Embed Size (px)
Citation preview

Plant Syst. Evol. 223:109-123 (2000) Plant Systematics and Evolution
© Springer-Verlag 2000 Printed in Austria
Reproductive biology of Wahlenbergia (Campanulaceae) endemic to Robinson Crusoe Island (Chile)
G. J. Anderson 1, G. Bernardello 2, P. Lopez S. s, D. J. Crawford 4, and T. F. Stuessy 5
1Department of Ecology and Evolutionary Biology, Storrs, CT, USA 2Instituto Multidisciplinario de Biologia Vegetal, C6rdoba, Argentina 3Concepcidn, Chile 4Department of Evolution, Ecology and Organismal Biology, Ohio State University, Columbus, OH, USA Slnstitute of Higher Plant Systematics and Evolution, Institute of Botany, University of Vienna, Austria
Received August 31, 1999 Accepted March 6, 2000
Abstract. The reproductive biology of W. berteroi, W. fernandeziana, and a putative hybrid between 147. fernandeziana and W. grahamiae, endemic to Robinson Crusoe Island (Juan Fernfindez archi- pelago, Chile) was studied. Flowers are hermaph- roditic, protandrous, offer nectar, and exhibit secondary pollen presentation involving pollen collecting hairs on the style. These features imply allogamy and biotic pollination. However, male and female phases overlap and no effective polli- nators were observed. Experimental data indicate these taxa are self-compatible and facultatively autogamous, a conclusion also suggested by the pollen/ovule ratios. Selfing is accomplished when the stigmatic lobes reflex and touch the style, except for W. berteroi where they do not reflex completely. Autogamy is accomplished in the latter when pollen grains deposited on the inner surface of the corolla throat by the "pollen brush" are gathered by stigmatic lobes when shaken by wind. The degree of autogamy, and perhaps self-compatibil- ity, seems to be inconstant, as implied by the variable natural seed set (overall range 21-188 seeds per fruit). A mixed mating system - primarily outcrossing/entomophilous, but also autogamous - must have been present in the continental ancestors of these taxa. Autogamy promoting self-fertiliza- tion is important now - on an island with scarce
pollinators - and in the past - when the first founders arrived.
Key words: Angiosperms, Campanulaceae, Wak- lenbergia-Breeding system, pollination, pollen collecting hairs, autogamy, self-compatibility, nectar, island biology, Juan Fernfindez Islands.
The genus Wahlenbe~ia (subfam. Campanu- loideae) includes 267 species primarily distrib- uted in the southern hemisphere (Lammers 1996). It is ecologically extremely variable, occupying a variety of habitats from sea level t o 4,000 m in altitude. Several of the species in this genus are endemic to the Juan Fernfindez archipelago, located 667 km W of continental Chile. The archipelago consists of three islands: Robinson Crusoe (=Masa t ie r ra ) and Santa Clara to the east, and Alexander Selkirk (=Masafuera ) , 100 km further west. These islands are biogeographically interesting because of their relative geologic youth (5.8 mil- lion years for Santa Clara Island, ca. 4 million years for Robinson Crusoe Island, 1-2 million years for Alexander Selkirk Island; Stuessy et al. 1984) and the high level of endemism

110 G.J. Anderson et al.: Reproductive biology of Wahlenbergia
among vascular plants (ca. 62.5% of the flora; Stuessy et al. 1992, Marticorena et al. 1998).
Five species of Wahlenbergia, all endemic, are recognized (Lammers 1996): three on Robinson Crusoe Island (W. fernandeziana A. DC., W. grahamiae Hemsl., and W. berteroi Hook. & Arn., which also occurs on Santa Clara), and two on Alexander Selkirk Island (W. masafuerae (Phil.) Skottsb. and W. tube- rosa Hook. f.). In addition, a putative hybrid between W. fernandeziana and W. grahamiae was reported as well (Lammers 1996). There is a living specimen of this putative hybrid cultivated in the National Forestry Corpora- tion (CONAF) gardens in the single perma- nent human settlement (San Juan Bautista, Robinson Crusoe Island). Lammers (1996) provided evidence of monophyly of these species that are supported by data on chro- mosome number (Sanders et al. 1983, Spooner et al. 1987, Crawford et al. 1990, Sun et al. 1990), allozyme variation (Crawford et al. 1990), and morphology (Lammers 1996).
The Wahlenbergia species from the Juan Fernfindez are proposed to have arisen auto- chthonously, perhaps originally from a species similar to W. scopulicola Smith, from north- eastern Australia (Lammers 1996). Lammers (1996) proposed Robinson Crusoe Island as the probable site of first colonization and origin of the non-cormose clade (W. fernan- deziana and W. grahamiae). Several traits suggest that W. fernandeziana is the ancestral species on the islands (Crawford et al. 1990). Dispersal of the non-cormose clade to Alex- ander Selkirk Island was followed by evolution of the cormose clade (W. tuberosa and W. masafuerae). The establishment of W. ber- teroi (also cormose) on Robinson Crusoe and Santa Clara Islands is suggested to have occurred as a tertiary event (Lammers 1996). The non-cormose clade and W. tuberosa are found at higher inland elevations, while W. berteroi and W. masafuerae are restricted to lower coastal elevations (Lammers 1996).
Although vegetative propagation can occur in Wahlenbergia via stolons, suckers, and pieces of detached roots, reproductive events
are usually sexual (Lothian 1947). The litera- ture on the reproductive biology in the family is voluminous and dates back at least 250 years to Linnaeus (cf. Shetler 1979). However, for several genera including Wahlenbergia, there is little reproductive information. Beyond a few scattered data mainly on floral visitors (e.g. Thomson 1880, Mfiller 1883, Haviland 1884, Knuth 1909, Lothian 1947, Carolin 1960, Thulin 1975, Moeliono and Tuyn 1960), the only detailed study is of W. aIbomarginata Hook. from New Zealand (Lloyd and Yates 1982).
Carl Skottsberg (1928) summarized obser- vations on pollination and reproductive biol- ogy of the Juan Fern~mdez Wahlenbergia species as part of study of the flora of the islands. The only publication on reproductive biology since Skottsberg's are brief comments on W. berteroi (Crawford et al, 1990) and on the hybrid (Ricci and Eaton 1994, as W. larrainii).
The reproductive biology of the Campa- nulaceae as a whole is complex. Protandry and entomophily are widespread and are combined with an unusual mechanism of secondary pollen presentation via pollen collecting hairs on the terminal 1/3 of the style (cf. Gadella 1964, Shetler 1979, Leins and Erbar 1990, Erbar and Leins 1995, Nyman 1993a, Yeo 1993). This syndrome can function with self- pollination to facilitate autogamy or with effective pollinators to promote allogamy (cf. Shetler 1979, Nyman 1993b). Given these complexities, detailed study of specific taxa are needed to clarify the reproductive biology. Furthermore, the knowledge of the reproduc- tive biology, besides being fundamental for systematic and evolutionary studies (Anderson 1995), is essential to successful conservation efforts (Holsinger 1991, Bernardello etal. 1999), particularly of endangered species where there are very few populatiov/s - as is the case of these endemics.
In the course of three recent expeditions to Robinson Crusoe Island, we studied the reproductive biology of W. fernandeziana and W. berteroi in the field. We chose to study

G. J. Anderson et al.: Reproductive biology of Wahlenbergia 111
W. fernandeziana and W. berteroi in particular because they represent different clades of the genus in the archipelago that are accessible. Unfortunately, W. grahamiae is extremely rare and known for a few specimens (Lammers 1996), so we were not able to study it. Both species are shrubby with flowers arranged in showy terminal, cymose inflorescences. Wah- lenbergia berteroi grows on exposed rock outcrops, cliffs, and ledges near the sea (occa- sionally at elevations up to 300 m), whereas W. fernandeziana is more frequent at 100- 800 m above sea level, most often in crevices on steep or vertical rock escarpments (Lam- mers 1996). Populat ions of W. berteroi in particular are limited. We were able to locate only two populations, each of which included only 3-7 individuals. Although this species may be found elsewhere on the islands in small numbers, it certainly is not as common as W. fernandeziana, which has some populat ions with nearly 100 individuals. In addition, we analyzed one specimen of the hybrid. This individual, planted in the C O N A F garden from seeds harvested in the field, is presumably the same as that studied by Ricci and Eaton (1994).
We addressed the following questions: (1) What is the functional structure of the flowers? (2) Are there floral visitors? (3) Are the taxa self-compatible? (4) Are self-pollina- tion and autogamy possible, and if so, is self- or cross-pollination the dominant mode? And (5) what is the implication of these data for the future conservation of these endemic species?
Material and methods
Eight natural populations and one cultivated hybrid plant were studied on three field trips to Robinson Crusoe Island, Chile, during January 1991 and 1996, and December 1996/January 1997 (see Appendix I, vouchers are deposited at CONN). The taxonomic treatment of Lammers (1996) was followed in the delimitation of the taxa.
Floral anatomy: Flowers were fixed in 70% ethanol, dehydrated in an ethyl alcohol-xylol series, and embedded in Paraplast. Serial cross- and longi-
sections were cut at 10 btm, mounted, stained with safranin-fast green-hematoxylin and observed with a compound microscope. Photomicrographs were taken with Kodak T-Max film and drawings made with the help of a camera lucida.
Pollen/ovule (P/O) ratio and pollen viabili- ty: Pollen quantity was estimated using Anderson and Symon's (1989) modification of Lloyd's (1965) technique. Ovules were counted with a dissecting scope. Buds examined for P/O ratios were near anthesis, so pollen was mature but anthers had not dehisced. The mean P/O was generated from individual ratios of separate buds. A minimum of three buds per specimen was examined.
Pollen viability was estimated with percent stainability of 100 grains from each of 10 flowers (a minimum of three specimens except in the case of the hybrid) using aniline blue in lactophenol (Hauser and Morrison 1964). Observations on pollen viability were done in greenhouse plants grown at the University of Connecticut produced from seeds collected in the field.
Experimental crosses: Branches with tagged buds were bagged in the field with nylon mesh bags to exclude potential visitors. Flowers were emasculated in bud prior to anther dehiscence. Crosses were performed by applying pollen held among the style hairs (using the styles themselves as pollen applicators) to receptive stigmata. After 48 hours, flowers were collected and fixed in 70% ethanol. Natural autogamy was checked by bag- ging young tagged buds and collecting the unma- nipulated flowers four days after opening. Open pollinated control flowers were randomly collected. Tanglefoot ® was applied in some cases to the treated branches in an attempt to exclude ants from the experiments. To count seeds from open pollinated flowers, mature (but not yet dehiscing) fruits were selected and crushed in a Petri dish. Mature seeds were counted under a dissecting microscope.
Pollen tube growth: The gynoecia were dissect- ed from the flowers and were softened with 8 N NaOH for one hour at 60 °C in a water bath, rinsed, and stained in aniline blue -0.1 N K3PO4 for two hours (Martin 1959), then flattened in glycerin on a slide. Pollen tubes were examined under a epifluorescence microscope.
Floral visitors: Approximately 10 plants of W. berteroi and 30 of W. fernandeziana were observed in the field for more than 50 hours, during our three
112 G.J. Anderson et al.: Reproductive biology of Wahlenbergia
field trips. Periods of observation ranged from 20 to 40 minutes all during daylight hours (from 08:00 to 17:00 hr). Visitors were recorded and some were captured for identification. A similar procedure was followed with the single cultivated specimen of the hybrid.
Results
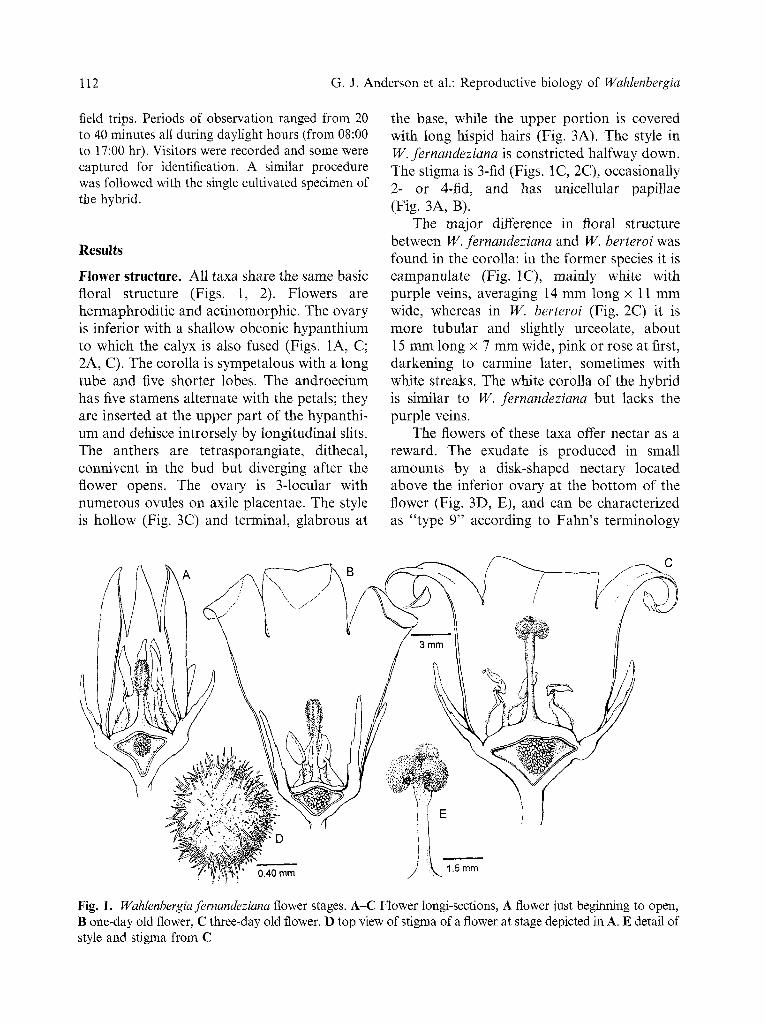
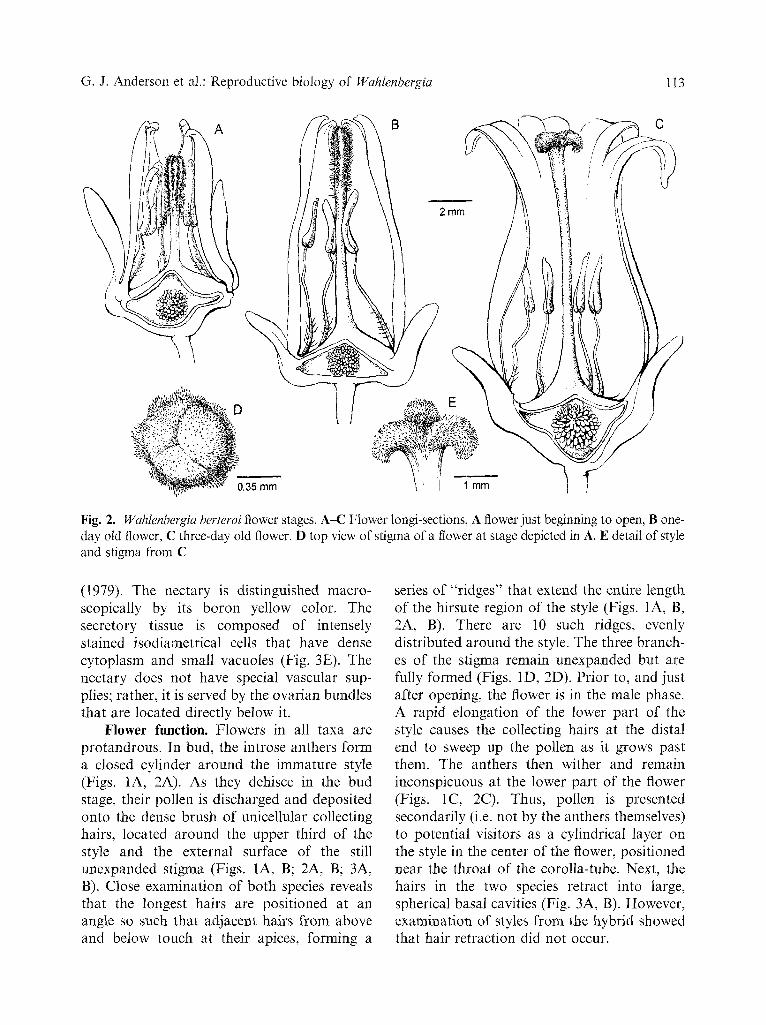
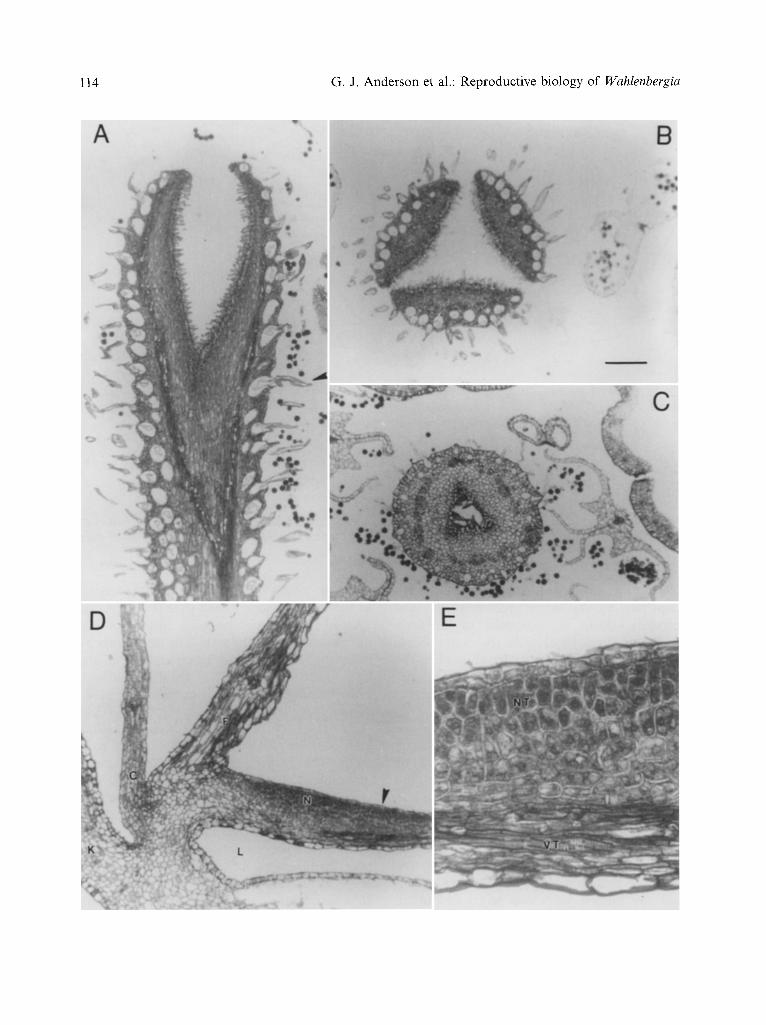
Flower structure. All taxa share the same basic floral structure (Figs. 1, 2). Flowers are hermaphroditic and actinomorphic. The ovary is inferior with a shallow obconic hypanthium to which the calyx is also fused (Figs. 1A, C; 2A, C). The corolla is sympetalous with a long tube and five shorter lobes. The androecium has five stamens alternate with the petals; they are inserted at the upper part of the hypanthi- um and dehisce introrsely by longitudinal slits. The anthers are tetrasporangiate, dithecal, connivent in the bud but diverging after the flower opens. The ovary is 3-1ocular with numerous ovules on axile placentae. The style is hollow (Fig. 3C) and terminal, glabrous at
the base, while the upper portion is covered with long hispid hairs (Fig. 3A). The style in W. fernandeziana is constricted halfway down. The stigma is 3-fid (Figs. 1C, 2C), occasionally 2- or 4-rid, and has unicellular papillae (Fig. 3A, B).
The major difference in floral structure between W. fernandeziana and W. berteroi was found in the corolla: in the former species it is campanulate (Fig. 1C), mainly white with purple veins, averaging 14 mm long x 11 mm wide, whereas in W. berteroi (Fig. 2C) it is more tubular and slightly urceolate, about 15 mm long x 7 mm wide, pink or rose at first, darkening to carmine later, sometimes with white streaks. The white corolla of the hybrid is similar to W. fernandeziana but lacks the purple veins.
The flowers of these taxa offer nectar as a reward. The exudate is produced in small amounts by a disk-shaped nectary located above the inferior ovary at the bot tom of the flower (Fig. 3D, E), and can be characterized as "type 9" according to Fahn's terminology
C
13
'~'" 0 .40 m m .5 mm
Fig. 1. Wahlenbergia fernandeziana flower stages. A-C Flower longi-sections, A flower just beginning to open, B one-day old flower, C three-day old flower. D top view of stigma of a flower at stage depicted in A. E detail of style and stigma from C
G. J. Anderson et al.: Reproductive biology of WahIenbergia 113
2~
" . : : 5:"-.~ : ". ~ ' : ": . ~
"~.~i ~ .i, S "~''~ ,! 0.35 rnrn
[
Fig. 2. Wahlenbergia berteroi flower stages. A-C Flower longi-sections, A flower just beginning to open, B one- day old flower, C three-day old flower. D top view of stigma of a flower at stage depicted in A. E detail of style and stigma from C
(1979). The nectary is distinguished macro- scopically by its boron yellow color. The secretory tissue is composed of intensely stained isodiametrical cells that have dense cytoplasm and small vacuoles (Fig. 3E). The nectary does not have special vascular sup- plies; rather, it is served by the ovarian bundles that are located directly below it.
Flower function. Flowers in all taxa are protandrous. In bud, the introse anthers form a closed cylinder around the immature style (Figs. IA, 2A). As they dehisce in the bud stage, their pollen is discharged and deposited onto the dense brush of unicellular collecting hairs, located around the upper third of the style and the external surface of the still unexpanded stigma (Figs. 1A, B; 2A, B; 3A, B). Close examination of both species reveals that the longest hairs are positioned at an angle so such that adjacent hairs from above and below touch at their apices, forming a
series of "ridges" that extend the entire length of the hirsute region of the style (Figs. 1A, B, 2A, B). There are 10 such ridges, evenly distributed around the style. The three branch- es of the stigma remain unexpanded but are fully formed (Figs. 1D, 2D). Prior to, and just after opening, the flower is in the male phase. A rapid elongation of the lower part of the style causes the collecting hairs at the distal end to sweep up the pollen as it grows past them. The anthers then wither and remain inconspicuous at the lower part of the flower (Figs. 1C, 2C). Thus, pollen is presented secondarily (i.e. not by the anthers themselves) to potential visitors as a cylindrical layer on the style in the center of the flower, positioned near the throat of the corolla-tube. Next, the hairs in the two species retract into large, spherical basal cavities (Fig. 3A, B). However, examination of styles from the hybrid showed that hair retraction did not occur.
114 G.J . Anderson et al.: Reproductive biology of Wahlenbergia
A B
G. J. Anderson et al.: Reproductive biology of Wahlenbergia 115
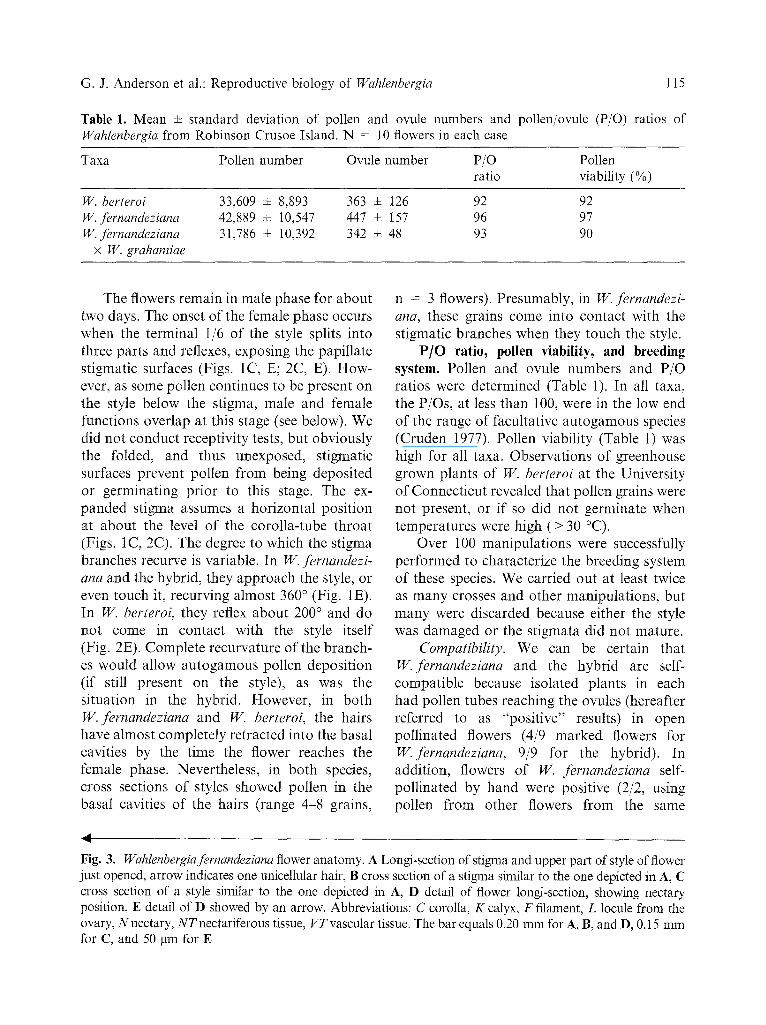
Table 1. Mean m standard deviation of pollen and ovule numbers and pollen/ovule (P/O) ratios of Wahlenbergia from Robinson Crusoe Island. N = 10 flowers in each case
Taxa Pollen number Ovule number P/O Pollen ratio viability (%)
W. berteroi 33,609 + 8,893 363 ± 126 92 92 FV. fernandeziana 42,889 ± 10,547 447 ± 157 96 97 W. fernandeziana 31,786 ± 10,392 342 ± 48 93 90
x W. grahamiae
The flowers remain in male phase for about two days. The onset of the female phase occurs when the terminal 1/6 of the style splits into three parts and reflexes, exposing the papillate stigmatic surfaces (Figs. 1C, E; 2C, E). How- ever, as some pollen continues to be present on the style below the stigma, male and female functions overlap at this stage (see below). We did not conduct receptivity tests, but obviously the folded, and thus unexposed, stigmatic surfaces prevent pollen f rom being deposited or germinat ing prior to this stage. The ex- panded stigma assumes a horizontal posi t ion at about the level of the corolla-tube throat (Figs. ] C, 2C). The degree to which the stigma branches recurve is variable. In W. fernandez# ana and the hybrid, they approach the style, or even touch it, recurving almost 360 ° (Fig. 1E). In W. berteroi, they reflex about 200 ° and do not come in contact with the style itself (Fig. 2E). Complete recurvature of the branch- es would allow au togamous pollen deposit ion (if still present on the style), as was the situation in the hybrid. However, in both W. fernandeziana and W. berteroi, the hairs have almost completely retracted into the basal cavities by the time the flower reaches the female phase. Nevertheless, in both species, cross sections of styles showed pollen in the basal cavities of the hairs (range 4-8 grains,
n = 3 flowers). Presumably, in W. fernandez# ana, these grains come into contact with the stigmatic branches when they touch the style.
P/O ratio, pollen viability, and breeding system. Pollen and ovule numbers and P/O ratios were determined (Table 1). In all taxa, the P/Os, at less than 100, were in the low end of the range of facultative au togamous species (Cruden 1977). Pollen viability (Table 1) was high for all taxa. Observations of greenhouse grown plants of W. berteroi at the University of Connect icut revealed that pollen grains were not present, or if so did not germinate when temperatures were high (> 30 °C).
Over 100 manipula t ions were successfully performed to characterize the breeding system of these species. We carried out at least twice as many crosses and other manipulat ions, but many were discarded because either the style was damaged or the st igmata did not mature.
Compatibility. We can be certain that W. fernandeziana and the hybrid are self- compatible because isolated plants in each had pollen tubes reaching the ovules (hereafter referred to as "posit ive" results) in open poll inated flowers (4/9 marked flowers for W. fernandeziana, 9/9 for the hybrid). In addition, flowers of W. fernandeziana self- pollinated by hand were positive (2/2, using pollen f rom other flowers f rom the same
Fig. 3. Wahlenbergiafernandeziana flower anatomy. A Longi-section of stigma and upper part of style of flower just opened, arrow indicates one unicellular hair, B cross section of a stigma similar to the one depicted in A, C cross section of a style similar to the one depicted in A, D detail of flower longi-section, showing nectary position, E detail of D showed by an arrow. Abbreviations: C corolla, K calyx, F filament, L locule from the ovary, N nectary, NT nectariferous tissue, VT vascular tissue. The bar equals 0.20 mm for A, B, and D, 0.15 mm for C, and 50 gm for E

116 G.J. Anderson et al.: Reproductive biology of Wahlenbergia
plant). The 4/7 positive results for tests of autogamy (which might be tests of geitonoga- my because introduced ants were able to get into even tightly sealed bags; of course, geito- nogamy tests self-compatibility), as well as the emasculated, open pollinated flowers tests (4/4 positive) are also evidence of self-compatibility in the hybrid. The latter are likely autogamous, or may be geitonogamous. They cannot be allogamous because there is only one plant of the hybrid known, and this single plant is separated by very long distances from any other members of the genus. Similarly, several of the emasculated, open pollinated flowers of W.fernandeziana with pollen tubes (7 of the 10 positive results), were from an isolated plant. Many manipulations were carried out with W. berteroi, but these suffered an inordinate amount of damage. However, two of the three flowers tested for autogamy (buds bagged and harvested subsequent to stigmata maturation) were positive in a setting with plants separated sufficiently that ants could not carry pollen between plants, and, as below, no other regular floral visitors were observed.
Allogamy. There is only plant known; thus, there possible. The successful tests
one living hybrid is no allogamy for open pollina-
tion on W. fernandeziana all involved an isolated plant, so these do not offer any insight to inter-plant pollen transport. However, the tests on emasculated, open pollinated flowers of this species also included some individuals from larger populations. In these, only 3 out of 29 flowers included pollen tubes, but, of course, any of these could have been the result of geitonogamy as well. More open pollinated flowers of W. berteroi were positive (14/20), and these involved plants in small populations (lighthouse = 3 plants, #3069; near the land- ing strip = 7 plants, #3095; Appendix 1), so this could be evidence of inter-plant transfer of pollen. As we discuss below, however, this is not likely given the virtual absence of floral visitors other than introduced ants. Thus, these few positive results might be a manifestation of autogamy, or possibly, geitonogamous trans- fer by ants.
Visitors. Although we observed that indi- viduals of all taxa bloom profusely and regularly produce nectar, visitation by insects is very rare. As there are few pollen-collecting visitors, pollen may remain in the older flowers in female phase, attached to the style hairs, as we observed in the hybrid. In more than 50 hours of observation, we detected one fly on W. berteroi, one moth on W. fernandeziana and the hybrid, and two halictid bees on flowers of the cultivated hybrid (Table 2). Note that the only places we observed the presumably recently introduced bees (Engel, in press) was near the only settlement, village of San Juan Bautista. Two of the three captured bees had pollen on their bodies, indicating they could act as effective pollinators - except for their lack of abundance. And, our focus here is on the reproductive biology of the native plants and pollinators; thus, the bee and ants (see below) are considered irrelevant.
Ants were also observed to visit the flowers of the hybrid (at the CONAF station), of one population of W. berteroi, and two of W. fer- nandeziana (Table 2). The ants were identified by E. O. Wilson as Linepithema humile (Mayr), native to Argentina, but common world-wide invasives. Given their introduced status, they aie not considered of consequence in the evolution of the reproductive system. Howev- er, like the introduced halictid bee, they could be of consequence for conservation - i.e., positively or negatively, affecting sexual repro- duction. The comments that follow are rele- vant in that context.
Thus, overall, we have no good evidence for any native biotic pollinators, but the nature of pollen (sticky), the secondary pollen pre- sentation (the stylar brush), and the shape of the flowers (bells), make abiotic (wind) polli- nation unlikely as well.
Seed set. The fruit is a capsule that bears light small, smooth-coated seeds (0.7-0.9 mm long) probably wind dispersed. If we are correct in assuming facultative autogamy, the data on seed numbers indicate that the level of autogamy is variable. For instance, in W. berteroi, we obtained the following seed

G. J. Anderson et al.: Reproductive biology of Wahlenbergia
Table 2. Insect floral visitors to Wahlenbergia on Robinson Crusoe Island
117
Plant taxa Location Date Order Family Pollen presence
W. berteroi Lighthouse 1 / 1 7 / 9 6 Hymenoptera Formicidae 1 No Landing strip 12/31/96 Diptera Undetermined Not collected Lighthouse 1 / 2 / 9 7 Hymenoptera Formicidae Not collected
W. femandeziana Lighthouse 1 / 1 3 / 9 6 Hymenoptera Formicidae Not collected Lighthouse 1 / 1 7 / 9 6 Lepidoptera Pyralidae No Lighthouse 1 / 1 7 / 9 6 Hymenoptera Formicidae~ No Lighthouse 1 / 2 / 9 7 Hymenoptera Formicidae Not collected Pirfimide 1 / 3 / 9 7 Hymenoptera Formicidae Not collected
W. fernandeziana CONAF 1/12/96 Hymenoptera Halictidae 2 No x W. grahamiae CONAF 1/20/96 Hymenoptera Halictidae 2 Yes (many)
CONAF 1/23/96 Hymenoptera Halictidae 2 Yes (few grains) CONAF 1/24/96 Lepidoptera Moth Not collected CONAF 1/10/97 Hymenoptera Formicidae Not collected
~ Identified as same species under the genus Lassioglossum (subgen. Dialictus) 2 Identified as same species: Linepithema humile
set per fruit (n = 5) in open pollinated flowers: 188, 90, 29, 27, 22, as well as in the hybrid: 125, 54, 44, 28, and 21, and in W.fernandeziana: 98, 74, 51, 42, and 23.
Discussion
The basic floral morphology of the three taxa we studied is concordant with previous obser- vations (Lammers 1996), and is shared gener- ally by Wahlenbergia (e.g. Lothian 1947, Moeliono and Tuyn 1960, Thulin 1975, Ros- atti 1986). Interestingly, the association be- tween corolla type and style constriction observed by Smith (1992) in some Australian species is manifest in the Juan Fernfindez species too. That is, styles from narrower corollas are not constricted (e.g.W. berteroi), whereas those from broader corollas (e.g. IV. fernandeziana) are constricted. The loca- tion of the nectary above the ovary is typical of several other families with inferior ovaries as well (e.g., Myrtaceae and Cucurbitaceae, Fahn 1979). This nectary position has been cited for Campanula (Fahn 1979), but this is the first report for Wahlenbergia. Protandry is wide- spread in the family (e.g. Shetler 1979, Nyman
1992, Inoue et al. 1996) and has been reported for other Wahlenbergia as well (e.g. Haviland 1884, Lothian 1947, Moeliono and Tuyn 1960, Thulin 1975, Lloyd and Yates 1982), but is not as pronounced in the Fernandezian species, because male and female phases overlap.
The pollen collecting hairs in Wahlenbergia are typical of the Campanulaceae (cf. Uphof o 1962; Shetler 1979, 1982; Leins and Erbar 1990; Nyman 1993a, b). This unusual mecha- nism of secondary pollen presentation is anal- ogous to the well-known type in Campanula (Carolin 1960; Gadella 1964; Shetler 1979, 1982; Erbar and Leins 1989; Yeo 1993; Nyman 1993a, b), first described by Sprengel in 1793 and observed by Lanais in 1738 (Shetler 1979). These hairs not only serve as pollen collectors, but also in Campanula have been shown to serve as a mechanism controlling stigma mat- uration and promoting cross-pollination (Ny- man 1993b). The length of the male phase is significantly shortened when the hairs are stimulated by contact (Richardson and Ste- phenson 1989, Nyman 1993b). Such stimula- tion can cause the hairs to retract (Shetler 1979, 1982) making the pollen unavailable for transfer, be it by subsequent pollinators or by

118 G.J. Anderson et al.: Reproductive biology of Wahlenbergia
selfing via the recurvature of the style (Nyman 1993b). Different species display variation in this mechanism, perhaps reflecting differences in mating systems. Hair retraction in some self- incompatible species is fixed, thereby dramat- ically reducing transfer of self-pollen to the stigma. In self-compatible species, hairs may not retract unless visited, acting as some sort of fertility insurance, promoting autogamous pollination if allogamy fails (Nyman 1993a). Hair retraction was observed in both W. fer- nandeziana and W. berteroi and thus appears not to be dependent on contact, as it occurred even without floral visitors. Of particular interest is that pollen production can be almost nil, especially when temperatures are very high (greenhouse experiments), and yet hair retrac- tion still occurs. This may indicate that in these species, retraction is also not associated with pollen production. Hairs on mature styles of the hybrid did not retract; this could reflect the syndrome of the very rare W. grahamiae, the other parent or perhaps be a manifestation of hybrid breakdown.
Wahlenbergia fernandeziana, W. berteroi, and the studied hybrid, are self-compatible. This conclusion agrees with that previously reported for W. berteroi and the hybrid (Craw- ford et al. 1990, Ricci and Eaton 1994, respec- tively), as well as for some other species in the genus (Lothian 1947, Moeliono and Tuyn 1960, Lloyd and Yates 1982). Given the species are self-compatible, autogamy is possible, as reported for several other Wahlenbergia (Mtil- ler 1883, Knuth 1909, Lothian 1947, Moeliono and Tuyn 1960, Thulin 1975). Our data support facultative autogamy. Thus, the variable seed set numbers could indicate variance in the degree of self-compatibility, or perhaps they may simply reflect differences in plant vigor, water availability, etc. Greenhouse experiments to test the duration of pollen viability in W. berteroi proved unsuccessful, as pollen production was low and plants failed to produce mature seed. This is likely due to unfavorable environmental conditions, given that in other taxa, high temperatures cause aberrant meiosis (e.g. Impatiens balsamina L., Nakamura 1935).
Most features of the breeding system - protandry, nectar as reward, secondary pollen presentation - reduce self-pollination or sug- gest allogamy and biotic pollinators. In spite of these many features, autogamy prevails in several Wahlenbergia species (Lothian 1947, Thulin 1975, Petterson 1997), as well as W. fernandeziana and the hybrid. Autogamy is effected by self-compatibility, partial over- lapping of male and female phases, and recurvature of stigmatic branches to the style. In the hybrid, as the hairs do not retract, pollen is readily available for self-pollination. And in W. fernandeziana, although the hairs retract, the pollen grains that stay in the hair bases may come in contact with the stigma when it recurves, allowing selfing.
In W. berteroi, although the same degree of stigmatic recurvature does not take place and this type of autogamy is not possible, most of the flowers observed have pollen on the stigmatic surfaces. We suggest an unusual method of autogamy, as follows. Pollen grains adhere to the corolla-tube throat, usually at the level where the mature stigma is located. As anthers dehisce in the bud stage, the corolla throat closely surrounds the anther cylinder which in turn, surrounds the style (Fig. 2B). Following anther dehiscence, both corolla and style elongate, and some pollen is deposited on the inner surface of the narrow urceolate corolla-throat (Fig. 2C). The ring of pollen in the throat is easy to see because its white color stands out against the pink corolla. As pollen stays viable throughout the receptive stage of the stigma (as shown in our bagging experi- ments), transfer from corolla to stigmata could occur when flowers are shaken by the ever- present wind, particularly on the exposed sea cliffs where W. berteroi grows. This mechanism circumvents the likely fixed hair retraction on the styles of this species as well. This autoga- mous wind-aided mechanism of pollination, although unusual, appears effective in this case. A similar situation was reported in Iris versicolor L. from Kent Island, New Bruns- wick, by Zink and Wheelright (1997). They observed that facultative self-pollination was

G. J. Anderson et al.: Reproductive biology of Wahlenbergia 119
promoted by wind that forced reflexed stigmas to brush against petals of the same flowers where pollen had settled.
Even though the reproductive mechanisms of W. fernandeziana and W. berteroi are close, phylogenetic analysis of these species indicates that W. fernandeziana is ancestral to W. ber- teroi (Lammers 1996). The narrowing of the floral tube in W. berteroi may have evolved to promote selfing in the presence of a non- recurving stigma that shows less recurvature. Given the virtual lack of dedicated pollinating insects (Skottsberg 1928, Bernardello et al. 1999, Anderson et al. in press), and in partic- ular, specialists that would respond to color differences, the darker purple variants of this species must be explained as co-variants linked to some other feature.
In general, Wahlenbergia has been consid- ered insect-pollinated (Thulin 1975, Lloyd and Yates 1982, Petterson 1997). The nectar chem- ical composition of all the taxa studied here is hexose dominant (Bernardello et al. in press). Such nectar is often associated with exposed nectaries and visits by short-tongued bees and flies (Baker and Baker 1983). However, that is not possible on the Juan Fernfindez where insect pollinators are extremely uncommon (Skottsberg 1928, Bernardello et al. 1999, Anderson et al. in press). The paradox of the majority of floral features suited to entomoph- fly, and a lack of native pollinating insects arises frequently in the Juan Fern/mdez flora (Ber- nardello et al. 1999, Anderson et al. in press). Such a syndrome may well characterize many other archipelagos as well where the insect fauna is limited (Bernardello and Anderson in prep.). Hummingbirds might visit the flowers for nectar (Colwell 1989), but they have never been recorded on any of these species, and the nectar quantity available is small as well.
A primarily outcrossing entomophilous system may well have been present in the continental ancestors of these evolutionary young island species (Thulin 1975, Lloyd and Yates 1982, Petterson 1997) - perhaps in addition to self-compatibility and facultative autogamy. These reproductive options, pro-
moting self-fertilization, would obviously be important to the first Wahlenbergia colonists on the islands, but remain so today - on islands with virtually no insect pollinators. Data on genetic diversity of three populations of W. berteroi showed no allozyme variation, as is the situation in other highly selfing species on the archipelago (Crawford et al. 1990). Of course, there are also few plants of this species still extant. The comparatively much more abun- dant W. fernandeziana (Crawford et al. 1990) exhibited relatively high enzyme diversity at both the species and population levels. This fact is difficult to explain from the available data that indicate a primarily autogamous system. A similar situation was observed in Dendroseris litoralis Skottsb. (Asteraceae) that has quite high allozyme diversity (Crawford et al. 1998) despite the fact that it is very rare (Stuessy et al. 1998), is self-compatible, and facultatively aut- ogamous (unpubl. data).
The degree of autogamy and allogamy can vary greatly within a single plant species, depending of factors such as pollinator abun- dance, degree of dichogamy, or duration of pollen viability (e.g. Horowitz and Harding 1972, Schaal and Leverich 1980, Wyatt 1984, Nyman 1991). In some species, evolution has encouraged a strategy of mixed mating sys- tems, combining abilities for outcrossing as well as selfing (Richards 1997). The stability of mixed mating systems can promote fitness benefits in offspring generated through out- crossing as well as selfing (Holsinger 1992). By the time of arrival on this island, Wahlenbergia ancestors probably had a mixed mating sys- tem, but the absence of pollinators may have forced the autogamous system present today.
Conservation. The seed set produced by these taxa is quite variable, with some fruits bearing comparatively few seeds, implying that pollination may be inefficient to some extent and/or that inbreeding depression is being expressed in terms of limiting seed set. This is understandable on the basis of a breeding system with many allogamous features that is now largely autogamous. Furthermore, we have not seen seedlings in the wild or individ-

120 G.J. Anderson et al.: Reproductive biology of WahIenbergia
uals propagating vegetatively. The marginally successful breeding system and the permanent- ly windy and cool to cold eroded habitat where these plants live makes survival difficult. In addition, a new pressure has been added: we noted that introduced locusts virtually de- stroyed a population of W. berteroi (#3095, near the landing strip), eating stems, leaves, fruits, and seeds.
Both environmental and reproductive con- straints seem to be reducing the fitness of these taxa. Wahlenbergia grahamiae is already ex- tremely rare on the island (Lammers 1996), the hybrid is known from only one cultivated plant, and there are very few, and then small, known populations of W. berteroi. From the genetic point of view, there are differences between the species as well: the variation of W. berteroi is zero, whereas that of W. fernan- deziana is comparatively higher (Crawford et al. 1990).
In order to conserve these plants, programs must involve a combination of environmental and reproductive measures. The populations of introduced insects and weeds must be controlled. And, experimentally produced all- ogamous seeds would enhance diversity in a restoration program. However, one recent
insect introduction might be valuable. Our field work, and that of others (pers. comm. of H. Toro) verify that there is now a known bee species on Robinson Crusoe Island (Anderson and Bernardello in prep.). This small halictid bee could prove to be of benefit for promotion of allogamy with species like those of Wahlen- bergia. And, given the lack of any significant native insect fauna, it does not pose a danger - at least to Wahlenbergia (it could compete for nectar with the two hummingbird species).
We thank the National Science Foundation, University of Connecticut Research Foundation and the Department of Ecology and Evolutionary Biology, and CONICET, CONICOR and SECYT- Universidad Nacional de C6rdoba (Argentina) for support, Mary J. Spring for preparing figures 1 and 2, Jane O'Donnell for moth and fly identification, Edward O. Wilson for the ant identification, Michael S. Engel for the bee identification, Thomas G. Lammers for helpful comments on the manu- script, Richard Jensen for statistical comments, Maryke Schlehofer for helping in many ways, and Adriana P6rez for slide preparation. The CONAF (Corporaci6n Nacional Forestal, Chile) is sincerely acknowledged for help, specially from Iv/in Leiva (Head). Assistance was provided by the Servicio Meteoroldgico from Robinson Crusoe Island.
Appendix I Wahlenbergia specimens studied. All collected on Robinson Crusoe Island, Ai;chipelago Juan Fernfindez, Chile, by G. J. Anderson et al. Abbreviations: M morphology, F fluorescence microscopy, P/O pollen/ ovule count, PV pollen viability, A anatomy, N nectar. Elevation in meters above sea level
Species Location Elevation Collection Date Study number
W. berteroi Lighthouse 10 3069 1/28/96 F, P/O, M Near landing
strip 0 3095 12/31/96 F, S, A Lighthouse 10 3096, 4006, 1/1/97 F, P/O
4007
2026 4005 4014
W. fernandeziana Villagra 450 Lighthouse 10 Corrales de 210
Molina Villagra 385 Villagra 450 Villagra 415 Villagra 380
4025 4026, 4033 4034
4027
1/14/96 P/O, M, A 1/2/97 F, P/O 1/3/97 F, P/O, PV, A
1/5/97 V 1/5/97 F,P/O 1/5/97 F 1/5/97 F

G. J. Anderson et al.: Reproductive biology of Wahlenbergia
Appendix (Continued)
121
Species Location Elevation Collection Date Study number
W. fernandeziana CONAF 60 2008 1/12/96 x W. grahamiae
CONAF 60 3097, 1/2/97 4002, 4003, 4004
P/O, PV, M, A
F, P/O, S
References
Anderson G. J. (1995) Systematics and reproduc- tive biology. In: Hoch P. C., Stephenson A. G. (eds.) Experimental and molecular approaches to plant biosystematics, Monogr. Syst. Bot. 53, pp. 263-272. Missouri Botanical Garden, St. Louis.
Anderson G. J., Bernardello G., Lopez S. P., Stuessy T. F., Crawford D. J. (2000) Dioecy and wind pollination in Pernettya rigida (Ericaceae) of the Juan Fernfindez Islands. Bot. J. Linn. Soc. (in press).
Anderson G. J., Symon D. E. (1989) Functional dioecy and andromonoecy in Solanum. Evolu- tion 43: 204-219.
Baker H. G., Baker I. (1983) Floral nectar sugar constituents in relation to pollinator type. In: Jones C. E., Little R. J. (eds.) Handbook of experimental pollination biology. Van Nostrand Reinhold, New York, pp. 117-141.
Bernardello G., Anderson G. J., Lopez S. P., Cleland M. A., Stuessy T. F., Crawford D. J. (1999) Reproductive biology of Lactoris fernan- deziana (Lactoridaceae). Amer. J. Bot. 86: 829- 840.
Bernardello G., Galetto L., Anderson G. J. (2000) Floral nectary structure and nectar chemical composition of some species from Robinson Crusoe Island (Chile). Canad. J. Bot. (in press).
Carolin R. C. (1960) The structures involved in the presentation of pollen to visiting insects in the order Campanales. Proc. Linn. Soc. NSW 85: 197-207.
Colwell R. K. (1989) Hummingbirds of the Juan Fernfindez Islands: natural history, evolution and population status. Ibis 131: 548-566.
Crawford D. J., Sang T., Stuessy T. F., Kim S-C., Silva O. M. (1998) Dendroseris (Asteraceae: Lactuceae) and Robinsonia (Asteraceae: Senecio- neae) on the Juan Fernandez Islands: similarities
and differences in biology and phylogeny. In: Stuessy T. F., Ono M. (eds.) Evolution and speciation of island plants. Cambridge Univer- sity Press, Cambridge, pp. 97-119.
Crawford D. J., Stuessy T. F., Lammers T. G., Silva O. M., Pacheco P. (1990) Allozyme vari- ation and evolutionary relationships among three species of Wahlenbergia (Campanulaceae) in the Juan Fernandez Islands. Bot. Gaz. 151: 119-124.
Cruden R. W. (1977) Pollen ovule ratios: a conservative indicator of breeding systems in flowering plants. Evolution 31: 32-46.
Engel M. S. (2000) A new Lassioglossum from the Juan Fernfindez Islands (Hymenoptera: Halicti- dae). Revista Chilena Entomol. (in press).
Erbar C., Leins P. (1989) On the early floral development and the mechanisms of secondary pollen presentation in Campanula, Jasione and Lobelia. Bot. Jahr. Syst. 11 I: 29-55.
Erbar C., Leins P. (1995) Portioned pollen release and the syndromes of secondary pollen presen- tation in the Campanulales-Asterales-complex. Flora 190: 323-338.
Fahn A. (1979) Secretory tissues in plants. Aca- demic Press, London.
Gadella T. W. J. (1964) Some cytological observa- tions in the genus Campanula. II. Proc. Royal Netherl. Acad. Sci. Ser. C 66: 270-283.
Hauser E. J. P., Morrison J. P. (1964) The cytochemical reduction of nitro blue tetrazolium as an index of pollen viability. Amer. J. Bot. 51: 748-752.
Haviland E. (1884) Occasional notes on plants indigenous in the immediate neighborhood of Sydney. Proc. Linn. Soc. NSW 9: 1171-1174.
Holsinger K. (1991) Conservation of genetic diver- sity in rare and endangered plants. In: Dudley E. C. (ed.) The unity of evolutionary biology: the proceedings of the fourth International Congress

122 G.J . Anderson et al.: Reproductive biology of Wahlenbergia
of systematic and evolutionary biology. Diosco- rides Press, Portland, pp. 626-633.
Holsinger K. (1992) Ecological models of plant mating systems and the evolutionary stability of mixed mating systems. In: Wyatt R. (ed.) Ecol- ogy and evolution of plant reproduction. Chap- man and Hall, London New York, pp. 169-191.
Horowitz A., Harding J. (1972) Genetics of Lupi- nus. V. Intraspecific variability for reproductive traits in Lupinus nanus. Bot. Gaz. 133: 155-165.
Inoue K., Maki M., Masuda M. (1996) Evolution of Campanula flowers in relation to insect pollinators on islands. In: Lloyd D. G., Barrett S. C. H. (eds.) Floral biology: studies on floral evolution in animal-pollinated plants. Chapman and Hall, New York, pp. 377-400.
Knuth P. (1909) Handbook of flower pollination, 3. University Press, Oxford.
Lammers T. G. (1996) Phylogeny, biogeography, and systematics of the Wahlenbergia fernandez# ana complex (Campanulaceae: Campanuloi- deae). Syst. Bot. 21: 397-415.
Leins P., Erbar C. (1990) On the mechanisms of secondary pollen presentation in the Campanu- lales-Asterales-complex. Bot. Acta 103: 87-92.
Lloyd D. G. (1965) Evolution of self-compatibility and racial differentiation in Leavenworthia (Cru- ciferae). Contr. Gray Herb. Harvard Univ. 195: 1-134.
Lloyd D. G., Yates J. M. A. (1982) Intrasexual selection and the segregation of pollen and stigmas in hermaphrodite plants, exemplified by Wahlenbergia albomarginata (Campanulaceae). Evolution 36: 903-913.
Lothian N. (1947) Critical notes on the genus Wahlenbergia Schrader, with descriptions of new species in the Australian region. Proc. Linn. Soc. NSW 71: 201-235.
Marticorena C., Stuessy T. F., Baeza C. M. (1998) Catalogue of the vascular flora of the Robinson Crusoe or Juan Fern/mdez Islands, Chile. Ga- yana Bot. 55: 187-211.
Martin F. W. (1959) Staining and observing pollen tubes in the style by means of fluorescence. Stain Technol. 34: 125-128.
Moeliono B., Tuyn P. (1960) Campanulaceae. In: Steenis C. G. G. J. Van (ed.) Flora Malesiana, Djakarta. 6: 107-141.
Mfiller H. (1883) The fertilization of flowers. (Translated by Thompson, D'A. W.). Macmillan and Co, London.
Nakamura M. (1935) On the irregular meiosis of the pollen mother cells of I. balsamina Linn. by the effect of artificial high temperature. Japan. J. Bot. 11: 118-123.
Nyman Y. (1991) Crossing experiments within the Campanula dichotoma group (Campanulaceae). Plant Syst. Evol. 177: 185-192.
Nyman Y. (1992) Pollination mechanisms in six Campanula species (Campanulaceae). Plant Syst. Evol. 181: 97-108.
Nyman Y. (1993a) The pollen-collecting hairs of Campanula (Campanulaceae). I. Morphological variation and the retractive mechanism. Amer. J. Bot. 80: 1427-1436.
Nyman Y. (1993b) The pollen-collecting hairs of Campanula (Campanulaceae). II. Function and adaptive significance in relation to pollination. Amer. J. Bot. 80: 1437-1443.
Petterson J. A. (1997) Revision of the genus Wahlenbergia (Campanulaceae) in New Zealand. N. Zeal. J. Bot. 35: 9-54.
Ricci M., Eaton L. (1994) The rescue of Wahlen- bergia larrainii in Robinson Crusoe Island, Chile. Biol. Conserv. 68: 89-93.
Richards A. J. (1997) Plant breeding systems. 2nd edn. Chapman and Hall, London.
Richardson T. E., Stephenson A. G. (1989) Pollen removal and pollen deposition affect the dura- tion of the staminate and pistillate phases in Campanula rapunculoides. Amer. J. Bot. 76: 532- 538.
Rosatti T. J. (1986) Sphenocleaceae and Campa- nulaceae. J. Arnold Arbor. 67: 38-45.
Sanders R. W., Stuessy T. F., Rodriguez R. (1983) Chromosome numbers from the flora of the Juan Fernandez Islands. Amer. J. Bot. 70: 799- 810.
Schaal B. A., Leverich W. J. (1980) Pollination and banner markings in Lupinus texensis (Legumi- nosae). Southw. Nat. 25: 280-282.
Shetler S. G. (1962) Notes on the life history of Campanula americana, the tall bellflower. Mich. Bot. 1: 9-14.
Shetler S. G. (1979) Pollen-collecting hairs of Campanula (Campanulaceae), I: Historical re- view. Taxon 28: 205-215.
Shetler S. G. (1982) Variation and evolution of the Nearctic harebells (Campanula subsect. Hetero- phylla). J. Cramer, Vaduz.
Skottsberg C. (1928) Pollinationsbiologie und Samenverbreitung auf den Juan Fernandez

G. J. Anderson et al.: Reproductive biology of Wahlenbergia 123
Inseln. In: Skottsberg C. (ed.) The natural history of the Juan Fernandez and Easter Islands, 2, Botany, Almqvist & Wiskell, Uppsala, pp. 503- 547.
Smith P. J. (1992) A revision of the genus Wahlen- bergia (Campanulaceae) in Australia. Telopea 5: 91-175.
Spooner D. M., Stuessy T. F., Crawford D. J., Silva O. M. (1987) Chromosome numbers from the flora of the Juan Fernandez Islands. II. Rhodora 89: 351-356.
Stuessy T. F., Crawford D. J., Marticorena C. (1990) Patterns of phylogeny in the endemic vascular flora of the Juan Fernandez Islands, Chile. Syst. Bot. 15: 338-346.
Stuessy T. F., Marticorena C., Rodriguez R., Crawford D. J., Silva O. M. (1992) Endemism in the vascular flora of the Juan Fernandez Islands. Aliso 13: 297-307.
Stuessy T. F., Sanders R. W., Silva M. (1984) Phytogeography and evolution of the flora of the Juan Fernandez Islands: a progress report. In: Radovsky F. J., Raven P. H., Sohmer S. H. (eds.) Biogeography of the tropical Pacific. Assoc. Syst. Coll. and B.P. Bishop Museum, Lawrence, Kansas, pp. 55-69.
Stuessy T. F., Swenson U., Crawford D. J., Anderson G., Silva O. M. (1998) Plant conser- vation in the Juan Fernandez Archipelago, Chile. Aliso 16: 89-101.
Sun B. Y., Huessy T. D., Crawford D. J. (1990) Chromosome counts from the flora of the Juan Fernandez Islands, Chile. III. Pacific Science 44: 258-264.
Thomson G. M. (1880) On the fertilization, etc., of New Zealand flowering plants. Trans. N. Zeal. Inst. 13: 241-291.
Thulin M. (1975) The genus Wahlenbergia s. lat. (Campanulaceae) in tropical Africa and Mada- gascar. Symb. Botanicae Upsal. 21: 1-223.
Uphof J. C. (1962) Plant hairs. In: Zimmerman W., Ozenda P. G. (eds.) Encyclopedia of Plant Anatomy, 2nd edn., 4. Borntraeger, Berlin, pp. 1-206.
Wyatt R. (1984) The evolution of self-pollination in granite outcrop species of Arenaria (Caryophyll- aceae). I. Morphological correlates. Evolution 38:80d~816.
Yeo P. F. (1993) Secondary pollen presentation. Plant Syst. Evol., Suppl. 6. Springer, New York, pp. 1-268.
Zink R. A., Wheelwright N. T. (1997) Facultative self-pollination in island irises. Amer. Midl. Nat. 137: 72-78.
Addresses of the authors: Gregory J. Anderson, Department of Ecology and Evolutionary Biology, Storrs, CT 06269-3043, USA; Gabriel Bernardello, Instituto Multidisciplinario de Biologia Vegetal, C. C. 495, 5000 Cdrdoba, Argentina; Patricio Lopez S., Casilla 1340, Concepcidn, Chile; Daniel J. Crawford, Department of Evolution, Ecology, and Organismal Biology, Ohio State University, Columbus, OH 43210, USA; Tod F. Stuessy, Institute of Higher Plant Systematics and Evolu- tion, Institute of Botany, University of Vienna, Rennweg 14, Vienna A-1030, Austria.





























