Embed Size (px)
Citation preview

408
Journal of Andrology, Vol. 24, No. 3, May/June 2003Copyright q American Society of Andrology
Regulation of Sulfated Glycoprotein-1 and Cathepsin DExpression in Adult Rat Epididymis
LOUIS HERMO AND SERO ANDONIAN
From the Department of Anatomy and Cell Biology, McGill University, Montreal, Quebec, Canada.
ABSTRACT: Endocytosis, whereby proteins are internalizedfrom the epididymal lumen to be eventually degraded in lyso-somes, is one of the major functions of the epididymal epithelialcells in maintaining a proper luminal milieu conducive for spermmaturation. In the present study, using light microscope immu-nocytochemical methods, we examined the regulation of 2 lyso-somal enzymes, sulfated glycoprotein-1 (SGP-1) and cathepsinD, in adult rat epididymides fixed in Bouin fixative and embeddedin paraffin. After orchidectomy (O) with or without testosterone(T) supplementation, efferent duct ligation (EDL), or hypophysec-tomy (H), lysosomes of principal cells were intensely reactivewith the anti-SGP-1 antibody, as were narrow, clear, and basalcells, with staining patterns similar to that of control animals.These experimental procedures also had no effect on cathepsinD expression in all cell types, except for clear cells of the corpusand cauda epididymidis, which after orchiedectomy and hypoph-ysectomy, became intensely reactive, unlike their completely un-reactive state in control animals. In O1T animals, as well as inEDL animals, clear cells remained unreactive. These data taken
together suggest that expression of SGP-1 is not under the con-trol of testicular or pituitary factors, as is also the case for ca-thepsin D expression by principal, narrow, and basal cells. How-ever, specific inhibition of cathepsin D expression by testosteroneor one of its metabolites appears to occur in clear cells of thecorpus and cauda epididymidis. Furthermore, in addition to small,typical lysosomes, principal cells also revealed large supranucle-ar and infranuclear spherical structures that were immunoreac-tive with both anti-SGP-1 and anti-cathepsin D antibodies, sug-gesting their lysosomal nature. With electron microscopy, thesestructures appeared electron-lucent and contained membranousprofiles embedded in an electron-dense, granular background.Such images suggest that the various experimental proceduresadversely affect the expression of several other lysosomal en-zymes in principal cells, leading to a lysosomal phenotype similarto that observed in various lysosomal storage diseases.
Key words: Light microscopy, orchidectomy, ligation, hypophysec-tomy, immunocytochemistry.
J Androl 2003;24:408-422
Throughout the entire epididymal duct, endocytosis isan important function of the epithelial cells, which
together with the synthesis and secretion of various sub-stances, leads to the proper conditioning of the luminalenvironment, which is essential for the maturation, via-bility, and storage of sperm (Cornwall et al, 2002; Hermoand Robaire, 2002). Endocytosis involves the uptake of avariety of substances from the epididymal lumen, manyof which are proteins, the origin of which is in part Ser-toli-cell derived, and in part from the sperm surface (Her-mo et al, 1994; Cooper, 1986). The endocytosis of pro-teins from the lumen into the epididymal epithelial cellsinvolves their receptor-mediated uptake via coated pits,followed by their appearance in the endocytic apparatus,which includes coated vesicles (100 nm in diameter), en-dosomes, multivesicular bodies, and lysosomes (Robaireand Hermo, 1988), as has also been shown for many othercell types (Murphy, 1991; Dunn and Maxfield, 1992). The
Supported by a grant from the Canadian Institutes of Health Research.Correspondence to: Dr Louis Hermo, Department of Anatomy and Cell
Biology, McGill University, 3640 University Street Room 1/33, Montreal,Quebec, Canada, H3A 2B2 (e-mail: [email protected]).
Received for publication; accepted for publication.
latter are the end points of endocytosis, at which proteinsare degraded into amino acids to be reused by the cells.
In the epididymis, the epithelial cells lining the ductinclude principal, narrow, clear, and basal cells, with eachpartaking in endocytosis in varying degrees (Moore andBedford, 1979; Hermo and Robaire, 2002). In principalcells, lysosomes have a distinct morphological appearanceand integral membrane proteins in the different epididy-mal regions (Hamilton, 1975; Robaire and Hermo, 1988;Suarez-Quian et al, 1992). In addition, various lysosomalenzymes are expressed in lysosomes of the epididymalepithelial cells, with some showing cell type-specific andregion-specific variations, suggesting substrate specificitywith regard to the turnover of proteins within the lyso-somes of these cells (Hermo et al, 1992; Tomomasa et al,1994; Abou-Haila et al, 1996).
The regulation of many epididymal epithelial functionshas been documented over the years to be dependent onandrogens, and recently, on estrogen, especially in effer-ent ducts, where it plays a role in the absorption of fluidfrom the lumen (Cornwall and Hann, 1995; Orgebin-Cristet al, 1996; Hess et al, 2002). Androgens also regulatethe expression and activity of several lysosomal enzymes(Cornwall et al, 2002). However, studies of the regulation

409Hermo and Andonian · Regulation of SGP-1 and Cathepsin D
of lysosomal enzyme expression in individual cell typesand regions of the epididymis have not been performed,with few exceptions (Luedtke et al, 2000).
Sulfated glycoprotein-1 (SGP-1), also referred to asprosaposin, has been noted in the epididymis by meansof Northern blot analysis and in situ hybridization (Syl-vester et al, 1989; Sun et al, 1994). Immunocytochemicalstudies with light and electron microscopy have localizedSGP-1 to the lysosomes of epithelial cells (Hermo et al,1992). SGP-1 is proteolytically cleaved in lysosomes intosaposins A, B, C, and D, which are sphingolipid-bindingproteins that function as activators for lysosomal enzymesinvolved in the hydrolysis of sphingolipids (Kretz et al,1990; O’Brien and Kishimoto, 1991). On the other hand,cathepsins are lysosomal proteolytic enzymes present incells of many tissues, and play a role in the intracellulardegradation of exogenous and endogenous proteins (Kir-schke et al, 1980; Kominami et al, 1991). Cathepsin D,an aspartyl endopeptidase, has a molecular weight of 42kd and an optimal activity of pH 3.8 (Srivastava and Nin-joor, 1982). In humans, cathepsin D has been localized tolysosomes of epithelial cells of the corpus epididymidis(Raczek et al, 1995), whereas in rats it is expressed in anepididymal cell type and region-specific manner (Igdouraet al, 1995).
In a previous study we noted that SGP-1 expression inthe efferent ducts was not dependent on luminal or cir-culating testicular factors, but was regulated by a pituitaryfactor (Rosenthal et al, 1995). Thus, it is of interest todetermine whether this is also true for SGP-1 expressionin the different cell types and regions of the epididymis.Likewise, it had been demonstrated that enzymatic activ-ity of cathepsin D in the epididymis increased after or-chidectomy but decreased after testosterone treatment(Mayorga and Bertini, 1982). However, the specific celltypes and regions of the epididymis affected were notdetermined.
Thus, the purpose of the present study was to examinethe regulation of SGP-1 and cathepsin D expression inthe epididymis after various experimental protocols onadult rats. Both enzymes have been localized by us incontrol animals in earlier studies (Hermo et al, 1992; Ig-doura et al, 1995). The protocols included orchidectomywith or without immediate testosterone replacement, ef-ferent duct ligation, and hypophysectomy. The epididy-mides of rats were fixed with Bouin fixative, embeddedin paraffin, and sections of the tissues were subsequentlyused for immunocytochemical analysis with light micros-copy.
Materials and MethodsAnimals and ProtocolsAdult male Sprague-Dawley rats (350–450 g) 3–4 months ofage were obtained from Charles River Laboratory Ltd (St Con-
stant, Quebec) and used for all aspects of this study. The animalswere subsequently subdivided into 6 groups. The first group con-sisted of untreated control animals. Bilateral ligation of the ef-ferent ducts constituted the second group. After an i.p. injectionof sodium pentobarbital (Somnitol, MTC Pharmaceuticals, Ham-ilton, Ontario), the testes and epididymides of adult rats wereexposed through an incision of the anterior abdominal wall. Asilk ligature was placed around both right and left efferent ductsat a proximal and distal site, after which they were cut in theinterval between the 2 ligatures. Care was taken to avoid ligatingthe adjacent blood vessels entering the testis. The animals (4 perinterval) were killed at 3, 7, 14, and 21 days following surgery.Bilateral orchidectomy constituted the third group. After anes-thesia, both testes of each rat were removed after a ligature wasplaced around the efferent ducts and testicular blood vessels. Theanimals (4 per interval) were killed at 3, 7, 14, and 21 days aftersurgery. Bilaterally orchidectomized rats that received 3 6.2-cmtestosterone-filled implants constituted the fourth group. Testos-terone-filled polydimethyl-siloxane (silastic) implants were pre-pared according to the method described by Stratton et al (1973)and have well-characterized steroid release rates (Brawer et al,1983). Subsequent to anesthesia, both testes were removed fromeach rat and the implants were placed s.c. immediately afterorchidectomy. The rats (4 per interval) were killed at 3, 7, 14and 21 days after surgery.
The fifth group consisted of hypophysectomized rats with 4rats per interval being killed at 7, 14, 21, and 28 days afterhypophysectomy. The sixth group consisted of 4 sham-operatedanimals, 2 of which received 3 empty 6.2-cm-long implants,with all rats being killed 14 days after initiation of the experi-ment.
All experimentation was carried out with minimal stress anddiscomfort being placed on the animals both during and aftersurgery according to the guidelines and approval of the univer-sity animal care committee.
Tissue Preparation for Light MicroscopyImmunocytochemistryAt the end of each treatment described above, the epididymidesof each Sprague-Dawley rat were fixed by perfusion with Bouinfixative via the abdominal aorta for 10 minutes. Following per-fusion, the epididymides were removed and cut so that sectionswould include all the major regions of the epididymis (ie, initialsegment, intermediate zone, caput, corpus, and cauda; Hermo etal, 1991b). The tissue was then immersed in Bouin fixative for72 hours, after which it was dehydrated and embedded in par-affin.
Light Microscopy ImmunostainingSections (5 mm thick) were cut and mounted on glass slides.They were then deparaffined with xylene and hydrated in gradedconcentrations of ethanol (from 100% to 50%). During hydra-tion, immersing the tissues in 70% ethanol containing 1% lith-ium carbonate for 5 minutes neutralized residual picric acid. In-activation of any endogenous peroxidase activity, use of glycinesolution in order to block free aldehyde groups, blocking withgoat serum, and washing with Tween buffer solution was per-formed as described previously (Hermo et al, 1992).
410 Journal of Andrology · May/June 2003
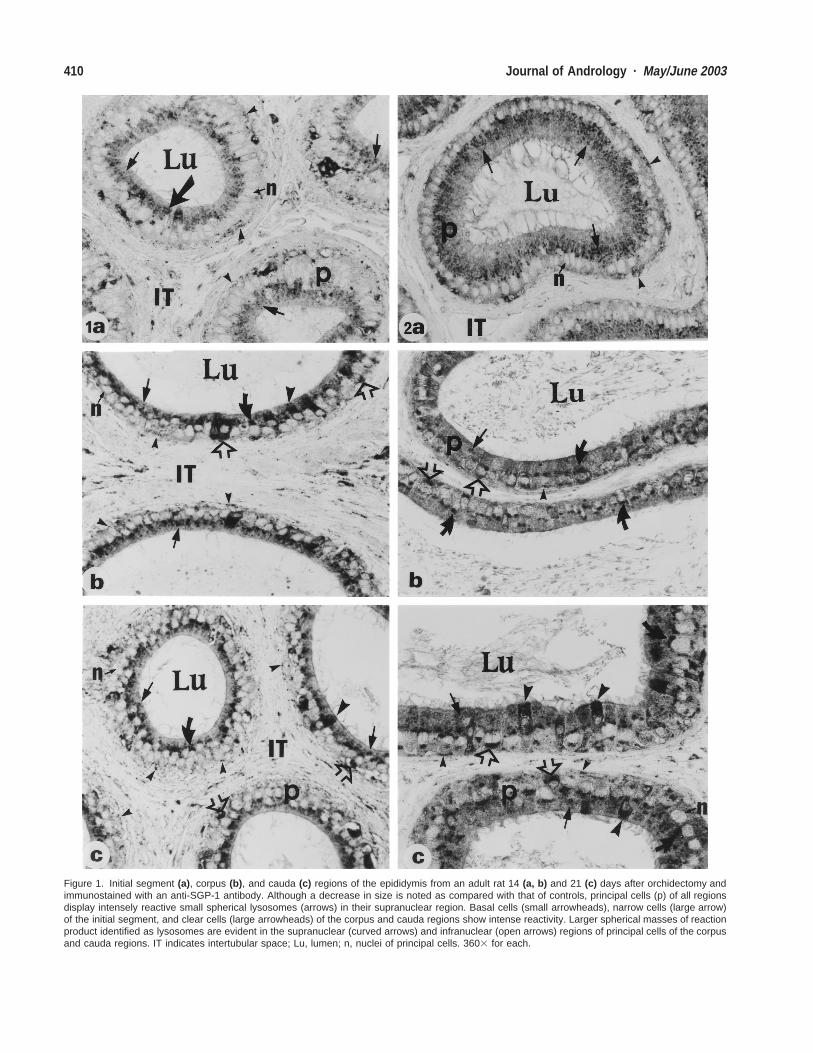
Figure 1. Initial segment (a), corpus (b), and cauda (c) regions of the epididymis from an adult rat 14 (a, b) and 21 (c) days after orchidectomy andimmunostained with an anti-SGP-1 antibody. Although a decrease in size is noted as compared with that of controls, principal cells (p) of all regionsdisplay intensely reactive small spherical lysosomes (arrows) in their supranuclear region. Basal cells (small arrowheads), narrow cells (large arrow)of the initial segment, and clear cells (large arrowheads) of the corpus and cauda regions show intense reactivity. Larger spherical masses of reactionproduct identified as lysosomes are evident in the supranuclear (curved arrows) and infranuclear (open arrows) regions of principal cells of the corpusand cauda regions. IT indicates intertubular space; Lu, lumen; n, nuclei of principal cells. 3603 for each.

411Hermo and Andonian · Regulation of SGP-1 and Cathepsin D
←
Figure 2. Initial segment (a), caput (b), and cauda (c) regions of the epididymis from an adult rat 14 days after orchidectomy and immediate testosteronesupplementation, immunostained with anti-SGP-1 antibody. Small intensely reactive lysosomes (arrows) are evident in the cytoplasm of principal cells(p) of all regions. Basal cells (small arrowheads) and clear cells (large arrowheads) are also reactive. Note larger spherical lysosomes in the supra-nuclear (curved arrows) and infranuclear (open arrows) regions of principal cells of the caput and cauda regions. IT indicates intertubular space; Lu,lumen; n, nuclei of principal cells. (a) and (b) 3753 each; (c) 4253.
A dilution factor of 1:20 and 1:100 in Tris-buffered saline(TBS) was used for the polyclonal anti-cathepsin D and anti-SGP-1 antibody, respectively. Dr M.D. Griswold initially pro-vided us with the anti-SGP-1 antibody, whereas Dr C.R. Moralesprovided us with anti-SGP-1 antibody more recently. Both an-tibodies are well-characterized and described by Sylvester et al(1989) and Morales et al (2000). The anti-cathepsin D antibodieswere purchased from Calbiochem (La Jolla, CA) and DAKO(Carpinteria, CA), and we had used them in a previous study ofthe epididymis (Igdoura et al, 1995). Each tissue section wasincubated in the primary antibody for 1.5 hours. After incuba-tion, the sections were immersed in Tween, blocked with goatserum, and subsequently incubated with goat anti-rabbit immu-noglobulin G (IgG) conjugated to peroxidase (Sigma, St LouisMO) at a dilution of 1:250 in TBS and incubated for 30 minutesat 358C in a humidified incubator. After incubation with a sec-ondary antibody, the tissue was washed by immersion in 4 wellsof Tween buffer solution for 2 minutes each.
The final reaction product was obtained by incubating theslides for 10 minutes in 250 mL of TBS containing 0.03% hy-drogen peroxide, 0.1 M imidazole, and 0.05% diaminobenzidinetetrahydrochloride (DAB) pH 7.4. The sections were counter-stained with 0.1% methylene blue (2 minutes) and then dehy-drated in a graded series of ethanol solutions (30 seconds each)and xylene (3 minutes). Cover slips were mounted onto glassslides using Permount. Incubation with normal rabbit serum ata dilution of 1:100 in TBS and incubation of tissues in secondaryantibody alone, without primary antibody, served as controls.
Routine Electron Microscopy AnalysisAn additional 9 adult male Sprague-Dawley rats (350–450 g)were used in this study for routine electron microscopy analysis.Three of the animals served as controls, and whereas another 3were bilaterally orchidectomized, the others had their efferentducts ligated on both sides; all animals were killed 14 days later.The epididymides of these animals were fixed by perfusion with2.5% glutaraldehyde in sodium cacodylate buffer via the abdom-inal aorta for 10 minutes. Thereafter, the tissue was removed,cut into the appropriate epididymal regions, and remained inbuffer overnight. On the following day, the tissue was postfixedin potassium ferrocyanide–reduced osmium tetroxide (Karnov-sky, 1971), dehydrated in alcohol and propylene oxide, and em-bedded in Epon; thick and thin sections were cut and treated asdescribed in our previous studies, with thin sections being ex-amined with a Philips electron microscope (Hermo et al, 1991a).
ResultsEffects of Orchidectomy on Expression of SGP-1 With orWithout Testosterone Supplementation,Hypophysectomy, and Efferent Duct LigationIn untreated control animals, as demonstrated in our pre-vious study, SGP-1 was expressed in lysosomes of prin-
cipal cells of all epididymal regions, and expression wasalso noted in narrow, basal, and clear cells of each epi-didymal region where these cells were located (Hermo etal, 1992). At all time points after orchidectomy (up to 21days), principal cells of all epididymal regions continuedto show small, intensely reactive spherical granules in thesupranuclear region of their cytoplasm. Indeed, despitethe reduction in size of the epididymal tubules and epi-thelial cells at the late time points after orchidectomy,these granules continued to be readily apparent and re-active for anti-SGP-1 antibody (Fig. 1a–c). Such granuleswere punctate in appearance and corresponded in positionand size to similar structures identified as lysosomes byroutine cytochemical and immunocytochemical methodsusing light and electron microscopy (Hermo et al, 1991b,1992; Robaire and Hermo, 1988; Hermo and Robaire,2002). In addition, large spherical masses of reactionproduct were observed supranuclearly in close proximityto the nucleus of some principal cells. Such masses werenot evident in control animals, and they were especiallyevident at the 14- and 21-day intervals in the caput, cor-pus, and cauda regions of the epididymis (Fig. 1b and c).Distinct large, spherical masses of reaction product alsowere evident in the infranuclear region of principal cellsclosely apposed to the nucleus, an observation that wasnot observed in control animals. As with the supranuclearmasses, they were especially evident at the 14- and 21-day intervals in the caput, corpus, and cauda regions ofthe epididymis (Fig. 1b and c). Because such sphericalmasses were reactive with both anti-SGP-1 and cathepsinD antibodies, it would appear that they correspond to ly-sosomal elements and will be referred to as large lyso-somal elements.
Basal cells also continued to express SGP-1 at all timepoints after orchidectomy, with the reaction product beingseen as small, punctate granules at the base of the epi-thelium or as thin, dense bands encompassing the nucleusof the hemispherical basal cells and stretching along theirthin, elongated processes (Fig. 1a–c). In the initial seg-ment, narrow cells also displayed intense reactivity foranti-SGP-1 antibody (Fig. 1a), as did clear cells of thecaput, corpus (Fig. 1b), and cauda (Fig. 1c) regions. Thereaction product in these cells often appeared evenly dis-tributed throughout their cytoplasm.
Orchidectomy followed by immediate testosterone sup-plementation (up to 21 days) also revealed no majorchanges to the expression of SGP-1 in epithelial cells of

412 Journal of Andrology · May/June 2003
the entire epididymis. Principal cells displayed small, in-tensely reactive spherical lysosomes in all epididymal re-gions, and narrow, basal, and clear cells remained intense-ly reactive (Fig. 2a–c). The addition of testosterone toorchidectomized rats restored the epididymal tubule andsize of the epithelial cells of the different regions to nor-mal size. However, principal cells, in addition to smallspherical lysosomes, continued to show large, sphericalsupranuclear and infranuclear lysosomal elements next tothe nucleus (Fig. 2b and c).
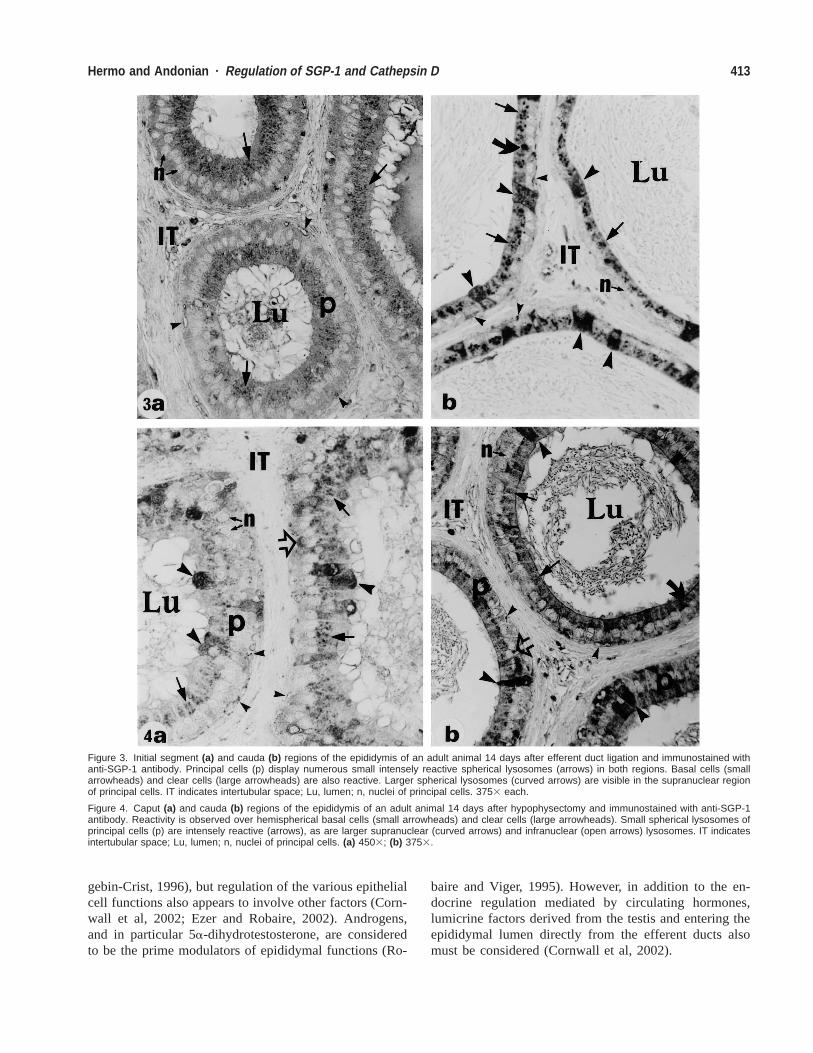
At the various time points after efferent duct ligation(up to 21 days), numerous intensely reactive spherical ly-sosomes of principal cells were observed in all epididy-mal regions; narrow, clear, and basal cells also remainedintensely reactive (Fig. 3a and b). As with orchidectomy,the large, spherical lysosomal elements in the supranucle-ar and infranuclear regions of principal cells of the epi-didymis were still prominent at the later time points afterefferent duct ligation in the caput, corpus, and cauda re-gions of the epididymis (Fig. 3b).
After hypophysectomy (4 weeks), changes in SGP-1expression were not noted in any cell type of any givenepididymal region (Fig. 4a and b). The presence of su-pranuclear and infranuclear large, spherical lysosomal el-ements was, however, evident in principal cells of thecaput, corpus, and cauda epididymal regions (Fig. 4).
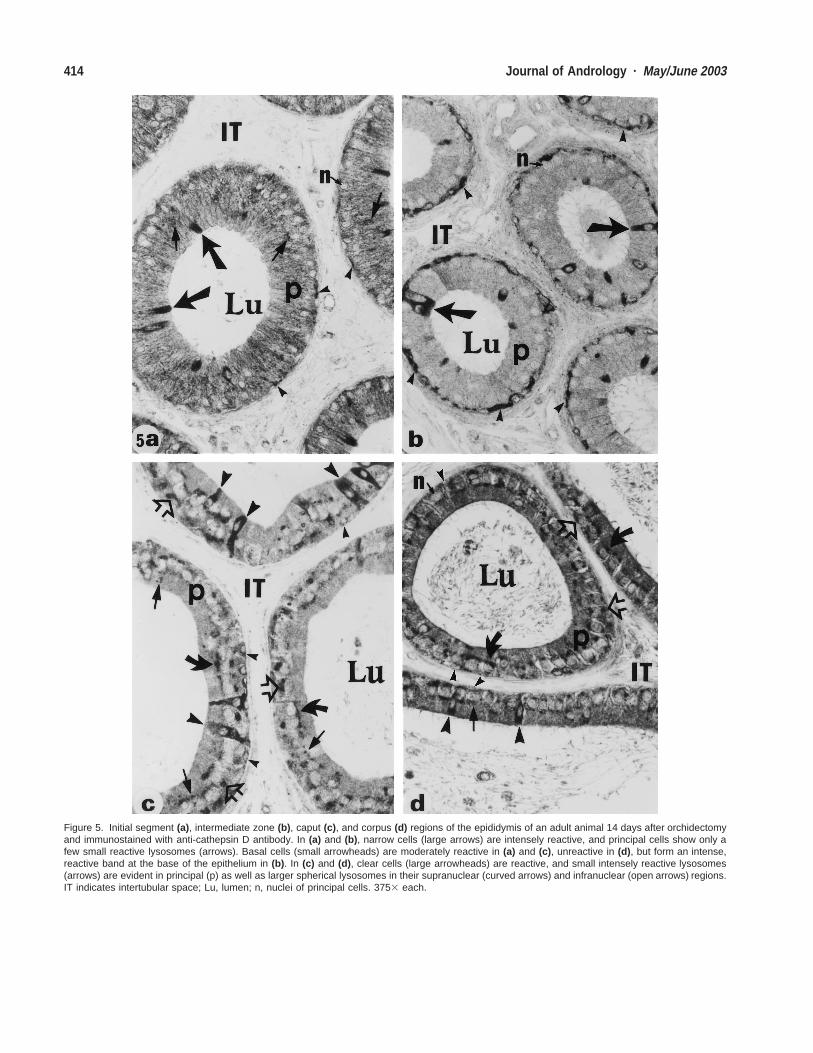
Expression of Cathepsin D in Epithelial Cells of theEpididymis After Experimental TreatmentsAfter orchidectomy, no major changes in cathepsin D ex-pression were noted in principal cells of the entire epi-didymis. There were few small reactive lysosomes in theproximal initial segment (Fig. 5a) and intermediate zone(Fig. 5b) and more in the caput (Fig. 5c), corpus (Fig.5d), and cauda (Fig. 6a) regions, as noted in control an-imals (Table). However, with SGP-1 after various treat-ments, large, spherical lysosomal elements were noted inthe supranuclear and infranuclear regions of the cyto-plasm of principal cells in the caput, corpus, and caudaregions (Fig. 5c and d; Fig. 6a). Comparable to controlanimals, narrow cells were intensely reactive for anti-ca-thepsin D antibody in the initial segment (Fig. 5a) andintermediate zone (Fig 5b) at all time points after orchi-dectomy (Table). Basal cells were intensely reactive andprominent in the intermediate zone (Fig. 5b) and mod-erately reactive in the initial segment (Fig. 5a) and caput(Fig. 5c) regions, but unreactive in the corpus (Fig. 5d)and cauda (Fig. 6a) epididymidis (Table). Clear cells con-tinued to be intensely reactive in the caput region (Fig.5c), comparable to that noted in control animals. How-ever, after orchidectomy, at various time points clear cellsbecame intensely reactive in the corpus (Fig. 5d) and cau-da (Fig. 6a) regions, where they were consistently unre-active in control animals (Table).
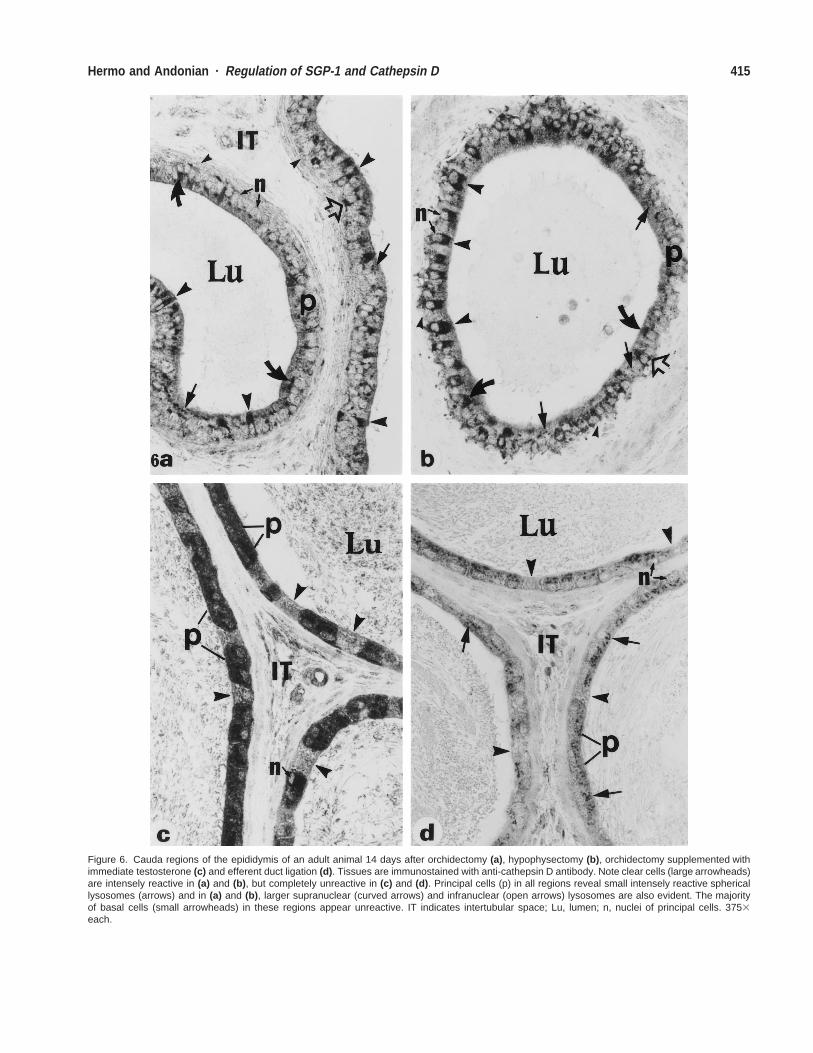
After hypophysectomy, cathepsin D expression was un-altered and comparable to that of control animals in prin-cipal, narrow, and basal cells (Fig. 6b, Table). As notedafter orchidectomy, large, spherical supranuclear and in-franuclear lysosomal elements were observed in principalcells (Fig. 6b). In addition, clear cells that were unreactivein control animals were intensely reactive after all timepoints after hypophysectomy (Fig. 6b, Table).
Orchidectomized animals supplemented immediatelywith testosterone (Fig. 6c), as well as efferent duct-ligatedanimals (Fig. 6d) also revealed no changes in cathepsin Dexpression in principal, narrow, and basal cells comparedwith that of control animals at all time points examined(Table). However, with both treatments, clear cells in cor-pus and cauda epididymides were completely unreactive(Fig. 6c and d), as was the case in control animals (Table).
When tissue sections were treated with normal rabbitserum or without the primary antibody, there was a com-plete absence of reaction over the epithelium or intertu-bular space. We have already published photographs ofcontrols for SGP-1 and cathepsin D (Hermo et al, 1992;Igdoura et al, 1995), and similar images were consistentlynoted in all our many control preparations.
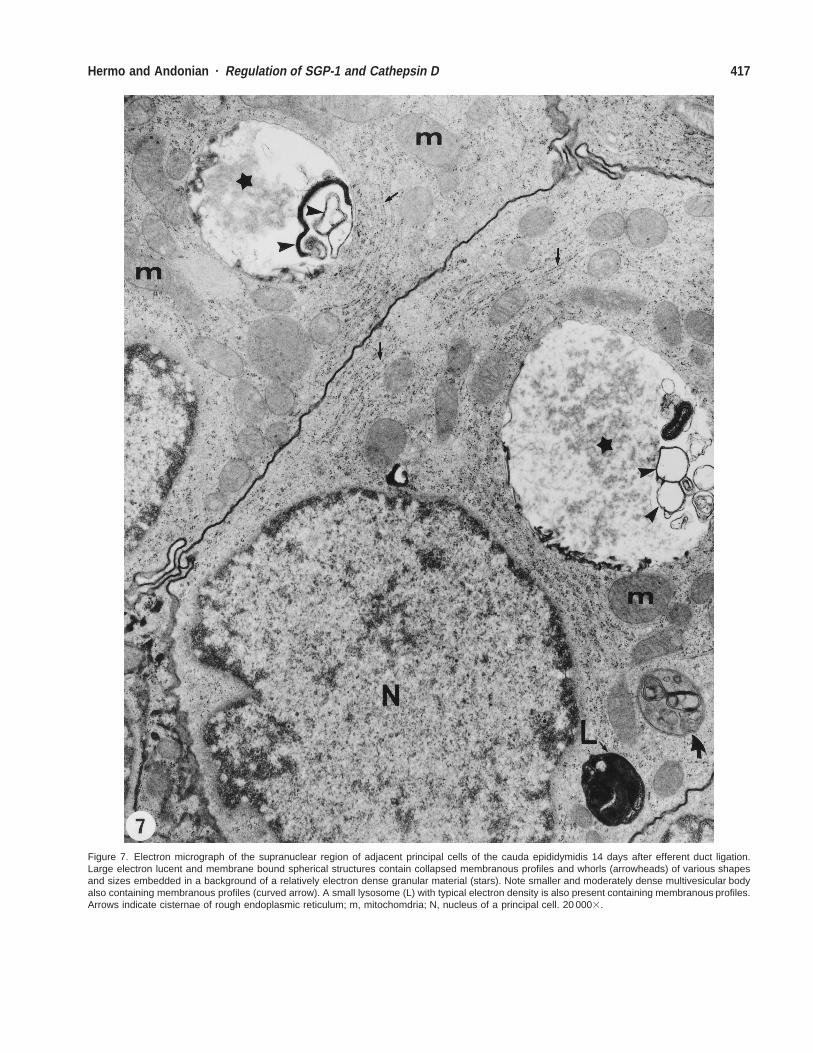
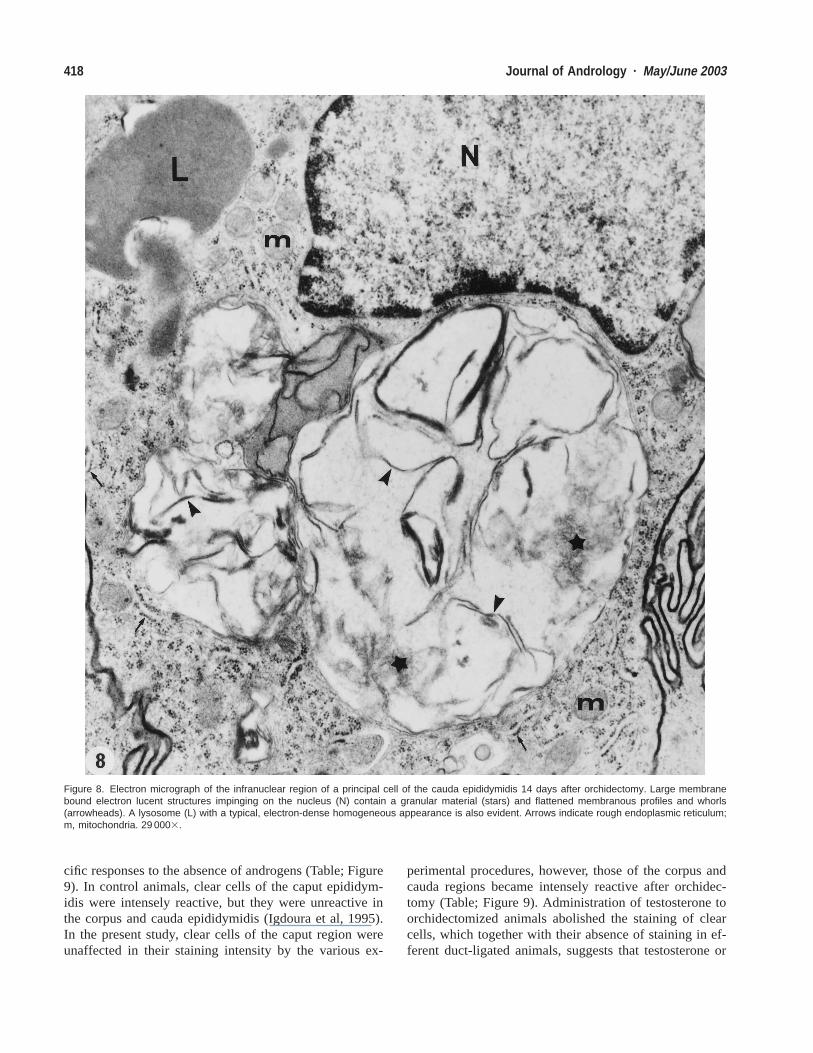
Electron Microscopic Observations of Principal CellsAfter Orchidectomy and Efferent Duct LigationTo further understand the nature of the large supranuclearand infranuclear lysosomes, electron microscopy wasused. At 14 days after efferent duct ligation or orchidec-tomy, principal cells of the caput, corpus, and cauda ep-ididymidis revealed large membrane-bound electron-lu-cent, spherical structures (Figs. 7 and 8). Such structurescontained flattened membranous profiles and whorls ofvarious sizes and shapes embedded in a relatively elec-tron-dense, granular material (Figs. 7 and 8). Occupyinga position next to the nucleus, these structures were large,at times approximating that of the nucleus. They were notthat abundant and whereas 1 or 2 could be seen in someprincipal cells, other cells showed none. Adjacent to thesestructures were typical, electron-dense lysosomes, as wellas multivesicular bodies (Figs. 7 and 8); their size andrelative numbers did not differ from those noted in controlanimals. These large supranuclear and infranuclear struc-tures were immunostained with both anti-SGP-1 and ca-thepsin D antibodies as seen with light microscopy, in-dicating that they corresponded to lysosomal elements.
Discussion
Regulation of SGP-1 Expression in the EpididymisIt is well established that many epididymal functions areunder the control of androgens (Orgebin-Crist et al, 1975;Robaire and Hermo, 1988; Robaire and Viger, 1995; Or-
413Hermo and Andonian · Regulation of SGP-1 and Cathepsin D
Figure 3. Initial segment (a) and cauda (b) regions of the epididymis of an adult animal 14 days after efferent duct ligation and immunostained withanti-SGP-1 antibody. Principal cells (p) display numerous small intensely reactive spherical lysosomes (arrows) in both regions. Basal cells (smallarrowheads) and clear cells (large arrowheads) are also reactive. Larger spherical lysosomes (curved arrows) are visible in the supranuclear regionof principal cells. IT indicates intertubular space; Lu, lumen; n, nuclei of principal cells. 3753 each.
Figure 4. Caput (a) and cauda (b) regions of the epididymis of an adult animal 14 days after hypophysectomy and immunostained with anti-SGP-1antibody. Reactivity is observed over hemispherical basal cells (small arrowheads) and clear cells (large arrowheads). Small spherical lysosomes ofprincipal cells (p) are intensely reactive (arrows), as are larger supranuclear (curved arrows) and infranuclear (open arrows) lysosomes. IT indicatesintertubular space; Lu, lumen; n, nuclei of principal cells. (a) 4503; (b) 3753.
gebin-Crist, 1996), but regulation of the various epithelialcell functions also appears to involve other factors (Corn-wall et al, 2002; Ezer and Robaire, 2002). Androgens,and in particular 5a-dihydrotestosterone, are consideredto be the prime modulators of epididymal functions (Ro-
baire and Viger, 1995). However, in addition to the en-docrine regulation mediated by circulating hormones,lumicrine factors derived from the testis and entering theepididymal lumen directly from the efferent ducts alsomust be considered (Cornwall et al, 2002).
414 Journal of Andrology · May/June 2003
Figure 5. Initial segment (a), intermediate zone (b), caput (c), and corpus (d) regions of the epididymis of an adult animal 14 days after orchidectomyand immunostained with anti-cathepsin D antibody. In (a) and (b), narrow cells (large arrows) are intensely reactive, and principal cells show only afew small reactive lysosomes (arrows). Basal cells (small arrowheads) are moderately reactive in (a) and (c), unreactive in (d), but form an intense,reactive band at the base of the epithelium in (b). In (c) and (d), clear cells (large arrowheads) are reactive, and small intensely reactive lysosomes(arrows) are evident in principal (p) as well as larger spherical lysosomes in their supranuclear (curved arrows) and infranuclear (open arrows) regions.IT indicates intertubular space; Lu, lumen; n, nuclei of principal cells. 3753 each.
415Hermo and Andonian · Regulation of SGP-1 and Cathepsin D
Figure 6. Cauda regions of the epididymis of an adult animal 14 days after orchidectomy (a), hypophysectomy (b), orchidectomy supplemented withimmediate testosterone (c) and efferent duct ligation (d). Tissues are immunostained with anti-cathepsin D antibody. Note clear cells (large arrowheads)are intensely reactive in (a) and (b), but completely unreactive in (c) and (d). Principal cells (p) in all regions reveal small intensely reactive sphericallysosomes (arrows) and in (a) and (b), larger supranuclear (curved arrows) and infranuclear (open arrows) lysosomes are also evident. The majorityof basal cells (small arrowheads) in these regions appear unreactive. IT indicates intertubular space; Lu, lumen; n, nuclei of principal cells. 3753each.
416 Journal of Andrology · May/June 2003
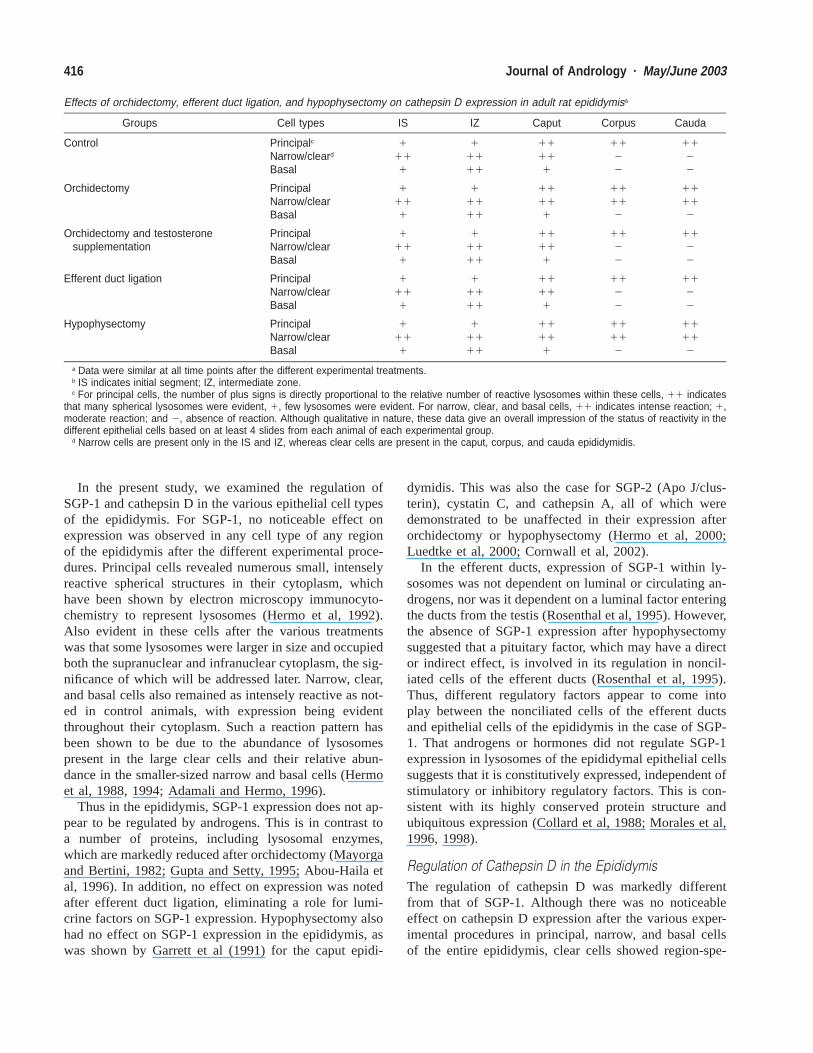
Effects of orchidectomy, efferent duct ligation, and hypophysectomy on cathepsin D expression in adult rat epididymisa
Groups Cell types IS IZ Caput Corpus Cauda
Control Principalc
Narrow/cleard
Basal
1111
11111
11111
1122
1122
Orchidectomy PrincipalNarrow/clearBasal
1111
11111
11111
11112
11112
Orchidectomy and testosteronesupplementation
PrincipalNarrow/clearBasal
1111
11111
11111
1122
1122
Efferent duct ligation PrincipalNarrow/clearBasal
1111
11111
11111
1122
1122
Hypophysectomy PrincipalNarrow/clearBasal
1111
11111
11111
11112
11112
a Data were similar at all time points after the different experimental treatments.b IS indicates initial segment; IZ, intermediate zone.c For principal cells, the number of plus signs is directly proportional to the relative number of reactive lysosomes within these cells, 11 indicates
that many spherical lysosomes were evident, 1, few lysosomes were evident. For narrow, clear, and basal cells, 11 indicates intense reaction; 1,moderate reaction; and 2, absence of reaction. Although qualitative in nature, these data give an overall impression of the status of reactivity in thedifferent epithelial cells based on at least 4 slides from each animal of each experimental group.
d Narrow cells are present only in the IS and IZ, whereas clear cells are present in the caput, corpus, and cauda epididymidis.
In the present study, we examined the regulation ofSGP-1 and cathepsin D in the various epithelial cell typesof the epididymis. For SGP-1, no noticeable effect onexpression was observed in any cell type of any regionof the epididymis after the different experimental proce-dures. Principal cells revealed numerous small, intenselyreactive spherical structures in their cytoplasm, whichhave been shown by electron microscopy immunocyto-chemistry to represent lysosomes (Hermo et al, 1992).Also evident in these cells after the various treatmentswas that some lysosomes were larger in size and occupiedboth the supranuclear and infranuclear cytoplasm, the sig-nificance of which will be addressed later. Narrow, clear,and basal cells also remained as intensely reactive as not-ed in control animals, with expression being evidentthroughout their cytoplasm. Such a reaction pattern hasbeen shown to be due to the abundance of lysosomespresent in the large clear cells and their relative abun-dance in the smaller-sized narrow and basal cells (Hermoet al, 1988, 1994; Adamali and Hermo, 1996).
Thus in the epididymis, SGP-1 expression does not ap-pear to be regulated by androgens. This is in contrast toa number of proteins, including lysosomal enzymes,which are markedly reduced after orchidectomy (Mayorgaand Bertini, 1982; Gupta and Setty, 1995; Abou-Haila etal, 1996). In addition, no effect on expression was notedafter efferent duct ligation, eliminating a role for lumi-crine factors on SGP-1 expression. Hypophysectomy alsohad no effect on SGP-1 expression in the epididymis, aswas shown by Garrett et al (1991) for the caput epidi-
dymidis. This was also the case for SGP-2 (Apo J/clus-terin), cystatin C, and cathepsin A, all of which weredemonstrated to be unaffected in their expression afterorchidectomy or hypophysectomy (Hermo et al, 2000;Luedtke et al, 2000; Cornwall et al, 2002).
In the efferent ducts, expression of SGP-1 within ly-sosomes was not dependent on luminal or circulating an-drogens, nor was it dependent on a luminal factor enteringthe ducts from the testis (Rosenthal et al, 1995). However,the absence of SGP-1 expression after hypophysectomysuggested that a pituitary factor, which may have a director indirect effect, is involved in its regulation in noncil-iated cells of the efferent ducts (Rosenthal et al, 1995).Thus, different regulatory factors appear to come intoplay between the nonciliated cells of the efferent ductsand epithelial cells of the epididymis in the case of SGP-1. That androgens or hormones did not regulate SGP-1expression in lysosomes of the epididymal epithelial cellssuggests that it is constitutively expressed, independent ofstimulatory or inhibitory regulatory factors. This is con-sistent with its highly conserved protein structure andubiquitous expression (Collard et al, 1988; Morales et al,1996, 1998).
Regulation of Cathepsin D in the EpididymisThe regulation of cathepsin D was markedly differentfrom that of SGP-1. Although there was no noticeableeffect on cathepsin D expression after the various exper-imental procedures in principal, narrow, and basal cellsof the entire epididymis, clear cells showed region-spe-
417Hermo and Andonian · Regulation of SGP-1 and Cathepsin D
Figure 7. Electron micrograph of the supranuclear region of adjacent principal cells of the cauda epididymidis 14 days after efferent duct ligation.Large electron lucent and membrane bound spherical structures contain collapsed membranous profiles and whorls (arrowheads) of various shapesand sizes embedded in a background of a relatively electron dense granular material (stars). Note smaller and moderately dense multivesicular bodyalso containing membranous profiles (curved arrow). A small lysosome (L) with typical electron density is also present containing membranous profiles.Arrows indicate cisternae of rough endoplasmic reticulum; m, mitochomdria; N, nucleus of a principal cell. 20 0003.
418 Journal of Andrology · May/June 2003
Figure 8. Electron micrograph of the infranuclear region of a principal cell of the cauda epididymidis 14 days after orchidectomy. Large membranebound electron lucent structures impinging on the nucleus (N) contain a granular material (stars) and flattened membranous profiles and whorls(arrowheads). A lysosome (L) with a typical, electron-dense homogeneous appearance is also evident. Arrows indicate rough endoplasmic reticulum;m, mitochondria. 29 0003.
cific responses to the absence of androgens (Table; Figure9). In control animals, clear cells of the caput epididym-idis were intensely reactive, but they were unreactive inthe corpus and cauda epididymidis (Igdoura et al, 1995).In the present study, clear cells of the caput region wereunaffected in their staining intensity by the various ex-
perimental procedures, however, those of the corpus andcauda regions became intensely reactive after orchidec-tomy (Table; Figure 9). Administration of testosterone toorchidectomized animals abolished the staining of clearcells, which together with their absence of staining in ef-ferent duct-ligated animals, suggests that testosterone or

419Hermo and Andonian · Regulation of SGP-1 and Cathepsin D
Figure 9. Schematic diagram of the staining pattern for cathepsin D inthe initial segment (IS), intermediate zone (IZ), caput (Cap), corpus (Cor),and cauda (Cau) epididymidis after efferent duct ligation (EDL), orchi-dectomy (O), orchidectomy and testosterone supplementation (O1T)and hypophysectomy (H). The same pattern of staining is noted at alltime points after experimental treatments. Principal cells are representedas columnar cells with microvillar brush border and are present in allepididymal regions. Narrow cells, found only in the initial segment andintermediate zone, are represented as attenuated cells at the right marginof the epithelial section. Clear cells, present only in the caput, corpus,and cauda epididymidis, are represented as columnar cells without mi-crovilli. The hemispherical basal cells are noted in all epididymal regions.The small dark dots correspond to the relative number of reactive lyso-somes within principal cells. The larger dark dots represent the largerspherical lysosomes noted in the supranuclear and infranuclear regionsof principal cells after the various experimental treatments. In narrow,clear, and basal cells, black staining over the cytoplasm represents anintense reaction, whereas moderate staining represents a moderate re-action, and no staining indicates the absence of a reaction. The intensereaction is the result of these cells containing numerous lysosomes, re-sulting in what appears as a uniform staining over the cytoplasm. Withthe exception of the large spherical supranuclear and infranuclear lyso-somes, the reaction depicted in the EDL/O1T column also reflects thatseen in the control group. Note that clear cells of the caput epididymidisare unchanged in their expression of cathepsin D after all experimental
←
treatments. However, clear cells of the corpus and cauda epididymidis,although unreactive in the control, EDL, and O1T groups, become in-tensely reactive in the O and H groups, suggesting that testosterone orone of its metabolites down-regulates its expression in control animals.
one of its metabolites inhibits the expression of cathepsinD in clear cells of these regions (Table; Figure 9). It isinteresting that the enzymatic activity of cathepsin D inthe epididymis has been shown to be increased abovenormal levels after orchidectomy and decreased after tes-tosterone treatment (Mayorga and Bertini, 1982), coincid-ing with our present immunocytochemical data. However,our study indicates that the specific cell type affected isthe clear cell and that it is affected in a region-specificmanner. Other proteins that demonstrated greater enzy-matic activity in the epididymis after androgen withdraw-al, often in a region-specific manner, include glucuroni-dase, RNase II, transforming growth factor b-1, gammaglutamyl-transpeptidase III, and SGP-2 (Mayorga andBertini, 1982; Cyr and Robaire, 1992; Palladino and Hin-ton, 1994; Gupta and Setty, 1995; Desai and Kondaiah,2000).
Whereas androgens play a prominent role in regulatinga variety of epididymal functions, a role for estrogens hasalso been implicated in the efferent ducts and epididymis,where estrogen receptors (ERs) a and b have been located(Meistrich et al, 1975; Orgebin-Crist et al, 1983; Fisheret al, 1997; Hess et al, 1997). In fact, in mice, the corpusand cauda epididymidis revealed a much denser labelingof the nuclei of clear cells with 3H-estradiol than withtestosterone, suggesting that estradiol may serve to mod-ulate clear cell function in the epididymis (Schleicher etal, 1984).
In the present study, cathepsin D expression in clearcells after orchidectomy became intense, but supplement-ing testosterone to these animals resulted in a decrease instaining, as was observed in control animals. It could beargued that estrogen rather than testosterone may suppressthe expression of cathepsin D in clear cells of the corpusand cauda epididymidis. With acid phosphatase activityI, Nikkanen and Vanha-Perttula (1977) also noted thatthere was an increase in activity with estrogen treatmentin the epididymis. Future studies employing ER a and bmouse knockout models in conjunction with immunocy-tochemistry would help resolve the role of estrogen oncathepsin D expression in clear cells, studies that are be-yond the scope of the present investigation.
In the present study, we acknowledge that the immu-nostaining pattern of SGP-1 and cathepsin D reflectssteady state protein levels and that our methods do notindicate differences in protein production. In addition, itis possible that protein degradation occurs following ourdifferent treatments, but that the proteins (degradation

420 Journal of Andrology · May/June 2003
fragments) remain immunoreactive. If this is the case, bi-ological function of each protein could be affected butbased on immunocytochemistry, no changes in expressionwould be detected. Another approach such as Westernblot analysis would reveal protein degradation and couldbe addressed in future studies along with quantitativeelectron microscopy immunocytochemistry. However,both of these approaches are beyond the scope of thepresent study.
Electron Microscopic Observations on Lysosomes ofPrincipal Cells After Various Experimental ProceduresOrchidectomy and hypophysectomy causes the epididy-mal weight to decrease (Robaire et al, 1977), accompa-nied by a decrease in luminal diameter of the tubules anddecrease in epithelial cell height (Orgebin-Crist et al,1975; Delongeas et al, 1987 Hermo and Papp, 1996).Principal cells are particularly sensitive to androgen lev-els, showing morphological changes such as accumulationof lysosomes, unlike that noted for the other epithelialcells, which appear to be unaffected (Moore and Bedford,1979).
For SGP-1 and cathepsin D alike, after all procedures,large spherical structures became evident in the supra-nuclear and infranuclear regions of principal cells. Suchstructures were highly reactive for anti-SGP-1 and anti-cathepsin D antibodies, suggesting that they correspondedto lysosomal elements. Using electron microscopy, thesestructures corresponded to large, membrane-bound spher-ical structures with electron-lucent content in which mem-branous profiles of various shapes and sizes and a gran-ular material were evident. Such structures were not notedin control animals.
Although it is not fully clear as to why such structuresbecome prominent, one hypothesis that could be proposedis the adverse effect that orchidectomy, efferent duct li-gation, and hypophysectomy have on expression of a va-riety of proteins and genes in the absence of androgens,luminal factors, or both emanating from the testis (Ro-baire and Viger, 1995; Cornwall et al, 2002; Ezer andRobaire, 2002). In fact, several lysosomal enzymes suchas acid phosphatase, N-acetyl-beta-D-glucosaminidase,beta-glucuronidase, N-acetylhexosaminidase, and arylsul-phatase, show a decreased enzymatic activity after orchi-dectomy (Mayorga and Bertini, 1982; Gupta and Setty,1995; Abou-Haila et al, 1996). Thus the reduction in thisactivity could result in the accumulation of substrates nor-mally acted on by these enzymes and result in a pheno-type as noted for various lysosomal storage diseases(Neufeld et al, 1975; Hammel and Alroy, 1995; Trasleret al, 1998). The latter have been shown to result in adramatic increase in number and size of lysosomes and achange in their appearance and location within principalcells (Adamali et al, 1999a,b). However, in the present
study, the effect was not noted to be as dramatic as it wasfor a given lysosomal gene knockout. This may be ex-plained by enzymatic activities of these enzymes beingmerely reduced after the various procedures and not com-pletely abolished (Mayorga and Bertini, 1982). Neverthe-less, a somewhat similar phenotype for lysosomes in prin-cipal cells appears to occur as a result of the adverseeffects of orchidectomy, efferent duct ligation, and hy-pophysectomy on expression of different proteins andgenes in the epididymis. Finally, while the significancefor up-regulation of cathepsin D after orchidectomy andhypophysectomy is not known, it may be suggested thatcathepsin D expression compensates for the reduction inactivity and expression of other lysosomal enzymes re-ported to occur after orchidectomy (Mayorga and Bertini,1982; Gupta and Setty, 1995; Abou-Haila et al, 1996).
AcknowledgmentsThe technical assistance of Lisa Discepola, Stephen Tepper, and JeannieMui is gratefully acknowledged. We thank Dr M.D. Griswold (Washing-ton State University, Pullman) and Dr C.R. Morales (Department of Anat-omy and Cell Biology, McGill University) for their generous supply ofthe anti-SGP-1 antibody.
ReferencesAbou-Haila A, Tulsiani DRP, Skudlarek MD, Orgebin-Crist M-C. Andro-
gen regulation of molecular forms of beta-D-glucuronidase in themouse epididymis: comparison with liver and kidney. J Androl. 1996;17:194–207.
Adamali HI, Hermo L. Apical and narrow cells are distinct cell typesdiffering in their structure, distribution and functions in the adult ratepididymis. J Androl. 1996;17:208–222.
Adamali HI, Somani IH, Huang J-Q, Mahuran D, Gravel RA, Trasler JM,Hermo L. I. Abnormalities in cells of the testis, efferent ducts andepididymis in juvenile and adult mice with beta-hexosaminidase Aand B deficiency. J Androl. 1999a;20:779–802.
Adamali HI, Somani IH, Huang J-Q, Mahuran D, Gravel RA, Trasler JM,Hermo L. II. Characterization and development of the regional- andcellular-specific abnormalities in the epididymis of mice with beta-hexosaminidase A deficiency. J Androl. 1999b;20:803–824.
Brawer JR, Schipper H, Robaire B. Effects of long term androgen andestradiol exposure on the hypothalamus. Endocrinology. 1983;112:194–199.
Collard MW, Sylvester SR, Tsuruta JK, Griswold MD. Biosynthesis andmolecular cloning of sulfated glycoprotein 1 secreted by rat Sertolicells: sequence similarity with the 70-kilodalton precursor to sulfatide/GM1 activator. Biochemistry. 1988;27:4557–4564.
Cooper TG. The Epididymis, Sperm Maturation and Fertilization. Berlin:Springer-Verlag; 1986.
Cornwall GA, Hann SR. Specialized gene expression in the epididymis.J Androl. 1995;16:379–383.
Cornwall GA, Lareyre J-J, Matusik RJ, Hinton BT, Orgebin-Crist M-C.Gene expression and epididymal function. In: Robaire B, Hinton BT,eds. The Epididymis: From Molecules To Clinical Practice. NewYork: Kluwer Academic/Plenum; 2002:169–199.
Cyr DG, Robaire B. Regulation of sulfated glycoprotein-2 (clusterin)

421Hermo and Andonian · Regulation of SGP-1 and Cathepsin D
messenger ribonucleic acid in the rat epididymis. Endocrinology.1992;130:2160–2166.
Delongeas JL, Gelly JL, Leheup B, Grignon G. Influence of testicularsecretions on differentiation in the rat epididymis: ultrastructural stud-ies after castration, efferent duct ligation and cryptorchidism. Exp CellBiol. 1987;55:74–82.
Desai KV, Kondaiah P. Androgen ablation results in differential regulationof transforming growth factor-beta isoforms in rat male accessory sexorgans and epididymis. J Mol Endocrinol. 2000;24:253–260.
Dunn KW, Maxfield FR. Delivery of ligands from sorting endosomes tolate endosomes occurs by maturation of sorting endosomes. J CellBiol. 1992;117:301–310.
Ezer N, Robaire B. Androgenic regulation of the structure and functionsof the epididymis. In: Robaire B, Hinton BT, eds. The Epididymis:From Molecules To Clinical Practice. New York: Kluwer Academic/Plenum; 2002:297–316.
Fisher JS, Millar MR, Majdic G, Saunders PT, Fraser HM, Sharpe RM.Immunolocalisation of oestrogen receptor-alpha within the testis andexcurrent ducts of the rat and marmoset monkey from perinatal lifeto adulthood. J Endocrinol. 1997;153:485–495.
Garrett SH, Garrett JE, Douglass J. In situ histochemical analysis of re-gion-specific gene expression in the adult rat epididymis. Mol ReprodDev. 1991;30:1–17.
Gupta G, Setty BS. Activities and androgenic regulation of lysosomalenzymes in the epididymis of rhesus monkey. Endocr Res. 1995;21:733–741.
Hamilton DW. Structure and function of the epithelium lining the ductuliefferentes, ductus epididymis and ductus deferens in the rat. In: GreepRO, Astwood EB, eds. Handbook of Physiology. Section 7, Vol 5.Washington, DC: American Physiological Society; 1975:303–317.
Hammel I, Alroy J. The effect of lysosomal storage diseases on secretorycells: an ultrastructural study of pancreas as an example. J SubmicroscCytol Pathol. 1995;27:143–60.
Hermo L, Dworkin J, Oko R. Role of epithelial clear cells of the ratepididymis in the disposal of the contents of cytoplasmic dropletsdetached from spermatozoa. Am J Anat. 1988;183:107–124.
Hermo L, Green H, Clermont Y. Golgi apparatus of epithelial principalcells of the epididymal initial segment of the rat: structure, relation-ship with endoplasmic reticulum, and role in the formation of secre-tory vesicles. Anat Rec. 1991a;229:159–176.
Hermo L, Morales CR, Oko R. Immunocytochemical localization of sul-fated glycoprotein-1 (SGP-1) and identification of its transcripts inepithelial cells of the extratesticular duct system of the rat. Anat Rec.1992;232:401–422.
Hermo L, Oko R, Morales CR. Secretion and endocytosis in the malereproductive tract: a role in sperm maturation. Int Rev Cytol. 1994;154:105–189.
Hermo L, Papp S. Effects of ligation, orchidectomy, and hypophysectomyon expression of the Yf subunit of GST-P in principal and basal cellsof the adult rat epididymis and on basal cell shape and overall ar-rangement. Anat Rec. 1996;244:59–69.
Hermo L, Robaire B. Epididymal cell types and their function. In: Ro-baire B, Hinton BT, eds. The Epididymis: From Molecules To ClinicalPractice. New York: Kluwer Academic/Plenum; 2002:81–102.
Hermo L, Wright J, Oko R, Morales CR. Role of epithelial cells of themale excurrent duct system of the rat in the endocytosis or secretionof sulfated glycoprotein-2 (clusterin). Biol Reprod. 1991b;44:1113–1131.
Hermo L, Xiaohong S, Morales CR. Circulating and luminal testicularfactors affect LRP-2 and Apo J expression in the epididymis follow-ing efferent duct ligation. J Androl. 2000;21:122–144.
Hess RA, Gist DH, Bunick D, Lubahn DB, Farrell A, Bahr J, Cooke PS,Greene GL. Estrogen receptor (alpha and beta) expression in the ex-
current ducts of the adult male rat reproductive tract. J Androl. 1997;18:602–611.
Hess RA, Bunick D, Lubahn DB, Zhou Q, Bouma J. Morphologic chang-es in efferent ductules and epididymis in estrogen receptor-alphaknockout mice. J Androl. 2000;21:107–121.
Hess RA, Zhou Q, Nie R. The role of estrogens in the endocrine andparacrine regulation of the efferent ductules, epididymis and vas de-ferens. In: Robaire B, Hinton BT, eds. The Epididymis: From Mole-cules To Clinical Practice. New York: Kluwer Academic/Plenum;2002:317–337.
Igdoura SA, Morales CR, Hermo L. Differential expression of cathepsinsB and D in testis and epididymis of adult rats. J Histochem Cytochem.1995;43:545–557.
Karnovsky MJ. Use of ferrocyanide-reduced osmium tetroxide in electronmicroscopy. In: Proceedings of the 11th Meeting of the AmericanSociety of Cell Biology New Orleans, La; 1971; abstract 284.
Kirschke H, Langner J, Reimann S, Weideranders B, Ansorge S, BohleyP. Lysosomal cysteine proteinases. In: Evered D, Whelan J, eds. Pro-tein Degradation in Health and Disease. Ciba Foundation Sympo-sium 75. Amsterdam: Excerpta Medica; 1980:15–35.
Kominami E, Ueno T, Muno D, Katunuma N. The selective role of ca-thepsins B and D in the lysosomal degradation of exogenous andendogenous proteins. FEBS Lett. 1991;287:189–192.
Kretz KA, Carson GS, Morimoto S, Kishimoto Y, Fluharty AL, O’BrienJS. Characterization of a mutation in a family with saposin B defi-ciency: a glycosylation site defect. Proc Natl Acad Sci USA. 1990;87:2541–2544.
Luedtke CC, Andonian S, Igdoura S, Hermo L. Cathepsin A is expressedin a cell- and region-specific manner in the testis and epididymis andis not regulated by testicular or pituitary factors. J Histochem Cyto-chem. 2000;48:1131–1146.
Mayorga LS, Bertini F. Effect of androgens on the activity of acid hy-drolases in rat epididymis. Int J Androl. 1982;5:345–352.
Meistrich ML, Hughes TH, Bruce WR. Alteration of epididymal spermtransport and maturation in mice by oestrogen and testosterone. Na-ture. 1975;258:145–147.
Moore HD, Bedford JM. The differential absorptive activity of epithelialcells of the rat epididymis before and after castration. Anat Rec. 1979;193:313–327.
Morales CR, El-Alfy M, Zhao Q, Igdoura SA. Expression and tissuedistribution of rat sulfated glycoprotein-1 (prosaposin). J HistochemCytochem. 1996;44:327–337.
Morales CR, Hay N, El-Alfy M, Zhao Q. Distribution of mouse sulfatedglycoprotein-1 (prosaposin) in the testis and other tissues. J Androl.1998;19:156–164.
Morales CR, Zhao Q, El-Alfy M, Suzuki K. Targeted disruption of themouse prosaposin gene affects the development of the prostate glandand other male reproductive organs. J Androl. 2000;21:765–775.
Murphy RF. Maturation models for endosome and lysosome biogenesis.Trends Cell Biol. 1991;1:77–82.
Neufeld EF, Lim TW, Shapiro LJ. Inherited disorders of lysosomal me-tabolism. Ann Rev Biochem. 1975;44:345–357.
Nikkanen V, Vanha-Perttula T. Acid phosphatase of the rat epididymis.III. Histochemical and biochemical responses in experimental condi-tions. Andrologia. 1977;9:357–370.
O’Brien JS, Kishimoto Y. Saposin proteins: structure, function, and rolein human lysosomal storage disorders. FASEB J. 1991;5:301–308.
Orgebin-Crist M-C. Androgens and epididymal function. In: Bhasin S,Gabelnick H, Spieler G, Swerdloff R, Wand C, eds. Pharmacology,Biology, and Clinical Application of Androgens. New York: Wiley-Liss; 1996:27–38.
Orgebin-Crist M-C, Danzo BJ, Davies J. Endocrine control of the devel-opment and maintenance of sperm fertilizing ability in the epididymis.In: Greep RO, Astwood EB, eds. Handbook of Physiology. Section

422 Journal of Andrology · May/June 2003
7, Vol 5. Washington, DC: American Physiological Society; 1975:319–338.
Orgebin-Crist MC, Eller BC, Danzo BJ. The effects of estradiol, tamox-ifen, and testosterone on the weights and histology of the epididymisand accessory sex organs of sexually immature rabbits. Endocrinol-ogy. 1983;113:1703–1715.
Palladino MA, Hinton BT. Expression of multiple gamma-glutamyl trans-peptidase messenger ribonucleic acid transcripts in the adult rat epi-didymis is differentially regulated by androgens and testicular factorsin a region-specific manner. Endocrinology. 1994;135:1146–1156.
Raczek S, Yeung CH, Hasilik A, Robenek H, Hertle L, Schulze H, CooperTG. Immunocytochemical localisation of some lysosomal hydrolases,their presence in luminal fluid and their directional secretion by hu-man epididymal cells in culture. Cell Tissue Res. 1995;280:415–425.
Robaire B, Ewing LL, Zirkin BR, Irby DC. Steroid d4-5a-reductase and3a-hydroxysteroid dehydrogenase in the rat epididymis. Endocrinol-ogy. 1977;101:1379–1390.
Robaire B, Hermo L. Efferent ducts, epididymis, and vas deferens: struc-ture, functions, and their regulation. In: Knobil E, Neill J, eds. ThePhysiology of Reproduction. New York: Raven Press; 1988:999–1080.
Robaire B, Viger RS. Regulation of epididymal epithelial cell functions.Biol Reprod. 1995;52:226–236.
Rosenthal AL, Igdoura SA, Morales CR, Hermo L. Hormonal regulationof sulfated glycoprotein-1 synthesis by nonciliated cells of the efferentducts of adult rats. Mol Reprod Dev. 1995;40:69–83.
Schleicher G, Drews U, Stumpf WE, Sar M. Differential distribution ofdihydrotestosterone and estradiol binding sites in the epididymis ofthe mouse. An autoradiographic study. Histochemistry. 1984;81:139–147.
Srivastava PN, Ninjoor V. Isolation and characterization of cathepsin Dfrom rabbit testis. Biochem Biophys Res Commun. 1982;109:63–69.
Stratton ID, Ewing LL, Desjardins C. Efficacy of testosterone-filled po-lydimethylsiloxane implants in maintaining plasma testosterone inrabbits. J Reprod Fertil. 1973;35:235–244.
Suarez-Quian CA, Jelesoff N, Byers SW. Lysosomal integral membraneproteins exhibit region and cell type specific distribution in the epi-didymis of the adult rat. Anat Rec. 1992;232:85–96.
Sun Y, Witte DP, Grabowski GA. Developmental and tissue-specific ex-pression of prosaposin mRNA in murine tissues. Am J Pathol. 1994;145:1390–1398.
Sylvester SR, Morales C, Oko R, Griswold MD. Sulfated glycoprotein-1(saposin precursor) in the reproductive tract of the male rat. Biol Re-prod. 1989;41:941–948.
Tomomasa H, Waguri S, Umeda T, Koiso K, Kominami E, Uchiyama Y.Lysosomal cysteine proteinases in rat epididymis. J Histochem Cy-tochem. 1994;42:417–425.
Trasler J, Saberi F, Somani IH, et al. Characterization of the testis andepididymis in mouse models of human Tay Sachs and Sandhoff dis-eases and partial determination of accumulated gangliosides. Endo-crinology. 1998;139:3280–3288.





























