Embed Size (px)
Citation preview

RESURGENCE OF INTEGRATED BEHAVIORAL UNITS
GUSTAVO BACHA-MENDEZ1, ALLISTON K. REID
2, AND ADELA MENDOZA-SOYLOVNA1
1FACULTAD DE PSICOLOGIAUNIVERSIDAD NACIONAL AUTONOMA DE MEXICO
2DEPARTMENT OF PSYCHOLOGYWOFFORD COLLEGE, SPARTANBURG, SC
Two experiments with rats examined the dynamics of well-learned response sequences whenreinforcement contingencies were changed. Both experiments contained four phases, each of whichreinforced a 2-response sequence of lever presses until responding was stable. The contingencies thenwere shifted to a new reinforced sequence until responding was again stable. Extinction-inducedresurgence of previously reinforced, and then extinguished, heterogeneous response sequences wasobserved in all subjects in both experiments. These sequences were demonstrated to be integratedbehavioral units, controlled by processes acting at the level of the entire sequence. Response-levelprocesses were also simultaneously operative. Errors in sequence production were strongly influencedby the terminal, not the initial, response in the currently reinforced sequence, but not by the previouslyreinforced sequence. These studies demonstrate that sequence-level and response-level processes canoperate simultaneously in integrated behavioral units. Resurgence and the development of integratedbehavioral units may be dissociated; thus the observation of one does not necessarily imply the other.
Key words: behavioral unit, extinction, functional response unit, response sequence, resurgence,sequence learning, lever press, rats
_______________________________________________________________________________
In his search for the fundamental units ofbehavior, Skinner (1935) defined a responseunit as a functional class of events subject tocontrol by reinforcing stimuli. A lever presscould become a behavioral unit, but so couldinterresponse times (IRTs) (e.g., Morse, 1966;Platt, 1973; Zeiler, 1977) and integratedsequences of simple responses (e.g., Fetter-man & Stubbs, 1982; Grayson & Wasserman,1979; Hawkes & Shimp, 1975; Reid, Chadwick,Dunham, & Miller, 2001; Schwartz, 1981, 1982,1986; Shimp, 1976, 1979). Zeiler (1977)identified three different kinds of responseunits that can be distinguished: formal, condi-tionable, and theoretical units. The formal
response unit is the operational definition ofthe measured response—that which the ex-perimenter prescribes as prerequisite for re-inforcer presentation. Conditionable responseunits are operants (Skinner, 1938). If a behav-ior is required for reinforcer presentation andreinforcement increases its probability, then itis a conditionable response unit. Finally, Zeiler(1977) defined theoretical response units asfollows:
The term response unit may also be used to referto something inferred rather than observeddirectly. A response, a stimulus–response re-lation, or some cognitive activity, can bepostulated to underlie observed performance.Inferred units are being used when it isasserted that organisms learn turning re-sponses, or to approach certain locations ina maze, or interresponse times, or entiresequences of behavior. (pp. 222–223)
Thus, theoretical response units are condition-able units (operants) that answer the questionof how reinforcement organizes behavior andwhat behavior is strengthened. They mayinclude self-imposed behavioral patterns last-ing several seconds (Shimp, 1976, 1979) orcombinations of simpler operants (Zeiler,1986a, 1986b).
Identification of formal and conditionableresponse units is unambiguous. The identifi-cation of theoretical response units is more
Please direct correspondence to the first author atLaboratorio de Adaptacion Animal, Facultad de Psicologıa,UNAM, Ave. Universidad #3004, Colonia Copilco Universi-dad, CP: 04510. Mexico, D. F. or Email at [email protected]. Direct correspondence to the secondauthor at Department of Psychology, Wofford College, 429N. Church St., Spartanburg, SC 29303 or Email at [email protected].
The first experiment was part of the requirements forthe degree of Licenciatura for the third author. Portions ofthis work were presented at the XV Congreso Mexicano deAnalisis de la Conducta in Puerto Vallarta, Jalisco, Mexicoin October 2003. Special thanks to Livia Sanchez for herhelp in carrying out the second experiment, to DianaEspinosa for her comments, to Ixel Alonso for her helpwith data analysis, and to Gregory Lieving for his insightfulreview of a previous draft.
doi: 10.1901/jeab.2007.55-05
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 2007, 87, 5–24 NUMBER 1 (JANUARY)
5

difficult because theoretical units are inferredover repeated observations of behavior ratherthan observed directly. This study concerns theidentification of theoretical response units asintegrated response sequences influenced byreinforcement contingencies.
Several different terms have been used torepresent the concept of theoretical responseunits, beginning with Skinner’s (1935, 1938)functional response units that may change overtime (see also Arbuckle & Lattal, 1988; Reid etal., 2001; Schneider & Morris, 1992). Shimp(1976) used the term functional units of analysis.Schwartz (1981, 1982, 1986) has argued thatcontingent reinforcement could turn an arbi-trary sequence of discrete key pecks into anintegrated behavioral unit, which he has alsocalled functional behavioral units or complexbehavioral units.
How can these theoretical behavioral unitsbe identified and measured? Four techniqueshave been used in recent years, although wewill argue that not all of these techniques areadequate.
1. One technique of identifying theoreticalbehavioral units was explained clearly byZeiler (1977), although it had been im-plemented earlier (e.g., Findley, 1962;Kelleher, Fry, & Cook, 1964). This tech-nique studies the hypothesized unitdirectly by first imposing the hypothesizedunit as a formal unit, and then demons-trating that it is conditionable. For exam-ple, to test the claim that certain reinforce-ment schedules can lead to the develop-ment of IRTs as theoretical behavioralunits, one could differentially reinforcea particular IRT and demonstrate thatIRT is controlled by reinforcement in thesame way as expected for simple responses.
2. Stability and replicability in the temporalpattern of reinforced behavior (overa period of several seconds) may indicatea self-imposed organization of behaviorunder the control of reinforcement con-tingencies (e.g., Hawkes & Shimp, 1975;Pisacreta, 1982; Reid et al., 2001; Shimp,1976, 1979; Terrace, 2001). If one demon-strates that this temporal pattern is alsounder stimulus control, it would provideeven stronger evidence that the behavior-al pattern functions as a behavioral unit.For example, Reid et al. (2001) examined
the development of integrated behavioralunits by observing the development ofself-imposed organization of behaviorwhen differential reinforcement was pro-vided for sequences of two lever presses ina free-operant procedure. They conclud-ed that integrated behavioral units con-sisting of two lever presses had formed bydemonstrating that the regularities inIRTs and transition probabilities betweenlevers were stable, replicable, and underthe stimulus control produced by demar-cating stimuli and by local reinforcementprobability.
3. Several researchers have concluded thatsequences of two or three lever presses orkey pecks had become organized intointegrated (theoretical) behavioral unitsby demonstrating that the generalizedmatching law was better described bysequence matching than by matching ofindividual responses (Fetterman &Stubbs, 1982; Reid et al., 2001; Schneider& Davison, 2005; Schneider & Morris,1992; Schwartz, 1986; Stubbs, Fetterman,& Dreyfus, 1987). The matching relationdescribes the influence of relative re-inforcement rate on relative responserate, but what ‘‘response’’ is controlledby the relative reinforcement rate? Stubbset al. demonstrated that when behaviorbecomes organized in such a way thatmatching of response sequences is ob-served when matching of individual re-sponses is not, then one can concludethat the response sequence is the func-tional response unit even if the individualkey peck had been the behavioral unitearlier in training.
4. The most common technique of identify-ing and measuring theoretical behavioralunits inherently assumes that reinforce-ment strengthens behavioral units wheth-er the units are key pecks, lever presses, orintegrated sequences of heterogeneousresponses (cf. Baum, 2002). A behavioralunit that has been reinforced frequentlyshould have substantial strength asa whole, allowing it to resist disruptionby a change in contingency (Schwartz,1981, 1982). Therefore, the techniqueinvolves changing the reinforcement con-tingencies and measuring whether theputative behavioral unit maintains its ‘‘in-
6 GUSTAVO BACHA-MENDEZ et al.

tegrity’’—maintaining the same behavior-al sequence and same temporal structure(Reed, Schachtman, & Hall, 1991). If itdoes maintain its integrity, yet its frequen-cy of production changes in accordancewith the reinforcement contingency, thenone can conclude that the pattern hasbecome a theoretical behavioral unit. Thistechnique has been widely implementedin research involving behavioral units(e.g., Grayson & Wasserman, 1979; Neur-inger, Kornell, & Olufs, 2001; Reed et al.,1991; Schwartz, 1981, 1982, 1986; Wasser-man, Nelson, & Larew, 1980; Zeiler, 1977,1986b).
Techniques 1 and 4 require the demonstra-tion that the behavioral unit is the same beforeand after a change in contingency—that it hasmaintained its integrity. For example, if thechange in contingency is a shift to extinction,then ‘‘to the extent that a stereotyped se-quence has become a behavioral unit, underextinction it should either occur or not, but ifit occurs, it should occur in the samestereotyped way that it does during reinforce-ment conditions’’ (Schwartz, 1981, p. 34).
These two techniques are problematic be-cause reinforced behavior sequences canoccur repetitively in the same order and withthe same temporal structure without thesequence achieving the status of an integratedbehavioral unit (Reid, Dixon, & Gray, inpress). For example, behavioral chains arecontrolled by reinforcement contingencies,and repetitions of behavioral chains are oftenhighly regular in their order and temporalstructure. Rather than being integrated struc-tures, behavioral chains are individual oper-ants linked together by response-producedstimuli. Under the right conditions, a discrim-inative stimulus can set the occasion for thefirst response in the chain, and the response-produced stimuli may allow the chain toprogress to the end. The important distinctionbetween integrated behavioral units and be-havioral chains is the level at which reinforce-ment is presumed to operate. With behavioralchains, reinforcement is presumed to influ-ence each response of the sequence separately(Catania, 1971; Skinner, 1938). With integrat-ed behavioral units, reinforcement is pre-sumed to operate at the level of the integratedunit, independent of its constituent responses.
Reid et al. (in press) argued that in order todemonstrate that a response sequence hasbecome an integrated behavioral unit, onemust demonstrate control by processes actingat the level of the purported unit (which wecall ‘‘sequence-level processes’’) rather thancontrol only by response-level processes (suchas all those involved in behavioral chains). Anexample of such a demonstration was outlinedas technique 3 above: pitting sequence match-ing against response matching and observingthat the matching law holds only with se-quences, not with individual responses (asdone originally by Stubbs et al., 1987, andmore comprehensively by Schneider & Davi-son, 2005). Sequence matching is a goodexample of a sequence-level process notaccounted for by more parsimonious re-sponse-level processes. Two examples of re-sponse-level processes are the effects of dis-criminative stimuli on the choice of the nextresponse, and the observation that reinforce-ment strengthens individual responses ina sequence depending upon their ordinalposition in the sequence or their temporalcontiguity with reinforcement (Catania, 1971).
Like sequence matching, extinction-in-duced resurgence appears to be anotherexample of behavior potentially influencedby a sequence-level process. Resurgence isobserved only after the resurgent responsehas been exposed to a history of reinforce-ment, and then followed by a period ofextinction (or very low reinforcement proba-bility) (Cleland, Guerin, Foster, & Temple,2001; Epstein, 1985; Lieving & Lattal, 2003).Presumably, the resurgent response has beenstrengthened sufficiently by reinforcement tomake it the most probable response followingextinction conditions. Prior studies of extinc-tion-induced resurgence have demonstratedresurgence of simple responses, rather thancomplex sequences that might involve theintegration of simpler responses. Nevertheless,to the extent that resurgence implies a historyof strengthening by reinforcement, a demon-stration of resurgence of entire responsesequences may potentially indicate that re-inforcement has strengthened response se-quences as an integrated behavioral unit. Ifso, then resurgence procedures may providea much-needed technique to study the de-velopment of integrated behavioral units, notonly their identification as units.
RESURGENCE OF INTEGRATED BEHAVIORAL UNITS 7

The main purpose of Experiment 1 was toassess whether extinction-induced resurgencecould reliably demonstrate that well-learnedresponse sequences had become integratedbehavioral units. At first glance, resurgence ofcomplete response sequences would appear tobe a sequence-level process that cannot beaccounted for by response-level processes.Thus, we also asked whether resurgencealways, never, or only in certain situations,indicates the existence of behavioral units.
An additional goal of this study concernedthe traditional ‘‘problem of behavioral units’’:As quantitative models of behavior are de-veloped that accurately predict behavior mea-sured as discrete responses (such as thematching law), how can we handle thetransition to new integrated behavioral units(such as those observed in sequence match-ing)? A learning model that assumes that theindividual key peck or lever press is theresponse unit that gains strength throughreinforcement will have considerable prob-lems unless it includes some mechanism forchanging the nature of the response unit.Presumably, once integrated behavioral unitsdevelop, the processes controlling their pro-duction are no longer limited to the response-level processes that affected the simplerresponses. Sequence-level processes presum-ably begin to control behavior, and reinforce-ment is assumed to strengthen the entiresequence as a new integrated behavioral unit.Therefore, our goal was to see if sequence-levelprocesses and response-level processes cansimultaneously control behavior, or whetherintegrated behavioral units, once organizedand controlled by reinforcement, precludecontrol by response-level processes.
EXPERIMENT 1METHOD
Subjects
Four naıve Wistar female rats, approximately4 months old at the beginning of the study,were maintained at 85% of their free-feedingweights with continuous access to water intheir individual home cages.
Apparatus
Two Med Associates (St. Albans, VT, USA)rat operant chambers measuring 26 cm wide,32 cm long, and 30 cm high were used. A
liquid dispenser (Med Associates ENV-200M),located through a 3-cm circular opening in thecenter of the control panel 2.5 cm above thefloor, provided access to a cup containing0.01 ml condensed milk diluted 50% withwater. The arm of the dispenser remained inthe up position until activated by a short pulseto fill the cup, which then remained availablefor an unlimited time period. The controlpanel also contained two response levers,located 9 cm to the left and right of thedispenser and 5.5 cm above the floor, andrequired a force of 0.15 N to operate. A 2.5-cmwhite 28-V lamp was located 5 cm above eachlever. A 28-V houselight was centered near thetop of the back wall opposite the controlpanel. Each operant chamber was locatedinside a sound-attenuating cubicle with a ven-tilation fan that masked extraneous noises. Inan adjacent room, a computer with Med-PCsoftware and a Med Associates interfacecontrolled each session and stored each eventand its time of occurrence.
Procedure
Training. After the reduction in bodyweights, subjects were trained to press leversby a successive approximations procedure.They then were exposed for three sessionsto a discrete-trials procedure that reinforcedthe completion of two lever presses, with-out regard to which levers were used. For thenext five sessions, reinforcement was contin-gent upon either heterogeneous sequence(i.e., LR and RL). Homogeneous sequenceswere unreinforced and immediately beganthe next trial. Sessions terminated after 30reinforcers or after 30 min, whichever camefirst.
Experimental procedure. All experimental ses-sions contained 50 trials. Each trial beganby turning on the lamps above the twolevers, and ended either when the subjectcompleted two responses or 1 min elapsedwithout completing two responses. Whenthe response sequence was correct, access tothe milk solution was made available byrefilling the cup, and the lamps were turnedoff for 3 s. If the response sequence wasincorrect or if 1 min elapsed without thecompletion of two responses, all lamps wereturned off for a timeout period of 10 s. Thehouselight remained on at all times exceptduring timeout.
8 GUSTAVO BACHA-MENDEZ et al.

In each experimental phase, each subjectwas trained to emit a particular two-responsesequence. This target sequence was constantthroughout all trials of the session. When thestability criteria were achieved, each subjectwas shifted to the next phase, which requireda different target sequence. The order ofexposure to these target sequences was coun-terbalanced across subjects (see Table 1). Withheterogeneous target sequences, respondingwas considered stable within a phase after atleast 25 sessions and when subjects were clearlydiscriminating between the two heterogeneoussequences (i.e., there was no overlap over fiveconsecutive sessions in the numbers of the twoheterogeneous sequences). Homogeneous se-quences are learned quickly, so we requireda minimum of 10 sessions when homogeneoussequences were the targets. Changes in phasealways began with the next session, neverwithin a session.
RESULTS AND DISCUSSION
Figure 1 shows the frequencies of each ofthe four possible response sequences (RR, LR,RL, and LL) for each subject in Phase 1 acrossblocks of five sessions. In order to more clearlyidentify differences in the curves, homoge-neous and heterogeneous sequences are de-picted in separate panels. The left panelsdepict the frequencies of heterogeneous se-quences (RL, LR) and identify the reinforcedsequence. The right panels depict the fre-quencies of the unreinforced homogeneoussequences in the same sessions. The left panelsshow a clear differentiation between the twoheterogeneous sequences as they came undercontrol of the reinforcement contingencies.For all subjects, the frequency of the rein-forced sequence systematically increased whilethe frequency of the unreinforced heteroge-neous sequence systematically decreased. In
order to encourage the formation of integrat-ed behavioral units consisting of the rein-forced sequence, we continued reinforcingthis sequence for many more sessions thannecessary for clear differentiation between thetwo heterogeneous sequences.
The homogeneous sequences depicted inthe right panels of Figure 1 were unreinforcederrors. In 3 of the 4 subjects, the mostcommon error was to repeat the last responserequired in the reinforced sequence. Forexample, Subject R21’s LR sequence wasreinforced and the most common error wasRR. Subjects rarely produced the other erro-neous homogeneous sequence.
Figure 2 shows the same analysis for Phase 2in which the opposite heterogeneous se-quence was reinforced. The number of ses-sions required for subjects to produce this newreinforced sequence more often than thepreviously reinforced heterogeneous sequence(now undergoing extinction) was substantiallygreater than that observed in the previousphase for all subjects. Phase 1 required 3–15sessions to observe clear differentiation in thefrequencies of the reinforced and unrein-forced heterogeneous sequences. However,Phase 2 often required 35–40 sessions toproduce this differentiation. Extinction ofthe previously reinforced sequence also re-quired many sessions for 3 of the 4 subjects.This slow rate of learning may have been dueto the extended training in Phase 1 thatencouraged the formation of integrated be-havioral units. Consistent with this hypothesis,subject R23 showed the fastest learning inPhase 2 and was exposed to the smallestnumber of sessions in Phase 1.
In contrast, the right panels of Figure 2show an immediate change in the types ofhomogeneous sequence errors produced. InPhase 1, subjects tended to repeat the lastresponse of the reinforced sequence. Thoseparticular erroneous sequences disappearedabruptly with the transition to the newlyreinforced sequence, even though subjectswere yet to emit many reinforced sequences.Instead, the most frequent homogeneoussequences were those that repeated the lastresponse required in the new reinforcedsequence. This observation has been docu-mented before (e.g., Grayson & Wasserman,1979), and it is consistent with previousreports that the terminal response in a se-
Table 1
Order of phases for each subject in Experiment 1. Eachphase required subjects to complete a particularreinforced sequence of left (L) and right (R) lever presses.
Phase: 1st 2nd 3rd 4th
Subject: Target Sequence:R21 LR RL RR LLR22 LR RL LL RRR23 RL LR RR LLR24 RL LR LL RR
RESURGENCE OF INTEGRATED BEHAVIORAL UNITS 9

Fig. 1. Each graph shows the average number of each response sequence observed in Phase 1 for each subject inExperiment 1, depicted over blocks of five sessions. The two heterogeneous sequences are depicted in the left panels, andthe homogeneous sequences are depicted in the right panels. Phase 1 reinforced a heterogeneous sequence, depicted inthe left panels as LR+ or RL+. Error bars represent one standard error.
10 GUSTAVO BACHA-MENDEZ et al.

Fig. 2. Each graph shows the average number of each response sequence observed in Phase 2 for each subject inExperiment 1, depicted over blocks of five sessions. The two heterogeneous sequences are depicted in the left panels, andthe homogeneous sequences are depicted in the right panels. Phase 2 reinforced a heterogeneous sequence, depicted inthe left panels as LR+ or RL+. Error bars represent one standard error.
RESURGENCE OF INTEGRATED BEHAVIORAL UNITS 11

quence is more sensitive to changes in re-inforcement contingency than are earlierresponses in the sequence (e.g., Corbit &Balleine, 2003; Reid, 1994; Reid et al., in press;Williams, 1999; Williams, Ploog, & Bell, 1995).Yet this observation is counterintuitive becauseit conflicts with predictions based on rein-forcement contiguity: The extensive reinforce-ment (from Phase 1) of the old sequenceshould have increased its resistance to change,especially of the terminal response mostcontiguous with reinforcement. As a result,one would expect persistence errors of theterminal response to be observed when thereinforcement contingency was changed.These persistence errors were not observed.Thus, error production did not appear to beaffected by resistance to change of the pre-viously learned sequence. Greater sensitivity ofthe terminal response to changes in reinforce-ment contingency was a stronger determinantof sequence errors than was the reinforcementhistory of the prior sequence (see also Reid,1994; Reid et al., in press).
The left panels of Figure 3 depict all foursequences observed in Phase 3, in whichpreviously reinforced heterogeneous se-quences were extinguished and a new homo-geneous sequence was reinforced. Recall thatPhases 3 and 4 lasted only 10 sessions eachbecause homogenous sequences are learnedquickly. Therefore, Figure 3 shows the fre-quencies of each sequence across individualsessions, rather than across blocks of sessions.We were interested in the changes that occursoon after the transition to the new phase, sothe first three sessions are depicted foremphasis. The last three sessions of thecondition are also depicted to show thesteady-state levels obtained for each sequence.The first three sessions in the left panels showa rapid increase observed in the newly re-inforced homogeneous sequence and a rapiddecrease in the previously reinforced hetero-geneous sequence. This rapid decrease standsin sharp contrast to the very slow extinctioncurves depicted in Figure 2, which were alsoheterogeneous sequences. Extinction of het-erogeneous sequences had required 35–40sessions when the new target was a heteroge-neous sequence (Phase 2), but only 3–6sessions were required when the new targetwas a homogeneous sequence (Phase 3). Thesteady-state level of responding at the end of
the phase was nearly perfect accuracy on thereinforced homogeneous sequence for allsubjects. With each subject, the sequencereinforced extensively in Phase 1 occurred atnegligible rates in Phase 3. Therefore, therewas no indication of extinction-induced re-surgence.
The results of Phase 4 are depicted in theright panels of Figure 3. In Phase 4, thepreviously reinforced homogeneous sequencewas no longer reinforced, and the otherhomogeneous sequence was reinforced. Incontrast to acquisition in Phase 3, at the startof Phase 4 resurgence of a previously learnedheterogeneous sequence substantially retard-ed learning of the new homogeneous se-quence. For all subjects the heterogeneoussequence that had been reinforced in Phase 2(see Figure 2) showed substantial resurgencein Phase 4 (identified in Figure 3 by arrows),even becoming the dominant sequence for 3subjects during the first three sessions of thephase. In all subjects, this sequence occurredat frequencies much higher than the currentlyreinforced sequence. For all subjects resur-gence of complex heterogeneous sequencesoccurred after a 10-day period of extinction(Phase 3) during which the frequency of thissequence was approximately zero. Ironically,the resurgent sequence was the same sequencethat had shown such a slow increase infrequency in Phase 2.
Transitions to each new phase includedextinction of the previously reinforced re-sponse; thus, it is important to demonstratethat the observed increase in sequence pro-duction, which we identify as extinction-in-duced resurgence, was not simply the resultof increased extinction-induced variability. Amethod providing a quantitative comparisonof resurgence versus increased variabilitywould be useful as a measure of the relativestrength of resurgence. Extinction-inducedvariability presumably would affect both het-erogeneous sequences equally, since theirdifficulty levels should be equal. Thus, thesimplest approach is to compare the numberof observations of the two sequences at eachof the first few sessions of the phase (whenthe transient effects of resurgence wouldbe expected to be observed). Figure 4 com-pares the frequency of occurrence of theresurgent sequence with that of the otherheterogeneous sequence, averaged across sub-
12 GUSTAVO BACHA-MENDEZ et al.

Fig. 3. The left panels depict all four sequence types observed in Phase 3, and the right panels depict Phase 4, for eachsubject in Experiment 1. Each graph shows the frequencies of each sequence across individual sessions, rather thanacross blocks of sessions. The first three sessions and the last three sessions of each phase are depicted in order toemphasize acquisition and the final steady-state levels of each sequence. Phases 3 and 4 reinforced homogeneoussequences, depicted in each panel as LL+ or RR+. Extinction-induced resurgence of a previously reinforced responsesequence is identified in the right panels by arrows.
RESURGENCE OF INTEGRATED BEHAVIORAL UNITS 13

jects, for the first three sessions of Phase 4.This average is representative of individualsubjects because every subject showed thesame effect. We examined the differencebetween the frequencies of the resurgentsequence and the other sequence using a 23 4 (Sequence 3 Session) mixed analysis ofvariance (ANOVA) with the first four sessionsas a repeated-measure factor across the 4subjects. Overall, the resurgent sequenceoccurred significantly more often than theother heterogeneous sequence, F(1,6) 525.725, p 5 .002, g2 5 .811. Therefore,resurgence was not an artifact of extinction-induced variability.
We repeatedly observed that the mostcommon error was to repeat the last responserequired in the new reinforced sequence. Theordinal position of responses within thesequence differentially affected the types oferrors produced—clearly a response-level pro-cess. Was this response-level process responsi-ble for resurgence? That is, did resurgencedepend upon the resurgent sequence sharingthe terminal (or initial) response with thenewly reinforced sequence? Resurgence wasobserved in all 4 subjects, yet only 2 subjects(R21 and R24) shared the same terminalresponse in the resurgent sequence and thecurrently reinforced sequence. Thus, resur-gence would not have been expected with theother 2 subjects, yet it was observed. Further-more, only 2 subjects (R22 and R23) shared
the same initial response in the resurgentsequence and the currently reinforced se-quence, yet resurgence was observed in allsubjects. The same logic holds with thepotential influence of the previously rein-forced sequence: only 2 subjects shared thesame initial or terminal response in theresurgent sequence and the previously rein-forced sequence, yet resurgence of the se-quence reinforced in Phase 2 was observed inall subjects. Therefore, we found no evidencethat resurgence was due to a response-levelprocess in this experiment. Naturally, Experi-ment 1 does not rule out the possibility ofresponse-level influence on resurgence inother situations.
Extinction-induced resurgence was observedin all subjects in Phase 4, but why did nosubjects show resurgence in Phase 3? Phase 3provided the same opportunities to observeextinction-induced resurgence that were pro-vided in Phase 4: the opportunity for therecurrence of a previously reinforced behavior(the heterogeneous sequence reinforced ex-tensively in Phase 1) under the condition thatreinforcer delivery ceases for a more recentlyreinforced behavior (the heterogeneous se-quence reinforced in Phase 2, which was nolonger reinforced in Phase 3) (Cleland et al.,2001; Epstein, 1985). To the extent thatresurgence of these sequences implies theexistence of integrated behavioral units, it isimportant to understand why the units wouldbe observed in one phase but not in anapparently similar phase. Understanding thecauses of resurgence may elucidate how in-tegrated behavioral units develop. Experiment2 continued this exploration by asking howresurgence of sequences depends upon thenumber and order of exposure to the re-inforcement conditions of Experiment 1.Experiment 2 repeated the study with a differ-ent order of reinforced sequences for eachsubject, examined which conditions produceextinction-induced resurgence, and examinedthe implications for the development of in-tegrated behavioral units. Our main concernwas not in resurgence, per se, but in what itcan teach us about the development of in-tegrated behavioral units. We wanted toestablish whether extinction-induced resur-gence is, in fact, a sequence-level process,and whether sequence-level and response-levelprocesses can coexist.
Fig. 4. The frequency of occurrence of the resurgentsequence is compared to the frequency of the otherheterogeneous sequence, averaged across subjects, foreach of the first three sessions of Phase 4 in Experiment 1.Error bars represent one standard error.
14 GUSTAVO BACHA-MENDEZ et al.

EXPERIMENT 2METHOD
Subjects
Four naıve Wistar female rats, approximately4 months old at the beginning of the study,were maintained at 85% of their free-feedingweights with continuous access to water intheir individual home cages.
Apparatus
The same rat chambers used in Experiment1 were used in the current experiment withone modification. The liquid dispensers werereplaced with standard pellet dispensers (MedAssociates, ENV 203M) that delivered 45-mgNoyes (Formula A/I) pellets into an un-illuminated receptacle (Med Associates, ENV-200-R2M), accessible through a 5-cm squareopening.
Procedure
The procedures used for training and thefour phases of this experiment were the sameas those used in Experiment 1 with twoexceptions. As illustrated in Table 2, the orderof reinforced sequences across phases wasdifferent. Experiment 1 did not includetransitions between previously reinforced ho-mogeneous sequences and newly reinforcedheterogeneous sequences. This type of transi-tion was included in the current experiment.The other difference in procedure concernedthe number of sessions in each phase. Astability criterion was in effect for Experiment1 that allowed different subjects to receivedifferent amounts of exposure to each re-inforced sequence in Phases 1 and 2. In thecurrent experiment, the number of sessions ineach phase was held constant across subjects.Phases that reinforced heterogeneous se-quences lasted 40 sessions, and those reinfor-cing homogeneous sequences lasted 10 ses-sions for all subjects.
RESULTS AND DISCUSSION
Figure 5 shows the frequencies of each ofthe four possible response sequences for eachsubject in Phase 1 across blocks of five sessions.As in the earlier figures, homogeneous andheterogeneous sequences are depicted inseparate panels to more clearly identify differ-ences in the curves. As the left panels show, allsubjects differentiated between the reinforcedand unreinforced heterogeneous sequences.The right panels depict homogeneous se-quences, which were always errors in thisphase. In 2 of the subjects (A3 and A4), thedifferences in frequency between the twohomogeneous sequences demonstrated a tran-sient tendency to repeat the last responserequired in the heterogeneous target se-quence. Subjects rarely produced the othererroneous homogeneous sequence. Theseobservations are consistent with the results ofPhases 1 and 2 of Experiment 1, but the effectswere larger and more enduring in the earlierexperiment.
The left panels of Figure 6 depict the resultsof Phase 2, in which previously reinforcedheterogeneous sequences were extinguishedand a new homogeneous sequence was re-inforced. Recall that Phases 2 and 3 lasted only10 sessions because homogenous sequencesare learned quickly. Therefore, Figure 6 showsthe frequencies of each sequence across in-dividual sessions, rather than across blocks ofsessions. All subjects learned the new homo-geneous target sequence rapidly, producingthis reinforced sequence in nearly every trialby Session 10, ending the phase. The pre-viously reinforced heterogeneous sequencewas effectively extinguished.
The right panels of Figure 6 depict theresults of Phase 3, which provided the firstopportunity to replicate the extinction-in-duced resurgence observed only in Phase 4of Experiment 1. All subjects showed resur-gence of the heterogeneous sequence that hadbeen reinforced in Phase 1. Resurgence inSubject A1 was short-lived, but the other 3subjects showed stronger, more persistentresurgence. Resurgence was observed in eachsubject regardless of whether the resurgentsequence shared the same initial or terminalresponse as the currently reinforced sequenceor as the previously reinforced sequence inthe prior phase. Therefore, the ordinal posi-tion of responses within the sequences was not
Table 2
Order of phases for each subject in Experiment 2. Eachphase required subjects to complete a particularreinforced sequence of left (L) and right (R) lever presses.
Phase: 1st 2nd 3rd 4th
Subject: Target Sequence:A1 RL LL RR LRA2 RL RR LL LRA3 LR LL RR RLA4 LR RR LL RL
RESURGENCE OF INTEGRATED BEHAVIORAL UNITS 15

Fig. 5. Each graph shows the average number of each response sequence observed in Phase 1 for each subject inExperiment 2, depicted over blocks of five sessions. The two heterogeneous sequences are depicted in the left panels, andthe homogeneous sequences are depicted in the right panels. Phase 1 reinforced a heterogeneous sequence, depicted inthe left panels as LR+ or RL+. Error bars represent one standard error.
16 GUSTAVO BACHA-MENDEZ et al.

Fig. 6. The left panels depict all four sequence types observed in Phase 2, and the right panels depict Phase 3, for eachsubject in Experiment 2. Each graph shows the frequencies of each sequence across individual sessions, rather thanacross blocks of sessions. The first three sessions and the last three sessions of each phase are depicted in order toemphasize acquisition and the final steady-state levels of each sequence. Phases 2 and 3 reinforced homogeneoussequences, depicted in each panel as LL+ or RR+. Extinction-induced resurgence of a previously reinforced responsesequence is identified in the right panels by arrows.
RESURGENCE OF INTEGRATED BEHAVIORAL UNITS 17

a causal factor in extinction-induced resur-gence.
Once again, it is important to demonstratethat the observed increase in sequence pro-duction, which we identify as extinction-in-duced resurgence, was not simply the result ofincreased extinction-induced variability. Fig-ure 7 compares the frequency of occurrenceof the resurgent sequence with that of theother heterogeneous sequence, averagedacross subjects, for the first three sessions ofPhase 3. This average is representative ofindividual subjects because every subjectshowed the same effect. We examined thedifference between the frequencies of theresurgent sequence and the other sequenceusing a 2 3 4 (Sequence 3 Session) mixedanalysis of variance (ANOVA) with the firstfour sessions as a repeated-measure factoracross the 4 subjects. Overall, the resurgentsequence occurred significantly more oftenthan the other heterogeneous sequence,F(1,6) 5 13.201, p 5 .011, g2 5 .688.Therefore, resurgence was not an artifact ofextinction-induced variability.
Figure 8 depicts the results of Phase 4 inwhich reinforcement was shifted from homo-geneous to heterogeneous sequences. Thecurves in the left panels were similar to thecurves in the left panels of Figure 5, in whichsubjects first learned to differentiate betweenthe two types of heterogeneous sequences. Butin Figure 8, for the first 5–10 sessions all 4
subjects produced the unreinforced heteroge-neous sequence more often than the currentlyreinforced sequence. When one value isgreater than another, we can ask what processenhanced the larger value, or we could askwhat process diminished the smaller value, orboth. The higher frequency of the unrein-forced heterogeneous sequence may indicateresurgence of the sequence reinforced inPhase 1 (three, not two, phases earlier). Forall subjects, this was the same sequence thatresurged in Phase 3. Alternatively, or perhapsin addition, the currently reinforced hetero-geneous sequence may have begun the phasewith low frequencies because it had neverbefore been reinforced and had always pro-duced timeouts. The current procedure doesnot allow us to separate these two interpreta-tions, so we cannot be certain that resurgencewas obtained in this phase.
Recall that in Phases 1 and 2 of Experiment1, subjects tended to repeat the last responseof the currently reinforced sequence, such thatthe target sequence LR yielded a substantialnumber of RR errors. This same effect can beobserved in the current study by examinationof the right panels of Figure 8 depictinghomogeneous sequences. All homogeneoussequences can be considered errors becausethe only reinforced sequence was heteroge-neous. Nevertheless, all subjects producedmore of the homogeneous sequence thatshared the same response as the terminalresponse of the currently reinforced heteroge-neous sequence, replicating the results ofExperiment 1. This elevated level was notsimply a carryover effect from the previousphase because the opposite homogeneoussequence had been previously reinforced withsubjects A2 and A3. These homogeneoussequences are unlikely to reflect extinction-induced resurgence because these particularsequences had not been reinforced and thenextinguished in either Phase 1 or Phase 2 forsubjects A1 or A4, even though they had beenwith subjects A2 and A3. Thus, the simplestexplanation is that the most common error inthe homogeneous sequences was to repeat thelast response required in the currently rein-forced sequence, not in the previously rein-forced sequence—a response-level processobserved in every subject in both experiments.
This experiment was designed to explorefurther the relationship between resurgence of
Fig. 7. The frequency of occurrence of the resurgentsequence is compared to the frequency of the otherheterogeneous sequence, averaged across subjects, foreach of the first three sessions of Phase 3 in Experiment 2.Error bars represent one standard error.
18 GUSTAVO BACHA-MENDEZ et al.
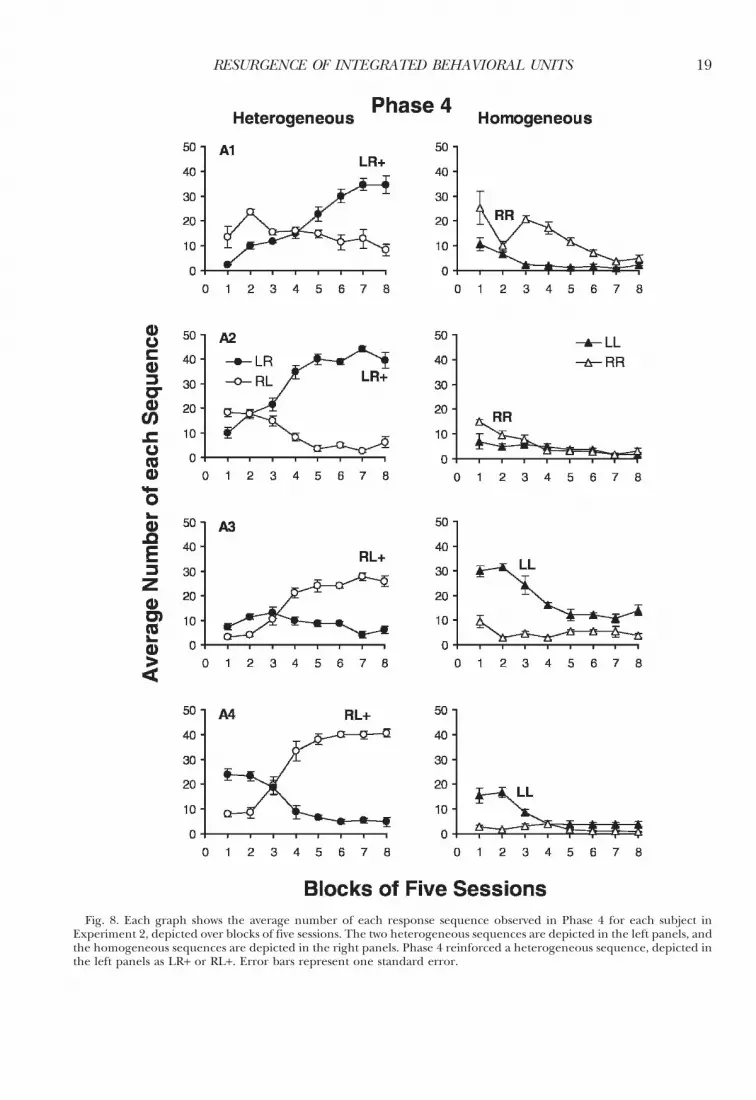
Fig. 8. Each graph shows the average number of each response sequence observed in Phase 4 for each subject inExperiment 2, depicted over blocks of five sessions. The two heterogeneous sequences are depicted in the left panels, andthe homogeneous sequences are depicted in the right panels. Phase 4 reinforced a heterogeneous sequence, depicted inthe left panels as LR+ or RL+. Error bars represent one standard error.
RESURGENCE OF INTEGRATED BEHAVIORAL UNITS 19

response sequences and the development ofintegrated behavioral units. We changed theorder of conditions from that of Experiment 1to see if resurgence would occur in a differentphase. Resurgence occurred exclusively inPhase 4 in Experiment 1, but it occurred inPhase 3 of the current study. There wasinconclusive evidence that these same se-quences may have resurged in Phase 4 as well,even though they were not the most recentlyreinforced candidate sequences. Resurgencedid not appear to be due to any response-levelprocesses in this experiment, thus replicatingthe results from Experiment 1. Response-levelprocesses were observed: The most commonerror for all subjects was to repeat the lastresponse required in the currently reinforcedsequence, not in the previously reinforcedsequence. This counterintuitive observationreplicates the types of errors observed inExperiment 1.
GENERAL DISCUSSION
All subjects in both experiments producedextinction-induced resurgence of a heteroge-neous response sequence. In all subjects inboth experiments, the most frequent errors insequence production were sequences thatrepeated the last response required in thecurrently reinforced sequence, but not in thepreviously reinforced sequence. Thus, errorproduction appeared to be controlled bya response-level process because the ordinalposition of responses in the sequence differ-entially affected the types of errors pro-duced. If the observed resurgence can bedemonstrated to be a sequence-level process,then these experiments appear to be the firstdemonstration that both sequence-level andresponse-level processes can operate simulta-neously within clearly established theoreticalbehavioral units (not to be confused withtraditional molar vs. molecular explanations ofbehavior on reinforcement schedules, inwhich theoretical behavioral units are notmeasured).
Does this resurgence demonstrate that thesequences had become integrated behavioralunits? Using the criteria for identifying theo-retical behavioral units proposed by Zeiler(1977), the first technique discussed in theintroduction, one would conclude that thesequences were, indeed, integrated theoretical
behavioral units because when reinforceddirectly, the resurgent sequences were differ-entially controlled by reinforcement (theywere conditionable units). In addition, theymaintained their ‘‘integrity’’ following shifts inreinforcement contingency with each subjectin each experiment (technique 4) because outof all possibilities, the resurgent sequenceswere precisely those that were reinforced twophases earlier.
Because the two techniques above arelimited in their ability to distinguish betweenintegrated behavioral units and behavioralchains, Reid et al. (in press) proposed analternative procedure for identifying integrat-ed behavioral units that provides a systematicmethod of evaluating the integrity of behav-ioral units. Their procedure asks: Were se-quence-level processes actually involved, or wasthe behavior pattern (i.e., resurgence) duesimply to one or more response-level pro-cesses? Their procedure of separating re-sponse-level from sequence-level processesprovides more precision than the techniquesdescribed earlier for identifying integratedbehavioral units. It begins with the creationof a decision tree representing the possibledecisions made at each point in the responsesequence.
Figure 9 depicts the conditional decisiontree representing each of the possible se-quences for both experiments. Because thesequences consisted of only two responses,only two decision levels were possible (one toselect the initial response and one for theterminal response). The tree is composed ofthree identical subtrees, each representing thechoice of pressing the left or right lever. Thefirst decision level, labeled ‘‘A’’, represents thechoice of the initial response in the sequence.This choice is likely to be affected by cuesindicating the beginning of the trial, such asprior food delivery or timeout. At the secondlevel, ‘‘B’’ and ‘‘C’’ represent the points atwhich the terminal response of the sequence isselected. This later decision is not likely to beaffected by the same beginning-of-trial cues,but other discriminative stimuli present in theenvironment (including the subject’s previousresponse and spatial location in the chamber)may influence lever selection. The completedsequences are represented at the bottom ofthe figure, to be followed by reinforcement ortimeout.
20 GUSTAVO BACHA-MENDEZ et al.

For each subject in both experiments, theobserved extinction-induced resurgence canbe represented as a single change at a partic-ular choice point. As an example, considerSubject R21 in Experiment 1 as it was shiftedfrom a reinforced sequence of RR in Phase 3to a new reinforced sequence LL in Phase 4.Before the new sequence was learned, thesubject produced extinction-induced resur-gence of the sequence RL, which had beenthe reinforced sequence in Phase 2. Thetransition from RR to RL represented a changeonly in the response produced in the terminalposition. In Figure 9, this change is represent-ed by choice point C. That is, resurgence forthis rat in this phase is represented as a re-sponse-level change at a single choice point, C.
This analysis allows one to answer questionsrelated to the separation of response-levelfrom sequence-level processes. For example:Did resurgence in each condition dependupon whether the extinguished sequenceand the resurgent sequence involved a changein the initial versus terminal response posi-tion? Resurgence in Experiment 1 involveda change in the terminal position for R21
(RRRRL, represented by choice point C) andR24 (LLRLR, choice point B) and a change inthe initial position for R22 (LLRRL, point A)and R23 (RRRLR, point A). Similarly, re-surgence observed in Phase 3 of Experiment 2involved a change in the terminal position forSubjects A2 (RRRRL, point C) and A3(LLRLR, point B) and a change in the initialposition for A1 (LLRRL, point A) and A4(RRRLR, point A). Therefore, resurgence inboth experiments was produced by changes atall three possible choice points, some pre-sumably influenced by beginning-of-trial cuesand others not. Therefore, resurgence did notdepend upon whether it involved a change inthe initial versus terminal response position.Resurgence did not depend upon this re-sponse-level process.
If only one or a limited number of possiblechoice points had been involved, then onemight conclude that resurgence was a re-sponse-level process, or the result of a limitedcombination of response-level processes. How-ever, this analysis demonstrates that extinc-tion-based resurgence of these response se-quences was a sequence-level process involvingchanges at every possible choice point. Itcannot be accounted for by proposing a moreparsimonious combination of response-levelprocesses. We are faced with two possibleexplanations in each experiment: either (a)subjects repeated a previously reinforced andwell-learned response sequence as an integrat-ed behavioral unit (a sequence-level process),or (b) every subject in both experimentsproduced that one particular series of in-dividual response decisions in which everydecision conformed to their previously rein-forced sequence, no matter what the responsepattern had been in the intervening phase.Clearly, the latter explanation is highly im-probable and unparsimonious. Sequence-levelprocesses were clearly operative. We canconfidently conclude that these well-learnedresponse sequences had formed integratedtheoretical behavioral units.
Sequence-level processes were operative, butresponse-level processes were simultaneouslyoperative within these integrated behavioralunits. Recall that the most frequent errors insequence production were sequences thatrepeated the last response required in thecurrently reinforced sequence (but not in thepreviously reinforced sequence). Thus, error
Fig. 9. This conditional decision tree represents theresponse-by-response choices as responses are producedto form response sequences. The letter A representsthe choice point for the initial response of the se-quence. Letters B and C represent the choice points forthe second, terminal response that terminates the two-response sequence.
RESURGENCE OF INTEGRATED BEHAVIORAL UNITS 21

production appeared to be controlled bya simple response-level process for all subjects.This process appears to be related to theassignment-of-credit problem during acquisi-tion (Staddon & Zhang, 1991), rather than tothe strength or persistence of responding inthe previous phase. The ordinal position ofresponses in the sequence differentially affect-ed the types of errors produced. To ourknowledge, this is the first demonstration thatresponse-level processes continue to be oper-ative after integrated behavioral units haveformed. Thus, reinforcement can work simul-taneously at the level of the simple responseand at the level of the integrated behavioralunit. This observation provides a potentialsolution to the traditional ‘‘problem’’ ofbehavioral units mentioned earlier: As quanti-tative models of behavior are developed thataccurately represent behavior measured asdiscrete responses, how can we handle thetransition to new integrated behavioral units?A learning model that assumes that theindividual key peck or lever press is theresponse unit that gains strength throughreinforcement may have considerable prob-lems unless it includes some mechanism forchanging the nature of the response unit. Thecurrent evidence implies that the processesacting on individual responses continue tooperate even when new behavioral units form,and new sequence-level processes can beadded to the determinants of the behaviorstream. The ‘‘problem’’ of the behavioral unitmay not require extensive modification ofexisting quantitative models of behavior.
Is resurgence related to the strength of thebehavioral unit? The notion of a behavioralunit is strongly tied to the assumption thatthese units gain strength when followed byreinforcement (Baum, 2002). Since the re-surgent sequences were integrated behavioralunits, we can ask whether resurgence was morelikely to be observed following phases thatprovided more reinforcement for the se-quences. Or, were additional factors (otherthan reinforcement history) involved in theproduction of resurgence?
In Experiment 1 (see Figure 1) Phase 1provided extensive reinforcement for a hetero-geneous sequence, and each subject pro-duced the reinforced sequence with highaccuracy. Therefore, the reinforced sequenceshould have become an integrated beha-
vioral unit with considerable strength. Indeed,when the contingency was shifted to Phase 2(see Figure 2), acquisition of the new se-quence appeared to be retarded most forthe subjects with the longest reinforcementhistory for the old sequence (Phase 1).Behavioral units with such strength should behighly likely to show resurgence, but noresurgence of these sequences was observedin Phase 3. By comparing the reinforcementhistories of the heterogeneous sequences inPhase 1 (Figure 1) to that of Phase 2 (seeFigure 2), it seems clear that the strength ofthe reinforced heterogeneous sequence inPhase 2 would be weaker than in Phase 1. Yetit was this presumably weaker sequence thatresurged strongly in Phase 4 with everysubject.
Experiment 2 also reinforced a heteroge-neous sequence in Phase 1 (see Figure 5) foran extended period to encourage the forma-tion of integrated behavioral units with con-siderable strength. It was precisely thesesequences that resurged in Phase 3 (Figure 6).How can these apparently conflicting resultsbetween experiments be reconciled? A richerreinforcement history of a complex behavioralunit does not necessarily lead to greaterprobability of its resurgence. For example, weobtained resurgence of heterogeneous se-quences in both studies, but no resurgenceof homogeneous sequences was obtained(even with nearly perfect accuracy duringtraining). Extinction-induced resurgence ap-pears to depend upon additional factorsunrelated to the strength of the behavioralunit. Possible examples may include thedifficulty of detecting changes in reinforce-ment contingency, and the difficulty of dis-criminating between reinforced and unrein-forced response sequences. Because suchadditional factors were not measured in thisstudy, the variables that influence the proba-bility of resurgence remain elusive. For re-searchers interested in studying resurgence,the current procedure provides a rich oppor-tunity to tease out the particular sets ofconditions that produce resurgence.
The development of integrated behavioralunits does not necessarily mean that resur-gence will be observed, even though it wasobserved in certain conditions of this study.What about the converse question: Doesobserved resurgence prove the existence of
22 GUSTAVO BACHA-MENDEZ et al.

integrated behavioral units? Not necessarily.Recall that simple behavioral chains can beconfused with integrated behavioral units, yetno integration across responses exists inchains. Under the right conditions, resurgenceof extended chains of responses might beobserved, but that does not mean they arebehavioral units (Reid et al., in press). Theconditions that would encourage resurgenceof behavioral chains are those with strongbeginning-of-trial cues and response-producedstimuli, similar to those in the proceduresand apparatus used by Schwartz (1981) andPisacreta (1982). Long chains involvingmany heterogeneous responses have beenrepeatedly demonstrated (e.g., Kelleher,1966), perhaps because behavioral chains donot require integration across constituentresponses. Therefore, we would expect futureresearchers to observe resurgence of responsesequences longer and more complex thanthose capable of being integrated into behav-ioral units.
REFERENCES
Arbuckle, J. L., & Lattal, K. A. (1988). Changes infunctional response units with briefly delayed re-inforcement. Journal of the Experimental Analysis ofBehavior, 49, 249–263.
Baum, W. M. (2002). From molecular to molar: Aparadigm shift in behavior analysis. Journal of theExperimental Analysis of Behavior, 78, 95–116.
Catania, A. C. (1971). Reinforcement schedules: The roleof responses preceding the one that produces thereinforcer. Journal of the Experimental Analysis ofBehavior, 15, 271–287.
Cleland, B. S., Guerin, B., Foster, T. M., & Temple, W.(2001). On terms: Resurgence. The Behavior Analyst,24, 255–260.
Corbit, L. H., & Balleine, B. W. (2003). Instrumental andPavlovian incentive processes have dissociable effectson components of a heterogeneous instrumentalchain. Journal of Experimental Psychology: Animal Behav-ior Processes, 29, 99–106.
Epstein, R. (1985). Extinction-induced resurgence: Pre-liminary investigations and possible applications. ThePsychological Record, 35, 143–153.
Fetterman, J. G., & Stubbs, D. A. (1982). Matching,maximizing, and the behavioral unit: Concurrentreinforcement of response sequences. Journal of theExperimental Analysis of Behavior, 37, 97–114.
Findley, J. D. (1962). An experimental outline for buildingand exploring multi-operant behavioral repertoires.Journal of the Experimental Analysis of Behavior, 5,113–166.
Grayson, R. J., & Wasserman, E. A. (1979). Conditioning oftwo-response patterns of key pecking in pigeons.Journal of the Experimental Analysis of Behavior, 31,23–29.
Hawkes, L., & Shimp, C. P. (1975). Reinforcement ofbehavioral patterns: Shaping a scallop. Journal of theExperimental Analysis of Behavior, 23, 3–16.
Kelleher, R. T. (1966). Chaining and conditioned re-inforcement. In W. K. Honig (Ed.), Operant behavior:Areas of research and application (pp. 160–212). Engle-wood Cliffs, NJ: Prentice-Hall.
Kelleher, R. T., Fry, W., & Cook, L. (1964). Adjusting fixed-ratio schedules in the squirrel monkey. Journal of theExperimental Analysis of Behavior, 7, 69–77.
Lieving, G. A., & Lattal, K. A. (2003). Recency, repeatabil-ity, and reinforcer retrenchment: An experimentalanalysis of resurgence. Journal of the ExperimentalAnalysis of Behavior, 80, 217–233.
Morse, W. H. (1966). Intermittent reinforcement. In W. K.Honig (Ed.), Operant behavior: Areas of research andapplication (pp. 52–108). Englewood Cliffs, NJ: Pren-tice-Hall.
Neuringer, A., Kornell, N., & Olufs, M. (2001). Stabilityand variability in extinction. Journal of ExperimentalPsychology: Animal Behavior Processes, 27, 79–94.
Pisacreta, R. (1982). Some factors that influence theacquisition of complex, stereotyped, response se-quences in pigeons. Journal of the Experimental Analysisof Behavior, 37, 359–369.
Platt, J. R. (1973). Percentile reinforcement: Paradigms forexperimental analysis of response shaping. In G. H.Bower (Ed.), Psychology of learning and motivation:Advances in research and theory: Vol. 7 (pp. 271–296).New York: Academic Press.
Reed, P., Schachtman, T. R., & Hall, G. (1991). Effect ofsignaled reinforcement on the formation of behav-ioral units. Journal of Experimental Psychology: AnimalBehavior Processes, 17, 475–485.
Reid, A. K. (1994). Learning new response sequences.Behavioural Processes, 32, 147–162.
Reid, A. K., Chadwick, C. Z., Dunham, M., & Miller, A.(2001). The development of functional responseunits: The role of demarcating stimuli. Journal of theExperimental Analysis of Behavior, 76, 303–320.
Reid, A. K., Dixon, R. A., & Gray, S. (in press).Variation and selection in response structures. InN. Innis (Ed.), Reflections on adaptive behavior: Essaysin honor of J. E. R. Staddon. Cambridge, MA: MITPress.
Schneider, S. M., & Davison, M. (2005). Demarcatedresponse sequences and generalized matching. Beha-vioural Processes, 70, 51–61.
Schneider, S. M., & Morris, E. K. (1992). Sequences ofspaced responses: Behavioral units and the role ofcontiguity. Journal of the Experimental Analysis ofBehavior, 58, 537–555.
Schwartz, B. (1981). Reinforcement creates behavioralunits. Behaviour Analysis Letters, 1, 33–41.
Schwartz, B. (1982). Interval and ratio reinforcementof a complex, sequential operant in pigeons.Journal of the Experimental Analysis of Behavior, 37,349–357.
Schwartz, B. (1986). Allocation of complex, sequentialoperants on multiple and concurrent schedules ofreinforcement. Journal of the Experimental Analysis ofBehavior, 45, 283–295.
Shimp, C. P. (1976). Short-term memory in the pigeon:Relative recency. Journal of the Experimental Analysis ofBehavior, 25, 55–61.
RESURGENCE OF INTEGRATED BEHAVIORAL UNITS 23

Shimp, C. P. (1979). The local organization of behaviour:Method and theory. In M. D. Zeiler, & P. Harzem(Eds.), Reinforcement and the organization of behavior (pp.261–298). Chichester, England: Wiley.
Skinner, B. F. (1935). The generic nature of the conceptsof stimulus and response. Journal of General Psychology,12, 40–65.
Skinner, B. F. (1938). Behavior of organisms: An experimentalanalysis. New York: Appleton-Century.
Staddon, J. E. R., & Zhang, Y. (1991). On the assignment-of-credit problem in operant learning. In M. L.Commons, S. Grossberg, & J. E. R. Staddon (Eds.),Neural network models of conditioning and action (pp.279–293). Hillsdale, NJ: Erlbaum.
Stubbs, D. A., Fetterman, J. G., & Dreyfus, L. R. (1987).Concurrent reinforcement of response sequences. InM. L. Commons, J. E. Mazur, J. A. Nevin, & H. Rachlin(Eds.), Quantitative analyses of behavior: Vol. 5. The effectof delay and of intervening events on reinforcement value(pp. 205–224). Hillsdale, NJ: Erlbaum.
Terrace, H. (2001). Chunking and serially organizedbehavior in pigeons, monkeys, and humans. In R. G.Cook (Ed.), Avian visual cognition. Retrieved August 3,2006, from Tufts University, Department of Psychol-ogy Avian Visual Cognition Laboratory web site:www.pigeon.psy.tufts.edu/avc/terrace.
Wasserman, E. A., Nelson, K. R., & Larew, M. B. (1980).Memory for sequences of stimuli and responses.Journal of the Experimental Analysis of Behavior, 34,49–59.
Williams, B. A. (1999). Value transmission in discrimina-tion learning involving stimulus chains. Journal of theExperimental Analysis of Behavior, 72, 177–185.
Williams, B. A., Ploog, B. O., & Bell, M. C. (1995). Stimulusdevaluation and extinction of chain schedule per-formance. Animal Learning and Behavior, 23, 104–114.
Zeiler, M. D. (1977). Schedules of reinforcement. In W. K.Honig, & J. E. R. Staddon (Eds.), Handbook of operantbehavior (pp. 201–232). Englewood Cliffs, NJ: Prentice-Hall.
Zeiler, M. D. (1986a). Behavioral units: A historicalintroduction. In T. Thompson, & M. D. Zeiler(Eds.), Analysis and integration of behavioral units (pp.1–12). Hillsdale, NJ: Erlbaum.
Zeiler, M. D. (1986b). Behavior units and optimality. In T.Thompson, & M. D. Zeiler (Eds.), Analysis andintegration of behavioral units (pp. 81–114). Hillsdale,NJ: Erlbaum.
Received: July 7, 2005Final acceptance: August 9, 2006
24 GUSTAVO BACHA-MENDEZ et al.





























