Embed Size (px)
Citation preview

1
Replicating Minicircles: Overcoming the Limitations of Transient and of Stable Expression Systems
In "Minicircle and Plasmid DNA Vectors - The Future of non-viral and viral Gene-Transfer", Schleef (Ed.) Wiley-VCH Verlag
K. Nehlsen1), S. Broll1,2), R. Kandimalla3), N. Heinz4), M. Heine5), S. Binius1), A. Schambach4) and J. Bode4*)
1) Helmholtz Center for Infection Research, Department Molecular Biotechnology, Inhoffenstraße 7, D-38124 Braunschweig
2) Leibniz Universität Hannover, Dezernat 4 - Forschung und Technologietransfer / Nationale Forschungsförderung.
3) Department of Pathology, Josephine Nefkens Institute Erasmus MC 3000 CA, Dr. Molewaterplein 50 Rotterdam, The Netherlands Germany
4) Hannover Medical School (MHH), Carl-Neuberg-Strasse 1, D-30625 Hannover, Institute for Experimental Haematology OE 6960, Room J11 01 6530; Tel.: +49 511-532-5136; Fax: +49 3212 106 7542; [email protected];
*) Corresponding Author
5) Rentschler Biotechnologie GmbH Erwin-Rentschler-Straße 21, 88471 Laupheim
Keywords: minicircles; nonviral episomes; ARS assay; oriP; S/MAR
Abbreviations used: BPV, bovine papillomavirus; BUR, DNA base-unpairing region; CHO, Chinese hamster ovary; CUE Core-Unpairing Element; CS, constitutive S/MAR; DS, dyad symmetry element; EBV, Epstein–Barr virus; eGFP, enhanced green fluorescent protein; egfp, the corresponding coding region; FACS, fluorescence-activated cell sorting; FISH, fluorescence in situ hybridization; Flp, flippase (site specific recombinase); FR, Family of Repeats (OriP); FRT, Flp-recognition target; GANC, ganciclovir; GOI, gene of interest; GOD, gene on duty; HDACi, histone deacetylase inhibitor; HMT, histone-methyltransferase; IR, initiator of replication; IRE, inverted repeat, LRT, long terminal repeat; LUC, luciferase; MC, minicircle; MP, miniplasmid; Ori, origin of replication; MRE, mirror-repeat; Ori, origin of replication; OriP, origin of plasmid replication; ORC, origin-recognition complex; PD, population doubling; pEpi, plasmid-episomal; pFAR, plasmid free of antibiotic resistance genes; PP, parental plasmid / educt for MC preparation; RMCE, (Flp-)recombinase-mediated cassette exchange; S/MAR, scaffold/matrix attachment region; SIDD, stress-induced duplex destabilization; SV40, simian virus 40; UE, DNA Unpairing Element; TIC, teratoma-initiating cell.

2
ABSTRACT A - Gene therapy: Call for new vector vehicles
• Nonviral vectors avoiding genomic disturbances • Independent expression units: chromatin domains
o S/MARs: a unifying principle o S/MAR actions are multifold and context-dependent o Stress-induced duplex destabilization (SIDD), a unifying property of S/MARs o Chromosome-based expression strategies: Episomes and/or predetermined
integration sites (RMCE)
B - Replicating nonviral episomes • Can the yeast-ARS principle be verified for mammalian cells? • ARS and S/MARs: common (SIDD-) properties • S/MAR plasmids: verification of the concept
o Transcription into the S/MAR: directionality and rate o Cell and nuclear permeation
� Transduction principles o Nuclear association sites o RMCE-based elaboration following establishment
• Remaining shortcomings and their solution o Establishment and maintenance: the EBV paradigm
� Complementarity of “molecular glue” and initiator of replication (IR-) functions
� Two variants of the L1 transposon system � Can replication-support elements be shuffled between the EBNA1- and
S/MAR vectors? � Selection principles overcoming the need of antibiotics � Targets for DNA methylation: role of CpGs � pEPIto
o Vector-size limitations (?) C - Minimalization approaches
• Oligomerizing S/MAR modules: pMARS and its properties • Replicating minicircles, a solution with great promise
o Establishment and maintenance parameters o Clonal behavior o Bi-MC systems o MC-size reduction: “In vivo evolution” o Transcriptional termination and polyadenylation: an intricate interplay o Episomal status: Proof and persistence
• Emerging extensions and refinements o Combination of excision- and RMCE-strategies o MC withdrawal at will o Pronuclear injection and somatic cell nuclear transfer o From cells to organs
SUMMARY AND OUTLOOK

3
ABSTRACT
Based on a 2 kb S/MAR- (Scaffold/Matrix Attachment Region) element, the first nonviral
autonomously replicating nonviral episome could be introduced in 1999. S/MAR-binding proteins
such as SAF-A/hnRNP-U were shown to act as „molecular glue” to provide maintenance functions.
These actions enabled the association with replication factories of the host cell and thereby a
once-per-cell-cycle replication of the supercoiled DNA circles. In case of the plasmid episome the
requirement of a selection agent for its establishment, its continued silencing, and a limited cloning
capacity remained the limiting parameters until 2006, when these restrictions could be overcome
by deleting the prokaryotic vector backbone. The remaining ~4 kb ´minicircle´ (“MC”, later reduced
to a ~3 kb derivative, “M18”), consists of only one active transcription unit in addition to the S/MAR
and is devoid of prokaryotic CpGs. In contrast to the “parental plasmid” precursors (PPs) it can be
established in the absence of drug selection, and it replicates stably without signs of integration.
Other than conventional minicircles that are maintained only in non-dividing tissues, this is the first
example suitable for the modification of dividing cells due to its authentic segregation. Supported
by its minimized size, and in accord with the “pFAR”-principle, the vector is no target for epigenetic
defense mechanisms; after its establishment it is efficiently retained in the host cell nucleus. Stable
clones can be derived, stored for subsequent purposes and used to generate cell lines with
predictable characteristics. In addition, several MCs can be established side-by side allowing the
regulated expression of multi-subunit proteins. While the minicircle preparation process could
continuously be refined in various cooperations, MC generation has also become possible in situ,
i.e. in the recipient cell itself. At present this "all-in-one” concept mainly serves exploratory
purposes to pre-select suitable candidates for MC production routines leading to MCs of
unprecedented purity and and with an authentic superhelical (ccc-)status.

4
A – GENE THERAPY: CALL FOR NEW VECTOR VEHICLES
General problems that have hampered gene therapy approaches concern the inability of targeting
vectors to appropriate genomic sites. Such an option would guarantee adequate gene expression,
and tolerance by the host.
In the absence of certain drawbacks viruses might be the preferred systems. Although they
have the natural inclination to invade human cells and to deposit their genome in highly expressed
loci their cloning capacity is usually restricted while their preparation is demanding and evaluation
is laborious. For retroviruses (except the genus Lentiviridae) gene transfer is restricted to dividing
cells and expression is difficult to maintain over extended times. To circumvent unanticipated
complications of this kind chromosomal organization principles gain increasing attention for an
appropriate design of second generation nonviral “chromosome-based vectors” [1].
• Nonviral vectors avoiding genomic disturbances
In this field the limited performance and shutdown of conventional transgene expression units are
important limitations that have to be overcome for many potential gene therapy applications [2,3].
Until recently, virtually all stable transfection procedures involved the transfer of linearized DNA.
The integration of these specimens depends on the eventual occurrence of a genomic break in
processes that are often associated with unpredictable rearrangements due to cell-intrinsic
nonhomologous end-joining (NHEJ-) related repair activities. Silencing phenomena have been
attributed to host defense mechanisms directed against the bacterial backbone of traditional
vectors that include elements such as unmethylated CpG motifs [2], a prokaryotic origin of
replication and antibiotic resistance genes [3]. While these sequences are required for the
production of plasmid DNA (pDNA), each raises serious biological safety problems due to the
dissemination of antibiotic resistance genes via horizontal gene transfer and a residual activity of
bacterial genes in the recipient [4]. This becomes particularly obvious in animal models for which
intramuscular injections of pDNA raise immune responses. The corresponding findings led
regulatory agencies to restrict the co-transfer of these components, especially antibiotic resistance

5
markers. These facts have motivated developments considering the organization of vector
backbones into host-like chromatin structures [5-10].
• Independent expression units: chromatin domains
Eukaryotic chromosomes are organized into a series of discrete higher order chromatin domains,
each of which is delimited by two boundary elements, so called scaffold/matrix attachment regions
(S/MARs; Fig. 1). These S/MARs associate with ubiquitous protein components of the nuclear
skeleton (listed in Fig. 1B), most prominently complexes of scaffold attachment factor A (SAF-A),
which form the base of a chromatin loop creating independent units of gene activity [10].
S/MARs, a unifying principle Naked transgenes are known to preferentially integrate into
heterochromatic areas [11]. However, if transfected as a domain, (S/MAR1 – GOI – S/MAR2) , the
resulting clones show elevated, comparable expression levels that are maintained for extended
periods of time [12]. This effect has been called “transcriptional augmentation” [5] as it is different
from enhancement by the following criteria:
- traditionally, S/MAR actions have only been observed after integration, whereas an
enhancer is active in transient and stable expression systems;
- the presence and activity of S/MARs in episomes suggests their dependence on an
authentic chromatin structure, which can only be attained during replication. Since the same
principles should exist for nonviral episomes it appears that the pathway leading to an ordered
chromatin organization (replication as part of the genome of the host cell or as an independent
unit) is of secondary importance;
S/MARs per se do not enhance transcriptional levels but rather prevent silencing. This is supported
by our observation that highly expressed genomic sites are no further improved by the presence of
these elements [13]. Stringent selection procedures have even led us to conclude that highly
expressed loci are governed by pre-existent genomic S/MARs [14]

6
Under these circumstances S/MARs clearly reveal “insulator functions” the effect (but not
the molecular basis) of which is comparable to the classical insulator cHS4 (a prototype insulator
from the chicken beta-like globin gene cluster) at some but not at all genomic sites [13]. If
subjected to the classical tests underlying the definition of S/MARs, cHS4 is clearly different, which
can be explained by the fact that it associates with a particular protein, the CCCTC-binding factor
CTCF that forming bridges to the nucleolar surface, which is mediated by nucleophosmin ]13].
Whereas S/MARs shield a gene from silencing, their insulator functions do not necessarily
share enhancer blocking activity with cHS4. Although extended boundaries consisting of
“constitutive S/MARs” clearly prevent interactions across domain borders, this is not the case for
“facultative S/MARs” that are much shorter and depend on the simultaneous presence of an
additional associating factor such as YY1 (otherwise called NMP1 [6], i.e. nuclear matrix protein 1
or SATB1; [7,8,15]). At promoter-upstream positions or as part of an early intron they may even be
required for enhancer actions, for instance by introducing loops that enable the apposition of a
promoter with its coordinated enhancer. Prominent examples are again the huIFN-ß gene [7,8] or
the mouse immunoglobulin κ- and µ- chain genes [16,17]. By necessity, intronic S/MARs have to
be transcribed. Since they do not impede passage of Pol II, their occupation must be regulated.
In yet another scenario transcribed S/MARs occur in intergenic regions. An element of this
type coincides with the replication origin of the chicken alpha-globin domain, which, in normal and
transformed erythroblasts, becomes part of a full-domain transcript [18]. After the transcription
process has led to opening of the domain in dedicated cells, the element re-attaches to the matrix
separating the individual transcription units. Finally, extended S/MARs coinciding with the domain
borders usually define the termini of a replicon [19], whereas the function of short S/MARs with a
role in S phase is modulated by transcription.
The rules underlying such an event could be studied on retroviral integrates, which have the
particular advantage that, at low MOIs (multiplicities of infection), they cleanly integrate as single
copies. Therefore this provirus model enables the study of single-copy inserts with defined ends
(LTRs) at otherwise unperturbed genomic integration sites. Except from the basal vector carrying a
4.3 kb transcription unit, derivatives were transduced, each containing an 800 bp huIFN-ß sub-

7
S/MAR insert (element “IV” in Fig. 1A) at a different position. Whereas the S/MAR-IV insert
impeded transcription at distances below 2.5 kb downstream from the promoter, it strongly
supported transcriptional initiation in case the distance exceeded 4.5 kb, i.e. at localizations within
the LTRs or ahead from the 3´-LTR [20]. These findings could be accommodated in the classical
twin supercoiled domain model of transcription, which comprises a over-wound domain in front of
and an under-wound (negatively supercoiled) one behind RNA polymerase [5].
S/MAR-actions are multifold and context-dependent Our findings that an S/MAR fragment
supports transcriptional initiation when placed at a certain distance downstream from a promoter
has since been exploited for a variety retroviruses and cell types (compiled in Tab. 1). The
transcription of these proviruses is known to become down-regulated by negative regulatory
factors associating with silencer elements within the LTRs or the tRNA primer binding site.
Initial experiments relied on Mo-MuLV vectors for which silencing of a reporter gene is
accompanied by 3´LTR methylation. In a pilot study ([21]; Tab. 1) the 800bp S/MAR-IV was placed,
in both orientations, either into the LTR (generating a proviral double-S/MAR status resembling a
chromatin domain) or next to the 3´LTR upstream end. While the experiments revealed an
unanticipated orientation effect (activity in the “+”, but not the “-“ direction; [21,26,27]), the location
of the S/MAR at or within the 3´LTR (plus the 5´LTR) was of minor relevance as anticipated by the
above pilot studies: in both cases the expression remained stable for more than four months, and
no LTR methylation was observed. This fact directly supports observations that S/MARs prevent
methylation in transcriptionally active loci [22]. Since the single-S/MAR setup with element IV next
to the 3´LTR enabled higher virus titers, all subsequent studies relied on this situation.
These experiments were extended to various other cell types and retroviruses with S/MAR-
IV alone or in direct contact to a double-copy core sequence from the prototype cHS4 insulator.
While some combination of the two elements seemed beneficial in one system [28] S/MAR-IV
alone seemed largely superior in another [29]. Studies on a non-S/MAR reference revealed further
mechanistic details: by assessing the acetylation status of histone H3 (i.e. a prototype euchromatin
marker, cf. Fig. 2) a significant provirus deacetylation occurred with time indicating silencing within

8
the stem cell. At early stages this effect could be reversed by a histone-deacetylase inhibitor
(HDACi), i.e. Trichostatin A (TSA). Contrary to HDACi actions, increased CpG methylation became
evident only at a later stage at which reactivation attempts using either TSA or the methyl-
transferase inhibitor 5´-azactidine (5´-azaC) remained inefficient. These observations confirm a
current model implying that, while silencing is initiated by histone-deacetylation, the silenced state
may become locked, by DNA methylation, only at later time points (Fig. 2).
An observation deserving further attention is the fact that, while S/MAR IV acts in an
orientation-dependent fashion in three reports, in the latter example [29] the same element is
effective regardless of its orientation. While the molecular basis for these particular differences
remains undetermined, they nevertheless confirm the context-dependent action of facultative
S/MARs. At a later point examples will illuminate the way S/MARs can modulate the superhelical
status of neighboring regulatory elements, depending both on their sequence and associated
structural features.
Stress-induced duplex destabilization (SIDD), a uni fying property of S/MARs S/MARs have
been operationally defined according to the protocols that led to their detection [13,30-32]. The
respective elements have been implicated in a variety of biological activities, all of which are
compatible with an affinity for the nuclear matrix. Besides insulator-, augmentation- and enhancer-
support functions these include the long-term maintenance of high transcription levels by
counteracting histone- and DNA methylation steps, the support of histone acetylation, and
accessory origin-of-replication functions.
In spite of this wide spectrum of activities, all S/MARs have one property in common: they
consist of a more or less regular succession of DNA-unpairing elements (UEs) which initiate
double strand separation under negative superhelical tension (Fig. 1A and [32]). These UEs
together constitute a base-unpairing region (BUR) with an architecture enabling the
accommodation of prototype nuclear matrix proteins [33].
UE properties were first analyzed for the standard pBr322 plasmid [34] for which the SIDD
profile reflected preferential opening of the intrinsic Ori. Subsequent analyses on pro- and

9
eukaryotic DNA were performed at a standard superhelical density of σ=−0.05 as first determined
for the bacterial plasmid. It is of note that site-specific nucleases have opened the possibility to
excise pieces of genomic DNA between integrase target sites, which have been strategically
positioned within a eukaryotic chromatin domain [35]. Since an integrase-mediated excision
process preserves the preexisting superhelicity within the resulting circle, σ- values for eukaryotic
genomic loci can be determined with precision. Results so far demonstrate a similar range for
active eukaryotic loci.
Our first studies on the structure/function relationships of S/MARs concerned the domain
organization of the human interferon ß (huIFN-ß) gene located at position 9p22 on the short arm of
chromosome 9 (Fig. 1) . Apparently, the 14 bp domain is flanked two ~5kb constitutive S/MARs
comprising the 2.2 kb EcoR1 fragment “E” (upstream border) and most of the ~ 4.5 kb Hind III
fragment that had been localized before by the classical scaffold-reassociation assays [36]. Other
marks are certain intense and widely-spaced individual peaks, which triggered in-depth
investigations by Klar et al. [7,8]. They showed an association of these sequences that could later
be associated with DNAseI hypersensitive sites with regulatory potential (the mentioned
“facultative S/MARs”).
SIDD analyses and functional tests on S/MARs from mammals and plants explain the
cross-species activity of these elements: without exception, active S/MARs are BURs with a
related architecture: all of these comprise a register of UEs that obey certain rules regarding the
minimum number, spacing and threshold destabilization. Together these features mediate the
association of a multifunctional protein called SAF-A, SP120 or hnRNP U [37,38]. Assembly as a
multimeric complex results from cooperative interactions with the S/MAR (“mass binding”, cf. Fig.
1A). These properties could be reproduced in an in vitro assay where SAF-A - S/MAR association
occurred in the presence of nonspecific competitor DNA and were assigned to a short stretch of
amino acids in the N-terminal region, designated SAF-box/SAP domain. SAF-A recognizes AT-rich
sequences (“AT patches”) that are common for S/MARs. Apart from this the co-purification of SAF-
A with proteins such as histone acetyltransferase (HAT p300/KAT3B, introduced in Fig. 2 as an
enzyme modulating histone H3-structure and function) indicates that SAF-A serves as a platform

10
for the assembly of factors modulating S/MAR functions. The presence of an RNA binding motif
(RGG box) in its C-terminal domain is in accord with its designation as a member of the hnRNP
family of proteins involved in the processing of pre-mRNAs .
Chromosome-based expression strategies: Episomes an d/or predetermined integration
sites (RMCE) Two of our central approaches addressing the design of chromosome-based vectors
are outlined in Fig. 3. Both concepts comply with a set of rules that have been covered extensively
in a recent review [1] :
- The incorporation of S/MARs which, due to their strand-separation potential support
transcription, provides accessory functions to origins of replication and enhances the efficiency
of recombinases [14] ;
- Flp-recombinase target sites (FRTs) are recombined in the presence of the “flippase” (Flp),
provided that they are identical and thereby able to cross-interact [1,39].
Site-specific recombinases (SSRs) have opened new options for the systematic modification of
eukaryotic genomes. In case two identical, equally oriented 48 bp target-sites are parts of a given
DNA segment, the intervening sequence will be quantitatively excised. If applied to the S/MAR-
plasmid (the so called “parental plasmid”, PP) in Fig. 3B the procedure generates two daughter
molecules, a miniplasmid (MP) accommodating prokaryotic vector parts and accessory sequences,
and a minicircle (MC), which exclusively consists of the desired functional eukaryotic sequences.
According to the above definitions the MC represents a minimal model for a functional chromatin
domain, though at an extrachromosomal location.
The formal reversion of this excision process would be the addition of MC and MP entities
(i.e. re-formation of the PP) but also of oligomeric derivatives containing products arising from MC
x MC or MP x MP recombination. Since these “reverse-type” reactions are bimolecular processes
that have to occur against kinetic and entropic barriers [40] they would have to be enforced by
extreme educt concentrations in the presence of Flp activity. This is the likely reason that
complications of this type have not been encountered.

11
B – REPLICATING NON-VIRAL EPISOMES
Nonviral gene delivery strategies are usually based on bacterial plasmid-DNA (pDNA) carrying the
gene of interest. Already in 2007 pDNA contributed to 26 % of all clinical trials. Due to its relative
safety, simplicity, and reliability, naked DNA received particular attention for transfer into muscle
tissue. Efforts to improve the efficiency of non-viral gene vehicles require a better understanding of
delivery kinetics for different types of DNA into clinically relevant cells. Three DNA species have
been compared: linearized plasmid DNA (l-DNA) formulated by single-site digestion of c-DNA,
reduced-size linear gene cassettes generated by PCR (pcr-DNA) and a covalently closed circular
(ccc-) vector with a certain superhelical status. The latter specimen deserves particular attention as
it surpasses linear DNA regarding transcriptional potential [41], resists integration in diploid cell
genomes [42] and facilitates the transfer across cellular membranes
Another step forward concerns nonviral circular episomes that could be converted into
minicircles following the general scheme depicted in Fig. 2B (and detailed in Chapter C). To this
end site specific recombinases (the Tyr-dependent recombinases Cre and Flp or Ser-dependent
variants, such as ΦC31 integrase and ParA resolvase) could successfully be applied [4].
Resolvases are sometimes preferred since absence of accessory factors leads them to operate in
an irreversible fashion. Before addressing the advantages of nonviral, replicating S/MAR-
minicircles we will briefly describe the properties of S/MAR plasmids which enable the generation
of ARS-type vectors.
• Can the yeast-ARS principle be verified for mamm alian cells?
In yeast an origin of replication is specified by ~125 base pair DNA-segments called autonomously
replicating sequences (ARS). ARS elements are putative origins of replication, which cause
plasmids including an ARS to be maintained autonomously in the absence of integration or other
sequence rearrangements. A closer inspection revealed an 11-bp core sequence (ACS, ARS
consensus sequence), which is part of the recognition site for the origin recognition complex
5µm

12
(ORC). However, while central properties of the ORC are evolutionarily conserved, the replication
promoting sequences are not. Thus, the nature of replication origins in metazoan genomes has
remained largely elusive.
A more direct access to mammalian Oris was expected from screening chromosomal DNA
for sequences which might confer the ability of autonomous replication in homologous mammalian
cells. For mouse genomic DNA this approach led to several, apparently functional DNA segments,
which later turned out to have mere plasmid-DNA amplification capacity. A variable subpopulation
of episomes could subsequently be ascribed to concatemeric integrates which recombined yielding
extrachromosomal circles with limited persistence.
• ARS and S/MARs: common (SIDD-) properties
There are definite relationships between ARS elements and S/MARs. An early report goes back to
Amati and Gasser [43] who demonstrated specific sequences bound to the yeast nuclear scaffold
to provide ARS functions. Certain regions with scaffold association potential could be shown to
include the 11 bp ARS consensus, suggesting that scaffold binding is related to ARS activity. A
later report by Ak and Benham [44] confirms that highly conserved properties of yeast origins
concern S/MAR-like characteristics, in particular. a definite susceptibility to superhelically driven
DNA duplex destabilization. It is suggested that these features, in conjunction with other
characteristics, might be exploited for the localization of Oris in the yeast genome. These ideas
gained support by Li et al. [45] investigating the ARS properties of S/MARs from tobacco in the
yeast system. In fact, two out of six elements complied with the relevant criteria. This confirms
relationships between scaffold attachment and replication potential also for higher eukaryotes.
Other replication minimal models go back to viruses, such as SV40, BPV or EBV that
replicate episomally in mammalian cells. Also there replication initiation is supported by an easily
melting DNA tract, i.e. a base-unpairing region (BUR). Conformational coupling permits the energy
absorbed by base-unpairing to be delivered to a DNA unwinding element (DUE) where it serves to
establish secondary structures such as hairpins or stem-loops. Once more these are prerequisites
for an ORC initiating replication at the origin recognition element (ORE; [46]).

13
For more than a decade vectors sharing functions with natural chromosomes were thought
to solve problems related to safety and reproducibility. These vehicles do not require viral factors
for their function, and should be stably maintained in the cell for many generations in the absence
of continued selection. In case of linear minichromosomes three functional elements are required:
telomeres, centromeres and an Ori. While functional telomeres and centromeres could be
provided, the megabase-size of these entities per se granted the occurrence of Ori-characteristics,
although these features had to remain largely undefined owing to Ori-extension. Most of these
approaches suffered from the long-term instability of artificial chromosomes (ACs), however, which
motivated the systematic exploitation of ARS-principles for mammalian cells.
• S/MAR plasmids: verification of the concept
The established strand-separation potential of S/MARs lends support to the idea that there is a
regular association of these elements with origins of replication as exemplified by the dihydrofolate
reductase domain [47]. This assumption led to the generation of an S/MAR plasmid with replication
potential in a variety of eukaryotic cell systems [48]. Available evidence indicates that it is the 2 kb
fragment of the huIFN-β 5´ S/MAR (Fig. 1A) that recruits components of the cellular replication
apparatus to support authentic replication and segregation [49]. After its establishment in the
nuclear architecture depending on an initial phase under selection pressure, the replication
apparatus of the host cell is utilized in a way that S/MAR episomes replicate once during the early
S phase of the cell cycle in synchrony with the cellular genome. Quite unexpected at first, this
vector does not require specific DNA sequences to accommodate the origin recognition complex in
vivo. This indicates that the site on the episome where replication initiates is determined by
epigenetic principles [50].
Transcription into the S/MAR: directionality and ra te A stringent prerequisite for an S/MAR
taking over Ori functions is its combination with an active transcription unit to enforce strand
separation. Fig. 3B indicates that transcription has to run into the S/MAR causing its over-winding
within the positive superhelical part of the classical twin-domain model. In this situation histones

14
will be driven off by the tracking protein but will re-associate and reform nucleosomes within the
under-wound (negatively supercoiled) domain [51]. At what time point the underwound and over-
wound parts of the plasmid will compensate each other is hard to decide due to the dynamic
(binding-)properties of the interposed S/MAR region.
The Fig. 4 experiment [52] demonstrates that the direction of transcription is the
prerequisite for efficient episomal persistence, at least for the prototype vector pEpi. Here we made
use of our toolbox, i.e. Flp- or Cre- recombinase in conjunction with two oppositely oriented
recognition sites at the flanking the transcription cassette to invert this unit in a remarkably slow [1]
recombination reaction. After terminating the process 10 out of 15 clones were found with an
inversely oriented unit (“i”), i.e. a transcriptional direction that poses the S/MAR in the negative
superhelical domain. Four constructs maintained the original orientation (“o”) and one harbored
both orientations (“i/o”). It is of note that within this collection the episomal state was apparent only
for the original S/MAR plasmids, while constructs with an inversely-oriented transcription unit
yielded Southern blots with a considerable background in the absence of a clear-cut signal.
Concerns that mechanistic particularities of the recombinase-mediated inversion process might
have triggered integration were invalidated by the observation that corresponding results were
obtained in case plasmids with either the “o”- or the “i”- orientation were applied in separate
experiments [53].
So far all studies agree in that the “o”-orientation is more efficient. This simplistic statement
is refined by a recent report on a sophisticated pEpi derivative that yielded fewer, but obviously
episomal copies in the “i”-orientation [54].
Cell and nuclear permeation Only part of naked DNA that gets in contact with the outer cellular
membrane can actually enter the cell. In order to improve gene transfer efficiency and to obtain
adequate expression, transfection agents and electroporation/nucleofection procedures are now in
common use. The apparently higher efficiency of the second class of methods is, at least in part,
due to introduction of strand breaks into both the circular vector and the genomic DNA, which
trigger non-homologous end-joining (NHJE) and thereby integration [55,56]. For obvious reasons

15
these approaches should be avoided when it comes to vectors for which performance depends on
an authentic ccc- status (see the PP and MC species in Fig. 3B). This may turn out to be different
for femto-second laser pulse transfer techniques which are under intense present development
[57]. The following section still has to rely on nonviral carriers such as cationic lipids and polymers
that interact with the anionic DNA via charged moieties, thereby forming compact, nano-sized
particles suitable for cellular uptake.
- Transduction principles Hsu and Uludağ [58] have applied four gene carriers,
polyethyleneimine (PEI), poly-L-Lysine (PLL), palmitic acid-grafted PLL (PLL-PA), and
Lipofectamine-2000 to test the delivery and expression for each of three DNAs, a ccc-plasmid, a
linearized version thereof (l-DNA) and a shorter l-DNA variant, obtained by PCR amplification of its
center section (pcr-DNA).
ccc-DNA exhibits a higher intracellular diffusion capacity facilitating nuclear targeting and/or
expression compared to its linearized forms. On balance, pcr-DNA bears only the promoter-GOI
unit in the absence of prokaryotic vector sequences and may have an improved potential to
traverse the nuclear membrane. Unfortunately, all forms of l-DNA are prone to intracellular
nuclease attacks unless they are capped, i.e. provided with hairpin structures at both ends to
comply with the Minimalistic Immunogenically Defined Gene Expression (“MIDGE”-) principle [3].
The results show no obvious difference in the morphology of particles regarding interaction,
binding kinetics, dissociation characteristics and DNA uptake depending on either DNA structure or
transfection reagent. Using PEI, however, the best expression was observed for ccc-DNA followed
by l-DNA and pcrDNA. Although the latter specimen was delivered to the same extent,
exonucleolytic actions may have invaded its functional core invalidating the (otherwise desired)
absence of prokaryotic vector parts.
These results are in accord with a superior expression of ccc-DNA. Recent indications from
yet another system let it seem likely that this status also facilitates passage of the nuclear
membrane: While the technically demanding injection of linear expression constructs into the pro-
nuclei of fertilized mammalian eggs is the traditional method for generating transgenic embryos, an
effective nuclear transfer can also be achieved upon cytoplasmic injection of ccc-specimens. This

16
originally unexpected phenomenon indicates that the circular superhelical status enables a specific
nuclear transfer route of yet undetermined nature [59]. Still another factor has been associated with
the kind of promoter(s) on the vector. Exemplified by the SV40 unit, association of ubiquitous
transcription factors and the subsequent exposure of their NLS signals were found to facilitate
passage of the nuclear membrane [60]. The specificity of this effect could best demonstrated by
controls (such as the CMV promoter) that are devoid of such an activity.
Nuclear association sites Nonviral episomes are able to recruit the replication apparatus of the
host cell. Contrary to their viral counterparts, they do not need external accessory factors. Again, it
is the S/MAR providing the link to the nuclear matrix. In this position it does not only enable use of
the endogenous transcription factories, but it also counteracts silencing.
Transcriptionally active genes replicate early in S phase, possibly supported by certain
transcription factors [61]. Chromatin-DNA interactions obey a “histone code”, i.e. particular patterns
of covalent histone tail modifications, which, together with DNA methylation patterns, is part of the
epigenetic code. While it is accepted that histone tails are modified by processes like methylation,
acetylation, ADP-ribosylation, ubiquitination, sumoylation and phosphorylation, functional details
have only been worked out in specific cases (Fig. 2). Of primary diagnostic value are acetylation
and methylation processes concerning the core histones, H3 and H4. A number of diagnostic
immunoprecipitation kits have become available to this end.
Lysine N-ε-acetylation is a dynamic, reversible and tightly regulated modification with a
major role in chromatin remodeling and in the regulation of gene expression, especially at the level
of transcription. For H3 the process occurs at several different lysine positions in the N-terminal
domain where it is performed by histone acetyltransferases (HATs/KATs) such as
CBP/p300/KAT3B (Fig. 2).
For the non-viral episome pEpi histone H3 acetylation was found to be enriched on the
expression unit while the same gene residing on an integrated control underwent histone H3K9 tri-
methylation (K9me3), and thereby silencing [62]. This study showed S/MAR episomes to
preferentially interact with early replication sites that are spread throughout the nucleoplasm.

17
Immobilization of an episome at these sites is the likely consequence of S/MAR-mediated binding
to nucleoskeletal structures [63]. Later work extended these findings by a systematic exploitation of
alterations in the H3 methylation status at lysines 4 (K4me, K4me3) and -36 (K36me3) for both the
pEpi vector and its S/MAR-free, integrating precursor construct, pGFP-C1:
- whereas pGFP-C1 is mostly decorated with K9me3 as mentioned, pEpi-eGFP is
preferentially associated with modifications typical of active chromatin. For K4 the
modifications are enriched on the S/MAR, but K36me3 was uniformly distributed over the
entire vector;
- for pEpi the pattern remained stable throughout the G1-, and G2-phases in accord with a
persistent association with early replicating (perichromatic) domains accommodating replication
and transcription machineries and RNA processing factors;
- activating histone modifications are removed during mitosis, at a time when the association
with the host chromosomes is initiated.
To enable tracing the episome and its chromatin status in vivo Tessadori et al. [64] have prepared
a pEpi-derivative, “pELO64”, with a tandem array of 64 LacO sites. The lacO/lacR technology of
Belmont [65], i.e. the transient expression of a mCherry-lacR fusion served the visualization of
constructs in the living cell. After establishment (3 weeks of continuous culture in selection
medium) immobility of episomes could be confirmed and shown to last tens of minutes. The
absence of a “corralled” local low amplitude movement, which is otherwise typical for clustered
vectors, suggests that episomes become individually and firmly bound to host chromatin.
Despite their immobility, episomes re-locate to positions closer to the nuclear center if their
gene expression is stimulated by the addition of a histone-deacetylase inhibitor (TSA) and the
inhibition of DNA de-methylation (5-aza-dC) or, even more convincingly, by targeting a VP16
domain to pELO64. The latter treatment showed that transcriptional activation mediates a
relocation of the signals towards the nuclear center (64). Together, these results prove that the
regulatory mechanisms for episomal genes comply with those of the host genes.

18
RMCE- based elaboration following establishment Establishment of nonviral episomes and
their viral equivalents in the nuclear architecture usually occurs at low copy numbers (4-8 [46]) and
rates (usually <5%). After establishment the episomes are stably maintained for hundreds of
generations. As mentioned, major efforts have been invested to overcome these limitations, among
which the excision of prokaryotic vector parts, i.e. the generation of minicircles (MCs) appears as
the most promising one [66]. Since a time-efficient MC preparation proved to be a major
bottleneck, certain exploratory studies still rely on SMAR-plasmid precursors, such as the pilot
study in Fig. 5. This experiment explores the potential RMCE- (recombinase-mediated cassette
exchange) for replacing, within an established nonviral episome, a marker gene expression
cassette (here: luciferase) for a cassette bearing a gene of interest (exemplified by GFP).
RMCE (Fig. 3A) is a relevant extension of the Flp/FRT-methodology [67], which is
thoroughly addressed in a parallel review [1]. RMCE is based on sets of heterospecific, 48 bp FRT
sites differing in the sequence of the 8 bp spacer, which separates two inverted 13 bp Flp-binding
elements. The term “heterospecific” implies negligible or no cross-interactions between two spacer
variants (FRTmut x FRTwt) but maximal recombination between identical sites (FRTmut x FRTmut =
FRTwt x FRTwt). Under these conditions, a cassette FRTmut – GOI1 - FRTwt remains perfectly stable
even in the continued presence of Flp activity [1]. Only in case a second cassette of the same
architecture is provided as part of a donor plasmid and at a molecular excess, this donor cassette
(FRTmut – GOI2 - FRTwt) can be “flipped in”, cleanly replacing the resident one, which is either
genomically anchored (Fig. 3) or part of an established episome (Fig.5)
The feasibility of the latter concept has been shown by exchanging a luciferase gene (here part
of an established S/MAR-plasmid) for an eGFP gene that enters the scene as part of a (non-
replicating) donor plasmid. In the present experiment gain of eGFP fluorescence for 7% of the cells
in the absence of selection reveals a rather high RMCE-rate when compared with integrated
acceptor sites [13,67]. This renders the concept suitable even for successive RMCE-steps. While
these data already indicate a remarkable accessibility of the episomal target, a more uniform and
rather stable population can be obtained after FAC-sorting the fluorescent cells. Of particular note
is the fact that, whereas copy numbers vary by just a factor of two among the clones, expression

19
levels may differ by two orders of magnitude [46,52]. In the case of S/MAR plasmids this may be
ascribed, in part, to silencing with time. In case of minicircles, later experiments will demonstrate,
however, that a similar diversity is governed by clonal properties [52]: while all acceptor sites are
early replicating and rather highly expressed, their occupancy is stable in agreement with their
characterization as described under “Nuclear association sites”.
• Remaining shortcomings and their solution
While the topic “limited establishment of nonviral episomes” has been introduced above, this
parameter has to be judged by comparison with viral paradigms which have the reputation of being
highly efficient.
Establishment and maintenance: the EBV paradigm So far the greatest progress toward the
development of an efficient episomal gene therapy vector goes back to plasmids based on
Epstein-Barr virus (EBV), a member of the gamma subfamily of herpesviruses [68]. Molecular
details of EBV replication are well understood: the EBV latent origin has been identified and named
OriP (origin of plasmid replication). To initiate replication, OriP requires the presence of a trans-
acting factor, EBV nuclear antigen-1 (EBNA-1). OriP extends over 1.7-kb and comprises two
functional elements, the family of repeats (FR) and the dyad symmetry (DS) element. The latter is
a 120-bp region containing two EBNA-1-binding sites separated by 9-bp. The FR element in turn
comprises twenty EBNA-1 binding sites. It has been demonstrated that it enables EBV genome
retention as an episome. EBNA1 complexes on both, DS and FR, interact by a DNA looping
mechanism in a way reminding of nuclear matrix proteins like SAF-A, which has been implicated
in the replication of S/MAR-based episomes (Fig. 1B).
The FR element increases transcription rates depending on the number of EBNA-1-binding
sites. Detailed investigations on Raji cells showed that the reduction of repeat-numbers on an
EBV-derived plasmid decreases the formation of antibiotic-resistant colonies by three orders of
magnitude as the plasmid is lost. For living cells it could be shown that EBNA-1 mediates the
segregation of OriP plasmids and a high mitotic stability by their attachment to metaphase

20
chromosomes. In summary, EBNA-1 mediates replication by binding to two elements, an initiator of
replication (the “DS-“, otherwise called “IR”-element) and to FR, the latter enforcing its retention.
- Complementarity of “molecular glue” and initiato r of replication (IR-) functions “Raji ori”, a
second Ori in the EBV genome permits licensed DNA synthesis over restricted time intervals but
fails in long-term ARS assays [68,69]. While, for oriP, 90–99% of newly introduced plasmids do not
support initial DNA synthesis, for Raji ori, the range is 99.99–99.999%. This situation can be
alleviated, however, if a floxed (i.e. reversibly mounted) DS element is provided in cis. Under these
circumstances, long-term extrachromosomal replication becomes possible and it remains stable,
even in case the DS unit is removed after establishment. These observations confirm that episomal
retention depends on a combination of “molecular glue” (FR-type) and “initiator of replication (DS-
/IR-type) functions and that the initial replication rates (enabled by DS) are of primary relevance.
Again, this observation serves as a guideline for further improvements of the nonviral counterpart.
- Two variants of an L1 transposon system A revealing side-by-side comparison of EBV-type
and S/MAR based episomes was recently reported by Rangasami [70]. Both constructs were
applied to generate L1 transcripts that are spliced and reversely transcribed to generate a
transposon that becomes genomically integrated (retro-transposon principle).
Unexpectedly, chromosomal integrations of the EBNA1-based primary vector were quite
common, explaining a significant loss of copies/expression over 50 days, possibly due to the
repetitive nature of L1 sequences. Using the S/MAR-based counterpart, the L1 cassette proved
increased, prolonged retrotransposition efficiency due to higher expression levels under otherwise
identical conditions. Virtually all cells maintained episomal vector copy numbers for at least 50
days illustrating the stability of the respective L1 transposition construct as part of the nonviral
system for cultured human cell lines.
These differences did not follow the initial transgene copy number per cell, which was about
two for the S/MAR-L1 construct but exceeded 40 in case of EBNA1-L1.
- Can replication-support elements be shuffled bet ween the EBNA1- and S/MAR-vectors?
Considering the beneficial actions of S/MARs in an episomal context with regard to vector
maintenance and transgene expression in mammalian cells, Giannakopoulos et al. [71] introduced

21
the standard 2kb element into an EBNA1 episome in order to improve its maintenance, which
otherwise diminished over time. Unexpectedly, this step completely eliminated the capacity of the
resulting Pcmv-OriP-EBNA1 vector to replicate as an episome. The comparative calculation of SIDD
profiles for a series of constructs suggested that the S/MAR and the DS element are in mutual
competition with DS being on the looser side. As a result, its strongly reduced duplex
destabilization will have affected the vector’s replicative potential. This situation changes if an
alternative replication of Initiation Region (the IR-element from the β-globin gene locus) takes the
position of the EBV-Ori: an Pcmv-eGFP-S/MAR-IR insert not only restores the replicative capacity
but it also enables an enhanced episome retention. These data underline a synergism between the
S/MAR- and the IR-elements with regard to vector-retention and -replication potential. While the
conventional S/MAR vectors have Ori-support (otherwise called “molecular glue-”) functions,
initiation of replication occurs “all over the place” [50]. A strictly localized initiation, however, is
likely to be superior for effective vector propagation.
Recently, these experiences were used to improve episomal maintenance in human
hematopoietic progenitor cells. To enhance the vector’s potential the ß-globin IR element was
mounted as before, while transcription through the eGFP marker was driven by either the
EF1/HTLV or the SFFV promoter. SIDD analyses of the respective S/MAR plasmids anticipated
that these changes would preserve the function of both elements, S/MAR and IR. In fact, after a
single initial sorting step all vectors were quantitatively maintained as stable episomes in mobilized
CD34+ peripheral blood cells (A. Athanassiadou, submitted).
- Selection principles overcoming the need of anti biotics While there is little chance to recover
cells bearing plasmid-derived vectors in the absence of selection, several concepts are underway
to avoid the risk of antibiotic resistance marker dissemination, i.e.
- providing a growth advantage independent of drug selection markers [72];
- using FAC-sorting instead; this approach is necessarily restricted to cells that can be kept in
culture [73];

22
- applying novel approaches to produce plasmids Free of Antibiotic Resistance genes, called
pFARs. The strategy is based on the suppression of a chromosomal nonsense mutation by a
plasmid-borne function [3].
Previous experiments have shown that a prototype S/MAR-plasmid vector encoding the luciferase
reporter gene enabled transgene expression for at least 6 months following hydrodynamic delivery
to mice livers. After partial hepatectomy, however, no detectable vector replication was seen to
persist. To deal with this phenomenon, Wong et al. [72] have developed an in vivo selection
strategy providing liver cells with a survival advantage. Accordingly, the vector was modified to
express the Bcl-2 gene conferring resistance to apoptosis in the presence of a Fas-activating
antibody. In fact, this Bcl-2-luciferase S/MAR plasmid enabled episomal replication and sustained
luciferase expression for more than three month. Quantitative PCR was performed at the end of
this period to compare the copy number of plasmid molecules revealing a tenfold increase for the
S/MAR vector relative to a non-S/MAR control. This confirms, for the first time, the ability of S/MAR
plasmids to replicate and establish mitotic stability at a detectable level after application to an adult
organism, as long as there is a selective advantage.
From a Molecular Biologists point of view, transcription units and promoters of bacterial or
fungal origin are common initiators of inactivating methylation reactions in mammalians. In case
drug resistance genes are applied, their expression next to the GOI contributes to promoter-
interferences and silencing effects [14]. Along these lines, attention is paid to the work by Gossen
and colleagues [73] demonstrating that the method to recover stably transfected cells has a
profound impact on transgene expression patterns. Standard antibiotic selection was directly
compared to FACS methods regarding the establishment of stable cells and proved that only the
second approach could overcome phenomena associated with the spontaneous resistance to drug
selection markers to provide uniform and stabile gene expression patterns. This was therefore the
method of choice for a stringent side-by side comparison of PPs and their MC derivatives (see
below).
Although conventional plasmids encoding antibiotic (aminoside-)inactivating proteins have
been approved for certain clinical applications, pFAR constructs gain increasing attention

23
representing the approach to overcome risks associated with the dissemination of antibiotic
resistance markers (or contaminating antibiotics): whereas luciferase activities decreased within
three weeks after intradermal electrotransfer of conventional plasmids, sustained levels were noted
for a pFAR derivative. Thus, novel strategies have become available for the efficient production of
biosafe plasmids, which has already proven its potential in several organs and tissues.
In this context it should be noted that the chromosomal vector strategies introduced in Fig.
3 are variants of a pFAR design relying either on the clean exchange of a genomic target by an
eukaryotic expression cassette (RMCE) or on the excision of accessory sequences after plasmid
production in bacteria (minicircle concept)
- Targets for DNA methylation: role of CpGs It is an accepted fact that eukaryotes have evolved
elaborate defense systems to fight the expression of ectopic transcription units in order to protect
the integrity of their genomes. In mammals, the insertion of retroviral DNA, the incorporation of
repeat arrays and the co-introduction of prokaryotic vector parts are major triggers of
transcriptional silencing processes. Additional defense strategies go back to the fact that
dinucleotide frequencies in mammals differ from those of other organisms. Of particular relevance
is a relatively low content of CpG dinucleotides, which, in addition, are often methylated (mCpG). In
most bacterial genomes, however, the occurrence of CpGs is in accord with statistical
expectations, and cytosines remain normally unmethylated.
Although the CpG content of mammalian genomes is low, regions exist where these motifs
reach statistical levels. These “CpG islands” are associated with many genes and are protected
from methylation (and thereby from subsequent meC → U transition) by interaction with
transcription factors. While silencing is commonly accompanied by the methylation of CpGs, these
events may depend on a prior methylation of histone H3 at Lys-9 [74]. In the context of Fig. 2 it is
of note that, for ES cells, promoters positive for H3K27me3 are fourfold more likely to acquire DNA
methylation. Such a methylation center can trigger chromatin condensation spreading to a
downstream promoter to provide it with a heterochromatic structure – at least in the cases where
such a process is not blocked by an intervening insulator element [13]. Regarding human
organisms, another level of defense is associated with the innate immune system, which has

24
evolved mechanisms to discriminate bacterial from intrinsic DNA via Toll-like receptor 9 (TLR 9-)
signaling.
- pEPIto Returning to the class of replicating nonviral episomes, consideration of a common
heterochromatization route led to developing a size- and CpG-reduced derivative of the pEpi
S/MAR plasmid, pEPIto [2]. Traditional pEpi-type vectors comprise a pUC-Ori for bacterial
propagation, the S/MAR, a second mammalian SV40-O/P driven transcription unit serving
selection purposes in bacteria (kana) and mammalian cells (G418) in addition to 206 CpGs. In
contrast, in the pEPito backbone there remain 37 CpGs, an R6KOri for bacterial propagation, an
ampicillin selection gene and the common 2 kb S/MAR. A second transcription unit (eGFP-IRES-
BSD) encoding both the fluorescence marker and a drug selection (BSD-) function provides
additional options.
Side-by side controls proved both, increased transgene expression levels and colony-
forming efficiencies, for pEPito in vitro, in addition to a more persistent expression profile in vivo.
While, in the present setup, the establishment efficiency for pEpi-1-replicons was as low as 0.25%
in a colony-forming assay, it was six-fold higher (~ 1,8%) for the pEPito-construct, both controlled
by a CMV promoter. Although the effect of unmodified CpGs is an accepted contribution to
silencing and while it may have determined the outcome of this study, there is a recent report [75]
that CpG content is of minor or no relevance in the context of a minicircle, which mostly consists of
eukaryotic sequences. In particular, CpG islands are clearly exempt from negative actions, which is
particularly obvious in the case of an UCOE. Such a Ubiquitous Chromatin Opening Element
confers resistance to DNA methylation–mediated silencing in line with its origin from two
divergently transcribed promoters that are embedded in an extended methylation-free CpG island
[76].
In summary, to date all findings are compatible with heterochromatization steps initiating at
bacterial vector elements. Apart from CpGs tracts, there seem to be other distinguishing features
between bacterial and mammalian genomes which may trigger structure-based discrimination
steps [77]. Among these is the relative occurrence and secondary structure forming potential of
inverted and mirror repeats. These IREs or MREs can form cruciform- or intramolecular triplexes

25
(H-DNA) in negatively supercoiled domains. Since supercoiling triggers strand separation for B-
DNA, it is one of the prerequisites for these and related structural transitions. Whereas in E.coli
IREs are mostly restricted to transcriptional termination sites, MREs comply with statistical
expectations. Sequences with H-forming potential on the other hand are only typical for humans.
Together these considerations suggest the existence of a multi-facetted structural code that is
recognized in a foreign host to delimit the expression of foreign genes.
Vector-size limitations (?) Over the years evidence for an inverse relation between episome size
and -stability has accumulated. This became particularly evident during cloning, electroporation-
mediated vector transfer, FACsorting routines, persistence of the superhelical state during freeze-
thawing cycles and long-term stability in mammalian cells. For pEpi-type vectors the performance
was best if their size did not exceed 10 kb as it strongly deteriorates above ~15kb.
Regarding these observations the description of a 156 kb iBAC-S/MAR-vector (pEPHZ-LDLR) by
Lufino et al. [78] came as a surprise, the more as the authors demonstrated that such a construct
enables infectious delivery while retaining a 135 kb transcription unit. CHO cells were infected
exploiting the high transgene capacity of herpes simplex virus type 1 (HSV-1). Infected cells were
kept in selective media for two weeks after which 108 early stage single clones could be isolated,
most of which did not survive. Ten growing clonal lines could be maintained and screened, by
plasmid rescue, for their episomal status, which succeeded in three cases. Two clones were kept
in the absence of continued selection and shown to preserve a low-copy number episomal status
for at least 100 cell generations. While these observations indicate that there is no stringent upper
vector size limit impairing episome function, they nevertheless confirm very low rates of
establishment in case a certain size limit is exceeded.
C – MINIMALIZATION APPROACHES

26
Following these tendencies, minimal sizes for nonviral extrachromosomal entities should be of
value. This not only concerns the class of pure minicircles that are derived from so called “parental
plasmids” (PPs) by excision of redundant auxiliary sequences, but also their replicating S/MAR
variants (Fig. 3B). Based on their intrinsic molecular glue- and replication-support activities nonviral
episomes, and minicircles in particular, are in the position to utilize the host´s replication and
segregation machinery.
Another minimalization option was shown to reside in the S/MAR element itself. For
unknown reasons it is so far almost exclusively the 2kb sub-S/MAR sequence from the huIFN-β
domain (Fig. 1) that has been used to provide replication potential – sometimes at the expense of
long-term stability and an increased tendency to integrate. Since the rules determining S/MARs are
known (Figs 1, 6, 7), the stage was set for systematic minimalization efforts at this level. The
relevant principles can be summarized as follows:
- S/MARs occur only in eukaryotic genomes, where they serve a variety functions.
They represent DNA elements, between a few hundred to several thousand base-pairs in
length, which are operationally defined by their affinity for the nuclear scaffold or -matrix. In
the present context the term “scaffold” denotes the common protein network serving
primarily structural support functions, whereas the term “matrix” stands for the entire
complement of proteins resisting a given nuclear extraction routine. A potential overlap of
these functions is reflected by the consensus term “scaffold-matrix attachment region”
(S/MAR). Since S/MARs do not share obvious sequence motifs, an important component
determining their performance is thought to rely on structural particularities;
- a given S/MAR may have predominantly context-dependent (facultative) activities or
constitutive (domain bordering) functions. While the first group can even be shorter than
originally claimed, consisting of a single strongly-destabilized UE of or slightly above 170 bp
plus accessory transcription factor binding motifs [79], the second group comprises an
extended register of moderately-destabilized UEs that have to obey certain structural rules
[5]. This architecture mediates an association with the ubiquitous components of the

27
nuclear scaffold (exemplified by SAF-A), which may acquire secondary nuclear matrix
components.
Finally, several minimalization principles (deletion of prokaryotic vector parts and selection markers
or minimal arrays of UEs) can be combined anticipating that this will enable a second-generation
minicircle with superior establishment and stability, both regarding its physical status and
expression characteristics.
• Oligomerizing S/MAR modules: pMARS and its properti es
The approach by Jenke et al. [80] relies on an oligomerization strategy to investigate the S/MAR
potential of a 155-bp module, i.e. the most destabilized UE in the 2 kb standard element, and of
155n-oligomers. Initial scaffold-reassociation studies in vitro confirmed the continuous increase of
activities when oligomerizing the monomer and showed that the binding strength of the original 2
kb template was approached at the tetramer level (620 bp; Fig. 6C). In this and other examples the
term “activity” is not restricted to affinity parameters but it extends to biological functions such as
transcriptional “augmentation” (shielding a transgene insert from an heterochromatic environment),
the effect of histone hyperacetylation and, in the present case, the capability of episomal
replication in the context of a plasmid. An association of these minimized vector derivatives with
nuclear matrix components could be demonstrated by FISH analyses and by in vivo crosslinking
using cis-diammineplatinum(II)-dichloride (cis-DDP) [63]. FISH analyses per se proved a non-
covalent association with the mitotic chromosome for the S/MAR-tetramer, but not the dimer,
which, using the cisDDP protocol, could be ascribed to SAF-A.
Fig. 7 illustrates a rather dramatic effect of this minimalization during an early time interval.
Transfected cells were sorted out after 5 population doublings (5 days) and kept in continuous
culture for another two weeks before FACScans were recorded. Their comparison shows that after
this interval 31% of pEpi-expressing cells had persisted, while for pMARs 64% of the recipient cells
were still active. In both cases the loss of fluorescence may be due to inadequate establishment
and -maintenance, to silencing or a combination of both. These functions may have been improved
for the size-reduced pMARs derivative, but in case of pEPI (and certain derivatives) there is

28
evidence for yet another contribution: within the standard 2 kb S/MAR polyadenylation of the
growing mRNA occurs at a cryptic signal, whereas the 620 bp tetramer permits the transcription
apparatus to transverse the entire element before being processed at the authentic SV40
polyadenylation signal located at its 3´terminal end (Fig. 11 will deal with these phenomena in a
wider context). Together these findings show a complex interplay of various expression parameters
associated with the extension and stability of the transcript. Recall that this transcript has to cover
S/MAR sequences beyond the translational stop signal.
• Replicating minicircles, a solution with great prom ise
While there is significant progress in the modification, by episomal DNA, of slowly-dividing tissues
like liver, muscle and brain, maintenance problems have so far limited the use of nonviral
episomes for dividing cells, for instance of the hematopoietic system. For liver, the most advanced
vehicles appear to be “minicircles”, small circular vectors that are exclusively composed from
eukaryotic sequences. In contrast to linear DNA, minicircles do not form concatemers and are less
prone to integration. It is also known that, owing to their superhelical status, they are superior
transcriptional templates DNA [41]. Based on this rationale M. A. Kay and co-workers have
demonstrated that transgene expression levels in minicircles can be 45-560- [66,81], or even 10-
1000fold [82] higher and also more persistent than conventional plasmids. These vehicles were
therefore subjected to a critical test to prove their episomal state, i.e. a 2/3 hepatectomy upon
which almost every hepatocyte undergoes one or two cell doublings until the liver mass is
reconstituted. The results show that minicircles per se are not functionally attached to
chromosomal DNA and they anticipate the category of problems that have to be overcome in case
replicating S/MAR variants are used for the modification of proliferating cells [72]. Studies on
S/MAR-plasmids predict that, to be effective, minicircles will not only need replication potential but
also the capacity to become established in the nuclear architecture. To provide these properties
both molecular glue and initiator of replication functions are required, which come to life only in the
appropriate superhelical (ccc-) context.
By subjecting a pEpi-derivative to the machinery required for excising its plasmid parts
while maintaining the ccc-status, we could introduce the first minicircle that met a major part of

29
these requirements [83]. Since we have established an Flp-recombinase based toolbox to permit
the inversion, excision as well as (RMCE-mediated) integration of appropriately flanked expression
cassettes, we preferred the Flp/FRT system over alternatives that have been applied to the same
end.
Establishment and maintenance parameters Following these considerations a side-by-side test
of pEpi-type S/MAR-plasmids and their minicircle derivatives was performed [83]. To enable a
stringent comparison, we applied a single FACsorting step to obtain two populations of 100%
fluorescent cells (Fig. 8), the stability of which could then be followed for extended periods of time
(here: 50 population doublings, i.e. approximately 50 days). If the sorting was performed after an
initial 5 days period, establishment of the minicircle was almost complete, evidenced by the fact
that there was just a slight further decay, i.e. the level of expressing cells remained at about 70%.
Sorting at this point maximized the difference to the S/MAR plasmid, which, as a possible
consequence of continuous silencing, was completely lost during 30 PDs. Although the outcome of
such a comparison is clearly coined by the time point of sorting, major intrinsic differences between
both systems become obvious:
- At first view, the apparent deficiency of the plasmid derivative can be overcome by
drug selection starting as late as at 12 PDs, however:
- analyses on the recovered population, which persists due to the presence of a neor
gene, indicate that ~40% of cells have lost the episome by integration [52] .
Episome establishment and maintenance is a complex process based on epigenetic parameters
and stochastic events of largely unknown nature. A positive effect of histone deacetylase inhibitors
such as Trichostatin A (TSA) and butyrate was provisionally ascribed to arresting cells in G1 and
G2 just ahead nuclear membrane breakdown in early metaphase. This idea had to be abandoned,
however, because this effect did not arise using alternative synchronization approaches. Attention
was therefore paid to the role of open chromatin structures, which persisted after time-limited
histone hyperacetylation [46]. Meanwhile this concept is systematically pursued for improving the
establishment of minicircles. The insert of Fig. 8 shows a corresponding experiment.

30
Clonal behavior It has been noted before that the expression profiles of both, S/MAR-plasmids
and minicircles cover 2-3 orders of magnitude (Fig. 9, top), which has to be interpreted considering
the fact that this range goes back to just a two-fold variation of copy numbers (typically 4-8). For
S/MAR plasmids a partial explanation of this behavior is transcriptional suppression as it occurs at
the episomal state to be increased after its integration. Evidence for such a process comes from
the continuous shift of pEpi-expression profiles to low level positions, which can at least partially be
reversed by the action of a HDACi [83]. Minicircles, in contrast, usually show persistent expression,
which remains unaffected by HDACis. In this case the model suggests that variation simply reflects
the clonal behavior of cells after these entities have been firmly established [46].
Proof of this concept required the isolation of single clones and demonstration of their
persistence. Fig. 9 reports the outcome of these experiments, which gave raise to clones with a
sharp, symmetrical expression profile (M23 and L2 in Fig.9). Besides, there were some clones with
a double-maximum, which retained this property after sub-cloning (clone H11). Additional proof for
stability of M23 and L2 could be provided by an extended freezing - re-thawing cycle during which
both the expression profile and the episomal status were maintained.
Bi-MC systems These data encouraged the use of minicircle clones for expression purposes, the
more as these combine properties of transient and of stable expression systems. Proof-of-principle
comes from two-minicircle transfer experiments in which one entity encodes the light (L-) chain of
antibody and the other its heavy-(H-) chain counterpart. Usually, a certain overexpression of the
(secretable) L-chain proves beneficial because this entity provides chaperone functions when it
comes to correct folding of the H-chain (that would otherwise plug the endoplasmic reticulum
followed by cell death).
Significant conclusion on the system’s properties could first be drawn from the fact that bi-
MC clones can be established either by a synchronous transfer or by successive transfer steps, i.e.
transfection of the H-chain minicircle at a time where the L-construct has already been established.
Comparable efficiencies of both protocols indicate dynamic properties for nuclear substructures

31
exposing a number of sites that are suitable for establishment at any given time-point. This finding
invalidates an alternative explanation, i.e. saturation of binding sites already at low vector copy
numbers.
This model can be expanded by auto-regulatory principles. These emerged from the
observation that, regardless at which ratio H- and L-vectors have been transfected, both entities
are stably maintained at a certain ratio suggesting “survival of the fittest” (Fig. 10). Based on these
concepts technically more demanding setups can be overcome, which enforce certain expression
ratios by the choice of appropriate promoters, integration sites or the use of IRES elements to
create co-expression units with different properties.
MC-size reduction: “ In vivo evolution” The benefit of “survival of the fittest-”, otherwise called “in
vivo evolution-” principles emerged also at yet another level. In spite of their superior persistence
and expression properties minicircles showed instability in the long run, i.e. after >20 weeks of
continuous cultivation. We used the incidental observation of independent, but identical S/MAR-
internal deletion events within the 4.1 kb minicircle as they occurred during the long-term
cultivation of CHO-strains (cf. clone M18 in Fig. 11B). The size-reduced S/MAR was recovered by
PCR and used to construct an S/MAR-minimized parental plasmid analogue (cf. Fig. 3B).
Processing this PP by Flp-mediated excision led to a 2.9 kb minicircle with a largely reduced 733
bp S/MAR-insert. Relative to the 4.1 kb precursor this step again caused a dramatic improvement
of expression characteristics, both regarding its level and the stable persistence of the “M18”
minicircle [46]. The fact that the parental plasmid precursor of M18 outperformed pMARS regarding
its long-term stability led us to abandon the idea to generate minicircles from artificial S/MARs with
internal sequence repeats.
While vector stability per se may contribute to high level expression, the relevance of
authentic transcriptional termination/polyadenylation has already emerged before. Northern blots in
Fig. 11C demonstrate prematurely-terminated transcripts within the extended, 2kb S/MAR for both
pEpi and its 4.1 kb minicircle derivative, but an authentic usage of the SV40poly(A) signal for the
short-S/MAR versions pMARs and “M18” (Fig. 11C). Corresponding SIDD profiles in Fig. 11D

32
provide evidence that the deletion that gave raise to “M18” has inactivated (but not removed) the
cryptic internal polyadenylation signal and, at the same time, re-activated the genuine SV40
poly(A) sequence. Obviously, the same poly(A) signal is less destabilized if it is part of the
extended 2 kb S/MAR (see the respective UEs in the Fig. 11D SIDD profiles). This difference is
ascribed to a competition of the SV40 derived poly(A ) signal with the large number of UEs in the 2
kb S/MAR, which reduces its strand-separation- together with its secondary structure forming
potential. Benham [34,84] has shown that for higher eukaryotes poly(A) consensus sequences are
only used if they coincide with a region of significant strand separation potential, whereas in yeast
the requirements are restricted to the strand-separation requirements. Only the absence of an
extended, competing BUR will therefore permit the SV40poly(A) signal to adopt the secondary
structure enabling its recognition by the polyadenylation machinery [85].
Transcriptional termination and polyadenylation: an intricate interplay To date all nonviral
replicating episomes rely on an S/MAR element that is transcribed over at least part of its length. In
any case this process extends the primary transcript in a way that may affect mRNA stability and
gene expression. An evaluation and optimization of these facts is stringently required, unless the
desired GOI is accommodated in a separate transcription unit, cf. Fig. 10, where the L- or the H-
encoding “gene of interest” (GOI) performs functions apart from the “gene on duty” (GOD, here a
fluorescent marker). Meanwhile this configuration has proven its value by consistent results in
various experimental setups. The performance of different GOI-S/MAR combinations on the other
hand is hard to predict. An example is the first version of pMARS (Fig. 7) that had to undergo
rectifications before its fluorescence could be evaluated [46].
Polyadenylated mRNAs contain variable extensions between the coding region and the
poly(A) tail. The length of both the 3′ untranslated region and the poly(A) tail relates to the
translational efficiency and the stability of mRNA. Besides, polyadenylation is intimately linked to
transcription termination in a mutual, reciprocating way: co-transcriptional cleavage within the
downstream RNA Pol II transcription termination region depends on the presence of an upstream
AATAAA-type element. Termination, in turn, is the prerequisite for subsequent pre-mRNA 3′ end

33
processing/polyadenylation guided by the AATAAA tract. This interdependence has opened the
chance to preserve the essential S/MAR passage by the transcription complex while
polyadenylation can be directed to a location close to the end of the coding region. The concept
was verified for a β-globin transcription (HBB) unit for which the proximal termination region was
relocated to a position behind the S/MAR while preserving the polyadenylation signal. In fact, the
resulting construct enabled replication and retention of the episome as anticipated in the absence
of any unwanted mRNA extension [54,71].
Whereas a low-rate transcription through the S/MAR suffices for the establishment and
maintenance of plasmid episomes, the same study indicates that higher rates are associated with
elevated episome copy numbers opening yet another regulatory option.
Episomal status: Proof and persistence The criteria that are sometimes used to establish the
episomal status are subject to considerable contention [83]. Among these are
- plasmid-rescue, i.e. a re-transfer of circular episomes from mammalian cells to E.coli. This
procedure is not feasible for minicircles, which, according to the pFAR- concept do not
comprise the necessary bacterial DNA components. Although plasmid rescue can verify the
principal presence of plasmid derivatives, it does in no way prove an episomal status for all
transgenes in the recipient cell. Finally, circular plasmid entities may originate from multimeric
integrated concatemers in conjunction with intramolecular recombination events [86].
- Full-length PCR amplification, which can only serve as a preliminary indication for the presence
of episomes. Again, an identical effect may go back to multiple transgenes that have integrated
in a concatemeric head-to-tail fashion. This status is a typical concomitant of the classical Ca++-
phosphate transfection procedure.
- Linear amplification-mediated (LAM-) PCR [87], a technique originally developed to
characterize retroviral insertion sites. The sensitivity of the method results from pre-
amplification of vector-genome junctions and its efficiency is boosted by magnetic capture,
dsDNA synthesis, restriction, linker ligation and nested PCR steps. A subsequent study
addressed the episomal status of integration-deficient lentiviral (IDLV-) vectors based on the

34
fact that LAM-PCR would detect LTR- and non-LTR mediated integration. While such an event
could be traced [88], it is a rare exception. At present these reports are leading to an increasing
number of LAM-PCR applications in order to verify the performance of LTR-based episomes
(so-called “LTR-circles”; (ESGCT 2011).
- A clear-cut Southern-blot signal is a more stringent criterion as additional bordering
fragments would arise in case of integration. But there are typical and frequently neglected
shortcomings as demonstrated in Fig. 12. These examples clearly show that analyses on
clonal mixtures can be meaningless. On the other hand the episomal state becomes obvious
for single clones such as M23 and H11 (cf. also insert to Fig. 9).
- The traditional extraction procedure according to Hirt leads to the enrichment of non-
integrated DNA - at least at early passages. The efficiency of this protocol may decrease with
time since repeated rounds of replication can give raise to extrachromosomal chains
(concatenates) even in case of the viral systems [89].
- The conclusions from Southern blot- and Hirt- extraction procedures can be reinforced by
ATP-dependent nuclease treatment. More commonly known as “Plasmid-Safe”, this system
relies on a selective DNase that is mostly used for the removal of contaminating bacterial
chromosomal DNA from plasmid preparations [90]. The enzyme rapidly degrades linear DNA
under conditions that leave duplex circular DNA intact, justifying its classification as an
exonuclease. Paradoxically, however, it acts as an endonuclease on single-stranded DNA. The
latter activity is inhibited by the presence of linear duplex-DNA explaining the order of preferred
DNA substrates:
linear dsDNA > linear ssDNA > circular ssDNA >> nic ked circular DNA > circular dsDNA
Consequently, an adequate first step prior to Southern blots would be digestion of genomic
DNA by a MC-non-cutting restriction enzyme. Post-treatment of Hirt extracts with the nuclease
would indicate the presence of circular dsDNA and thereby enhance the stringency of the
assay. Apart from these approaches the ultimate criterion appears to be
- FISH-visualization of transgenes on metaphase spreads, which has proven its potential
before [49,52]. The approach generates multiple focused fluorescent spots in association with

35
the chromosomes when we have to deal with intact episomes. Such an association is lost if the
preparation involves shear forces [52]. Alternatively, we find a single intense doublet indicating
the typical co-integration of multiple copies subsequent to DNA transfer [49]. For the maxicircle
(and the respective parental plasmids) there are obvious examples where intense doublets,
one signal on each chromatid, indicate integration events that occurred during continued
cultivation and replication (Fig. 13).
In conclusion, while full-length PCR may serve as a preliminary hint, the Southern-blot and Hirt
extraction protocols may strengthen the evidence for the episomal status in case they are
combined with ATP-dependent nuclease treatment. Though more demanding, the most
comprehensive information on copy number and -status is enabled by metaphase-FISH.
A final comment addresses the apparent discrepancy between the results of Southern-blots
and metaphase-FISH data in Figs. 12 and 13, since the blots reveal a considerable contribution of
non-episomal minicircles in contrast to the fluorescence microscope data that are compatible with
an exclusively episomal status. Non-episomal specimens can arise from rigid treatments at stages,
prior to establishment. These treatments may involve FACS sorting – [91] as well as clone picking-
routines. Following these considerations it is recommended to allow an extended period of time for
establishment and/or to adapt the relevant parameters. Results in Fig. 9 strongly suggest that
clones that have been established after reaching their ultimate chromatin structure resist even
vigorous treatments.
• Emerging extensions and refinements
While the history of authentic nonviral replicating episomes (S/MAR-plasmids) goes back to 1999
[48] and while its purification can rely on standard procedures, the generation, identification and
recovery of minicircle derivatives is much more demanding as it has to rely on several dedicated
routines. These routines may even enhance the danger of secondary rearrangements, due to
S/MAR-intrinsic instabilities.
Common protocols involve the action of site-specific nucleases of either the threonine-
(lambda- Cre-, Flp-) or the serine (ΦC31-, and ParA-) family. These activities can be provided

36
- either from inducible, genomically anchored genes of modified E.coli strains; induction is
either performed by a temperature shift [83] or a metabolite, typically arabinose [82].
- alternatively, the recombinase is located in the (MP-)part of the parental plasmid, which
accommodates the auxiliary functions (miniplasmid-section).
Whereas our initial experiments firmly relied on the first concept (E.coli strain MM294 with a
genomic copy of heat-inducible Flp [83]), our efforts have recently shifted to the second option.
Due to the fact that for bacteria mRNA is translated into protein as soon as it is transcribed, this
altered concept led to vast improvements regarding yield and also purity (S. Binius, Dissertation
TU Braunschweig, 2011). Another important consideration motivating this change is the possibility
to select the bacterial strain according to the requirements, especially regarding its propensity to
generate unwanted concatemeric side products, which (among others) depends on RecA+ and
RecF+ functions. A reverse switch of strategies (transfer of auxiliary genes from the parental
plasmid to genomic locations) was taken by Kay et al [82]. Their refined protocol has to
accommodate three auxiliary genes (two for recombinases, and one encoding the homing
endonucleases I-SceI), excluding alternative options.
A more recent approach addresses the creation of minicircles from plasmid precursors in
vitro. The procedure involves the simple excision of unwanted sequences by restriction enzymes
and subsequent end-ligation. While this protocol per se would fail to produce active minicircles due
to the absence of a superhelical structure, the appropriate topological state can be adjusted by in
vitro treatments, either using histone-like proteins followed by topoisomerase [92] or by gyrase
([93] Dissertation M. Heine, TU Braunschweig 2012). Such a procedure may profit from the fact
that superhelicity can be controlled. After optimization, this may positively influence early
transcription and replication steps and thereby establishment in the host.
Combination of excision- and RMCE strategies Following a long standing tradition our focus is
on Flp-dependent recombinase protocols as these open a variety of simultaneous or successive
modification steps (see, for instance, the RMCE example in Fig. 5 and the toolboxes described in
ref. [39, 67]). Concerns that the Flp-mediated generation would suffer from the principal

37
reversibility of Flp reactions did not hold in this case, since excision is strongly preferred over
addition due to kinetic and thermodynamic principles 40].
As an intermediary solution we have developed a protocol that allows minicircle generation
directly in the recipient eukaryotic cell. In contrast to the replicating MC, the MP product will be lost
in dividing tissues by dilution. Currently, this procedure enables a wide range of pilot studies. In
case of success the relevant experiment can be repeated using a parental plasmid derivative that
enables MC generation in E.coli followed by purification routines (S. Binius, Dissertation TU
Braunschweig, 2011). Work is underway to recover the MC-sequences from an auto-processing
PP system which proved effective in vivo. These sequences will then be incorporated into the
standard backbone permitting large-scale MC production in E.coli (ref. [4] and contributions of
these authors in the present book edition).
Central features of our processing systems for eukaryotic cells are summarized in Fig. 14.
A PSV40-Flp-expression unit becomes active as soon as the parental plasmid is transferred to the
recipient cell. The educt is then processed by Flp-catalyzed crossover between two FRT wild type
sites (red half arrows). This excision separates Flp from its promoter, which now serves to drive a
positive/negative selection marker and/or the egfp reporter gene to provide the following functions:
- an auto-limiting feature (no additional Flp-activity is generated after this step);
- if placed appropriately, the reporter gene becomes activated and permits quantification of
the excision reaction; controls have proven that this process proceeds to completion (A.
Oumard, N. Heinz et al., in progress).
To add RMCE options comparable to Fig. 5, we will provide the PP with a third, heterospecific FRT
site (yellow half arrow in Fig. 14). This site does not interfere with the excision reaction but enables
subsequent exchange of a cassette and thereby the introduction of new functions into an
established episome (cf. Fig. 5). In the given example, a positive-negative selection marker (for
instance the hygtk fusion gene) may have served to support establishment of the minicircle in the
presence of Hygromycin and to enrich successful RMCE events by counter-selection (here in the
presence of GANC). Advantages of this process arise from the facts that (i) no potentially

38
debilitating, sorting steps are required at early time points and (ii) RMCE will establish a pFAR
situation.
Due to the chronological sequence of events that led to the development of minicircles from
the original S/MAR-plasmids, most accessory options have originally been explored at the level of
(parental-) plasmids before being transferred to the ultimate vectors. In this context the exploitation
of selection procedures for episomal establishment and supplementation of S/MAR activities by
initiator-of-replication (IR-) are of prime importance (see chapter “Establishment and maintenance:
The EBV paradigm”). On balance, certain intrinsic differences between the PP- and MC systems
have to be taken in consideration, exemplified by the following paragraph.
MC withdrawal at will For several applications it would be valuable to have a tool available for
withdrawing episomal vectors at will. Since the Yamanaka group [94] could apply the combined
expression of reprogramming factors to establish induced pluripotent stem (iPS-) cells the
development of novel approaches for their delivery in a reversible, dose-controlled fashion has
become an active area of research. Current refinements concern the targeted differentiation of
iPSCs into functional somatic cells, and the approaches for a conditional elimination of certain
iPSC-progenitors that might otherwise raise to teratoma-intiating cells (TICs),.
Due to the risk of insertional mutagenesis, viral transduction routes become increasingly
replaced by nonviral methods in order to induce the iPSC status. Among the alternatives is a
recent report building on a standard minicircle, which, in contrast to a regular plasmids, has
enabled the generation of iPSC clones from human adipose stem cells [95]. The superior
performance of the MC is ascribed to higher transfer efficiency together with stronger and more
persistent expression characteristics. Current extensions of this concept rely on replication-
competent S/MAR minicircles as these permit a prolonged expression in diving cells. We might be
able to terminate this phase at will, given the availability of strategies that permit withdrawal of the
expression unit. A candidate approach might exploit the role of active transcription into the S/MAR
but so far there are indications that traversal of the S/MAR is only required for episomal
establishment rather than long-term maintenance. Two recent reports address this question, but
lead to opposite conclusions.

39
The first study [96] concerns persistence of episomes in CHO-K1 cells in case transcription
of the egfp gene is regulated using the Tet-On system, which permits transcription only in the
presence of doxycycline. Removal of the antibiotic is shown to cause three-fold reduction of
expressing cells and an about twelve-fold reduction in fluorescence activity. Although the data
indicate some leakiness, these observations led to the conclusion that these phenomena were
governed by vector loss.
In the second example [64] pEpi derivatives carrying a tandem array of lac operator
sequences are used to enable visualization and modulation of the episome´s chromatin status. For
CHO-K1 cells carrying established episomes, only 5% express the reporter gene at a detectable
level. Treatment with inhibitors of DNA-methyltransferases (5-aza-dC) and/or histone-deacetylases
(TSA) resulted in an almost 4-fold increase in the percentage of cells with detectable eGFP
expression. This indicated that decreasing fluorescence had to be ascribed to silencing rather than
vector loss.
Our lab has applied the capacity of retroviral particles to transfer mRNA (RMT), episomal
DNA (RET) and membrane- as well as intracellular proteins (RPT). These processes rely on
systematic blocks within the regular RV life cycle, and are covered by the term retroviral “pseudo-
transduction” [97]. In the present context RET is of particular interest, since episomal intermediates
persist due to inactivated integrase functions (cf. the retroviral delivery of LTR-circles indicated in
Fig. 1). Episomes of this origin have been successfully applied for gene expression over a limited
period of time. [88,98]. Although we anticipated that these entities would gain replication capacity
by the introduction of an S/MAR we had to find a rapid disappearance of reporter gene
fluorescence. This phenomenon could be reproduced using a fully synthetic 2-LTR minicircle-
analogue that could be transferred, by lipofection, at elevated copy numbers. Induction of histone
hyperacetylation by butyrate resulted in a transient recovery of fluorescence for a cell population
with the 2-LTR circle in an episomal state. Since these observations could be repeated in several
subsequent inactivation/activation cycles, they are consistent with the idea that, once established,
lack of transcription does not lead to vector-loss (A. Oumard, unpublished). It is of note that LTR-

40
dependent silencing is a well known phenomenon that has been ascribed to negative regulatory
factors associating with its 5´end.
A similar conclusion goes back to minicircles encoding a GFP reporter and a separate milk-
specific expression cassette. After establishment these MCs were stably transmitted for more than
three month in monoclonal primary bovine fibroblast lines even in the absence of continued
selection. Serum starvation greatly reduced GFP fluorescence, which, however, could be fully
restored after serum was re-added to the medium. These data confirm that established minicircles
are not lost during periods without transcription ([99] and in preparation), which is the prerequisite
for cell modifications to survive early embryonic development phases in the absence of gene
activity.
Only further studies can show whether or not early stages of establishment are different for
minicircles and S/MAR-plasmids, in that the latter group meets more stringent requirements for its
persistence as indicated by differences under the conditions of the Fig. 8, 11 and 13. To overcome
barriers of this type, methods are being developed that will disable minicircle persistence by the
recombinase-mediated excision of “floxed” or “flirted” (loxP- or FRT-flanked) S/MAR inserts [1].
Pronuclear injection and somatic cell nuclear trans fer Very little has been reported on attempts
to produce animals that ubiquitously express episomes. Manzini et al. [100] generated transgenic
pig fetuses by sperm mediated gene transfer (SMGT) and showed the episome to confer
expression of the transgene marker GFP in most cells and tissues. To our knowledge, though, no
live animals with episomes have been generated. Recent experiments try to fill this gap, and will
allow addressing the question as to whether episomes are stably and ubiquitously expressed and
passed on to the next generation through the germ line.
Here two approaches are of prime relevance. Somatic cell nuclear transfer (SCNT) is a two-
step process in which a gene construct is first introduced into somatic cells, followed by transfer
into enucleated oocytes. Since the birth of “Dolly”, the first animal to be cloned from an adult cell in
1997, it is established that physiologically normal beings can be generated by SCNT. Since then
the procedure has been verified for the major livestock species including cattle, goats, pigs and

41
deer in addition to laboratory rodents. Ongoing studies use minicircles for the generation of cows
expressing transgenic proteins in milk as mentioned above [99]. To this end MCs with a GFP
reporter gene and a lactation-specific expression cassette under the control of the murine whey
acidic protein (WAP) are transfected into bovine fibroblasts. Following molecular characterization,
these cells are applied as donors for SCNT to generate transgenic offspring.
Another approach starts with the pronuclear transfer of minicircles and other supercoiled
DNAs enabling ectopic gene expression in embryos, used to study reprogramming events during
early ontogenesis. While these expectations can be met by the classical, demanding pronuclear
injection technology, identical results were obtained by simple cytoplasmic injection of vectors with
ccc-status [59]. This reveals a nuclear transfer mechanism of yet unknown nature. While the
conventional technology had to live with random aspects as it resulted in the integration of one or
multiple copies of a gene into one or several unspecified genomic loci these can, at least in
principle, be overcome by minicircle transfer. Present emphasis is posed on a better understanding
of the molecular interactions by which episomes become productively established such that
minicircles gain replication potential in early embryos.
From cells to organs S/MAR vectors devoid of extraneous bacterial sequences could be applied
to provide high and sustained transgene expression in the recipient cells. A common in vivo model
relies on liver into which either S/MAR plasmids or S/MAR minicircles can be transferred by
hydrodynamic injection. While the expression from a prototype S-MAR plasmid dropped to 10% of
its initial level within 25 days, in case of the minicircle its luciferase expression remained for the
entire three-month period of the experiment. At this time it was approximately two orders of
magnitude higher than for both the S/MAR-free minicircle control and the S/MAR parental plasmid
[101].
Partial hepatectomy on S/MAR minicircle treated mice caused a rapid drop of expression
due to the lack of vector replication but there are present approaches to overcome this
phenomenon by providing a survival advantage associated with the MC. Already now the ongoing
studies underline the utility of minicircles for persistent, atoxic gene expression in the liver. They

42
clearly demonstrate the benefit of an intrinsic S/MAR also for expression parameters not directly
related to active replication [81].
SUMMARY AND OUTLOOK
Recombinant viruses are widely utilized as vectors for gene transfer. They have, however, certain
intrinsic drawbacks including a limited opportunity for repeated administrations due to acute
inflammatory and delayed immune responses. For vectors that integrate foreign DNA into the
genome insertional mutagenesis has become a major issue, which has directed attention to viral
episomes such as OriP vectors that rely on the replication machinery of the Epstein-Barr virus. In
this and other cases viral proteins (here: EBNA1) are able to mimic the function of chromosomal
proteins in order to exploit the replication functions of the host cell.
Since each viral vector carries a potential risk, intense efforts have been launched to create
artificial chromosomes using telomeres, centromeres and intrinsic Ori functions. These systems
have suffered from considerable instability in addition to the fact that, with a few exceptions,
functional mammalian Oris have remained barely defined due to their extended and multifaceted
structure. This is definitely different for yeast where Autonomously Replicating Sequences (ARS)
have been defined permitting the facile construction of episomes with a function that is largely
restricted to this species. A property common to yeast and mammalian Oris is the association with
a Scaffold/Matrix Attachment Region (S/MAR), an element than can confer a multitude of activities
to vectors provided that these obey chromosomal organization principles. A parameter of particular
relevance is the topological status, which provides structural imprints adapting the recognition
potential and function of S/MARs to a given situation. The observation that an S/MAR, tuned by an
adjacent transcription unit, can be used to provide plasmids with replication potential led to the first
nonviral plasmid episome (pEpi) in 1999. pEpi, in fact, was the first example of a vector with ARS-
like functions in mammalians.
Regarding the associated properties, an S/MAR episome resembles EBV vectors in that it
provides both “molecular glue” (MG-) and “initiator of replication (IR-) functions. EBV builds on

43
interaction of the viral EBNA1 protein with the repetitive EBV-derived “family of repeats” (FR) to
address chromosomes and replication machineries of the host cell. For the nonviral homologue the
respective functions go back to the S/MAR in association with the cellular SAF-A/hnRNP-U protein.
In both cases complex formation is governed by the repetitive structure of the DNA site, which
leads the partner protein to oligomerize and to enter strong, but reversible “mass-binding”
interactions. The IR-function on the other hand is well characterized for EBV, where it localizes to a
dyad-symmetry (DS) element accommodating the EBNA-dimer. Unexpected at first, S/MARs do
not harbor a defined initiator of bidirectional replication element but rather induce structural
changes permitting replication initiation over the plasmid’s entire length. Present efforts are
directed to combining S/MAR- and IR- functions on a single, nonviral episome, which however is
only one among a variety of approaches described in this review to optimize this vector class.
A relevant improvement that is well underway is the conversion of S/MAR plasmids into
“minicircles”. Minicircles per se are already well established vectors, which, due to the deletion of
prokaryotic plasmid parts, resist host-defense actions that would otherwise lead to silencing.
Minicircles have proven considerable therapeutic potential after hydrodynamic gene transfer or jet
injection into barely dividing tissues such as liver and muscle. This review demonstrates that these
entities can be supplemented by an S/MAR, which, per se, largely improves the expression
properties of this novel vehicle. In conjunction with a transcription unit it provides replication
functions and nuclear establishment, which, although largely superior to S/MAR plasmids, remains
a target for further optimization.
The term “establishment” covers the interval between vector transduction and its functional
association with the nuclear substructures providing replication potential. After this stage the vector
has reached its ultimate chromatin structure, which permits its long term maintenance with an
efficiency that is unprecedented by other types of episomes. In the present article we have shown
that this stable association permits the isolation of clonal cell lines with predictable properties going
back to one or even several distinct minicircles that can be accommodated in parallel and lend
themselves to further modification in situ.

44
In conclusion, S/MAR-minicircles combine the properties of efficient transient expression
systems (facilitated membrane transfer and physical stability leading to an extended transcriptional
burst) with those of stable expression systems. The transition between both phases is smooth such
that comprehensive procedures for a wide variety of purposes can be envisaged. Current
bottlenecks on the way to S/MAR-MCs with adequate purity and an established ccc-status have
been identified and enable industrial scale routine preparation (102).
Acknowledgements
Our particular thanks go to all colleagues who communicated ideas and contributed to this project
[44, 49, 54, 59, 64, 71]. We gratefully acknowledge support by Martin Schleef (PlasmidFactory
Bielefeld) who provided a replicating S/MAR minicircle (corresponding to our vector M18) produced
from our components and ideas for upcoming joint projects. Work in the authors lab at Hannover
Med. School has been supported by the the CliniGene Network of Excellence
(EuropeanCommission FP6 Research Program, contract LSHBCT-2006-018933), the Excellence
Initiative “REBIRTH” (From Regenerative BIology to Reconstructive THerapy, the SFB 738
(Optimierung konventioneller und innovativer Transplantate) and ReGene (Regenerative Medizin
und Biologie) grants, provided to the consortium by the BMBF.

45
REFERENCES
[1] Turan, S. and Bode, J. Site-specific recombinases: from tag-and-target- to tag-and-exchange-based genomic modifications FASEB J. (2011) 25:4088-4107 http://dx.doi.org/10.1096/fj.11-186940
[2] Haase, R., Argyros, O., Wong, S. P.,Harbottle, R. P. et al., pEPito: a significantly improved non-viral episomal expression vector for mammalian cells. BMC Biotechnol. 2010, 10:20.
http://www.biomedcentral.com/1472-6750/10/20
[3] Marie, C., Vandermeulen, G., Quiviger, M., Richard, M. ,et al., pFARs, Plasmids free of antibiotic resistancemarkers, display high-level transgene expression in muscle, skin and tumour cells. J. Gene Med. 2010, 12, 323–332. doi: 10.1002/jgm.1441.
[4] Mayrhofer, P., Schleef, M., Jechlinger, W.,Use of Minicircle Plasmids for Gene Therapy in Walther, W., Stein, U. S. (Eds.), Methods in Molecular Biology, Gene Therapy of Cancer, Humana Press 2009, pp. 87-104. DOI: 10.1007/978-1-59745-561-9_4.
[5] Bode, J., Benham, C., Knopp, A., Mielke, C., Transcriptional augmentation: modulation of gene expression by scaffold/matrix attached regions (S/MAR elements). Crit. Rev. Eukaryot. Gene Expr. 2000, 10, 73 – 90.
[6] Guo, B., P. R. Odgren, A. J. van Wijnen, T. J. Last, et al., The nuclear matrix protein NMP-1 is the transcription factor YY1. Proc. Natl. Acad. Sci. USA 1995. 92, 10526–10530.
[7] Klar, M., Stellamanns, E., Ak, P., Gluch, A. et al., Dominant genomic structures: detection and potential signal functions, Gene 2005, 364, 79-89. http://dx.doi.org/10.1016/j.gene.2005.07.023
[8] Klar. M., Bode, J., Enhanceosome formation over the interferon-beta promoter underlies a remote-control mechanism mediated by YY1 and YY2. Mol. Cell. Biol. 2005, 25, 10159-10170. http://dx.doi.org/10.1016/j.jmb.2005.11.073
[9] Johnson, C. N. , Levy, L. S., Matrix attachment regions as targets for retroviral integration. Virol. J. 2005 2: 68. doi:10.1186/1743-422X-2-68.
[10] Gluch, A., Vidakovic, M. , Bode, J. Scaffold/matrix Attachment Regions (S/MARs): Relevance for Disease and Therapy, in Protein-Protein Interactions as New Drug Targets / Handbook of Experimental Pharmacology 186, Klussmann E., Scott JD (Eds), Springer 2008, pp 67-103. http://www.springer.com/series/164
[11] Festenstein, R., Tolaini,M., Corbella, P., Mamalaki, C., et al., Locus control region function and heterochromatin-induced position effect variegation. Science 1996, 271, 1123-1125.
[12] Bode J, Bartsch J, Boulikas T, Iber M, et al., Transcription-Promoting Genomic Sites in Mammalia: Their Elucidation and Architectural Principles. In: “Textbook of Gene Therapy and Molecular Biology”, Gene Therapy and Molecular Biology Publications, 1998 T. Boulikas, (Ed.), Gene Therapy Press, Athens, Greece pp 551-580.

46
[13] Goetze, S., Baer, A., Winkelmann, S., Nehlsen, K., Genomic bordering elements: their performance at pre-defined genomic loci. Mol. Cell. Biol. 2005, 25, 2260-2272.
[14] Qiao, J., Oumard, A., Wegloehner, W. , Bode, J., Novel tag-and-exchange (RMCE) strategies generate master cell clones with predictable and stable transgene expression properties. J. Mol. Biol. 2009, 390, 579–594. doi:10.1016/j.jmb.2009.05.012
[15] Cai, S., Han, H. J., Kohwi-Shigematsu, T., Tissue-specific nuclear architecture and gene expression regulated by SATB1. Nat. Genet. 2003, 34 (1): 42–51. doi:10.1038/ng1146
[16] Cockerill, P. N., Garrard, W. T., Chromosomal loop anchorage of the kappa immunoglobulin gene occurs next to the enhancer in a region containing topoisomerase II sites. Cell 1986, 44, 273-282.
[17] Cockerill, P. N., Yuen, M. H., Garrard, W. T., The enhancer of the immunoglobulin heavy chain locuIs is flanked by presumptive chromosomal loop anchorage elements. J. Biol. Chem. 1987, 262, 5394-5397.
[18] de Moura Gallo, C. V., Vassetzky, Y. S., Targa, F. R., Georgiev, G. P. et al., The presence of sequence-specific protein binding sites correlate with replication activity and matrix binding in a 1.7 Kb-long DNA fragment of the chicken alpha-globin gene domain. Biochem. Biophys. Res. Commun. 1991, 179(1):512-519.
[19] Handeli S., Klar A., Meuth M., Cedar H., Mapping replication units in animal cells. Cell 1989, 57(6), 909-920.
[20] Schuebeler, D., Mielke, C., Maass, K., Bode, J., Scaffold/Matrix-attached regions act upon transcription in a context-dependent manner. Biochemistry 1996, 35:11160-11169.
[21] Agarwal, M., Austin, T. W., Morel, F., Chen, J. et al., Scaffold Attachment Region-Mediated Enhancement of Retroviral Vector Expression in Primary T Cells. J. Virol. 1998, 72(5), 3720–3728.
[22] Auten, J., Agarwal, M., Chen, J., Sutton, R. et al., Effect of Scaffold Attachment Region on Transgene Expression in Retrovirus Vector-Transduced Primary T Cells and Macrophages. Hum. Gene Ther. 1999. 10, 1389-1399.
[23] Dang, Q., Auten, J., Plavec,I., Human beta interferon scaffold attachment region inhibits de novo methylation and confers long-term, copy number-dependent expression to a retroviral vector, J. Virol. 2000, 74, 2671–2678.
[24] Murray, L., Travis, M., Luens-Abitorabi, K., et al., Addition of the human interferon beta scaffold attachment region to retroviral vector backbones increases the level of in vivo transgene expression among progeny of engrafted human hematopoietic stem cells. Hum. Gene Ther. 2000, 11, 2039-2050.
[25] Kurre, P., Morris, J., Thomasson, B., Kohn, D. B. et al., Scaffold attachment region–containing retrovirus vectors improve long-term proviral expression after transplantation of GFP-modified CD34+ baboon repopulating cells. Blood 2003, 102(9), 3117-3119.

47
[26] Ramezani, A., Hawley, T. S., Hawley, R.G., Performance- and safety-enhanced lentiviral vectors containing the human interferon-beta scaffold attachment region and the chicken beta-globin insulator, Blood 2003, 101, 4717–4724.
[27] Ma, Y., Ramezani, A., Lewis, R., Hawley, R. G., Thomson, J. A., High-Level Sustained Transgene Expression in Human Embryonic Stem Cells Using Lentiviral Vectors. Stem Cells 2003, 21, 111-117.
[28] Moreno, R., Martinez, I., Petriz, J., González, J. R. et al., Boundary sequences stabilize transgene expression from subtle position effects in retroviral vectorsBlood Cells, Molecules, and Diseases 43 (2009) 214–220
[29] Moreno, R., Martínez, I., Petriz, J., Nadal, M. et al., The β-Interferon Scaffold Attachment Region Confers High-Level Transgene Expression and Avoids Extinction by Epigenetic Modifications of Integrated Provirus in Adipose Tissue-Derived Human Mesenchymal Stem Cells. Tissue Engineering (C) ahead of print. doi:10.1089/ten.tec.2010.0383.
[30] Kay, V., Bode, J., Detection of scaffold-attached regions (SARs) by in vitro techniques; activities of these elements in vivo. In: Methods in Molecular and Cellular Biology 1995, 5, 186-194 - Methods for studying DNA-protein interactions - an overview, Papavassiliou A. G., King, S. L., (eds.) Inc. Wiley-Liss.
[31] Goetze S, Y. Huesemann, A. Baer, Bode J., Functional characterization of transgene integration patterns by halo-FISH: electroporation versus retroviral infection. Biochemstry 2003, 42, 7035-7043.
[32] Bode, J., Winkelmann, S., Goetze, S., Spiker, S., et al., Correlations between scaffold/matrix attachment region (S/MAR) binding activity and DNA duplex destabilization energy. J. Mol. Biol. 2006, 358, 597-613.
http://dx.doi.org/10.1016/j.jmb.2005.11.073
[33] Bode, J., Goetze, S., Heng, H., Krawetz, S. A., Benham, C., From DNA structure to gene expression: mediators of nuclear compartmentalization and -dynamics. Chromosome Res. 2003, 11, 435-445. http://dx.doi.org/10.1023/A:1024918525818
[34] Benham, C. J., Duplex destabilization in superhelical DNA is predicted to occur at specific transcriptional regulatory regions. J. Mol. Biol. 1996, 255, 425-434.
[35] Kouzine, F., Liu, J., Sanford, S., Chung, H. J. et al., The dynamic response of upstream DNA to transcription-generated torsional stress. Nature Struct. Mol. Biol. 2004, 11, 1092 – 1100.
doi:10.1038/nsmb848.
[36] Bode, J., Maass, K. (1988) The chromatin domain surrounding the human interferon-beta gene as defined by scaffold-attached regions (SARs). Biochemistry 1988, 27, 4706-4711.
[37] Kipp, M.,Gohring, F., Ostendorp,T., vanDrunen, C. M. et al., SAF-Box, a conserved protein domain that specifically recognizes scaffold attachment region DNA. Mol. Cell. Biol. 2000, 20, 7480–7489.

48
[38] Kawano, S., Miyaji, M., Ichiyasu, S., ,Tsutsui, K. M. et al., Regulation of DNA Topoisomerase II through RNA-dependent Association with Heterogeneous Nuclear Ribonucleoprotein U (hnRNP-U). J. Biol. Chem. 2010, 285(34), 26451–26460.
[39] Bode J, Schlake T, Iber M, Schuebeler D, et al., The transgeneticist´s toolbox: novel methods for the targeted modification of eukaryotic genomes. Biol. Chem. 2000, 381, 801-813.
[40] Baer, A., Bode, J., Coping with kinetic and thermodynamic barriers: RMCE, an efficient strategy for the targeted integration of transgenes”. Curr. Opin. Biotechnol. 2001, 12, 473-480. http://dx.doi.org/10.1016/S0958-1669(00)00248-2
[41] Weintraub, H., Cheng, P. F., Conrad, K., Expression of transfected DNA depends on DNA topology. Cell 1986, 46, 115-122.
[42] Oumard A, Qiao J, Jostock T, Li T , Bode J., Recommended Method for chromosome Exploitation: RMCE-based cassette-exchange systems in animal cell biotechnology. Cytotechnology 2006, 50, 93–108. doi: 10.1007/s10616-006-6550-0.
[43] Amati, B. B., Gasser, S. M., Chromosomal ARS and CEN elements bind specifically to the yeast nuclear scaffold. Cell 1988, 54, 987-978.
[44] Ak, P., Benham, C. J., Susceptibility to superhelically driven DNA duplex destabilization: a highly conserved property of yeast replication origins. PLoS Comput. Biol. 2005, e7, 41–46.
[45] Li, H., Yang, Y., Zhang, K. Zheng, C., Identification and analysis of ARS function of six plant MARs. Chin. Sci. Bull. 2004, 49(5), 462-466. DOI: 10.1007/BF02900965
[46] Broll S, Oumard A, Hahn K, Schambach A , Bode, J., Minicircle Performance Depending on S/MAR-Nuclear Matrix Interactions. J. Mol. Biol. 2010, 395(5), 950-965.
http://dx.doi.org/10.1016/j.jmb.2009.11.066
[47] Dijkwel, P. A. , Hamlin, J. L., Matrix attachment regions are positioned near replication initiation sites, genes, and an interamplicon junction in the amplified dihydrofolate reductase domain of Chinese hamster ovary cells. Mol Cell Biol. 1988, 8(12): 5398-5409.
[48] Piechaczek, C., Fetzer, C., Baiker, A., Bode, et al.., A vector based on the SV40 origin of replication and chromosomal S/MARs replicates episomally in CHO cells. Nucleic Acid Res 1999, 27, 426-428.
[49] Baiker, A., Maercker, C., Piechaczek, C., Schmidt, S. B. A., et al., Mitotic stability of a human scaffold/matrix attached region containing episomal vectors is provided by association with nuclear matrix. Nat. Cell Biol., 2000, 2, 182-184.
[50] Schaarschmidt, D., Baltin, J., Stehle, I. M., Lipps, H. J., et al., An episomal mammalian replicon: sequenceindependent binding of the origin recognition complex. EMBO J 2004, 23, 191-201.
[51] Bode J, Fetzer CP, Nehlsen K, Scinteie M, et al. The hitchhiking principle: optimizing episomal vectors for the use in gene therapy and biotechnology. Gene Ther. Mol. Biol 2001, 6, 33-46.

49
[52] Nehlsen, K., Molekulare Grundlagen der episomalen Replikation: Charakterisierung zirkulärer, nichtviraler Vektoren. Dissertation Technische Universität Braunschweig 2004. http://opus.tu-bs.de/opus/volltexte/2004/587/pdf/Dissertation.pdf
[53] Stehle, I. M., Scinteie, M. F., Baiker, A., et al., Exploiting a minimal system to study the epigenetic control of DNA replication: the interplay between transcription and replication. Chromosome Res. 2003, 11, 413-421.
[54] Sgourou, A., Routledge, S., Spathas, D., Athanassiadou, A. et al. Physiological levels of HBB transgene expression from S/MAR element-based replicating episomal vectors. J. Biotechnol. 2009, 143, 85–94, doi:10.1016/j.jbiotec.2009.06.018.
[55] Meaking, W.S., Edgerton,J., Wharton, C. W. , Meldrum, R. A., Electroporation-induced damage in mammalian cell DNA. Biochim. Biophys. Acta 1995, 1264, 357-362.
[56] Nickoloff, J. A., Spirio, L. N., Reynolds, R. J. A. Comparison of Calcium Phosphate Coprecipitation and Electroporation - Implications for Studies on the Genetic Effects of DNA Damage. Mol. Biotech. 1998, 10, 93-101.
[57] Mthunzi, P., Dholakia, K., Gunn-Moore, F., Phototransfection of mammalian cells using femtosecond laser pulses: optimization and applicability to stem cell differentiation. J. Biomed. Optics 2010, 15(4), 041507-1 - 041507-7.
[58] Hsu, C. Y. , Uludağ, H., Effects of size and topology of DNA molecules on intracellular delivery with non-viral gene carriers. BMC Biotechnol. 2008, 8:23.
[59] Iqbal, K., Barg-Kues, B., Broll, S., Bode. J., et al., Cytoplasmic injection of circular plasmids allows targeted expression in mammalian embryos. BioTechniques 2009, 47, 959–968
http://dx.doi.org/10.2144/000113270.
[60] Vacik, J., Dean, B. S., Zimmer, W. E. and Dean, D. A., Cell-specific nuclear import of plasmid DNA. Gene Ther. 1999, 6, 1006-1014.
[61] van der Vliet, P. C., Roles of Transcription Factors in DNA Replication in DNA replication in eukaryotic cells. CSH Monographs 1996, 31 (1996): DNA Replication in Eukaryotic Cells 31 1996 DOI: 10.1101/087969459.31.87
[62] Stehle, I. M., Postberg, J., Rupprecht, S., Cremer, T. et al., Establishment and mitotic stability of an extra-chromosomal mammalian replicon. BMC Cell Biol 2007, 8:33.
[63] Jenke, B. H., Fetzer, C. P., Jönsson, F., Fackelmayer, et al.., An episomally replicating vector binds to the nuclear matrix protein SAF-A in vivo. EMBO Rep. 2002, 3, 349–354
[64] Tessadori , F., Zeng, K., Manders, E., Riool, M. et al., Stable S/MAR-based episomal vectors are regulated at the chromatin level. Chromosome Res. 2010, 18(7):757-775.
DOI 10.1007/s10577-010-9165-4

50
[65] Belmont A. S., Visualizing chromosome dynamics with GFP. Trends Cell Biol 2001, 11, 250–257.
[66] Chen, Z. Y., He, C. Y., Erhardt, A., Kay, M. A., Minicircle DNA vectors devoid of bacterial DNA result in persistent and high-level transgene expression in vivo. Mol Ther 2003, 8,
495-500.
[67] Turan, S., Kuehle, J., Schambach, A., Baum, et al., RMCE-multiplexing: versatile extensions of the Flp-Recombinase-Mediated Cassette-Exchange technology. J. Mol. Biol. 2010, 402 , 52-69. http://dx.doi.org/10.1016/j.jmb.2010.07.015
[68] Kumar, P., Saha, A., Robertson E. S., Epstein-Barr Virus Hijacks Cell-Cycle Machinery. Microbe 2010, 5(6) 251-256.
[69] Wang, C. Y., Sugden, B., Identifying a property of origins of DNA synthesis required to support plasmids stably in human cells. Proc. Natl Acad. Sci. USA 2008, 105, 9639–9644.
[70] Rangasami, D., An S/MAR-based L1 retrotransposition cassette mediates sustained levels of insertional mutagenesis without suffering from epigenetic silencing of DNA methylation. Epigenetics 2010, 5(7), 601-611.
[71] Giannakopoulos, A., Stavrou, E. F.,Zarkadis, I., Zoumbos, N. et al., The Functional Role of S/MARs in Episomal Vectors as Defined by the Stress-Induced Destabilization Profile of the Vector Sequences J. Mol. Biol. 2009, 387, 1239–1249.
[72] Wong, S. P., Argyros, O., Coutelle, C., Harbottle, R. P., Non-viral S/MAR vectors replicate episomally in vivo when provided with a selective advantage. Gene Ther. 2010, 18, 82-87.
[73] Kaufman, W. L., Kocman, I., Agrawal, V., Rahn, H. P. et al.., Homogeneity and persistence of transgene expression by omitting antibiotic selection in cell line isolation. Nucleic Acids Res. 2008, 36, e111
[74] Fuks, F., Hurd, P.J., Wolf, D., Nan, X., Bird, A.P., Kouzarides, T., The methyl-CpG-binding protein MeCP2 links DNA methylation to histone methylation. J. Biol. Chem. 2003, 278, 4035–4040.
[75] Chen, Z. Y., Riu, E., He, C. Y., Xu, H., Kay, M. A., Silencing of episomal transgene expression in liver by plasmid bacterial backbone DNA is independent of CpG methylation. Mol. Ther 2008, 16, 548-556.
[76] Williams, S., Mustoe, T., Mulcahy, T., Griffiths, M. et al., CpG-island fragments from the HNRPA2B1/CBX3 genomic locus reduce silencing and enhance transgene expression from the hCMV promoter/enhancer in mammalian cells. BMC Biotech. 2005, 5:17. doi:10.1186/1472-6750-5-17
[77] Schroth, G. P., Ho, P. S., Occurrence of potential cruciform and H-DNA forming sequences in genomic DNA. Nucleic Acids Research, 1995, 23(11), 1977-1983.

51
[78] Lufino, M. M., Manservigi, R. Wade-Martins, R., An S/MAR-based infectious episomal genomic DNA expression vector provides long-term regulated functional complementation of LDLR deficiency. Nucleic Acids Res 2007, 35: e98.
[79] Bode, J., Kohwi, Y., Dickinson, L., Joh, T., Klehr, D., Mielke, C., Kohwi-Shigematsu, T., Biological significance of unwinding capability of nuclear matrix-associating DNAs. Science 1992, 255, 195-197.
[80] Jenke, A. C., Stehle, I. M., Herrmann, F., Eisenberger, T. et al., Nuclear scaffold/matrix attached region modules linked to a transcription unit are sufficient for replication and maintenance of a mammalian episome. Proc. Nat.l Acad Sci. USA 2004, 101, 11322–11327.
[81] Chen, Z. Y. , Yant, S. R., He, C. Y., Meuse, L. et al., Linear DNAs concatemerize in vivo and result in sustained transgene expression in mouse liver. Mol. Ther. 2001, 3, 403-410.
[82] Kay, M., He, C. H. Chen, Z. Y., A robust system for production of minicircle DNA vectors Nature Biotech. 2010, 28(12), 1287-1291. doi:10.1038/nbt.1708
[83] Nehlsen, K., Broll, S., Bode, J., Replicating minicircles: Generation of nonviral episomes for the efficient modification of dividing cells. Gene Ther. Mol. Biol. 2006, 10, 233-244.
[84] Benham C, T. Kohwi-Shigematsu, Bode, J., Stress Induced Duplex DNA Destabilization in Scaffold/ Matrix Attachment Regions. J. Mol. Biol. 1997, 272, 181-196.
http://dx.doi.org/10.1006/jmbi.1997.1385.
[85] Takagaki, Y. Manley, J. L., Complex protein interactions within the human polyadenylation machinery identify a novel component. Mol. Cell. Biol. 2000, 20(5), 1515–1525.
[86] Wegner, M., Zastrow, G., Klavinius, A., Schwender, S. et al., Cis-acting sequences from mouse rDNA promote plasmid DNA amplification and persistence in mouse cells: implication of HMG-I in their function. Nucleic Acids Res. 1989, 17, 9909-9932.
[87] Schmidt, M., Schwarzwaelder, K., Bartholomae, C., Zaoui, K. et al., High-resolution insertion-site analysis by linear amplification–mediated PCR (LAM-PCR). Nature Meth. 2007, 4(12), 1051-1057.
[88] Wanisch, K., Yáñez-Muñoz, R. J., Integration-deficient lentiviral vectors: A slow coming of age. Mol. Ther. 2009, 17(8), 1316-1332.
[89] Klehr, D., Bode, J., Comparative evaluation of bovine papilloma virus (BPV): vectors for the study of gene expression in mammalian cells. Mol. Gen. (Life Science Adv.) 1988, 7, 47-52.
[90] Friedman, E. A., Smiths, H. O., An Adenosine Triphosphate-dependent deoxyribonuclease from Hemophilus influenzae - Substrate specificity. J. Biol. Chem. 1972, 247(9), 2859-2865.
[91] Shapiro, H. M., Practical Flow Cytometry: 3rd ed. Wiley-Liss, Inc., 1995, pp 366.

52
[92] Barth, M. C., Dederich, D. A., Dedon, P- C., An improved method for large-scale preparation of negatively and positively supercoiled plasmid DNA. Biotechniques 2009, 47, 633-635.
[93] Zhixing, Y., Nahon, J. L., DNA gyrase improves DNA transformation of E. coli cells with large recombinant plasmids Nucleic Acids Res., 1995, 23(16), 3353-3354.
[94] Takahashi, K.,Yamanaka, S., Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126(4), 663-676.
doi:10.1016/j.cell.2006.07.024.
[95] Jia, F., Wilson, K. D., Sun, N., Gupta, D. M. et al. (2010) A nonviral minicircle vector for deriving human iPS cells, Nature Meth. 2010, 7 (3), 197-199. doi:10.1038/nmeth.1426
[95] Rupprecht, S., Hagedorn, C., Seruggia, D., Magnusson, T. et al., Controlled removal of a nonviral episomal vector from transfected cells. Gene 2010, 466, 36–42.
[96] Galla, M., Will, E., Kraunus, J., Lei Chen, L., Baum, C., Retroviral Pseudotransduction Technique for Targeted Cell Manipulation. cell manipulation. Molecular Cell 2004, 16, 309–315.
[97] Yáñez-Muñoz, R. J., Balaggan, K. S., MacNeil, A., Howe, S., et al., Effective gene therapy with nonintegrating lentiviral vectors. Nat. Med. 2006, 12, 348-353. doi:10.1038/nm1365.
[98] S. Wagner, J. Mccracken1, S. Bruszies, J. Oswald et al., Minicircle episomal plasmids are stably transmitted in bovine fibroblasts. Transgenic Res. 2010, 19,152–153. DOI 10.1007/s11248-009-9326-4
[99] Manzini, S., Vargiolu, A., Stehle, I. M., Bacci, M. L. et al., Genetically modified pigs produced with a nonviral episomal vector. Proc. Natl. Acad. Sci. 2006 103, 17672-17677.
[100] Argyros, O., Wong, S. P., Constantinos, F., Tolmachov, O. et al., Development of S/MAR minicircles for enhanced and persistent transgene expression in the mouse liver J. Mol. Med. 89(5),515-29 . DOI: 10.1007/s00109-010-0713-3.
[102] Heinz, N., Broll, S., Schleef, M., Baum, C. et al. (2011) Filling a gap: S/MAR-based replicating minicircles, CliniBook - Nonviral Platform; Clinigene Network 2011, in press. http://www.clinigene.eu/assets/files/pdf/Annonce%20CliniGenebookBat2.pdf

53

54
FIGURE LEGENDS
Fig. 1: Chromatin domains, the smallest autoregulat ory expression units
A. Characterization of boundary elements (“constitutive S/MARs”/CSs) and of domain-internal
context-dependent elements (“facultative S/MARs”). CSs are characterized by their attachment
functions (hooked symbols) whereas facultative S/MARs mostly coincide with DNAse I
hypersensitive sites (HS). Both types can be identified in SIDD profiles indicating the energy
required for DNA strand separation at any location along the X-axis [kb]. Negative peaks (so called
“unpairing elements”, UEs) that reach a G value of zero (0 kcal/mol) are expected to open at a
given superhelical tension (usually σ = -0.05). Exemplified by the human interferon-ß (huIFN-ß-)
domain the structure of constitutive S/MARs (base-unpairing regions/BURs at the domain borders)
is associated with an oligomeric complex of scaffold-attachment factor A (SAF-A) and accessory
proteins (here: the histone acetyltransferase p300, green labels). Other imprints in the SIDD profile
mark the polyadenylation site (T), which is involved in transcriptional termination, and a breakpoint
junction (A1235), i.e. a site involved in genomic deletions [5].
B. Interpretation of these data in the framework of a popular chromatin domain model. Each
domain is delimited by extended, constitutive S/MAR elements, which, in case of the huIFN-ß
domain, extend over ~ 4 kb each, covering the ~2.2 kb EcoRI fragment “S/MAR E” (upstream) and
the ~ 3kb Hind III fragment “b” (downstream). The minicircle (MC), which is in the focus of this
review, can be considered as a simplistic domain model in which a single constitutive S/MAR
delimits a chromatin loop. Whereas these extended elements remain scaffold-associated
independent of the organism’s cell type, the association of restricted, “facultative” S/MARs is
governed by the cell- and stage-specific association of specialized factors, exemplified by YY1
(originally termed “nuclear matrix protein 1” / NMP-1) [6-8]. Facultative S/MARs can serve
enhancer-accessory functions rather than acting as an insulatos. Common to both classes is the
ease of strand separation, which causes spots of DNAse I hypersensitivity at the transition points
to regular B-type DNA. These “HS sites” proved to be preferred integration targets for retroviruses
[9] that are released from a pre-integration complex (PIC; see right-hand symbols characterizing
the delivery of a circular provirus-precursor).

55
Fig. 2: Components of the epigenetic code exemplifi ed by histone H3
H3 lysines (residues K4/-9/-14/-18/-27/-36 and -79), indicating the transcriptional status of
chromatin sites are predominantly located in its N-terminal tail domain. They are modified by either
methyltransferases (HMT, now named “KMT”*)) or by acetyl transferases (HAT/KAT) and –
deacetylases (HDACs). Activating modifications are indicated in green, italicized letters whereas
inactivating modifications are symbolized by regular letters in red. It is of note that the degree of
modification may induce different responses (K9- and K27 methylation reactions) whereas in other
cases activation or inactivation depends on the chromatin context. H3-modifications inducing an
inactive state occur in a stepwise fashion: Lysine residues K9 or K27 have to be de-acetylated in
order to permit mono-methylation, which is still compatible with the active states. Subsequently, a
methyltransferase induces the di- or even tri-methylated state. By recruiting the heterochromatin
protein HP1, gene silencing is initiated at H3K9me where it is caused by DNA methyltransferases
(KDM3/-6) to yield the triacetylated forms K9/-27. What follows is a complex, barely understood
multi-component interplay involving, for instance, H3K9me3, Suv39h and DNA methyltransferases.
*) The rationalized “KMT” nomenclature follows doi:10.1016/j.cell.2007.10.039.
Fig. 3: Chromosome-based expression principles: two current approaches and their
combination
In both branches of this representation, chromosome-based vectors comply with guidelines 1-3
(top). Both approaches (RMCE, left branch and minicircles/MCs, right branch) share the following
principles:
- eukaryotic expression units are delivered in the form of an autonomous chromatin domain,
characterized by either two bordering S/MAR elements (hooked boxes, cf. Fig. 1) or, in case of a
circular entity, by a single one (Fig. 1B);
- the application of a site-specific recombinase (Flp) inducing the recombination between two
identical Flp recombinase target (FRT-) sites (red half-arrows). If these sites are located on a
single DNA molecule, as in branch B, the result is excision - here cleavage of a “parental plasmid”

56
(PP) to yield two circular derivatives, the “miniplasmid” (MP, not shown) and the minicircle (MC);
further details will be explained in Fig 14. While the latter contains the eukaryotic vector parts, the
MP comprises prokaryotic and auxiliary sequences, which are lost as the cell divides;
- if two identical sets of different (“heterospecific”) FRTs are part of a genomic target and of a
donor plasmid as in branch A, the genomic cassette (Fmut -hygtk-F wt, here the classical setup F3-
hygtk-F) is cleanly replaced by a matching donor cassette (F3-GOI-F). This Flp-recombinase
mediated cassette exchange (Flp-RMCE) process avoids co-introduction of the prokaryotic vector
parts (dashed line), i.e. the cassette on the donor plasmid takes the place of the selection marker
cassette in a so called “flp-in” process.
“+/-“ symbolizes a fused selection gene, typically composed of a positive (hyg) and a negative
(HSV-tk) selection marker.
Fig. 4: Episomal persistence depends on the relativ e orientation between a transcription
unit and the S/MAR
A derivative of the parental S/MAR plasmid (PP) depicted in Fig. 3B was prepared such that the
GFP transcription unit was flanked by two recombination target sites in inverse orientation. During
a pulse of recombinase activity the transcription unit became inverted for 9 out of 15 clones; one
clone (EG78.1) contained constructs with both orientations (compare PCR analyses in the top
section with scheme in the insert).
Subsequent Southern analyses, performed after vector linearization by HindIII, indicated full
length episomes only in case of the functional original orientation (“o”) for which the transcript
extended into the S/MAR; the signal at 7.3kb reflects the size of an intact episome. In cases where
PCR analyses revealed inversion (“i”) of the transcription unit the blots were dominated by plasmid
loss and/or multiple integration events.
Fig. 5: An established episomal vector permits RMCE -based modifications in situ

57
Following the principles in branch “A” of Fig. 3, a cassette (here a luciferase expression unit
flanked by two different, “heterospecific” FRTs) can be exchanged by a cassette obeying the same
architectural principles. An authentic modification requires the prior establishment of the target
vector (here: a pEpi-type episome with a luciferase expression cassette), which is replaced by a
secondary cassette (here: egfp) provided by a donor plasmid. RMCE is driven by molar excess of
both the donor and the recombinase [1]. In the given example the exchange reaches 7%. RMCE
events can be enriched by FAC-sorting, which results in a collection of mostly stable clones (here:
70%). Decreased expression levels can be ascribed to the silencing effects of plasmid episomes
since the reaction is not reversed in the absence of Flp recombinase.
Fig. 6: S/MAR activities of eukaryotic base-unpairi ng regions (BURs) can be deduced from
SIDD profiles
DNA duplex destabilization properties of a DNA segment can be visualized after virtual cloning of
the element in question into a standard plasmid (here the PTZ-18R vector, a pUC derivative). This
corresponds to the context in which the experimental strengths of scaffold binding of many of the
S/MAR fragments have been assessed.
A - Sequence of pTZ_18R, which per se provides three unpairing elements (UEs) at the promoter
and the terminator of the ampicllin resistance gene, as well as at the phage f1 Ori; these signals
serve as internal standards. Besides the prototype SIDD profile [map position vs. G(x)], the
respective probability profile (map position versus p) is indicated. This representation is usually of
lower complexity (here: a single peak at the position of AmpT) indicating the site where strand
unpairing initiates.
B - The ~2 kb S/MAR insert providing the pEpi vector with replication potential, inserted into the
pTZ_18R backbone. Note the repetitive pattern of UEs, which obeys certain spacing criteria.
Obviously, the strongest S/MAR-UE (the “CUE” at position 800) surpasses the AmpT-associated
peak regarding its strand separation potential
C - S/MAR activity of the most strongly destabilized UE within the 2kb BUR in part B, determined
by an in vitro assay for the oligomer series M1 (monomer) to M4 (tetramer)

58
D - SIDD profile for the tetramer (M4) in the common pTZ_18R context. Note that the insert
competes with the AmpP- and f1-Ori-associated UEs but barely with the otherwise dominant “core-
unpairing element” (CUE ) derived from AmpT.
Fig. 7: Minimalization approaches - pMARS and Minic ircles
pMARS is a derivative with a functional, minimal S/MAR composed from four units of the most
effective UE (element M4 in Fig. 6D
A – Upper scheme anticipates the conversion of a pEpi-derived parental plasmid (circle) into two
derivatives, the minicircle (derived from the lower half) and the miniplasmid (from the upper half,
containing the selection gene and the prokaryotic Ori)
A – Bottom: FACS profile (eGFP-fluorescence versus size) for the pEpi-type vector containing the
full 2 kb S/MAR and a corresponding profile for the pMARS derivative, i.e. the tetrameric synthetic
sub-S/MAR “M4”.
B, C – FACsorting for the pEpi derivative (~2 kb S/MAR insert) and the minimized pMARs version
(4 x 155 bp “M4-“ insert)
Fig. 8: Establishment process of a 6.4 kb S/MAR pla smid and its 4.1 kb minicircle derivative,
both containing the same ~2 kb S/MAR sequence
Long-term expression of replicating episomes in CHO-K1 cells after 5-days (5 population
doublings, PDs) of establishment. After this period fluorescent cells were recovered by FAC-
sorting. 10 days (~10 PDs); after sorting 70% of fluorescent cells remained for the minicircle (MC),
but less than 10% for the parental plasmid (here: the pEpi-vector shown in Figure 7A). If, at this
time, the latter population is subjected to G418 selection, the remaining fluorescent subpopulation
becomes dominant reaching 60% after 50 PDs in which case, however, FISH analyses proved
integration events for 40 % of the cells (cf. Fig. 13). It is of note that presence of the HDACi
butyrate (“+butyrate”) for a 24 h period - followed by release from this treatment 18 h prior to
transfection of the S/MAR plasmid - significantly slows down the decay of the fluorescent cell

59
population. Due to the pFAR principle selection is neither possible nor required in case of the
minicircle as a substantial proportion (65% in this experiment) of cells maintains fluorescence
during the entire time interval.
Insert: Butyrate pre-treatment (“+B”) improves establishment rates also for the MC. Under the
given conditions [46] the population of fluorescent cells stabilized at 20% of eGFP-expressing cells
if transfection was performed without prior butyrate treatment (“-B”) but at 33-40% if transfection
followed a period in the presence of the HDACi. Controls demonstrated that the butyrate effect was
not due to synchronizing cells at the G0/G1 case (the value determined after 24-36 h in 5 mM
butyrate) but rater the consequence of a persistently altered epigenetic status initiated by, for
instance, histone hyperacetylation.
Fig. 9: Individual MC-clones established in the nuc lear architecture
MCs of 4.1 kb have been transferred, by lipofection, into CHO recipient cells. FACScan (eGFP
expression profiles) are shown for a cell population containing clones with 4–8 (average 5) copies
of the MC (“mixture”). Authentic single clones (M23, L2 and H11) were analyzed correspondingly
37 days post-transfection. Clones M23 and L2 are associated with symmetrical FACS profiles. The
Gaussian distribution found here is compatible with a unique (class of) association site(s).
Interestingly, this pattern resists freeze–thawing cycles (freezing at day 37 post-lipofection, storage
for 14 weeks, and FACScan 28 days after renewed continuous culture) as shown in the bottom
row. KWT, control from non-transfected CHOK1 cells.
The time course indicated on top (arrow) symbolizes the relevant stages during the MC life
cycle between preparation/transfer and withdrawal. The present study comprises the maintenance-
and expression stages following establishment.
Fig. 10: Co-transfer of minicircles accommodating h eavy-chain (H) and light-chain (L)
antibody genes supports survival of cells with an o ptimum expression ratio
Left: L- and H- genes are provided as transcription cassettes that are incorporated separate from
the fluorescence marker genes (rfp or egfp, respectively).

60
Right: FACS analysis after 20 generations – if red fluorescence is plotted versus green
fluorescence, surviving cells accumulate along a diagonal representing a constant expression ratio.
The upper-right quadrant contains 83% of all cells and served to recover high-producer strains (R.
K. Masters thesis, TU Braunschweig, 2008).
Fig. 11: AN S/MAR-internal auto-deletion process an d its functional relevance
A, B - After long-term cultivation, PCR revealed a reduced size (2.9 instead of 4.1 kb) for 2 out of
13 clones (exemplified by “M18” in B). According to sequence analyses these clones underwent
identical deletions (dashed red lines in A). The yellow star marks a premature transcription
termination site functioning in the context of the extended, 2kb S/MAR element but not after the
S/MAR-internal deletion as shown in section C.
C - Northern blot analyses were performed on authentic polyadenylated mRNA and hybridized with
a labeled eGFP-DNA probe. It is seen that transcript length depends exclusively on the extension
of the S/MAR, which is reduced (prematurely terminated) in case of the full 2 kb fragment (4.1 kb
MC and pEpi, lanes 1 and 3). In contrast, it is full-size for MC-derivative M18 (lane 2), its PP-
precursor (lane 4) and the plasmid-vector pMARS (lane 5).
D - SIDD profiles provide an explanation for premature termination since in these cases the
destabilization of the SV40 poly(A) sequence (marked by the left-hand circle) is reduced by
competition with the extended ~2kb BUR. This is depicted for pEPI (top SIDD profile) relative to the
M18 minicircle derivative (bottom profile [46]).
E - S/MAR size-reduction leads to improved expression (and long-term stability; [46]), a plausible
consequence of authentic transcript termination and –polyadenylation (red star-symbol in A).
Fig. 12: Southern blots for a number of expressing CHO-K1 clones
Clones have been pre-sorted, by FACS, 7 PD after DNA transfer. An aliquot of the clone mixture
(T) and single clones were cultured before high molecular weight DNA was harvested from 1×106
cells. Genomic DNA was cut with the MC-single cutter BstZ17I, subjected to Southern blot

61
analyses and visualized by a radioactive egfp-probe. The selection of single clones comprises
clones M23 and H11, which have been analyzed for expression and long-term persistence as
shown in Fig.11.
Fig. 13: Metaphase-FISH analyses
FISH-analyses were performed 55 PDs after transfection. Sections MC(1) and MC(2) demonstrate
the presence of individual minicircles while sections pEpi(1) and pEpi(2) give evidence of
integration events (intense doublets across the chromosome arms are found in 40% of the cells).
Note that parental plasmids and minicircles are presented at different magnification while the
overview (“wt”) corresponds to ~half the magnification chosen for the pEpi slides. See ref. [83] for
technical details.
Fig. 14: Multiple options for generating and elabor ating Minicircles (MCs) in situ, i.e. in the
recipient eukaryotic cell
The replication potential of this vector class is due to the properties of a scaffold/matrix attachment
region (S/MAR), which enables non-covalent anchoring the MC to chromosomes in the host cell.
MCs and active precursors (PPs) are the first true mammalian equivalents to yeast ARS plasmids.
- MCs can be generated from parental plasmids in situ using the Flp recombinase encoded
on the vector backbone (blue arrow). After recombining two identical target sites (FRTs, red half-
arrows) it is separated from the promoter becoming part of the MP.
- The MP has no replication capacity and, in contrast to the MC, is lost as the cell divides.
MCs consists only of eukaryotic sequences, among these the gene-of-interest (here: eGFP, green
arrow) and a 730 bp S/MAR (derivative “M18”).
- The additional inclusion of a heterospecific FRT (yellow half-arrow) permits subsequent
elaboration of the MC after its establishment in the nuclear architecture (principle in Fig. 5). In the
given example RMCE is applied to introduce the GOI, which removes and takes the place of a
positive/negative selection marker. Note that the functions of the respective coding regions (+/-

62
selection marker or GOI), individual promoters, IRES elements or fusion genes have not been
specified as these may vary from case to case. Optimal versions are about to enter large-scale
minicircle production routines as described in the text.

63
Table 1: Context-dependent and anti-silencing actio ns of an 800 bp S/MAR element (origin: 5´boundary of the huIFNβ gene domain). The element is used alone or in combination with the cHS4 element (an insulator from the chicken - beta globin gene cluster)
1 directional effect argues against enhancer mechanism; 2 H3 deacetylation precedes methylation in control. CpG methylation, does not establish but rather fixes inact. state
[29] -- (!) S/MAR alone superior. No 5´LTR CpG-meth. at day
180. 2 human mesench.
stem cells (ASCs) VSV-G IV(+/-c)HS4
[28] S/MAR-[cHS4]2 -3´LTR most efficient (elements
“oppositely oriented”) K562, HEK293, KB3.1
lines pSFb91
(SFFVxMESV) IV(+/-cHS4)
[26,
27] ++ Optimal expression in presence of SAR-cHS4-3´LTR human hematop.
stem cell line KG1a
and progenitor cells
Lentiviral
VSV-G IV (+/-cHS4)
[25] Retransplant mixed (+/- S/MAR) population.
S/MAR increases expression in all hematopoietic
lineages 2-9x for 6-12 mo Baboon marrow cells
prestimulated/
transplanted
Onco-RV vector
(Phoenix GALV) IV
[24] 4x long-term increase of transgene express./cell mobilized CD34+
HSCs MoMuLV/MSCV IV
[23] Prevents 5´LTR de-novo methylation (100%);
continued stable expression; level parallels copy
No. human T-cells
(CEMSS) MoMuLV IV
[22] HIV replication inhibited in CD4+ T but not
monocytes. Effect due to 2-10x expression of
RevM10 in T . No of expr. Cells and level ↑ human CD4+/CD8+ T;
primary MΦ MoMuLV IV
[21] ++1 Similar function within or ahead from 3´LTR.
Prevents silencing, not: IS-dependence; HIV
replication in CD4+ 100x more inhibited huPBL; CD4+/8+ T
(resting!) MoMuLV IV
Ref. Orien-
tation
dep.
Activity Cell type RV vector
system Insulator
[S/MAR IV /
cHS4]

64
A1235 T
HS-sites
huIFN-β
Mass- Binding
Nuclear Scaffold/Matrix : - Lamins- Matrins (ARBP/meCP2, Calmodulin, DNA-Polß, HAT, HDA, HMG, HAT, HDA, HMG 1/2,
Nucleolin, NuMa, PARP, SAF-A/hnRNP-U , SAF-B, Topo II, ssDNA-Binders);- Transcriptional modulators (SATB1) and specific transcription factors (YY1/2).
P
enh
MC
I I I
Nehlsen 2011, Fig. 1

65
Nehlsen2011, Fig. 2
me2me2meme2me2
me5)me3)me2)me1)
K79K36K27K14 / K18K9K4
ac12)ac11)ac10)
me3, me3me34)9)me38)me37)me36)
M1 A136
N-terminal tail
• de-acetylation →→
• tri-methylation →
• DNA-methylation
ac13)
1) KMT/Set1; 2) KMT1/KMT8/G9a; 3) KMT6; 4) KMT3; 5) KMT4; 6) KDM5; 7) KDM3; 8) KDM6; 9) KDM4; 10) KAT13; 11) KAT2/12; 12) KAT3/12; 13) KAT3A/B

66
II - Nonviral Episomes
Flp - excision
Guidelines and Strategies: 1 – No random integration (→ RMCE or episomal DNA)
2 – Never co-introduce prokaryotic (plasmid-) DNA
3 – No co-expressed gene / sel. marker besides the GOI
*
*
amp r
Flp
off eGFP
„Minicircle“ A - Establishment
B - Maintenance
*Epi-retroviral transfer
A B
Nehlsen 2011, Fig. 3
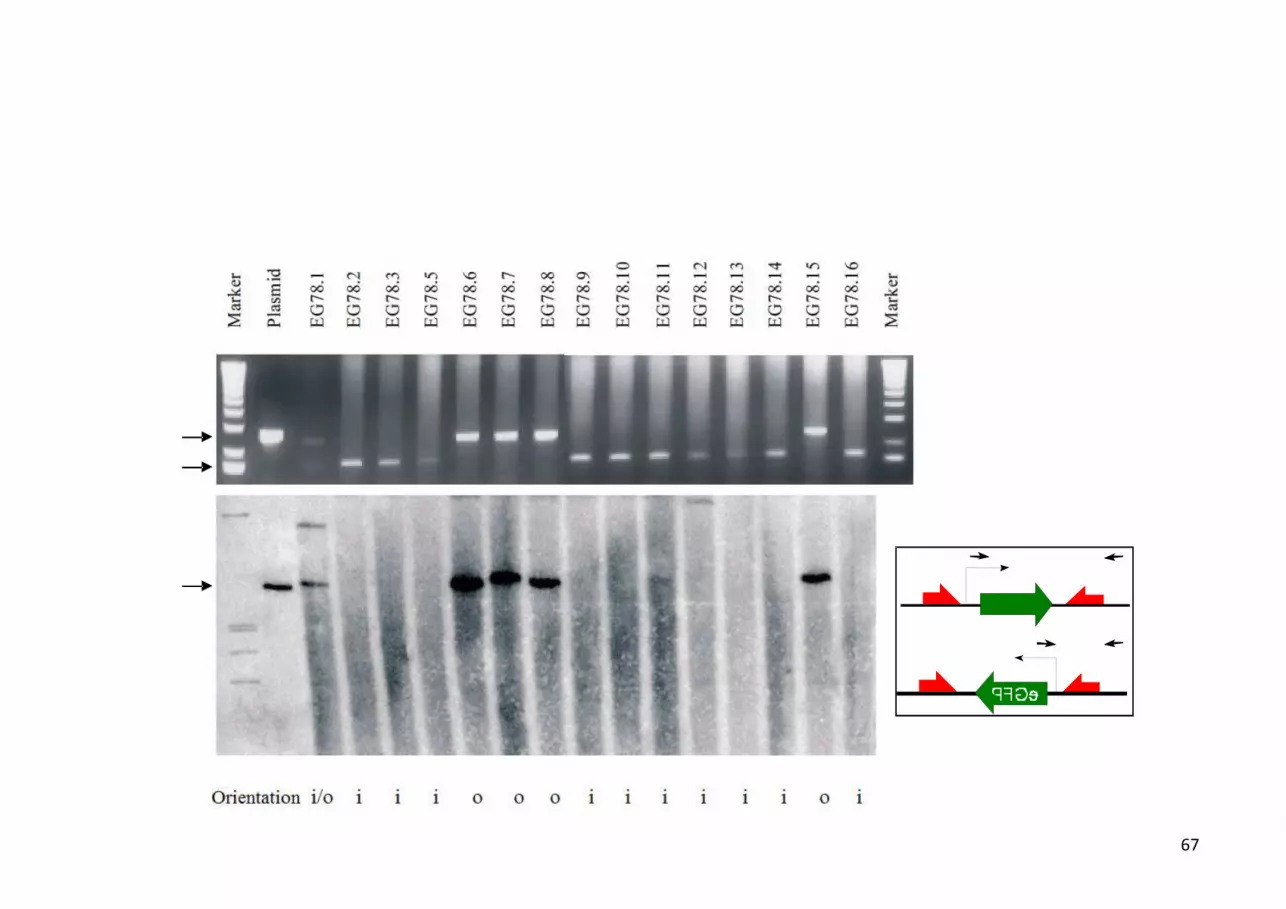
67

68

69
A B
C D
Nehlsen 2011, Fig. 6

70
pEpi-FGSARF
31%
A B
C pMARS
64%
Nehlsen 2011, Fig. 7

71
parental pl.
(minicircle)
PDs
+ butyrate.
Nehlsen 2011, Fig. 8

72
Preparation → Transfer → Establishment → Maintenance → Expression → Withdrawal
Nehlsen 2011, Fig. 9
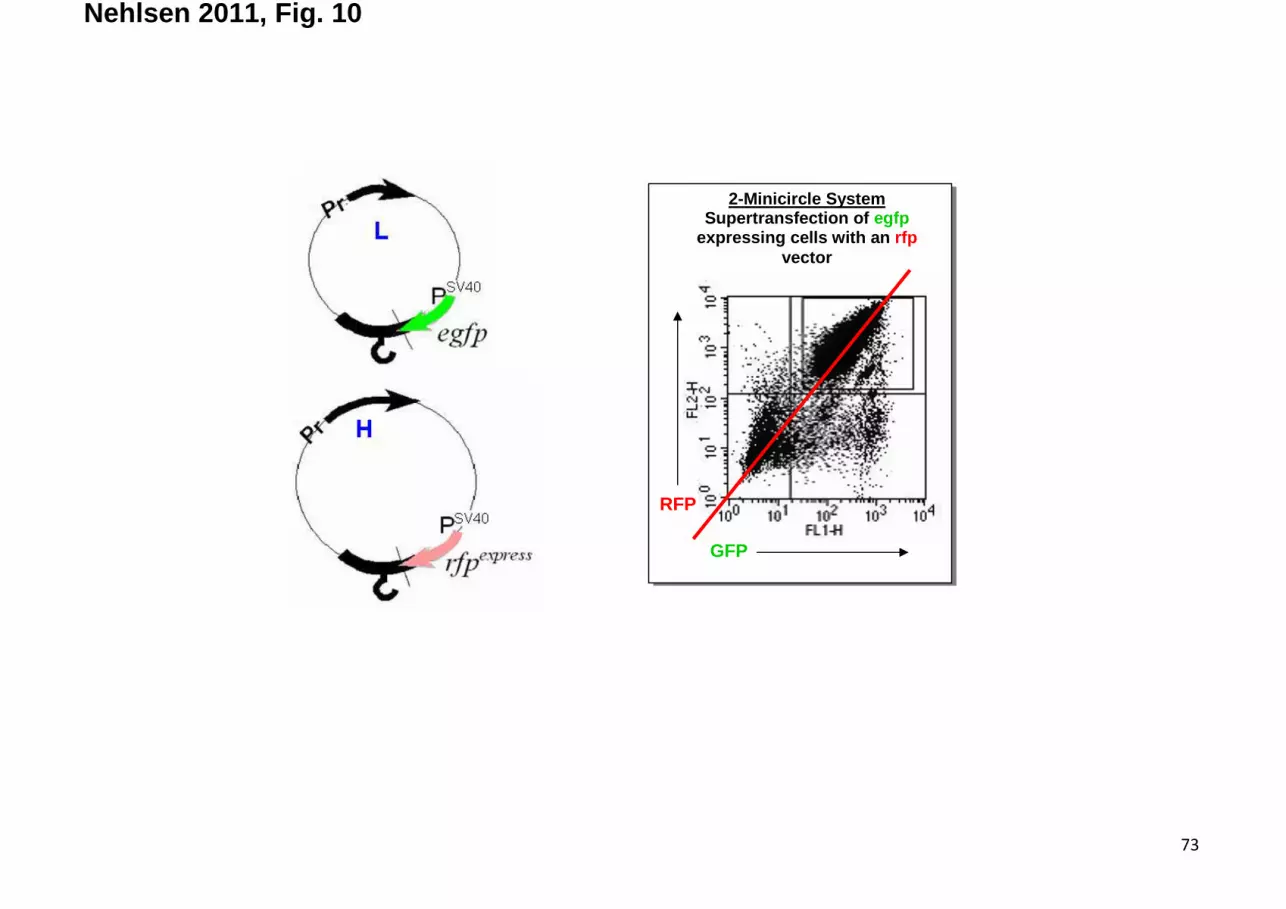
73
GFP
2-Minicircle System Supertransfection of egfp
expressing cells with an rfp vector
RFP
Nehlsen 2011, Fig. 10

74
PSV4
0
S/MAR
R
eGF
polyA
* *
∆∆∆∆1.3 kb1.3 kb1.3 kb1.3 kb
A
C
B
D E
pMARS pEPI
Nehlsen 2011, Fig. 11

75
Nehlsen 2011, Fig. 12

76
MC (2)
5µm
5µm
Nehlsen 2011, Fig. 13
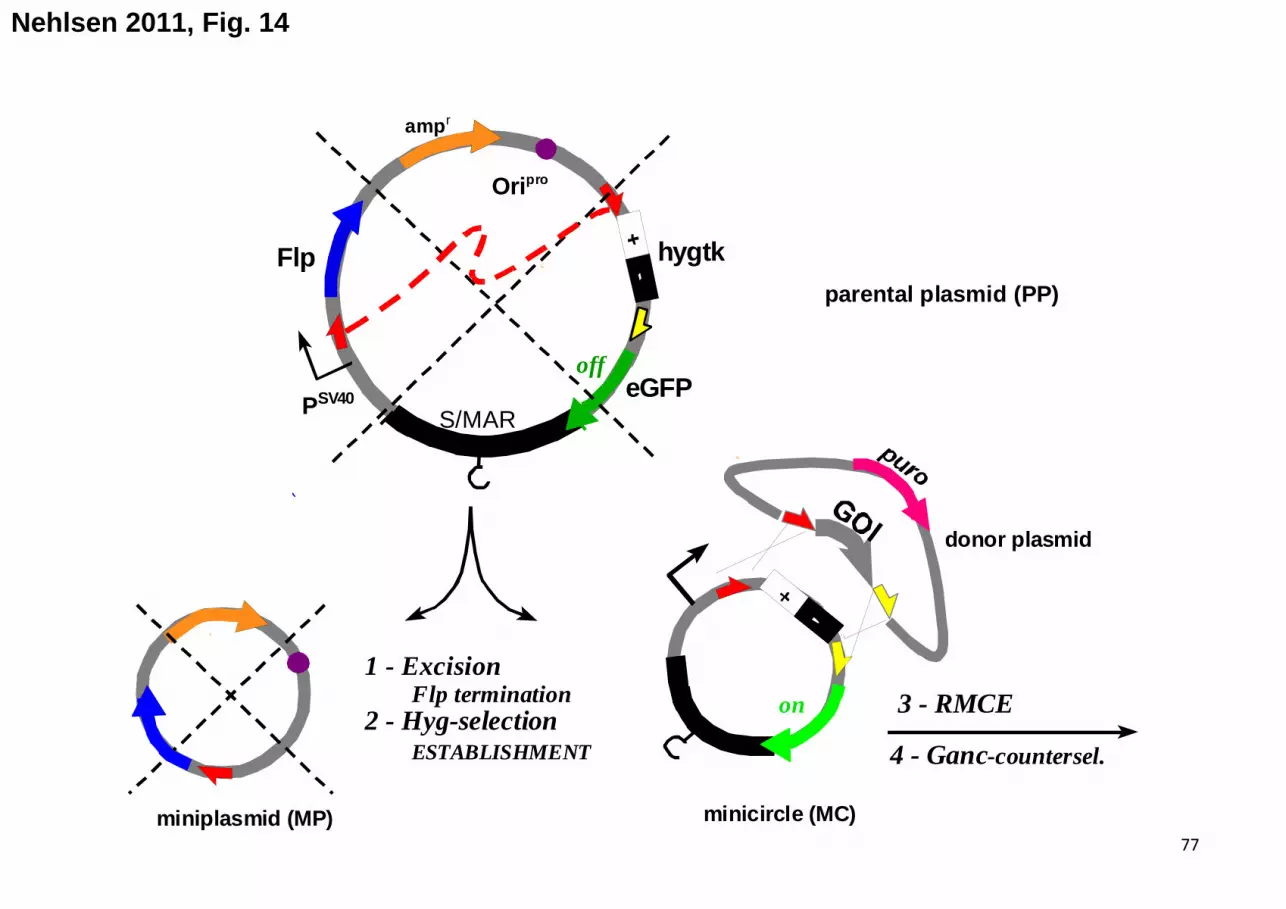
77
Nehlsen 2011, Fig. 14
Ori pro
amp r
minicircle (MC)miniplasmid (MP)
PSV40S/MAR
Flp
3 - RMCE1 - Excision ÷÷÷÷ Flp termination2 - Hyg-selection ÷÷÷÷ ESTABLISHMENT
eGFP
hygtk
donor plasmid
4 - Ganc-countersel.
off
on
parental plasmid (PP)





























