Embed Size (px)
Citation preview

Copyright � 2009 by the Genetics Society of AmericaDOI: 10.1534/genetics.109.105841
Reproductive Value and Fluctuating Selectionin an Age-Structured Population
Steinar Engen,*,1 Russell Lande†,‡ and Bernt-Erik Sæther‡
*Centre for Conservation Biology, Department of Mathematical Sciences and ‡Centre for Conservation Biology, Department of Biology,Norwegian University of Science and Technology, N-7491 Trondheim, Norway and †Division of Biology, Imperial College London,
Silwood Park Campus, Ascot, Berkshire SL5 7PY, United Kingdom
Manuscript received June 3, 2009Accepted for publication July 15, 2009
ABSTRACT
Fluctuations in age structure caused by environmental stochasticity create autocorrelation and transientfluctuations in both population size and allele frequency, which complicate demographic andevolutionary analyses. Following a suggestion of Fisher, we show that weighting individuals of differentage by their reproductive value serves as a filter, removing temporal autocorrelation in populationdemography and evolution due to stochastic age structure. Assuming weak selection, random mating, anda stationary distribution of environments with no autocorrelation, we derive a diffusion approximation forevolution of the reproductive value weighted allele frequency. The expected evolution obeys an adaptivetopography defined by the long-run growth rate of the population. The expected fitness of a genotype isits Malthusian fitness in the average environment minus the covariance of its growth rate with that of thepopulation. Simulations of the age-structured model verify the accuracy of the diffusion approximation.We develop statistical methods for measuring the expected selection on the reproductive value weightedallele frequency in a fluctuating age-structured population.
THE evolutionary dynamics of age-structured pop-ulations were formalized by Charlesworth
(1980, 1994) and Lande (1982) on the basis of earlierideas of Fisher (1930, 1958), Medawar (1946, 1952),and Hamilton (1966), showing that the strength ofselection on genes affecting the vital rates of survival orfecundity depends on their age of action (reviewed byde Jong 1994; Charlesworth 2000). Fisher definedthe reproductive value of individuals of a given ageas their expected contribution to future populationgrowth, determined by the age-specific vital rates. Thishas the property that in a constant environment thetotal reproductive value in a population always in-creases at a constant rate. The total population size,however, undergoes transient fluctuations as the stableage distribution is approached, and the total popula-tion size only asymptotically approaches a constantgrowth rate (Caswell 2001).
Environmental stochasticity creates continual fluctu-ations in age structure, producing temporal autocorre-lation in population size and in allele frequencies,which seriously complicate demographic and evolution-ary analyses. Fisher (1930, 1958, p. 35) suggested foranalysis of genetic evolution that individuals should beweighted by their reproductive value to compensate fordeviations from the stable age distribution. Here we
apply this suggestion to study weak fluctuating selec-tion in an age-structured population in a stochasticenvironment.
One of the central conceptual paradigms of evolu-tionary biology was described by Wright (1932). Hisadaptive topography represents a population as a pointon a surface of population mean fitness as a functionof allele frequencies. Assuming weak selection, ran-dom mating, and loose linkage (implying approximateHardy–Weinberg equilibrium within loci and linkageeqilibrium among loci), natural selection in a constantenvironment causes the population to evolve uphill ofthe mean fitness surface (Wright 1937, 1945, 1969;Arnold et al. 2001; Gavrilets 2004). Evolution by nat-ural selection thus tends to increase the mean fitness ofa population in a constant environment.
Lande (2007, 2008) generalized Wright’s adaptivetopography to a stochastic environment, allowing density-dependent population growth but assuming density-independent selection, showing that the expectedevolution maximizes the long-run growth rate of thepopulation at low density, r � r � s2
e=2. Here r is pop-ulation growth rate at low density in the averageenvironment and s2
e is the environmental variance inpopulation growth rate among years, which are stan-dard parameters in stochastic demography (Cohen
1977, 1979; Tuljapurkar 1982; Caswell 2001; Lande
et al. 2003). In this model of stochastic evolution theadaptive topography describing the expected evolutionis derived by expressing r and s2
e as functions of allele
1Corresponding author: Department of Mathematical Sciences, Norwe-gian University of Science and Technology, N-7491 Trondheim,Norway. E-mail: [email protected]
Genetics 183: 629–637 (October 2009)

frequencies with parameters being the mean Malthu-sian fitnesses of the genotypes and their temporalvariances and covariances. These results are based ondiffusion approximations for the coupled stochasticprocesses of population size and allele frequencies in afluctuating environment.
Diffusion approximations are remarkably accuratefor many problems in evolution and ecology (Crow andKimura 1970; Lande et al. 2003). Because a diffusionprocess is subject to white noise with no temporalautocorrelation, the approximation is most accurate ifthe noise in the underlying biological process is approx-imately uncorrelated among years. Despite temporalautocorrelation in total population size produced byage-structure fluctuations, the stochastic demographyof age-structured populations over timescales of ageneration or more can nevertheless be accuratelyapproximated by a diffusion process (Tuljapurkar
1982; Lande and Orzack 1988; Engen et al. 2005a,2007). The success of the diffusion approximation fortotal population size occurs because the noise in thetotal reproductive value is nearly white, with no tempo-ral autocorrelation to first order, and the log of totalpopulation size fluctuates around the log of reproduc-tive value with a return time to equilibrium on the orderof a few generations (Engen et al. 2007). Hence thediffusion approximation is well suited to describe thestochastic dynamics of total reproductive value as well astotal population size.
This article extends Lande’s (2008) model of fluctu-ating selection without age structure by deriving adiffusion approximation for the evolution of an age-structured population in a stochastic environment.Assuming weak selection at all ages, random mating,and a stationary distribution of environments with notemporal autocorrelation, we show that the main resultsof the model remain valid, provided that the modelparameters are expressed in terms of means, variances,and covariances of age-specific vital rates and that allelefrequencies are defined by weighting individuals ofdifferent age by their reproductive value, as suggestedby Fisher (1930, 1958). We perform simulations toverify the accuracy of the diffusion approximation andoutline statistical methods for estimating the expectedselection acting on the reproductive value weightedallele frequency.
STOCHASTIC DEMOGRAPHY ANDREPRODUCTIVE VALUE
In a stage-structured population with ni individuals instage i let the dynamics of the population column vectorn be governed by the stochastic projection matrix Lgiving the population vector in the next year Ln. Theexpected projection matrix in the average environmentis denoted as l. Projection matrices for an age-structuredpopulation have nonzero elements in the first row
representing age-specific annual fecundities and onthe subdiagonal representing age-specific annual sur-vival probabilities (Leslie 1945, 1948). For a stage-structured population nonzero elements may also occuron the diagonal representing stage-specific annualsurvival without transition and below the subdiagonalor above the diagonal representing nonadjacent stagetransitions (Lefkovitch 1965; Caswell 2001).
The stochastic projection matrices are assumed to beindependent and identically distributed through time.Subscripts l and L are used to indicate dependence onthe expected matrix l and the stochastic matrix L. Letthe column vector ul and row vector vl with componentsuli and vli, i ¼ 1; 2; . . . ; k, be the right and left eigen-vector of l associated with the dominant real eigenvaluell, defined by lul¼ llul and vll¼ llvl. If the eigenvectorsare scaled so that
Pi uli ¼ 1 and
Pi ulivli ¼ 1, then ul is
the stable age distribution and vl is the vector ofreproductive values for the stages.
The total reproductive value in the population isVL ¼ vl n ¼
Pi nivli . In a constant environment, the
total reproductive value grows at a constant rate by anannual multiplicative factor ll even under departuresfrom the stable age distribution, although the totalpopulation size grows at this rate only asymptotically asa stable age distribution is approached (Fisher 1930;Leslie 1948; Caswell 2001). Engen et al. (2007)showed that the first-order approximation to the annualchange in reproductive value is
D ln VL � ln ll 1 ln½1 1 l�1l vleLul �; ð1Þ
where eL¼ L� l. This shows that the log of reproductivevalue approximates a random walk with no temporalautocorrelation in the noise. First-order Taylor expan-sion of the noise terms yields the mean and variance ofannual changes in total reproductive value under smallnoise
E ½D lnVL� ¼ rL ¼ rl �s2
L
2; ð2Þ
where rl ¼ ln ll is the growth rate (or Malthusianparameter) in the average environment, rL is the long-run growth rate, and
s2L ¼ var½D lnVL � ¼ l�2
l
Xijab
vliulj vlaulbcov½eLij ; eLab � ð3Þ
is the environmental variance in total reproductivevalue (Tuljapurkar 1982; Caswell 2001; Lande et al.2003; Engen et al. 2007). Here eLij denotes the elementin the ith row and jth column of eL. The total re-productive value equals the total population size NL ifthe population is exactly at its stable age distribution,and Engen et al. (2007) showed that ln NL undergoesstationary fluctuations around ln VL with a return timeto equilibrium of about one generation. That theprocess ln VL has approximately white noise explains
630 S. Engen, R. Lande and B.-E. Sæther

the success of the diffusion approximation for ln VL
(and ln NL), identified simply as the Wiener process withinfinitesimal mean and variance rL and s2
L (Lande andOrzack 1988).
Note that the stable age distribution and reproductivevalue vector appearing in the approximations are thoseassociated with the dominant eigenvalue of the ex-pected projection matrix l ¼ EL and not the expectedvalue of the vectors associated with the stochastic matrixL (Tuljapurkar 1982; Lande and Orzack 1988). Thisoccurs because the theory assumes small noise, explor-ing by first-order expansions how the stochastic dynam-ics deviate from the dynamics in a constant environmentdetermined by the dominant eigenvalue of l and thecorresponding eigenvectors ul and vl.
Analyzing allele frequency evolution requires consid-eration of two or more correlated age-structured pro-cesses. Along with the stochastic projection matrix Ldescribed above, let M be the stochastic projectionmatrix for a different genotype with expectation m inthe average environment. Generally, the environmentsmay have distinct effects on different genotypes so thatthe environmental correlations between correspondingelements in L and M are less than one. In addition to thelong-run growth rates and environmental variances forthe two processes, we then also need to incorporate inthe model an environmental covariance between the twoprocesses that can be expressed using the above first-order approximation to the noise term in Equation 1 as
cov½D lnVL ; D lnVM � ¼ ðll lmÞ�1Xijab
vliulj vmaumbcov½eLij ; eMab �:
ð4Þ
We employ the simplified notation C(L, M) for thiscovariance so that the environmental variances associ-ated with the two stochastic projection matrices are s2
L ¼C(L, L) and s2
M ¼ C(M, M). The joint process forln VL and ln VM can then be approximated by a two-dimensional Wiener process with infinitesimal covari-ance C(L, M).
The approximations in Equations 2 and 3, firstderived by Tuljapurkar (1982) for population sizeand here derived more simply using reproductive value,are asymptotically exact as the temporal variances invital rates approach zero. However, these approxima-tions have good accuracy for coefficients of variation invital rates up to 30% and for larger coefficients ofvariation in vital rates that have small sensitivities viuj
(Lande et al. 2003). Lande and Orzack (1988) firstemployed these results in a diffusion approximation forthe total population size in a stochastic environment.Engen et al. (2007) pointed out that these approxima-tions were more accurate for the total reproductivevalue that contains all information on future populationsize as originally shown by Fisher (1930) for a constantenvironment. This result was based on showing that thelog of total population size fluctuates around the log of
total reproductive value with a characteristic return timeto equilibrium of about one generation. The accuracy ofthese diffusion models, which has been confirmed bystochastic simulations (Engen et al. 2005a), occurs becausethe process ln VL approximates a random walk, which isknown to be accurately described as a Brownian motion(or Wiener) diffusion process (Karlin and Taylor 1981).This argument also justifies using a two-dimensionaldiffusion for (ln VL, ln VM) with the covariance given byEquation 4 derived from a first-order approximation tothe noise term in Equation 1 as done for the variances.
STOCHASTIC EVOLUTION WITHOUTAGE STRUCTURE
For a population without age structure in a constantenvironment, with no density regulation, weak selec-tion, and random mating, classical theory for continu-ous-time models reveals that the population size N growsapproximately as
dN
dt¼ rN ; ð5Þ
where r ¼P
i
Pj pipj rij is the mean Malthusian fitness in
the population, pi is the frequency of allele Ai, and rij isthe Malthusian fitness of genotype AiAj. The correspond-ing rate of allele frequency evolution is approximately
dpi
dt¼ piðri � rÞ ¼ pið1� piÞ
2
@r
@pi; ð6Þ
where ri ¼P
j pj rij is the mean fitness of allele Ai
(Fisher 1930; Crow and Kimura 1970). The final formof Equation 6 is Wright’s (1937, 1969) adaptive topog-raphy in continuous time.
Lande (2008) showed that the above results hold alsofor fluctuating selection with environmental variancesand covariances in genotypic fitnesses, provided that inEquation 5 and in the final form of Equation 6 r isreplaced by the long-run growth rate of the population,r ¼ r � s2
e=2, where s2e is the environmental variance in
population growth rate. This result is derived from thestochastic fitness rij 1 dBij(t)/dt, where the Bij(t) areBrownian motions with E[dBij(t)] ¼ 0 and E[dBij(t)dBab(t)] ¼ cijabdt (Karlin and Taylor 1981). The sto-chastic differential equation for Ni is then dNi ¼riNidt 1 Ni
Pj pj dBijðtÞ. Using the Ito stochastic calculus
(Turelli 1977), the infinitesimal covariance betweenln Ni and ln Nj is
Pab papbciajb and the environmental
variance in population growth rate takes the form
s2e ¼
1
dt
Xij
pipjcov½dlnNi ; dlnNj � ¼Xijab
pipjpapbcijab :
ð7Þ
Finally, for simplicity, consider only two alleles A0
and A1, writing 2Ni ¼ 2Npi for the abundance of Ai andp0 ¼ p ¼ 1 � q for the frequency of A0. The expected
Fluctuating Selection and Age Structure 631

change in allele frequency can be expressed using anexpected selection coefficient, w0 � w1, in a formanalogous to the classical deterministic model (Crow
and Kimura 1970),
Edp
dt¼ pð1� pÞðw0 � w1Þ; ð8Þ
where the expected fitness of allele Ai, wi ¼P
j pj wij , isthe weighted average expected fitness of genotypescontaining it, wij ¼ rij �
Pab papbcijab . The expected
fitness of a genotype can therefore be defined as itsMalthusian fitness in the average environment minusthe covariance of its growth rate with that of thepopulation (Lande 2008).
On the logit scale, y¼ ln[p/(1� p)], the infinitesimalmean defining the diffusion approximation for y takesits simplest form,
mðyÞ ¼X
a
ðr0a � r1aÞpa �1
2
Xab
ðc0a0b � c1a1bÞpapb ð9Þ
with indexes a and b summed over 0 and 1 so that pa ¼p1�aqa and similarly for the infinitesimal variance
nðyÞ ¼X
ab
ðc0a0b 1 c1a1b � 2c0a1bÞpapb : ð10Þ
Equations 9 and 10 define the diffusion approximationfor y by substituting p¼ ey/(1 1 ey). The diffusion for p isfound by the reverse transformation (Karlin andTaylor 1981), giving the infinitesimal mean andvariance
aðpÞ ¼ pð1� pÞmðyÞ1 1
2pð1� pÞð1� 2pÞnðyÞ ð11Þ
bðpÞ ¼ p2ð1� pÞ2nðyÞ: ð12Þ
In stochastic models of genetic drift in a finitepopulation, when using the Ito calculus to computethe infinitesimal mean and variance of a diffusionapproximation, it is often assumed that the environ-ment influences each genotype identically (Engen et al.2005a; Shpak 2007). However, in models of fluctuatingselection (Lande 2007, 2008) stochastic environmentsexert distinct influences on the demography of differ-ent genotypes, producing a positive environmentalvariance contributing to the infinitesimal varianceb(p) driving stochastic changes in allele frequencieseven in populations sufficiently large to neglect geneticdrift. The next section demonstrates that this model canbe applied to an age-structured population by interpret-ing model parameters in terms of age-specific vital ratesand calculating allele frequency by weighting individu-als of different age by their reproductive value. The finalsection outlines statistical methods for estimating theexpected selection coefficient acting on the reproduc-tive value weighted allele frequency.
STOCHASTIC EVOLUTION WITH AGE STRUCTURE
We proceed to analyze the stochastic model withmultiple alleles Ai when the population has stages 1,2, . . . , k. Individuals of the ordered genotype AiAj definea projection matrix Lij ¼ Lji that fluctuates throughtime with mean lij and no temporal autocorrelation. Fora Leslie matrix model (Caswell 2001), the elements inthe first row of the matrices expressing fecundity arehalf the mean number of offspring produced by thegenotype at each age in the given year, that is, the meannumber of copies of each allele transmitted to offspring.The subdiagonal elements are their survival probabili-ties. The long-run growth rate associated with thestochastic matrix for each genotype Lij, denoted as rij ,is that of a hypothetical pure population of genotypeAiAj. Each genotype AiAj also has an environmentalvariance C(Lij, Lij), and every pair of genotypes AiAj andAaAb has an environmental covariance C(Lij, Lab).Fluctuating selection is generated by the environmentalfluctuations in the matrices Lij provided that these arenot all perfectly correlated, while differences betweenthe expected matrices lij cause selection in the averageenvironment.
We assume random mating among reproductiveindividuals of all stages and weak selection at all stagessuch that differences among corresponding elements ofthe matrices Lij among genotypes are small. All stagestherefore remain close to Hardy–Weinberg equilib-rium. The population also is assumed to undergodensity-independent growth and to be sufficiently largeto ignore random genetic drift, so that genotypic pro-jection matrices are independent of population density.Let Xi denote the (column) vector describing thenumber of Ai alleles in individuals at each stage. Thestochastic projection matrices for allelic numbers Xi arethen approximately Li ¼
Pj pjLij with mean li ¼P
j pj lij . The (row) vector of reproductive values forallele Ai is the left eigenvector, vi, associated with thedominant eigenvalue of li.
The total reproductive value of alleles Ai in the age-structured population is Vi ¼ viXi ¼
Pa viaXia , where
the summation over a covers all component stages of thevectors. When the projection matrix Xi fluctuates intime, the dynamics of the log of total reproductive valuehave no temporal autocorrelation to the first order, as inthe case of no age structure. Over a period short enoughfor pi to remain nearly constant, the joint process ln Vi
for all the alleles can accordingly be described as amultivariate Wiener process with infinitesimal meansthat are the long-run growth rates ri associated with thematrices Li ¼
Pj pjLij and infinitesimal covariances
C(Li, Lj) as above.Under weak selection at all stages, the dominant
eigenvalues and eigenvectors are similar for all matriceslij. The Malthusian fitness of allele Ai in the averageenvironment, given by the dominant eigenvalue of the
632 S. Engen, R. Lande and B.-E. Sæther
matrix li ¼P
pj lij , can then be approximated by a linearfunction in the pj, that is, ri �
Pj pj rij . This linear
approximation is very accurate under weak selection,justifying the application of Fisher’s formula (the firstform of Equation 6) to an age-structured populationin a constant environment, as proven rigorously byCharlesworth (1980, 1994). From Equation 4 it also fol-lows that CðLi ; LjÞ �
Pab papbciajb , where ciajb¼ C(Lia, Ljb).
The assumption of weak selection is required toapproximate the growth rate ri of the total reproductivevalue of Ai alleles as a linear function of allele frequen-cies pi, as in deterministic and stochastic models with noage structure (Equations 6 and 8). Hence, we requirethat the matrices lij for the different genotypes aresufficiently close for this approximation to be valid. Inpractice, when estimates of the matrices are availableand these indicate that selection may be too strong forthe theory to be applicable, it is recommended to checknumerically the linearity of the ri as functions of the pj.
Adopting the above approximations, the stochasticprocess for the reproductive values ðV1; V2; . . . ; VkÞ,conditioned on the allele frequencies, is the same as themodel for ðN1;N2; . . . ;NkÞ without age structure out-lined in a previous section. The total reproductive valuein the population, V ¼
Pi Vi , thus grows at the rate
r ¼P
ij pipj rij . Furthermore, for given allele frequenciespi, the reproductive value weighted allele frequencies
Pi ¼ Vi
.Xj
Vj ð13Þ
have the same dynamics as in Lande (2008). Completeformulas for the joint dynamics of population size andallele frequencies in terms of infinitesimal means,variances, and covariances are given by Lande (2008,Equations 3a, 3b, and 4a–4e).
For an age-structured population, the expectedevolution is
EdPi
dt¼ pið1� piÞ
2
@r
@pi: ð14Þ
On the left side the expected evolution of Pi refers toreproductive value weighted frequencies defined byEquation 13, while on the right side the pi are un-weighted allele frequencies from individual counts.Transient fluctuations due to temporal autocorrelationsin the log of allelic numbers, ln Ni, caused by stochasticage structure are to first order absent from the dynamicsof the log of total reproductive value of the allele, ln Vi.The allele frequency, pi, therefore fluctuates around thereproductive value weighted allele frequency Pi. Onaverage, unconditionally with respect to age structure,EdPi¼ Edpi, and the above model therefore also defines,in the unconditional sense, the adaptive topography forthe pi. Conditioned on the age structure, the expectedevolution of Pi obeys Wright’s adaptive topography,whereas the expected evolution of pi differs somewhat.If the population is exactly at the stable age distribution,
then Pi¼ pi and EdPi¼ Edpi. Given an age structure thatdeviates from the stable age distribution, only Pi isexpected to follow Wright’s adaptive topography, whilepi instead evolves to track the path of Pi.
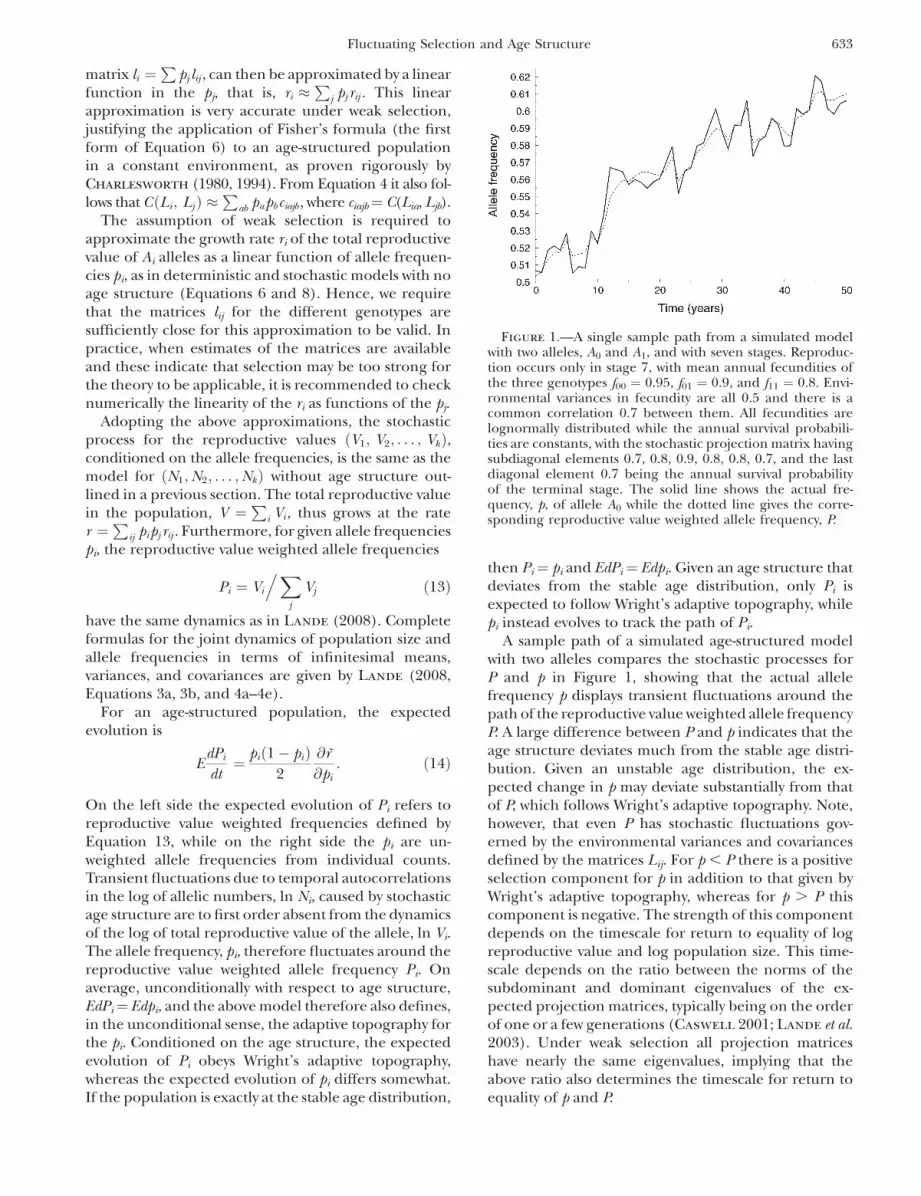
A sample path of a simulated age-structured modelwith two alleles compares the stochastic processes forP and p in Figure 1, showing that the actual allelefrequency p displays transient fluctuations around thepath of the reproductive value weighted allele frequencyP. A large difference between P and p indicates that theage structure deviates much from the stable age distri-bution. Given an unstable age distribution, the ex-pected change in p may deviate substantially from thatof P, which follows Wright’s adaptive topography. Note,however, that even P has stochastic fluctuations gov-erned by the environmental variances and covariancesdefined by the matrices Lij. For p , P there is a positiveselection component for p in addition to that given byWright’s adaptive topography, whereas for p . P thiscomponent is negative. The strength of this componentdepends on the timescale for return to equality of logreproductive value and log population size. This time-scale depends on the ratio between the norms of thesubdominant and dominant eigenvalues of the ex-pected projection matrices, typically being on the orderof one or a few generations (Caswell 2001; Lande et al.2003). Under weak selection all projection matriceshave nearly the same eigenvalues, implying that theabove ratio also determines the timescale for return toequality of p and P.
Figure 1.—A single sample path from a simulated modelwith two alleles, A0 and A1, and with seven stages. Reproduc-tion occurs only in stage 7, with mean annual fecundities ofthe three genotypes f00 ¼ 0.95, f01 ¼ 0.9, and f11 ¼ 0.8. Envi-ronmental variances in fecundity are all 0.5 and there is acommon correlation 0.7 between them. All fecundities arelognormally distributed while the annual survival probabili-ties are constants, with the stochastic projection matrix havingsubdiagonal elements 0.7, 0.8, 0.9, 0.8, 0.8, 0.7, and the lastdiagonal element 0.7 being the annual survival probabilityof the terminal stage. The solid line shows the actual fre-quency, p, of allele A0 while the dotted line gives the corre-sponding reproductive value weighted allele frequency, P.
Fluctuating Selection and Age Structure 633

These results indicate that an accurate diffusionapproximation for the reproductive value weightedallele frequency P can be obtained by using the in-finitesimal mean and variance from the model withoutage structure (Equations 11 and 12) by substituting Pfor p throughout. Parameters of the model must also beexpressed using statistics of the age-specific vital rates,identifying the Malthusian fitness of a genotype in theaverage environment, rij, as log of the leading eigen-value of its mean projection matrix, lij, and identifyingthe environmental covariance between genotypes cijab asC(Lij, Lab). The adaptive topography (Equation 6 with rin place of r) and the expected fitness of an allele or agenotype (Equation 8), derived by Lande (2008) for adiploid population without age structure, then remainvalid for the age-structured model applied to thereproductive value weighted allele frequency.
To test the accuracy of the diffusion approximationwe compared it to simulations of age-structured pop-ulations for two distinct cases: the transitional probabil-ity distribution for allele frequency under fluctuatingselection of a consistent direction leading to quasi-fixation and selection of fluctuating direction pro-ducing a stationary distribution of allele frequency(Haldane and Jayakar 1963; Lande 2008). Simulatedquantiles of p for an advantageous allele on the way tofixation closely match those from the diffusion approx-imation for reproductive weighted allele frequency P, as
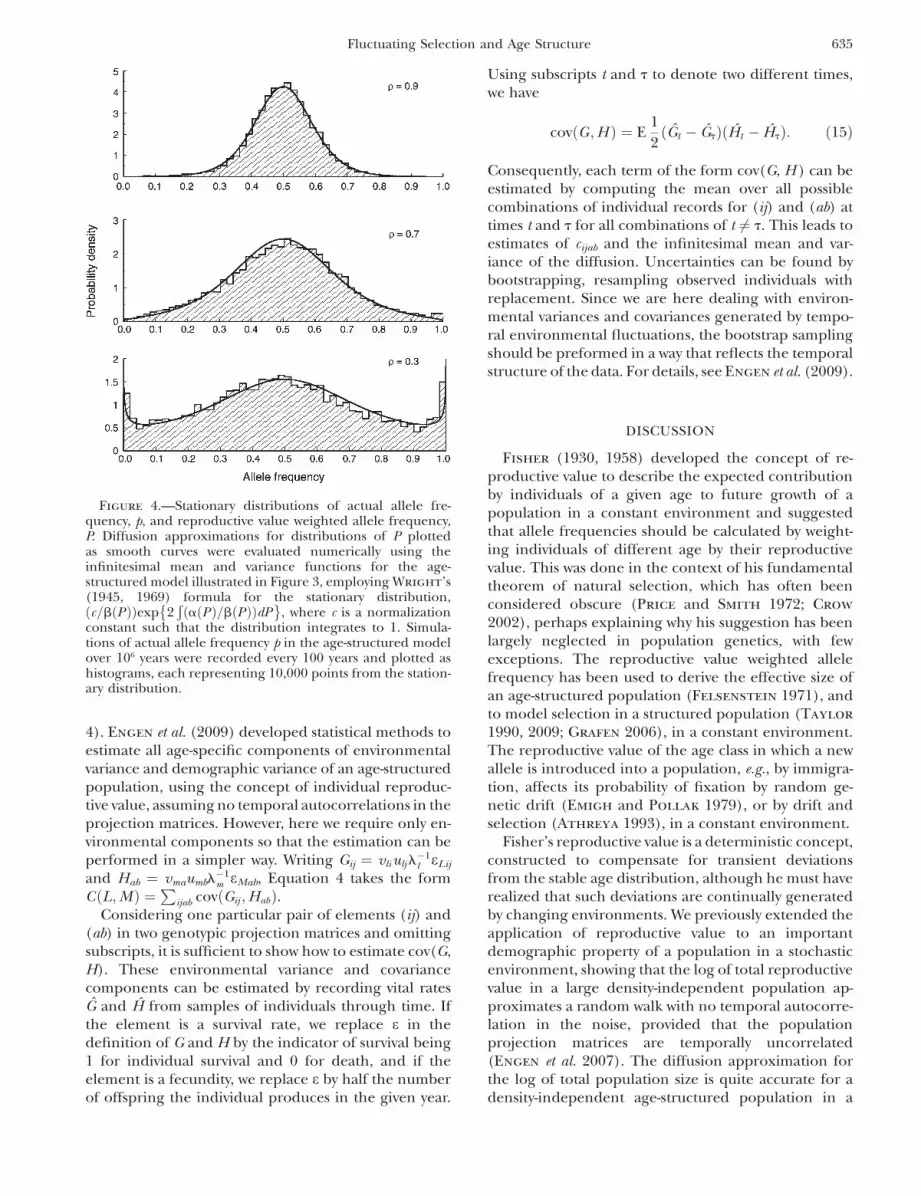
shown in Figure 2. Histograms of simulated stationarydistributions of allele frequency p also agree well withstationary distributions of the reproductive valueweighted frequency P derived from the diffusion ap-proximation, as illustrated in Figures 3 and 4 for amodel in which all three genotypes have the sameexpected life history, but the heterozygote has theadvantage of a smaller environmental variance infecundity than the homozygotes.
MEASUREMENT OF SELECTION
Measuring the expected selection coefficient actingon the reproductive value weighted allele frequency(Equation 8) requires estimation of parameters in theinfinitesimal mean and variance of the diffusion ap-proximation (Equations 9–12). The first step is toestimate the mean projection matrix for each genotypein the average environment by recording mean valuesof vital rates through time. From each mean projectionmatrix lij an estimate of the log of the dominant ei-genvalue rij and the corresponding eigenvectors can becomputed. With known allele frequencies this gives thefirst term of Equation 9 and it remains to estimate theenvironmental variances and covariances cijab (Equation
Figure 2.—Simulations from a model with five stages, withreproduction only in the terminal stage and mean annual fe-cundities of the genotypes f00 ¼ 1.00, f01 ¼ 0.9, and f11 ¼ 0.8.Environmental variances in annual fecundities are all 0.2 andthere is a common correlation 0.5 between them. All fecund-ities are lognormally distributed. The annual survival proba-bilities are constant with subdiagonal elements 0.7, 0.9, 0.9,0.7 and 0.6 for the last diagonal element. The solid line showsquantiles for the actual allele frequency p from 10,000 simu-lations of the full age-structured model, while the dotted linesare quantiles for the reproductive value weighted allele fre-quency P from 10,000 simulations of the diffusion approxima-tion.
Figure 3.—Infinitesimal mean a(P) and variance b(P) inthe diffusion approximation for reproductive value weightedallele frequency P (Equations 11 and 12) in a model with fivestages. All three genotypes have the same expected projectionmatrices. The annual survival probabilities are constant withsubdiagonal elements 0.7, 0.9, 0.9, 0.7 and 0.6 for the last di-agonal element. Only the terminal stage reproduces with ex-pected annual fecundity 0.9. Environmental variances infecundity are 0.3 for the homozygotes and 0.1 for the hetero-zygote. The correlations r between any two fecundities are thesame and vary among the graphs, r ¼ 0.9 (solid line), 0.7(dashed line), and 0.3 (dotted line).
634 S. Engen, R. Lande and B.-E. Sæther
4). Engen et al. (2009) developed statistical methods toestimate all age-specific components of environmentalvariance and demographic variance of an age-structuredpopulation, using the concept of individual reproduc-tive value, assuming no temporal autocorrelations in theprojection matrices. However, here we require only en-vironmental components so that the estimation can beperformed in a simpler way. Writing Gij ¼ vliulj l
�1l eLij
and Hab ¼ vmaumbl�1m eMab, Equation 4 takes the form
CðL;M Þ ¼P
ijab covðGij ;HabÞ.Considering one particular pair of elements (ij) and
(ab) in two genotypic projection matrices and omittingsubscripts, it is sufficient to show how to estimate cov(G,H). These environmental variance and covariancecomponents can be estimated by recording vital ratesG and H from samples of individuals through time. Ifthe element is a survival rate, we replace e in thedefinition of G and H by the indicator of survival being1 for individual survival and 0 for death, and if theelement is a fecundity, we replace e by half the numberof offspring the individual produces in the given year.
Using subscripts t and t to denote two different times,we have
covðG ;H Þ ¼ E1
2ðGt � GtÞðHt � HtÞ: ð15Þ
Consequently, each term of the form cov(G, H ) can beestimated by computing the mean over all possiblecombinations of individual records for (ij) and (ab) attimes t and t for all combinations of t 6¼ t. This leads toestimates of cijab and the infinitesimal mean and var-iance of the diffusion. Uncertainties can be found bybootstrapping, resampling observed individuals withreplacement. Since we are here dealing with environ-mental variances and covariances generated by tempo-ral environmental fluctuations, the bootstrap samplingshould be preformed in a way that reflects the temporalstructure of the data. For details, see Engen et al. (2009).
DISCUSSION
Fisher (1930, 1958) developed the concept of re-productive value to describe the expected contributionby individuals of a given age to future growth of apopulation in a constant environment and suggestedthat allele frequencies should be calculated by weight-ing individuals of different age by their reproductivevalue. This was done in the context of his fundamentaltheorem of natural selection, which has often beenconsidered obscure (Price and Smith 1972; Crow
2002), perhaps explaining why his suggestion has beenlargely neglected in population genetics, with fewexceptions. The reproductive value weighted allelefrequency has been used to derive the effective size ofan age-structured population (Felsenstein 1971), andto model selection in a structured population (Taylor
1990, 2009; Grafen 2006), in a constant environment.The reproductive value of the age class in which a newallele is introduced into a population, e.g., by immigra-tion, affects its probability of fixation by random ge-netic drift (Emigh and Pollak 1979), or by drift andselection (Athreya 1993), in a constant environment.
Fisher’s reproductive value is a deterministic concept,constructed to compensate for transient deviationsfrom the stable age distribution, although he must haverealized that such deviations are continually generatedby changing environments. We previously extended theapplication of reproductive value to an importantdemographic property of a population in a stochasticenvironment, showing that the log of total reproductivevalue in a large density-independent population ap-proximates a random walk with no temporal autocorre-lation in the noise, provided that the populationprojection matrices are temporally uncorrelated(Engen et al. 2007). The diffusion approximation forthe log of total population size is quite accurate for adensity-independent age-structured population in a
Figure 4.—Stationary distributions of actual allele fre-quency, p, and reproductive value weighted allele frequency,P. Diffusion approximations for distributions of P plottedas smooth curves were evaluated numerically using theinfinitesimal mean and variance functions for the age-structured model illustrated in Figure 3, employing Wright’s(1945, 1969) formula for the stationary distribution,ðc=bðPÞÞexp 2
ÐðaðPÞ=bðPÞÞdP
� �, where c is a normalization
constant such that the distribution integrates to 1. Simula-tions of actual allele frequency p in the age-structured modelover 106 years were recorded every 100 years and plotted ashistograms, each representing 10,000 points from the station-ary distribution.
Fluctuating Selection and Age Structure 635

random environment (Lande and Orzack 1988;Engen et al. 2005a). This occurs because this processactually approximates a diffusion for the log of totalreproductive value (Engen et al. 2007). For a given totalreproductive value, the log of total population size hasno additional predictive power, fluctuating around thelog of total reproductive value with a timescale of aboutone generation for return to equality of these twovariables.
Similarly, for evolution in an age-structured popula-tion, this article demonstrates that the actual allelefrequency p fluctuates around the reproductive valueweighted allele frequency P on a timescale of about onegeneration, due to stochastic fluctuations in the agestructure and temporal autocorrelation in p that isabsent to first order from the fluctuations in P (Figure1). These results together with Fisher’s suggestionnaturally lead to the derivation of a diffusion approxi-mation for the reproductive value weighted allelefrequency, P, in an age-structured population subjectto density-independent fluctuating selection (Equa-tions 11 and 12). The accuracy of this method isillustrated by comparisons of the diffusion approxima-tion with simulations of age-structured models (Figures2 and 4).
Distinguishing between total population size N andtotal reproductive value V, and between the unweightedand reproductive value weighted allele frequencies, pand P, is important for analyzing selection and evolutionin a fluctuating environment, despite the timescale forautocorrelation in N and p being only about one gen-eration. Many organisms of interest, such as largevertebrates or perennial plants, have generation timesof several years or more, and for such species investigatorstypically measure age-specific components of fitness onan annual basis. Age-specific vital rates have been com-bined into total lifetime fitness on the basis of simpli-fying assumptions of constant population size and/orconstant age structure (Fisher 1930; Charlesworth
1980; Lande 1982; Clutton-Brock 1988). Alterna-tively, analysis has focused on time series of age-averagedselection (Grant and Grant 2002; Sheldon et al. 2003;Garant et al. 2004) or time-averaged age-specific se-lection (Pelletier et al. 2007). Recent extension of thePrice equation to include fluctuating age structure re-mains genetically ambiguous (Coulson and Tuljapurkar
2008). In contrast, using reproductive value weighting,and measuring the fitness of a genotype by the de-mographic growth rate of its total reproductive value,overcomes the problems caused by autocorrelation andcorrectly combines all components of fitness in a fluc-tuating age-structured population.
Assuming random mating and weak selection at allages, the evolution of reproductive value weighted allelefrequency, P, in an age-structured population follows thediffusion approximation derived by Lande (2008) forevolution of allele frequencies in a population without
age structure in a fluctuating environment. The ex-pected evolution of P obeys a generalization of Wright’sadaptive topography (Equation 6), maximizing thelong-run growth rate of the population, r ¼ r � s2
e=2,as a function of allelic frequencies, where r is the meanMalthusian fitness in the average environment and s2
e isthe environmental variance in population growth rate.Contrary to common belief, the expected fitness of agenotype within a population is its Malthusian fitness inthe average environment minus the covariance of itsgrowth rate with that of the population (Equation 8).For an age-structured population the model parametersmust be expressed using basic statistics of age-specificvital rates of the genotypes. The Malthusian fitness of agenotype in the average environment is log of theleading eigenvalue of its mean projection matrix, andthe variances and covariances of genotypic growth ratesdepend on patterns of variability in their vital rates(Equation 4).
The present theory assumes large population size anddensity-independent vital rates, which together ensurethat the stochastic projection matrices are not influ-enced by the population vector. Genetic drift in a smallage-structured population can be represented by anadditional term in the infinitesimal variance of thediffusion approximation (Engen et al. 2005b; Shpak
2007). The model can then be used to analyze theprobability of fixation and the time to fixation as theboundaries now become accessible to the diffusionprocess. Density-dependent selection in a fluctuatingenvironment has been analyzed to study evolutionsubject to life-history trade-offs in simple models withno age structure (Lande et al. 2009). Extending modelsof density-dependent selection to age-structured pop-ulations may prove difficult because density regulationof population size generally produces complex inter-actions among the age classes (Lande et al. 2006). Onlyone special form of density regulation, where the pop-ulation vector exerts an identical multiplicative effecton all elements of the projection matrices in a given year(Desharnais and Cohen 1986), would preserve thedynamics of the age distribution and allele frequencies,leaving our results unchanged. The present resultsnevertheless provide a necessary step toward a moregeneral understanding of life-history evolution in fluc-tuating environments.
This work was supported by the Norwegian University of Science andTechnology through a grant to the Centre for Conservation Biology,The Research Council of Norway (Storforsk: Population genetics in anecological perspective), and the Royal Society of London.
LITERATURE CITED
Arnold, S. J., M. F. Pfrender and A. G. Jones, 2001 The adaptivelandscape as conceptual bridge between micro- and macro-evolu-tion. Genetica 112–113: 9–23.
Athreya, K. B., 1993 Rates of decay for the survival probability of amutant gene. II. The multitype case. J. Math. Biol. 32: 45–53.
636 S. Engen, R. Lande and B.-E. Sæther

Caswell, H., 2001 Matrix Population Models, Ed. 2. Sinauer Associ-ates, Sunderland, MA.
Charlesworth, B., 1980 Evolution in Age-Structured Populations.Cambridge University Press, Cambridge, UK.
Charlesworth, B., 1994 Evolution in Age-Structured Populations, Ed.2. Cambridge University Press, Cambridge, UK.
Charlesworth, B., 2000 Fisher, Medawar, Hamilton and the evolu-tion of aging. Genetics 156: 927–931.
Clutton-Brock, T. H., 1988 Reproductive Success. University of Chi-cago Press, Chicago.
Cohen, J. E., 1977 Ergodicity of age structure in populations withMarkovian vital rates, III: finite-state moments and growth rate;an illustration. Adv. Appl. Probab. 9: 462–475.
Cohen, J. E., 1979 Comparative statistics and stochastic dynamics ofage-structured populations. Theor. Popul. Biol. 16: 159–171.
Coulson, T., and S. Tuljapurkar, 2008 The dynamics of a quanti-tative trait in an age-structured population living in a variableenvironment. Am. Nat. 172: 599–612.
Crow, J. F., 2002 Perspective: here’s to Fisher, additive genetic var-iance, and the fundamental theorem of natural selection. Evolu-tion 56: 1313–1316.
Crow, J. F., and M. Kimura, 1970 An Introduction to Population Genet-ics Theory. Burgess/Alpha Editions, Minneapolis.
de Jong, G., 1994 The fitness of fitness concepts and the descriptionof natural selection. Q. Rev. Biol. 69: 3–29.
Desharnais, R. A., and J. E. Cohen, 1986 Life not lived due to dis-equilibrium in heterogeneous age-structured populations. The-or. Popul. Biol. 29: 385–406.
Emigh, T. H., and E. Pollak, 1979 Fixation probabilities and effec-tive population numbers in diploid populations with overlappinggenerations. Theor. Popul. Biol. 15: 86–107.
Engen, S., R. Lande, B.-E. Sæther and H. Weimerskirch,2005a Extinction in relation to demographic and environmen-tal stochasticity in age-structured models. Math. Biosci. 195: 210–227.
Engen, S., R. Lande and B.-E. Sæther, 2005b Effective size of a fluc-tuating age-structured population. Genetics 170: 941–954.
Engen, S., R. Lande and B.-E. Sæther, 2007 Using reproductivevalue to estimate key parameters in density-independent age-structured populations. J. Theor. Biol. 244: 308–317.
Engen, S., R. Lande, B.-E. Sæther and F. S. Dobson, 2009 Individualannual fitness, reproductive value, and the stochastic dynamics ofage-structured populations. Am. Nat. (in press)
Felsenstein, J., 1971 Inbreeding and variance effective numbers inpopulations with overlapping generations. Genetics 68: 581–597.
Fisher, R. A., 1930 The Genetical Theory of Natural Selection. Claren-don Press, Oxford.
Fisher, R. A., 1958 The Genetical Theory of Natural Selection, Ed. 2.Dover, New York.
Garant, D., L. E. B. Kruuk, R. H. McCleery and B. C. Sheldon,2004 Evolution in a changing environment: a case study withgreat tit fledging mass. Am. Nat. 164: E115–E129.
Gavrilets, S., 2004 Fitness Landscapes and the Origin of Species.Princeton University Press, Princeton, NJ.
Grafen, A., 2006 A theory of Fisher’s reproductive value. J. Math.Biol. 53: 15–60.
Grant, P. R., and B. R. Grant, 2002 Unpredictable evolution in a30-year study of Darwin’s finches. Science 296: 707–711.
Haldane, J. B. S., and S. D. Jayakar, 1963 Polymorphism due toselection of varying direction. J. Genet. 58: 237–242.
Hamilton, W. D., 1966 The moulding of senescence by natural se-lection. J. Theor. Biol. 12: 12–45.
Karlin, S., and H. M. Taylor, 1981 A Second Course in StochasticProcesses. Academic Press, New York.
Lande, R., 1982 A quantitative genetic theory of life history evolu-tion. Ecology 63: 607–615.
Lande, R., 2007 Expected relative fitness and the adaptive topogra-phy of fluctuating selection. Evolution 61: 1835–1846.
Lande, R., 2008 Adaptive topography of fluctuating selection in aMendelian population. J. Evol. Biol. 21: 1096–1105.
Lande, R., and S. H. Orzack, 1988 Extinction dynamics of age-structured populations in a fluctuating environment. Proc. Natl.Acad. Sci. USA 85: 7418–7421.
Lande, R., S. Engen and B.-E. Sæther, 2003 Stochastic PopulationDynamics in Ecology and Conservation. Oxford University Press,Oxford.
Lande, R., S. Engen, B.-E. Sæther and T. Coulson,2006 Estimating density dependence from time series of popu-lation age structure. Am. Nat. 168: 76–87.
Lande, R., S. Engen and B.-E. Sæther, 2009 An evolutionary max-imum principle for density-dependent population dynamics in afluctuating environment. Philos. Trans. R. Soc. Lond. 364: 1511–1518.
Lefkovitch, L. P., 1965 The study of population growth in organ-isms grouped by stages. Biometrics 21: 1–18.
Leslie, P. H., 1945 On the use of matrices in certain populationmathematics. Biometrika 33: 183–212.
Leslie, P. H., 1948 Some further notes on the use of matrices inpopulation mathematics. Biometrika 35: 213–245.
Medawar, P. B., 1946 Old age and natural death. Mod. Q. 1: 30–56.Medawar, P. B., 1952 An Unsolved Problem of Biology. H. K. Lewis,
London.Pelletier, F., T. Clutton-Brock, J. Pemberton, S. Tujapurkar and
T. Coulson, 2007 The evolutionary demography of ecologicalchange: linking trait variation and population growth. Science315: 1571–1574.
Price, G. R., and C. A. B. Smith, 1972 Fisher’s Malthusian param-eter and reproductive value. Ann. Hum. Genet. 36: 1–7.
Sheldon, B. C., L. E. B. Kruuk and J. Merila, 2003 Natural selec-tion and inheritance of breeding time and clutch size in the col-lared flycatcher. Evolution 57: 406–427.
Shpak, M., 2007 Selection against demographic stochasticity in age-structured populations. Genetics 177: 2181–2194.
Taylor, P. D., 1990 Allele-frequency change in a class-structuredpopulation. Am. Nat. 135: 95–106.
Taylor, P., 2009 Decompositions of Price’s formula in an inhomo-geneous population. J. Evol. Biol. 22: 201–213.
Tuljapurkar, S. D., 1982 Population dynamics in variable environ-ments. II. Correlated environments, sensitivity analysis anddynamics. Theor. Popul. Biol. 21: 114–140.
Turelli, M., 1977 Random environments and stochastic calculus.Theor. Popul. Biol. 12: 140–178.
Wright, S., 1932 The roles of mutation, inbreeding, cross-breedingand selection in evolution. Proc. IV Int. Congr. Genet. 1: 356–366.
Wright, S., 1937 The distribution of gene frequencies in popula-tions. Proc. Natl. Acad. Sci. USA 23: 305–320.
Wright, S., 1945 The differential equation of the distribution ofgene frequencies. Proc. Natl. Acad. Sci. USA 31: 383–389.
Wright, S., 1969 Evolution and the Genetics of Populations: The Theoryof Gene Frequencies, Vol. 2. University of Chicago Press, Chicago.
Communicating editor: J. Wakeley
Fluctuating Selection and Age Structure 637

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.





























