors in the skin of
patients with palmoplantar pustulosis. Br
J Dermatol 2002;146:383–91.
[67] Nakazato Y, Tamura N, Ohkuma A, et al. Idio-
pathic pure sudomotor failure: anhidrosis due to
deficits in cholinergic transmission. Neurology
2004;63:1476–80.
[68] Haider A, Solish N. Focal hyperhidrosis: diagno-
sis and treatment. Can Med Assoc J 2005;172:
69–75.
[69] Goldstein DS, Pechnik S, Moak J, et al. Painful
sweating. Neurology 2004;63:1471–5.
[70] Takeda H, Kondo S. Immunohistochemical study
of angiotensin receptors in normal human sweat
glands and eccrine poroma. Br J Dermatol 2001;
144:1189–92.
[71] Steckelings UM, Wollschlager T, Peters J, et al.
Human skin: source and target organ for angioten-
sin II. Exp Dermatol 2004;13:148–54.
[72] Millington GWM. Propiomelanocortin (POMC):
the cutaneous roles of its melanocortin products
and receptors. Clin Exp Dermatol 2006;31:
407–12.
[73] BohmM, Schiller M, Stander S, et al. Evidence for
expression of melanocortin-1 receptor in human
sebocytes in vitro and in vivo. J Invest Dermatol
2002;118:533–9.
[74] Stander S, BohmM, Brzoska T, et al. Expression of
melanocortin-1 receptor in normal, malformed and
neoplastic skin glands and hair follicles. Exp Der-
matol 2002;11:42–51.
[75] Zhang L, Li WH, Anthonavage M, et al. Melano-
cortin-5 receptor: a marker of human sebocyte dif-
ferentiation. Peptides 2006;27:413–20.
[76] Thiboutot D, Sivarajah A, Gilliand K, et al. The
melanocortin 5 receptor is expressed in human
sebaceous glands and rat preputial glands. J Invest
Dermatol 2000;115:614–9.
[77] Gatting SJ. Targeting melanocortin receptors as
potential novel therapeutics. Pharmacol Ther
2006;111:1–15.
[78] Kummer W, Herbst WM, Heym C. Vasoactive
intestinal polypeptide receptor-like immunoreac-
tivity in human sweat glands. Neurosci Lett 1990;
110:239–43.
[79] Heinz-Erian P, Paul S, Said SI. Receptor for vaso-
active intestinal peptide on isolated human sweat
glands. Peptides 1986;7(Suppl 1):151–4.
[80] Wollina U, Bocker T, Castelli E. Talgdrusentumo-
ren. In: Garbe C, Dummer R, Kaufmann R, et al,
editors. Dermatologische Onkologie. Berlin:
Springer-Verlag; 1997. p. 532–47.
[81] Wollina U, Paus R, Feldrappe S. Sequential
expression of glutathione-S-transferase isoenzymes
during hair anagen growth in mice and their
relationship to phosphotyrosinase and VIP recep-
tor. Histol Histopathol 1995;10:39–45.

588 WOLLINA et al
[82] Slominski A, Pisarchik A, Tobin DJ, et al. Differen-
tial expression of a cutaneous corticotropin-releasing
hormone system. Endocrinology 2004;145:941–50.
[83] Ganceviciene R, Marciukaitiene I, Graziene V,
et al. New accents in the pathogenesis of acne vula-
ris. Acta Medica, in press.
[84] Zouboulis CC, SeltmannH,Hiroi N, et al. Cortico-
tropin-releasing hormone: an autocrine hormone
that promotes lipogenesis in human sebocytes.
Proc Natl Acad Sci USA 2002;99:7148–53.
[85] KrauseK, Schnitger A, Fimmel S, et al. Corticotro-
pin-releasing hormone skin signalling is receptor-
mediated and is predominant in the sebaceous
glands. Horm Metab Res 2007;39:166–70.
[86] Muller FB, Muller-Rover S, Korge BP, et al. Adre-
nomedullin: expression and possible role in human
skin and hair growth. Br JDermatol 2003;148:30–8.
[87] Deguchi M, Niwa M, Shigematsu K, et al. Specific
[125I] Bolton-Hunter substance P binding sites in
human and rat skin. Neurosci Lett 1989;99:287–92.
[88] Staniek V, Misery L, Peguet-Navarro J, et al. Ex-
pression of gastrin-releasing peptide receptor in
human skin. Acta Derm Venereol 1996;76:282–6.
[89] Misery L, Bourchanny D, Kanitakis J, et al. Mod-
ulation of substance P and somatostatin receptors
in cutaneous lymphocytic inflammatory and
tumoral infiltrates. J Eur Acad Dermatol Venereol
2001;15:238–41.
[90] Krause S, Behrends J, Borowski A, et al. Blockade
of interleukin-13-mediated cell activationbyanovel
inhibitory antibody to human IL-13 receptor
alpha1. Mol Immunol 2006;43:1799–807.
[91] Fichtner-Feigl S, Strober W, Kawakami K, et al.
IL-13 signaling through the IL13alpha2 receptor
is involved in induction of TGF-beta1 production
and fibrosis. Nat Med 2006;12:99–106.
[92] Arima K, Sato K, Tanaka G, et al. Characteriza-
tion of the interaction between interleukin-13 and
interleukin-13 receptors. J Biol Chem 2005;280:
24915–22.
[93] Akaiwa M, Yu B, Umeshita-Suyama R, et al. Lo-
calization of human interleukin 13 receptor in
non-haematopoietic cells. Cytokine 2001;13:75–84.
[94] MatsushitaM,YamamotoT,NishiokaK.Upregu-
lation of interleukin-13 and its receptor in a murine
model of bleomycine-induce scleroderma. Int Arch
Allergy Immunol 2004;135:348–56.
[95] Ahmed AA, Nordlind K, Schultzberg M, et al.
Proinflammatory cytokines and their correspond-
ing receptor proteins in eccrine sweat glands in
normal and cutaneous leishmaniasis human skin.
An immunohistochemical study. Exp Dermatol
1996;5:230–5.
[96] Furue M, Kato M, Nakamura K, et al. Dysregu-
lated expression of transforming growth factor
beta and its type-I and type-II receptors in basal-
cell carcinoma. Int J Cancer 1997;71:505–9.
[97] Wollina U, Lange D, Ten Dijke P, et al. Eccrine
sweat glands: expression of transforming growth
factor-beta and bone morphogenetic protein type
I receptors and their intracellular Smad proteins.
Acta Derm Venereol 1999;79:183–6.
[98] Wollina U, Lange D, Funa K, et al. Expression of
transforming growth factor beta isoforms and their
receptors during hair growth phases in mice. Histol
Histopathol 1996;11:431–6.
[99] Zouboulis CC. Human skin: an independent
peripheral endocrine organ. Horm Res 2000;54:
230–42.
[100] Zouboulis CC. The human skin as a hormone
target and an endocrine gland. Hormones 2004;3:
9–26.





























