Embed Size (px)
Citation preview

This article was originally published in a journal published byElsevier, and the attached copy is provided by Elsevier for the
author’s benefit and for the benefit of the author’s institution, fornon-commercial research and educational use including without
limitation use in instruction at your institution, sending it to specificcolleagues that you know, and providing a copy to your institution’s
administrator.
All other uses, reproduction and distribution, including withoutlimitation commercial reprints, selling or licensing copies or access,
or posting on open internet sites, your personal or institution’swebsite or repository, are prohibited. For exceptions, permission
may be sought for such use through Elsevier’s permissions site at:
http://www.elsevier.com/locate/permissionusematerial

Autho
r's
pers
onal
co
py
Restraint stress decreases virus-induced pro-inflammatory cytokinemRNA expression during acute Theiler's virus infection
W. Mi a, b, T.W. Prentice c, C.R. Young a, c, R.R. Johnson c, A.N. Sieve c,M.W. Meagher c, C.J.R. Welsh a, b, d,⁎
a Department of Veterinary Integrative Biosciences, College of Veterinary Medicine and Biomedical Sciences, Texas A&M University,College Station, TX 77843, USA
b Genetics Program, Texas A&M University, College Station, TX 77843, USAc Department of Psychology, College of Liberal Arts, Texas A&M University, College Station, TX 77843, USA
d Department of Veterinary Pathobiology, College of Veterinary Medicine, Texas A&M University, College Station, TX 77843, USA
Received 2 December 2005; received in revised form 13 May 2006; accepted 17 May 2006
Abstract
Stressful life events have been associated with the onset and/or exacerbation of multiple sclerosis (MS). Our previous studies haveindicated that restraint stress (RS) reduces inflammation and virus-induced chemokine expression in the Theiler's virus-induceddemyelination (TVID) model of MS. Here we report that RS significantly reduced the virus-induced interferon-γ mRNA levels in the brain.Additionally, mRNA levels of lymphotoxin-β, tumor necrosis factor-α, and interferon-γ in the brain were negatively correlated with viraltiters in the brain. These results indicated an immunosuppressive effect of stress during early TVID causing impaired viral clearance, whichmay be a potential exacerbating factor for later demyelination.© 2006 Elsevier B.V. All rights reserved.
Keywords: Theiler's virus; Restraint stress; Cytokines
1. Introduction
The etiology of multiple sclerosis (MS) remains uncertainand multiple factors, such as genetic predisposition, andexternal environmental factors, including infectious patho-gens, psychological stressors, geographical locations, havebeen reported as being linked to the pathogenesis of MS. Inparticular, several viruses have been associated with theneurological disorder (Gilden and Lipton, 2005). Theiler'smurine encephalomyelitis virus-induced demyelination(TVID) has served for several decades as an animal modelto investigate infectious mechanisms mediating central
nervous system (CNS) demyelination in human MS (Lipton,1975). Intracerebral inoculation of Theiler's virus, a naturalmurine pathogen, produces a biphasic CNS disease insusceptible mice. The acute phase of disease involves a mildencephalomyelitis with predominant neuronal infection,whereas the chronic phase is characterized by inflammatorydemyelination in white matter with glial and microglialinfection (Lipton, 1975). Susceptible strains of mice fail toclear virus and a persistent CNS infection is established thatis essential for the induction of demyelination in the spinalcord (Lipton et al., 1984; Rodriguez et al., 1983; Oleszak etal., 2004). Although it is evident that the susceptibility toTheiler's virus infection and demyelination is geneticallyregulated (Brahic and Bureau, 1998), the immune responseplays a critical role in determining the viral load and thedegree of viral persistence, thus effecting the progression andseverity of demyelinating disease (Aubagnac et al., 1999;Rodriguez et al., 1996).
Journal of Neuroimmunology 178 (2006) 49–61www.elsevier.com/locate/jneuroim
⁎ Corresponding author. Department of Veterinary Integrative Bios-ciences, College of Veterinary Medicine and Biomedical Sciences, TexasA&M University, College Station, TX 77843-4458, USA. Tel.: +1 979 8624974; fax: +1 979 847 8981.
E-mail address: [email protected] (C.J.R. Welsh).
0165-5728/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.jneuroim.2006.05.010

Autho
r's
pers
onal
co
py
Acute encephalomyelitis in TVID is primarily mediatedby both innate and adaptive anti-viral immunity viainfiltrating NK cells (Paya et al., 1989) and CD4 + andCD8 + T cells (reviewed in Oleszak et al., 2004). Adaptiveimmunity contributes to the late demyelination, by delayed-type hypersensitivity (DTH) responses to viral antigens viaCD4 + T cells (Borrow et al., 1993; Gerety et al., 1994).Autoimmunity also contributes to the late demyelinatingdisease (Miller et al., 1997; Borrow et al., 1998; Dal Cantoet al., 2000).
In response to Theiler's virus infection, a variety ofinflammatory mediators, such as cytokines, chemokinesand enzymes are produced. Cytokines play a crucial role inthe regulation of the immune response in clearing the virusfrom the CNS and mediating demyelination. It has beenshown that a panel of predominantly pro-inflammatorycytokines, such as interferon-γ (IFN-γ), tumor necrosisfactor-α (TNF-α), interleukin (IL)-6, and interleukin (IL)-12, was induced during acute encephalomyelitis in allstrains of mice, but higher levels in susceptible mice thanin resistant mice (Chang et al., 2000; Sato et al., 1997;Theil et al., 2000). Some Th1 cytokines (i.e. IFN-γ),exhibit anti-viral activity in TVID as observed in knockoutmice (Rodriguez et al., 2003). In addition, Th1 cytokinesmay further activate and regulate other immune cells, suchas antigen presenting cells and microglia, thus promotingprotection against viral infection (reviewed by Oleszak etal., 2004).
After the first 2 weeks of infection, Th1 cytokine levelsdecline in resistant mice, which coincides with the clearanceof virus from the CNS; whereas in susceptible strains ofmice, Th1 cytokines remain at relatively high levels and arepartly responsible for the progression of demyelination(Begolka et al., 1998; Chang et al., 2000; Murray et al.,2002; Sato et al., 1997; Trottier et al., 2004). In contrast, theanti-inflammatory (Th2) cytokines (IL-10 and TGF-β) havebeen shown to alleviate demyelination in TVID (Drescher etal., 2000), and experimental allergic encephalomyelitis(Begolka et al., 1998). Interestingly, the cytokine shiftbetween Th1 and Th2 has also been correlated with thefluctuation in clinical manifestations of relapse-remitting MSand primary progressive MS (reviewed in Oleszak et al.,2004).
Increasing evidence demonstrates that stressful life eventsplay a role in both the onset and relapse of MS (Mohr et al.,2004). The stress response involves a complex interactionbetween the neuroendocrine and the immune systems,resulting in alterations in both innate and adaptive immunityto various pathogens and stimuli. These alterations enhancethe risk or vulnerability to inflammatory and autoimmunediseases, as well as infectious disease. The mechanisms bywhich stress modulates the pathogenesis or progression ofautoimmune diseases are not fully understood. Two majorpathways, the hypothalamic-pituitary-adrenal (HPA) axisand the systemic/adrenomedullary sympathetic nervoussystem (SNS) are activated in response to stress, resulting
in the production of glucocorticoids and catecholaminesfrom the two systems. Glucocorticoids and catecholaminesas well as other stress hormones, opioids, growth hormones,prolactin, profoundly impact the function of the immunesystem (Pruett, 2003). Among these stress hormones, therole of glucocorticoids in the immune response has beenextensively studied. It has been widely used as animmunosuppressor or antiinflammatory agent in the therapyof many autoimmune diseases, including MS.
On the other hand, the effectors of immune cells, such ascytokines, secreted in response to stress stimulation, caninfluence the neuroendocrine system via cytokine receptors,thus providing links between the two systems (reviewed inHaddad et al., 2002). Therefore, cytokines appear to be apivotal factor in the pathogenesis of MS and the stressresponse.
Previous studies from our laboratory have shown thatstress increased plasma corticosterone (Campbell et al.,2001), decreased NK cell activity (Welsh et al., 2004),reduced chemokine expression and CNS inflammation (Miet al., 2004) during the acute infection with Theiler's virus.In the current study, we further examine the cytokineexpression following restraint stress and Theiler's virusinfection and the stress-induced Th1/Th2 cytokine shift.These studies should enable us to gain some insight into thecomplex the role of environmental factors, stressor and virus,on the pathogenesis of MS.
2. Method and materials
2.1. Subjects and grouping
3-week-old male CBA mice were obtained from HarlanLaboratories (Harlan Sprague Dawley, Inc., Indianapolis,Indiana) and were maintained on a 12-h light/dark cycle. Themicewere group housed 3–4 per cage and counterbalanced byweight upon arrival and were then allowed to habituate for1 week before the initiation of any experimental procedures.
2.2. Experimental Design
A 2 (infected vs. non-infected) by 2 (restraint vs. non-restraint) factorial design was employed. At 4 weeks of age,cages were randomly assigned to one of the followingtreatment conditions: (1) Non-infected/Non-restrained (NI/NR): mice remained undisturbed in their home cages; (2)Non-infected/Restrained (NI/R): each mouse was subjectedto restraint stress in their home cages; (3) Infected/Non-restrained (I/NR): mice were infected intracerebrally withBeAn virus; or (4) Infected/Restrained (I/R): each mousewas intracerebrally infected with virus and subjected torestraint stress. In the first experiment (N=32, NI/NR=8, NI/R=8, I/NR=8, I/R=8), mice were terminated at day 2 p.i.,and in the second experiment (N=62, NI/NR=13, NI/R=13,I/NR=13, I/R=23), mice were terminated at day 7 p.i. Forthe viral titers, TNF-α protein and correlation and regression
50 W. Mi et al. / Journal of Neuroimmunology 178 (2006) 49–61

Autho
r's
pers
onal
co
py
experiment, we used a total of 16 mice, 4 for each condition(NI/NR, NI/R, I/NR, I/R).
2.3. Restraint stress paradigm
Restraint was applied by placing the mice into well-ventilated restraining tubes (2.7-cm internal diameter and14-cm length) for 12 h (from 9 pm–9 am) each for 3 or8 consecutive nights (Campbell et al., 2001; Mi et al.,2004; Sheridan et al., 1991). Restraint stress was initiated1 day prior to infection and continued for 2 or 7subsequent sessions.
2.4. Virus and viral inoculation
The BeAn strain of Theiler's virus was obtained from Dr.H.L. Lipton, Department of Microbiology-Immunology,University of Illinois at Chicago, IL and was propagatedand amplified in L2 cells as previously described (Welsh etal., 1987). Following isoflurane (Vedco Inc., St. Joseph, MO)anesthetization, 5×10 4 p.f.u. (plaque forming units) ofBeAn strain in a 20 μl volume was inoculated into the rightmid-parietal cortex at a depth of approximately 1.5 mm. Thenon-infected mice received a similar injection of 20 μl ofsterile phosphate-buffered saline (PBS).
2.5. Termination of mice
Mice were sacrificed by i.p. injection of pentobarbital ateither day 2 p.i. or day 7 p.i. Mice were perfused with diethylpyrocarbonate (DEPC, Sigma)-treated PBS into the rightventricle of the heart. The brains and spleens were asepticallyremoved, weighed, and then flash frozen in liquid nitrogenand stored at −80 °C until used.
2.6. Ribonuclease protection assay (RPA)
Total RNAwas extracted from the brain and spleen usingTRIzol (Invitrogen, Carlsbad, CA) according to themanufacturer's instructions. Two Multi-Probe TemplateSets, mCK-2b (IL-12p35, IL-12p40, IL-10, IL-1α, IL-1β,IL-1Receptor antagonist (IL-1Ra), IL-18/interferon-gamma-inducing-factor (IGIF), IL-6, IFN-γ, migration inhibitoryfactor (MIF), and two housekeeping genes L32 (ribosomeRNA) and GAPDH (glyceraldehyde-3-phosphate dehydro-genase) and mCK-3b TNF-β, lymphotoxin-β (LT-β), IL-6,IFN-γ, IFN-β, transforming growth factor β1, 2, 3 (TGF-β1,2, 3) and housekeeping genes, L32 and GAPDH) (BDPharMingen, San Diego, CA) were used to examine cytokineexpression in the tissues. The probe was labeled with 32P-UTP (800 Ci/mmol, 10 μCi/μl, PerkinElmer Life Sciences,Inc., Boston, MA). RPA were conducted using RPA III andMAXIscript kit (Ambion, Austin, TX), according to themanufacturer's instructions.
Briefly, 25 μg RNAwas hybridized overnight (16 h) withlabeled probe and the reactions were subsequently digested
with RNase A/T1 Mix, followed by precipitation withInactivation/Precipitation III solution (Ambion, Austin, TX).The protected RNAprobe complexes were resolved on 5%acrylamide/8 M urea/1×TBE gel. Then the gel was dried for2 h on a gel drier (Gel Drier, Model 583, Bio-Rad), andexposed to a Fujifilm Phosphor Imaging Plate in BAScassette 2325 (Fuji Medical Systems USA, Inc.) for 14–16 h.
The plate was scanned with a Fujifilm Bio-imagingAnalyzer BAS-1800II (Kanagawa, Japan) and the photo-stimulated luminescence (PSL) was analyzed with anImageGauge software (Version 3.12, Science Lab, FujiPhoto Film Co. Ltd). The densitometry of each band wasrepresented by the PSL reading and then normalized with ahousekeeping gene (L32 or GAPDH).
2.7. Western blot analysis
While isolating the RNA from the aqueous phase of thehomogenates using TRIzol, the remaining organic phasewas used for further protein isolation as described in themanufacturer's instructions (Invitrogen, Carlsbad, CA).Western blot analyses for TNF-α were conducted in 4–20% Tris–HCl Ready Gels (BioRad, Laboratories, Hercu-les, CA). 25 μg protein was loaded into each lane forWestern blotting. Following electrophoresis, proteins weretransferred to a nitrocellulose membrane (Hybond-C super,Amersham, Illinois, USA) on a Trans-Blot SD Semi-dryTransfer Cell and the non-specific binding sites wereblocked with 5% non-fat dry milk in Tween–Tris bufferedsaline (TTBS, 0.1% Tween-20 in 100 mM Tris–Cl [pH7.5], 0.9% NaCl). Then the membrane was incubated for2 h at room temperature with rabbit anti-mouse TNF-α polyclonal antibody (CHEMICON International, Inc.)0.15 μg/mL in TTBS or rabbit anti-GAPDH monoclonalantibody (Advanced ImmunoChemical Inc., Long Beach,CA, 1:300 in TTBS) for loading control. After threewashes with TTBS, the membrane was incubated for30 min at room temperature with biotinylated donkey anti-rabbit IgG secondary antibody (CHEMICON International,Inc., 1:10,000 dilution in TTBS) followed by three TTBSwashes. The membrane was then incubated with strepta-vidin–horseradish peroxidase conjugate (Amersham Bios-ciences, UK, 1:1000 in TTBS) and the protein antibodycompounds were revealed with a SuperSignal West PicoChemiluminescent Substrate kit (PIERCE, Rockford, IL)after four washes with TTBS and exposed to XOMAT film(Kodak, Rochester, NY). Molecular weights were deter-mined using MultiMark (Invitrogen, Carlsbad, CA).Western blots were scanned for quantification with ImageJ 1.33U (NIH, USA). The expression of TNF-α wasnormalized with GAPDH.
2.8. Plaque assays
To correlate the cytokine expression with the viral load inthe brains of infected mice, the brains were dissected and cut
51W. Mi et al. / Journal of Neuroimmunology 178 (2006) 49–61

Autho
r's
pers
onal
co
py
longitudinally along the mid line of the brain, the left andright hemispheres were alternatively collected for either RPAor viral titration by a plaque assay. Briefly, the brain wasaseptically removed, weighed, and homogenized in Dulbec-co's Modified Eagle Medium (DMEM, Invitrogen Carlsbad,CA). Cellular debris was removed by centrifugation and thetiter of virus in the supernatant was determined by standardplaque assays on L2 cells.
3. Real-time RT-PCR
We analyzed mRNA levels for IFN-γ, LT-β, and TNF-α in a subset of the brain RNA samples by real-time RT-PCR. Total RNA (1 μg) was reverse-transcribed into cDNAusing SuperScript II Reverse Transcriptase (Invitrogen,Carlsbad, CA) according to manufacturer's instructionsusing oligo(dT) or random hexamer primers. Real-time RT-PCR was performed with a GeneAmp 5700 SequenceDetection System (Perkin-Elmer, Norwalk, CT) and SYBRGreen PCR Master Mix (Applied Biosystems, Foster City,CA). Reactions were performed in MicroAmp Optical 96-Well Plates (Applied Biosystems, Foster City, CA) in avolume of 25 μL. Data were analyzed using GeneAmp 5700SDS Software (Perkin-Elmer, Norwalk, CT). For eachsample, the cytokine cycle threshold (Ct) values werenormalized against β-actin Ct values and mRNA levelswere quantitated relative to the control (non-infected, non-restrained) group using the method described by Livak andSchmittgen (2001). Amplification specificity was deter-mined using dissociation curve analysis and/or by directsequencing of the PCR product. PCR primers were designedusing Primer Express software (Applied Biosystems, FosterCity, CA). The following primers were used: 5′β-actin: GCAACG AGC GGT TCC G; 3′β- actin: TCC AGC CTT CCTTCT TGG G; 5′IFN-γ: CAG CAA CAG CAA GGC GAAA; 3′IFN-γ: CTG GAC CTG TGG GTT GTT GAC; 5′LT-β:GAG AGG GTC TAC GTT AAC ATC AGT CA; 3′ LT-β:CGC CCC GAA GAA GGT CTT; 5′TNF-α: TGA TCCGCG ACG TGG AA; 3′TNF-α: CCG CCT GGA GTT CTGGAA.
3.1. Statistical analysis
A 2-way ANOVA was conducted entering restraint andinfection as between subject variables. Duncan's multiplerange tests and means comparisons were used for post hocanalyses. A p value of 0.05 or less was considered significantin all cases.
4. Results
4.1. Cytokine expression in the brain following Theiler'svirus infection and restraint stress
To assess the potential effect of restraint stress on theimmune response in the CNS following Theiler's virus
infection, brains were collected at days 2 and 7 p.i. and themRNA cytokine levels were determined by RNase protec-tion assay (Figs. 1 and 2). A series of ANOVAs wereconducted for each cytokine to determine the effect ofinfection, restraint and the interaction between the twovariables at the different time points.
As summarized in Table 1, at day 2 p.i., infection inducedmRNA cytokine elevation of LT-β, TNF-α, IFN-γ, and IL-6all F's(1,30)>4.200, p's<0.05. Although not statisticallysignificant, IL-12p40 was marginally elevated due toinfection (p=0.07). At day 7 p.i., a main effect of infectionwas observed for LT-β, TNF-α, IFN-γ, IFN-β, IL- 1α, IL-1β, IL-6, IL-12p35, and IL-12p40 all F's(1,52)>4.016,p's<0.05 (Table 1).
A smaller number of cytokines were affected by restraintstress than by infection. Restraint increased mRNA levels forTNF-β, but decreased LT-β in the brain at day 2 p.i., all F's(1,30)>6.196, p's<0.05. At day 7 p.i., restraint increasedmRNA for TNF-β, IFN-β, and IL-1α, all F's(1,52)>3.959,p's<0.05. (Table 1).
As depicted in Fig. 2 and Table 1, an interaction betweeninfection and restraint was observed, with restraint signifi-cantly decreasing the infection-induced increase of IFN-γ atday 2 p.i. (F(1,30)=4.664, p<0.05). Although the maineffect of restraint on IFN-β was not significant, post hoc
Fig. 1. Phosphor image of a representative RPA for cytokine mRNA in thebrain following restraint or/and BeAn infection at day 7 post infection using(A) mCK-2b and (B) mCK-3b kits. Total RNAwas isolated from the brainsand spleens of CBA mice at day 7 p.i. 25 μg RNAwas hybridized with a32P-labeled chemokine probe set as described in Materials and methods. Thehousekeeping genes GAPDH was used for the normalization of sampleloading and technique error. Lanes 1, 2, 3, 4 represent the brain samples ofNI/NR (Non-infected/Non-restrained), NI/R (Non-infected/Restrained), I/NR (Infected/Non-restrained) and I/R (Infected/Restrained), respectively,collected at day 7 post infection.
52 W. Mi et al. / Journal of Neuroimmunology 178 (2006) 49–61

Autho
r's
pers
onal
co
py
analysis revealed higher levels in the I/R mice than I/NRmice at day 7 p.i., p<0.05.
We used real-time RT-PCR to confirm cytokine RNAexpression levels for TNF-α, LT-β and IFN-γ, using a subset(N=15) of the RNA samples from brains at day 7 p.i. (Fig.5). In the brain, PCR confirmed that infection with Theiler'svirus increased production of TNF-α, LT-β and IFN-γ (allF's(1,13)>21.451, p's<0.001.). A statistically significantinteraction also was observed, such that restraint increasedLT-β in non-infected mice, but decreased it in infected mice,F(1,13)=12.493, p<0.005.
4.2. Cytokine expression in the spleen following Theiler'svirus infection and restraint stress
To further examine the effect of restraint stress on theperipheral immune response following infection, we evalu-ated the mRNA cytokine levels in the spleen using RPA atdays 2 and 7 p.i. (Figs. 3 and 4 and summarized in Table 2).
ANOVA revealed a significant main effect of infection oncytokine production, such that Theiler's virus inducedincreases in mRNA for TNF-α, TNF-β, IFN-γ, IFN-β andIL-12p40 at day 2 p.i. (all F's (1,30)>4.214, p's<0.05) andLT-β, IL-1β, IL-12p40, and IL- 18 at day 7 p.i. (all F's(1,46)>6.026, p's<0.05 ) (Table 2).
A notable effect of stress on the cytokine expression in thespleen was observed at day 2 p.i., demonstrating thatrestraint resulted in reductions in mRNA levels of TNF-α,TNF-β, LT-β, IFN-γ, IFN-β, and IL-6, and an increase in IL-18, all F's(1,30)>7.252, p's<0.01. In contrast, restraintproduced a less remarkable suppressive effect of stress on
Fig. 2. The effect of restraint stress on cytokine mRNA levels in the brainduring acute Theiler's virus infection. Using RPA, the levels of cytokinemRNAwere derived from the densitometric ratio of the individual cytokineto the housekeeping gene GAPDH and multiplied by 1000. At day 2 p.i.,IFN-γ, LT-β, IL-12p40, IL-6 and IFN-β were increased by TMEV infection,and the expression of IFN-γ, LT-β, IL-12p40 were decreased followingrestraint stress. The cytokines remained at an elevated level at day 7 p.i., andrestraint stress reduced the expression of IFN-γ, LT-β, IL-12p40 and IL-6,however, IFN-β was increased by stress. The values were represented bymean±S.E.M.
Fig. 2 (continued).
53W. Mi et al. / Journal of Neuroimmunology 178 (2006) 49–61

Autho
r's
pers
onal
co
py
cytokine expression at day 7 p.i. At this time point only adown-regulation of IFN-β and an up-regulation of IL-18 bystress was detected, F(1,30)=4.482, p<0.05. In addition,stress also elevated a Th2 cytokine, IL-10, in the spleen atday 2 p.i., F(1, 28)=5.091, p's<0.05, but this increase didnot reach significance at day 7 p.i., p=0.10 (Fig. 4).
Restraint decreased the virus-induced mRNA cytokineexpression of TNF-α at day 2 p.i., F(1,30)=4.755, p<0.05(Fig. 5). Interestingly, by day 7 p.i., the interaction wasreversed, such that restraint increased TNF-α expression ininfected mice, but decreased it in non-infected mice F(1,51)=4.574, p<0.05. The same pattern was observed for TNF-β at day 7 p.i., F(1,51)=4.185, p<0.05. These resultssuggest that stress exerts a greater effect on the peripheralimmune function by down-regulating pro-inflammatorycytokines during the earlier stage than later stage inrestrained animals (Table 2).
4.3. The expression of TNF-α protein level in the brain byWestern blotting
TNF-α is an essential cytokine involved in the innate anti-viral response, and our previous study showed that serumTNF-α was elevated in all restrained animals at days 1 and 2p.i., irrespective of infection status (Welsh et al., 2004). Inthe current study, we further examined the TNF-α proteinlevels using Western blotting.
A main effect of infection was observed on the TNF-α protein level in the brain, F(1,12)=5.958, p<0.04. Aninteraction between infection and restraint did not reachsignificance; however, restraint stress significantly decreasedthe TNF-α in the brain of infected mice, as confirmed by posthoc mean comparison (Fig. 6). The result indicated a
difference in TNF-α response in the circulation (systemic)and the CNS (local) following Theiler's virus infection andrestraint stress.
4.4. The correlation of pro-inflammatory cytokines produc-tion and viral clearance in the brain
Pro-inflammatory cytokines play an important role inboth viral clearance and activation of the immune response toviral infection. To determine whether the level of cytokinescorrelates with the level of virus, we set up a separateexperiment, which consisted of two groups: infected/non-restrained and infected/restrained groups. This allowed us toexamine the cytokine expression and the viral titers in thebrain from the same animal. At day 7 p.i., the mice wereterminated and the brain was dissected out and cut into twohalves along the mid sagittal plane. Then we alternativelytook the left or right half of the brain for assaying ofcytokines by RPA and the remaining half of the brain fordetermination of viral titers by a plaque assay.
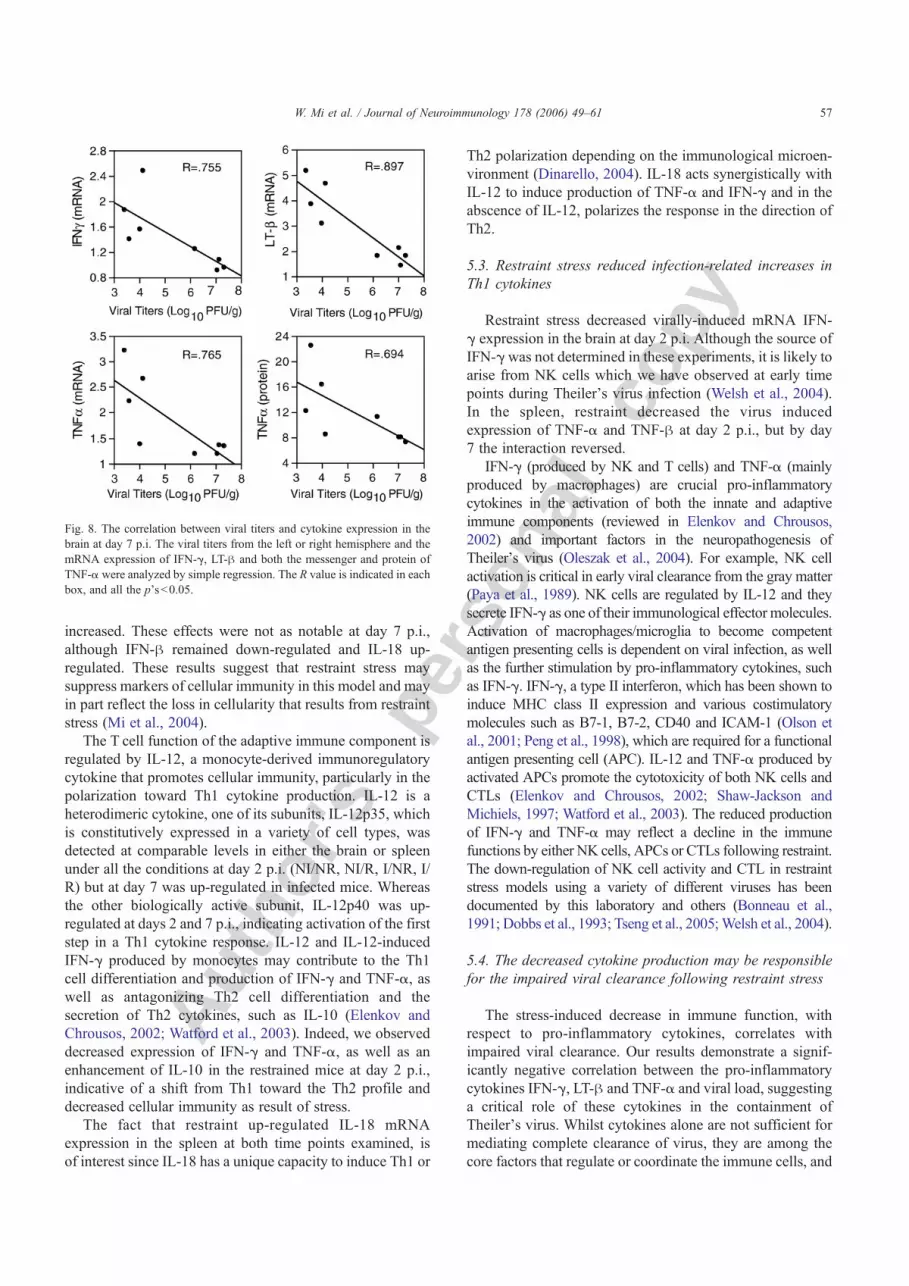
Restraint appeared to impair the viral clearance in thebrain at day 7 p.i., resulting in higher viral titers in therestrained mice (p=0.001 ) as depicted in Fig. 7. Individualcytokine levels were correlated with viral titers by simpleregression. The mRNA levels for the pro-inflammatorycytokines LT-β, TNF-α and IFN-γ were negatively corre-lated with viral titers in the brain, wherein a higher viral level
Table 1Cytokine expression in the brain following restraint and Theiler's virusinfection
Braincytokines
Day 2 post infection Day 7 post infection
Infection RS Inf+RS Infection RS Inf+RS
IL-1α + +IL-1β +IL-6 + (+)IL-10IL-12p35 +IL-12p40 (+) +IL-18/IGIFIFN-β see text + +IFN-γ + see text +Lt-β + − +TNF-α + +TNF-β + +
A summary of the alterations in cytokine mRNA levels in the brainfollowing Theiler's virus infection and restraint stress. The results are shownas the main effect of either infection (INF) or restraint stress (RS), with +/−representing an increase or a decrease in cytokine mRNA in the brain,respectively. Results shown in parentheses indicate trends that were notstatistically significant (p<0.10). Interactions between the two variables aredescribed in the text.
Fig. 3. Phosphor image of a representative RPA for cytokine mRNA in thespleen following restraint and/or BeAn infection at day 7 post infectionusing (A) mCK-2b and (B) mCK-3b kits. Total RNAwas isolated from thebrains and spleens of CBA mice at day 7 p.i. 25 μg RNA was hybridizedwith a 32P-labeled chemokine probe set as described in Materials andmethods. The housekeeping gene L32 was used for the normalization ofsample loading and technique error. Lanes 1, 2, 3, 4 represent the brainsamples of NI/NR (Non-infected/Non-restrained), NI/R (Non-infected/Restrained), I/NR (Infected/Nonrestrained) and I/R (Infected/Restrained),respectively, collected at day 7 p.i.
54 W. Mi et al. / Journal of Neuroimmunology 178 (2006) 49–61

Autho
r's
pers
onal
co
py
consistently correlated with a lower cytokine expression, allp's<0.05. In addition, there was a marginal correlationbetween the TNF-α protein level and the viral titers in thebrain, wherein an increase in TNF-α was associated with alower viral titers in the brain, p=0.056, R=0.694 (Fig. 8).These results indicate the potential role of pro-inflammatorycytokines in viral clearance.
5. Discussion
The current study is a continuation of our investigation ofthe effects of restraint stress on the neuropathogenesis ofTheiler's virus infection. Experimental infection with Thei-
ler's virus in the brain elicits both innate and adaptiveimmunity, which are characterized by CNS mononuclearinfiltration consisting of NK cells, macrophages, virus-specific CD4 + and CD8 + T cells, as well as activation ofthe CNS residential glia. The temporal mRNA cytokineanalysis conducted in this study provides insights into theeffects of restraint stress on both the central and peripheralimmune response to Theiler's virus infection. However, weonly examined whole brain and spleen preparations and, withthe exception of TNF-α, only mRNA cytokine levels and theresults should be viewed with these caveats in mind.Comparable results were obtained using RPA and real-timeRT-PCR for RNA expression levels of TNF-α, LT-β and IFN-γ in the brain.
5.1. Infection with Theiler's virus results in expression ofpro-inflammatory cytokines in the brain and spleen
The pro-inflammatory cytokines: TNF-α, LT-β, IFN-γ and IL-6 were induced in the brain as early as day 2 p.i. and
Fig. 4. The effect of restraint stress on cytokine mRNA levels in the spleenduring acute Theiler's virus infection. Using RPA, the levels of cytokinemRNAwere derived from the densitometric ratio of the individual cytokineto the housekeeping gene L32 and multiplied by 1000. At day 2 p.i., restraintstress significantly decreased Theiler's virus-induced expression of proin-flammatory cytokines TNF-α, TNF-β, LT-β, IFN-β, IFN-γ, IL-6 and IL-12p40; however, an anti-inflammatory cytokine, IL-10 declined followingrestraint stress. The values were represented by mean±S.E.M.
Fig. 4 (continued).
55W. Mi et al. / Journal of Neuroimmunology 178 (2006) 49–61

Autho
r's
pers
onal
co
py
continued to be expressed at day 7 p.i., indicative of immuneactivation of macrophage/microglial cells and NK cellswithin the CNS. By day 7 p.i. the panel of cytokines induced
by infection, increased to include: IFN-β, IL-1α, IL1-β, IL-12p35, IL-12p40, LT-β, TNF-α, IFN-γ. In SJL mice asimilar pattern of cytokine profiles were seen during earlyinfection with certain notable differences (Chang et al.,2000). SJL mice developed high levels of IL-10 and TGF-β that we did not observe in CBA mice. High levels of TGF-β have been suggested to account in part for the delayedappearance and activity of cytotoxic T cells into the brain ofSJL mice (Oleszak et al., 2004).
In the spleen, Theiler's virus infection induced a similarprofile of pro-inflammatory cytokines: TNF-α, TNF-β,IFN-γ, IFN-β, IL-12p40 and IL-6. However, fewercytokines were expressed in the spleen at day 7 p.i.compared to day 2 p.i.
5.2. Restraint stress alone reduces more cytokines in thespleen than the brain
A reduction in the constitutive expression of LT-β byrestraint was documented in the brain at day 2 p.i. and anincrease in TNF-β. By day 7 p.i. restraint increased TNF-β,IFN-β and IL-1α in the brain. As compared to the brain, agreater number of cytokines in the spleen, TNF-α, TNF-β,IL-6, IFN-γ, IFN-β, and LT-β, were profoundly reduced byrestraint at day 2 p.i., whereas IL-18 and IL-10 were
Table 2Cytokine expression in the spleen following restraint and Theiler's virusinfection
Spleencytokines
Day 2 post infection Day 7 post infection
Infection RS Inf+RS Infection RS Inf+RS
IL-1αIL-1β +IL-6 − (−)IL-10 +IL-12p35IL-12p40 + +IL-18/IGIF + + +IFN-β + − −IFN-γ + −Lt-β − +TNF-α + − see text see textTNF-β + − see text
A summary of the alterations in cytokine mRNA levels in the spleenfollowing Theiler's virus infection and restraint stress. The results are shownas the main effect of either infection (INF) or restraint stress (RS), with +/−representing an increase or a decrease in cytokine mRNA in the spleen,respectively. Results shown in parentheses indicate trends that were notstatistically significant (p<0.10). Interactions between the two variables aredescribed in the text.
Fig. 5. The effect of restraint stress on relative mRNA levels for IFN-γ, LT-β, and TNF-α in the brain at day 7 p.i. measured by real-time RT-PCR.Levels of cytokine mRNAwere normalized to β-actin mRNA, and for eachexperimental group are presented relative to the control (non-infected, non-restrained) group. This data confirmed that infection with Theiler's virusincreased mRNA levels for all three cytokines. The values are represented bymean±S.E.M.
Fig. 6. The protein TNF-α level in the brain at day 7 p.i. by Western blot.TNF-α protein expression was increased by Theiler's virus infection, anddecreased by restraint stress. The TNF-α level was derived from thedensitometric ratio of TNF-α and GAPDH. The value was represented bymean±S.E.M.
Fig. 7. The impact of restraint stress on viral titers in the brain at day 7 p.i. byplaque assay. The viral titer was calculated by the logarithm of plaqueforming assay (PFU) per gram of brain tissue. The values were representedby mean±S.E.M. (*) indicates p<0.0001.
56 W. Mi et al. / Journal of Neuroimmunology 178 (2006) 49–61
Autho
r's
pers
onal
co
pyincreased. These effects were not as notable at day 7 p.i.,although IFN-β remained down-regulated and IL-18 up-regulated. These results suggest that restraint stress maysuppress markers of cellular immunity in this model and mayin part reflect the loss in cellularity that results from restraintstress (Mi et al., 2004).
The T cell function of the adaptive immune component isregulated by IL-12, a monocyte-derived immunoregulatorycytokine that promotes cellular immunity, particularly in thepolarization toward Th1 cytokine production. IL-12 is aheterodimeric cytokine, one of its subunits, IL-12p35, whichis constitutively expressed in a variety of cell types, wasdetected at comparable levels in either the brain or spleenunder all the conditions at day 2 p.i. (NI/NR, NI/R, I/NR, I/R) but at day 7 was up-regulated in infected mice. Whereasthe other biologically active subunit, IL-12p40 was up-regulated at days 2 and 7 p.i., indicating activation of the firststep in a Th1 cytokine response. IL-12 and IL-12-inducedIFN-γ produced by monocytes may contribute to the Th1cell differentiation and production of IFN-γ and TNF-α, aswell as antagonizing Th2 cell differentiation and thesecretion of Th2 cytokines, such as IL-10 (Elenkov andChrousos, 2002; Watford et al., 2003). Indeed, we observeddecreased expression of IFN-γ and TNF-α, as well as anenhancement of IL-10 in the restrained mice at day 2 p.i.,indicative of a shift from Th1 toward the Th2 profile anddecreased cellular immunity as result of stress.
The fact that restraint up-regulated IL-18 mRNAexpression in the spleen at both time points examined, isof interest since IL-18 has a unique capacity to induce Th1 or
Th2 polarization depending on the immunological microen-vironment (Dinarello, 2004). IL-18 acts synergistically withIL-12 to induce production of TNF-α and IFN-γ and in theabscence of IL-12, polarizes the response in the direction ofTh2.
5.3. Restraint stress reduced infection-related increases inTh1 cytokines
Restraint stress decreased virally-induced mRNA IFN-γ expression in the brain at day 2 p.i. Although the source ofIFN-γwas not determined in these experiments, it is likely toarise from NK cells which we have observed at early timepoints during Theiler's virus infection (Welsh et al., 2004).In the spleen, restraint decreased the virus inducedexpression of TNF-α and TNF-β at day 2 p.i., but by day7 the interaction reversed.
IFN-γ (produced by NK and T cells) and TNF-α (mainlyproduced by macrophages) are crucial pro-inflammatorycytokines in the activation of both the innate and adaptiveimmune components (reviewed in Elenkov and Chrousos,2002) and important factors in the neuropathogenesis ofTheiler's virus (Oleszak et al., 2004). For example, NK cellactivation is critical in early viral clearance from the gray matter(Paya et al., 1989). NK cells are regulated by IL-12 and theysecrete IFN-γ as one of their immunological effector molecules.Activation of macrophages/microglia to become competentantigen presenting cells is dependent on viral infection, as wellas the further stimulation by pro-inflammatory cytokines, suchas IFN-γ. IFN-γ, a type II interferon, which has been shown toinduce MHC class II expression and various costimulatorymolecules such as B7-1, B7-2, CD40 and ICAM-1 (Olson etal., 2001; Peng et al., 1998), which are required for a functionalantigen presenting cell (APC). IL-12 and TNF-α produced byactivated APCs promote the cytotoxicity of both NK cells andCTLs (Elenkov and Chrousos, 2002; Shaw-Jackson andMichiels, 1997; Watford et al., 2003). The reduced productionof IFN-γ and TNF-α may reflect a decline in the immunefunctions by either NK cells, APCs or CTLs following restraint.The down-regulation of NK cell activity and CTL in restraintstress models using a variety of different viruses has beendocumented by this laboratory and others (Bonneau et al.,1991; Dobbs et al., 1993; Tseng et al., 2005;Welsh et al., 2004).
5.4. The decreased cytokine production may be responsiblefor the impaired viral clearance following restraint stress
The stress-induced decrease in immune function, withrespect to pro-inflammatory cytokines, correlates withimpaired viral clearance. Our results demonstrate a signif-icantly negative correlation between the pro-inflammatorycytokines IFN-γ, LT-β and TNF-α and viral load, suggestinga critical role of these cytokines in the containment ofTheiler's virus. Whilst cytokines alone are not sufficient formediating complete clearance of virus, they are among thecore factors that regulate or coordinate the immune cells, and
Fig. 8. The correlation between viral titers and cytokine expression in thebrain at day 7 p.i. The viral titers from the left or right hemisphere and themRNA expression of IFN-γ, LT-β and both the messenger and protein ofTNF-α were analyzed by simple regression. The R value is indicated in eachbox, and all the p's<0.05.
57W. Mi et al. / Journal of Neuroimmunology 178 (2006) 49–61

Autho
r's
pers
onal
co
py
changes in their level may be indicative of the degree ofactivation of the immune system.
IFN-γ is an important immune mediator for viralclearance in the CNS by inducing type II nitric oxidesynthase (iNOS) from glia and type I nitric oxide synthase(nNOS) from neurons. IFN-γ has been shown to be criticallyimportant in the pathogenesis of several different viralinfections, such as measles, herpes simplex virus (reviewedin Chesler and Reiss, 2002). The crucial role of IFN-γ inTheiler's virus infection has been demonstrated previously,whereby the administration of monoclonal antibody to IFN-γ increases viral persistence and demyelination in bothsusceptible SJL/J and resistant C57BL/10NJ strains of mice(Rodriguez et al., 1995). In addition, persistent viralinfection and demyelination has been recorded in C57BL/6IFN-γ −/− knockout mice, but not in wild type mice (Murrayet al., 2002). Furthermore, IFN-γ was shown to have potentanti-viral activity in vitro in cerebrovascular endothelial cellsinfected with Theiler's virus (Welsh et al., 1995).
TNF-α confers anti-viral activity, especially whensynergized with other cytokines, such as IFNs (Ito andO'Malley, 1987; Wong and Goeddel, 1986). The effect ofTNF-α in viral infection of the CNS may be mediated by theup-regulation of TNF-α receptor, as reported in an in vitrostudy using mouse hepatitis virus (MHV)-JHM strain(Rempel et al., 2005). In the Theiler's virus model, TNF-α has anti-viral activity in vitro (Welsh et al., 1995). TNF-α has also been shown to be toxic for oligodendrocytes andis consequently implicated in demyelination (Selmaj andRaine, 1988). In the acute infection, TNF-α and IFN-γ areproduced by CTL, which mediate the non-cytotoxicmechanisms, suggesting a potential role for TNF-α in theelimination of virus (Lyman et al., 2004).
LT-β, a membrane form of lymphotoxin expressed onactivated T, B and NK cells, plays a critical role in theresistance to intracellular pathogens, including viruses(Benedict et al., 2001; Lin et al., 2003; Marshall et al.,1999; Soderberg et al., 2004), bacteria (Ehlers et al., 2003)and prions (Mabbott et al., 2003; Oldstone et al., 2002). LT-β induces anti-viral activity to Theiler's virus infection bymounting effective CTL activity enabling viral clearanceduring the early stage, thus diminishing demyelination at thelater stage. Compared to the soluble LT-α, LT-β plays aminor role in demyelination (Lin et al., 2003).
The decreased expression of pro-inflammatory cytokinesinduced by stress may reflect decreased activation ofimmune cells and/or CNS glia or alternatively the decreasein recruitment of immune cells to the CNS. It has been shownthat stress may alter the cellular composition in circulationand redistribution of immune cells in lymphoid organs usinga variety of experimental stressors, such as restraint,immobilization and social stress (Dhabhar et al., 1995;Fleshner et al., 1995; Stefanski et al., 2003). This effect wasalso observed in the Theiler's virus restraint model by ourlaboratory, demonstrating that restraint leads to a decreasednumber of circulating lymphocytes (Welsh et al., 2004) and
decreased inflammation in the CNS (Campbell et al., 2001;Mi et al., 2004). Additionally, we previously showed thatstress results in decreased expression of the chemokines, Ltn,IP-10 and RANTES, which are potential chemoattractantsfor both NK cell and T cells (Mi et al., 2004). Therefore, thereduction in cellularity, as well as the decline in recruitmentto the inflamed sites following restraint stress may beadditional reasons for the reduction in cytokine expression inthe brain or spleen.
5.5. Restraint stress increased the mRNA levels of IFN-β ininfected mice
An increase of IFN-β, a type I IFN, was observed in thebrain and spleen following Theiler's virus infection. IFN-β is a potential anti-viral cytokine and an inducer of NKcell-mediated cytolysis (Biron, 1998). However, it has alsobeen used as a therapeutic agent for both MS andexperimental autoimmune encephalomyelitis (EAE)—ananimal model of MS. The mode of action of IFN-β ismediated in part by suppressing both IL-12 production anddownstream Th1 cytokines (Klimstra et al., 2000). Unlikethe pro-inflammatory cytokines, the level of IFN-β waselevated in the brain of I/R mice. Restraint-inducedincreases in type I IFNs have also been documented inother viral infections, such as influenza virus (Hunzeker etal., 2004). The rise of IFN-β was correlated with impairedviral clearance brought upon by stress at day 7 p.i., whichmay possibly be explained by a compensatory response todecreased levels of anti-viral IFN-γ and other pro-inflammatory cytokines. Since restraint stressed mice haveincreased levels of virus in the CNS, this may act as anincreased stimulus for the production of IFN-β.
5.6. Alteration in cytokine expression is mediated by bothhormonal and neuronal mechanisms
It is well recognized that glucocorticoids regulate theTh1/Th2 balance by suppressing IL-12, thus removing itsinhibitory effect on the production of Th2 cytokines, such asIL-10 (Elenkov and Chrousos, 2002). Although we did notobserve a restraint-induced decrease in IL-12, we did noteelevated IL-10 in restrained mice, which may be explainedby the restraint-induced elevation of circulating corticoste-rone as reported previously (Campbell et al., 2001; Sieve etal., 2004). Glucocorticoids are known to induce IL-10production, as well as other Th2 cytokines, IL-4 and IL-13 inrat CD4 + T cells (Ramierz et al., 1996). T regulatory cells(TR 1 cell), have been shown to be a potential source of IL-10 following glucocorticoid exposure, which is mediated byup-regulation of a different transcription factor, FOXP3(Karagiannidis et al., 2004). Moreover, glucocorticoid canpromote IL-10 production by both macrophages anddendritic cells (Franchimont, 2004); thus, together with theTR 1 cell, these cells may be the major source of IL-10 in thestress response. However, these results do not eliminate the
58 W. Mi et al. / Journal of Neuroimmunology 178 (2006) 49–61

Autho
r's
pers
onal
co
py
possibility that restraint-induced catacholamines (CAs) mayalso be involved in this Th1/Th2 regulation, in addition to thehypothesis of peripheral-derived immune regulation. Be-cause the spleen is predominantly innervated by sympatheticnerve fibers, activation of the SNS results in the release ofnorepinepherine in either a synaptic or a nonsynapticmanner in the spleen (Elenkov et al., 2000), for example,during restraint stress. Catecholamines inhibit IL-12 andsubsequent Th1 cytokine production, but stimulate IL-10production via β2-adrenergic receptors, thus providingadditional machinery for the Th1/Th2 transition (Elenkovand Chrousos, 2002).
5.7. Differential immunoregulatory mechanism may beinvolved in the CNS and peripheral immune organs duringstress
Generally, our data indicate similarities between cytokineprofiles in the brain and the spleen following infection, witha greater number of Th1 cytokines detected in the brain. Thiscould be due to several mechanisms, including either morepotential cellular sources for cytokine production in thebrain, such as astrocytes, microglia, infiltrating inflammatorycells; or a higher viral load in the CNS than in the spleen (Miet al., in press).
A notable discrepancy between the immune response inthe brain and spleen is demonstrated by the differentialexpression of IL-10 in the two organs. The increase in IL-10caused by stress was only found in the spleen at day 2 p.i.,and not at day 7 p.i. The elevation of IL-10 appears to beassociated with the decline in Th1 cytokines—in the spleen.An IL-10 response was never detected in the brain. However,the down-regulation of Th1 cytokines were present up to day7 p.i. in the brain. It is plausible that two mechanisms mayaccount for the reduced Th1 response following stress; eitherby reducing the expression of the upstream cytokine, IL-12p40, or by producing the Th1 inhibitory cytokine, IL-10.However, the skew in Th2 response was only noticed in thespleen at day 2 p.i., suggesting the inhibitive role of Th2cytokine on Th1 cytokine production was transient in thisrestraint model. The absence of Th2 response in the brain ateither time point examined may suggest an additionalmechanism that a direct down-regulation of Th1 cytokines,rather than inhibition by Th2 cytokines, plays a major role inthe stress-induced immune response to Theiler's virusinfection. We speculate that a regulatory effect by Th2cytokines during restraint stress may occur in the periphery.
In summary, restraint stress of mice infected withTheiler's virus, produced decreased pro-inflammatory cyto-kine expression in the brain and spleen, as compared to thenon-restrained mice, and caused an impaired viral clearancewhich may account for subsequent prolonged or persistentviral infection and enhanced demyelination (Sieve et al.,2004). The stress-induced regulatory mechanism in viralinfection and the cell type involved in the process warrantfurther investigation.
Acknowledgments
This research was funded by grants to C.J.R.W. and M.W.M. from the National Multiple Sclerosis Society RG 3128and NIH/NINDS R01 39569, and supported, in part by aTexas A&M University LTSF Training Grant to W.M.Wentao Mi performed this work in partial fulfillment of therequirements for the PhD degree at Texas A&M University,College Station, Texas. The authors would also like to thankDr. Paul Spencer and Dr. Rick Metz for advice on real-timeRT-PCR.
References
Aubagnac, S., Brahic, M., Bureau, J.F., 1999. Viral load and a locus onchromosome 11 affect the late clinical disease caused by Theiler's virus.J. Virol. 73, 7965–7971.
Begolka, W.S., Vanderlugt, C.L., Rahbe, S.M., Miller, S.D., 1998.Differential expression of inflammatory cytokines parallels progressionof central nervous system pathology in two clinically distinct models ofmultiple sclerosis. J. Immunol. 161, 4437–4446.
Benedict, C.A., Banks, T.A., Senderowicz, L., Ko, M., Britt, W.J., Angulo,A., Ghazal, P., Ware, C.F., 2001. Lymphotoxins and cytomegaloviruscooperatively induce interferon-beta, establishing host-virus detente.Immunity 15, 617–626.
Biron, C.A., 1998. Role of early cytokines, including alpha and betainterferons (IFNalpha/ beta), in innate and adaptive immune responses toviral infections. Semin. Immunol. 10, 383–390.
Bonneau, R.H., Sheridan, J.F., Feng, N.G., Glaser, R., 1991. Stress-inducedsuppression of herpes simplex virus (HSV)-specific cytotoxic Tlymphocyte and natural killer cell activity and enhancement of acutepathogenesis following local HSV infection. Brain Behav. Immun. 5,170–192.
Borrow, P., Welsh, C.J., Nash, A.A., 1993. Study of the mechanisms bywhich CD4+T cells contribute to protection in Theiler's murineencephalomyelitis. Immunology 80, 502–506.
Borrow, P., Welsh, C.J., Dean, D., Tonks, P., Blakemore, W.F., Nash, A.A.,1998. Investigation of the role of autoimmune responses to myelin in thepathogenesis of TMEV-induced demyelinating disease. Immunology 93,478–484.
Brahic, M., Bureau, J.F., 1998. Genetics of susceptibility to Theiler's virusinfection. BioEssays 20, 627–633.
Campbell, T., Meagher, M.W., Sieve, A., Scott, B., Storts, R., Welsh, T.H.,Welsh, C.J., 2001. The effects of restraint stress on the neuropathogen-esis of Theiler's virus infection: I. Acute disease. Brain Behav. Immun.15, 235–254.
Chang, J.R., Zaczynska, E., Katsetos, C.D., Platsoucas, C.D., Oleszak, E.L.,2000. Differential expression of TGF-beta, IL-2, and other cytokines inthe CNS of Theiler's murine encephalomyelitis virus-infected suscep-tible and resistant strains of mice. Virology 278, 346–360.
Chesler, D.A., Reiss, C.S., 2002. The role of IFN-gamma in immuneresponses to viral infections of the central nervous system. CytokineGrowth Factor Rev. 13, 441–454.
Dal Canto, M.C., Calenoff, M.A., Miller, S.D., Vanderlugt, C.L., 2000.Lymphocytes from mice chronically infected with Theiler's murineencephalomyelitis virus produce demyelination of organotypic culturesafter stimulation with the major encephalitic epitope of myelinproteolipid protein. Epitope spreading in TMEV infection has functionalactivity. J. Neuroimmunol. 104, 79–84.
Dinarello, C.A., 2004. Interleukin-18 and the treatment of rheumatoidarthritis. Rheum. Dis. Clin. North Am. 30, 417–434.
Dhabhar, F.S., Miller, A.H., McEwen, B.S., Spencer, R.L., 1995. Effects ofstress on immune cell distribution. Dynamics and hormonal mechan-isms. J. Immunol. 154, 5511–5527.
59W. Mi et al. / Journal of Neuroimmunology 178 (2006) 49–61

Autho
r's
pers
onal
co
py
Dobbs, C.M., Vasquez, M., Glaser, R., Sheridan, J.F., 1993. Mechan-isms of stress-induced modulation of viral pathogenesis andimmunity. J. Neuroimmunol. 48, 151–160.
Drescher, K.M., Murray, P.D., Lin, X., Carlino, J.A., Rodriguez, M.,2000. TGF-beta 2 reduces demyelination, virus antigen expression,and macrophage recruitment in a viral model of multiple sclerosis.J. Immunol. 164, 3207–3213.
Ehlers, S., Holscher, C., Scheu, S., Tertilt, C., Hehlgans, T., Suwinski, J.,Endres, R., Pfeffer, K., 2003. The lymphotoxin beta receptor is criticallyinvolved in controlling infections with the intracellular pathogensMycobacterium tuberculosis and Listeria monocytogenes. J. Immunol.170, 5210–5218.
Elenkov, I.J., Chrousos, G.P., 2002. Stress hormones, proinflammatory andantiinflammatory cytokines, and autoimmunity. Ann. N.Y. Acad. Sci.966, 290–303.
Elenkov, I.J., Wilder, R.L., Chrousos, G.P., Vizi, E.S., 2000. Thesympathetic nerve-an integrative interface between two supersystems:the brain and the immune system. Pharmacol. Rev. 52, 595–638.
Fleshner, M., Hermann, J., Lockwood, L.L., Laudenslager, M.L., Watkins,L.R., Maier, S.F., 1995. Stressed rats fail to expand the CD45RC+CD4+(Th1-like) T cell subset in response to KLH: possible involvement ofIFN-gamma. Brain Behav. Immun. 9, 101–112.
Franchimont, D., 2004. Overview of the actions of glucocorticoids on theimmune response: a good model to characterize new pathways ofimmunosuppression for new treatment strategies. Ann. N. Y. Acad. Sci.1024, 124–137.
Gerety, S.J., Karpus, W.J., Cubbon, A.R., Goswami, R.G., Rundell, M.K.,Peterson, J.D., Miller, S.D., 1994. Class II-restricted T cell responses inTheiler's murine encephalomyelitis virus-induced demyelinating dis-ease. V. Mapping of a dominant immunopathologic VP2 Tcell epitope insusceptible SJL/J mice. J. Immunol. 152, 908–918.
Gilden, D.H., Lipton, H.L., 2005. Infectious causes of multiple sclerosis.Lancet Neurol. 4, 195–202.
Haddad, J.J., Saade, N.E., Safieh-Garabedian, B., 2002. Cytokines andneuro-immune-endocrine interactions: a role for the hypothalamic-pituitary-adrenal revolving axis. J. Neuroimmunol. 133 (1-2), 1–19(Dec, Review).
Hunzeker, J., Padgett, D.A., Sheridan, P.A., Dhabhar, F.S., Sheridan, J.F.,2004. Modulation of natural killer cell activity by restraint stress duringan influenza A/PR8 infection in mice. Brain Behav. Immun. 18,526–535.
Ito, M., O'Malley, J.A., 1987. Anti-viral effects of recombinant humantumor necrosis factor. Lymphokine Res. 6, 309–318.
Karagiannidis, C., Akdis, M., Holopainen, P., Woolley, N.J., Hense, G.,Ruckert, B., Mantel, P.Y., Menz, G., Akdis, C.A., Blaser, K., Schmidt-Weber, C.B., 2004. Glucocorticoids up-regulate FOXP3 expressionand regulatory T cells in asthma. J. Allergy Clin. Immunol. 114,1425–1433.
Klimstra, W.B., Nguyen, K.B., Biron, C.A., Johnston, R.E., Karp, C.L.,2000. Interferon beta in multiple sclerosis: is IL-12 suppression the key?J. Virol. 74, 3366–3378.
Lin, X., Ma, X., Rodriguez, M., Feng, X., Zoecklein, L., Fu, Y.X., Roos,R.P., 2003. Membrane lymphotoxin is required for resistance toTheiler's virus infection. Int. Immunol. 15, 955–962.
Lipton, H.L., 1975. Theiler's virus infection in mice: an unusual biphasicdisease process leading to demyelination. Infect. Immun. 11,1147–1155.
Lipton, H.L., Kratochvil, J., Sethi, P., Dal Canto, M.C., 1984. Theiler's virusantigen detected in mouse spinal cord 2 1/2 years after infection.Neurology 34, 1117–1119.
Livak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expressiondata using real-time quantitative PCR and the 2(-Delta Delta C(T))method. Methods 25, 402–408.
Lyman, M.A., Myoung, J., Mohindru, M., Kim, B.S., 2004. Quantitative,not qualitative, differences in CD8(+) T cell responses to Theiler'smurine encephalomyelitis virus between resistant C57BL/6 andsusceptible SJL/J mice. Eur. J. Immunol. 34, 2730–2739.
Mabbott, N.A., Young, J., McConnell, I., Bruce, M.E., 2003.Follicular dendritic cell dedifferentiation by treatment with aninhibitor of the lymphotoxin pathway dramatically reduces scrapiesusceptibility. J. Virol. 77, 6845–6854.
Marshall, W.L., Brinkman, B.M., Ambrose, C.M., Pesavento, P.A.,Uglialoro, A.M., Teng, E., Finberg, R.W., Browning, J.L., Goldfeld,A.E., 1999. Signaling through the lymphotoxin-beta receptor stimulatesHIV-1 replication alone and in cooperation with soluble or membrane-bound TNF-alpha. J. Immunol. 162, 6016–6023.
Mi, W., Belyavskyi, M., Johnson, R.R., Sieve, A.N., Storts, R., Meagher, M.W., Welsh, C.J., 2004. Alterations in chemokine expression followingTheiler's virus infection and restraint stress. J. Neuroimmunol. 151,103–115.
Mi, W., Young, C.R., Storts, R., Steelman, A., Meagher, M.W., Welsh, C.J.,in press. Stress alters pathogenecity and facilitates systemic dissemina-tion of Theiler's virus. Microb. Pathog.
Miller, S.D., VanDerlugt, C.L., Begolka, W.S., Pao, W., Yauch, R.L.,Neville, K.L., Katz-Levy, Y., Carrizosa, A., Kim, B.S., 1997. Persistentinfection with Theiler's virus leads to CNS autoimmunity via epitopespreading. Nat. Med. 3, 1133–1136.
Mohr, D.C., Hart, S.L., Julian, L., Cox, D., Pelletier, D., 2004. Associationbetween stressful life events and exacerbation in Multiple Sclerosis: ameta analysis. BMJ 328 (7442), 731–735.
Murray, P.D., McGavern, D.B., Pease, L.R., Rodriguez, M., 2002. Cellularsources and targets of IFN-gamma-mediated protection against viraldemyelination and neurological deficits. Eur. J. Immunol. 32, 606–615.
Oldstone, M.B., Race, R., Thomas, D., Lewicki, H., Homann, D., Smelt, S.,Holz, A., Koni, P., Lo, D., Chesebro, B., Flavell, R., 2002.Lymphotoxin-alpha-and lymphotoxin-beta-deficient mice differ insusceptibility to scrapie: evidence against dendritic cell involvement inneuroinvasion. J. Virol. 76, 4357–4363.
Oleszak, E.L., Chang, J.R., Friedman, H., Katsetos, C.D., Platsoucas, C.D.,2004. Theiler's virus infection: a model for multiple sclerosis. Clin.Microbiol. Rev. 17, 174–207.
Olson, J.K., Girvin, A.M., Miller, S.D., 2001. Direct activation of innate andantigen presenting functions of microglia following infection withTheiler's virus. J. Virol. 75, 9780–9789.
Paya, C.V., Patick, A.K., Leibson, P.J., Rodriguez, M., 1989. Role of naturalkiller cells as immune effectors in encephalitis and demyelinationinduced by Theiler's virus. J. Immunol. 143, 95–102.
Peng, Z.C., Kristensson, K., Bentivoglio, M., 1998. Distribution andtemporal regulation of the immune response in the rat brain tointracerebroventricular injection of interferon-gamma. Exp. Neurol.154, 403–417.
Pruett, S., 2003. Stress and the immune system. Pathophysiology 9, 133–153.Ramierz, F., Fowell, D.J., Puklavec, M., Simmonds, S., Mason, D., 1996.
Glucocorticoids promote a TH2 cytokine response by CD4+ T cells invitro. J. Immunol. 156, 2406–2412.
Rempel, J.D., Quina, L.A., Blakely-Gonzales, P.K., Buchmeier, M.J., Gruol,D.L., 2005. Viral induction of central nervous system innate immuneresponses. J. Virol. 79, 4369–4381.
Rodriguez, M., Leibowitz, J.L., Lampert, P.W., 1983. Persistent infection ofoligodendrocytes in Theiler's virus-induced encephalomyelitis. Ann.Neurol. 13, 426–433.
Rodriguez, M., Pavelko, K., Coffman, R.L., 1995. Gamma interferon iscritical for resistance to Theiler's virus-induced demyelination. J. Virol.69, 7286–7290.
Rodriguez, M., Pavelko, K.D., Njenga, M.K., Logan, W.C., Wettstein, P.J.,1996. The balance between persistent virus infection and immune cellsdetermines demyelination. J. Immunol. 157, 5699–5709.
Rodriguez, A., Angel, S.O., Pinto, D.C., Vila, I., Acosta, A., Amendoeira, R.R., Lannes-Vieira, J., Rodrigues, M.R., 2003. Interferon-gammaindependent oxidation of melatonin by macrophages. Vaccine 21,1327–1335.
Sato, S., Reiner, S.L., Jensen, M.A., Roos, R.P., 1997. Central nervoussystem cytokine mRNA expression following Theiler's murineencephalomyelitis virus infection. J. Neuroimmunol. 76, 213–223.
60 W. Mi et al. / Journal of Neuroimmunology 178 (2006) 49–61

Autho
r's
pers
onal
co
py
Selmaj, K.W., Raine, C.S., 1988. Tumor necrosis factor mediates myelin andoligodendrocyte damage in vitro. Ann. Neurol. 23, 339–346.
Shaw-Jackson, C., Michiels, T., 1997. Infection of macrophages by Theiler'smurine encephalomyelitis virus is highly dependent on their activationor differentiation state. J. Virol. 71, 8864–8867.
Sheridan, J.F., Feng, N.G., Bonneau, R.H., Allen, C.M., Huneycutt, B.S.,Glaser, R., 1991. Restraint stress differentially affects anti-viral cellularand humoral immune responses in mice. J. Neuroimmunol. 31,245–255.
Sieve, A.N., Steelman, A.J., Young, C.R., Storts, R., Welsh, T.H., Welsh, C.J., Meagher, M.W., 2004. Chronic restraint stress during early Theiler'svirus infection exacerbates the subsequent demyelinating disease in SJLmice. J. Neuroimmunol. 155, 103–118.
Soderberg, K.A., Linehan, M.M., Ruddle, N.H., Iwasaki, A., 2004.MAdCAM-1 expressing sacral lymph node in the lymphotoxin beta-deficient mouse provides a site for immune generation following vaginalherpes simplex virus-2 infection. J. Immunol. 173, 1908–1913.
Stefanski, V., Peschel, A., Reber, S., 2003. Social stress affects migration ofblood T cells into lymphoid organs. J. Neuroimmunol. 138, 17–24.
Theil, D.J., Tsunoda, I., Libbey, J.E., Derfuss, T.J., Fujinami, R.S.,2000. Alterations in cytokine but not chemokine mRNA expressionduring three distinct Theiler's virus infections. J. Neuroimmunol.104, 22–30.
Trottier, M., Schlitt, B.P., Kung, A.Y., Lipton, H.L., 2004. Transition fromacute to persistent Theiler's virus infection requires active viral
replication that drives proinflammatory cytokine expression and chronicdemyelinating disease. J. Virol. 78, 12480–12488.
Tseng, R.J., Padgett, D.A., Dhabhar, F.S., Engler, H., Sheridan, J.F., 2005.Stress induced modulation of NK activity during influenza viralinfection: role of glucocorticoids and opioids. Brain Behav. Immun.19, 153–164.
Watford, W.T., Moriguchi, M., Morinobu, A., O'Shea, J.J., 2003. Thebiology of IL-12: coordinating innate and adaptive immune responses.Cytokine Growth Factor Rev. 14, 361–368.
Welsh, C.J., Tonks, P., Nash, A.A., Blakemore, W.F., 1987. The effect ofL3T4 T cell depletion on the pathogenesis of Theiler's murineencephalomyelitis virus infection in CBA mice. J. Gen. Virol. 68,1659–1667.
Welsh, C.J., Sapatino, B.V., Rosenbaum, B.A., Smith III, R., 1995.Characteristics of cloned cerebrovascular endothelial cells followinginfection with Theiler's virus: I. Acute infection. J. Neuroimmunol. 62,119–125.
Welsh, C.J., Bustamante, L., Nayak, M., Welsh, T.H., Dean, D.D., Meagher,M.W., 2004. The effects of restraint stress on the neuropathogenesis ofTheiler's virus infection: II. NK cell function and cytokine levels inacute disease. Brain Behav. Immun. 18, 166–174.
Wong, G.H., Goeddel, D.V., 1986. Tumor necrosis factors alpha and betainhibit virus replication and synergize with interferons. Nature 323,819–822.
61W. Mi et al. / Journal of Neuroimmunology 178 (2006) 49–61





























