Embed Size (px)
Citation preview

Article
Rice Resistance Buffers against the Induced Enhancement ofBrown Planthopper Fitness by Some Insecticides
Finbarr G Horgan 123 Ainara Pentildealver-Cruz 45 and Maria Liberty P Almazan 5
Citation Horgan FG
Pentildealver-Cruz A Almazan MLP
Rice Resistance Buffers against the
Induced Enhancement of Brown
Planthopper Fitness by Some
Insecticides Crops 2021 1 166ndash184
httpsdoiorg103390
crops1030016
Academic Editors Fulai Liu
Mohammad Anwar Hossain and
Mohammad Golam Mostofa
Received 5 November 2021
Accepted 30 November 2021
Published 3 December 2021
Publisherrsquos Note MDPI stays neutral
with regard to jurisdictional claims in
published maps and institutional affil-
iations
Copyright copy 2021 by the authors
Licensee MDPI Basel Switzerland
This article is an open access article
distributed under the terms and
conditions of the Creative Commons
Attribution (CC BY) license (https
creativecommonsorglicensesby
40)
1 EcoLaVerna Integral Restoration Ecology Bridestown Kildinan T56 P499 County Cork Ireland2 Escuela de Agronomiacutea Facultad de Ciencias Agrarias y Forestales Universidad Catoacutelica del Maule
Casilla 7-D 3349001 Curicoacute Chile3 Centre for Pesticide Suicide Prevention UniversityBHF Centre for Cardiovascular Science
University of Edinburgh Edinburgh EH16 4TJ UK4 Institut de Geacuteneacutetique Environnement et Protection des Plantes (IGEPP)
Institut National de Recherche pour lrsquoAgriculture lrsquoAlimentation et lrsquoEnvironnement (INRAE) Institut AgroUniversity of Rennes CEDEX 49045 Angers France ainarapenalveragrocampus-ouestfr
5 International Rice Research Institute Makati 1226 Philippines mlalmazanirriorg Correspondence fhorganecolavernaorg
Abstract The brown planthopper Nilaparvata lugens (Staringl)[BPH] is a damaging pest of rice inAsia Insecticides and rice varietal resistance are widely implemented BPH management prac-tices However outbreaks of BPH have been linked to excessive insecticide usemdashchallengingthe compatibility of these two management practices IR62 is a variety with resistance againstBPH the whitebacked planthopper Sogatella furcifera Horvaacuteth [WBPH] and the green leafhopperNephotettix virescens (Distant)[GLH] We compared BPH responses to IR62 and to the susceptiblevariety IR64 treated with buprofezin carbofuran cartap hydrochloride cypermethrin deltamethrinfipronil or thiamethoxam + chlorantraniliprole In greenhouse bioassays cypermethrin fiproniland thiamethoxam + chlorantraniliprole reduced egg laying on both varieties and together withbuprofezin reduced nymph survival to zero Buprofezin carbofuran and cartap hydrochloride stim-ulated egg laying and carbofuran increased nymph biomass but these effects were reduced on IR62Planthopper populations were ten times higher on deltamethrin-treated rice than untreated rice in ascreenhouse experiment Host resistance failed to buffer against this insecticide-induced resurgencein BPH and WBPH However IR62 reduced the effect in GLH Rice treated with cypermethrin andfipronil had reduced yields compared to untreated controls suggesting possible phytotoxic effectsWe found little evidence of synergies between the two management practices but host resistance didbuffer against the undesirable effects of some insecticides
Keywords cypermethrin deltamethrin hormesis resurgence secondary outbreaks yield
1 Introduction
The brown planthopper Nilaparvata lugens (Staringl)[BPH] is among the most damagingpests of rice in Asia [1] BPH occurs throughout the year in tropical Asia where farmerscan produce between one and three rice crops per year At optimal temperatures theplanthopper completes about three generations per crop thereby rapidly building uppopulations in regions of intensive rice production [12] Furthermore BPH populationsrespond to high levels of nitrogenous fertilizers by increasing feeding efficiency growthrates development rates and reproductive outputmdashthereby further enhancing populationgrowth [34] During the northern hemisphere spring planthoppers migrate from tropicalAsia to Korea Japan and Northeastern China where large numbers of planthoppers cansometimes overwhelm natural enemies thereby causing extensive damage to early-seasonrice crops in these regions [5ndash7]
The management of BPH has relied heavily on two principal methods firstly publicresearch has focused on developing rice varieties with resistance (ie host plant resistance
Crops 2021 1 166ndash184 httpsdoiorg103390crops1030016 httpswwwmdpicomjournalcrops
Crops 2021 1 167
[HPR]) to the planthopper based on conventional crosses between naturally resistanttraditional landraces and high-yielding rice varieties [8] Resistant varieties have beendeployed to farmersrsquo fields since the 1970s however BPH populations can often rapidlyadapt to this resistancemdasha phenomenon known as lsquovirulence adaptationrsquo [9ndash11] Currentlythere are several resistance genes available from rice landraces and wild rice species suchthat virulence adaptation can now be better managed [12] The second most prominentBPH management method is the use of chemical insecticides [21314] The development ofinsecticides is mainly conducted through private companies Beginning in the early 2000slarge increases in the trade and use of insecticides have been reported throughout Asia [2]Furthermore trends suggest that farmers increasingly apply insecticides prophylacticallyand these applications are not in response to observed high pest densities as recommendedby Integrated Pest Management (IPM) principals [21516] Several detailed studies haveshown that some commonly-used insecticides actually enhance the growth rates of BPHpopulations and in many cases will ultimately lead to localized outbreaks of BPH infarmersrsquo fields [17ndash27] The deployment of resistant rice and the prophylactic applicationof insecticides might therefore be antagonistic to achieving successful BPH managementFurthermore where farmers conduct prophylactic insecticide applications HPR cannotachieve one of its principal objectives that of reducing pesticide inputs and maintaininga healthy rice environment [12] Indeed in such cases resistance can become simply anlsquoinsurancersquo against insecticide-induced BPH outbreaks and may inadvertently contributeto an overuse of insecticides
The mechanisms by which insecticides lead to BPH resurgence have received consid-erable research attention Recently Wu et al (2020) [27] reviewed much of the availableinformation Resurgence has been noted across a wide range of insecticide products thathave two main effects in the rice ecosystem Firstly certain insecticides enhance the feedingrates growth rates development reproductive output and migration potential of BPHeither through direct effects on planthopper physiology known as hormesis or indirectlyby inducing susceptibility in the host plant (ie by lowering phenol concentrations andenhancing sugar mobilization) [1828] Secondly many insecticides will have a greatereffect in reducing natural enemy populations than in reducing BPH populations therebydepleting the pest regulation capacity of the rice ecosystem [29ndash34] The higher susceptibil-ity of natural enemies to insecticides compared to BPH and other pests has been attributedto a greater mobility of natural enemies and thereby higher probabilities of exposureto the insecticides furthermore many predatory and parasitic arthropods have slowerpopulation growth rates than BPH [35] which reduces the potential for natural enemies todevelop insecticide resistance
Horgan (2018) [12] reviewed studies where insecticides were applied to both resistantand susceptible rice varieties to compare effects In many cases insecticides reduced theeffectiveness of resistance however relative resistance was often maintained across testvarietiesmdashalbeit with some minor exceptions [283637] However only ten publishedstudies have examined the combined effects of host resistance and insecticides on BPHpopulation development [18203137ndash43] a further three studies examined effects on thewhite-backed planthopper Sogatella furcifera Horvaacuteth [WBPH] an increasingly commonpest of rice in Asia [283644] Only seven of these studies have been published since 2000and to our knowledge there have been no published studies since 2018 that compare theactions of insecticides on BPH-susceptible and resistant rice (based on records retrievedusing Google Scholar) As a consequence information on interactions between currently-effective BPH resistance genes and commonly used insecticides is largely unavailableBased on the materials (rice varieties and insecticides) used in previous studies there isclearly a need to further examine the possible synergistic andor antagonistic effects ofcombining host resistance and insecticides for BPH management
In the present study we examine the effects of seven widely available insecticideson the integrity of resistance in rice against BPH We selected a rice variety IR62 withrelatively strong resistance against BPH populations from across Asia [1145] This variety
Crops 2021 1 168
possesses the Bph3 and Bph32 resistance genes derived from the Indian landrace PTB33Because of its relatively high effectiveness IR62 and doner varieties with the Bph3 locuscontaining either the Bph3 or Bph32 resistance genes have been extensively studied inrecent years [46ndash48] We also selected insecticides that are currently used to manage insectherbivores in rice in Asia Many of the products are not specifically recommended for useagainst BPH however resurgence is often the result of applications against other insectpests (ie BPH is a secondary pest) and particularly against rice leaffolders [49] Weexamined the indirect plant-mediated effects of insecticides on BPH because our focuswas on possible insecticide-plant interactions as well as on direct insecticide effects onBPH Therefore we conducted our experiments in greenhouses and in a screenhousethat were largely free of natural enemies Furthermore because of possible phytotoxiceffects of insecticides on rice plants [5051] and the potential role of such phytotoxicityin determining BPH-rice interactions [49] we also assessed insecticide effects on plantgrowth and yields in the screenhouse We discuss our results in terms of applying HPRand insecticide treatments according to IPM principles
2 Materials and Methods21 Herbivore Species
BPH were obtained from colonies reared at the International Rice Research Institute(IRRI) in the Philippines Planthoppers were initially collected in 2009 from rice fields inLaguna Province The founder population (ca 500 individuals) was placed in wire meshcages of 120 times 60 times 60 cm (H timesW times L) under greenhouse conditions (temperatures [T]25ndash37 C 70ndash90 relative humidity [RH] 12h12h daynight [DN]) and was continuouslyreared on ge30 days after sowing [DAS] rice plants (variety = TN1) Feeding plants werereplaced every two weeks The colony received periodic (each year) introgressions ofwild-caught individuals from the same collection sites BPH from the Laguna Provincehave been screened for virulence and are largely adapted to feed on rice varieties with theBph1 bph2 bph5 bph7 Bph18 Bph25 and Bph26 genes [1145] The colonies were also testedfor levels of resistance to insecticides and have moderate resistance to imidacloprid andhigh resistance to BPMC [52]
22 Plant Materials
We used two varieties in our experiments IR62 was selected as a variety with highresistance to BPH populations throughout South and Southeast Asia [53] Resistance inIR62 was derived from PTB33 and is attributed to the Bph3Bph32 resistance genes [4853]BPH has reduced feeding lower weight gain slower development and reduced egg-layingon IR62 compared to susceptible varieties [4] The variety also has moderate resistanceto WBPH and to the green leafhopper Nephotettix virescens (Distant) [GLH] [4554] Fur-thermore resistance derived from PTB33 and associated with the Bph3 locus appears tobe highly durable since BPH populations with virulence against PTB33-derived varietiesincluding IR62 are still rare despite over 30 years of deployment in regions of Mindanao(Philippines) and in Cambodia [55] The variety IR64 was selected as a susceptible vari-ety IR64 has the Bph1 gene for resistance but BPH populations throughout Asia haveadapted to this gene [1153] Nevertheless IR64 has additional quantitative resistance thatcontributes to tolerance (ie a relatively high capacity to withstand damage) of BPH inthe variety [56] IR64 was released by IRRI in 1985 and is considered a mega-variety inAsia [57] Because of its popularity the variety has been enhanced using marker-assistedbreeding to incorporate traits such as flood tolerance (ie IR64-Sub1 released in 2009) [57]For our experiments seeds of both varieties were acquired from the IRRI-germplasm bank
23 Insecticides
We used seven insecticides in our experiments The insecticides were selected basedon their availability at IRRI Buprofezin (2-tert-butylimini-3-isopropyl-5-phenyl-3456-tetrahydro-2H-135-thiadiazin-5-one) (25SC 200 mL haminus1) contains the growth regulator
Crops 2021 1 169
thiadezine and is used against planthoppers and leafhoppers in rice It is the recommendedinsecticide to combat high densities of planthoppers in rice because it has relatively lowtoxicity to natural enemies [58] We used two synthetic pyrethroids cypermethrin (cyan-(3-pheboxyphenyl)-methyl-(3-CS2-dichloroethenyl)-22-dimethyl cyclopropane carboxy-late) (10EC 800 mL haminus1) and deltamethrin ((S)-cyano-3-phenoxybenzyl (1R3R)-3-(22-dibromovinyl)-22-dimethyl-cyclopropane-carboxylate) (25EC 500 mL haminus1) cypermethrinhas been commonly used in rice production to combat rice tungro virus that is transmittedby rice leafhoppers [59] Deltamethrin is a broad-spectrum insecticide that is also some-times used to manage leafhoppers Both pyrethroids were previously noted to cause BPHresurgence [396061] and are currently not recommended for use in rice [62] Neverthe-less cypermethrin and deltamethrin are still commonly used on rice in some countriesincluding Indonesia and Bangladesh [131461] For this reason we included both productsin our experiments We used the carbamate insecticide carbofuran (22-Dimethyl-23-dihydro-1-benzofuran-7-flmethyl-carbamate) (3G 3 kg haminus1) as a systemic insecticidethat is sometimes recommended for control of rice pests including planthoppers [6364]Carbofuran has been banned in the European Union the USA and Canada but is still usedin much of Asia [65] We used thiamethoxam 20 + chlorantraniliprole 20 (3-[(2-chloro-13-thiazol-5-yl)methyl]-5-methyl-135-oxadiazinan-4-ylidene nitramide + 5-bromo-N-[4-chloro-2-methyl-6-(methylcarbamoyl)phenyl]-2-(3-chloropyridin-2-yl)pyrazole-3-carboxamide) (40 WG75 g haminus1) ie a combined neonicotinoid and diamide product as a further systemic in-secticide The product is absorbed through the plant to give broad-spectrum control of ricepests [66] Finally we used fipronil (5-Amino-1-[26-dichloro-4-(trifluoromethyl)phenyl]-4-(trifluoromethanesulfinyl)-1H-pyrazole-3-carbonitrile) (50SC 500 mL haminus1) and car-tap hydrochloride (S-[3-carbamoylsulfanyl-2-(dimethylamino)propyl] carbamothioate-hydrochloride) (a nereistoxin analogue) (50SP 400 g haminus1) as two broad-spectrum in-secticides commonly used to control leaffolders in Asian rice [67ndash70] All insecticideswere applied at recommended rates with quantities estimated based on the surface soilarea of each experimental unit Insecticides were applied by trained pesticide applicatorsusing standard protective equipment in a well-ventilated environment (an insect screen-house) Any evaluations of treated rice plants were conducted at a minimum of 3 dayspost-application
24 Effects of HPR and Insecticides on BPH Preferences
We assessed the effects of the insecticides on oviposition by BPH in a multichoice andin binary choice bioassays For both tests seedlings of IR62 and IR64 were planted individ-ually in 0 pots (5times 5 cm Htimes Diameter) in a greenhouse (T = 25ndash37 C RH = 70ndash90 DN= 12 h12 h) At 27 days after sowing (DAS) the seedlings were treated with one of eachof the seven insecticides as described above or were sprayed with water (ie untreatedcontrol) We used water as a control because some of the products were diluted with waterPlants were returned to the greenhouse at 30 DAS For each variety in the multichoicebioassay eight plants at 30 DASmdashone for each treatmentmdashwere placed together in a wiremesh cage of 120 times 60 times 60 cm (H times W times L) The plants were arranged in a circularconfiguration each separated by 10 cm such that adjacent plants did not touch each otheror the cage walls The cages were replicated five times for each rice variety with plantspositioned randomly during each replication Twenty gravid female N lugens were placedat the center of each cage and allowed to lay eggs for 3 days after which all adults weremanually removed from the cages Eggs were allowed to develop in the plants for a further4 days after which the plants were collected wrapped in moistened tissue paper andstored in a refrigerator Within 5 days all plants were dissected under a stereo microscope(Nikon SMZ-2B Tokyo Japan) to count the egg clusters Egg clusters were removed fromthe plant tissues and the numbers of eggs per cluster were recorded The total number ofeggs was estimated as the sum of eggs per plant from all clusters
For the binary choice tests one plant (either IR62 or IR64) for each insecticide treatmentwas placed together with a control (untreated) plant of the same variety in an acetate cage
Crops 2021 1 170
of 30 times 25 times 50 cm (H timesW times L) (ie two plants per cage) This produced 14 treatmenttimes variety combinations (7 insecticides times 2 varieties) Each combination of plant pairswas replicated three times with cages positioned randomly on greenhouse benches andreplicates of each combination pair placed in separate greenhouse compartments Fivegravid females were placed between the two pots in each cage and allowed to move withinthe cage and lay eggs After 3 days all adults were manually removed from the cages Theplants were collected 4 days after removal of the adults to allow the eggs to develop Plantswere placed in moist tissue paper and stored in a refrigerator until clusters and eggs werecounted under a stereomicroscope (as described above) For both bioassays the replicateswere each placed in separate greenhouse compartments (ie 5 or 3 blocks)
25 Effects of HPR and Insecticides on BPH Fitness
We conducted two BPH antibiosis tests in the greenhouse We examined egg layingby gravid females on each variety and for each insecticide treatment using an ovipositiontest At 30 DAS treated and control plants were individually caged by placing an acetatetube 45 times 5 cm (H times Diameter) with a mesh top over each pot A recently mated gravidBPH was placed inside each cage through a slit in the acetate and was allowed to feed andoviposit After 3 days the females were manually removed from the cages Plants were leftfor 4 days to allow eggs to develop The plants were then collected and dissected under astereo microscope to count the clusters and eggs (as described above)
We examined the effects of host resistance and insecticide treatments on BPH nymphsin a survival test At 30 DAS treated and control plants were individually caged asdescribed above Recently emerged nymphs (le2 instar) were collected from the colonyusing a suction-pooter and placed inside the cages at a density of ten per cage through aslit in the acetate The nymphs were allowed to feed and develop for 15 days after whichthe number of survivors and their developmental stages were recorded For both bioassaysplants were dried in a forced draft oven at 60 C for 7 days and were weighed For thesurvival bioassay the nymphs were also dried and weighed to estimate dry biomass andthereby control for varying consumption of plant liquids Both bioassays were replicatedfive times (ie 8 treatments times 2 varieties times 5 replicates = 80 pots per bioassay) with eachreplicate placed in a separate greenhouse compartment (ie 5 blocks)
26 Effects of HPR and Insecticides in a Screenhouse Environment
The combined effects of insecticide treatments and HPR were investigated using riceplants grown in an insect-proof screenhouse The screenhouse consisted of six concretebays (dimensions 266times 25 m (LtimesW)) filled with flooded paddy soil Temperatures in thescreenhouse varied between 21 and 34 C and RH varied between 70 and 90 Rainwatercould pass through the screenhouse roof during the experiment 160 mm of precipitationwas recorded representing the Philippines dry season The screenhouse had no artificiallighting (ie DN = 12 h12 h)
In one of the bays translucent plastic barriers were erected to delimitate five exper-imental blocks of 150 times 300 m Each block was further divided into eight cubicles of075 m2 using the same plastic materials The barriers were supported by bamboo stakesand reached a height of 150 m and a depth into the paddy soil of approx 030 m After thebarriers were installed two 23 DAS seedlings each of IR62 and IR64 were randomly trans-planted to each cubicle at 25 times 25 cm spacing We used a standard fertilizer regime to sim-ulate farmersrsquo fields The seedlings were treated with ammonium sulphate at 100 kg haminus1
split equally into three applications (ie one basal and two top dressings) Solophos andmuriate of potash was also applied at 20 kg haminus1 as a basal application Plants were contin-uously watered and the cubicles were manually weeded until rice plants were harvestedCubicles in each of the five blocks received one of the eight treatments (seven insecticidesand a control) Treatments were randomly assigned to cubicles within blocks The insecti-cide applications were made at 30 and 60 DAS to simulate farmer practices under intensive
Crops 2021 1 171
rice production systems where farmers apply insecticides ge two times [14] however inour experiment we did not mix insecticides
Rice herbivores including BPH WBPH and GLH occurred at low densities in thescreenhouse Therefore at the opposite end of the screenhouse (separated by a distanceof 20 m) a bay with gt50 DAS TN1 rice plants was infested with high densities of BPH(approx 200 BPH plantminus1) This acted as a potential source of planthoppers to attackthe treated IR62 and IR64 plants in our experiment Natural enemies were largely absentfrom the screenhouse because of the insect-proof cladding (1 mm times 1 mm mesh bore)Microparasitoids (Oligosita spp and Anagrus spp) that could pass through the screenoccasionally attacked planthopper eggs in the screenhouse (estimated 5ndash10 of eggsparasitized unpublished data)
The IR62 and IR64 plants were continuously monitored to assess grain maturationWhen about 85 of the rice grain was mature any arthropods on the plants were collectedusing a vacuum sampler and stored in plastic vials The plants were then destructivelyharvested by carefully pulling the plant from the soil to maintain intact roots The plantroots were washed under running water and each plant was placed inside a paper bagThe plants and arthropods were then dried in a forced draft oven at 60 C for 7 days andweighed Arthropods were separated by species and counted The plants were separatedinto roots shoots and panicles and the numbers of reproductive and vegetative tillerswere recorded Rice grain was removed from the panicles and separated as filled andnon-filled grain The dried grain was counted and weighed
27 Data Analyses
The numbers of egg clusters and eggs laid in rice plants during the multichoice testwere analyzed using univariate general linear models (GLM) on ranked (within replicate)data Separate analyses were conducted for IR62 and IR64 plants Data from the binarychoice bioassays were analyzed using paired t-tests The numbers of egg clusters and eggslaid in treated and non-treated plants during the oviposition bioassay were compared usingunivariate GLM with plant biomass initially included as a covariate but later removedbecause it had no significant effect Cluster and egg numbers were ranked within blocksbefore analyses Average cluster size was estimated by dividing the number of eggs in eachplant by the number of clusters Cluster size was analyzed using univariate GLM removingthe block effect Similarly the proportions of surviving nymphs and nymph biomass fromthe survival bioassay were analyzed using univariate GLMs initially including plantbiomass as a covariate Data from the survival bioassay were ranked before analysesToo few nymphs survived under some treatments to examine development rates or thesex of surviving adults We estimated the combined effects of HPR and each insecticidelsquoxrsquo as -((fitness on untreated IR64 minus fitness on x-treated IR62)fitness on untreated IR64)To estimate the effect of the insecticide alone we removed the effect on HPR which weestimated as fitness on untreated IR62 minus fitness on untreated IR64 Fitness was assessedfrom the oviposition bioassay as the numbers of eggs laid and from the survival bioassayas nymph biomass Where nymph mortality on insecticide-treated IR64 plants approached100 HPR effects were regarded as insignificant
We also examined combined and insecticide effects from the screenhouse data basedon densities of BPH and WBPH (see below) Effects were calculated using insecticide andHPR combinations for each replicated block (N = 5) Estimated effects were comparedacross treatments using univariate GLMs Because three herbivores (BPH WBPH andGLH) occurred in our screenhouse plots we used a split-plot multivariate GLM to analyzeherbivore densities Total planthopper and leafhopper biomass and density and plantgrowth parameters (tiller number biomass yield grain number proportion of grainunfilled and grain size) were analyzed using split-plot univariate GLMs The covariateplant biomass was initially included in analyses of the effects on herbivores but had noeffect and was subsequently removed Similarly the covariate herbivore biomass wasinitially included in analyses of plant growth parameters but had no effect and was
Crops 2021 1 172
subsequently removed The combined effects of insecticide and HPR and the separateeffects of each insecticide (calculated as indicated above) on BPH and WBPH were alsoanalyzed using univariate GLM
Post-hoc Tukey tests were applied to all significant treatment factors Residuals wereplotted following all parametric analyses and were found to be normal and homogenous
3 Results31 Effects of HPR and Insecticides on BPH Preferences
Treatments affected the number of clusters and eggs that were laid in IR62 plants(ranked clusters and eggs F732 = 4738 p lt 0001) and in IR64 plants (clusters F732 = 6229p lt 0001 eggs F732 = 7757 p lt 0001) (Figure 1AB) Fewer eggs were laid on IR64 plantstreated with the pyrethroids (cypermethrin and deltamethrin) fipronil and thiamethoxam-chlorantraniliprole compared to plants treated with buprofezin carbofuran and cartaphydrochloride (Figure 1B) Cypermethrin-treated IR62 plants also had fewer eggs thanbuprofezin-treated IR62 plants (Figure 1A) Although BPH females tended to lay fewereggs in many of the insecticide-treated rice plants in the binary choice experiments therewere no statistically significant differences between the numbers of eggs laid in treated-IR62(t ranges from minus0746 to 3186 p gt 005) or treated-IR64 (t ranges from minus0108 to 2789p gt 005) compared to corresponding non-treated plants (Figure 1CD)
Crops 2021 1 FOR PEER REVIEW 7
herbivore densities Total planthopper and leafhopper biomass and density and plant growth parameters (tiller number biomass yield grain number proportion of grain un-filled and grain size) were analyzed using split-plot univariate GLMs The covariate plant biomass was initially included in analyses of the effects on herbivores but had no effect and was subsequently removed Similarly the covariate herbivore biomass was initially included in analyses of plant growth parameters but had no effect and was subsequently removed The combined effects of insecticide and HPR and the separate effects of each insecticide (calculated as indicated above) on BPH and WBPH were also analyzed using univariate GLM
Post-hoc Tukey tests were applied to all significant treatment factors Residuals were plotted following all parametric analyses and were found to be normal and homogenous
3 Results 31 Effects of HPR and Insecticides on BPH Preferences
Treatments affected the number of clusters and eggs that were laid in IR62 plants (ranked clusters and eggs F732 = 4738 p lt 0001) and in IR64 plants (clusters F732 = 6229 p lt 0001 eggs F732 = 7757 p lt 0001) (Figure 1AB) Fewer eggs were laid on IR64 plants treated with the pyrethroids (cypermethrin and deltamethrin) fipronil and thiameth-oxam-chlorantraniliprole compared to plants treated with buprofezin carbofuran and cartap hydrochloride (Figure 1B) Cypermethrin-treated IR62 plants also had fewer eggs than buprofezin-treated IR62 plants (Figure 1A) Although BPH females tended to lay fewer eggs in many of the insecticide-treated rice plants in the binary choice experiments there were no statistically significant differences between the numbers of eggs laid in treated-IR62 (t ranges from minus0746 to 3186 p gt 005) or treated-IR64 (t ranges from minus0108 to 2789 p gt 005) compared to corresponding non-treated plants (Figure 1CD)
Figure 1 Results from multichoice (AB) and binary choice (CD) bioassays conducted in the green-house Bioassays were conducted separately for IR62 (AC) and IR64 (BD) Plants were treated with one of seven insecticides (A to G) as identified in the legend Control plants were sprayed with water Error bars are presented (N = 5 for (A) and (B) N = 3 for (C) and (D)) Lowercase letters (andashc) indicate homogenous treatment groups (Tukey p lt 005)
A B C D E F G0
100
200
300
400
Treatment
A B C D E F G
Non treatedTreated
A B C D E F G Control
Num
ber o
f egg
s pe
r pla
nt
0
100
200
300
400
A B C D E F G Control
IR62 IR64
A
C
B
D
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliproleH Control
a abab
c
abc
abc bc bc
c
aaa
ab
ab
bb b
Figure 1 Results from multichoice (AB) and binary choice (CD) bioassays conducted in thegreenhouse Bioassays were conducted separately for IR62 (AC) and IR64 (BD) Plants were treatedwith one of seven insecticides (A to G) as identified in the legend Control plants were sprayed withwater Error bars are presented (N = 5 for (AB) N = 3 for (CD)) Lowercase letters (andashc) indicatehomogenous treatment groups (Tukey p lt 005)
32 Effects of HPR and Insecticides on BPH Fitness
More eggs were laid on IR64 plants than on IR62 plants but BPH produced similarnumbers of egg clusters on both plants The lower numbers of eggs in IR62 were due tosmaller clusters on the variety (Table 1) The numbers of eggs and clusters were affectedby insecticide treatment with fewer eggs and clusters on cypermethrin-treated fipronil-treated and thiamethoxam-chlorantraniliprole-treated plants than on plants under the
Crops 2021 1 173
remaining treatments (Table 1) The treatment-effects were not affected by host variety (ieinteractions were non-significant Table 1) Insecticide treatments had no effect on clustersize in the bioassay (Table 1) There was no significant effect of combined insecticide andHPR (F628 = 0449 p = 0840) on observed changes in egg laying during the bioassay butthe estimated insecticide effect was greater for thiamethoxam-chlorantraniliprole fiproniland cypermethrin than for buprofezin cartap hydrochloride and carbofuran (F628 = 3140p = 0018 Figure 2A)
Table 1 Results from greenhouse oviposition and survival tests with IR62 (resistant) and IR64 (susceptible) each treatedwith one of seven insecticides Numbers are means plusmn SEM (N = 5)
Treatments Egg Clusters(Number Plantminus1) 12
Eggs(Number Plantminus1) 12
Cluster Size(Number of Eggs) 1
Nymph Survival(Proportions) 12
Nymph Biomass (mgDry Weight Plantminus1) 12
IR62Buprofezin 1240 plusmn 140 b 5040 plusmn 904 b 393 plusmn 036 000 plusmn 000 a 000 plusmn 000 a
Carbofuran 1300 plusmn 089 b 5380 plusmn 421 b 417 plusmn 026 064 plusmn 064 d 154 plusmn 154 d
Cartaphydrochloride 1360 plusmn 075 b 5200 plusmn 486 b 382 plusmn 029 028 plusmn 028 b 151 plusmn 151 b
Cypermethrin 620 plusmn 169 a 2000 plusmn 608 a 316 plusmn 023 002 plusmn 002 a 002 plusmn 002 a
Deltamethrin 1360 plusmn 160 b 4340 plusmn 566 b 338 plusmn 069 040 plusmn 040 c 120 plusmn 120 c
Fipronil 520 plusmn 146 a 2240 plusmn 735 a 393 plusmn 064 000 plusmn 000 a 000 plusmn 000 a
Thiamethoxam-chlorantraniliprole 380 plusmn 086 a 1700 plusmn 435 a 447 plusmn 032 000 plusmn 000 a 000 plusmn 000 a
Control 1260 plusmn 103 b 4900 plusmn 716 b 388 plusmn 054 080 plusmn 080 d 292 plusmn 292 d
IR64Buprofezin 2100 plusmn 164 10940 plusmn 1346 556 plusmn 117 000 plusmn 000 000 plusmn 000Carbofuran 1820 plusmn 404 9880 plusmn 2939 524 plusmn 074 086 plusmn 086 420 plusmn 420
Cartaphydrochloride 2140 plusmn 619 11460 plusmn 3141 549 plusmn 064 010 plusmn 010 045 plusmn 045
Cypermethrin 380 plusmn 233 1980 plusmn 1155 550 plusmn 058 000 plusmn 000 000 plusmn 000Deltamethrin 1560 plusmn 209 7500 plusmn 1581 473 plusmn 069 040 plusmn 040 201 plusmn 201
Fipronil 380 plusmn 086 2100 plusmn 552 546 plusmn 080 000 plusmn 000 000 plusmn 000Thiamethoxam-chlorantraniliprole 260 plusmn 075 800 plusmn 212 373 plusmn 133 000 plusmn 000 000 plusmn 000
Control 1740 plusmn 364 7520 plusmn 2344 426 plusmn 073 086 plusmn 086 360 plusmn 360F-treatment 3 19898 19898 0541 ns 133811 108451
F-variety 3 2596 ns 7607 7471 1080 ns 0008 ns
F-interaction 3 1840 ns 2034 ns 0610 ns 4902 9496 1 Lowercase letters (andashd) indicate homogenous treatment groups (for IR62 and IR64 combined) 2 Data were ranked within blocks beforeanalysis 3 DF = 164 for varieties 764 for treatments and interactions ns = p gt 005 = p le 001 = p le 0001
Nymph survival and biomass were not affected by rice variety (Table 1) Buprofezinfipronil thiamethoxam-chlorantraniliprole and cypermethrin reduced survival to 0 in thebioassay and all insecticides except carbofuran resulted in lower survival and biomass thanon untreated plants (Table 1) There were significant interactions because of higher nymphmortality and a resulting lower biomass on carbofuran-treated IR62 compared to untreatedIR62 but lower mortality and a higher biomass of nymphs on carbofuran-treated IR64than on untreated IR64 (Table 1) There was no significant effect of insecticide treatment(F628 = 0946 p = 0479) on observed changes in nymph biomass during the bioassaybut the estimated combined effects of pesticide and HPR was greater for thiamethoxam-chlorantraniliprole fipronil cypermethrin and buprofezin than for cartap hydrochlorideand carbofuran (F628 = 5699 p = 0001) (Figure 2B)
Crops 2021 1 174
Crops 2021 1 FOR PEER REVIEW 9
Figure 2 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) egg laying and (B) nymph biomass in greenhouse bioas-says Effects are indicated as proportional changes in fitness parameters compared to untreated IR64 Negative numbers indicate reduced fitness Standard errors are indicated (N = 5) Lowercase letters (ab) indicated homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 1)
Nymph survival and biomass were not affected by rice variety (Table 1) Buprofezin fipronil thiamethoxam-chlorantraniliprole and cypermethrin reduced survival to 0 in the bioassay and all insecticides except carbofuran resulted in lower survival and biomass than on untreated plants (Table 1) There were significant interactions because of higher nymph mortality and a resulting lower biomass on carbofuran-treated IR62 compared to untreated IR62 but lower mortality and a higher biomass of nymphs on carbofuran-treated IR64 than on untreated IR64 (Table 1) There was no significant effect of insecticide treatment (F628 = 0946 p = 0479) on observed changes in nymph biomass during the bio-assay but the estimated combined effects of pesticide and HPR was greater for thiameth-oxam-chlorantraniliprole fipronil cypermethrin and buprofezin than for cartap hydro-chloride and carbofuran (F628 = 5699 p = 0001) (Figure 2B)
33 Effects of HPR and Insecticides on Planthoppers and Leafhoppers Plants in the screenhouse were colonized by BPH WBPH and GLH Densities for
each species were similar on IR62 and IR64 (Wilkrsquos λ = 0994 p = 0952) There was a sig-nificant treatment effect on all three species (Wilkrsquos λ = 0586 p = 0049) because of high densities on deltamethrin-treated plants (Table 2) The numbers and biomass of all three species combined were also greater on deltamethrin-treated plants than on the remaining treatments (Table 2) Host resistance reduced GLH densities under the deltamethrin treat-ment but had no apparent effect on densities under the other treatments (Table 2) There were significant combined effects of HPR and insecticide on the numbers of BPH (F624 = 2686 p = 0039) and WBPH (F624 = 2482 p = 0052) this was almost exclusively due to the resurgence effect of deltamethrin on the planthoppers (BPH F624 = 2686 p = 0039 WBPH F624 = 2482 p = 0052) (Figure 3)
Δ eg
gs(P
ropo
rtion
)
-10
-08
-06
-04
-02
00
02
04
06
Treatment
A B C D E F G
Δ N
ymph
bio
mas
s(P
ropo
rtion
)
-10
-08
-06
-04
-02
00
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
ab
a a
b
a
b b
a a a a a a a a
b b bb
bb
Figure 2 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone(P grey bars) on (A) egg laying and (B) nymph biomass in greenhouse bioassays Effects are indicated as proportionalchanges in fitness parameters compared to untreated IR64 Negative numbers indicate reduced fitness Standard errors areindicated (N = 5) Lowercase letters (ab) indicated homogenous treatment groups based on Tukey pairwise comparisons(p lt 005) (see also Table 1)
33 Effects of HPR and Insecticides on Planthoppers and Leafhoppers
Plants in the screenhouse were colonized by BPH WBPH and GLH Densities foreach species were similar on IR62 and IR64 (Wilkrsquos λ = 0994 p = 0952) There was asignificant treatment effect on all three species (Wilkrsquos λ = 0586 p = 0049) because of highdensities on deltamethrin-treated plants (Table 2) The numbers and biomass of all threespecies combined were also greater on deltamethrin-treated plants than on the remainingtreatments (Table 2) Host resistance reduced GLH densities under the deltamethrintreatment but had no apparent effect on densities under the other treatments (Table 2)There were significant combined effects of HPR and insecticide on the numbers of BPH(F624 = 2686 p = 0039) and WBPH (F624 = 2482 p = 0052) this was almost exclusively dueto the resurgence effect of deltamethrin on the planthoppers (BPH F624 = 2686 p = 0039WBPH F624 = 2482 p = 0052) (Figure 3)
Crops 2021 1 175
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants underseven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density(Number Plantminus1) 1
WBPH Density(Number Plantminus1) 1
GLH Density(Number Plantminus1) 1
Density ofPlanthoppers and
Leafhoppers(Number Plantminus1) 1
Biomass ofPlanthoppers and
Leafhoppers(mg Dry Weight) 1
IR62Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a
Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartaphydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a
Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a
Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a
Thiamethoxam-chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a
IR64Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartaphydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123
Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns
F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Crops 2021 1 FOR PEER REVIEW 10
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants under seven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density (Num-
ber Plantminus1) 1 WBPH Density
(Number Plantminus1) 1 GLH Density
(Number Plantminus1) 1
Density of Planthoppers and Leafhoppers (Number
Plantminus1) 1
Biomass of Planthoppers and Leafhoppers (mg Dry
Weight) 1 IR62
Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartap hydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a Thiamethoxam-
chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a IR64
Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132 Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartap hydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123 Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113 Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052 Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087 F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns 1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatments and interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in the screenhouse Effects are indicated as proportional changes in planthopper densities compared to untreated IR64 Negative numbers indicate reduced densities positive numbers are increased den-sities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
Δ BP
H(N
umbe
r pla
nt-1
)
0
5
10
15
20
Treatment
A B C D E F G
Δ W
BPH
(Num
ber p
lant
-1)
0
20
30
40
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
a a
aa a a
a a a
a
a a
b
b
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimatedpesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in thescreenhouse Effects are indicated as proportional changes in planthopper densities compared to
Crops 2021 1 176
untreated IR64 Negative numbers indicate reduced densities positive numbers are increaseddensities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenoustreatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
34 Direct and Indirect Effects of Insecticides on Rice Development
IR62 plants had more tillers and attained a greater biomass than IR64 plants in thescreenhouse (Table 3) IR62 grain matured faster and plants were harvested earlier than theIR64 plants (Table 3) IR62 plants also produced more grain (number of grain) and hadhigher yields than IR64 but had smaller grain and more unfilled grain than IR64 (Table 3)
Table 3 Rice growth parameters for IR62 (resistant) and IR64 (susceptible) under seven insecticide treatments and forcontrol non-treated plants in a screenhouse environment Numbers are means plusmn SEM
Treatment Tiller Number 1 Plant Biomass(g Dry Weight) 1
Time to Harvest(Days) 1
Weightof Filled Grain(g Dry Weight) 1
Number ofFilled Grain
(1000s) 1
Unfilled Grain(Proportion) 1
1000 GrainWeight 1
IR62Buprofezin 2660 plusmn 206 ab 4643 plusmn 385 ab 7990 plusmn 168 ab 5776 plusmn 683 ab 361plusmn 061 ab 011 plusmn 003 a 2008 plusmn 042 ab
Carbofuran 2900 plusmn 106 ab 5546 plusmn 357 b 7720 plusmn 080 ab 7448 plusmn 691 b 446plusmn 061 ab 011 plusmn 001 a 2103 plusmn 054 b
Cartaphydrochloride 2820 plusmn 327 ab 4896 plusmn 470 ab 7920 plusmn 153 b 6430 plusmn 712 ab 415plusmn 064 ab 012 plusmn 001 b 2023 plusmn 065 a
Cypermethrin 2870 plusmn 301 a 4957 plusmn 562 a 7760 plusmn 103 b 6032 plusmn 822 a 410plusmn 067 ab 014 plusmn 002 a 1965 plusmn 076 a
Deltamethrin 2620 plusmn 345 a 4592 plusmn 588 a 7840 plusmn 108 ab 5698 plusmn 832 ab 379plusmn 047 ab 012 plusmn 001 a 2030 plusmn 063 ab
Fipronil 2380 plusmn 176 ab 4607 plusmn 657 ab 7700 plusmn 179 ab 5266 plusmn 390 a 318 plusmn 032 a 013 plusmn 002 a 2128 plusmn 047 b
Thiamethoxam-chlorantraniliprole 2650 plusmn 250 b 4547 plusmn 429 b 7720 plusmn 102 a 5777 plusmn 688 ab 408 plusmn 061 b 013 plusmn 002 a 2019 plusmn 067 ab
Control 3120 plusmn 247 b 5401 plusmn 555 b 7720 plusmn 080 ab 7045 plusmn 520 b 396plusmn 036 ab 012 plusmn 002 a 2076 plusmn 050 ab
IR64Buprofezin 2340 plusmn 250 5064 plusmn 747 8350 plusmn 124 5080 plusmn 952 270 plusmn 054 008 plusmn 001 2277 plusmn 039Carbofuran 2040 plusmn 097 4591 plusmn 141 8540 plusmn 073 4951 plusmn 312 263 plusmn 023 006 plusmn 000 2295 plusmn 036
Cartaphydrochloride 2120 plusmn 075 4336 plusmn 285 8520 plusmn 140 4273 plusmn 579 238 plusmn 026 015 plusmn 007 2050 plusmn 234
Cypermethrin 1990 plusmn 089 3968 plusmn 272 8710 plusmn 135 3920 plusmn 552 250 plusmn 028 008 plusmn 002 2171 plusmn 070Deltamethrin 1980 plusmn 181 4325 plusmn 357 8460 plusmn 068 4710 plusmn 496 277 plusmn 024 008 plusmn 001 2314 plusmn 012
Fipronil 2120 plusmn 174 4697 plusmn 311 8450 plusmn 089 4888 plusmn 392 274 plusmn 027 008 plusmn 001 2247 plusmn 042Thiamethoxam-chlorantraniliprole 2520 plusmn 287 5404 plusmn 465 8380 plusmn 196 5594 plusmn 546 347 plusmn 053 010 plusmn 001 2265 plusmn 072
Control 2160 plusmn 076 4678 plusmn 342 8600 plusmn 096 4940 plusmn 543 267 plusmn 029 008 plusmn 001 2283 plusmn 049F-treatment 4100 ns 3410 2253 ns 5150 2555 3923 4924
F-variety 168232 7665 547500 104356 118242 32308 106597 F-interaction 5980 6113 4824 5760 2921 2769 2552
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Insecticides affected plant growth directly and indirectly (through increased planthop-per densities) Plants treated with cypermethrin and deltamethrin had fewer tillers had alower biomass and under cypermethrin had lower yields than untreated plants (Table 3)Plants treated with fipronil also had reduced yields due to lower grain numbers comparedto untreated plants (Table 3) Although yields were not affected treatment with cartaphydrochloride increased the proportion of unfilled grain compared to untreated plantsof both varieties Cartap hydrochloride and cypermethrin also caused a delay in grainmaturation on IR62 and IR64 respectively relative to thiamethoxam-chlorantraniliproletreated plants (Table 3)
There were several significant interactions tiller numbers on both plants were similarunder buprofezin fipronil and thiamethoxam-chlorantraniliprole treatments but wereotherwise higher on IR62 (Table 3) biomass was lower on IR64 under carbofuran andcypermethrin treatments but was otherwise similar on both plants (Table 3) buprofezindelayed grain maturation on IR62 but maturation was otherwise faster on IR62 comparedto IR64 (Table 3) yields under buprofezin fipronil and thiamethoxam-chlorantraniliprolewere similar for IR62 and IR64 but otherwise yields were higher on IR62 (Table 3) grain
Crops 2021 1 177
numbers were higher on IR62 than IR64 except under fipronil treatments (Table 3) theproportions of grain unfilled were higher on IR62 but were similar between varieties underthe fipronil treatment (Table 3) 1000-grain weight was higher in IR64 but similar on bothvarieties when treated with cartap hydrochloride (Table 3)
4 Discussion
Our results using a highly resistant rice variety and some commonly used insecticidesindicate a range of possible effects on planthoppers and rice plants by combining these twomanagement practices
41 Synergies
The most predictable effect of combining HPR and insecticides is a synergy in reduc-ing BPH numbers This is suggested because planthoppers are weakened by the hostrsquosresistance and should therefore be more susceptible to the toxic effects of insecticides [40]Using spray towers over potted plants Heinrichs et al (1984) [40] examined the combinedeffects of a range of insecticides (that included carbofuran) with resistance by comparingBPH mortality on six resistant and two susceptible rice varieties Their results indicatedthat mortality was greatest when insecticides were applied to BPH on resistant varietiesThe effects were similar when the study was repeated with WBPH for three insecticides(including carbofuran) on the resistant variety N22 (Wbph1 gene) [40] In our study hostresistance reduced egg laying and nymph biomass on non-treated IR62 compared to non-treated IR64 Egg laying was also reduced on plants treated with cypermethrin fipronilor thiamethoxam-chlorantraniliprole (Table 1) Based on the results of our choice andno-choice oviposition bioassays cypermethrin acted mainly as a deterrent of gravid fe-males whereas fipronil and thiamethoxam-chlorantraniliprole directly reduced egg layingDespite similar BPH responses to both the insecticides and resistant host there were no ap-parent synergies between any of these insecticides and resistance in reducing egg numbersin the oviposition bioassay In contrast to these three insecticides our bioassays indicatedthat buprofezin carbofuran and cartap hydrochloride can lead to higher numbers of eggson treated plants compared to untreated plants (Table 1) These effects were not statisticallysignificant and may be exaggerated because bioassays were conducted over relativelyshort time scales For example planthoppers will sometimes respond to stresses such ashigh temperatures by increasing oviposition rates [7172] Furthermore the stimulation ofegg-laying by these chemicals was not apparent in our screenhouse experiment (Table 2)Nevertheless our results do indicate that host resistance reduced the magnitude of anystimulatory effect on oviposition egg numbers on buprofezin-treated carbofuran-treatedand cartap hydrochloride-treated IR64 in the no-choice bioassay were twice as high as onthe corresponding IR62 plants In cases such as these the resistance of IR62 functions as abuffer or lsquoinsurancersquo against possible undesired pesticide effects [1238]
Although buprofezin appeared to stimulate oviposition the growth hormone had astrong impact on nymph development leading to zero survival of exposed nymphs in ourgreenhouse bioassay Cypermethrin fipronil and thiamethoxam-chlorantraniliprole alsoreduced nymph survival to zero in the greenhouse bioassay (Table 1) Where insecticideshave such high efficiencies any fitness losses to BPH due to host resistance then becomeredundant Unlike buprofezin carbofuran also stimulated nymph biomass on IR64 in thesurvival bioassay (Table 1) however nymph biomass was lower on carbofuran-treated IR62than on non-treated IR62 indicating that the hostrsquos resistance buffered against undesiredstimulatory effects on BPH and may have functioned synergistically to augment mortalityand slow BPH growth Such buffering allows insecticides to be used against other pestswithout inducing secondary outbreaks of non-target pests In our screenhouse experimentherbivore densities were generally low on both IR62 and IR64 for most treatments (exceptdeltamethrin) and were below threshold densities for insecticide applicationsmdashwhichare normally about 25ndash50 planthoppers per hill [1373] Under such low densities it wasnot possible to observe the impact of host resistance on planthopper population build-
Crops 2021 1 178
up There was some indication of higher densities on cartap hydrochloride-treated IR62in the screenhouse which may be related to the stimulatory effects of the product onegg laying and the low mortality of nymphs on cartap hydrochloride-treated plants asobserved in the greenhouse bioassays (Table 1) Field studies from the Philippines haveindicated that IR62 effectively reduces BPH densities during wet and dry seasons [4]However farmers are rarely aware of which anti-herbivore resistance their rice varietiespossess and frequently apply pesticides regardless of resistance thereby reducing theirfarm profits [3841] For example Joshi et al (1992) [41] reported widespread outbreaks ofBPH on IR62 in Mindanao Philippines after applications of decamethrin (a pyrethroid) tothe resistant variety Where such resurgence-causing insecticides are applied resistanceseems to be often incapable of reducing the stimulatory effects of the insecticide and itloses its capacity to buffer against outbreaks
42 Antagonisms
Several insecticides are associated with increasing densities of planthoppers andwith resurgence outbreaks in farmersrsquo fields [27293041496174] Indeed patches oflsquohopperburnrsquo (ie dead rice plants resulting from planthopper damage) are often anindicator of high insecticide use in tropical Asia [214] In many cases these resurgence-causing insecticides were once effective in reducing planthopper densities but over timepopulations developed resistance against the products The resulting low toxicity canthen have a stimulatory effect on planthopper feeding and development [2123242743]This may be through hormesis which is a direct response by the planthopper to low-levelexposure to the toxic chemical or it may result from physiological changes to the rice plantsthat ultimately benefit the planthopper [1849] In our screenhouse study we induced aresurgence outbreak of BPH WBPH and to a lesser extent GLH on deltamethrin-treatedrice plants (Table 2) The combined densities of all three herbivores increased almost10 times on the treated plants Furthermore herbivore biomass increased by between fourand six times Although the resurgence effects of deltamethrin have been documentedin previous studies [2029ndash313975] we could not predict from our greenhouse studiesthat any such outbreak would occur Egg laying was unaffected by deltamethrin in ourchoice and no-choice experiments and nymph survival and development both declinedon deltamethrin-treated rice in the survival bioassay (Table 1) The massive increases inplanthopper densities on deltamethrin-treated plants in our study seem to result froma heightened attraction of adult BPH and WBPH to the plants [49] particularly sincenumbers were so low on all other plants including both treated and non-treated plantsHowever once on the plants planthoppers also apparently had enhanced fitness Cuonget al (1997) [31] observed higher population densities of BPH on deltamethrin-treatedrice in Vietnam albeit with lower-level increases on resistant varieties Furthermore in agreenhouse study Azzam et al (2009) [20] found that deltamethrin reduced pre-ovipositiontimes increased the duration of oviposition and increased egg hatchability and femalelongevity Nanthakumar et al (2012) [43] further showed that deltamethrin increased BPHfeeding efficiency on resistant and susceptible rice plants as determined through honeydewproduction and population size In studies with WBPH Salim and Heinrichs (1987) [36]showed deltamethrin to increase settling (antixenosis) feeding and population growthand Suri and Singh (2011) [28] showed deltamethrin to increase WBPH reproductive ratesand nymph survival In our study the observed resurgence of planthoppers was similaron both varieties indicating as was previously noted by Joshi et al (1992) [41] thatresurgence-causing pyrethroids are antagonistic to resistance in IR62
Though not statistically significant cartap hydrochloride also increased BPH numberson IR62 but not on IR64 in our screenhouse study This indicates that the product mayalso be antagonistic to the hostrsquos resistance mechanisms The stronger effect on IR62 maybe related to the differences between IR62 biomass growth rates and anatomy (ie alarger plant with higher tillering) compared to IR64 (Table 3) Once resistance is no longereffective because of chemically-enhanced BPH fitness these plant characteristics which
Crops 2021 1 179
have been related to observed preferences in BPH and other sap-sucking insects [349]could further accelerate the build-up of BPH populations on cartap hydrochloride-treatedplants To further test this idea experiments could be conducted using near-isogenic ricelines that differ mainly in the presence or absence of tagged resistance genes [76] Suchtests would avoid confounding differences in plant anatomy between test varieties IR62resistance to BPH has been well studied [4] The variety is also resistant to WBPH atolder plant stages [54] Therefore in our study deltamethrin was also antagonistic tothis WBPH-resistance (Table 2) However BPH feeding also facilitates WBPH populationgrowth [5477] and there may have been added indirect plant-mediated effects [77] thataugmented WBPH populations on both varieties in our experiment IR62 is also resistant toGLH [45] In our experiments GLH densities also increased on deltamethrin-treated plantsbut the effect was only apparent on IR64 Therefore GLH-resistance in IR62 apparentlybuffered against deltamethrin-associated increases in GLH on the variety as was alsonoted from field studies in Vietnam [31] Deltamethrin is currently not recommended foruse in rice [202878] however the product continues to be marketed in rice-dominatedregions throughout Asia [131479] Although cypermethrin has been associated with BPHoutbreaks and was consequently removed for use on rice in Thailand and Vietnam [62]we did not observe cypermethrin-induced resurgence in our screenhouse experimentNevertheless cypermethrin has been associated with resurgence of BPH in South Asia [61]and we have noted stimulatory effects of cypermethrin on other rice herbivores in thePhilippines (Horgan unpublished data)
43 Phytotoxicity
Phytotoxicity is relatively common following applications of insecticides [182850]Phytotoxic effects may change a plantrsquos physiology without consequences for crop yieldsHowever such changes to plant physiology could have indirect effects on herbivores Forexample deltamethrin methyl parathion and quinalphos produce increases in total sugarsand reducing sugars as well as increasing the protein contents of the rice leaf sheath andleaf blade [28] Several insecticides also reduce plant phenols in ricemdashwhich increases therice plantrsquos susceptibility to BPH and other insects [27] In our screenhouse experiment wenoted several changes to rice anatomy and yields Importantly we noted that cypermethrinand fipronil both reduced rice yields (Table 3) This was not due to indirect herbivoreeffects because planthopper and leafhopper densities were consistently low on rice treatedwith these insecticides The effects therefore probably resulted from direct phytotoxiceffects on the plants Furthermore these effects were most pronounced on treated IR62plants Other minor effects on grain production included increased proportions of unfilledgrain in cartap hydrochloride-treated plants and delays in grain maturation on cartaphydrochloride-treated and cypermethrin-treated plants (Table 3) Despite the high densitiesof BPH and WBPH on deltamethrin-treated plants yields were not significantly lower thanin control non-treated plants Both IR62 and IR64 have been noted for their high toleranceto BPH damage particularly under high nitrogenous fertilizer applications [3456] as usedin our screenhouse experiment Therefore where plants have high tolerance to damageapplying insecticides could also result in profitability losses on rice farms and in the caseof insecticide-induced pest population increases tolerance might buffer against subsequentyield losses There are few reports of the phytotoxicity of insecticides to rice carbofuran istoxic to rice at high concentrations but may also enhance the growth of rice plants throughinteractions with nitrogen-fixing bacteria in the rhizosphere [80] Future studies couldmore carefully assess field-grown plants for possible phytotoxic effects particularly ifinsecticides have the potential to directly reduce yields
44 Management implications
Global trends suggest that resistance in rice has failed to reduce insecticide appli-cations [2] This may be due to aggressive marketing by agrochemical companies andstrategies to increase pesticide sales such as promoting prophylactic applications [16] Be-
Crops 2021 1 180
cause we selected our test insecticides (except the pyrethroids) without any a priori criteriaour results suggest that many currently used insecticides probably have some negativeeffects on yield either directly through phytotoxic effects or indirectly by inducing pestresurgence Furthermore our results indicate that even where pests reach high densitiesas on deltamethrin-treated plants in the screenhouse these densities do not necessarilylead to significant yield reductions
Among the products we used buprofezin although it had little effect on oviposi-tion was effective in reducing nymph survival Furthermore there were only minorapparent phytotoxic effects on the rice plants Buprofezin has a low toxicity to naturalenemies in rice fields [58] and is therefore the most compatible of the insecticides withstrategies to increase rice field biodiversity and restore ecosystem services [32ndash348182]Thiamethoxam-chlorantraniliprole was also effective in reducing both oviposition andnymph survival and had no apparent toxic effects on rice plants However thiamethoxam-chlorantraniliprole and other neonicotinoids although moderately toxic can have severeeffects on beneficial hymenopteran parasitoids [83] although they have apparently lowtoxicity to predatory mirid bugs [84] The remaining insecticides either directly reducedyields or were associated with higher BPH biomass or densities in our experiments
We noted few synergies with HPR although nymph biomass andor survival waslower in cartap hydrochloride and carbofuran-treated IR62 plants compared to correspond-ing IR64 plants Meanwhile cartap hydrochloride and deltamethrin were antagonisticto resistance in IR62 in the screenhouse experiment Among the strongest effects of HPRwas to act as a buffer against GLH population build-up on deltamethrin-treated IR62 andagainst carbofuran-induced increases in BPH nymph biomass in the survival bioassayTherefore HPR may play a role in reducing the possibilities of secondary outbreaks ofBPH and other insects The combined use of HPR and targeted insecticides against BPHcreates redundancy in the rice field and counters measures to manage insecticide resistanceand virulence by unnecessarily exposing plant protection products to evolving pest pop-ulations HPR (and host plant tolerance) can be effective against target pests as revealedin previous studies [4] and could be incorporated into other pest management practicesparticularly agroecological practices such as ecological engineering with which it is highlycompatible [818586]
5 Conclusions
Based on our results and the results of previous studies HPR and the applicationof chemical insecticides against BPH rarely produce synergistic effects Antagonisticeffects where chemicals caused increases in BPH fitness or populations on resistant ricewere more apparent in our study In particular deltamethrin caused an outbreak ofBPH in our screenhouse study Nevertheless HPR can buffer against the insecticide-induced enhancement of herbivore fitness (hormesis) and potentially reduce possibilities ofsecondary outbreaks as was suggested from results with cartap hydrochloride in our studyBased on our results we suggest that carbofuran cartap hydrochloride cypermethrindeltamethrin and fipronil should be avoided in rice particularly in the Philippines (wherethis research was conducted) due to their resurgence-causing potential or because of directreductions to rice yields
Author Contributions Conceptualization FGH methodology FGH AP-C and MLPA formalanalysis FGH and AP-C investigation FGH AP-C and MLPA data curation FGHAP-C and MLPA writingmdashoriginal draft preparation FGH and AP-C writingmdashreview andediting FGH AP-C and MLPA visualization FGH supervision FGH and MLPA projectadministration FGH funding acquisition FGH All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by the Bill and Melinda Gates Foundation (Cereal SystemsInitiative for South Asia [CSISA] OPP52303) and the Global Rice Science Platform (GRiSP) underthe directorship of Achim Dobermann AP-C was partially funded by The Spanish Governmentthrough Becas Faro International
Crops 2021 1 181
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data presented in this study are available on reasonable requestfrom the corresponding author
Acknowledgments The authors thank Fame Ramal Carmencita Bernal Alberto Naredo ReyeulQuintana Vincent Virtudes Marol Recide Jenyrose Geronda and Ellen Genil for assistance duringthis study The contributions of Rodante Abas (sadly deceased) during three decades of entomologicalresearch at IRRI are gratefully acknowledged Rice lines were supplied through the Genebank of IRRI
Conflicts of Interest The authors declare no conflict of interest
References1 Bottrell DG Schoenly KG Resurrecting the ghost of green revolutions past The brown planthopper as a recurring threat to
high-yielding rice production in tropical Asia J Asia-Pac Entomol 2012 15 122ndash140 [CrossRef]2 Horgan FG Integrated pest management for sustainable rice cultivation A holistic approach In Achieving Sustainable Cultivation
of Rice Cultivation Pest and Disease Management Sasaki T Ed Burleigh Dodds Scientific Cambridge UK 2017 pp 309ndash3423 Horgan FG de Freitas TFS Crisol-Martiacutenez E Mundaca EA Bernal CC Nitrogenous fertilizer reduces resistance but
enhances tolerance to the brown planthopper in fast-growing moderately resistant rice Insects 2021 12 989 [CrossRef] [PubMed]4 Horgan FG Pentildealver Cruz A Bernal CC Ramal AF Almazan MLP Wilby A Resistance and tolerance to the brown
planthopper Nilaparvata lugens (Staringl) in rice infested at different growth stages across a gradient of nitrogen applicationsField Crops Res 2018 217 53ndash65 [CrossRef]
5 Hereward JP Cai X Matias AMA Walter GH Xu C Wang Y Migration dynamics of an important rice pest The brownplanthopper (Nilaparvata lugens) across AsiamdashInsights from population genomics Evol Appl 2020 13 2449ndash2459 [CrossRef]
6 Hu C Guo J Fu X Huang Y Gao X Wu K Seasonal migration pattern of Nilaparvata lugens (Hemiptera Delphacidae) overthe Bohai Sea in northern China J Econ Entomol 2018 111 2129ndash2135 [CrossRef]
7 Ma J Wang Y-C Hu Y-Y Lu M-H Wan G-J Chen F-J Liu W-C Zhai B-P Hu G Brown planthopper Nilaparvatalugens was concentrated at the rear of the typhoon Soudelor in eastern China in August 2015 Insect Sci 2018 25 916ndash926[CrossRef]
8 Fujita D Kohli A Horgan FG Rice resistance to planthoppers and leafhoppers Crit Rev Plant Sci 2013 32 162ndash191[CrossRef]
9 Fujii T Yoshida K Kobayashi T Myint KKM Yasui H Sanada-Morimura S Matsumura M Long-term virulencemonitoring of differential cultivars in Japanrsquos immigrant populations of Nilaparvata lugens (Hemiptera Delphacidae) in 2001ndash2009Appl Entomol Zool 2021 56 407ndash418 [CrossRef]
10 Myint KKM Yasui H Takagi M Matsumura M Virulence of long-term laboratory populations of the brown planthopperNilaparvata lugens (Stal) and whitebacked planthopper Sogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) on rice differentialvarieties Appl Entomol Zool 2009 44 149ndash153 [CrossRef]
11 Horgan FG Ramal AF Bentur JS Kumar R Bhanu KV Sarao PS Iswanto EH Van Chien H Phyu MHBernal CC et al Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia againstresistant rice varieties Crop Prot 2015 78 222ndash231 [CrossRef]
12 Horgan FG Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers Crop Prot2018 110 21ndash33 [CrossRef]
13 Uddin ABMA Islam KS Jahan M Ara A Afrin S Farmerrsquos perception about resurgence of brown planthopper NilaparvataLugens (Staringl) in Bangladesh Bangladesh Rice J 2020 23 21ndash33 [CrossRef]
14 Triwidodo H Brown planthoppers infestations and insecticides use pattern in Java Indonesia AGRIVITA J Agric Sci 202042 320ndash330 [CrossRef]
15 Veres A Wyckhuys KA Kiss J Toacuteth F Burgio G Pons X Avilla C Vidal S Razinger J Bazok R et al An update of theWorldwide Integrated Assessment (WIA) on systemic pesticides Part 4 Alternatives in major cropping systems Environ SciPollut Res 2020 27 29867ndash29899 [CrossRef]
16 Bakker L van der Werf W Tittonell P Wyckhuys KA Bianchi FJ Neonicotinoids in global agriculture Evidence for a newpesticide treadmill Ecol Soc 2020 25 26 [CrossRef]
17 Wu J-C Xu J-X Yuan S-Z Liu J-L Jiang Y-H Xu J-F Pesticide-induced susceptibility of rice to brown planthopperNilaparvata lugens Entomol Exp Appl 2001 100 119ndash126 [CrossRef]
18 Wu J-C Qiu H-M Yang G-Q Liu J-L Liu G-J Wilkens RM Effective duration of pesticide-induced susceptibility of riceto brown planthopper (Nilaparvata lugens Staringl Homoptera Delphacidae) and physiological and biochemical changes in riceplants following pesticide application Int J Pest Manag 2004 50 55ndash62 [CrossRef]
19 Wang A-H Wu J-C Yu Y-S Liu J-L Yue J-F Wang MY Selective insecticide-induced stimulation on fecundity andbiochemical changes in Tryporyza incertulas (Lepidoptera Pyralidae) J Econ Entomol 2005 98 1144ndash1149 [CrossRef] [PubMed]
Crops 2021 1 182
20 Azzam S Wang F Wu J-C Shen J Wang L-P Yang G-Q Guo Y-R Comparisons of stimulatory effects of a seriesof concentrations of four insecticides on reproduction in the rice brown planthopper Nilaparvata lugens Staringl (HomopteraDelphacidae) Int J Pest Manag 2009 55 347ndash358 [CrossRef]
21 Ge L-Q Hu J-H Wu J-C Yang G-Q Gu H Insecticide-induced changes in protein RNA and DNA contents in ovary andfat body of female Nilaparvata lugens (Hemiptera Delphacidae) J Econ Entomol 2009 102 1506ndash1514 [CrossRef]
22 Ling S Zhang J Hu L Zhang R Effect of fipronil on the reproduction feeding and relative fitness of brown planthopperNilaparvata lugens Appl Entomol Zool 2009 44 543ndash548 [CrossRef]
23 Ge L-Q Wang L-P Zhao K-F Wu J-C Huang L-J Mating pair combinations of insecticide-treated male and femaleNilaparvata lugens Staringl (Hemiptera Delphacidae) planthoppers influence protein content in the male accessory glands (MAGs)and vitellin content in both fat bodies and ovaries of adult females Pestic Biochem Physiol 2010 98 279ndash288 [CrossRef]
24 Ge L-Q Wu J-C Zhao K-F Chen Y Yang G-Q Induction of Nlvg and suppression of Nljhe gene expression in Nilaparvatalugens (Staringl) (Hemiptera Delphacidae) adult females and males exposed to two insecticides Pestic Biochem Physiol 201098 269ndash278 [CrossRef]
25 Hu J-H Wu J-C Yin J-L Gu H-N Physiology of insecticide-induced stimulation of reproduction in the rice brownplanthopper (Nilaparvata lugens (Staringl)) Dynamics of protein in fat body and ovary Int J Pest Manag 2010 56 23ndash30 [CrossRef]
26 Wang L-P Shen J Ge L-Q Wu J-C Yang G-Q Jahn GC Insecticide-induced increase in the protein content of maleaccessory glands and its effect on the fecundity of females in the brown planthopper Nilaparvata lugens Staringl (HemipteraDelphacidae) Crop Prot 2010 29 1280ndash1285 [CrossRef]
27 Wu J Ge L Liu F Song Q Stanley D Pesticide-induced planthopper population resurgence in rice cropping systems AnnuRev Entomol 2020 65 409ndash429 [CrossRef]
28 Suri KS Singh G Insecticide-induced resurgence of the whitebacked planthopper Sogatella furcifera (Horvath) (HemipteraDelphacidae) on rice varieties with different levels of resistance Crop Prot 2011 30 118ndash124 [CrossRef]
29 Fabellar LT Heinrichs EA Toxicity of insecticides to predators of rice brown planthoppers Nilaparvata lugens (Staringl) (HomopteraDelphacidae) Environ Entomol 1984 13 832ndash837 [CrossRef]
30 Fabellar LT Heinrichs EA Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators Crop Prot1986 5 254ndash258 [CrossRef]
31 Cuong NL Ben PT Phuong LT Chau LM Cohen MB Effect of host plant resistance and insecticide on brown planthopperNilaparvata lugens (Staringl) and predator population development in the Mekong Delta Vietnam Crop Prot 1997 16 707ndash715[CrossRef]
32 Preetha G Stanley J Suresh S Samiyappan R Risk assessment of insecticides used in rice on mirid bug Cyrtorhinuslividipennis Reuter the important predator of brown planthopper Nilaparvata lugens (Stal) Chemosphere 2010 80 498ndash503[CrossRef] [PubMed]
33 Tanaka K Endo S Kazano H Toxicity of insecticides to predators of rice planthoppers Spiders the mirid bug and the dryinidwasp Appl Entomol Zool 2000 35 177ndash187 [CrossRef]
34 Zhang X Xu Q Lu W Liu F Sublethal effects of four synthetic insecticides on the generalist predator Cyrtorhinus lividipennisJ Pest Sci 2015 88 383ndash392 [CrossRef]
35 Settle WH Ariawan H Astuti ET Cahyana W Hakim AL Hindayana D Lestari AS Managing tropical rice peststhrough conservation of generalist natural enemies and alternative prey Ecology 1996 77 1975ndash1988 [CrossRef]
36 Salim M Heinrichs EA Insecticide-induced changes in the levels of resistance of rice cultivars to the whitebacked planthopperSogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) Crop Prot 1987 6 28ndash32 [CrossRef]
37 Yin J-L Xu H-W Wu J-C Hu J-H Yang G-Q Cultivar and insecticide applications affect the physiological development ofthe brown planthopper Nilaparvata lugens (Staringl) (Hemiptera Delphacidae) Environ Entomol 2014 37 206ndash212 [CrossRef]
38 Cheng C Chang W Studies on varietal resistance to the brown planthopper in Taiwan In Brown Planthopper Threat toRice Production in Asia International Rice Research Institute Ed International Rice Research Institute Manila Philippines1979 pp 251ndash271
39 Reissig WH Heinrichs EA Valencia SL Insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on ricevarieties with different levels of resistance Environ Entomol 1982 11 165ndash168 [CrossRef]
40 Heinrichs EA Fabellar LT Basilio RP Wen T-C Medrano F Susceptibility of rice planthoppers Nilaparvata lugens andSogatella furcifera (Homoptera Delphacidae) to insecticides as influenced by level of resistance in the host plant Environ Entomol1984 13 455ndash458 [CrossRef]
41 Joshi R Shepard B Kenmore P Lydia R Insecticide-induced resurgence of brown planthopper (BPH) on IR62 Int Rice ResNewsl 1992 17 9ndash10
42 Zhu Z-R Cheng J Jiang M-X Zhang X-X Complex influence of rice variety fertilization timing and insecticide onpopulation dynamics of Sogatella furcifera (Horvath) Nilaparvata lugens (Staringl) (Homoptera Delphacidae) and their natural enemiesin rice in Hangzhou China J Pest Sci 2004 77 65ndash74 [CrossRef]
43 Nanthakumar M Jhansi Lakshmi V Shashi Bhushan V Balachandran SM Mohan M Decrease of rice plant resistance andinduction of hormesis and carboxylesterase titre in brown planthopper Nilaparvata lugens (Staringl) by xenobiotics Pestic BiochemPhysiol 2012 102 146ndash152 [CrossRef]
Crops 2021 1 183
44 Yao H-W Jiang C-Y Ye G-Y Cheng J-A Studies on cultivar-induced changes in insecticide susceptibility and its relatedenzyme activities of the white-backed planthopper Sogatella furcifera (Horvath)(Homopter Delphacidae) Insect Sci 2002 9 9ndash15
45 Horgan FG Srinivasan TS Bentur JS Kumar R Bhanu KV Sarao PS Van Chien H Almazan MLP Bernal CCRamal AF et al Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers Implicationsfor rice breeding and gene deployment Agronomy 2017 7 62 [CrossRef]
46 Jairin J Phengrat K Teangdeerith S Vanavichit A Toojinda T Mapping of a broad-spectrum brown planthopper resistancegene Bph3 on rice chromosome 6 Mol Breed 2007 19 35ndash44 [CrossRef]
47 Ren J Gao F Wu X Lu X Zeng L Lv J Su X Luo H Ren G Bph32 a novel gene encoding an unknown SCRdomain-containing protein confers resistance against the brown planthopper in rice Sci Rep 2016 6 37645 [CrossRef]
48 Nguyen CD Zheng S-H Sanada-Morimura S Matsumura M Yasui H Fujita D Substitution mapping and characterizationof brown planthopper resistance genes from indica rice variety lsquoPTB33prime (Oryza sativa L) Breed Sci 2021 in press [CrossRef]
49 Chelliah S Heinrichs EA Factors affecting insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on RiceEnviron Entomol 1980 9 773ndash777 [CrossRef]
50 Moore MT Kroumlger R Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growthArch Environ Contam Toxicol 2010 59 574ndash581 [CrossRef]
51 Kim J Hwang I Han M Jang B Effect of insecticide and fungicide on phytotoxicity of herbicide in rice Korean J Weed Sci1986 6 67ndash75
52 Horgan FG Garcia CPF Haverkort F de Jong PW Ferrater JB Changes in insecticide resistance and host range performanceof planthoppers artificially selected to feed on resistant rice Crop Prot 2020 127 104963 [CrossRef] [PubMed]
53 Khush GS Virk PS IR Varieties and Their Impact International Rice Research Institute Manila Philippines 200554 Srinivasan TS Almazan MLP Bernal CC Ramal AF Subbarayalu MK Horgan FG Interactions between nymphs of
Nilaparvata lugens and Sogatella furcifera (Hemiptera Delphacidae) on resistant and susceptible rice varieties Appl Entomol Zool2016 51 81ndash90 [CrossRef]
55 Pentildealver Cruz A Arida A Heong KL Horgan FG Aspects of brown planthopper adaptation to resistant rice varieties withthe Bph3 gene Entomol Exp Appl 2011 141 245ndash257 [CrossRef]
56 Alam SN Cohen MB Detection and analysis of QTLs for resistance to the brown planthopper Nilaparvata lugens in adoubled-haploid rice population Theor Appl Genet 1998 97 1370ndash1379 [CrossRef]
57 Mackill DJ Khush GS IR64 A high-quality and high-yielding mega variety Rice 2018 11 18 [CrossRef] [PubMed]58 Heinrichs EA Basilio RP Valencia SL Buprofezin a selective insecticide for the management of rice planthoppers (Ho-
moptera Delphacidae) and leafhoppers (Homoptera Cicadellidae) Environ Entomol 1984 13 515ndash521 [CrossRef]59 Satapathy MK Anjaneyulu A Use of cypermethrin a synthetic pyrethroid in the control of rice tungro virus disease and its
vector Trop Pest Manag 1984 30 170ndash178 [CrossRef]60 Vorley W Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown
planthopper (BPH) in Kedah Malaysia Int Rice Res Newsl 1987 10 19ndash2061 Uddin A Islam KS Jahan M Ara A Khan MAI Factor influencing the resurgence of brown planthopper in Bangladesh
SAARC J Agric 2020 18 117ndash128 [CrossRef]62 Escalada MM Luecha M Heong KL Social impacts of planthopper outbreaks Case study from Thailand In Rice Planthoppers
Ecology Management Socio Economics and Policy Heong KL Cheng J Escalada MM Eds Zhejiang University PressHangzhou China 2015 pp 209ndash226
63 Zhang CP He HM Yu JZ Hu XQ Zhu YH Wang Q Residues of carbosulfan and its metabolites carbofuran and3-hydroxy carbofuran in rice field ecosystem in China J Environ Sci Health 2016 51 351ndash357 [CrossRef] [PubMed]
64 Samanta S Barman M Samanta NRA Bio-efficacy trials of carbofuran 3 CG against insect pests of rice J Entomol ZoolStud 2020 8 1529ndash1533
65 Donley N The USA lags behind other agricultural nations in banning harmful pesticides Environ Health 2019 18 44 [CrossRef][PubMed]
66 Adams A Gore J Musser F Cook D Catchot A Walker T Dobbins C Efficacy of selected insecticides applied to hybridrice seed J Econ Entomol 2015 109 200ndash206 [CrossRef]
67 Kumar R Singh B Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) Field Bull Environ Contam Toxicol2013 90 482ndash488 [CrossRef]
68 Dhaka S Prajapati C Singh D Singh R Field evaluation of insecticides and bio-pesticides against rice leaf folder CnaphalocrocisMedinalis (Guenee) Ann Plant Prot Sci 2011 19 324ndash326
69 Matsumura M Takeuchi H Satoh M Sanada-Morimura S Otuka A Watanabe T Van Thanh D Species-specific insecticideresistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in East and South-eastAsia Pest Manag Sci 2008 64 1115ndash1121 [CrossRef] [PubMed]
70 Tang J Li J Shao Y Yang B Liu Z Fipronil resistance in the whitebacked planthopper (Sogatella furcifera) Possible resistancemechanisms and cross-resistance Pest Manag Sci 2010 66 121ndash125 [CrossRef]
71 Horgan FG Arida A Ardestani G Almazan MLP Temperature-dependent oviposition and nymph performance revealdistinct thermal niches of coexisting planthoppers with similar thresholds for development PLoS ONE 2020 15 e0235506[CrossRef] [PubMed]
Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]

Crops 2021 1 167
[HPR]) to the planthopper based on conventional crosses between naturally resistanttraditional landraces and high-yielding rice varieties [8] Resistant varieties have beendeployed to farmersrsquo fields since the 1970s however BPH populations can often rapidlyadapt to this resistancemdasha phenomenon known as lsquovirulence adaptationrsquo [9ndash11] Currentlythere are several resistance genes available from rice landraces and wild rice species suchthat virulence adaptation can now be better managed [12] The second most prominentBPH management method is the use of chemical insecticides [21314] The development ofinsecticides is mainly conducted through private companies Beginning in the early 2000slarge increases in the trade and use of insecticides have been reported throughout Asia [2]Furthermore trends suggest that farmers increasingly apply insecticides prophylacticallyand these applications are not in response to observed high pest densities as recommendedby Integrated Pest Management (IPM) principals [21516] Several detailed studies haveshown that some commonly-used insecticides actually enhance the growth rates of BPHpopulations and in many cases will ultimately lead to localized outbreaks of BPH infarmersrsquo fields [17ndash27] The deployment of resistant rice and the prophylactic applicationof insecticides might therefore be antagonistic to achieving successful BPH managementFurthermore where farmers conduct prophylactic insecticide applications HPR cannotachieve one of its principal objectives that of reducing pesticide inputs and maintaininga healthy rice environment [12] Indeed in such cases resistance can become simply anlsquoinsurancersquo against insecticide-induced BPH outbreaks and may inadvertently contributeto an overuse of insecticides
The mechanisms by which insecticides lead to BPH resurgence have received consid-erable research attention Recently Wu et al (2020) [27] reviewed much of the availableinformation Resurgence has been noted across a wide range of insecticide products thathave two main effects in the rice ecosystem Firstly certain insecticides enhance the feedingrates growth rates development reproductive output and migration potential of BPHeither through direct effects on planthopper physiology known as hormesis or indirectlyby inducing susceptibility in the host plant (ie by lowering phenol concentrations andenhancing sugar mobilization) [1828] Secondly many insecticides will have a greatereffect in reducing natural enemy populations than in reducing BPH populations therebydepleting the pest regulation capacity of the rice ecosystem [29ndash34] The higher susceptibil-ity of natural enemies to insecticides compared to BPH and other pests has been attributedto a greater mobility of natural enemies and thereby higher probabilities of exposureto the insecticides furthermore many predatory and parasitic arthropods have slowerpopulation growth rates than BPH [35] which reduces the potential for natural enemies todevelop insecticide resistance
Horgan (2018) [12] reviewed studies where insecticides were applied to both resistantand susceptible rice varieties to compare effects In many cases insecticides reduced theeffectiveness of resistance however relative resistance was often maintained across testvarietiesmdashalbeit with some minor exceptions [283637] However only ten publishedstudies have examined the combined effects of host resistance and insecticides on BPHpopulation development [18203137ndash43] a further three studies examined effects on thewhite-backed planthopper Sogatella furcifera Horvaacuteth [WBPH] an increasingly commonpest of rice in Asia [283644] Only seven of these studies have been published since 2000and to our knowledge there have been no published studies since 2018 that compare theactions of insecticides on BPH-susceptible and resistant rice (based on records retrievedusing Google Scholar) As a consequence information on interactions between currently-effective BPH resistance genes and commonly used insecticides is largely unavailableBased on the materials (rice varieties and insecticides) used in previous studies there isclearly a need to further examine the possible synergistic andor antagonistic effects ofcombining host resistance and insecticides for BPH management
In the present study we examine the effects of seven widely available insecticideson the integrity of resistance in rice against BPH We selected a rice variety IR62 withrelatively strong resistance against BPH populations from across Asia [1145] This variety
Crops 2021 1 168
possesses the Bph3 and Bph32 resistance genes derived from the Indian landrace PTB33Because of its relatively high effectiveness IR62 and doner varieties with the Bph3 locuscontaining either the Bph3 or Bph32 resistance genes have been extensively studied inrecent years [46ndash48] We also selected insecticides that are currently used to manage insectherbivores in rice in Asia Many of the products are not specifically recommended for useagainst BPH however resurgence is often the result of applications against other insectpests (ie BPH is a secondary pest) and particularly against rice leaffolders [49] Weexamined the indirect plant-mediated effects of insecticides on BPH because our focuswas on possible insecticide-plant interactions as well as on direct insecticide effects onBPH Therefore we conducted our experiments in greenhouses and in a screenhousethat were largely free of natural enemies Furthermore because of possible phytotoxiceffects of insecticides on rice plants [5051] and the potential role of such phytotoxicityin determining BPH-rice interactions [49] we also assessed insecticide effects on plantgrowth and yields in the screenhouse We discuss our results in terms of applying HPRand insecticide treatments according to IPM principles
2 Materials and Methods21 Herbivore Species
BPH were obtained from colonies reared at the International Rice Research Institute(IRRI) in the Philippines Planthoppers were initially collected in 2009 from rice fields inLaguna Province The founder population (ca 500 individuals) was placed in wire meshcages of 120 times 60 times 60 cm (H timesW times L) under greenhouse conditions (temperatures [T]25ndash37 C 70ndash90 relative humidity [RH] 12h12h daynight [DN]) and was continuouslyreared on ge30 days after sowing [DAS] rice plants (variety = TN1) Feeding plants werereplaced every two weeks The colony received periodic (each year) introgressions ofwild-caught individuals from the same collection sites BPH from the Laguna Provincehave been screened for virulence and are largely adapted to feed on rice varieties with theBph1 bph2 bph5 bph7 Bph18 Bph25 and Bph26 genes [1145] The colonies were also testedfor levels of resistance to insecticides and have moderate resistance to imidacloprid andhigh resistance to BPMC [52]
22 Plant Materials
We used two varieties in our experiments IR62 was selected as a variety with highresistance to BPH populations throughout South and Southeast Asia [53] Resistance inIR62 was derived from PTB33 and is attributed to the Bph3Bph32 resistance genes [4853]BPH has reduced feeding lower weight gain slower development and reduced egg-layingon IR62 compared to susceptible varieties [4] The variety also has moderate resistanceto WBPH and to the green leafhopper Nephotettix virescens (Distant) [GLH] [4554] Fur-thermore resistance derived from PTB33 and associated with the Bph3 locus appears tobe highly durable since BPH populations with virulence against PTB33-derived varietiesincluding IR62 are still rare despite over 30 years of deployment in regions of Mindanao(Philippines) and in Cambodia [55] The variety IR64 was selected as a susceptible vari-ety IR64 has the Bph1 gene for resistance but BPH populations throughout Asia haveadapted to this gene [1153] Nevertheless IR64 has additional quantitative resistance thatcontributes to tolerance (ie a relatively high capacity to withstand damage) of BPH inthe variety [56] IR64 was released by IRRI in 1985 and is considered a mega-variety inAsia [57] Because of its popularity the variety has been enhanced using marker-assistedbreeding to incorporate traits such as flood tolerance (ie IR64-Sub1 released in 2009) [57]For our experiments seeds of both varieties were acquired from the IRRI-germplasm bank
23 Insecticides
We used seven insecticides in our experiments The insecticides were selected basedon their availability at IRRI Buprofezin (2-tert-butylimini-3-isopropyl-5-phenyl-3456-tetrahydro-2H-135-thiadiazin-5-one) (25SC 200 mL haminus1) contains the growth regulator
Crops 2021 1 169
thiadezine and is used against planthoppers and leafhoppers in rice It is the recommendedinsecticide to combat high densities of planthoppers in rice because it has relatively lowtoxicity to natural enemies [58] We used two synthetic pyrethroids cypermethrin (cyan-(3-pheboxyphenyl)-methyl-(3-CS2-dichloroethenyl)-22-dimethyl cyclopropane carboxy-late) (10EC 800 mL haminus1) and deltamethrin ((S)-cyano-3-phenoxybenzyl (1R3R)-3-(22-dibromovinyl)-22-dimethyl-cyclopropane-carboxylate) (25EC 500 mL haminus1) cypermethrinhas been commonly used in rice production to combat rice tungro virus that is transmittedby rice leafhoppers [59] Deltamethrin is a broad-spectrum insecticide that is also some-times used to manage leafhoppers Both pyrethroids were previously noted to cause BPHresurgence [396061] and are currently not recommended for use in rice [62] Neverthe-less cypermethrin and deltamethrin are still commonly used on rice in some countriesincluding Indonesia and Bangladesh [131461] For this reason we included both productsin our experiments We used the carbamate insecticide carbofuran (22-Dimethyl-23-dihydro-1-benzofuran-7-flmethyl-carbamate) (3G 3 kg haminus1) as a systemic insecticidethat is sometimes recommended for control of rice pests including planthoppers [6364]Carbofuran has been banned in the European Union the USA and Canada but is still usedin much of Asia [65] We used thiamethoxam 20 + chlorantraniliprole 20 (3-[(2-chloro-13-thiazol-5-yl)methyl]-5-methyl-135-oxadiazinan-4-ylidene nitramide + 5-bromo-N-[4-chloro-2-methyl-6-(methylcarbamoyl)phenyl]-2-(3-chloropyridin-2-yl)pyrazole-3-carboxamide) (40 WG75 g haminus1) ie a combined neonicotinoid and diamide product as a further systemic in-secticide The product is absorbed through the plant to give broad-spectrum control of ricepests [66] Finally we used fipronil (5-Amino-1-[26-dichloro-4-(trifluoromethyl)phenyl]-4-(trifluoromethanesulfinyl)-1H-pyrazole-3-carbonitrile) (50SC 500 mL haminus1) and car-tap hydrochloride (S-[3-carbamoylsulfanyl-2-(dimethylamino)propyl] carbamothioate-hydrochloride) (a nereistoxin analogue) (50SP 400 g haminus1) as two broad-spectrum in-secticides commonly used to control leaffolders in Asian rice [67ndash70] All insecticideswere applied at recommended rates with quantities estimated based on the surface soilarea of each experimental unit Insecticides were applied by trained pesticide applicatorsusing standard protective equipment in a well-ventilated environment (an insect screen-house) Any evaluations of treated rice plants were conducted at a minimum of 3 dayspost-application
24 Effects of HPR and Insecticides on BPH Preferences
We assessed the effects of the insecticides on oviposition by BPH in a multichoice andin binary choice bioassays For both tests seedlings of IR62 and IR64 were planted individ-ually in 0 pots (5times 5 cm Htimes Diameter) in a greenhouse (T = 25ndash37 C RH = 70ndash90 DN= 12 h12 h) At 27 days after sowing (DAS) the seedlings were treated with one of eachof the seven insecticides as described above or were sprayed with water (ie untreatedcontrol) We used water as a control because some of the products were diluted with waterPlants were returned to the greenhouse at 30 DAS For each variety in the multichoicebioassay eight plants at 30 DASmdashone for each treatmentmdashwere placed together in a wiremesh cage of 120 times 60 times 60 cm (H times W times L) The plants were arranged in a circularconfiguration each separated by 10 cm such that adjacent plants did not touch each otheror the cage walls The cages were replicated five times for each rice variety with plantspositioned randomly during each replication Twenty gravid female N lugens were placedat the center of each cage and allowed to lay eggs for 3 days after which all adults weremanually removed from the cages Eggs were allowed to develop in the plants for a further4 days after which the plants were collected wrapped in moistened tissue paper andstored in a refrigerator Within 5 days all plants were dissected under a stereo microscope(Nikon SMZ-2B Tokyo Japan) to count the egg clusters Egg clusters were removed fromthe plant tissues and the numbers of eggs per cluster were recorded The total number ofeggs was estimated as the sum of eggs per plant from all clusters
For the binary choice tests one plant (either IR62 or IR64) for each insecticide treatmentwas placed together with a control (untreated) plant of the same variety in an acetate cage
Crops 2021 1 170
of 30 times 25 times 50 cm (H timesW times L) (ie two plants per cage) This produced 14 treatmenttimes variety combinations (7 insecticides times 2 varieties) Each combination of plant pairswas replicated three times with cages positioned randomly on greenhouse benches andreplicates of each combination pair placed in separate greenhouse compartments Fivegravid females were placed between the two pots in each cage and allowed to move withinthe cage and lay eggs After 3 days all adults were manually removed from the cages Theplants were collected 4 days after removal of the adults to allow the eggs to develop Plantswere placed in moist tissue paper and stored in a refrigerator until clusters and eggs werecounted under a stereomicroscope (as described above) For both bioassays the replicateswere each placed in separate greenhouse compartments (ie 5 or 3 blocks)
25 Effects of HPR and Insecticides on BPH Fitness
We conducted two BPH antibiosis tests in the greenhouse We examined egg layingby gravid females on each variety and for each insecticide treatment using an ovipositiontest At 30 DAS treated and control plants were individually caged by placing an acetatetube 45 times 5 cm (H times Diameter) with a mesh top over each pot A recently mated gravidBPH was placed inside each cage through a slit in the acetate and was allowed to feed andoviposit After 3 days the females were manually removed from the cages Plants were leftfor 4 days to allow eggs to develop The plants were then collected and dissected under astereo microscope to count the clusters and eggs (as described above)
We examined the effects of host resistance and insecticide treatments on BPH nymphsin a survival test At 30 DAS treated and control plants were individually caged asdescribed above Recently emerged nymphs (le2 instar) were collected from the colonyusing a suction-pooter and placed inside the cages at a density of ten per cage through aslit in the acetate The nymphs were allowed to feed and develop for 15 days after whichthe number of survivors and their developmental stages were recorded For both bioassaysplants were dried in a forced draft oven at 60 C for 7 days and were weighed For thesurvival bioassay the nymphs were also dried and weighed to estimate dry biomass andthereby control for varying consumption of plant liquids Both bioassays were replicatedfive times (ie 8 treatments times 2 varieties times 5 replicates = 80 pots per bioassay) with eachreplicate placed in a separate greenhouse compartment (ie 5 blocks)
26 Effects of HPR and Insecticides in a Screenhouse Environment
The combined effects of insecticide treatments and HPR were investigated using riceplants grown in an insect-proof screenhouse The screenhouse consisted of six concretebays (dimensions 266times 25 m (LtimesW)) filled with flooded paddy soil Temperatures in thescreenhouse varied between 21 and 34 C and RH varied between 70 and 90 Rainwatercould pass through the screenhouse roof during the experiment 160 mm of precipitationwas recorded representing the Philippines dry season The screenhouse had no artificiallighting (ie DN = 12 h12 h)
In one of the bays translucent plastic barriers were erected to delimitate five exper-imental blocks of 150 times 300 m Each block was further divided into eight cubicles of075 m2 using the same plastic materials The barriers were supported by bamboo stakesand reached a height of 150 m and a depth into the paddy soil of approx 030 m After thebarriers were installed two 23 DAS seedlings each of IR62 and IR64 were randomly trans-planted to each cubicle at 25 times 25 cm spacing We used a standard fertilizer regime to sim-ulate farmersrsquo fields The seedlings were treated with ammonium sulphate at 100 kg haminus1
split equally into three applications (ie one basal and two top dressings) Solophos andmuriate of potash was also applied at 20 kg haminus1 as a basal application Plants were contin-uously watered and the cubicles were manually weeded until rice plants were harvestedCubicles in each of the five blocks received one of the eight treatments (seven insecticidesand a control) Treatments were randomly assigned to cubicles within blocks The insecti-cide applications were made at 30 and 60 DAS to simulate farmer practices under intensive
Crops 2021 1 171
rice production systems where farmers apply insecticides ge two times [14] however inour experiment we did not mix insecticides
Rice herbivores including BPH WBPH and GLH occurred at low densities in thescreenhouse Therefore at the opposite end of the screenhouse (separated by a distanceof 20 m) a bay with gt50 DAS TN1 rice plants was infested with high densities of BPH(approx 200 BPH plantminus1) This acted as a potential source of planthoppers to attackthe treated IR62 and IR64 plants in our experiment Natural enemies were largely absentfrom the screenhouse because of the insect-proof cladding (1 mm times 1 mm mesh bore)Microparasitoids (Oligosita spp and Anagrus spp) that could pass through the screenoccasionally attacked planthopper eggs in the screenhouse (estimated 5ndash10 of eggsparasitized unpublished data)
The IR62 and IR64 plants were continuously monitored to assess grain maturationWhen about 85 of the rice grain was mature any arthropods on the plants were collectedusing a vacuum sampler and stored in plastic vials The plants were then destructivelyharvested by carefully pulling the plant from the soil to maintain intact roots The plantroots were washed under running water and each plant was placed inside a paper bagThe plants and arthropods were then dried in a forced draft oven at 60 C for 7 days andweighed Arthropods were separated by species and counted The plants were separatedinto roots shoots and panicles and the numbers of reproductive and vegetative tillerswere recorded Rice grain was removed from the panicles and separated as filled andnon-filled grain The dried grain was counted and weighed
27 Data Analyses
The numbers of egg clusters and eggs laid in rice plants during the multichoice testwere analyzed using univariate general linear models (GLM) on ranked (within replicate)data Separate analyses were conducted for IR62 and IR64 plants Data from the binarychoice bioassays were analyzed using paired t-tests The numbers of egg clusters and eggslaid in treated and non-treated plants during the oviposition bioassay were compared usingunivariate GLM with plant biomass initially included as a covariate but later removedbecause it had no significant effect Cluster and egg numbers were ranked within blocksbefore analyses Average cluster size was estimated by dividing the number of eggs in eachplant by the number of clusters Cluster size was analyzed using univariate GLM removingthe block effect Similarly the proportions of surviving nymphs and nymph biomass fromthe survival bioassay were analyzed using univariate GLMs initially including plantbiomass as a covariate Data from the survival bioassay were ranked before analysesToo few nymphs survived under some treatments to examine development rates or thesex of surviving adults We estimated the combined effects of HPR and each insecticidelsquoxrsquo as -((fitness on untreated IR64 minus fitness on x-treated IR62)fitness on untreated IR64)To estimate the effect of the insecticide alone we removed the effect on HPR which weestimated as fitness on untreated IR62 minus fitness on untreated IR64 Fitness was assessedfrom the oviposition bioassay as the numbers of eggs laid and from the survival bioassayas nymph biomass Where nymph mortality on insecticide-treated IR64 plants approached100 HPR effects were regarded as insignificant
We also examined combined and insecticide effects from the screenhouse data basedon densities of BPH and WBPH (see below) Effects were calculated using insecticide andHPR combinations for each replicated block (N = 5) Estimated effects were comparedacross treatments using univariate GLMs Because three herbivores (BPH WBPH andGLH) occurred in our screenhouse plots we used a split-plot multivariate GLM to analyzeherbivore densities Total planthopper and leafhopper biomass and density and plantgrowth parameters (tiller number biomass yield grain number proportion of grainunfilled and grain size) were analyzed using split-plot univariate GLMs The covariateplant biomass was initially included in analyses of the effects on herbivores but had noeffect and was subsequently removed Similarly the covariate herbivore biomass wasinitially included in analyses of plant growth parameters but had no effect and was
Crops 2021 1 172
subsequently removed The combined effects of insecticide and HPR and the separateeffects of each insecticide (calculated as indicated above) on BPH and WBPH were alsoanalyzed using univariate GLM
Post-hoc Tukey tests were applied to all significant treatment factors Residuals wereplotted following all parametric analyses and were found to be normal and homogenous
3 Results31 Effects of HPR and Insecticides on BPH Preferences
Treatments affected the number of clusters and eggs that were laid in IR62 plants(ranked clusters and eggs F732 = 4738 p lt 0001) and in IR64 plants (clusters F732 = 6229p lt 0001 eggs F732 = 7757 p lt 0001) (Figure 1AB) Fewer eggs were laid on IR64 plantstreated with the pyrethroids (cypermethrin and deltamethrin) fipronil and thiamethoxam-chlorantraniliprole compared to plants treated with buprofezin carbofuran and cartaphydrochloride (Figure 1B) Cypermethrin-treated IR62 plants also had fewer eggs thanbuprofezin-treated IR62 plants (Figure 1A) Although BPH females tended to lay fewereggs in many of the insecticide-treated rice plants in the binary choice experiments therewere no statistically significant differences between the numbers of eggs laid in treated-IR62(t ranges from minus0746 to 3186 p gt 005) or treated-IR64 (t ranges from minus0108 to 2789p gt 005) compared to corresponding non-treated plants (Figure 1CD)
Crops 2021 1 FOR PEER REVIEW 7
herbivore densities Total planthopper and leafhopper biomass and density and plant growth parameters (tiller number biomass yield grain number proportion of grain un-filled and grain size) were analyzed using split-plot univariate GLMs The covariate plant biomass was initially included in analyses of the effects on herbivores but had no effect and was subsequently removed Similarly the covariate herbivore biomass was initially included in analyses of plant growth parameters but had no effect and was subsequently removed The combined effects of insecticide and HPR and the separate effects of each insecticide (calculated as indicated above) on BPH and WBPH were also analyzed using univariate GLM
Post-hoc Tukey tests were applied to all significant treatment factors Residuals were plotted following all parametric analyses and were found to be normal and homogenous
3 Results 31 Effects of HPR and Insecticides on BPH Preferences
Treatments affected the number of clusters and eggs that were laid in IR62 plants (ranked clusters and eggs F732 = 4738 p lt 0001) and in IR64 plants (clusters F732 = 6229 p lt 0001 eggs F732 = 7757 p lt 0001) (Figure 1AB) Fewer eggs were laid on IR64 plants treated with the pyrethroids (cypermethrin and deltamethrin) fipronil and thiameth-oxam-chlorantraniliprole compared to plants treated with buprofezin carbofuran and cartap hydrochloride (Figure 1B) Cypermethrin-treated IR62 plants also had fewer eggs than buprofezin-treated IR62 plants (Figure 1A) Although BPH females tended to lay fewer eggs in many of the insecticide-treated rice plants in the binary choice experiments there were no statistically significant differences between the numbers of eggs laid in treated-IR62 (t ranges from minus0746 to 3186 p gt 005) or treated-IR64 (t ranges from minus0108 to 2789 p gt 005) compared to corresponding non-treated plants (Figure 1CD)
Figure 1 Results from multichoice (AB) and binary choice (CD) bioassays conducted in the green-house Bioassays were conducted separately for IR62 (AC) and IR64 (BD) Plants were treated with one of seven insecticides (A to G) as identified in the legend Control plants were sprayed with water Error bars are presented (N = 5 for (A) and (B) N = 3 for (C) and (D)) Lowercase letters (andashc) indicate homogenous treatment groups (Tukey p lt 005)
A B C D E F G0
100
200
300
400
Treatment
A B C D E F G
Non treatedTreated
A B C D E F G Control
Num
ber o
f egg
s pe
r pla
nt
0
100
200
300
400
A B C D E F G Control
IR62 IR64
A
C
B
D
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliproleH Control
a abab
c
abc
abc bc bc
c
aaa
ab
ab
bb b
Figure 1 Results from multichoice (AB) and binary choice (CD) bioassays conducted in thegreenhouse Bioassays were conducted separately for IR62 (AC) and IR64 (BD) Plants were treatedwith one of seven insecticides (A to G) as identified in the legend Control plants were sprayed withwater Error bars are presented (N = 5 for (AB) N = 3 for (CD)) Lowercase letters (andashc) indicatehomogenous treatment groups (Tukey p lt 005)
32 Effects of HPR and Insecticides on BPH Fitness
More eggs were laid on IR64 plants than on IR62 plants but BPH produced similarnumbers of egg clusters on both plants The lower numbers of eggs in IR62 were due tosmaller clusters on the variety (Table 1) The numbers of eggs and clusters were affectedby insecticide treatment with fewer eggs and clusters on cypermethrin-treated fipronil-treated and thiamethoxam-chlorantraniliprole-treated plants than on plants under the
Crops 2021 1 173
remaining treatments (Table 1) The treatment-effects were not affected by host variety (ieinteractions were non-significant Table 1) Insecticide treatments had no effect on clustersize in the bioassay (Table 1) There was no significant effect of combined insecticide andHPR (F628 = 0449 p = 0840) on observed changes in egg laying during the bioassay butthe estimated insecticide effect was greater for thiamethoxam-chlorantraniliprole fiproniland cypermethrin than for buprofezin cartap hydrochloride and carbofuran (F628 = 3140p = 0018 Figure 2A)
Table 1 Results from greenhouse oviposition and survival tests with IR62 (resistant) and IR64 (susceptible) each treatedwith one of seven insecticides Numbers are means plusmn SEM (N = 5)
Treatments Egg Clusters(Number Plantminus1) 12
Eggs(Number Plantminus1) 12
Cluster Size(Number of Eggs) 1
Nymph Survival(Proportions) 12
Nymph Biomass (mgDry Weight Plantminus1) 12
IR62Buprofezin 1240 plusmn 140 b 5040 plusmn 904 b 393 plusmn 036 000 plusmn 000 a 000 plusmn 000 a
Carbofuran 1300 plusmn 089 b 5380 plusmn 421 b 417 plusmn 026 064 plusmn 064 d 154 plusmn 154 d
Cartaphydrochloride 1360 plusmn 075 b 5200 plusmn 486 b 382 plusmn 029 028 plusmn 028 b 151 plusmn 151 b
Cypermethrin 620 plusmn 169 a 2000 plusmn 608 a 316 plusmn 023 002 plusmn 002 a 002 plusmn 002 a
Deltamethrin 1360 plusmn 160 b 4340 plusmn 566 b 338 plusmn 069 040 plusmn 040 c 120 plusmn 120 c
Fipronil 520 plusmn 146 a 2240 plusmn 735 a 393 plusmn 064 000 plusmn 000 a 000 plusmn 000 a
Thiamethoxam-chlorantraniliprole 380 plusmn 086 a 1700 plusmn 435 a 447 plusmn 032 000 plusmn 000 a 000 plusmn 000 a
Control 1260 plusmn 103 b 4900 plusmn 716 b 388 plusmn 054 080 plusmn 080 d 292 plusmn 292 d
IR64Buprofezin 2100 plusmn 164 10940 plusmn 1346 556 plusmn 117 000 plusmn 000 000 plusmn 000Carbofuran 1820 plusmn 404 9880 plusmn 2939 524 plusmn 074 086 plusmn 086 420 plusmn 420
Cartaphydrochloride 2140 plusmn 619 11460 plusmn 3141 549 plusmn 064 010 plusmn 010 045 plusmn 045
Cypermethrin 380 plusmn 233 1980 plusmn 1155 550 plusmn 058 000 plusmn 000 000 plusmn 000Deltamethrin 1560 plusmn 209 7500 plusmn 1581 473 plusmn 069 040 plusmn 040 201 plusmn 201
Fipronil 380 plusmn 086 2100 plusmn 552 546 plusmn 080 000 plusmn 000 000 plusmn 000Thiamethoxam-chlorantraniliprole 260 plusmn 075 800 plusmn 212 373 plusmn 133 000 plusmn 000 000 plusmn 000
Control 1740 plusmn 364 7520 plusmn 2344 426 plusmn 073 086 plusmn 086 360 plusmn 360F-treatment 3 19898 19898 0541 ns 133811 108451
F-variety 3 2596 ns 7607 7471 1080 ns 0008 ns
F-interaction 3 1840 ns 2034 ns 0610 ns 4902 9496 1 Lowercase letters (andashd) indicate homogenous treatment groups (for IR62 and IR64 combined) 2 Data were ranked within blocks beforeanalysis 3 DF = 164 for varieties 764 for treatments and interactions ns = p gt 005 = p le 001 = p le 0001
Nymph survival and biomass were not affected by rice variety (Table 1) Buprofezinfipronil thiamethoxam-chlorantraniliprole and cypermethrin reduced survival to 0 in thebioassay and all insecticides except carbofuran resulted in lower survival and biomass thanon untreated plants (Table 1) There were significant interactions because of higher nymphmortality and a resulting lower biomass on carbofuran-treated IR62 compared to untreatedIR62 but lower mortality and a higher biomass of nymphs on carbofuran-treated IR64than on untreated IR64 (Table 1) There was no significant effect of insecticide treatment(F628 = 0946 p = 0479) on observed changes in nymph biomass during the bioassaybut the estimated combined effects of pesticide and HPR was greater for thiamethoxam-chlorantraniliprole fipronil cypermethrin and buprofezin than for cartap hydrochlorideand carbofuran (F628 = 5699 p = 0001) (Figure 2B)
Crops 2021 1 174
Crops 2021 1 FOR PEER REVIEW 9
Figure 2 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) egg laying and (B) nymph biomass in greenhouse bioas-says Effects are indicated as proportional changes in fitness parameters compared to untreated IR64 Negative numbers indicate reduced fitness Standard errors are indicated (N = 5) Lowercase letters (ab) indicated homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 1)
Nymph survival and biomass were not affected by rice variety (Table 1) Buprofezin fipronil thiamethoxam-chlorantraniliprole and cypermethrin reduced survival to 0 in the bioassay and all insecticides except carbofuran resulted in lower survival and biomass than on untreated plants (Table 1) There were significant interactions because of higher nymph mortality and a resulting lower biomass on carbofuran-treated IR62 compared to untreated IR62 but lower mortality and a higher biomass of nymphs on carbofuran-treated IR64 than on untreated IR64 (Table 1) There was no significant effect of insecticide treatment (F628 = 0946 p = 0479) on observed changes in nymph biomass during the bio-assay but the estimated combined effects of pesticide and HPR was greater for thiameth-oxam-chlorantraniliprole fipronil cypermethrin and buprofezin than for cartap hydro-chloride and carbofuran (F628 = 5699 p = 0001) (Figure 2B)
33 Effects of HPR and Insecticides on Planthoppers and Leafhoppers Plants in the screenhouse were colonized by BPH WBPH and GLH Densities for
each species were similar on IR62 and IR64 (Wilkrsquos λ = 0994 p = 0952) There was a sig-nificant treatment effect on all three species (Wilkrsquos λ = 0586 p = 0049) because of high densities on deltamethrin-treated plants (Table 2) The numbers and biomass of all three species combined were also greater on deltamethrin-treated plants than on the remaining treatments (Table 2) Host resistance reduced GLH densities under the deltamethrin treat-ment but had no apparent effect on densities under the other treatments (Table 2) There were significant combined effects of HPR and insecticide on the numbers of BPH (F624 = 2686 p = 0039) and WBPH (F624 = 2482 p = 0052) this was almost exclusively due to the resurgence effect of deltamethrin on the planthoppers (BPH F624 = 2686 p = 0039 WBPH F624 = 2482 p = 0052) (Figure 3)
Δ eg
gs(P
ropo
rtion
)
-10
-08
-06
-04
-02
00
02
04
06
Treatment
A B C D E F G
Δ N
ymph
bio
mas
s(P
ropo
rtion
)
-10
-08
-06
-04
-02
00
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
ab
a a
b
a
b b
a a a a a a a a
b b bb
bb
Figure 2 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone(P grey bars) on (A) egg laying and (B) nymph biomass in greenhouse bioassays Effects are indicated as proportionalchanges in fitness parameters compared to untreated IR64 Negative numbers indicate reduced fitness Standard errors areindicated (N = 5) Lowercase letters (ab) indicated homogenous treatment groups based on Tukey pairwise comparisons(p lt 005) (see also Table 1)
33 Effects of HPR and Insecticides on Planthoppers and Leafhoppers
Plants in the screenhouse were colonized by BPH WBPH and GLH Densities foreach species were similar on IR62 and IR64 (Wilkrsquos λ = 0994 p = 0952) There was asignificant treatment effect on all three species (Wilkrsquos λ = 0586 p = 0049) because of highdensities on deltamethrin-treated plants (Table 2) The numbers and biomass of all threespecies combined were also greater on deltamethrin-treated plants than on the remainingtreatments (Table 2) Host resistance reduced GLH densities under the deltamethrintreatment but had no apparent effect on densities under the other treatments (Table 2)There were significant combined effects of HPR and insecticide on the numbers of BPH(F624 = 2686 p = 0039) and WBPH (F624 = 2482 p = 0052) this was almost exclusively dueto the resurgence effect of deltamethrin on the planthoppers (BPH F624 = 2686 p = 0039WBPH F624 = 2482 p = 0052) (Figure 3)
Crops 2021 1 175
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants underseven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density(Number Plantminus1) 1
WBPH Density(Number Plantminus1) 1
GLH Density(Number Plantminus1) 1
Density ofPlanthoppers and
Leafhoppers(Number Plantminus1) 1
Biomass ofPlanthoppers and
Leafhoppers(mg Dry Weight) 1
IR62Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a
Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartaphydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a
Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a
Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a
Thiamethoxam-chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a
IR64Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartaphydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123
Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns
F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Crops 2021 1 FOR PEER REVIEW 10
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants under seven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density (Num-
ber Plantminus1) 1 WBPH Density
(Number Plantminus1) 1 GLH Density
(Number Plantminus1) 1
Density of Planthoppers and Leafhoppers (Number
Plantminus1) 1
Biomass of Planthoppers and Leafhoppers (mg Dry
Weight) 1 IR62
Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartap hydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a Thiamethoxam-
chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a IR64
Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132 Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartap hydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123 Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113 Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052 Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087 F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns 1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatments and interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in the screenhouse Effects are indicated as proportional changes in planthopper densities compared to untreated IR64 Negative numbers indicate reduced densities positive numbers are increased den-sities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
Δ BP
H(N
umbe
r pla
nt-1
)
0
5
10
15
20
Treatment
A B C D E F G
Δ W
BPH
(Num
ber p
lant
-1)
0
20
30
40
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
a a
aa a a
a a a
a
a a
b
b
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimatedpesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in thescreenhouse Effects are indicated as proportional changes in planthopper densities compared to
Crops 2021 1 176
untreated IR64 Negative numbers indicate reduced densities positive numbers are increaseddensities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenoustreatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
34 Direct and Indirect Effects of Insecticides on Rice Development
IR62 plants had more tillers and attained a greater biomass than IR64 plants in thescreenhouse (Table 3) IR62 grain matured faster and plants were harvested earlier than theIR64 plants (Table 3) IR62 plants also produced more grain (number of grain) and hadhigher yields than IR64 but had smaller grain and more unfilled grain than IR64 (Table 3)
Table 3 Rice growth parameters for IR62 (resistant) and IR64 (susceptible) under seven insecticide treatments and forcontrol non-treated plants in a screenhouse environment Numbers are means plusmn SEM
Treatment Tiller Number 1 Plant Biomass(g Dry Weight) 1
Time to Harvest(Days) 1
Weightof Filled Grain(g Dry Weight) 1
Number ofFilled Grain
(1000s) 1
Unfilled Grain(Proportion) 1
1000 GrainWeight 1
IR62Buprofezin 2660 plusmn 206 ab 4643 plusmn 385 ab 7990 plusmn 168 ab 5776 plusmn 683 ab 361plusmn 061 ab 011 plusmn 003 a 2008 plusmn 042 ab
Carbofuran 2900 plusmn 106 ab 5546 plusmn 357 b 7720 plusmn 080 ab 7448 plusmn 691 b 446plusmn 061 ab 011 plusmn 001 a 2103 plusmn 054 b
Cartaphydrochloride 2820 plusmn 327 ab 4896 plusmn 470 ab 7920 plusmn 153 b 6430 plusmn 712 ab 415plusmn 064 ab 012 plusmn 001 b 2023 plusmn 065 a
Cypermethrin 2870 plusmn 301 a 4957 plusmn 562 a 7760 plusmn 103 b 6032 plusmn 822 a 410plusmn 067 ab 014 plusmn 002 a 1965 plusmn 076 a
Deltamethrin 2620 plusmn 345 a 4592 plusmn 588 a 7840 plusmn 108 ab 5698 plusmn 832 ab 379plusmn 047 ab 012 plusmn 001 a 2030 plusmn 063 ab
Fipronil 2380 plusmn 176 ab 4607 plusmn 657 ab 7700 plusmn 179 ab 5266 plusmn 390 a 318 plusmn 032 a 013 plusmn 002 a 2128 plusmn 047 b
Thiamethoxam-chlorantraniliprole 2650 plusmn 250 b 4547 plusmn 429 b 7720 plusmn 102 a 5777 plusmn 688 ab 408 plusmn 061 b 013 plusmn 002 a 2019 plusmn 067 ab
Control 3120 plusmn 247 b 5401 plusmn 555 b 7720 plusmn 080 ab 7045 plusmn 520 b 396plusmn 036 ab 012 plusmn 002 a 2076 plusmn 050 ab
IR64Buprofezin 2340 plusmn 250 5064 plusmn 747 8350 plusmn 124 5080 plusmn 952 270 plusmn 054 008 plusmn 001 2277 plusmn 039Carbofuran 2040 plusmn 097 4591 plusmn 141 8540 plusmn 073 4951 plusmn 312 263 plusmn 023 006 plusmn 000 2295 plusmn 036
Cartaphydrochloride 2120 plusmn 075 4336 plusmn 285 8520 plusmn 140 4273 plusmn 579 238 plusmn 026 015 plusmn 007 2050 plusmn 234
Cypermethrin 1990 plusmn 089 3968 plusmn 272 8710 plusmn 135 3920 plusmn 552 250 plusmn 028 008 plusmn 002 2171 plusmn 070Deltamethrin 1980 plusmn 181 4325 plusmn 357 8460 plusmn 068 4710 plusmn 496 277 plusmn 024 008 plusmn 001 2314 plusmn 012
Fipronil 2120 plusmn 174 4697 plusmn 311 8450 plusmn 089 4888 plusmn 392 274 plusmn 027 008 plusmn 001 2247 plusmn 042Thiamethoxam-chlorantraniliprole 2520 plusmn 287 5404 plusmn 465 8380 plusmn 196 5594 plusmn 546 347 plusmn 053 010 plusmn 001 2265 plusmn 072
Control 2160 plusmn 076 4678 plusmn 342 8600 plusmn 096 4940 plusmn 543 267 plusmn 029 008 plusmn 001 2283 plusmn 049F-treatment 4100 ns 3410 2253 ns 5150 2555 3923 4924
F-variety 168232 7665 547500 104356 118242 32308 106597 F-interaction 5980 6113 4824 5760 2921 2769 2552
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Insecticides affected plant growth directly and indirectly (through increased planthop-per densities) Plants treated with cypermethrin and deltamethrin had fewer tillers had alower biomass and under cypermethrin had lower yields than untreated plants (Table 3)Plants treated with fipronil also had reduced yields due to lower grain numbers comparedto untreated plants (Table 3) Although yields were not affected treatment with cartaphydrochloride increased the proportion of unfilled grain compared to untreated plantsof both varieties Cartap hydrochloride and cypermethrin also caused a delay in grainmaturation on IR62 and IR64 respectively relative to thiamethoxam-chlorantraniliproletreated plants (Table 3)
There were several significant interactions tiller numbers on both plants were similarunder buprofezin fipronil and thiamethoxam-chlorantraniliprole treatments but wereotherwise higher on IR62 (Table 3) biomass was lower on IR64 under carbofuran andcypermethrin treatments but was otherwise similar on both plants (Table 3) buprofezindelayed grain maturation on IR62 but maturation was otherwise faster on IR62 comparedto IR64 (Table 3) yields under buprofezin fipronil and thiamethoxam-chlorantraniliprolewere similar for IR62 and IR64 but otherwise yields were higher on IR62 (Table 3) grain
Crops 2021 1 177
numbers were higher on IR62 than IR64 except under fipronil treatments (Table 3) theproportions of grain unfilled were higher on IR62 but were similar between varieties underthe fipronil treatment (Table 3) 1000-grain weight was higher in IR64 but similar on bothvarieties when treated with cartap hydrochloride (Table 3)
4 Discussion
Our results using a highly resistant rice variety and some commonly used insecticidesindicate a range of possible effects on planthoppers and rice plants by combining these twomanagement practices
41 Synergies
The most predictable effect of combining HPR and insecticides is a synergy in reduc-ing BPH numbers This is suggested because planthoppers are weakened by the hostrsquosresistance and should therefore be more susceptible to the toxic effects of insecticides [40]Using spray towers over potted plants Heinrichs et al (1984) [40] examined the combinedeffects of a range of insecticides (that included carbofuran) with resistance by comparingBPH mortality on six resistant and two susceptible rice varieties Their results indicatedthat mortality was greatest when insecticides were applied to BPH on resistant varietiesThe effects were similar when the study was repeated with WBPH for three insecticides(including carbofuran) on the resistant variety N22 (Wbph1 gene) [40] In our study hostresistance reduced egg laying and nymph biomass on non-treated IR62 compared to non-treated IR64 Egg laying was also reduced on plants treated with cypermethrin fipronilor thiamethoxam-chlorantraniliprole (Table 1) Based on the results of our choice andno-choice oviposition bioassays cypermethrin acted mainly as a deterrent of gravid fe-males whereas fipronil and thiamethoxam-chlorantraniliprole directly reduced egg layingDespite similar BPH responses to both the insecticides and resistant host there were no ap-parent synergies between any of these insecticides and resistance in reducing egg numbersin the oviposition bioassay In contrast to these three insecticides our bioassays indicatedthat buprofezin carbofuran and cartap hydrochloride can lead to higher numbers of eggson treated plants compared to untreated plants (Table 1) These effects were not statisticallysignificant and may be exaggerated because bioassays were conducted over relativelyshort time scales For example planthoppers will sometimes respond to stresses such ashigh temperatures by increasing oviposition rates [7172] Furthermore the stimulation ofegg-laying by these chemicals was not apparent in our screenhouse experiment (Table 2)Nevertheless our results do indicate that host resistance reduced the magnitude of anystimulatory effect on oviposition egg numbers on buprofezin-treated carbofuran-treatedand cartap hydrochloride-treated IR64 in the no-choice bioassay were twice as high as onthe corresponding IR62 plants In cases such as these the resistance of IR62 functions as abuffer or lsquoinsurancersquo against possible undesired pesticide effects [1238]
Although buprofezin appeared to stimulate oviposition the growth hormone had astrong impact on nymph development leading to zero survival of exposed nymphs in ourgreenhouse bioassay Cypermethrin fipronil and thiamethoxam-chlorantraniliprole alsoreduced nymph survival to zero in the greenhouse bioassay (Table 1) Where insecticideshave such high efficiencies any fitness losses to BPH due to host resistance then becomeredundant Unlike buprofezin carbofuran also stimulated nymph biomass on IR64 in thesurvival bioassay (Table 1) however nymph biomass was lower on carbofuran-treated IR62than on non-treated IR62 indicating that the hostrsquos resistance buffered against undesiredstimulatory effects on BPH and may have functioned synergistically to augment mortalityand slow BPH growth Such buffering allows insecticides to be used against other pestswithout inducing secondary outbreaks of non-target pests In our screenhouse experimentherbivore densities were generally low on both IR62 and IR64 for most treatments (exceptdeltamethrin) and were below threshold densities for insecticide applicationsmdashwhichare normally about 25ndash50 planthoppers per hill [1373] Under such low densities it wasnot possible to observe the impact of host resistance on planthopper population build-
Crops 2021 1 178
up There was some indication of higher densities on cartap hydrochloride-treated IR62in the screenhouse which may be related to the stimulatory effects of the product onegg laying and the low mortality of nymphs on cartap hydrochloride-treated plants asobserved in the greenhouse bioassays (Table 1) Field studies from the Philippines haveindicated that IR62 effectively reduces BPH densities during wet and dry seasons [4]However farmers are rarely aware of which anti-herbivore resistance their rice varietiespossess and frequently apply pesticides regardless of resistance thereby reducing theirfarm profits [3841] For example Joshi et al (1992) [41] reported widespread outbreaks ofBPH on IR62 in Mindanao Philippines after applications of decamethrin (a pyrethroid) tothe resistant variety Where such resurgence-causing insecticides are applied resistanceseems to be often incapable of reducing the stimulatory effects of the insecticide and itloses its capacity to buffer against outbreaks
42 Antagonisms
Several insecticides are associated with increasing densities of planthoppers andwith resurgence outbreaks in farmersrsquo fields [27293041496174] Indeed patches oflsquohopperburnrsquo (ie dead rice plants resulting from planthopper damage) are often anindicator of high insecticide use in tropical Asia [214] In many cases these resurgence-causing insecticides were once effective in reducing planthopper densities but over timepopulations developed resistance against the products The resulting low toxicity canthen have a stimulatory effect on planthopper feeding and development [2123242743]This may be through hormesis which is a direct response by the planthopper to low-levelexposure to the toxic chemical or it may result from physiological changes to the rice plantsthat ultimately benefit the planthopper [1849] In our screenhouse study we induced aresurgence outbreak of BPH WBPH and to a lesser extent GLH on deltamethrin-treatedrice plants (Table 2) The combined densities of all three herbivores increased almost10 times on the treated plants Furthermore herbivore biomass increased by between fourand six times Although the resurgence effects of deltamethrin have been documentedin previous studies [2029ndash313975] we could not predict from our greenhouse studiesthat any such outbreak would occur Egg laying was unaffected by deltamethrin in ourchoice and no-choice experiments and nymph survival and development both declinedon deltamethrin-treated rice in the survival bioassay (Table 1) The massive increases inplanthopper densities on deltamethrin-treated plants in our study seem to result froma heightened attraction of adult BPH and WBPH to the plants [49] particularly sincenumbers were so low on all other plants including both treated and non-treated plantsHowever once on the plants planthoppers also apparently had enhanced fitness Cuonget al (1997) [31] observed higher population densities of BPH on deltamethrin-treatedrice in Vietnam albeit with lower-level increases on resistant varieties Furthermore in agreenhouse study Azzam et al (2009) [20] found that deltamethrin reduced pre-ovipositiontimes increased the duration of oviposition and increased egg hatchability and femalelongevity Nanthakumar et al (2012) [43] further showed that deltamethrin increased BPHfeeding efficiency on resistant and susceptible rice plants as determined through honeydewproduction and population size In studies with WBPH Salim and Heinrichs (1987) [36]showed deltamethrin to increase settling (antixenosis) feeding and population growthand Suri and Singh (2011) [28] showed deltamethrin to increase WBPH reproductive ratesand nymph survival In our study the observed resurgence of planthoppers was similaron both varieties indicating as was previously noted by Joshi et al (1992) [41] thatresurgence-causing pyrethroids are antagonistic to resistance in IR62
Though not statistically significant cartap hydrochloride also increased BPH numberson IR62 but not on IR64 in our screenhouse study This indicates that the product mayalso be antagonistic to the hostrsquos resistance mechanisms The stronger effect on IR62 maybe related to the differences between IR62 biomass growth rates and anatomy (ie alarger plant with higher tillering) compared to IR64 (Table 3) Once resistance is no longereffective because of chemically-enhanced BPH fitness these plant characteristics which
Crops 2021 1 179
have been related to observed preferences in BPH and other sap-sucking insects [349]could further accelerate the build-up of BPH populations on cartap hydrochloride-treatedplants To further test this idea experiments could be conducted using near-isogenic ricelines that differ mainly in the presence or absence of tagged resistance genes [76] Suchtests would avoid confounding differences in plant anatomy between test varieties IR62resistance to BPH has been well studied [4] The variety is also resistant to WBPH atolder plant stages [54] Therefore in our study deltamethrin was also antagonistic tothis WBPH-resistance (Table 2) However BPH feeding also facilitates WBPH populationgrowth [5477] and there may have been added indirect plant-mediated effects [77] thataugmented WBPH populations on both varieties in our experiment IR62 is also resistant toGLH [45] In our experiments GLH densities also increased on deltamethrin-treated plantsbut the effect was only apparent on IR64 Therefore GLH-resistance in IR62 apparentlybuffered against deltamethrin-associated increases in GLH on the variety as was alsonoted from field studies in Vietnam [31] Deltamethrin is currently not recommended foruse in rice [202878] however the product continues to be marketed in rice-dominatedregions throughout Asia [131479] Although cypermethrin has been associated with BPHoutbreaks and was consequently removed for use on rice in Thailand and Vietnam [62]we did not observe cypermethrin-induced resurgence in our screenhouse experimentNevertheless cypermethrin has been associated with resurgence of BPH in South Asia [61]and we have noted stimulatory effects of cypermethrin on other rice herbivores in thePhilippines (Horgan unpublished data)
43 Phytotoxicity
Phytotoxicity is relatively common following applications of insecticides [182850]Phytotoxic effects may change a plantrsquos physiology without consequences for crop yieldsHowever such changes to plant physiology could have indirect effects on herbivores Forexample deltamethrin methyl parathion and quinalphos produce increases in total sugarsand reducing sugars as well as increasing the protein contents of the rice leaf sheath andleaf blade [28] Several insecticides also reduce plant phenols in ricemdashwhich increases therice plantrsquos susceptibility to BPH and other insects [27] In our screenhouse experiment wenoted several changes to rice anatomy and yields Importantly we noted that cypermethrinand fipronil both reduced rice yields (Table 3) This was not due to indirect herbivoreeffects because planthopper and leafhopper densities were consistently low on rice treatedwith these insecticides The effects therefore probably resulted from direct phytotoxiceffects on the plants Furthermore these effects were most pronounced on treated IR62plants Other minor effects on grain production included increased proportions of unfilledgrain in cartap hydrochloride-treated plants and delays in grain maturation on cartaphydrochloride-treated and cypermethrin-treated plants (Table 3) Despite the high densitiesof BPH and WBPH on deltamethrin-treated plants yields were not significantly lower thanin control non-treated plants Both IR62 and IR64 have been noted for their high toleranceto BPH damage particularly under high nitrogenous fertilizer applications [3456] as usedin our screenhouse experiment Therefore where plants have high tolerance to damageapplying insecticides could also result in profitability losses on rice farms and in the caseof insecticide-induced pest population increases tolerance might buffer against subsequentyield losses There are few reports of the phytotoxicity of insecticides to rice carbofuran istoxic to rice at high concentrations but may also enhance the growth of rice plants throughinteractions with nitrogen-fixing bacteria in the rhizosphere [80] Future studies couldmore carefully assess field-grown plants for possible phytotoxic effects particularly ifinsecticides have the potential to directly reduce yields
44 Management implications
Global trends suggest that resistance in rice has failed to reduce insecticide appli-cations [2] This may be due to aggressive marketing by agrochemical companies andstrategies to increase pesticide sales such as promoting prophylactic applications [16] Be-
Crops 2021 1 180
cause we selected our test insecticides (except the pyrethroids) without any a priori criteriaour results suggest that many currently used insecticides probably have some negativeeffects on yield either directly through phytotoxic effects or indirectly by inducing pestresurgence Furthermore our results indicate that even where pests reach high densitiesas on deltamethrin-treated plants in the screenhouse these densities do not necessarilylead to significant yield reductions
Among the products we used buprofezin although it had little effect on oviposi-tion was effective in reducing nymph survival Furthermore there were only minorapparent phytotoxic effects on the rice plants Buprofezin has a low toxicity to naturalenemies in rice fields [58] and is therefore the most compatible of the insecticides withstrategies to increase rice field biodiversity and restore ecosystem services [32ndash348182]Thiamethoxam-chlorantraniliprole was also effective in reducing both oviposition andnymph survival and had no apparent toxic effects on rice plants However thiamethoxam-chlorantraniliprole and other neonicotinoids although moderately toxic can have severeeffects on beneficial hymenopteran parasitoids [83] although they have apparently lowtoxicity to predatory mirid bugs [84] The remaining insecticides either directly reducedyields or were associated with higher BPH biomass or densities in our experiments
We noted few synergies with HPR although nymph biomass andor survival waslower in cartap hydrochloride and carbofuran-treated IR62 plants compared to correspond-ing IR64 plants Meanwhile cartap hydrochloride and deltamethrin were antagonisticto resistance in IR62 in the screenhouse experiment Among the strongest effects of HPRwas to act as a buffer against GLH population build-up on deltamethrin-treated IR62 andagainst carbofuran-induced increases in BPH nymph biomass in the survival bioassayTherefore HPR may play a role in reducing the possibilities of secondary outbreaks ofBPH and other insects The combined use of HPR and targeted insecticides against BPHcreates redundancy in the rice field and counters measures to manage insecticide resistanceand virulence by unnecessarily exposing plant protection products to evolving pest pop-ulations HPR (and host plant tolerance) can be effective against target pests as revealedin previous studies [4] and could be incorporated into other pest management practicesparticularly agroecological practices such as ecological engineering with which it is highlycompatible [818586]
5 Conclusions
Based on our results and the results of previous studies HPR and the applicationof chemical insecticides against BPH rarely produce synergistic effects Antagonisticeffects where chemicals caused increases in BPH fitness or populations on resistant ricewere more apparent in our study In particular deltamethrin caused an outbreak ofBPH in our screenhouse study Nevertheless HPR can buffer against the insecticide-induced enhancement of herbivore fitness (hormesis) and potentially reduce possibilities ofsecondary outbreaks as was suggested from results with cartap hydrochloride in our studyBased on our results we suggest that carbofuran cartap hydrochloride cypermethrindeltamethrin and fipronil should be avoided in rice particularly in the Philippines (wherethis research was conducted) due to their resurgence-causing potential or because of directreductions to rice yields
Author Contributions Conceptualization FGH methodology FGH AP-C and MLPA formalanalysis FGH and AP-C investigation FGH AP-C and MLPA data curation FGHAP-C and MLPA writingmdashoriginal draft preparation FGH and AP-C writingmdashreview andediting FGH AP-C and MLPA visualization FGH supervision FGH and MLPA projectadministration FGH funding acquisition FGH All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by the Bill and Melinda Gates Foundation (Cereal SystemsInitiative for South Asia [CSISA] OPP52303) and the Global Rice Science Platform (GRiSP) underthe directorship of Achim Dobermann AP-C was partially funded by The Spanish Governmentthrough Becas Faro International
Crops 2021 1 181
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data presented in this study are available on reasonable requestfrom the corresponding author
Acknowledgments The authors thank Fame Ramal Carmencita Bernal Alberto Naredo ReyeulQuintana Vincent Virtudes Marol Recide Jenyrose Geronda and Ellen Genil for assistance duringthis study The contributions of Rodante Abas (sadly deceased) during three decades of entomologicalresearch at IRRI are gratefully acknowledged Rice lines were supplied through the Genebank of IRRI
Conflicts of Interest The authors declare no conflict of interest
References1 Bottrell DG Schoenly KG Resurrecting the ghost of green revolutions past The brown planthopper as a recurring threat to
high-yielding rice production in tropical Asia J Asia-Pac Entomol 2012 15 122ndash140 [CrossRef]2 Horgan FG Integrated pest management for sustainable rice cultivation A holistic approach In Achieving Sustainable Cultivation
of Rice Cultivation Pest and Disease Management Sasaki T Ed Burleigh Dodds Scientific Cambridge UK 2017 pp 309ndash3423 Horgan FG de Freitas TFS Crisol-Martiacutenez E Mundaca EA Bernal CC Nitrogenous fertilizer reduces resistance but
enhances tolerance to the brown planthopper in fast-growing moderately resistant rice Insects 2021 12 989 [CrossRef] [PubMed]4 Horgan FG Pentildealver Cruz A Bernal CC Ramal AF Almazan MLP Wilby A Resistance and tolerance to the brown
planthopper Nilaparvata lugens (Staringl) in rice infested at different growth stages across a gradient of nitrogen applicationsField Crops Res 2018 217 53ndash65 [CrossRef]
5 Hereward JP Cai X Matias AMA Walter GH Xu C Wang Y Migration dynamics of an important rice pest The brownplanthopper (Nilaparvata lugens) across AsiamdashInsights from population genomics Evol Appl 2020 13 2449ndash2459 [CrossRef]
6 Hu C Guo J Fu X Huang Y Gao X Wu K Seasonal migration pattern of Nilaparvata lugens (Hemiptera Delphacidae) overthe Bohai Sea in northern China J Econ Entomol 2018 111 2129ndash2135 [CrossRef]
7 Ma J Wang Y-C Hu Y-Y Lu M-H Wan G-J Chen F-J Liu W-C Zhai B-P Hu G Brown planthopper Nilaparvatalugens was concentrated at the rear of the typhoon Soudelor in eastern China in August 2015 Insect Sci 2018 25 916ndash926[CrossRef]
8 Fujita D Kohli A Horgan FG Rice resistance to planthoppers and leafhoppers Crit Rev Plant Sci 2013 32 162ndash191[CrossRef]
9 Fujii T Yoshida K Kobayashi T Myint KKM Yasui H Sanada-Morimura S Matsumura M Long-term virulencemonitoring of differential cultivars in Japanrsquos immigrant populations of Nilaparvata lugens (Hemiptera Delphacidae) in 2001ndash2009Appl Entomol Zool 2021 56 407ndash418 [CrossRef]
10 Myint KKM Yasui H Takagi M Matsumura M Virulence of long-term laboratory populations of the brown planthopperNilaparvata lugens (Stal) and whitebacked planthopper Sogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) on rice differentialvarieties Appl Entomol Zool 2009 44 149ndash153 [CrossRef]
11 Horgan FG Ramal AF Bentur JS Kumar R Bhanu KV Sarao PS Iswanto EH Van Chien H Phyu MHBernal CC et al Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia againstresistant rice varieties Crop Prot 2015 78 222ndash231 [CrossRef]
12 Horgan FG Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers Crop Prot2018 110 21ndash33 [CrossRef]
13 Uddin ABMA Islam KS Jahan M Ara A Afrin S Farmerrsquos perception about resurgence of brown planthopper NilaparvataLugens (Staringl) in Bangladesh Bangladesh Rice J 2020 23 21ndash33 [CrossRef]
14 Triwidodo H Brown planthoppers infestations and insecticides use pattern in Java Indonesia AGRIVITA J Agric Sci 202042 320ndash330 [CrossRef]
15 Veres A Wyckhuys KA Kiss J Toacuteth F Burgio G Pons X Avilla C Vidal S Razinger J Bazok R et al An update of theWorldwide Integrated Assessment (WIA) on systemic pesticides Part 4 Alternatives in major cropping systems Environ SciPollut Res 2020 27 29867ndash29899 [CrossRef]
16 Bakker L van der Werf W Tittonell P Wyckhuys KA Bianchi FJ Neonicotinoids in global agriculture Evidence for a newpesticide treadmill Ecol Soc 2020 25 26 [CrossRef]
17 Wu J-C Xu J-X Yuan S-Z Liu J-L Jiang Y-H Xu J-F Pesticide-induced susceptibility of rice to brown planthopperNilaparvata lugens Entomol Exp Appl 2001 100 119ndash126 [CrossRef]
18 Wu J-C Qiu H-M Yang G-Q Liu J-L Liu G-J Wilkens RM Effective duration of pesticide-induced susceptibility of riceto brown planthopper (Nilaparvata lugens Staringl Homoptera Delphacidae) and physiological and biochemical changes in riceplants following pesticide application Int J Pest Manag 2004 50 55ndash62 [CrossRef]
19 Wang A-H Wu J-C Yu Y-S Liu J-L Yue J-F Wang MY Selective insecticide-induced stimulation on fecundity andbiochemical changes in Tryporyza incertulas (Lepidoptera Pyralidae) J Econ Entomol 2005 98 1144ndash1149 [CrossRef] [PubMed]
Crops 2021 1 182
20 Azzam S Wang F Wu J-C Shen J Wang L-P Yang G-Q Guo Y-R Comparisons of stimulatory effects of a seriesof concentrations of four insecticides on reproduction in the rice brown planthopper Nilaparvata lugens Staringl (HomopteraDelphacidae) Int J Pest Manag 2009 55 347ndash358 [CrossRef]
21 Ge L-Q Hu J-H Wu J-C Yang G-Q Gu H Insecticide-induced changes in protein RNA and DNA contents in ovary andfat body of female Nilaparvata lugens (Hemiptera Delphacidae) J Econ Entomol 2009 102 1506ndash1514 [CrossRef]
22 Ling S Zhang J Hu L Zhang R Effect of fipronil on the reproduction feeding and relative fitness of brown planthopperNilaparvata lugens Appl Entomol Zool 2009 44 543ndash548 [CrossRef]
23 Ge L-Q Wang L-P Zhao K-F Wu J-C Huang L-J Mating pair combinations of insecticide-treated male and femaleNilaparvata lugens Staringl (Hemiptera Delphacidae) planthoppers influence protein content in the male accessory glands (MAGs)and vitellin content in both fat bodies and ovaries of adult females Pestic Biochem Physiol 2010 98 279ndash288 [CrossRef]
24 Ge L-Q Wu J-C Zhao K-F Chen Y Yang G-Q Induction of Nlvg and suppression of Nljhe gene expression in Nilaparvatalugens (Staringl) (Hemiptera Delphacidae) adult females and males exposed to two insecticides Pestic Biochem Physiol 201098 269ndash278 [CrossRef]
25 Hu J-H Wu J-C Yin J-L Gu H-N Physiology of insecticide-induced stimulation of reproduction in the rice brownplanthopper (Nilaparvata lugens (Staringl)) Dynamics of protein in fat body and ovary Int J Pest Manag 2010 56 23ndash30 [CrossRef]
26 Wang L-P Shen J Ge L-Q Wu J-C Yang G-Q Jahn GC Insecticide-induced increase in the protein content of maleaccessory glands and its effect on the fecundity of females in the brown planthopper Nilaparvata lugens Staringl (HemipteraDelphacidae) Crop Prot 2010 29 1280ndash1285 [CrossRef]
27 Wu J Ge L Liu F Song Q Stanley D Pesticide-induced planthopper population resurgence in rice cropping systems AnnuRev Entomol 2020 65 409ndash429 [CrossRef]
28 Suri KS Singh G Insecticide-induced resurgence of the whitebacked planthopper Sogatella furcifera (Horvath) (HemipteraDelphacidae) on rice varieties with different levels of resistance Crop Prot 2011 30 118ndash124 [CrossRef]
29 Fabellar LT Heinrichs EA Toxicity of insecticides to predators of rice brown planthoppers Nilaparvata lugens (Staringl) (HomopteraDelphacidae) Environ Entomol 1984 13 832ndash837 [CrossRef]
30 Fabellar LT Heinrichs EA Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators Crop Prot1986 5 254ndash258 [CrossRef]
31 Cuong NL Ben PT Phuong LT Chau LM Cohen MB Effect of host plant resistance and insecticide on brown planthopperNilaparvata lugens (Staringl) and predator population development in the Mekong Delta Vietnam Crop Prot 1997 16 707ndash715[CrossRef]
32 Preetha G Stanley J Suresh S Samiyappan R Risk assessment of insecticides used in rice on mirid bug Cyrtorhinuslividipennis Reuter the important predator of brown planthopper Nilaparvata lugens (Stal) Chemosphere 2010 80 498ndash503[CrossRef] [PubMed]
33 Tanaka K Endo S Kazano H Toxicity of insecticides to predators of rice planthoppers Spiders the mirid bug and the dryinidwasp Appl Entomol Zool 2000 35 177ndash187 [CrossRef]
34 Zhang X Xu Q Lu W Liu F Sublethal effects of four synthetic insecticides on the generalist predator Cyrtorhinus lividipennisJ Pest Sci 2015 88 383ndash392 [CrossRef]
35 Settle WH Ariawan H Astuti ET Cahyana W Hakim AL Hindayana D Lestari AS Managing tropical rice peststhrough conservation of generalist natural enemies and alternative prey Ecology 1996 77 1975ndash1988 [CrossRef]
36 Salim M Heinrichs EA Insecticide-induced changes in the levels of resistance of rice cultivars to the whitebacked planthopperSogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) Crop Prot 1987 6 28ndash32 [CrossRef]
37 Yin J-L Xu H-W Wu J-C Hu J-H Yang G-Q Cultivar and insecticide applications affect the physiological development ofthe brown planthopper Nilaparvata lugens (Staringl) (Hemiptera Delphacidae) Environ Entomol 2014 37 206ndash212 [CrossRef]
38 Cheng C Chang W Studies on varietal resistance to the brown planthopper in Taiwan In Brown Planthopper Threat toRice Production in Asia International Rice Research Institute Ed International Rice Research Institute Manila Philippines1979 pp 251ndash271
39 Reissig WH Heinrichs EA Valencia SL Insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on ricevarieties with different levels of resistance Environ Entomol 1982 11 165ndash168 [CrossRef]
40 Heinrichs EA Fabellar LT Basilio RP Wen T-C Medrano F Susceptibility of rice planthoppers Nilaparvata lugens andSogatella furcifera (Homoptera Delphacidae) to insecticides as influenced by level of resistance in the host plant Environ Entomol1984 13 455ndash458 [CrossRef]
41 Joshi R Shepard B Kenmore P Lydia R Insecticide-induced resurgence of brown planthopper (BPH) on IR62 Int Rice ResNewsl 1992 17 9ndash10
42 Zhu Z-R Cheng J Jiang M-X Zhang X-X Complex influence of rice variety fertilization timing and insecticide onpopulation dynamics of Sogatella furcifera (Horvath) Nilaparvata lugens (Staringl) (Homoptera Delphacidae) and their natural enemiesin rice in Hangzhou China J Pest Sci 2004 77 65ndash74 [CrossRef]
43 Nanthakumar M Jhansi Lakshmi V Shashi Bhushan V Balachandran SM Mohan M Decrease of rice plant resistance andinduction of hormesis and carboxylesterase titre in brown planthopper Nilaparvata lugens (Staringl) by xenobiotics Pestic BiochemPhysiol 2012 102 146ndash152 [CrossRef]
Crops 2021 1 183
44 Yao H-W Jiang C-Y Ye G-Y Cheng J-A Studies on cultivar-induced changes in insecticide susceptibility and its relatedenzyme activities of the white-backed planthopper Sogatella furcifera (Horvath)(Homopter Delphacidae) Insect Sci 2002 9 9ndash15
45 Horgan FG Srinivasan TS Bentur JS Kumar R Bhanu KV Sarao PS Van Chien H Almazan MLP Bernal CCRamal AF et al Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers Implicationsfor rice breeding and gene deployment Agronomy 2017 7 62 [CrossRef]
46 Jairin J Phengrat K Teangdeerith S Vanavichit A Toojinda T Mapping of a broad-spectrum brown planthopper resistancegene Bph3 on rice chromosome 6 Mol Breed 2007 19 35ndash44 [CrossRef]
47 Ren J Gao F Wu X Lu X Zeng L Lv J Su X Luo H Ren G Bph32 a novel gene encoding an unknown SCRdomain-containing protein confers resistance against the brown planthopper in rice Sci Rep 2016 6 37645 [CrossRef]
48 Nguyen CD Zheng S-H Sanada-Morimura S Matsumura M Yasui H Fujita D Substitution mapping and characterizationof brown planthopper resistance genes from indica rice variety lsquoPTB33prime (Oryza sativa L) Breed Sci 2021 in press [CrossRef]
49 Chelliah S Heinrichs EA Factors affecting insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on RiceEnviron Entomol 1980 9 773ndash777 [CrossRef]
50 Moore MT Kroumlger R Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growthArch Environ Contam Toxicol 2010 59 574ndash581 [CrossRef]
51 Kim J Hwang I Han M Jang B Effect of insecticide and fungicide on phytotoxicity of herbicide in rice Korean J Weed Sci1986 6 67ndash75
52 Horgan FG Garcia CPF Haverkort F de Jong PW Ferrater JB Changes in insecticide resistance and host range performanceof planthoppers artificially selected to feed on resistant rice Crop Prot 2020 127 104963 [CrossRef] [PubMed]
53 Khush GS Virk PS IR Varieties and Their Impact International Rice Research Institute Manila Philippines 200554 Srinivasan TS Almazan MLP Bernal CC Ramal AF Subbarayalu MK Horgan FG Interactions between nymphs of
Nilaparvata lugens and Sogatella furcifera (Hemiptera Delphacidae) on resistant and susceptible rice varieties Appl Entomol Zool2016 51 81ndash90 [CrossRef]
55 Pentildealver Cruz A Arida A Heong KL Horgan FG Aspects of brown planthopper adaptation to resistant rice varieties withthe Bph3 gene Entomol Exp Appl 2011 141 245ndash257 [CrossRef]
56 Alam SN Cohen MB Detection and analysis of QTLs for resistance to the brown planthopper Nilaparvata lugens in adoubled-haploid rice population Theor Appl Genet 1998 97 1370ndash1379 [CrossRef]
57 Mackill DJ Khush GS IR64 A high-quality and high-yielding mega variety Rice 2018 11 18 [CrossRef] [PubMed]58 Heinrichs EA Basilio RP Valencia SL Buprofezin a selective insecticide for the management of rice planthoppers (Ho-
moptera Delphacidae) and leafhoppers (Homoptera Cicadellidae) Environ Entomol 1984 13 515ndash521 [CrossRef]59 Satapathy MK Anjaneyulu A Use of cypermethrin a synthetic pyrethroid in the control of rice tungro virus disease and its
vector Trop Pest Manag 1984 30 170ndash178 [CrossRef]60 Vorley W Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown
planthopper (BPH) in Kedah Malaysia Int Rice Res Newsl 1987 10 19ndash2061 Uddin A Islam KS Jahan M Ara A Khan MAI Factor influencing the resurgence of brown planthopper in Bangladesh
SAARC J Agric 2020 18 117ndash128 [CrossRef]62 Escalada MM Luecha M Heong KL Social impacts of planthopper outbreaks Case study from Thailand In Rice Planthoppers
Ecology Management Socio Economics and Policy Heong KL Cheng J Escalada MM Eds Zhejiang University PressHangzhou China 2015 pp 209ndash226
63 Zhang CP He HM Yu JZ Hu XQ Zhu YH Wang Q Residues of carbosulfan and its metabolites carbofuran and3-hydroxy carbofuran in rice field ecosystem in China J Environ Sci Health 2016 51 351ndash357 [CrossRef] [PubMed]
64 Samanta S Barman M Samanta NRA Bio-efficacy trials of carbofuran 3 CG against insect pests of rice J Entomol ZoolStud 2020 8 1529ndash1533
65 Donley N The USA lags behind other agricultural nations in banning harmful pesticides Environ Health 2019 18 44 [CrossRef][PubMed]
66 Adams A Gore J Musser F Cook D Catchot A Walker T Dobbins C Efficacy of selected insecticides applied to hybridrice seed J Econ Entomol 2015 109 200ndash206 [CrossRef]
67 Kumar R Singh B Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) Field Bull Environ Contam Toxicol2013 90 482ndash488 [CrossRef]
68 Dhaka S Prajapati C Singh D Singh R Field evaluation of insecticides and bio-pesticides against rice leaf folder CnaphalocrocisMedinalis (Guenee) Ann Plant Prot Sci 2011 19 324ndash326
69 Matsumura M Takeuchi H Satoh M Sanada-Morimura S Otuka A Watanabe T Van Thanh D Species-specific insecticideresistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in East and South-eastAsia Pest Manag Sci 2008 64 1115ndash1121 [CrossRef] [PubMed]
70 Tang J Li J Shao Y Yang B Liu Z Fipronil resistance in the whitebacked planthopper (Sogatella furcifera) Possible resistancemechanisms and cross-resistance Pest Manag Sci 2010 66 121ndash125 [CrossRef]
71 Horgan FG Arida A Ardestani G Almazan MLP Temperature-dependent oviposition and nymph performance revealdistinct thermal niches of coexisting planthoppers with similar thresholds for development PLoS ONE 2020 15 e0235506[CrossRef] [PubMed]
Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]

Crops 2021 1 168
possesses the Bph3 and Bph32 resistance genes derived from the Indian landrace PTB33Because of its relatively high effectiveness IR62 and doner varieties with the Bph3 locuscontaining either the Bph3 or Bph32 resistance genes have been extensively studied inrecent years [46ndash48] We also selected insecticides that are currently used to manage insectherbivores in rice in Asia Many of the products are not specifically recommended for useagainst BPH however resurgence is often the result of applications against other insectpests (ie BPH is a secondary pest) and particularly against rice leaffolders [49] Weexamined the indirect plant-mediated effects of insecticides on BPH because our focuswas on possible insecticide-plant interactions as well as on direct insecticide effects onBPH Therefore we conducted our experiments in greenhouses and in a screenhousethat were largely free of natural enemies Furthermore because of possible phytotoxiceffects of insecticides on rice plants [5051] and the potential role of such phytotoxicityin determining BPH-rice interactions [49] we also assessed insecticide effects on plantgrowth and yields in the screenhouse We discuss our results in terms of applying HPRand insecticide treatments according to IPM principles
2 Materials and Methods21 Herbivore Species
BPH were obtained from colonies reared at the International Rice Research Institute(IRRI) in the Philippines Planthoppers were initially collected in 2009 from rice fields inLaguna Province The founder population (ca 500 individuals) was placed in wire meshcages of 120 times 60 times 60 cm (H timesW times L) under greenhouse conditions (temperatures [T]25ndash37 C 70ndash90 relative humidity [RH] 12h12h daynight [DN]) and was continuouslyreared on ge30 days after sowing [DAS] rice plants (variety = TN1) Feeding plants werereplaced every two weeks The colony received periodic (each year) introgressions ofwild-caught individuals from the same collection sites BPH from the Laguna Provincehave been screened for virulence and are largely adapted to feed on rice varieties with theBph1 bph2 bph5 bph7 Bph18 Bph25 and Bph26 genes [1145] The colonies were also testedfor levels of resistance to insecticides and have moderate resistance to imidacloprid andhigh resistance to BPMC [52]
22 Plant Materials
We used two varieties in our experiments IR62 was selected as a variety with highresistance to BPH populations throughout South and Southeast Asia [53] Resistance inIR62 was derived from PTB33 and is attributed to the Bph3Bph32 resistance genes [4853]BPH has reduced feeding lower weight gain slower development and reduced egg-layingon IR62 compared to susceptible varieties [4] The variety also has moderate resistanceto WBPH and to the green leafhopper Nephotettix virescens (Distant) [GLH] [4554] Fur-thermore resistance derived from PTB33 and associated with the Bph3 locus appears tobe highly durable since BPH populations with virulence against PTB33-derived varietiesincluding IR62 are still rare despite over 30 years of deployment in regions of Mindanao(Philippines) and in Cambodia [55] The variety IR64 was selected as a susceptible vari-ety IR64 has the Bph1 gene for resistance but BPH populations throughout Asia haveadapted to this gene [1153] Nevertheless IR64 has additional quantitative resistance thatcontributes to tolerance (ie a relatively high capacity to withstand damage) of BPH inthe variety [56] IR64 was released by IRRI in 1985 and is considered a mega-variety inAsia [57] Because of its popularity the variety has been enhanced using marker-assistedbreeding to incorporate traits such as flood tolerance (ie IR64-Sub1 released in 2009) [57]For our experiments seeds of both varieties were acquired from the IRRI-germplasm bank
23 Insecticides
We used seven insecticides in our experiments The insecticides were selected basedon their availability at IRRI Buprofezin (2-tert-butylimini-3-isopropyl-5-phenyl-3456-tetrahydro-2H-135-thiadiazin-5-one) (25SC 200 mL haminus1) contains the growth regulator
Crops 2021 1 169
thiadezine and is used against planthoppers and leafhoppers in rice It is the recommendedinsecticide to combat high densities of planthoppers in rice because it has relatively lowtoxicity to natural enemies [58] We used two synthetic pyrethroids cypermethrin (cyan-(3-pheboxyphenyl)-methyl-(3-CS2-dichloroethenyl)-22-dimethyl cyclopropane carboxy-late) (10EC 800 mL haminus1) and deltamethrin ((S)-cyano-3-phenoxybenzyl (1R3R)-3-(22-dibromovinyl)-22-dimethyl-cyclopropane-carboxylate) (25EC 500 mL haminus1) cypermethrinhas been commonly used in rice production to combat rice tungro virus that is transmittedby rice leafhoppers [59] Deltamethrin is a broad-spectrum insecticide that is also some-times used to manage leafhoppers Both pyrethroids were previously noted to cause BPHresurgence [396061] and are currently not recommended for use in rice [62] Neverthe-less cypermethrin and deltamethrin are still commonly used on rice in some countriesincluding Indonesia and Bangladesh [131461] For this reason we included both productsin our experiments We used the carbamate insecticide carbofuran (22-Dimethyl-23-dihydro-1-benzofuran-7-flmethyl-carbamate) (3G 3 kg haminus1) as a systemic insecticidethat is sometimes recommended for control of rice pests including planthoppers [6364]Carbofuran has been banned in the European Union the USA and Canada but is still usedin much of Asia [65] We used thiamethoxam 20 + chlorantraniliprole 20 (3-[(2-chloro-13-thiazol-5-yl)methyl]-5-methyl-135-oxadiazinan-4-ylidene nitramide + 5-bromo-N-[4-chloro-2-methyl-6-(methylcarbamoyl)phenyl]-2-(3-chloropyridin-2-yl)pyrazole-3-carboxamide) (40 WG75 g haminus1) ie a combined neonicotinoid and diamide product as a further systemic in-secticide The product is absorbed through the plant to give broad-spectrum control of ricepests [66] Finally we used fipronil (5-Amino-1-[26-dichloro-4-(trifluoromethyl)phenyl]-4-(trifluoromethanesulfinyl)-1H-pyrazole-3-carbonitrile) (50SC 500 mL haminus1) and car-tap hydrochloride (S-[3-carbamoylsulfanyl-2-(dimethylamino)propyl] carbamothioate-hydrochloride) (a nereistoxin analogue) (50SP 400 g haminus1) as two broad-spectrum in-secticides commonly used to control leaffolders in Asian rice [67ndash70] All insecticideswere applied at recommended rates with quantities estimated based on the surface soilarea of each experimental unit Insecticides were applied by trained pesticide applicatorsusing standard protective equipment in a well-ventilated environment (an insect screen-house) Any evaluations of treated rice plants were conducted at a minimum of 3 dayspost-application
24 Effects of HPR and Insecticides on BPH Preferences
We assessed the effects of the insecticides on oviposition by BPH in a multichoice andin binary choice bioassays For both tests seedlings of IR62 and IR64 were planted individ-ually in 0 pots (5times 5 cm Htimes Diameter) in a greenhouse (T = 25ndash37 C RH = 70ndash90 DN= 12 h12 h) At 27 days after sowing (DAS) the seedlings were treated with one of eachof the seven insecticides as described above or were sprayed with water (ie untreatedcontrol) We used water as a control because some of the products were diluted with waterPlants were returned to the greenhouse at 30 DAS For each variety in the multichoicebioassay eight plants at 30 DASmdashone for each treatmentmdashwere placed together in a wiremesh cage of 120 times 60 times 60 cm (H times W times L) The plants were arranged in a circularconfiguration each separated by 10 cm such that adjacent plants did not touch each otheror the cage walls The cages were replicated five times for each rice variety with plantspositioned randomly during each replication Twenty gravid female N lugens were placedat the center of each cage and allowed to lay eggs for 3 days after which all adults weremanually removed from the cages Eggs were allowed to develop in the plants for a further4 days after which the plants were collected wrapped in moistened tissue paper andstored in a refrigerator Within 5 days all plants were dissected under a stereo microscope(Nikon SMZ-2B Tokyo Japan) to count the egg clusters Egg clusters were removed fromthe plant tissues and the numbers of eggs per cluster were recorded The total number ofeggs was estimated as the sum of eggs per plant from all clusters
For the binary choice tests one plant (either IR62 or IR64) for each insecticide treatmentwas placed together with a control (untreated) plant of the same variety in an acetate cage
Crops 2021 1 170
of 30 times 25 times 50 cm (H timesW times L) (ie two plants per cage) This produced 14 treatmenttimes variety combinations (7 insecticides times 2 varieties) Each combination of plant pairswas replicated three times with cages positioned randomly on greenhouse benches andreplicates of each combination pair placed in separate greenhouse compartments Fivegravid females were placed between the two pots in each cage and allowed to move withinthe cage and lay eggs After 3 days all adults were manually removed from the cages Theplants were collected 4 days after removal of the adults to allow the eggs to develop Plantswere placed in moist tissue paper and stored in a refrigerator until clusters and eggs werecounted under a stereomicroscope (as described above) For both bioassays the replicateswere each placed in separate greenhouse compartments (ie 5 or 3 blocks)
25 Effects of HPR and Insecticides on BPH Fitness
We conducted two BPH antibiosis tests in the greenhouse We examined egg layingby gravid females on each variety and for each insecticide treatment using an ovipositiontest At 30 DAS treated and control plants were individually caged by placing an acetatetube 45 times 5 cm (H times Diameter) with a mesh top over each pot A recently mated gravidBPH was placed inside each cage through a slit in the acetate and was allowed to feed andoviposit After 3 days the females were manually removed from the cages Plants were leftfor 4 days to allow eggs to develop The plants were then collected and dissected under astereo microscope to count the clusters and eggs (as described above)
We examined the effects of host resistance and insecticide treatments on BPH nymphsin a survival test At 30 DAS treated and control plants were individually caged asdescribed above Recently emerged nymphs (le2 instar) were collected from the colonyusing a suction-pooter and placed inside the cages at a density of ten per cage through aslit in the acetate The nymphs were allowed to feed and develop for 15 days after whichthe number of survivors and their developmental stages were recorded For both bioassaysplants were dried in a forced draft oven at 60 C for 7 days and were weighed For thesurvival bioassay the nymphs were also dried and weighed to estimate dry biomass andthereby control for varying consumption of plant liquids Both bioassays were replicatedfive times (ie 8 treatments times 2 varieties times 5 replicates = 80 pots per bioassay) with eachreplicate placed in a separate greenhouse compartment (ie 5 blocks)
26 Effects of HPR and Insecticides in a Screenhouse Environment
The combined effects of insecticide treatments and HPR were investigated using riceplants grown in an insect-proof screenhouse The screenhouse consisted of six concretebays (dimensions 266times 25 m (LtimesW)) filled with flooded paddy soil Temperatures in thescreenhouse varied between 21 and 34 C and RH varied between 70 and 90 Rainwatercould pass through the screenhouse roof during the experiment 160 mm of precipitationwas recorded representing the Philippines dry season The screenhouse had no artificiallighting (ie DN = 12 h12 h)
In one of the bays translucent plastic barriers were erected to delimitate five exper-imental blocks of 150 times 300 m Each block was further divided into eight cubicles of075 m2 using the same plastic materials The barriers were supported by bamboo stakesand reached a height of 150 m and a depth into the paddy soil of approx 030 m After thebarriers were installed two 23 DAS seedlings each of IR62 and IR64 were randomly trans-planted to each cubicle at 25 times 25 cm spacing We used a standard fertilizer regime to sim-ulate farmersrsquo fields The seedlings were treated with ammonium sulphate at 100 kg haminus1
split equally into three applications (ie one basal and two top dressings) Solophos andmuriate of potash was also applied at 20 kg haminus1 as a basal application Plants were contin-uously watered and the cubicles were manually weeded until rice plants were harvestedCubicles in each of the five blocks received one of the eight treatments (seven insecticidesand a control) Treatments were randomly assigned to cubicles within blocks The insecti-cide applications were made at 30 and 60 DAS to simulate farmer practices under intensive
Crops 2021 1 171
rice production systems where farmers apply insecticides ge two times [14] however inour experiment we did not mix insecticides
Rice herbivores including BPH WBPH and GLH occurred at low densities in thescreenhouse Therefore at the opposite end of the screenhouse (separated by a distanceof 20 m) a bay with gt50 DAS TN1 rice plants was infested with high densities of BPH(approx 200 BPH plantminus1) This acted as a potential source of planthoppers to attackthe treated IR62 and IR64 plants in our experiment Natural enemies were largely absentfrom the screenhouse because of the insect-proof cladding (1 mm times 1 mm mesh bore)Microparasitoids (Oligosita spp and Anagrus spp) that could pass through the screenoccasionally attacked planthopper eggs in the screenhouse (estimated 5ndash10 of eggsparasitized unpublished data)
The IR62 and IR64 plants were continuously monitored to assess grain maturationWhen about 85 of the rice grain was mature any arthropods on the plants were collectedusing a vacuum sampler and stored in plastic vials The plants were then destructivelyharvested by carefully pulling the plant from the soil to maintain intact roots The plantroots were washed under running water and each plant was placed inside a paper bagThe plants and arthropods were then dried in a forced draft oven at 60 C for 7 days andweighed Arthropods were separated by species and counted The plants were separatedinto roots shoots and panicles and the numbers of reproductive and vegetative tillerswere recorded Rice grain was removed from the panicles and separated as filled andnon-filled grain The dried grain was counted and weighed
27 Data Analyses
The numbers of egg clusters and eggs laid in rice plants during the multichoice testwere analyzed using univariate general linear models (GLM) on ranked (within replicate)data Separate analyses were conducted for IR62 and IR64 plants Data from the binarychoice bioassays were analyzed using paired t-tests The numbers of egg clusters and eggslaid in treated and non-treated plants during the oviposition bioassay were compared usingunivariate GLM with plant biomass initially included as a covariate but later removedbecause it had no significant effect Cluster and egg numbers were ranked within blocksbefore analyses Average cluster size was estimated by dividing the number of eggs in eachplant by the number of clusters Cluster size was analyzed using univariate GLM removingthe block effect Similarly the proportions of surviving nymphs and nymph biomass fromthe survival bioassay were analyzed using univariate GLMs initially including plantbiomass as a covariate Data from the survival bioassay were ranked before analysesToo few nymphs survived under some treatments to examine development rates or thesex of surviving adults We estimated the combined effects of HPR and each insecticidelsquoxrsquo as -((fitness on untreated IR64 minus fitness on x-treated IR62)fitness on untreated IR64)To estimate the effect of the insecticide alone we removed the effect on HPR which weestimated as fitness on untreated IR62 minus fitness on untreated IR64 Fitness was assessedfrom the oviposition bioassay as the numbers of eggs laid and from the survival bioassayas nymph biomass Where nymph mortality on insecticide-treated IR64 plants approached100 HPR effects were regarded as insignificant
We also examined combined and insecticide effects from the screenhouse data basedon densities of BPH and WBPH (see below) Effects were calculated using insecticide andHPR combinations for each replicated block (N = 5) Estimated effects were comparedacross treatments using univariate GLMs Because three herbivores (BPH WBPH andGLH) occurred in our screenhouse plots we used a split-plot multivariate GLM to analyzeherbivore densities Total planthopper and leafhopper biomass and density and plantgrowth parameters (tiller number biomass yield grain number proportion of grainunfilled and grain size) were analyzed using split-plot univariate GLMs The covariateplant biomass was initially included in analyses of the effects on herbivores but had noeffect and was subsequently removed Similarly the covariate herbivore biomass wasinitially included in analyses of plant growth parameters but had no effect and was
Crops 2021 1 172
subsequently removed The combined effects of insecticide and HPR and the separateeffects of each insecticide (calculated as indicated above) on BPH and WBPH were alsoanalyzed using univariate GLM
Post-hoc Tukey tests were applied to all significant treatment factors Residuals wereplotted following all parametric analyses and were found to be normal and homogenous
3 Results31 Effects of HPR and Insecticides on BPH Preferences
Treatments affected the number of clusters and eggs that were laid in IR62 plants(ranked clusters and eggs F732 = 4738 p lt 0001) and in IR64 plants (clusters F732 = 6229p lt 0001 eggs F732 = 7757 p lt 0001) (Figure 1AB) Fewer eggs were laid on IR64 plantstreated with the pyrethroids (cypermethrin and deltamethrin) fipronil and thiamethoxam-chlorantraniliprole compared to plants treated with buprofezin carbofuran and cartaphydrochloride (Figure 1B) Cypermethrin-treated IR62 plants also had fewer eggs thanbuprofezin-treated IR62 plants (Figure 1A) Although BPH females tended to lay fewereggs in many of the insecticide-treated rice plants in the binary choice experiments therewere no statistically significant differences between the numbers of eggs laid in treated-IR62(t ranges from minus0746 to 3186 p gt 005) or treated-IR64 (t ranges from minus0108 to 2789p gt 005) compared to corresponding non-treated plants (Figure 1CD)
Crops 2021 1 FOR PEER REVIEW 7
herbivore densities Total planthopper and leafhopper biomass and density and plant growth parameters (tiller number biomass yield grain number proportion of grain un-filled and grain size) were analyzed using split-plot univariate GLMs The covariate plant biomass was initially included in analyses of the effects on herbivores but had no effect and was subsequently removed Similarly the covariate herbivore biomass was initially included in analyses of plant growth parameters but had no effect and was subsequently removed The combined effects of insecticide and HPR and the separate effects of each insecticide (calculated as indicated above) on BPH and WBPH were also analyzed using univariate GLM
Post-hoc Tukey tests were applied to all significant treatment factors Residuals were plotted following all parametric analyses and were found to be normal and homogenous
3 Results 31 Effects of HPR and Insecticides on BPH Preferences
Treatments affected the number of clusters and eggs that were laid in IR62 plants (ranked clusters and eggs F732 = 4738 p lt 0001) and in IR64 plants (clusters F732 = 6229 p lt 0001 eggs F732 = 7757 p lt 0001) (Figure 1AB) Fewer eggs were laid on IR64 plants treated with the pyrethroids (cypermethrin and deltamethrin) fipronil and thiameth-oxam-chlorantraniliprole compared to plants treated with buprofezin carbofuran and cartap hydrochloride (Figure 1B) Cypermethrin-treated IR62 plants also had fewer eggs than buprofezin-treated IR62 plants (Figure 1A) Although BPH females tended to lay fewer eggs in many of the insecticide-treated rice plants in the binary choice experiments there were no statistically significant differences between the numbers of eggs laid in treated-IR62 (t ranges from minus0746 to 3186 p gt 005) or treated-IR64 (t ranges from minus0108 to 2789 p gt 005) compared to corresponding non-treated plants (Figure 1CD)
Figure 1 Results from multichoice (AB) and binary choice (CD) bioassays conducted in the green-house Bioassays were conducted separately for IR62 (AC) and IR64 (BD) Plants were treated with one of seven insecticides (A to G) as identified in the legend Control plants were sprayed with water Error bars are presented (N = 5 for (A) and (B) N = 3 for (C) and (D)) Lowercase letters (andashc) indicate homogenous treatment groups (Tukey p lt 005)
A B C D E F G0
100
200
300
400
Treatment
A B C D E F G
Non treatedTreated
A B C D E F G Control
Num
ber o
f egg
s pe
r pla
nt
0
100
200
300
400
A B C D E F G Control
IR62 IR64
A
C
B
D
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliproleH Control
a abab
c
abc
abc bc bc
c
aaa
ab
ab
bb b
Figure 1 Results from multichoice (AB) and binary choice (CD) bioassays conducted in thegreenhouse Bioassays were conducted separately for IR62 (AC) and IR64 (BD) Plants were treatedwith one of seven insecticides (A to G) as identified in the legend Control plants were sprayed withwater Error bars are presented (N = 5 for (AB) N = 3 for (CD)) Lowercase letters (andashc) indicatehomogenous treatment groups (Tukey p lt 005)
32 Effects of HPR and Insecticides on BPH Fitness
More eggs were laid on IR64 plants than on IR62 plants but BPH produced similarnumbers of egg clusters on both plants The lower numbers of eggs in IR62 were due tosmaller clusters on the variety (Table 1) The numbers of eggs and clusters were affectedby insecticide treatment with fewer eggs and clusters on cypermethrin-treated fipronil-treated and thiamethoxam-chlorantraniliprole-treated plants than on plants under the
Crops 2021 1 173
remaining treatments (Table 1) The treatment-effects were not affected by host variety (ieinteractions were non-significant Table 1) Insecticide treatments had no effect on clustersize in the bioassay (Table 1) There was no significant effect of combined insecticide andHPR (F628 = 0449 p = 0840) on observed changes in egg laying during the bioassay butthe estimated insecticide effect was greater for thiamethoxam-chlorantraniliprole fiproniland cypermethrin than for buprofezin cartap hydrochloride and carbofuran (F628 = 3140p = 0018 Figure 2A)
Table 1 Results from greenhouse oviposition and survival tests with IR62 (resistant) and IR64 (susceptible) each treatedwith one of seven insecticides Numbers are means plusmn SEM (N = 5)
Treatments Egg Clusters(Number Plantminus1) 12
Eggs(Number Plantminus1) 12
Cluster Size(Number of Eggs) 1
Nymph Survival(Proportions) 12
Nymph Biomass (mgDry Weight Plantminus1) 12
IR62Buprofezin 1240 plusmn 140 b 5040 plusmn 904 b 393 plusmn 036 000 plusmn 000 a 000 plusmn 000 a
Carbofuran 1300 plusmn 089 b 5380 plusmn 421 b 417 plusmn 026 064 plusmn 064 d 154 plusmn 154 d
Cartaphydrochloride 1360 plusmn 075 b 5200 plusmn 486 b 382 plusmn 029 028 plusmn 028 b 151 plusmn 151 b
Cypermethrin 620 plusmn 169 a 2000 plusmn 608 a 316 plusmn 023 002 plusmn 002 a 002 plusmn 002 a
Deltamethrin 1360 plusmn 160 b 4340 plusmn 566 b 338 plusmn 069 040 plusmn 040 c 120 plusmn 120 c
Fipronil 520 plusmn 146 a 2240 plusmn 735 a 393 plusmn 064 000 plusmn 000 a 000 plusmn 000 a
Thiamethoxam-chlorantraniliprole 380 plusmn 086 a 1700 plusmn 435 a 447 plusmn 032 000 plusmn 000 a 000 plusmn 000 a
Control 1260 plusmn 103 b 4900 plusmn 716 b 388 plusmn 054 080 plusmn 080 d 292 plusmn 292 d
IR64Buprofezin 2100 plusmn 164 10940 plusmn 1346 556 plusmn 117 000 plusmn 000 000 plusmn 000Carbofuran 1820 plusmn 404 9880 plusmn 2939 524 plusmn 074 086 plusmn 086 420 plusmn 420
Cartaphydrochloride 2140 plusmn 619 11460 plusmn 3141 549 plusmn 064 010 plusmn 010 045 plusmn 045
Cypermethrin 380 plusmn 233 1980 plusmn 1155 550 plusmn 058 000 plusmn 000 000 plusmn 000Deltamethrin 1560 plusmn 209 7500 plusmn 1581 473 plusmn 069 040 plusmn 040 201 plusmn 201
Fipronil 380 plusmn 086 2100 plusmn 552 546 plusmn 080 000 plusmn 000 000 plusmn 000Thiamethoxam-chlorantraniliprole 260 plusmn 075 800 plusmn 212 373 plusmn 133 000 plusmn 000 000 plusmn 000
Control 1740 plusmn 364 7520 plusmn 2344 426 plusmn 073 086 plusmn 086 360 plusmn 360F-treatment 3 19898 19898 0541 ns 133811 108451
F-variety 3 2596 ns 7607 7471 1080 ns 0008 ns
F-interaction 3 1840 ns 2034 ns 0610 ns 4902 9496 1 Lowercase letters (andashd) indicate homogenous treatment groups (for IR62 and IR64 combined) 2 Data were ranked within blocks beforeanalysis 3 DF = 164 for varieties 764 for treatments and interactions ns = p gt 005 = p le 001 = p le 0001
Nymph survival and biomass were not affected by rice variety (Table 1) Buprofezinfipronil thiamethoxam-chlorantraniliprole and cypermethrin reduced survival to 0 in thebioassay and all insecticides except carbofuran resulted in lower survival and biomass thanon untreated plants (Table 1) There were significant interactions because of higher nymphmortality and a resulting lower biomass on carbofuran-treated IR62 compared to untreatedIR62 but lower mortality and a higher biomass of nymphs on carbofuran-treated IR64than on untreated IR64 (Table 1) There was no significant effect of insecticide treatment(F628 = 0946 p = 0479) on observed changes in nymph biomass during the bioassaybut the estimated combined effects of pesticide and HPR was greater for thiamethoxam-chlorantraniliprole fipronil cypermethrin and buprofezin than for cartap hydrochlorideand carbofuran (F628 = 5699 p = 0001) (Figure 2B)
Crops 2021 1 174
Crops 2021 1 FOR PEER REVIEW 9
Figure 2 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) egg laying and (B) nymph biomass in greenhouse bioas-says Effects are indicated as proportional changes in fitness parameters compared to untreated IR64 Negative numbers indicate reduced fitness Standard errors are indicated (N = 5) Lowercase letters (ab) indicated homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 1)
Nymph survival and biomass were not affected by rice variety (Table 1) Buprofezin fipronil thiamethoxam-chlorantraniliprole and cypermethrin reduced survival to 0 in the bioassay and all insecticides except carbofuran resulted in lower survival and biomass than on untreated plants (Table 1) There were significant interactions because of higher nymph mortality and a resulting lower biomass on carbofuran-treated IR62 compared to untreated IR62 but lower mortality and a higher biomass of nymphs on carbofuran-treated IR64 than on untreated IR64 (Table 1) There was no significant effect of insecticide treatment (F628 = 0946 p = 0479) on observed changes in nymph biomass during the bio-assay but the estimated combined effects of pesticide and HPR was greater for thiameth-oxam-chlorantraniliprole fipronil cypermethrin and buprofezin than for cartap hydro-chloride and carbofuran (F628 = 5699 p = 0001) (Figure 2B)
33 Effects of HPR and Insecticides on Planthoppers and Leafhoppers Plants in the screenhouse were colonized by BPH WBPH and GLH Densities for
each species were similar on IR62 and IR64 (Wilkrsquos λ = 0994 p = 0952) There was a sig-nificant treatment effect on all three species (Wilkrsquos λ = 0586 p = 0049) because of high densities on deltamethrin-treated plants (Table 2) The numbers and biomass of all three species combined were also greater on deltamethrin-treated plants than on the remaining treatments (Table 2) Host resistance reduced GLH densities under the deltamethrin treat-ment but had no apparent effect on densities under the other treatments (Table 2) There were significant combined effects of HPR and insecticide on the numbers of BPH (F624 = 2686 p = 0039) and WBPH (F624 = 2482 p = 0052) this was almost exclusively due to the resurgence effect of deltamethrin on the planthoppers (BPH F624 = 2686 p = 0039 WBPH F624 = 2482 p = 0052) (Figure 3)
Δ eg
gs(P
ropo
rtion
)
-10
-08
-06
-04
-02
00
02
04
06
Treatment
A B C D E F G
Δ N
ymph
bio
mas
s(P
ropo
rtion
)
-10
-08
-06
-04
-02
00
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
ab
a a
b
a
b b
a a a a a a a a
b b bb
bb
Figure 2 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone(P grey bars) on (A) egg laying and (B) nymph biomass in greenhouse bioassays Effects are indicated as proportionalchanges in fitness parameters compared to untreated IR64 Negative numbers indicate reduced fitness Standard errors areindicated (N = 5) Lowercase letters (ab) indicated homogenous treatment groups based on Tukey pairwise comparisons(p lt 005) (see also Table 1)
33 Effects of HPR and Insecticides on Planthoppers and Leafhoppers
Plants in the screenhouse were colonized by BPH WBPH and GLH Densities foreach species were similar on IR62 and IR64 (Wilkrsquos λ = 0994 p = 0952) There was asignificant treatment effect on all three species (Wilkrsquos λ = 0586 p = 0049) because of highdensities on deltamethrin-treated plants (Table 2) The numbers and biomass of all threespecies combined were also greater on deltamethrin-treated plants than on the remainingtreatments (Table 2) Host resistance reduced GLH densities under the deltamethrintreatment but had no apparent effect on densities under the other treatments (Table 2)There were significant combined effects of HPR and insecticide on the numbers of BPH(F624 = 2686 p = 0039) and WBPH (F624 = 2482 p = 0052) this was almost exclusively dueto the resurgence effect of deltamethrin on the planthoppers (BPH F624 = 2686 p = 0039WBPH F624 = 2482 p = 0052) (Figure 3)
Crops 2021 1 175
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants underseven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density(Number Plantminus1) 1
WBPH Density(Number Plantminus1) 1
GLH Density(Number Plantminus1) 1
Density ofPlanthoppers and
Leafhoppers(Number Plantminus1) 1
Biomass ofPlanthoppers and
Leafhoppers(mg Dry Weight) 1
IR62Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a
Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartaphydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a
Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a
Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a
Thiamethoxam-chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a
IR64Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartaphydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123
Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns
F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Crops 2021 1 FOR PEER REVIEW 10
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants under seven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density (Num-
ber Plantminus1) 1 WBPH Density
(Number Plantminus1) 1 GLH Density
(Number Plantminus1) 1
Density of Planthoppers and Leafhoppers (Number
Plantminus1) 1
Biomass of Planthoppers and Leafhoppers (mg Dry
Weight) 1 IR62
Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartap hydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a Thiamethoxam-
chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a IR64
Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132 Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartap hydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123 Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113 Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052 Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087 F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns 1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatments and interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in the screenhouse Effects are indicated as proportional changes in planthopper densities compared to untreated IR64 Negative numbers indicate reduced densities positive numbers are increased den-sities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
Δ BP
H(N
umbe
r pla
nt-1
)
0
5
10
15
20
Treatment
A B C D E F G
Δ W
BPH
(Num
ber p
lant
-1)
0
20
30
40
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
a a
aa a a
a a a
a
a a
b
b
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimatedpesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in thescreenhouse Effects are indicated as proportional changes in planthopper densities compared to
Crops 2021 1 176
untreated IR64 Negative numbers indicate reduced densities positive numbers are increaseddensities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenoustreatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
34 Direct and Indirect Effects of Insecticides on Rice Development
IR62 plants had more tillers and attained a greater biomass than IR64 plants in thescreenhouse (Table 3) IR62 grain matured faster and plants were harvested earlier than theIR64 plants (Table 3) IR62 plants also produced more grain (number of grain) and hadhigher yields than IR64 but had smaller grain and more unfilled grain than IR64 (Table 3)
Table 3 Rice growth parameters for IR62 (resistant) and IR64 (susceptible) under seven insecticide treatments and forcontrol non-treated plants in a screenhouse environment Numbers are means plusmn SEM
Treatment Tiller Number 1 Plant Biomass(g Dry Weight) 1
Time to Harvest(Days) 1
Weightof Filled Grain(g Dry Weight) 1
Number ofFilled Grain
(1000s) 1
Unfilled Grain(Proportion) 1
1000 GrainWeight 1
IR62Buprofezin 2660 plusmn 206 ab 4643 plusmn 385 ab 7990 plusmn 168 ab 5776 plusmn 683 ab 361plusmn 061 ab 011 plusmn 003 a 2008 plusmn 042 ab
Carbofuran 2900 plusmn 106 ab 5546 plusmn 357 b 7720 plusmn 080 ab 7448 plusmn 691 b 446plusmn 061 ab 011 plusmn 001 a 2103 plusmn 054 b
Cartaphydrochloride 2820 plusmn 327 ab 4896 plusmn 470 ab 7920 plusmn 153 b 6430 plusmn 712 ab 415plusmn 064 ab 012 plusmn 001 b 2023 plusmn 065 a
Cypermethrin 2870 plusmn 301 a 4957 plusmn 562 a 7760 plusmn 103 b 6032 plusmn 822 a 410plusmn 067 ab 014 plusmn 002 a 1965 plusmn 076 a
Deltamethrin 2620 plusmn 345 a 4592 plusmn 588 a 7840 plusmn 108 ab 5698 plusmn 832 ab 379plusmn 047 ab 012 plusmn 001 a 2030 plusmn 063 ab
Fipronil 2380 plusmn 176 ab 4607 plusmn 657 ab 7700 plusmn 179 ab 5266 plusmn 390 a 318 plusmn 032 a 013 plusmn 002 a 2128 plusmn 047 b
Thiamethoxam-chlorantraniliprole 2650 plusmn 250 b 4547 plusmn 429 b 7720 plusmn 102 a 5777 plusmn 688 ab 408 plusmn 061 b 013 plusmn 002 a 2019 plusmn 067 ab
Control 3120 plusmn 247 b 5401 plusmn 555 b 7720 plusmn 080 ab 7045 plusmn 520 b 396plusmn 036 ab 012 plusmn 002 a 2076 plusmn 050 ab
IR64Buprofezin 2340 plusmn 250 5064 plusmn 747 8350 plusmn 124 5080 plusmn 952 270 plusmn 054 008 plusmn 001 2277 plusmn 039Carbofuran 2040 plusmn 097 4591 plusmn 141 8540 plusmn 073 4951 plusmn 312 263 plusmn 023 006 plusmn 000 2295 plusmn 036
Cartaphydrochloride 2120 plusmn 075 4336 plusmn 285 8520 plusmn 140 4273 plusmn 579 238 plusmn 026 015 plusmn 007 2050 plusmn 234
Cypermethrin 1990 plusmn 089 3968 plusmn 272 8710 plusmn 135 3920 plusmn 552 250 plusmn 028 008 plusmn 002 2171 plusmn 070Deltamethrin 1980 plusmn 181 4325 plusmn 357 8460 plusmn 068 4710 plusmn 496 277 plusmn 024 008 plusmn 001 2314 plusmn 012
Fipronil 2120 plusmn 174 4697 plusmn 311 8450 plusmn 089 4888 plusmn 392 274 plusmn 027 008 plusmn 001 2247 plusmn 042Thiamethoxam-chlorantraniliprole 2520 plusmn 287 5404 plusmn 465 8380 plusmn 196 5594 plusmn 546 347 plusmn 053 010 plusmn 001 2265 plusmn 072
Control 2160 plusmn 076 4678 plusmn 342 8600 plusmn 096 4940 plusmn 543 267 plusmn 029 008 plusmn 001 2283 plusmn 049F-treatment 4100 ns 3410 2253 ns 5150 2555 3923 4924
F-variety 168232 7665 547500 104356 118242 32308 106597 F-interaction 5980 6113 4824 5760 2921 2769 2552
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Insecticides affected plant growth directly and indirectly (through increased planthop-per densities) Plants treated with cypermethrin and deltamethrin had fewer tillers had alower biomass and under cypermethrin had lower yields than untreated plants (Table 3)Plants treated with fipronil also had reduced yields due to lower grain numbers comparedto untreated plants (Table 3) Although yields were not affected treatment with cartaphydrochloride increased the proportion of unfilled grain compared to untreated plantsof both varieties Cartap hydrochloride and cypermethrin also caused a delay in grainmaturation on IR62 and IR64 respectively relative to thiamethoxam-chlorantraniliproletreated plants (Table 3)
There were several significant interactions tiller numbers on both plants were similarunder buprofezin fipronil and thiamethoxam-chlorantraniliprole treatments but wereotherwise higher on IR62 (Table 3) biomass was lower on IR64 under carbofuran andcypermethrin treatments but was otherwise similar on both plants (Table 3) buprofezindelayed grain maturation on IR62 but maturation was otherwise faster on IR62 comparedto IR64 (Table 3) yields under buprofezin fipronil and thiamethoxam-chlorantraniliprolewere similar for IR62 and IR64 but otherwise yields were higher on IR62 (Table 3) grain
Crops 2021 1 177
numbers were higher on IR62 than IR64 except under fipronil treatments (Table 3) theproportions of grain unfilled were higher on IR62 but were similar between varieties underthe fipronil treatment (Table 3) 1000-grain weight was higher in IR64 but similar on bothvarieties when treated with cartap hydrochloride (Table 3)
4 Discussion
Our results using a highly resistant rice variety and some commonly used insecticidesindicate a range of possible effects on planthoppers and rice plants by combining these twomanagement practices
41 Synergies
The most predictable effect of combining HPR and insecticides is a synergy in reduc-ing BPH numbers This is suggested because planthoppers are weakened by the hostrsquosresistance and should therefore be more susceptible to the toxic effects of insecticides [40]Using spray towers over potted plants Heinrichs et al (1984) [40] examined the combinedeffects of a range of insecticides (that included carbofuran) with resistance by comparingBPH mortality on six resistant and two susceptible rice varieties Their results indicatedthat mortality was greatest when insecticides were applied to BPH on resistant varietiesThe effects were similar when the study was repeated with WBPH for three insecticides(including carbofuran) on the resistant variety N22 (Wbph1 gene) [40] In our study hostresistance reduced egg laying and nymph biomass on non-treated IR62 compared to non-treated IR64 Egg laying was also reduced on plants treated with cypermethrin fipronilor thiamethoxam-chlorantraniliprole (Table 1) Based on the results of our choice andno-choice oviposition bioassays cypermethrin acted mainly as a deterrent of gravid fe-males whereas fipronil and thiamethoxam-chlorantraniliprole directly reduced egg layingDespite similar BPH responses to both the insecticides and resistant host there were no ap-parent synergies between any of these insecticides and resistance in reducing egg numbersin the oviposition bioassay In contrast to these three insecticides our bioassays indicatedthat buprofezin carbofuran and cartap hydrochloride can lead to higher numbers of eggson treated plants compared to untreated plants (Table 1) These effects were not statisticallysignificant and may be exaggerated because bioassays were conducted over relativelyshort time scales For example planthoppers will sometimes respond to stresses such ashigh temperatures by increasing oviposition rates [7172] Furthermore the stimulation ofegg-laying by these chemicals was not apparent in our screenhouse experiment (Table 2)Nevertheless our results do indicate that host resistance reduced the magnitude of anystimulatory effect on oviposition egg numbers on buprofezin-treated carbofuran-treatedand cartap hydrochloride-treated IR64 in the no-choice bioassay were twice as high as onthe corresponding IR62 plants In cases such as these the resistance of IR62 functions as abuffer or lsquoinsurancersquo against possible undesired pesticide effects [1238]
Although buprofezin appeared to stimulate oviposition the growth hormone had astrong impact on nymph development leading to zero survival of exposed nymphs in ourgreenhouse bioassay Cypermethrin fipronil and thiamethoxam-chlorantraniliprole alsoreduced nymph survival to zero in the greenhouse bioassay (Table 1) Where insecticideshave such high efficiencies any fitness losses to BPH due to host resistance then becomeredundant Unlike buprofezin carbofuran also stimulated nymph biomass on IR64 in thesurvival bioassay (Table 1) however nymph biomass was lower on carbofuran-treated IR62than on non-treated IR62 indicating that the hostrsquos resistance buffered against undesiredstimulatory effects on BPH and may have functioned synergistically to augment mortalityand slow BPH growth Such buffering allows insecticides to be used against other pestswithout inducing secondary outbreaks of non-target pests In our screenhouse experimentherbivore densities were generally low on both IR62 and IR64 for most treatments (exceptdeltamethrin) and were below threshold densities for insecticide applicationsmdashwhichare normally about 25ndash50 planthoppers per hill [1373] Under such low densities it wasnot possible to observe the impact of host resistance on planthopper population build-
Crops 2021 1 178
up There was some indication of higher densities on cartap hydrochloride-treated IR62in the screenhouse which may be related to the stimulatory effects of the product onegg laying and the low mortality of nymphs on cartap hydrochloride-treated plants asobserved in the greenhouse bioassays (Table 1) Field studies from the Philippines haveindicated that IR62 effectively reduces BPH densities during wet and dry seasons [4]However farmers are rarely aware of which anti-herbivore resistance their rice varietiespossess and frequently apply pesticides regardless of resistance thereby reducing theirfarm profits [3841] For example Joshi et al (1992) [41] reported widespread outbreaks ofBPH on IR62 in Mindanao Philippines after applications of decamethrin (a pyrethroid) tothe resistant variety Where such resurgence-causing insecticides are applied resistanceseems to be often incapable of reducing the stimulatory effects of the insecticide and itloses its capacity to buffer against outbreaks
42 Antagonisms
Several insecticides are associated with increasing densities of planthoppers andwith resurgence outbreaks in farmersrsquo fields [27293041496174] Indeed patches oflsquohopperburnrsquo (ie dead rice plants resulting from planthopper damage) are often anindicator of high insecticide use in tropical Asia [214] In many cases these resurgence-causing insecticides were once effective in reducing planthopper densities but over timepopulations developed resistance against the products The resulting low toxicity canthen have a stimulatory effect on planthopper feeding and development [2123242743]This may be through hormesis which is a direct response by the planthopper to low-levelexposure to the toxic chemical or it may result from physiological changes to the rice plantsthat ultimately benefit the planthopper [1849] In our screenhouse study we induced aresurgence outbreak of BPH WBPH and to a lesser extent GLH on deltamethrin-treatedrice plants (Table 2) The combined densities of all three herbivores increased almost10 times on the treated plants Furthermore herbivore biomass increased by between fourand six times Although the resurgence effects of deltamethrin have been documentedin previous studies [2029ndash313975] we could not predict from our greenhouse studiesthat any such outbreak would occur Egg laying was unaffected by deltamethrin in ourchoice and no-choice experiments and nymph survival and development both declinedon deltamethrin-treated rice in the survival bioassay (Table 1) The massive increases inplanthopper densities on deltamethrin-treated plants in our study seem to result froma heightened attraction of adult BPH and WBPH to the plants [49] particularly sincenumbers were so low on all other plants including both treated and non-treated plantsHowever once on the plants planthoppers also apparently had enhanced fitness Cuonget al (1997) [31] observed higher population densities of BPH on deltamethrin-treatedrice in Vietnam albeit with lower-level increases on resistant varieties Furthermore in agreenhouse study Azzam et al (2009) [20] found that deltamethrin reduced pre-ovipositiontimes increased the duration of oviposition and increased egg hatchability and femalelongevity Nanthakumar et al (2012) [43] further showed that deltamethrin increased BPHfeeding efficiency on resistant and susceptible rice plants as determined through honeydewproduction and population size In studies with WBPH Salim and Heinrichs (1987) [36]showed deltamethrin to increase settling (antixenosis) feeding and population growthand Suri and Singh (2011) [28] showed deltamethrin to increase WBPH reproductive ratesand nymph survival In our study the observed resurgence of planthoppers was similaron both varieties indicating as was previously noted by Joshi et al (1992) [41] thatresurgence-causing pyrethroids are antagonistic to resistance in IR62
Though not statistically significant cartap hydrochloride also increased BPH numberson IR62 but not on IR64 in our screenhouse study This indicates that the product mayalso be antagonistic to the hostrsquos resistance mechanisms The stronger effect on IR62 maybe related to the differences between IR62 biomass growth rates and anatomy (ie alarger plant with higher tillering) compared to IR64 (Table 3) Once resistance is no longereffective because of chemically-enhanced BPH fitness these plant characteristics which
Crops 2021 1 179
have been related to observed preferences in BPH and other sap-sucking insects [349]could further accelerate the build-up of BPH populations on cartap hydrochloride-treatedplants To further test this idea experiments could be conducted using near-isogenic ricelines that differ mainly in the presence or absence of tagged resistance genes [76] Suchtests would avoid confounding differences in plant anatomy between test varieties IR62resistance to BPH has been well studied [4] The variety is also resistant to WBPH atolder plant stages [54] Therefore in our study deltamethrin was also antagonistic tothis WBPH-resistance (Table 2) However BPH feeding also facilitates WBPH populationgrowth [5477] and there may have been added indirect plant-mediated effects [77] thataugmented WBPH populations on both varieties in our experiment IR62 is also resistant toGLH [45] In our experiments GLH densities also increased on deltamethrin-treated plantsbut the effect was only apparent on IR64 Therefore GLH-resistance in IR62 apparentlybuffered against deltamethrin-associated increases in GLH on the variety as was alsonoted from field studies in Vietnam [31] Deltamethrin is currently not recommended foruse in rice [202878] however the product continues to be marketed in rice-dominatedregions throughout Asia [131479] Although cypermethrin has been associated with BPHoutbreaks and was consequently removed for use on rice in Thailand and Vietnam [62]we did not observe cypermethrin-induced resurgence in our screenhouse experimentNevertheless cypermethrin has been associated with resurgence of BPH in South Asia [61]and we have noted stimulatory effects of cypermethrin on other rice herbivores in thePhilippines (Horgan unpublished data)
43 Phytotoxicity
Phytotoxicity is relatively common following applications of insecticides [182850]Phytotoxic effects may change a plantrsquos physiology without consequences for crop yieldsHowever such changes to plant physiology could have indirect effects on herbivores Forexample deltamethrin methyl parathion and quinalphos produce increases in total sugarsand reducing sugars as well as increasing the protein contents of the rice leaf sheath andleaf blade [28] Several insecticides also reduce plant phenols in ricemdashwhich increases therice plantrsquos susceptibility to BPH and other insects [27] In our screenhouse experiment wenoted several changes to rice anatomy and yields Importantly we noted that cypermethrinand fipronil both reduced rice yields (Table 3) This was not due to indirect herbivoreeffects because planthopper and leafhopper densities were consistently low on rice treatedwith these insecticides The effects therefore probably resulted from direct phytotoxiceffects on the plants Furthermore these effects were most pronounced on treated IR62plants Other minor effects on grain production included increased proportions of unfilledgrain in cartap hydrochloride-treated plants and delays in grain maturation on cartaphydrochloride-treated and cypermethrin-treated plants (Table 3) Despite the high densitiesof BPH and WBPH on deltamethrin-treated plants yields were not significantly lower thanin control non-treated plants Both IR62 and IR64 have been noted for their high toleranceto BPH damage particularly under high nitrogenous fertilizer applications [3456] as usedin our screenhouse experiment Therefore where plants have high tolerance to damageapplying insecticides could also result in profitability losses on rice farms and in the caseof insecticide-induced pest population increases tolerance might buffer against subsequentyield losses There are few reports of the phytotoxicity of insecticides to rice carbofuran istoxic to rice at high concentrations but may also enhance the growth of rice plants throughinteractions with nitrogen-fixing bacteria in the rhizosphere [80] Future studies couldmore carefully assess field-grown plants for possible phytotoxic effects particularly ifinsecticides have the potential to directly reduce yields
44 Management implications
Global trends suggest that resistance in rice has failed to reduce insecticide appli-cations [2] This may be due to aggressive marketing by agrochemical companies andstrategies to increase pesticide sales such as promoting prophylactic applications [16] Be-
Crops 2021 1 180
cause we selected our test insecticides (except the pyrethroids) without any a priori criteriaour results suggest that many currently used insecticides probably have some negativeeffects on yield either directly through phytotoxic effects or indirectly by inducing pestresurgence Furthermore our results indicate that even where pests reach high densitiesas on deltamethrin-treated plants in the screenhouse these densities do not necessarilylead to significant yield reductions
Among the products we used buprofezin although it had little effect on oviposi-tion was effective in reducing nymph survival Furthermore there were only minorapparent phytotoxic effects on the rice plants Buprofezin has a low toxicity to naturalenemies in rice fields [58] and is therefore the most compatible of the insecticides withstrategies to increase rice field biodiversity and restore ecosystem services [32ndash348182]Thiamethoxam-chlorantraniliprole was also effective in reducing both oviposition andnymph survival and had no apparent toxic effects on rice plants However thiamethoxam-chlorantraniliprole and other neonicotinoids although moderately toxic can have severeeffects on beneficial hymenopteran parasitoids [83] although they have apparently lowtoxicity to predatory mirid bugs [84] The remaining insecticides either directly reducedyields or were associated with higher BPH biomass or densities in our experiments
We noted few synergies with HPR although nymph biomass andor survival waslower in cartap hydrochloride and carbofuran-treated IR62 plants compared to correspond-ing IR64 plants Meanwhile cartap hydrochloride and deltamethrin were antagonisticto resistance in IR62 in the screenhouse experiment Among the strongest effects of HPRwas to act as a buffer against GLH population build-up on deltamethrin-treated IR62 andagainst carbofuran-induced increases in BPH nymph biomass in the survival bioassayTherefore HPR may play a role in reducing the possibilities of secondary outbreaks ofBPH and other insects The combined use of HPR and targeted insecticides against BPHcreates redundancy in the rice field and counters measures to manage insecticide resistanceand virulence by unnecessarily exposing plant protection products to evolving pest pop-ulations HPR (and host plant tolerance) can be effective against target pests as revealedin previous studies [4] and could be incorporated into other pest management practicesparticularly agroecological practices such as ecological engineering with which it is highlycompatible [818586]
5 Conclusions
Based on our results and the results of previous studies HPR and the applicationof chemical insecticides against BPH rarely produce synergistic effects Antagonisticeffects where chemicals caused increases in BPH fitness or populations on resistant ricewere more apparent in our study In particular deltamethrin caused an outbreak ofBPH in our screenhouse study Nevertheless HPR can buffer against the insecticide-induced enhancement of herbivore fitness (hormesis) and potentially reduce possibilities ofsecondary outbreaks as was suggested from results with cartap hydrochloride in our studyBased on our results we suggest that carbofuran cartap hydrochloride cypermethrindeltamethrin and fipronil should be avoided in rice particularly in the Philippines (wherethis research was conducted) due to their resurgence-causing potential or because of directreductions to rice yields
Author Contributions Conceptualization FGH methodology FGH AP-C and MLPA formalanalysis FGH and AP-C investigation FGH AP-C and MLPA data curation FGHAP-C and MLPA writingmdashoriginal draft preparation FGH and AP-C writingmdashreview andediting FGH AP-C and MLPA visualization FGH supervision FGH and MLPA projectadministration FGH funding acquisition FGH All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by the Bill and Melinda Gates Foundation (Cereal SystemsInitiative for South Asia [CSISA] OPP52303) and the Global Rice Science Platform (GRiSP) underthe directorship of Achim Dobermann AP-C was partially funded by The Spanish Governmentthrough Becas Faro International
Crops 2021 1 181
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data presented in this study are available on reasonable requestfrom the corresponding author
Acknowledgments The authors thank Fame Ramal Carmencita Bernal Alberto Naredo ReyeulQuintana Vincent Virtudes Marol Recide Jenyrose Geronda and Ellen Genil for assistance duringthis study The contributions of Rodante Abas (sadly deceased) during three decades of entomologicalresearch at IRRI are gratefully acknowledged Rice lines were supplied through the Genebank of IRRI
Conflicts of Interest The authors declare no conflict of interest
References1 Bottrell DG Schoenly KG Resurrecting the ghost of green revolutions past The brown planthopper as a recurring threat to
high-yielding rice production in tropical Asia J Asia-Pac Entomol 2012 15 122ndash140 [CrossRef]2 Horgan FG Integrated pest management for sustainable rice cultivation A holistic approach In Achieving Sustainable Cultivation
of Rice Cultivation Pest and Disease Management Sasaki T Ed Burleigh Dodds Scientific Cambridge UK 2017 pp 309ndash3423 Horgan FG de Freitas TFS Crisol-Martiacutenez E Mundaca EA Bernal CC Nitrogenous fertilizer reduces resistance but
enhances tolerance to the brown planthopper in fast-growing moderately resistant rice Insects 2021 12 989 [CrossRef] [PubMed]4 Horgan FG Pentildealver Cruz A Bernal CC Ramal AF Almazan MLP Wilby A Resistance and tolerance to the brown
planthopper Nilaparvata lugens (Staringl) in rice infested at different growth stages across a gradient of nitrogen applicationsField Crops Res 2018 217 53ndash65 [CrossRef]
5 Hereward JP Cai X Matias AMA Walter GH Xu C Wang Y Migration dynamics of an important rice pest The brownplanthopper (Nilaparvata lugens) across AsiamdashInsights from population genomics Evol Appl 2020 13 2449ndash2459 [CrossRef]
6 Hu C Guo J Fu X Huang Y Gao X Wu K Seasonal migration pattern of Nilaparvata lugens (Hemiptera Delphacidae) overthe Bohai Sea in northern China J Econ Entomol 2018 111 2129ndash2135 [CrossRef]
7 Ma J Wang Y-C Hu Y-Y Lu M-H Wan G-J Chen F-J Liu W-C Zhai B-P Hu G Brown planthopper Nilaparvatalugens was concentrated at the rear of the typhoon Soudelor in eastern China in August 2015 Insect Sci 2018 25 916ndash926[CrossRef]
8 Fujita D Kohli A Horgan FG Rice resistance to planthoppers and leafhoppers Crit Rev Plant Sci 2013 32 162ndash191[CrossRef]
9 Fujii T Yoshida K Kobayashi T Myint KKM Yasui H Sanada-Morimura S Matsumura M Long-term virulencemonitoring of differential cultivars in Japanrsquos immigrant populations of Nilaparvata lugens (Hemiptera Delphacidae) in 2001ndash2009Appl Entomol Zool 2021 56 407ndash418 [CrossRef]
10 Myint KKM Yasui H Takagi M Matsumura M Virulence of long-term laboratory populations of the brown planthopperNilaparvata lugens (Stal) and whitebacked planthopper Sogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) on rice differentialvarieties Appl Entomol Zool 2009 44 149ndash153 [CrossRef]
11 Horgan FG Ramal AF Bentur JS Kumar R Bhanu KV Sarao PS Iswanto EH Van Chien H Phyu MHBernal CC et al Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia againstresistant rice varieties Crop Prot 2015 78 222ndash231 [CrossRef]
12 Horgan FG Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers Crop Prot2018 110 21ndash33 [CrossRef]
13 Uddin ABMA Islam KS Jahan M Ara A Afrin S Farmerrsquos perception about resurgence of brown planthopper NilaparvataLugens (Staringl) in Bangladesh Bangladesh Rice J 2020 23 21ndash33 [CrossRef]
14 Triwidodo H Brown planthoppers infestations and insecticides use pattern in Java Indonesia AGRIVITA J Agric Sci 202042 320ndash330 [CrossRef]
15 Veres A Wyckhuys KA Kiss J Toacuteth F Burgio G Pons X Avilla C Vidal S Razinger J Bazok R et al An update of theWorldwide Integrated Assessment (WIA) on systemic pesticides Part 4 Alternatives in major cropping systems Environ SciPollut Res 2020 27 29867ndash29899 [CrossRef]
16 Bakker L van der Werf W Tittonell P Wyckhuys KA Bianchi FJ Neonicotinoids in global agriculture Evidence for a newpesticide treadmill Ecol Soc 2020 25 26 [CrossRef]
17 Wu J-C Xu J-X Yuan S-Z Liu J-L Jiang Y-H Xu J-F Pesticide-induced susceptibility of rice to brown planthopperNilaparvata lugens Entomol Exp Appl 2001 100 119ndash126 [CrossRef]
18 Wu J-C Qiu H-M Yang G-Q Liu J-L Liu G-J Wilkens RM Effective duration of pesticide-induced susceptibility of riceto brown planthopper (Nilaparvata lugens Staringl Homoptera Delphacidae) and physiological and biochemical changes in riceplants following pesticide application Int J Pest Manag 2004 50 55ndash62 [CrossRef]
19 Wang A-H Wu J-C Yu Y-S Liu J-L Yue J-F Wang MY Selective insecticide-induced stimulation on fecundity andbiochemical changes in Tryporyza incertulas (Lepidoptera Pyralidae) J Econ Entomol 2005 98 1144ndash1149 [CrossRef] [PubMed]
Crops 2021 1 182
20 Azzam S Wang F Wu J-C Shen J Wang L-P Yang G-Q Guo Y-R Comparisons of stimulatory effects of a seriesof concentrations of four insecticides on reproduction in the rice brown planthopper Nilaparvata lugens Staringl (HomopteraDelphacidae) Int J Pest Manag 2009 55 347ndash358 [CrossRef]
21 Ge L-Q Hu J-H Wu J-C Yang G-Q Gu H Insecticide-induced changes in protein RNA and DNA contents in ovary andfat body of female Nilaparvata lugens (Hemiptera Delphacidae) J Econ Entomol 2009 102 1506ndash1514 [CrossRef]
22 Ling S Zhang J Hu L Zhang R Effect of fipronil on the reproduction feeding and relative fitness of brown planthopperNilaparvata lugens Appl Entomol Zool 2009 44 543ndash548 [CrossRef]
23 Ge L-Q Wang L-P Zhao K-F Wu J-C Huang L-J Mating pair combinations of insecticide-treated male and femaleNilaparvata lugens Staringl (Hemiptera Delphacidae) planthoppers influence protein content in the male accessory glands (MAGs)and vitellin content in both fat bodies and ovaries of adult females Pestic Biochem Physiol 2010 98 279ndash288 [CrossRef]
24 Ge L-Q Wu J-C Zhao K-F Chen Y Yang G-Q Induction of Nlvg and suppression of Nljhe gene expression in Nilaparvatalugens (Staringl) (Hemiptera Delphacidae) adult females and males exposed to two insecticides Pestic Biochem Physiol 201098 269ndash278 [CrossRef]
25 Hu J-H Wu J-C Yin J-L Gu H-N Physiology of insecticide-induced stimulation of reproduction in the rice brownplanthopper (Nilaparvata lugens (Staringl)) Dynamics of protein in fat body and ovary Int J Pest Manag 2010 56 23ndash30 [CrossRef]
26 Wang L-P Shen J Ge L-Q Wu J-C Yang G-Q Jahn GC Insecticide-induced increase in the protein content of maleaccessory glands and its effect on the fecundity of females in the brown planthopper Nilaparvata lugens Staringl (HemipteraDelphacidae) Crop Prot 2010 29 1280ndash1285 [CrossRef]
27 Wu J Ge L Liu F Song Q Stanley D Pesticide-induced planthopper population resurgence in rice cropping systems AnnuRev Entomol 2020 65 409ndash429 [CrossRef]
28 Suri KS Singh G Insecticide-induced resurgence of the whitebacked planthopper Sogatella furcifera (Horvath) (HemipteraDelphacidae) on rice varieties with different levels of resistance Crop Prot 2011 30 118ndash124 [CrossRef]
29 Fabellar LT Heinrichs EA Toxicity of insecticides to predators of rice brown planthoppers Nilaparvata lugens (Staringl) (HomopteraDelphacidae) Environ Entomol 1984 13 832ndash837 [CrossRef]
30 Fabellar LT Heinrichs EA Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators Crop Prot1986 5 254ndash258 [CrossRef]
31 Cuong NL Ben PT Phuong LT Chau LM Cohen MB Effect of host plant resistance and insecticide on brown planthopperNilaparvata lugens (Staringl) and predator population development in the Mekong Delta Vietnam Crop Prot 1997 16 707ndash715[CrossRef]
32 Preetha G Stanley J Suresh S Samiyappan R Risk assessment of insecticides used in rice on mirid bug Cyrtorhinuslividipennis Reuter the important predator of brown planthopper Nilaparvata lugens (Stal) Chemosphere 2010 80 498ndash503[CrossRef] [PubMed]
33 Tanaka K Endo S Kazano H Toxicity of insecticides to predators of rice planthoppers Spiders the mirid bug and the dryinidwasp Appl Entomol Zool 2000 35 177ndash187 [CrossRef]
34 Zhang X Xu Q Lu W Liu F Sublethal effects of four synthetic insecticides on the generalist predator Cyrtorhinus lividipennisJ Pest Sci 2015 88 383ndash392 [CrossRef]
35 Settle WH Ariawan H Astuti ET Cahyana W Hakim AL Hindayana D Lestari AS Managing tropical rice peststhrough conservation of generalist natural enemies and alternative prey Ecology 1996 77 1975ndash1988 [CrossRef]
36 Salim M Heinrichs EA Insecticide-induced changes in the levels of resistance of rice cultivars to the whitebacked planthopperSogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) Crop Prot 1987 6 28ndash32 [CrossRef]
37 Yin J-L Xu H-W Wu J-C Hu J-H Yang G-Q Cultivar and insecticide applications affect the physiological development ofthe brown planthopper Nilaparvata lugens (Staringl) (Hemiptera Delphacidae) Environ Entomol 2014 37 206ndash212 [CrossRef]
38 Cheng C Chang W Studies on varietal resistance to the brown planthopper in Taiwan In Brown Planthopper Threat toRice Production in Asia International Rice Research Institute Ed International Rice Research Institute Manila Philippines1979 pp 251ndash271
39 Reissig WH Heinrichs EA Valencia SL Insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on ricevarieties with different levels of resistance Environ Entomol 1982 11 165ndash168 [CrossRef]
40 Heinrichs EA Fabellar LT Basilio RP Wen T-C Medrano F Susceptibility of rice planthoppers Nilaparvata lugens andSogatella furcifera (Homoptera Delphacidae) to insecticides as influenced by level of resistance in the host plant Environ Entomol1984 13 455ndash458 [CrossRef]
41 Joshi R Shepard B Kenmore P Lydia R Insecticide-induced resurgence of brown planthopper (BPH) on IR62 Int Rice ResNewsl 1992 17 9ndash10
42 Zhu Z-R Cheng J Jiang M-X Zhang X-X Complex influence of rice variety fertilization timing and insecticide onpopulation dynamics of Sogatella furcifera (Horvath) Nilaparvata lugens (Staringl) (Homoptera Delphacidae) and their natural enemiesin rice in Hangzhou China J Pest Sci 2004 77 65ndash74 [CrossRef]
43 Nanthakumar M Jhansi Lakshmi V Shashi Bhushan V Balachandran SM Mohan M Decrease of rice plant resistance andinduction of hormesis and carboxylesterase titre in brown planthopper Nilaparvata lugens (Staringl) by xenobiotics Pestic BiochemPhysiol 2012 102 146ndash152 [CrossRef]
Crops 2021 1 183
44 Yao H-W Jiang C-Y Ye G-Y Cheng J-A Studies on cultivar-induced changes in insecticide susceptibility and its relatedenzyme activities of the white-backed planthopper Sogatella furcifera (Horvath)(Homopter Delphacidae) Insect Sci 2002 9 9ndash15
45 Horgan FG Srinivasan TS Bentur JS Kumar R Bhanu KV Sarao PS Van Chien H Almazan MLP Bernal CCRamal AF et al Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers Implicationsfor rice breeding and gene deployment Agronomy 2017 7 62 [CrossRef]
46 Jairin J Phengrat K Teangdeerith S Vanavichit A Toojinda T Mapping of a broad-spectrum brown planthopper resistancegene Bph3 on rice chromosome 6 Mol Breed 2007 19 35ndash44 [CrossRef]
47 Ren J Gao F Wu X Lu X Zeng L Lv J Su X Luo H Ren G Bph32 a novel gene encoding an unknown SCRdomain-containing protein confers resistance against the brown planthopper in rice Sci Rep 2016 6 37645 [CrossRef]
48 Nguyen CD Zheng S-H Sanada-Morimura S Matsumura M Yasui H Fujita D Substitution mapping and characterizationof brown planthopper resistance genes from indica rice variety lsquoPTB33prime (Oryza sativa L) Breed Sci 2021 in press [CrossRef]
49 Chelliah S Heinrichs EA Factors affecting insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on RiceEnviron Entomol 1980 9 773ndash777 [CrossRef]
50 Moore MT Kroumlger R Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growthArch Environ Contam Toxicol 2010 59 574ndash581 [CrossRef]
51 Kim J Hwang I Han M Jang B Effect of insecticide and fungicide on phytotoxicity of herbicide in rice Korean J Weed Sci1986 6 67ndash75
52 Horgan FG Garcia CPF Haverkort F de Jong PW Ferrater JB Changes in insecticide resistance and host range performanceof planthoppers artificially selected to feed on resistant rice Crop Prot 2020 127 104963 [CrossRef] [PubMed]
53 Khush GS Virk PS IR Varieties and Their Impact International Rice Research Institute Manila Philippines 200554 Srinivasan TS Almazan MLP Bernal CC Ramal AF Subbarayalu MK Horgan FG Interactions between nymphs of
Nilaparvata lugens and Sogatella furcifera (Hemiptera Delphacidae) on resistant and susceptible rice varieties Appl Entomol Zool2016 51 81ndash90 [CrossRef]
55 Pentildealver Cruz A Arida A Heong KL Horgan FG Aspects of brown planthopper adaptation to resistant rice varieties withthe Bph3 gene Entomol Exp Appl 2011 141 245ndash257 [CrossRef]
56 Alam SN Cohen MB Detection and analysis of QTLs for resistance to the brown planthopper Nilaparvata lugens in adoubled-haploid rice population Theor Appl Genet 1998 97 1370ndash1379 [CrossRef]
57 Mackill DJ Khush GS IR64 A high-quality and high-yielding mega variety Rice 2018 11 18 [CrossRef] [PubMed]58 Heinrichs EA Basilio RP Valencia SL Buprofezin a selective insecticide for the management of rice planthoppers (Ho-
moptera Delphacidae) and leafhoppers (Homoptera Cicadellidae) Environ Entomol 1984 13 515ndash521 [CrossRef]59 Satapathy MK Anjaneyulu A Use of cypermethrin a synthetic pyrethroid in the control of rice tungro virus disease and its
vector Trop Pest Manag 1984 30 170ndash178 [CrossRef]60 Vorley W Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown
planthopper (BPH) in Kedah Malaysia Int Rice Res Newsl 1987 10 19ndash2061 Uddin A Islam KS Jahan M Ara A Khan MAI Factor influencing the resurgence of brown planthopper in Bangladesh
SAARC J Agric 2020 18 117ndash128 [CrossRef]62 Escalada MM Luecha M Heong KL Social impacts of planthopper outbreaks Case study from Thailand In Rice Planthoppers
Ecology Management Socio Economics and Policy Heong KL Cheng J Escalada MM Eds Zhejiang University PressHangzhou China 2015 pp 209ndash226
63 Zhang CP He HM Yu JZ Hu XQ Zhu YH Wang Q Residues of carbosulfan and its metabolites carbofuran and3-hydroxy carbofuran in rice field ecosystem in China J Environ Sci Health 2016 51 351ndash357 [CrossRef] [PubMed]
64 Samanta S Barman M Samanta NRA Bio-efficacy trials of carbofuran 3 CG against insect pests of rice J Entomol ZoolStud 2020 8 1529ndash1533
65 Donley N The USA lags behind other agricultural nations in banning harmful pesticides Environ Health 2019 18 44 [CrossRef][PubMed]
66 Adams A Gore J Musser F Cook D Catchot A Walker T Dobbins C Efficacy of selected insecticides applied to hybridrice seed J Econ Entomol 2015 109 200ndash206 [CrossRef]
67 Kumar R Singh B Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) Field Bull Environ Contam Toxicol2013 90 482ndash488 [CrossRef]
68 Dhaka S Prajapati C Singh D Singh R Field evaluation of insecticides and bio-pesticides against rice leaf folder CnaphalocrocisMedinalis (Guenee) Ann Plant Prot Sci 2011 19 324ndash326
69 Matsumura M Takeuchi H Satoh M Sanada-Morimura S Otuka A Watanabe T Van Thanh D Species-specific insecticideresistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in East and South-eastAsia Pest Manag Sci 2008 64 1115ndash1121 [CrossRef] [PubMed]
70 Tang J Li J Shao Y Yang B Liu Z Fipronil resistance in the whitebacked planthopper (Sogatella furcifera) Possible resistancemechanisms and cross-resistance Pest Manag Sci 2010 66 121ndash125 [CrossRef]
71 Horgan FG Arida A Ardestani G Almazan MLP Temperature-dependent oviposition and nymph performance revealdistinct thermal niches of coexisting planthoppers with similar thresholds for development PLoS ONE 2020 15 e0235506[CrossRef] [PubMed]
Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]

Crops 2021 1 169
thiadezine and is used against planthoppers and leafhoppers in rice It is the recommendedinsecticide to combat high densities of planthoppers in rice because it has relatively lowtoxicity to natural enemies [58] We used two synthetic pyrethroids cypermethrin (cyan-(3-pheboxyphenyl)-methyl-(3-CS2-dichloroethenyl)-22-dimethyl cyclopropane carboxy-late) (10EC 800 mL haminus1) and deltamethrin ((S)-cyano-3-phenoxybenzyl (1R3R)-3-(22-dibromovinyl)-22-dimethyl-cyclopropane-carboxylate) (25EC 500 mL haminus1) cypermethrinhas been commonly used in rice production to combat rice tungro virus that is transmittedby rice leafhoppers [59] Deltamethrin is a broad-spectrum insecticide that is also some-times used to manage leafhoppers Both pyrethroids were previously noted to cause BPHresurgence [396061] and are currently not recommended for use in rice [62] Neverthe-less cypermethrin and deltamethrin are still commonly used on rice in some countriesincluding Indonesia and Bangladesh [131461] For this reason we included both productsin our experiments We used the carbamate insecticide carbofuran (22-Dimethyl-23-dihydro-1-benzofuran-7-flmethyl-carbamate) (3G 3 kg haminus1) as a systemic insecticidethat is sometimes recommended for control of rice pests including planthoppers [6364]Carbofuran has been banned in the European Union the USA and Canada but is still usedin much of Asia [65] We used thiamethoxam 20 + chlorantraniliprole 20 (3-[(2-chloro-13-thiazol-5-yl)methyl]-5-methyl-135-oxadiazinan-4-ylidene nitramide + 5-bromo-N-[4-chloro-2-methyl-6-(methylcarbamoyl)phenyl]-2-(3-chloropyridin-2-yl)pyrazole-3-carboxamide) (40 WG75 g haminus1) ie a combined neonicotinoid and diamide product as a further systemic in-secticide The product is absorbed through the plant to give broad-spectrum control of ricepests [66] Finally we used fipronil (5-Amino-1-[26-dichloro-4-(trifluoromethyl)phenyl]-4-(trifluoromethanesulfinyl)-1H-pyrazole-3-carbonitrile) (50SC 500 mL haminus1) and car-tap hydrochloride (S-[3-carbamoylsulfanyl-2-(dimethylamino)propyl] carbamothioate-hydrochloride) (a nereistoxin analogue) (50SP 400 g haminus1) as two broad-spectrum in-secticides commonly used to control leaffolders in Asian rice [67ndash70] All insecticideswere applied at recommended rates with quantities estimated based on the surface soilarea of each experimental unit Insecticides were applied by trained pesticide applicatorsusing standard protective equipment in a well-ventilated environment (an insect screen-house) Any evaluations of treated rice plants were conducted at a minimum of 3 dayspost-application
24 Effects of HPR and Insecticides on BPH Preferences
We assessed the effects of the insecticides on oviposition by BPH in a multichoice andin binary choice bioassays For both tests seedlings of IR62 and IR64 were planted individ-ually in 0 pots (5times 5 cm Htimes Diameter) in a greenhouse (T = 25ndash37 C RH = 70ndash90 DN= 12 h12 h) At 27 days after sowing (DAS) the seedlings were treated with one of eachof the seven insecticides as described above or were sprayed with water (ie untreatedcontrol) We used water as a control because some of the products were diluted with waterPlants were returned to the greenhouse at 30 DAS For each variety in the multichoicebioassay eight plants at 30 DASmdashone for each treatmentmdashwere placed together in a wiremesh cage of 120 times 60 times 60 cm (H times W times L) The plants were arranged in a circularconfiguration each separated by 10 cm such that adjacent plants did not touch each otheror the cage walls The cages were replicated five times for each rice variety with plantspositioned randomly during each replication Twenty gravid female N lugens were placedat the center of each cage and allowed to lay eggs for 3 days after which all adults weremanually removed from the cages Eggs were allowed to develop in the plants for a further4 days after which the plants were collected wrapped in moistened tissue paper andstored in a refrigerator Within 5 days all plants were dissected under a stereo microscope(Nikon SMZ-2B Tokyo Japan) to count the egg clusters Egg clusters were removed fromthe plant tissues and the numbers of eggs per cluster were recorded The total number ofeggs was estimated as the sum of eggs per plant from all clusters
For the binary choice tests one plant (either IR62 or IR64) for each insecticide treatmentwas placed together with a control (untreated) plant of the same variety in an acetate cage
Crops 2021 1 170
of 30 times 25 times 50 cm (H timesW times L) (ie two plants per cage) This produced 14 treatmenttimes variety combinations (7 insecticides times 2 varieties) Each combination of plant pairswas replicated three times with cages positioned randomly on greenhouse benches andreplicates of each combination pair placed in separate greenhouse compartments Fivegravid females were placed between the two pots in each cage and allowed to move withinthe cage and lay eggs After 3 days all adults were manually removed from the cages Theplants were collected 4 days after removal of the adults to allow the eggs to develop Plantswere placed in moist tissue paper and stored in a refrigerator until clusters and eggs werecounted under a stereomicroscope (as described above) For both bioassays the replicateswere each placed in separate greenhouse compartments (ie 5 or 3 blocks)
25 Effects of HPR and Insecticides on BPH Fitness
We conducted two BPH antibiosis tests in the greenhouse We examined egg layingby gravid females on each variety and for each insecticide treatment using an ovipositiontest At 30 DAS treated and control plants were individually caged by placing an acetatetube 45 times 5 cm (H times Diameter) with a mesh top over each pot A recently mated gravidBPH was placed inside each cage through a slit in the acetate and was allowed to feed andoviposit After 3 days the females were manually removed from the cages Plants were leftfor 4 days to allow eggs to develop The plants were then collected and dissected under astereo microscope to count the clusters and eggs (as described above)
We examined the effects of host resistance and insecticide treatments on BPH nymphsin a survival test At 30 DAS treated and control plants were individually caged asdescribed above Recently emerged nymphs (le2 instar) were collected from the colonyusing a suction-pooter and placed inside the cages at a density of ten per cage through aslit in the acetate The nymphs were allowed to feed and develop for 15 days after whichthe number of survivors and their developmental stages were recorded For both bioassaysplants were dried in a forced draft oven at 60 C for 7 days and were weighed For thesurvival bioassay the nymphs were also dried and weighed to estimate dry biomass andthereby control for varying consumption of plant liquids Both bioassays were replicatedfive times (ie 8 treatments times 2 varieties times 5 replicates = 80 pots per bioassay) with eachreplicate placed in a separate greenhouse compartment (ie 5 blocks)
26 Effects of HPR and Insecticides in a Screenhouse Environment
The combined effects of insecticide treatments and HPR were investigated using riceplants grown in an insect-proof screenhouse The screenhouse consisted of six concretebays (dimensions 266times 25 m (LtimesW)) filled with flooded paddy soil Temperatures in thescreenhouse varied between 21 and 34 C and RH varied between 70 and 90 Rainwatercould pass through the screenhouse roof during the experiment 160 mm of precipitationwas recorded representing the Philippines dry season The screenhouse had no artificiallighting (ie DN = 12 h12 h)
In one of the bays translucent plastic barriers were erected to delimitate five exper-imental blocks of 150 times 300 m Each block was further divided into eight cubicles of075 m2 using the same plastic materials The barriers were supported by bamboo stakesand reached a height of 150 m and a depth into the paddy soil of approx 030 m After thebarriers were installed two 23 DAS seedlings each of IR62 and IR64 were randomly trans-planted to each cubicle at 25 times 25 cm spacing We used a standard fertilizer regime to sim-ulate farmersrsquo fields The seedlings were treated with ammonium sulphate at 100 kg haminus1
split equally into three applications (ie one basal and two top dressings) Solophos andmuriate of potash was also applied at 20 kg haminus1 as a basal application Plants were contin-uously watered and the cubicles were manually weeded until rice plants were harvestedCubicles in each of the five blocks received one of the eight treatments (seven insecticidesand a control) Treatments were randomly assigned to cubicles within blocks The insecti-cide applications were made at 30 and 60 DAS to simulate farmer practices under intensive
Crops 2021 1 171
rice production systems where farmers apply insecticides ge two times [14] however inour experiment we did not mix insecticides
Rice herbivores including BPH WBPH and GLH occurred at low densities in thescreenhouse Therefore at the opposite end of the screenhouse (separated by a distanceof 20 m) a bay with gt50 DAS TN1 rice plants was infested with high densities of BPH(approx 200 BPH plantminus1) This acted as a potential source of planthoppers to attackthe treated IR62 and IR64 plants in our experiment Natural enemies were largely absentfrom the screenhouse because of the insect-proof cladding (1 mm times 1 mm mesh bore)Microparasitoids (Oligosita spp and Anagrus spp) that could pass through the screenoccasionally attacked planthopper eggs in the screenhouse (estimated 5ndash10 of eggsparasitized unpublished data)
The IR62 and IR64 plants were continuously monitored to assess grain maturationWhen about 85 of the rice grain was mature any arthropods on the plants were collectedusing a vacuum sampler and stored in plastic vials The plants were then destructivelyharvested by carefully pulling the plant from the soil to maintain intact roots The plantroots were washed under running water and each plant was placed inside a paper bagThe plants and arthropods were then dried in a forced draft oven at 60 C for 7 days andweighed Arthropods were separated by species and counted The plants were separatedinto roots shoots and panicles and the numbers of reproductive and vegetative tillerswere recorded Rice grain was removed from the panicles and separated as filled andnon-filled grain The dried grain was counted and weighed
27 Data Analyses
The numbers of egg clusters and eggs laid in rice plants during the multichoice testwere analyzed using univariate general linear models (GLM) on ranked (within replicate)data Separate analyses were conducted for IR62 and IR64 plants Data from the binarychoice bioassays were analyzed using paired t-tests The numbers of egg clusters and eggslaid in treated and non-treated plants during the oviposition bioassay were compared usingunivariate GLM with plant biomass initially included as a covariate but later removedbecause it had no significant effect Cluster and egg numbers were ranked within blocksbefore analyses Average cluster size was estimated by dividing the number of eggs in eachplant by the number of clusters Cluster size was analyzed using univariate GLM removingthe block effect Similarly the proportions of surviving nymphs and nymph biomass fromthe survival bioassay were analyzed using univariate GLMs initially including plantbiomass as a covariate Data from the survival bioassay were ranked before analysesToo few nymphs survived under some treatments to examine development rates or thesex of surviving adults We estimated the combined effects of HPR and each insecticidelsquoxrsquo as -((fitness on untreated IR64 minus fitness on x-treated IR62)fitness on untreated IR64)To estimate the effect of the insecticide alone we removed the effect on HPR which weestimated as fitness on untreated IR62 minus fitness on untreated IR64 Fitness was assessedfrom the oviposition bioassay as the numbers of eggs laid and from the survival bioassayas nymph biomass Where nymph mortality on insecticide-treated IR64 plants approached100 HPR effects were regarded as insignificant
We also examined combined and insecticide effects from the screenhouse data basedon densities of BPH and WBPH (see below) Effects were calculated using insecticide andHPR combinations for each replicated block (N = 5) Estimated effects were comparedacross treatments using univariate GLMs Because three herbivores (BPH WBPH andGLH) occurred in our screenhouse plots we used a split-plot multivariate GLM to analyzeherbivore densities Total planthopper and leafhopper biomass and density and plantgrowth parameters (tiller number biomass yield grain number proportion of grainunfilled and grain size) were analyzed using split-plot univariate GLMs The covariateplant biomass was initially included in analyses of the effects on herbivores but had noeffect and was subsequently removed Similarly the covariate herbivore biomass wasinitially included in analyses of plant growth parameters but had no effect and was
Crops 2021 1 172
subsequently removed The combined effects of insecticide and HPR and the separateeffects of each insecticide (calculated as indicated above) on BPH and WBPH were alsoanalyzed using univariate GLM
Post-hoc Tukey tests were applied to all significant treatment factors Residuals wereplotted following all parametric analyses and were found to be normal and homogenous
3 Results31 Effects of HPR and Insecticides on BPH Preferences
Treatments affected the number of clusters and eggs that were laid in IR62 plants(ranked clusters and eggs F732 = 4738 p lt 0001) and in IR64 plants (clusters F732 = 6229p lt 0001 eggs F732 = 7757 p lt 0001) (Figure 1AB) Fewer eggs were laid on IR64 plantstreated with the pyrethroids (cypermethrin and deltamethrin) fipronil and thiamethoxam-chlorantraniliprole compared to plants treated with buprofezin carbofuran and cartaphydrochloride (Figure 1B) Cypermethrin-treated IR62 plants also had fewer eggs thanbuprofezin-treated IR62 plants (Figure 1A) Although BPH females tended to lay fewereggs in many of the insecticide-treated rice plants in the binary choice experiments therewere no statistically significant differences between the numbers of eggs laid in treated-IR62(t ranges from minus0746 to 3186 p gt 005) or treated-IR64 (t ranges from minus0108 to 2789p gt 005) compared to corresponding non-treated plants (Figure 1CD)
Crops 2021 1 FOR PEER REVIEW 7
herbivore densities Total planthopper and leafhopper biomass and density and plant growth parameters (tiller number biomass yield grain number proportion of grain un-filled and grain size) were analyzed using split-plot univariate GLMs The covariate plant biomass was initially included in analyses of the effects on herbivores but had no effect and was subsequently removed Similarly the covariate herbivore biomass was initially included in analyses of plant growth parameters but had no effect and was subsequently removed The combined effects of insecticide and HPR and the separate effects of each insecticide (calculated as indicated above) on BPH and WBPH were also analyzed using univariate GLM
Post-hoc Tukey tests were applied to all significant treatment factors Residuals were plotted following all parametric analyses and were found to be normal and homogenous
3 Results 31 Effects of HPR and Insecticides on BPH Preferences
Treatments affected the number of clusters and eggs that were laid in IR62 plants (ranked clusters and eggs F732 = 4738 p lt 0001) and in IR64 plants (clusters F732 = 6229 p lt 0001 eggs F732 = 7757 p lt 0001) (Figure 1AB) Fewer eggs were laid on IR64 plants treated with the pyrethroids (cypermethrin and deltamethrin) fipronil and thiameth-oxam-chlorantraniliprole compared to plants treated with buprofezin carbofuran and cartap hydrochloride (Figure 1B) Cypermethrin-treated IR62 plants also had fewer eggs than buprofezin-treated IR62 plants (Figure 1A) Although BPH females tended to lay fewer eggs in many of the insecticide-treated rice plants in the binary choice experiments there were no statistically significant differences between the numbers of eggs laid in treated-IR62 (t ranges from minus0746 to 3186 p gt 005) or treated-IR64 (t ranges from minus0108 to 2789 p gt 005) compared to corresponding non-treated plants (Figure 1CD)
Figure 1 Results from multichoice (AB) and binary choice (CD) bioassays conducted in the green-house Bioassays were conducted separately for IR62 (AC) and IR64 (BD) Plants were treated with one of seven insecticides (A to G) as identified in the legend Control plants were sprayed with water Error bars are presented (N = 5 for (A) and (B) N = 3 for (C) and (D)) Lowercase letters (andashc) indicate homogenous treatment groups (Tukey p lt 005)
A B C D E F G0
100
200
300
400
Treatment
A B C D E F G
Non treatedTreated
A B C D E F G Control
Num
ber o
f egg
s pe
r pla
nt
0
100
200
300
400
A B C D E F G Control
IR62 IR64
A
C
B
D
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliproleH Control
a abab
c
abc
abc bc bc
c
aaa
ab
ab
bb b
Figure 1 Results from multichoice (AB) and binary choice (CD) bioassays conducted in thegreenhouse Bioassays were conducted separately for IR62 (AC) and IR64 (BD) Plants were treatedwith one of seven insecticides (A to G) as identified in the legend Control plants were sprayed withwater Error bars are presented (N = 5 for (AB) N = 3 for (CD)) Lowercase letters (andashc) indicatehomogenous treatment groups (Tukey p lt 005)
32 Effects of HPR and Insecticides on BPH Fitness
More eggs were laid on IR64 plants than on IR62 plants but BPH produced similarnumbers of egg clusters on both plants The lower numbers of eggs in IR62 were due tosmaller clusters on the variety (Table 1) The numbers of eggs and clusters were affectedby insecticide treatment with fewer eggs and clusters on cypermethrin-treated fipronil-treated and thiamethoxam-chlorantraniliprole-treated plants than on plants under the
Crops 2021 1 173
remaining treatments (Table 1) The treatment-effects were not affected by host variety (ieinteractions were non-significant Table 1) Insecticide treatments had no effect on clustersize in the bioassay (Table 1) There was no significant effect of combined insecticide andHPR (F628 = 0449 p = 0840) on observed changes in egg laying during the bioassay butthe estimated insecticide effect was greater for thiamethoxam-chlorantraniliprole fiproniland cypermethrin than for buprofezin cartap hydrochloride and carbofuran (F628 = 3140p = 0018 Figure 2A)
Table 1 Results from greenhouse oviposition and survival tests with IR62 (resistant) and IR64 (susceptible) each treatedwith one of seven insecticides Numbers are means plusmn SEM (N = 5)
Treatments Egg Clusters(Number Plantminus1) 12
Eggs(Number Plantminus1) 12
Cluster Size(Number of Eggs) 1
Nymph Survival(Proportions) 12
Nymph Biomass (mgDry Weight Plantminus1) 12
IR62Buprofezin 1240 plusmn 140 b 5040 plusmn 904 b 393 plusmn 036 000 plusmn 000 a 000 plusmn 000 a
Carbofuran 1300 plusmn 089 b 5380 plusmn 421 b 417 plusmn 026 064 plusmn 064 d 154 plusmn 154 d
Cartaphydrochloride 1360 plusmn 075 b 5200 plusmn 486 b 382 plusmn 029 028 plusmn 028 b 151 plusmn 151 b
Cypermethrin 620 plusmn 169 a 2000 plusmn 608 a 316 plusmn 023 002 plusmn 002 a 002 plusmn 002 a
Deltamethrin 1360 plusmn 160 b 4340 plusmn 566 b 338 plusmn 069 040 plusmn 040 c 120 plusmn 120 c
Fipronil 520 plusmn 146 a 2240 plusmn 735 a 393 plusmn 064 000 plusmn 000 a 000 plusmn 000 a
Thiamethoxam-chlorantraniliprole 380 plusmn 086 a 1700 plusmn 435 a 447 plusmn 032 000 plusmn 000 a 000 plusmn 000 a
Control 1260 plusmn 103 b 4900 plusmn 716 b 388 plusmn 054 080 plusmn 080 d 292 plusmn 292 d
IR64Buprofezin 2100 plusmn 164 10940 plusmn 1346 556 plusmn 117 000 plusmn 000 000 plusmn 000Carbofuran 1820 plusmn 404 9880 plusmn 2939 524 plusmn 074 086 plusmn 086 420 plusmn 420
Cartaphydrochloride 2140 plusmn 619 11460 plusmn 3141 549 plusmn 064 010 plusmn 010 045 plusmn 045
Cypermethrin 380 plusmn 233 1980 plusmn 1155 550 plusmn 058 000 plusmn 000 000 plusmn 000Deltamethrin 1560 plusmn 209 7500 plusmn 1581 473 plusmn 069 040 plusmn 040 201 plusmn 201
Fipronil 380 plusmn 086 2100 plusmn 552 546 plusmn 080 000 plusmn 000 000 plusmn 000Thiamethoxam-chlorantraniliprole 260 plusmn 075 800 plusmn 212 373 plusmn 133 000 plusmn 000 000 plusmn 000
Control 1740 plusmn 364 7520 plusmn 2344 426 plusmn 073 086 plusmn 086 360 plusmn 360F-treatment 3 19898 19898 0541 ns 133811 108451
F-variety 3 2596 ns 7607 7471 1080 ns 0008 ns
F-interaction 3 1840 ns 2034 ns 0610 ns 4902 9496 1 Lowercase letters (andashd) indicate homogenous treatment groups (for IR62 and IR64 combined) 2 Data were ranked within blocks beforeanalysis 3 DF = 164 for varieties 764 for treatments and interactions ns = p gt 005 = p le 001 = p le 0001
Nymph survival and biomass were not affected by rice variety (Table 1) Buprofezinfipronil thiamethoxam-chlorantraniliprole and cypermethrin reduced survival to 0 in thebioassay and all insecticides except carbofuran resulted in lower survival and biomass thanon untreated plants (Table 1) There were significant interactions because of higher nymphmortality and a resulting lower biomass on carbofuran-treated IR62 compared to untreatedIR62 but lower mortality and a higher biomass of nymphs on carbofuran-treated IR64than on untreated IR64 (Table 1) There was no significant effect of insecticide treatment(F628 = 0946 p = 0479) on observed changes in nymph biomass during the bioassaybut the estimated combined effects of pesticide and HPR was greater for thiamethoxam-chlorantraniliprole fipronil cypermethrin and buprofezin than for cartap hydrochlorideand carbofuran (F628 = 5699 p = 0001) (Figure 2B)
Crops 2021 1 174
Crops 2021 1 FOR PEER REVIEW 9
Figure 2 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) egg laying and (B) nymph biomass in greenhouse bioas-says Effects are indicated as proportional changes in fitness parameters compared to untreated IR64 Negative numbers indicate reduced fitness Standard errors are indicated (N = 5) Lowercase letters (ab) indicated homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 1)
Nymph survival and biomass were not affected by rice variety (Table 1) Buprofezin fipronil thiamethoxam-chlorantraniliprole and cypermethrin reduced survival to 0 in the bioassay and all insecticides except carbofuran resulted in lower survival and biomass than on untreated plants (Table 1) There were significant interactions because of higher nymph mortality and a resulting lower biomass on carbofuran-treated IR62 compared to untreated IR62 but lower mortality and a higher biomass of nymphs on carbofuran-treated IR64 than on untreated IR64 (Table 1) There was no significant effect of insecticide treatment (F628 = 0946 p = 0479) on observed changes in nymph biomass during the bio-assay but the estimated combined effects of pesticide and HPR was greater for thiameth-oxam-chlorantraniliprole fipronil cypermethrin and buprofezin than for cartap hydro-chloride and carbofuran (F628 = 5699 p = 0001) (Figure 2B)
33 Effects of HPR and Insecticides on Planthoppers and Leafhoppers Plants in the screenhouse were colonized by BPH WBPH and GLH Densities for
each species were similar on IR62 and IR64 (Wilkrsquos λ = 0994 p = 0952) There was a sig-nificant treatment effect on all three species (Wilkrsquos λ = 0586 p = 0049) because of high densities on deltamethrin-treated plants (Table 2) The numbers and biomass of all three species combined were also greater on deltamethrin-treated plants than on the remaining treatments (Table 2) Host resistance reduced GLH densities under the deltamethrin treat-ment but had no apparent effect on densities under the other treatments (Table 2) There were significant combined effects of HPR and insecticide on the numbers of BPH (F624 = 2686 p = 0039) and WBPH (F624 = 2482 p = 0052) this was almost exclusively due to the resurgence effect of deltamethrin on the planthoppers (BPH F624 = 2686 p = 0039 WBPH F624 = 2482 p = 0052) (Figure 3)
Δ eg
gs(P
ropo
rtion
)
-10
-08
-06
-04
-02
00
02
04
06
Treatment
A B C D E F G
Δ N
ymph
bio
mas
s(P
ropo
rtion
)
-10
-08
-06
-04
-02
00
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
ab
a a
b
a
b b
a a a a a a a a
b b bb
bb
Figure 2 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone(P grey bars) on (A) egg laying and (B) nymph biomass in greenhouse bioassays Effects are indicated as proportionalchanges in fitness parameters compared to untreated IR64 Negative numbers indicate reduced fitness Standard errors areindicated (N = 5) Lowercase letters (ab) indicated homogenous treatment groups based on Tukey pairwise comparisons(p lt 005) (see also Table 1)
33 Effects of HPR and Insecticides on Planthoppers and Leafhoppers
Plants in the screenhouse were colonized by BPH WBPH and GLH Densities foreach species were similar on IR62 and IR64 (Wilkrsquos λ = 0994 p = 0952) There was asignificant treatment effect on all three species (Wilkrsquos λ = 0586 p = 0049) because of highdensities on deltamethrin-treated plants (Table 2) The numbers and biomass of all threespecies combined were also greater on deltamethrin-treated plants than on the remainingtreatments (Table 2) Host resistance reduced GLH densities under the deltamethrintreatment but had no apparent effect on densities under the other treatments (Table 2)There were significant combined effects of HPR and insecticide on the numbers of BPH(F624 = 2686 p = 0039) and WBPH (F624 = 2482 p = 0052) this was almost exclusively dueto the resurgence effect of deltamethrin on the planthoppers (BPH F624 = 2686 p = 0039WBPH F624 = 2482 p = 0052) (Figure 3)
Crops 2021 1 175
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants underseven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density(Number Plantminus1) 1
WBPH Density(Number Plantminus1) 1
GLH Density(Number Plantminus1) 1
Density ofPlanthoppers and
Leafhoppers(Number Plantminus1) 1
Biomass ofPlanthoppers and
Leafhoppers(mg Dry Weight) 1
IR62Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a
Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartaphydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a
Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a
Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a
Thiamethoxam-chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a
IR64Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartaphydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123
Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns
F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Crops 2021 1 FOR PEER REVIEW 10
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants under seven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density (Num-
ber Plantminus1) 1 WBPH Density
(Number Plantminus1) 1 GLH Density
(Number Plantminus1) 1
Density of Planthoppers and Leafhoppers (Number
Plantminus1) 1
Biomass of Planthoppers and Leafhoppers (mg Dry
Weight) 1 IR62
Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartap hydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a Thiamethoxam-
chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a IR64
Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132 Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartap hydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123 Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113 Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052 Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087 F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns 1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatments and interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in the screenhouse Effects are indicated as proportional changes in planthopper densities compared to untreated IR64 Negative numbers indicate reduced densities positive numbers are increased den-sities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
Δ BP
H(N
umbe
r pla
nt-1
)
0
5
10
15
20
Treatment
A B C D E F G
Δ W
BPH
(Num
ber p
lant
-1)
0
20
30
40
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
a a
aa a a
a a a
a
a a
b
b
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimatedpesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in thescreenhouse Effects are indicated as proportional changes in planthopper densities compared to
Crops 2021 1 176
untreated IR64 Negative numbers indicate reduced densities positive numbers are increaseddensities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenoustreatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
34 Direct and Indirect Effects of Insecticides on Rice Development
IR62 plants had more tillers and attained a greater biomass than IR64 plants in thescreenhouse (Table 3) IR62 grain matured faster and plants were harvested earlier than theIR64 plants (Table 3) IR62 plants also produced more grain (number of grain) and hadhigher yields than IR64 but had smaller grain and more unfilled grain than IR64 (Table 3)
Table 3 Rice growth parameters for IR62 (resistant) and IR64 (susceptible) under seven insecticide treatments and forcontrol non-treated plants in a screenhouse environment Numbers are means plusmn SEM
Treatment Tiller Number 1 Plant Biomass(g Dry Weight) 1
Time to Harvest(Days) 1
Weightof Filled Grain(g Dry Weight) 1
Number ofFilled Grain
(1000s) 1
Unfilled Grain(Proportion) 1
1000 GrainWeight 1
IR62Buprofezin 2660 plusmn 206 ab 4643 plusmn 385 ab 7990 plusmn 168 ab 5776 plusmn 683 ab 361plusmn 061 ab 011 plusmn 003 a 2008 plusmn 042 ab
Carbofuran 2900 plusmn 106 ab 5546 plusmn 357 b 7720 plusmn 080 ab 7448 plusmn 691 b 446plusmn 061 ab 011 plusmn 001 a 2103 plusmn 054 b
Cartaphydrochloride 2820 plusmn 327 ab 4896 plusmn 470 ab 7920 plusmn 153 b 6430 plusmn 712 ab 415plusmn 064 ab 012 plusmn 001 b 2023 plusmn 065 a
Cypermethrin 2870 plusmn 301 a 4957 plusmn 562 a 7760 plusmn 103 b 6032 plusmn 822 a 410plusmn 067 ab 014 plusmn 002 a 1965 plusmn 076 a
Deltamethrin 2620 plusmn 345 a 4592 plusmn 588 a 7840 plusmn 108 ab 5698 plusmn 832 ab 379plusmn 047 ab 012 plusmn 001 a 2030 plusmn 063 ab
Fipronil 2380 plusmn 176 ab 4607 plusmn 657 ab 7700 plusmn 179 ab 5266 plusmn 390 a 318 plusmn 032 a 013 plusmn 002 a 2128 plusmn 047 b
Thiamethoxam-chlorantraniliprole 2650 plusmn 250 b 4547 plusmn 429 b 7720 plusmn 102 a 5777 plusmn 688 ab 408 plusmn 061 b 013 plusmn 002 a 2019 plusmn 067 ab
Control 3120 plusmn 247 b 5401 plusmn 555 b 7720 plusmn 080 ab 7045 plusmn 520 b 396plusmn 036 ab 012 plusmn 002 a 2076 plusmn 050 ab
IR64Buprofezin 2340 plusmn 250 5064 plusmn 747 8350 plusmn 124 5080 plusmn 952 270 plusmn 054 008 plusmn 001 2277 plusmn 039Carbofuran 2040 plusmn 097 4591 plusmn 141 8540 plusmn 073 4951 plusmn 312 263 plusmn 023 006 plusmn 000 2295 plusmn 036
Cartaphydrochloride 2120 plusmn 075 4336 plusmn 285 8520 plusmn 140 4273 plusmn 579 238 plusmn 026 015 plusmn 007 2050 plusmn 234
Cypermethrin 1990 plusmn 089 3968 plusmn 272 8710 plusmn 135 3920 plusmn 552 250 plusmn 028 008 plusmn 002 2171 plusmn 070Deltamethrin 1980 plusmn 181 4325 plusmn 357 8460 plusmn 068 4710 plusmn 496 277 plusmn 024 008 plusmn 001 2314 plusmn 012
Fipronil 2120 plusmn 174 4697 plusmn 311 8450 plusmn 089 4888 plusmn 392 274 plusmn 027 008 plusmn 001 2247 plusmn 042Thiamethoxam-chlorantraniliprole 2520 plusmn 287 5404 plusmn 465 8380 plusmn 196 5594 plusmn 546 347 plusmn 053 010 plusmn 001 2265 plusmn 072
Control 2160 plusmn 076 4678 plusmn 342 8600 plusmn 096 4940 plusmn 543 267 plusmn 029 008 plusmn 001 2283 plusmn 049F-treatment 4100 ns 3410 2253 ns 5150 2555 3923 4924
F-variety 168232 7665 547500 104356 118242 32308 106597 F-interaction 5980 6113 4824 5760 2921 2769 2552
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Insecticides affected plant growth directly and indirectly (through increased planthop-per densities) Plants treated with cypermethrin and deltamethrin had fewer tillers had alower biomass and under cypermethrin had lower yields than untreated plants (Table 3)Plants treated with fipronil also had reduced yields due to lower grain numbers comparedto untreated plants (Table 3) Although yields were not affected treatment with cartaphydrochloride increased the proportion of unfilled grain compared to untreated plantsof both varieties Cartap hydrochloride and cypermethrin also caused a delay in grainmaturation on IR62 and IR64 respectively relative to thiamethoxam-chlorantraniliproletreated plants (Table 3)
There were several significant interactions tiller numbers on both plants were similarunder buprofezin fipronil and thiamethoxam-chlorantraniliprole treatments but wereotherwise higher on IR62 (Table 3) biomass was lower on IR64 under carbofuran andcypermethrin treatments but was otherwise similar on both plants (Table 3) buprofezindelayed grain maturation on IR62 but maturation was otherwise faster on IR62 comparedto IR64 (Table 3) yields under buprofezin fipronil and thiamethoxam-chlorantraniliprolewere similar for IR62 and IR64 but otherwise yields were higher on IR62 (Table 3) grain
Crops 2021 1 177
numbers were higher on IR62 than IR64 except under fipronil treatments (Table 3) theproportions of grain unfilled were higher on IR62 but were similar between varieties underthe fipronil treatment (Table 3) 1000-grain weight was higher in IR64 but similar on bothvarieties when treated with cartap hydrochloride (Table 3)
4 Discussion
Our results using a highly resistant rice variety and some commonly used insecticidesindicate a range of possible effects on planthoppers and rice plants by combining these twomanagement practices
41 Synergies
The most predictable effect of combining HPR and insecticides is a synergy in reduc-ing BPH numbers This is suggested because planthoppers are weakened by the hostrsquosresistance and should therefore be more susceptible to the toxic effects of insecticides [40]Using spray towers over potted plants Heinrichs et al (1984) [40] examined the combinedeffects of a range of insecticides (that included carbofuran) with resistance by comparingBPH mortality on six resistant and two susceptible rice varieties Their results indicatedthat mortality was greatest when insecticides were applied to BPH on resistant varietiesThe effects were similar when the study was repeated with WBPH for three insecticides(including carbofuran) on the resistant variety N22 (Wbph1 gene) [40] In our study hostresistance reduced egg laying and nymph biomass on non-treated IR62 compared to non-treated IR64 Egg laying was also reduced on plants treated with cypermethrin fipronilor thiamethoxam-chlorantraniliprole (Table 1) Based on the results of our choice andno-choice oviposition bioassays cypermethrin acted mainly as a deterrent of gravid fe-males whereas fipronil and thiamethoxam-chlorantraniliprole directly reduced egg layingDespite similar BPH responses to both the insecticides and resistant host there were no ap-parent synergies between any of these insecticides and resistance in reducing egg numbersin the oviposition bioassay In contrast to these three insecticides our bioassays indicatedthat buprofezin carbofuran and cartap hydrochloride can lead to higher numbers of eggson treated plants compared to untreated plants (Table 1) These effects were not statisticallysignificant and may be exaggerated because bioassays were conducted over relativelyshort time scales For example planthoppers will sometimes respond to stresses such ashigh temperatures by increasing oviposition rates [7172] Furthermore the stimulation ofegg-laying by these chemicals was not apparent in our screenhouse experiment (Table 2)Nevertheless our results do indicate that host resistance reduced the magnitude of anystimulatory effect on oviposition egg numbers on buprofezin-treated carbofuran-treatedand cartap hydrochloride-treated IR64 in the no-choice bioassay were twice as high as onthe corresponding IR62 plants In cases such as these the resistance of IR62 functions as abuffer or lsquoinsurancersquo against possible undesired pesticide effects [1238]
Although buprofezin appeared to stimulate oviposition the growth hormone had astrong impact on nymph development leading to zero survival of exposed nymphs in ourgreenhouse bioassay Cypermethrin fipronil and thiamethoxam-chlorantraniliprole alsoreduced nymph survival to zero in the greenhouse bioassay (Table 1) Where insecticideshave such high efficiencies any fitness losses to BPH due to host resistance then becomeredundant Unlike buprofezin carbofuran also stimulated nymph biomass on IR64 in thesurvival bioassay (Table 1) however nymph biomass was lower on carbofuran-treated IR62than on non-treated IR62 indicating that the hostrsquos resistance buffered against undesiredstimulatory effects on BPH and may have functioned synergistically to augment mortalityand slow BPH growth Such buffering allows insecticides to be used against other pestswithout inducing secondary outbreaks of non-target pests In our screenhouse experimentherbivore densities were generally low on both IR62 and IR64 for most treatments (exceptdeltamethrin) and were below threshold densities for insecticide applicationsmdashwhichare normally about 25ndash50 planthoppers per hill [1373] Under such low densities it wasnot possible to observe the impact of host resistance on planthopper population build-
Crops 2021 1 178
up There was some indication of higher densities on cartap hydrochloride-treated IR62in the screenhouse which may be related to the stimulatory effects of the product onegg laying and the low mortality of nymphs on cartap hydrochloride-treated plants asobserved in the greenhouse bioassays (Table 1) Field studies from the Philippines haveindicated that IR62 effectively reduces BPH densities during wet and dry seasons [4]However farmers are rarely aware of which anti-herbivore resistance their rice varietiespossess and frequently apply pesticides regardless of resistance thereby reducing theirfarm profits [3841] For example Joshi et al (1992) [41] reported widespread outbreaks ofBPH on IR62 in Mindanao Philippines after applications of decamethrin (a pyrethroid) tothe resistant variety Where such resurgence-causing insecticides are applied resistanceseems to be often incapable of reducing the stimulatory effects of the insecticide and itloses its capacity to buffer against outbreaks
42 Antagonisms
Several insecticides are associated with increasing densities of planthoppers andwith resurgence outbreaks in farmersrsquo fields [27293041496174] Indeed patches oflsquohopperburnrsquo (ie dead rice plants resulting from planthopper damage) are often anindicator of high insecticide use in tropical Asia [214] In many cases these resurgence-causing insecticides were once effective in reducing planthopper densities but over timepopulations developed resistance against the products The resulting low toxicity canthen have a stimulatory effect on planthopper feeding and development [2123242743]This may be through hormesis which is a direct response by the planthopper to low-levelexposure to the toxic chemical or it may result from physiological changes to the rice plantsthat ultimately benefit the planthopper [1849] In our screenhouse study we induced aresurgence outbreak of BPH WBPH and to a lesser extent GLH on deltamethrin-treatedrice plants (Table 2) The combined densities of all three herbivores increased almost10 times on the treated plants Furthermore herbivore biomass increased by between fourand six times Although the resurgence effects of deltamethrin have been documentedin previous studies [2029ndash313975] we could not predict from our greenhouse studiesthat any such outbreak would occur Egg laying was unaffected by deltamethrin in ourchoice and no-choice experiments and nymph survival and development both declinedon deltamethrin-treated rice in the survival bioassay (Table 1) The massive increases inplanthopper densities on deltamethrin-treated plants in our study seem to result froma heightened attraction of adult BPH and WBPH to the plants [49] particularly sincenumbers were so low on all other plants including both treated and non-treated plantsHowever once on the plants planthoppers also apparently had enhanced fitness Cuonget al (1997) [31] observed higher population densities of BPH on deltamethrin-treatedrice in Vietnam albeit with lower-level increases on resistant varieties Furthermore in agreenhouse study Azzam et al (2009) [20] found that deltamethrin reduced pre-ovipositiontimes increased the duration of oviposition and increased egg hatchability and femalelongevity Nanthakumar et al (2012) [43] further showed that deltamethrin increased BPHfeeding efficiency on resistant and susceptible rice plants as determined through honeydewproduction and population size In studies with WBPH Salim and Heinrichs (1987) [36]showed deltamethrin to increase settling (antixenosis) feeding and population growthand Suri and Singh (2011) [28] showed deltamethrin to increase WBPH reproductive ratesand nymph survival In our study the observed resurgence of planthoppers was similaron both varieties indicating as was previously noted by Joshi et al (1992) [41] thatresurgence-causing pyrethroids are antagonistic to resistance in IR62
Though not statistically significant cartap hydrochloride also increased BPH numberson IR62 but not on IR64 in our screenhouse study This indicates that the product mayalso be antagonistic to the hostrsquos resistance mechanisms The stronger effect on IR62 maybe related to the differences between IR62 biomass growth rates and anatomy (ie alarger plant with higher tillering) compared to IR64 (Table 3) Once resistance is no longereffective because of chemically-enhanced BPH fitness these plant characteristics which
Crops 2021 1 179
have been related to observed preferences in BPH and other sap-sucking insects [349]could further accelerate the build-up of BPH populations on cartap hydrochloride-treatedplants To further test this idea experiments could be conducted using near-isogenic ricelines that differ mainly in the presence or absence of tagged resistance genes [76] Suchtests would avoid confounding differences in plant anatomy between test varieties IR62resistance to BPH has been well studied [4] The variety is also resistant to WBPH atolder plant stages [54] Therefore in our study deltamethrin was also antagonistic tothis WBPH-resistance (Table 2) However BPH feeding also facilitates WBPH populationgrowth [5477] and there may have been added indirect plant-mediated effects [77] thataugmented WBPH populations on both varieties in our experiment IR62 is also resistant toGLH [45] In our experiments GLH densities also increased on deltamethrin-treated plantsbut the effect was only apparent on IR64 Therefore GLH-resistance in IR62 apparentlybuffered against deltamethrin-associated increases in GLH on the variety as was alsonoted from field studies in Vietnam [31] Deltamethrin is currently not recommended foruse in rice [202878] however the product continues to be marketed in rice-dominatedregions throughout Asia [131479] Although cypermethrin has been associated with BPHoutbreaks and was consequently removed for use on rice in Thailand and Vietnam [62]we did not observe cypermethrin-induced resurgence in our screenhouse experimentNevertheless cypermethrin has been associated with resurgence of BPH in South Asia [61]and we have noted stimulatory effects of cypermethrin on other rice herbivores in thePhilippines (Horgan unpublished data)
43 Phytotoxicity
Phytotoxicity is relatively common following applications of insecticides [182850]Phytotoxic effects may change a plantrsquos physiology without consequences for crop yieldsHowever such changes to plant physiology could have indirect effects on herbivores Forexample deltamethrin methyl parathion and quinalphos produce increases in total sugarsand reducing sugars as well as increasing the protein contents of the rice leaf sheath andleaf blade [28] Several insecticides also reduce plant phenols in ricemdashwhich increases therice plantrsquos susceptibility to BPH and other insects [27] In our screenhouse experiment wenoted several changes to rice anatomy and yields Importantly we noted that cypermethrinand fipronil both reduced rice yields (Table 3) This was not due to indirect herbivoreeffects because planthopper and leafhopper densities were consistently low on rice treatedwith these insecticides The effects therefore probably resulted from direct phytotoxiceffects on the plants Furthermore these effects were most pronounced on treated IR62plants Other minor effects on grain production included increased proportions of unfilledgrain in cartap hydrochloride-treated plants and delays in grain maturation on cartaphydrochloride-treated and cypermethrin-treated plants (Table 3) Despite the high densitiesof BPH and WBPH on deltamethrin-treated plants yields were not significantly lower thanin control non-treated plants Both IR62 and IR64 have been noted for their high toleranceto BPH damage particularly under high nitrogenous fertilizer applications [3456] as usedin our screenhouse experiment Therefore where plants have high tolerance to damageapplying insecticides could also result in profitability losses on rice farms and in the caseof insecticide-induced pest population increases tolerance might buffer against subsequentyield losses There are few reports of the phytotoxicity of insecticides to rice carbofuran istoxic to rice at high concentrations but may also enhance the growth of rice plants throughinteractions with nitrogen-fixing bacteria in the rhizosphere [80] Future studies couldmore carefully assess field-grown plants for possible phytotoxic effects particularly ifinsecticides have the potential to directly reduce yields
44 Management implications
Global trends suggest that resistance in rice has failed to reduce insecticide appli-cations [2] This may be due to aggressive marketing by agrochemical companies andstrategies to increase pesticide sales such as promoting prophylactic applications [16] Be-
Crops 2021 1 180
cause we selected our test insecticides (except the pyrethroids) without any a priori criteriaour results suggest that many currently used insecticides probably have some negativeeffects on yield either directly through phytotoxic effects or indirectly by inducing pestresurgence Furthermore our results indicate that even where pests reach high densitiesas on deltamethrin-treated plants in the screenhouse these densities do not necessarilylead to significant yield reductions
Among the products we used buprofezin although it had little effect on oviposi-tion was effective in reducing nymph survival Furthermore there were only minorapparent phytotoxic effects on the rice plants Buprofezin has a low toxicity to naturalenemies in rice fields [58] and is therefore the most compatible of the insecticides withstrategies to increase rice field biodiversity and restore ecosystem services [32ndash348182]Thiamethoxam-chlorantraniliprole was also effective in reducing both oviposition andnymph survival and had no apparent toxic effects on rice plants However thiamethoxam-chlorantraniliprole and other neonicotinoids although moderately toxic can have severeeffects on beneficial hymenopteran parasitoids [83] although they have apparently lowtoxicity to predatory mirid bugs [84] The remaining insecticides either directly reducedyields or were associated with higher BPH biomass or densities in our experiments
We noted few synergies with HPR although nymph biomass andor survival waslower in cartap hydrochloride and carbofuran-treated IR62 plants compared to correspond-ing IR64 plants Meanwhile cartap hydrochloride and deltamethrin were antagonisticto resistance in IR62 in the screenhouse experiment Among the strongest effects of HPRwas to act as a buffer against GLH population build-up on deltamethrin-treated IR62 andagainst carbofuran-induced increases in BPH nymph biomass in the survival bioassayTherefore HPR may play a role in reducing the possibilities of secondary outbreaks ofBPH and other insects The combined use of HPR and targeted insecticides against BPHcreates redundancy in the rice field and counters measures to manage insecticide resistanceand virulence by unnecessarily exposing plant protection products to evolving pest pop-ulations HPR (and host plant tolerance) can be effective against target pests as revealedin previous studies [4] and could be incorporated into other pest management practicesparticularly agroecological practices such as ecological engineering with which it is highlycompatible [818586]
5 Conclusions
Based on our results and the results of previous studies HPR and the applicationof chemical insecticides against BPH rarely produce synergistic effects Antagonisticeffects where chemicals caused increases in BPH fitness or populations on resistant ricewere more apparent in our study In particular deltamethrin caused an outbreak ofBPH in our screenhouse study Nevertheless HPR can buffer against the insecticide-induced enhancement of herbivore fitness (hormesis) and potentially reduce possibilities ofsecondary outbreaks as was suggested from results with cartap hydrochloride in our studyBased on our results we suggest that carbofuran cartap hydrochloride cypermethrindeltamethrin and fipronil should be avoided in rice particularly in the Philippines (wherethis research was conducted) due to their resurgence-causing potential or because of directreductions to rice yields
Author Contributions Conceptualization FGH methodology FGH AP-C and MLPA formalanalysis FGH and AP-C investigation FGH AP-C and MLPA data curation FGHAP-C and MLPA writingmdashoriginal draft preparation FGH and AP-C writingmdashreview andediting FGH AP-C and MLPA visualization FGH supervision FGH and MLPA projectadministration FGH funding acquisition FGH All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by the Bill and Melinda Gates Foundation (Cereal SystemsInitiative for South Asia [CSISA] OPP52303) and the Global Rice Science Platform (GRiSP) underthe directorship of Achim Dobermann AP-C was partially funded by The Spanish Governmentthrough Becas Faro International
Crops 2021 1 181
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data presented in this study are available on reasonable requestfrom the corresponding author
Acknowledgments The authors thank Fame Ramal Carmencita Bernal Alberto Naredo ReyeulQuintana Vincent Virtudes Marol Recide Jenyrose Geronda and Ellen Genil for assistance duringthis study The contributions of Rodante Abas (sadly deceased) during three decades of entomologicalresearch at IRRI are gratefully acknowledged Rice lines were supplied through the Genebank of IRRI
Conflicts of Interest The authors declare no conflict of interest
References1 Bottrell DG Schoenly KG Resurrecting the ghost of green revolutions past The brown planthopper as a recurring threat to
high-yielding rice production in tropical Asia J Asia-Pac Entomol 2012 15 122ndash140 [CrossRef]2 Horgan FG Integrated pest management for sustainable rice cultivation A holistic approach In Achieving Sustainable Cultivation
of Rice Cultivation Pest and Disease Management Sasaki T Ed Burleigh Dodds Scientific Cambridge UK 2017 pp 309ndash3423 Horgan FG de Freitas TFS Crisol-Martiacutenez E Mundaca EA Bernal CC Nitrogenous fertilizer reduces resistance but
enhances tolerance to the brown planthopper in fast-growing moderately resistant rice Insects 2021 12 989 [CrossRef] [PubMed]4 Horgan FG Pentildealver Cruz A Bernal CC Ramal AF Almazan MLP Wilby A Resistance and tolerance to the brown
planthopper Nilaparvata lugens (Staringl) in rice infested at different growth stages across a gradient of nitrogen applicationsField Crops Res 2018 217 53ndash65 [CrossRef]
5 Hereward JP Cai X Matias AMA Walter GH Xu C Wang Y Migration dynamics of an important rice pest The brownplanthopper (Nilaparvata lugens) across AsiamdashInsights from population genomics Evol Appl 2020 13 2449ndash2459 [CrossRef]
6 Hu C Guo J Fu X Huang Y Gao X Wu K Seasonal migration pattern of Nilaparvata lugens (Hemiptera Delphacidae) overthe Bohai Sea in northern China J Econ Entomol 2018 111 2129ndash2135 [CrossRef]
7 Ma J Wang Y-C Hu Y-Y Lu M-H Wan G-J Chen F-J Liu W-C Zhai B-P Hu G Brown planthopper Nilaparvatalugens was concentrated at the rear of the typhoon Soudelor in eastern China in August 2015 Insect Sci 2018 25 916ndash926[CrossRef]
8 Fujita D Kohli A Horgan FG Rice resistance to planthoppers and leafhoppers Crit Rev Plant Sci 2013 32 162ndash191[CrossRef]
9 Fujii T Yoshida K Kobayashi T Myint KKM Yasui H Sanada-Morimura S Matsumura M Long-term virulencemonitoring of differential cultivars in Japanrsquos immigrant populations of Nilaparvata lugens (Hemiptera Delphacidae) in 2001ndash2009Appl Entomol Zool 2021 56 407ndash418 [CrossRef]
10 Myint KKM Yasui H Takagi M Matsumura M Virulence of long-term laboratory populations of the brown planthopperNilaparvata lugens (Stal) and whitebacked planthopper Sogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) on rice differentialvarieties Appl Entomol Zool 2009 44 149ndash153 [CrossRef]
11 Horgan FG Ramal AF Bentur JS Kumar R Bhanu KV Sarao PS Iswanto EH Van Chien H Phyu MHBernal CC et al Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia againstresistant rice varieties Crop Prot 2015 78 222ndash231 [CrossRef]
12 Horgan FG Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers Crop Prot2018 110 21ndash33 [CrossRef]
13 Uddin ABMA Islam KS Jahan M Ara A Afrin S Farmerrsquos perception about resurgence of brown planthopper NilaparvataLugens (Staringl) in Bangladesh Bangladesh Rice J 2020 23 21ndash33 [CrossRef]
14 Triwidodo H Brown planthoppers infestations and insecticides use pattern in Java Indonesia AGRIVITA J Agric Sci 202042 320ndash330 [CrossRef]
15 Veres A Wyckhuys KA Kiss J Toacuteth F Burgio G Pons X Avilla C Vidal S Razinger J Bazok R et al An update of theWorldwide Integrated Assessment (WIA) on systemic pesticides Part 4 Alternatives in major cropping systems Environ SciPollut Res 2020 27 29867ndash29899 [CrossRef]
16 Bakker L van der Werf W Tittonell P Wyckhuys KA Bianchi FJ Neonicotinoids in global agriculture Evidence for a newpesticide treadmill Ecol Soc 2020 25 26 [CrossRef]
17 Wu J-C Xu J-X Yuan S-Z Liu J-L Jiang Y-H Xu J-F Pesticide-induced susceptibility of rice to brown planthopperNilaparvata lugens Entomol Exp Appl 2001 100 119ndash126 [CrossRef]
18 Wu J-C Qiu H-M Yang G-Q Liu J-L Liu G-J Wilkens RM Effective duration of pesticide-induced susceptibility of riceto brown planthopper (Nilaparvata lugens Staringl Homoptera Delphacidae) and physiological and biochemical changes in riceplants following pesticide application Int J Pest Manag 2004 50 55ndash62 [CrossRef]
19 Wang A-H Wu J-C Yu Y-S Liu J-L Yue J-F Wang MY Selective insecticide-induced stimulation on fecundity andbiochemical changes in Tryporyza incertulas (Lepidoptera Pyralidae) J Econ Entomol 2005 98 1144ndash1149 [CrossRef] [PubMed]
Crops 2021 1 182
20 Azzam S Wang F Wu J-C Shen J Wang L-P Yang G-Q Guo Y-R Comparisons of stimulatory effects of a seriesof concentrations of four insecticides on reproduction in the rice brown planthopper Nilaparvata lugens Staringl (HomopteraDelphacidae) Int J Pest Manag 2009 55 347ndash358 [CrossRef]
21 Ge L-Q Hu J-H Wu J-C Yang G-Q Gu H Insecticide-induced changes in protein RNA and DNA contents in ovary andfat body of female Nilaparvata lugens (Hemiptera Delphacidae) J Econ Entomol 2009 102 1506ndash1514 [CrossRef]
22 Ling S Zhang J Hu L Zhang R Effect of fipronil on the reproduction feeding and relative fitness of brown planthopperNilaparvata lugens Appl Entomol Zool 2009 44 543ndash548 [CrossRef]
23 Ge L-Q Wang L-P Zhao K-F Wu J-C Huang L-J Mating pair combinations of insecticide-treated male and femaleNilaparvata lugens Staringl (Hemiptera Delphacidae) planthoppers influence protein content in the male accessory glands (MAGs)and vitellin content in both fat bodies and ovaries of adult females Pestic Biochem Physiol 2010 98 279ndash288 [CrossRef]
24 Ge L-Q Wu J-C Zhao K-F Chen Y Yang G-Q Induction of Nlvg and suppression of Nljhe gene expression in Nilaparvatalugens (Staringl) (Hemiptera Delphacidae) adult females and males exposed to two insecticides Pestic Biochem Physiol 201098 269ndash278 [CrossRef]
25 Hu J-H Wu J-C Yin J-L Gu H-N Physiology of insecticide-induced stimulation of reproduction in the rice brownplanthopper (Nilaparvata lugens (Staringl)) Dynamics of protein in fat body and ovary Int J Pest Manag 2010 56 23ndash30 [CrossRef]
26 Wang L-P Shen J Ge L-Q Wu J-C Yang G-Q Jahn GC Insecticide-induced increase in the protein content of maleaccessory glands and its effect on the fecundity of females in the brown planthopper Nilaparvata lugens Staringl (HemipteraDelphacidae) Crop Prot 2010 29 1280ndash1285 [CrossRef]
27 Wu J Ge L Liu F Song Q Stanley D Pesticide-induced planthopper population resurgence in rice cropping systems AnnuRev Entomol 2020 65 409ndash429 [CrossRef]
28 Suri KS Singh G Insecticide-induced resurgence of the whitebacked planthopper Sogatella furcifera (Horvath) (HemipteraDelphacidae) on rice varieties with different levels of resistance Crop Prot 2011 30 118ndash124 [CrossRef]
29 Fabellar LT Heinrichs EA Toxicity of insecticides to predators of rice brown planthoppers Nilaparvata lugens (Staringl) (HomopteraDelphacidae) Environ Entomol 1984 13 832ndash837 [CrossRef]
30 Fabellar LT Heinrichs EA Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators Crop Prot1986 5 254ndash258 [CrossRef]
31 Cuong NL Ben PT Phuong LT Chau LM Cohen MB Effect of host plant resistance and insecticide on brown planthopperNilaparvata lugens (Staringl) and predator population development in the Mekong Delta Vietnam Crop Prot 1997 16 707ndash715[CrossRef]
32 Preetha G Stanley J Suresh S Samiyappan R Risk assessment of insecticides used in rice on mirid bug Cyrtorhinuslividipennis Reuter the important predator of brown planthopper Nilaparvata lugens (Stal) Chemosphere 2010 80 498ndash503[CrossRef] [PubMed]
33 Tanaka K Endo S Kazano H Toxicity of insecticides to predators of rice planthoppers Spiders the mirid bug and the dryinidwasp Appl Entomol Zool 2000 35 177ndash187 [CrossRef]
34 Zhang X Xu Q Lu W Liu F Sublethal effects of four synthetic insecticides on the generalist predator Cyrtorhinus lividipennisJ Pest Sci 2015 88 383ndash392 [CrossRef]
35 Settle WH Ariawan H Astuti ET Cahyana W Hakim AL Hindayana D Lestari AS Managing tropical rice peststhrough conservation of generalist natural enemies and alternative prey Ecology 1996 77 1975ndash1988 [CrossRef]
36 Salim M Heinrichs EA Insecticide-induced changes in the levels of resistance of rice cultivars to the whitebacked planthopperSogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) Crop Prot 1987 6 28ndash32 [CrossRef]
37 Yin J-L Xu H-W Wu J-C Hu J-H Yang G-Q Cultivar and insecticide applications affect the physiological development ofthe brown planthopper Nilaparvata lugens (Staringl) (Hemiptera Delphacidae) Environ Entomol 2014 37 206ndash212 [CrossRef]
38 Cheng C Chang W Studies on varietal resistance to the brown planthopper in Taiwan In Brown Planthopper Threat toRice Production in Asia International Rice Research Institute Ed International Rice Research Institute Manila Philippines1979 pp 251ndash271
39 Reissig WH Heinrichs EA Valencia SL Insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on ricevarieties with different levels of resistance Environ Entomol 1982 11 165ndash168 [CrossRef]
40 Heinrichs EA Fabellar LT Basilio RP Wen T-C Medrano F Susceptibility of rice planthoppers Nilaparvata lugens andSogatella furcifera (Homoptera Delphacidae) to insecticides as influenced by level of resistance in the host plant Environ Entomol1984 13 455ndash458 [CrossRef]
41 Joshi R Shepard B Kenmore P Lydia R Insecticide-induced resurgence of brown planthopper (BPH) on IR62 Int Rice ResNewsl 1992 17 9ndash10
42 Zhu Z-R Cheng J Jiang M-X Zhang X-X Complex influence of rice variety fertilization timing and insecticide onpopulation dynamics of Sogatella furcifera (Horvath) Nilaparvata lugens (Staringl) (Homoptera Delphacidae) and their natural enemiesin rice in Hangzhou China J Pest Sci 2004 77 65ndash74 [CrossRef]
43 Nanthakumar M Jhansi Lakshmi V Shashi Bhushan V Balachandran SM Mohan M Decrease of rice plant resistance andinduction of hormesis and carboxylesterase titre in brown planthopper Nilaparvata lugens (Staringl) by xenobiotics Pestic BiochemPhysiol 2012 102 146ndash152 [CrossRef]
Crops 2021 1 183
44 Yao H-W Jiang C-Y Ye G-Y Cheng J-A Studies on cultivar-induced changes in insecticide susceptibility and its relatedenzyme activities of the white-backed planthopper Sogatella furcifera (Horvath)(Homopter Delphacidae) Insect Sci 2002 9 9ndash15
45 Horgan FG Srinivasan TS Bentur JS Kumar R Bhanu KV Sarao PS Van Chien H Almazan MLP Bernal CCRamal AF et al Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers Implicationsfor rice breeding and gene deployment Agronomy 2017 7 62 [CrossRef]
46 Jairin J Phengrat K Teangdeerith S Vanavichit A Toojinda T Mapping of a broad-spectrum brown planthopper resistancegene Bph3 on rice chromosome 6 Mol Breed 2007 19 35ndash44 [CrossRef]
47 Ren J Gao F Wu X Lu X Zeng L Lv J Su X Luo H Ren G Bph32 a novel gene encoding an unknown SCRdomain-containing protein confers resistance against the brown planthopper in rice Sci Rep 2016 6 37645 [CrossRef]
48 Nguyen CD Zheng S-H Sanada-Morimura S Matsumura M Yasui H Fujita D Substitution mapping and characterizationof brown planthopper resistance genes from indica rice variety lsquoPTB33prime (Oryza sativa L) Breed Sci 2021 in press [CrossRef]
49 Chelliah S Heinrichs EA Factors affecting insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on RiceEnviron Entomol 1980 9 773ndash777 [CrossRef]
50 Moore MT Kroumlger R Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growthArch Environ Contam Toxicol 2010 59 574ndash581 [CrossRef]
51 Kim J Hwang I Han M Jang B Effect of insecticide and fungicide on phytotoxicity of herbicide in rice Korean J Weed Sci1986 6 67ndash75
52 Horgan FG Garcia CPF Haverkort F de Jong PW Ferrater JB Changes in insecticide resistance and host range performanceof planthoppers artificially selected to feed on resistant rice Crop Prot 2020 127 104963 [CrossRef] [PubMed]
53 Khush GS Virk PS IR Varieties and Their Impact International Rice Research Institute Manila Philippines 200554 Srinivasan TS Almazan MLP Bernal CC Ramal AF Subbarayalu MK Horgan FG Interactions between nymphs of
Nilaparvata lugens and Sogatella furcifera (Hemiptera Delphacidae) on resistant and susceptible rice varieties Appl Entomol Zool2016 51 81ndash90 [CrossRef]
55 Pentildealver Cruz A Arida A Heong KL Horgan FG Aspects of brown planthopper adaptation to resistant rice varieties withthe Bph3 gene Entomol Exp Appl 2011 141 245ndash257 [CrossRef]
56 Alam SN Cohen MB Detection and analysis of QTLs for resistance to the brown planthopper Nilaparvata lugens in adoubled-haploid rice population Theor Appl Genet 1998 97 1370ndash1379 [CrossRef]
57 Mackill DJ Khush GS IR64 A high-quality and high-yielding mega variety Rice 2018 11 18 [CrossRef] [PubMed]58 Heinrichs EA Basilio RP Valencia SL Buprofezin a selective insecticide for the management of rice planthoppers (Ho-
moptera Delphacidae) and leafhoppers (Homoptera Cicadellidae) Environ Entomol 1984 13 515ndash521 [CrossRef]59 Satapathy MK Anjaneyulu A Use of cypermethrin a synthetic pyrethroid in the control of rice tungro virus disease and its
vector Trop Pest Manag 1984 30 170ndash178 [CrossRef]60 Vorley W Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown
planthopper (BPH) in Kedah Malaysia Int Rice Res Newsl 1987 10 19ndash2061 Uddin A Islam KS Jahan M Ara A Khan MAI Factor influencing the resurgence of brown planthopper in Bangladesh
SAARC J Agric 2020 18 117ndash128 [CrossRef]62 Escalada MM Luecha M Heong KL Social impacts of planthopper outbreaks Case study from Thailand In Rice Planthoppers
Ecology Management Socio Economics and Policy Heong KL Cheng J Escalada MM Eds Zhejiang University PressHangzhou China 2015 pp 209ndash226
63 Zhang CP He HM Yu JZ Hu XQ Zhu YH Wang Q Residues of carbosulfan and its metabolites carbofuran and3-hydroxy carbofuran in rice field ecosystem in China J Environ Sci Health 2016 51 351ndash357 [CrossRef] [PubMed]
64 Samanta S Barman M Samanta NRA Bio-efficacy trials of carbofuran 3 CG against insect pests of rice J Entomol ZoolStud 2020 8 1529ndash1533
65 Donley N The USA lags behind other agricultural nations in banning harmful pesticides Environ Health 2019 18 44 [CrossRef][PubMed]
66 Adams A Gore J Musser F Cook D Catchot A Walker T Dobbins C Efficacy of selected insecticides applied to hybridrice seed J Econ Entomol 2015 109 200ndash206 [CrossRef]
67 Kumar R Singh B Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) Field Bull Environ Contam Toxicol2013 90 482ndash488 [CrossRef]
68 Dhaka S Prajapati C Singh D Singh R Field evaluation of insecticides and bio-pesticides against rice leaf folder CnaphalocrocisMedinalis (Guenee) Ann Plant Prot Sci 2011 19 324ndash326
69 Matsumura M Takeuchi H Satoh M Sanada-Morimura S Otuka A Watanabe T Van Thanh D Species-specific insecticideresistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in East and South-eastAsia Pest Manag Sci 2008 64 1115ndash1121 [CrossRef] [PubMed]
70 Tang J Li J Shao Y Yang B Liu Z Fipronil resistance in the whitebacked planthopper (Sogatella furcifera) Possible resistancemechanisms and cross-resistance Pest Manag Sci 2010 66 121ndash125 [CrossRef]
71 Horgan FG Arida A Ardestani G Almazan MLP Temperature-dependent oviposition and nymph performance revealdistinct thermal niches of coexisting planthoppers with similar thresholds for development PLoS ONE 2020 15 e0235506[CrossRef] [PubMed]
Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]

Crops 2021 1 170
of 30 times 25 times 50 cm (H timesW times L) (ie two plants per cage) This produced 14 treatmenttimes variety combinations (7 insecticides times 2 varieties) Each combination of plant pairswas replicated three times with cages positioned randomly on greenhouse benches andreplicates of each combination pair placed in separate greenhouse compartments Fivegravid females were placed between the two pots in each cage and allowed to move withinthe cage and lay eggs After 3 days all adults were manually removed from the cages Theplants were collected 4 days after removal of the adults to allow the eggs to develop Plantswere placed in moist tissue paper and stored in a refrigerator until clusters and eggs werecounted under a stereomicroscope (as described above) For both bioassays the replicateswere each placed in separate greenhouse compartments (ie 5 or 3 blocks)
25 Effects of HPR and Insecticides on BPH Fitness
We conducted two BPH antibiosis tests in the greenhouse We examined egg layingby gravid females on each variety and for each insecticide treatment using an ovipositiontest At 30 DAS treated and control plants were individually caged by placing an acetatetube 45 times 5 cm (H times Diameter) with a mesh top over each pot A recently mated gravidBPH was placed inside each cage through a slit in the acetate and was allowed to feed andoviposit After 3 days the females were manually removed from the cages Plants were leftfor 4 days to allow eggs to develop The plants were then collected and dissected under astereo microscope to count the clusters and eggs (as described above)
We examined the effects of host resistance and insecticide treatments on BPH nymphsin a survival test At 30 DAS treated and control plants were individually caged asdescribed above Recently emerged nymphs (le2 instar) were collected from the colonyusing a suction-pooter and placed inside the cages at a density of ten per cage through aslit in the acetate The nymphs were allowed to feed and develop for 15 days after whichthe number of survivors and their developmental stages were recorded For both bioassaysplants were dried in a forced draft oven at 60 C for 7 days and were weighed For thesurvival bioassay the nymphs were also dried and weighed to estimate dry biomass andthereby control for varying consumption of plant liquids Both bioassays were replicatedfive times (ie 8 treatments times 2 varieties times 5 replicates = 80 pots per bioassay) with eachreplicate placed in a separate greenhouse compartment (ie 5 blocks)
26 Effects of HPR and Insecticides in a Screenhouse Environment
The combined effects of insecticide treatments and HPR were investigated using riceplants grown in an insect-proof screenhouse The screenhouse consisted of six concretebays (dimensions 266times 25 m (LtimesW)) filled with flooded paddy soil Temperatures in thescreenhouse varied between 21 and 34 C and RH varied between 70 and 90 Rainwatercould pass through the screenhouse roof during the experiment 160 mm of precipitationwas recorded representing the Philippines dry season The screenhouse had no artificiallighting (ie DN = 12 h12 h)
In one of the bays translucent plastic barriers were erected to delimitate five exper-imental blocks of 150 times 300 m Each block was further divided into eight cubicles of075 m2 using the same plastic materials The barriers were supported by bamboo stakesand reached a height of 150 m and a depth into the paddy soil of approx 030 m After thebarriers were installed two 23 DAS seedlings each of IR62 and IR64 were randomly trans-planted to each cubicle at 25 times 25 cm spacing We used a standard fertilizer regime to sim-ulate farmersrsquo fields The seedlings were treated with ammonium sulphate at 100 kg haminus1
split equally into three applications (ie one basal and two top dressings) Solophos andmuriate of potash was also applied at 20 kg haminus1 as a basal application Plants were contin-uously watered and the cubicles were manually weeded until rice plants were harvestedCubicles in each of the five blocks received one of the eight treatments (seven insecticidesand a control) Treatments were randomly assigned to cubicles within blocks The insecti-cide applications were made at 30 and 60 DAS to simulate farmer practices under intensive
Crops 2021 1 171
rice production systems where farmers apply insecticides ge two times [14] however inour experiment we did not mix insecticides
Rice herbivores including BPH WBPH and GLH occurred at low densities in thescreenhouse Therefore at the opposite end of the screenhouse (separated by a distanceof 20 m) a bay with gt50 DAS TN1 rice plants was infested with high densities of BPH(approx 200 BPH plantminus1) This acted as a potential source of planthoppers to attackthe treated IR62 and IR64 plants in our experiment Natural enemies were largely absentfrom the screenhouse because of the insect-proof cladding (1 mm times 1 mm mesh bore)Microparasitoids (Oligosita spp and Anagrus spp) that could pass through the screenoccasionally attacked planthopper eggs in the screenhouse (estimated 5ndash10 of eggsparasitized unpublished data)
The IR62 and IR64 plants were continuously monitored to assess grain maturationWhen about 85 of the rice grain was mature any arthropods on the plants were collectedusing a vacuum sampler and stored in plastic vials The plants were then destructivelyharvested by carefully pulling the plant from the soil to maintain intact roots The plantroots were washed under running water and each plant was placed inside a paper bagThe plants and arthropods were then dried in a forced draft oven at 60 C for 7 days andweighed Arthropods were separated by species and counted The plants were separatedinto roots shoots and panicles and the numbers of reproductive and vegetative tillerswere recorded Rice grain was removed from the panicles and separated as filled andnon-filled grain The dried grain was counted and weighed
27 Data Analyses
The numbers of egg clusters and eggs laid in rice plants during the multichoice testwere analyzed using univariate general linear models (GLM) on ranked (within replicate)data Separate analyses were conducted for IR62 and IR64 plants Data from the binarychoice bioassays were analyzed using paired t-tests The numbers of egg clusters and eggslaid in treated and non-treated plants during the oviposition bioassay were compared usingunivariate GLM with plant biomass initially included as a covariate but later removedbecause it had no significant effect Cluster and egg numbers were ranked within blocksbefore analyses Average cluster size was estimated by dividing the number of eggs in eachplant by the number of clusters Cluster size was analyzed using univariate GLM removingthe block effect Similarly the proportions of surviving nymphs and nymph biomass fromthe survival bioassay were analyzed using univariate GLMs initially including plantbiomass as a covariate Data from the survival bioassay were ranked before analysesToo few nymphs survived under some treatments to examine development rates or thesex of surviving adults We estimated the combined effects of HPR and each insecticidelsquoxrsquo as -((fitness on untreated IR64 minus fitness on x-treated IR62)fitness on untreated IR64)To estimate the effect of the insecticide alone we removed the effect on HPR which weestimated as fitness on untreated IR62 minus fitness on untreated IR64 Fitness was assessedfrom the oviposition bioassay as the numbers of eggs laid and from the survival bioassayas nymph biomass Where nymph mortality on insecticide-treated IR64 plants approached100 HPR effects were regarded as insignificant
We also examined combined and insecticide effects from the screenhouse data basedon densities of BPH and WBPH (see below) Effects were calculated using insecticide andHPR combinations for each replicated block (N = 5) Estimated effects were comparedacross treatments using univariate GLMs Because three herbivores (BPH WBPH andGLH) occurred in our screenhouse plots we used a split-plot multivariate GLM to analyzeherbivore densities Total planthopper and leafhopper biomass and density and plantgrowth parameters (tiller number biomass yield grain number proportion of grainunfilled and grain size) were analyzed using split-plot univariate GLMs The covariateplant biomass was initially included in analyses of the effects on herbivores but had noeffect and was subsequently removed Similarly the covariate herbivore biomass wasinitially included in analyses of plant growth parameters but had no effect and was
Crops 2021 1 172
subsequently removed The combined effects of insecticide and HPR and the separateeffects of each insecticide (calculated as indicated above) on BPH and WBPH were alsoanalyzed using univariate GLM
Post-hoc Tukey tests were applied to all significant treatment factors Residuals wereplotted following all parametric analyses and were found to be normal and homogenous
3 Results31 Effects of HPR and Insecticides on BPH Preferences
Treatments affected the number of clusters and eggs that were laid in IR62 plants(ranked clusters and eggs F732 = 4738 p lt 0001) and in IR64 plants (clusters F732 = 6229p lt 0001 eggs F732 = 7757 p lt 0001) (Figure 1AB) Fewer eggs were laid on IR64 plantstreated with the pyrethroids (cypermethrin and deltamethrin) fipronil and thiamethoxam-chlorantraniliprole compared to plants treated with buprofezin carbofuran and cartaphydrochloride (Figure 1B) Cypermethrin-treated IR62 plants also had fewer eggs thanbuprofezin-treated IR62 plants (Figure 1A) Although BPH females tended to lay fewereggs in many of the insecticide-treated rice plants in the binary choice experiments therewere no statistically significant differences between the numbers of eggs laid in treated-IR62(t ranges from minus0746 to 3186 p gt 005) or treated-IR64 (t ranges from minus0108 to 2789p gt 005) compared to corresponding non-treated plants (Figure 1CD)
Crops 2021 1 FOR PEER REVIEW 7
herbivore densities Total planthopper and leafhopper biomass and density and plant growth parameters (tiller number biomass yield grain number proportion of grain un-filled and grain size) were analyzed using split-plot univariate GLMs The covariate plant biomass was initially included in analyses of the effects on herbivores but had no effect and was subsequently removed Similarly the covariate herbivore biomass was initially included in analyses of plant growth parameters but had no effect and was subsequently removed The combined effects of insecticide and HPR and the separate effects of each insecticide (calculated as indicated above) on BPH and WBPH were also analyzed using univariate GLM
Post-hoc Tukey tests were applied to all significant treatment factors Residuals were plotted following all parametric analyses and were found to be normal and homogenous
3 Results 31 Effects of HPR and Insecticides on BPH Preferences
Treatments affected the number of clusters and eggs that were laid in IR62 plants (ranked clusters and eggs F732 = 4738 p lt 0001) and in IR64 plants (clusters F732 = 6229 p lt 0001 eggs F732 = 7757 p lt 0001) (Figure 1AB) Fewer eggs were laid on IR64 plants treated with the pyrethroids (cypermethrin and deltamethrin) fipronil and thiameth-oxam-chlorantraniliprole compared to plants treated with buprofezin carbofuran and cartap hydrochloride (Figure 1B) Cypermethrin-treated IR62 plants also had fewer eggs than buprofezin-treated IR62 plants (Figure 1A) Although BPH females tended to lay fewer eggs in many of the insecticide-treated rice plants in the binary choice experiments there were no statistically significant differences between the numbers of eggs laid in treated-IR62 (t ranges from minus0746 to 3186 p gt 005) or treated-IR64 (t ranges from minus0108 to 2789 p gt 005) compared to corresponding non-treated plants (Figure 1CD)
Figure 1 Results from multichoice (AB) and binary choice (CD) bioassays conducted in the green-house Bioassays were conducted separately for IR62 (AC) and IR64 (BD) Plants were treated with one of seven insecticides (A to G) as identified in the legend Control plants were sprayed with water Error bars are presented (N = 5 for (A) and (B) N = 3 for (C) and (D)) Lowercase letters (andashc) indicate homogenous treatment groups (Tukey p lt 005)
A B C D E F G0
100
200
300
400
Treatment
A B C D E F G
Non treatedTreated
A B C D E F G Control
Num
ber o
f egg
s pe
r pla
nt
0
100
200
300
400
A B C D E F G Control
IR62 IR64
A
C
B
D
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliproleH Control
a abab
c
abc
abc bc bc
c
aaa
ab
ab
bb b
Figure 1 Results from multichoice (AB) and binary choice (CD) bioassays conducted in thegreenhouse Bioassays were conducted separately for IR62 (AC) and IR64 (BD) Plants were treatedwith one of seven insecticides (A to G) as identified in the legend Control plants were sprayed withwater Error bars are presented (N = 5 for (AB) N = 3 for (CD)) Lowercase letters (andashc) indicatehomogenous treatment groups (Tukey p lt 005)
32 Effects of HPR and Insecticides on BPH Fitness
More eggs were laid on IR64 plants than on IR62 plants but BPH produced similarnumbers of egg clusters on both plants The lower numbers of eggs in IR62 were due tosmaller clusters on the variety (Table 1) The numbers of eggs and clusters were affectedby insecticide treatment with fewer eggs and clusters on cypermethrin-treated fipronil-treated and thiamethoxam-chlorantraniliprole-treated plants than on plants under the
Crops 2021 1 173
remaining treatments (Table 1) The treatment-effects were not affected by host variety (ieinteractions were non-significant Table 1) Insecticide treatments had no effect on clustersize in the bioassay (Table 1) There was no significant effect of combined insecticide andHPR (F628 = 0449 p = 0840) on observed changes in egg laying during the bioassay butthe estimated insecticide effect was greater for thiamethoxam-chlorantraniliprole fiproniland cypermethrin than for buprofezin cartap hydrochloride and carbofuran (F628 = 3140p = 0018 Figure 2A)
Table 1 Results from greenhouse oviposition and survival tests with IR62 (resistant) and IR64 (susceptible) each treatedwith one of seven insecticides Numbers are means plusmn SEM (N = 5)
Treatments Egg Clusters(Number Plantminus1) 12
Eggs(Number Plantminus1) 12
Cluster Size(Number of Eggs) 1
Nymph Survival(Proportions) 12
Nymph Biomass (mgDry Weight Plantminus1) 12
IR62Buprofezin 1240 plusmn 140 b 5040 plusmn 904 b 393 plusmn 036 000 plusmn 000 a 000 plusmn 000 a
Carbofuran 1300 plusmn 089 b 5380 plusmn 421 b 417 plusmn 026 064 plusmn 064 d 154 plusmn 154 d
Cartaphydrochloride 1360 plusmn 075 b 5200 plusmn 486 b 382 plusmn 029 028 plusmn 028 b 151 plusmn 151 b
Cypermethrin 620 plusmn 169 a 2000 plusmn 608 a 316 plusmn 023 002 plusmn 002 a 002 plusmn 002 a
Deltamethrin 1360 plusmn 160 b 4340 plusmn 566 b 338 plusmn 069 040 plusmn 040 c 120 plusmn 120 c
Fipronil 520 plusmn 146 a 2240 plusmn 735 a 393 plusmn 064 000 plusmn 000 a 000 plusmn 000 a
Thiamethoxam-chlorantraniliprole 380 plusmn 086 a 1700 plusmn 435 a 447 plusmn 032 000 plusmn 000 a 000 plusmn 000 a
Control 1260 plusmn 103 b 4900 plusmn 716 b 388 plusmn 054 080 plusmn 080 d 292 plusmn 292 d
IR64Buprofezin 2100 plusmn 164 10940 plusmn 1346 556 plusmn 117 000 plusmn 000 000 plusmn 000Carbofuran 1820 plusmn 404 9880 plusmn 2939 524 plusmn 074 086 plusmn 086 420 plusmn 420
Cartaphydrochloride 2140 plusmn 619 11460 plusmn 3141 549 plusmn 064 010 plusmn 010 045 plusmn 045
Cypermethrin 380 plusmn 233 1980 plusmn 1155 550 plusmn 058 000 plusmn 000 000 plusmn 000Deltamethrin 1560 plusmn 209 7500 plusmn 1581 473 plusmn 069 040 plusmn 040 201 plusmn 201
Fipronil 380 plusmn 086 2100 plusmn 552 546 plusmn 080 000 plusmn 000 000 plusmn 000Thiamethoxam-chlorantraniliprole 260 plusmn 075 800 plusmn 212 373 plusmn 133 000 plusmn 000 000 plusmn 000
Control 1740 plusmn 364 7520 plusmn 2344 426 plusmn 073 086 plusmn 086 360 plusmn 360F-treatment 3 19898 19898 0541 ns 133811 108451
F-variety 3 2596 ns 7607 7471 1080 ns 0008 ns
F-interaction 3 1840 ns 2034 ns 0610 ns 4902 9496 1 Lowercase letters (andashd) indicate homogenous treatment groups (for IR62 and IR64 combined) 2 Data were ranked within blocks beforeanalysis 3 DF = 164 for varieties 764 for treatments and interactions ns = p gt 005 = p le 001 = p le 0001
Nymph survival and biomass were not affected by rice variety (Table 1) Buprofezinfipronil thiamethoxam-chlorantraniliprole and cypermethrin reduced survival to 0 in thebioassay and all insecticides except carbofuran resulted in lower survival and biomass thanon untreated plants (Table 1) There were significant interactions because of higher nymphmortality and a resulting lower biomass on carbofuran-treated IR62 compared to untreatedIR62 but lower mortality and a higher biomass of nymphs on carbofuran-treated IR64than on untreated IR64 (Table 1) There was no significant effect of insecticide treatment(F628 = 0946 p = 0479) on observed changes in nymph biomass during the bioassaybut the estimated combined effects of pesticide and HPR was greater for thiamethoxam-chlorantraniliprole fipronil cypermethrin and buprofezin than for cartap hydrochlorideand carbofuran (F628 = 5699 p = 0001) (Figure 2B)
Crops 2021 1 174
Crops 2021 1 FOR PEER REVIEW 9
Figure 2 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) egg laying and (B) nymph biomass in greenhouse bioas-says Effects are indicated as proportional changes in fitness parameters compared to untreated IR64 Negative numbers indicate reduced fitness Standard errors are indicated (N = 5) Lowercase letters (ab) indicated homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 1)
Nymph survival and biomass were not affected by rice variety (Table 1) Buprofezin fipronil thiamethoxam-chlorantraniliprole and cypermethrin reduced survival to 0 in the bioassay and all insecticides except carbofuran resulted in lower survival and biomass than on untreated plants (Table 1) There were significant interactions because of higher nymph mortality and a resulting lower biomass on carbofuran-treated IR62 compared to untreated IR62 but lower mortality and a higher biomass of nymphs on carbofuran-treated IR64 than on untreated IR64 (Table 1) There was no significant effect of insecticide treatment (F628 = 0946 p = 0479) on observed changes in nymph biomass during the bio-assay but the estimated combined effects of pesticide and HPR was greater for thiameth-oxam-chlorantraniliprole fipronil cypermethrin and buprofezin than for cartap hydro-chloride and carbofuran (F628 = 5699 p = 0001) (Figure 2B)
33 Effects of HPR and Insecticides on Planthoppers and Leafhoppers Plants in the screenhouse were colonized by BPH WBPH and GLH Densities for
each species were similar on IR62 and IR64 (Wilkrsquos λ = 0994 p = 0952) There was a sig-nificant treatment effect on all three species (Wilkrsquos λ = 0586 p = 0049) because of high densities on deltamethrin-treated plants (Table 2) The numbers and biomass of all three species combined were also greater on deltamethrin-treated plants than on the remaining treatments (Table 2) Host resistance reduced GLH densities under the deltamethrin treat-ment but had no apparent effect on densities under the other treatments (Table 2) There were significant combined effects of HPR and insecticide on the numbers of BPH (F624 = 2686 p = 0039) and WBPH (F624 = 2482 p = 0052) this was almost exclusively due to the resurgence effect of deltamethrin on the planthoppers (BPH F624 = 2686 p = 0039 WBPH F624 = 2482 p = 0052) (Figure 3)
Δ eg
gs(P
ropo
rtion
)
-10
-08
-06
-04
-02
00
02
04
06
Treatment
A B C D E F G
Δ N
ymph
bio
mas
s(P
ropo
rtion
)
-10
-08
-06
-04
-02
00
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
ab
a a
b
a
b b
a a a a a a a a
b b bb
bb
Figure 2 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone(P grey bars) on (A) egg laying and (B) nymph biomass in greenhouse bioassays Effects are indicated as proportionalchanges in fitness parameters compared to untreated IR64 Negative numbers indicate reduced fitness Standard errors areindicated (N = 5) Lowercase letters (ab) indicated homogenous treatment groups based on Tukey pairwise comparisons(p lt 005) (see also Table 1)
33 Effects of HPR and Insecticides on Planthoppers and Leafhoppers
Plants in the screenhouse were colonized by BPH WBPH and GLH Densities foreach species were similar on IR62 and IR64 (Wilkrsquos λ = 0994 p = 0952) There was asignificant treatment effect on all three species (Wilkrsquos λ = 0586 p = 0049) because of highdensities on deltamethrin-treated plants (Table 2) The numbers and biomass of all threespecies combined were also greater on deltamethrin-treated plants than on the remainingtreatments (Table 2) Host resistance reduced GLH densities under the deltamethrintreatment but had no apparent effect on densities under the other treatments (Table 2)There were significant combined effects of HPR and insecticide on the numbers of BPH(F624 = 2686 p = 0039) and WBPH (F624 = 2482 p = 0052) this was almost exclusively dueto the resurgence effect of deltamethrin on the planthoppers (BPH F624 = 2686 p = 0039WBPH F624 = 2482 p = 0052) (Figure 3)
Crops 2021 1 175
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants underseven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density(Number Plantminus1) 1
WBPH Density(Number Plantminus1) 1
GLH Density(Number Plantminus1) 1
Density ofPlanthoppers and
Leafhoppers(Number Plantminus1) 1
Biomass ofPlanthoppers and
Leafhoppers(mg Dry Weight) 1
IR62Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a
Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartaphydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a
Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a
Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a
Thiamethoxam-chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a
IR64Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartaphydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123
Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns
F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Crops 2021 1 FOR PEER REVIEW 10
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants under seven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density (Num-
ber Plantminus1) 1 WBPH Density
(Number Plantminus1) 1 GLH Density
(Number Plantminus1) 1
Density of Planthoppers and Leafhoppers (Number
Plantminus1) 1
Biomass of Planthoppers and Leafhoppers (mg Dry
Weight) 1 IR62
Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartap hydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a Thiamethoxam-
chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a IR64
Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132 Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartap hydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123 Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113 Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052 Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087 F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns 1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatments and interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in the screenhouse Effects are indicated as proportional changes in planthopper densities compared to untreated IR64 Negative numbers indicate reduced densities positive numbers are increased den-sities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
Δ BP
H(N
umbe
r pla
nt-1
)
0
5
10
15
20
Treatment
A B C D E F G
Δ W
BPH
(Num
ber p
lant
-1)
0
20
30
40
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
a a
aa a a
a a a
a
a a
b
b
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimatedpesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in thescreenhouse Effects are indicated as proportional changes in planthopper densities compared to
Crops 2021 1 176
untreated IR64 Negative numbers indicate reduced densities positive numbers are increaseddensities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenoustreatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
34 Direct and Indirect Effects of Insecticides on Rice Development
IR62 plants had more tillers and attained a greater biomass than IR64 plants in thescreenhouse (Table 3) IR62 grain matured faster and plants were harvested earlier than theIR64 plants (Table 3) IR62 plants also produced more grain (number of grain) and hadhigher yields than IR64 but had smaller grain and more unfilled grain than IR64 (Table 3)
Table 3 Rice growth parameters for IR62 (resistant) and IR64 (susceptible) under seven insecticide treatments and forcontrol non-treated plants in a screenhouse environment Numbers are means plusmn SEM
Treatment Tiller Number 1 Plant Biomass(g Dry Weight) 1
Time to Harvest(Days) 1
Weightof Filled Grain(g Dry Weight) 1
Number ofFilled Grain
(1000s) 1
Unfilled Grain(Proportion) 1
1000 GrainWeight 1
IR62Buprofezin 2660 plusmn 206 ab 4643 plusmn 385 ab 7990 plusmn 168 ab 5776 plusmn 683 ab 361plusmn 061 ab 011 plusmn 003 a 2008 plusmn 042 ab
Carbofuran 2900 plusmn 106 ab 5546 plusmn 357 b 7720 plusmn 080 ab 7448 plusmn 691 b 446plusmn 061 ab 011 plusmn 001 a 2103 plusmn 054 b
Cartaphydrochloride 2820 plusmn 327 ab 4896 plusmn 470 ab 7920 plusmn 153 b 6430 plusmn 712 ab 415plusmn 064 ab 012 plusmn 001 b 2023 plusmn 065 a
Cypermethrin 2870 plusmn 301 a 4957 plusmn 562 a 7760 plusmn 103 b 6032 plusmn 822 a 410plusmn 067 ab 014 plusmn 002 a 1965 plusmn 076 a
Deltamethrin 2620 plusmn 345 a 4592 plusmn 588 a 7840 plusmn 108 ab 5698 plusmn 832 ab 379plusmn 047 ab 012 plusmn 001 a 2030 plusmn 063 ab
Fipronil 2380 plusmn 176 ab 4607 plusmn 657 ab 7700 plusmn 179 ab 5266 plusmn 390 a 318 plusmn 032 a 013 plusmn 002 a 2128 plusmn 047 b
Thiamethoxam-chlorantraniliprole 2650 plusmn 250 b 4547 plusmn 429 b 7720 plusmn 102 a 5777 plusmn 688 ab 408 plusmn 061 b 013 plusmn 002 a 2019 plusmn 067 ab
Control 3120 plusmn 247 b 5401 plusmn 555 b 7720 plusmn 080 ab 7045 plusmn 520 b 396plusmn 036 ab 012 plusmn 002 a 2076 plusmn 050 ab
IR64Buprofezin 2340 plusmn 250 5064 plusmn 747 8350 plusmn 124 5080 plusmn 952 270 plusmn 054 008 plusmn 001 2277 plusmn 039Carbofuran 2040 plusmn 097 4591 plusmn 141 8540 plusmn 073 4951 plusmn 312 263 plusmn 023 006 plusmn 000 2295 plusmn 036
Cartaphydrochloride 2120 plusmn 075 4336 plusmn 285 8520 plusmn 140 4273 plusmn 579 238 plusmn 026 015 plusmn 007 2050 plusmn 234
Cypermethrin 1990 plusmn 089 3968 plusmn 272 8710 plusmn 135 3920 plusmn 552 250 plusmn 028 008 plusmn 002 2171 plusmn 070Deltamethrin 1980 plusmn 181 4325 plusmn 357 8460 plusmn 068 4710 plusmn 496 277 plusmn 024 008 plusmn 001 2314 plusmn 012
Fipronil 2120 plusmn 174 4697 plusmn 311 8450 plusmn 089 4888 plusmn 392 274 plusmn 027 008 plusmn 001 2247 plusmn 042Thiamethoxam-chlorantraniliprole 2520 plusmn 287 5404 plusmn 465 8380 plusmn 196 5594 plusmn 546 347 plusmn 053 010 plusmn 001 2265 plusmn 072
Control 2160 plusmn 076 4678 plusmn 342 8600 plusmn 096 4940 plusmn 543 267 plusmn 029 008 plusmn 001 2283 plusmn 049F-treatment 4100 ns 3410 2253 ns 5150 2555 3923 4924
F-variety 168232 7665 547500 104356 118242 32308 106597 F-interaction 5980 6113 4824 5760 2921 2769 2552
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Insecticides affected plant growth directly and indirectly (through increased planthop-per densities) Plants treated with cypermethrin and deltamethrin had fewer tillers had alower biomass and under cypermethrin had lower yields than untreated plants (Table 3)Plants treated with fipronil also had reduced yields due to lower grain numbers comparedto untreated plants (Table 3) Although yields were not affected treatment with cartaphydrochloride increased the proportion of unfilled grain compared to untreated plantsof both varieties Cartap hydrochloride and cypermethrin also caused a delay in grainmaturation on IR62 and IR64 respectively relative to thiamethoxam-chlorantraniliproletreated plants (Table 3)
There were several significant interactions tiller numbers on both plants were similarunder buprofezin fipronil and thiamethoxam-chlorantraniliprole treatments but wereotherwise higher on IR62 (Table 3) biomass was lower on IR64 under carbofuran andcypermethrin treatments but was otherwise similar on both plants (Table 3) buprofezindelayed grain maturation on IR62 but maturation was otherwise faster on IR62 comparedto IR64 (Table 3) yields under buprofezin fipronil and thiamethoxam-chlorantraniliprolewere similar for IR62 and IR64 but otherwise yields were higher on IR62 (Table 3) grain
Crops 2021 1 177
numbers were higher on IR62 than IR64 except under fipronil treatments (Table 3) theproportions of grain unfilled were higher on IR62 but were similar between varieties underthe fipronil treatment (Table 3) 1000-grain weight was higher in IR64 but similar on bothvarieties when treated with cartap hydrochloride (Table 3)
4 Discussion
Our results using a highly resistant rice variety and some commonly used insecticidesindicate a range of possible effects on planthoppers and rice plants by combining these twomanagement practices
41 Synergies
The most predictable effect of combining HPR and insecticides is a synergy in reduc-ing BPH numbers This is suggested because planthoppers are weakened by the hostrsquosresistance and should therefore be more susceptible to the toxic effects of insecticides [40]Using spray towers over potted plants Heinrichs et al (1984) [40] examined the combinedeffects of a range of insecticides (that included carbofuran) with resistance by comparingBPH mortality on six resistant and two susceptible rice varieties Their results indicatedthat mortality was greatest when insecticides were applied to BPH on resistant varietiesThe effects were similar when the study was repeated with WBPH for three insecticides(including carbofuran) on the resistant variety N22 (Wbph1 gene) [40] In our study hostresistance reduced egg laying and nymph biomass on non-treated IR62 compared to non-treated IR64 Egg laying was also reduced on plants treated with cypermethrin fipronilor thiamethoxam-chlorantraniliprole (Table 1) Based on the results of our choice andno-choice oviposition bioassays cypermethrin acted mainly as a deterrent of gravid fe-males whereas fipronil and thiamethoxam-chlorantraniliprole directly reduced egg layingDespite similar BPH responses to both the insecticides and resistant host there were no ap-parent synergies between any of these insecticides and resistance in reducing egg numbersin the oviposition bioassay In contrast to these three insecticides our bioassays indicatedthat buprofezin carbofuran and cartap hydrochloride can lead to higher numbers of eggson treated plants compared to untreated plants (Table 1) These effects were not statisticallysignificant and may be exaggerated because bioassays were conducted over relativelyshort time scales For example planthoppers will sometimes respond to stresses such ashigh temperatures by increasing oviposition rates [7172] Furthermore the stimulation ofegg-laying by these chemicals was not apparent in our screenhouse experiment (Table 2)Nevertheless our results do indicate that host resistance reduced the magnitude of anystimulatory effect on oviposition egg numbers on buprofezin-treated carbofuran-treatedand cartap hydrochloride-treated IR64 in the no-choice bioassay were twice as high as onthe corresponding IR62 plants In cases such as these the resistance of IR62 functions as abuffer or lsquoinsurancersquo against possible undesired pesticide effects [1238]
Although buprofezin appeared to stimulate oviposition the growth hormone had astrong impact on nymph development leading to zero survival of exposed nymphs in ourgreenhouse bioassay Cypermethrin fipronil and thiamethoxam-chlorantraniliprole alsoreduced nymph survival to zero in the greenhouse bioassay (Table 1) Where insecticideshave such high efficiencies any fitness losses to BPH due to host resistance then becomeredundant Unlike buprofezin carbofuran also stimulated nymph biomass on IR64 in thesurvival bioassay (Table 1) however nymph biomass was lower on carbofuran-treated IR62than on non-treated IR62 indicating that the hostrsquos resistance buffered against undesiredstimulatory effects on BPH and may have functioned synergistically to augment mortalityand slow BPH growth Such buffering allows insecticides to be used against other pestswithout inducing secondary outbreaks of non-target pests In our screenhouse experimentherbivore densities were generally low on both IR62 and IR64 for most treatments (exceptdeltamethrin) and were below threshold densities for insecticide applicationsmdashwhichare normally about 25ndash50 planthoppers per hill [1373] Under such low densities it wasnot possible to observe the impact of host resistance on planthopper population build-
Crops 2021 1 178
up There was some indication of higher densities on cartap hydrochloride-treated IR62in the screenhouse which may be related to the stimulatory effects of the product onegg laying and the low mortality of nymphs on cartap hydrochloride-treated plants asobserved in the greenhouse bioassays (Table 1) Field studies from the Philippines haveindicated that IR62 effectively reduces BPH densities during wet and dry seasons [4]However farmers are rarely aware of which anti-herbivore resistance their rice varietiespossess and frequently apply pesticides regardless of resistance thereby reducing theirfarm profits [3841] For example Joshi et al (1992) [41] reported widespread outbreaks ofBPH on IR62 in Mindanao Philippines after applications of decamethrin (a pyrethroid) tothe resistant variety Where such resurgence-causing insecticides are applied resistanceseems to be often incapable of reducing the stimulatory effects of the insecticide and itloses its capacity to buffer against outbreaks
42 Antagonisms
Several insecticides are associated with increasing densities of planthoppers andwith resurgence outbreaks in farmersrsquo fields [27293041496174] Indeed patches oflsquohopperburnrsquo (ie dead rice plants resulting from planthopper damage) are often anindicator of high insecticide use in tropical Asia [214] In many cases these resurgence-causing insecticides were once effective in reducing planthopper densities but over timepopulations developed resistance against the products The resulting low toxicity canthen have a stimulatory effect on planthopper feeding and development [2123242743]This may be through hormesis which is a direct response by the planthopper to low-levelexposure to the toxic chemical or it may result from physiological changes to the rice plantsthat ultimately benefit the planthopper [1849] In our screenhouse study we induced aresurgence outbreak of BPH WBPH and to a lesser extent GLH on deltamethrin-treatedrice plants (Table 2) The combined densities of all three herbivores increased almost10 times on the treated plants Furthermore herbivore biomass increased by between fourand six times Although the resurgence effects of deltamethrin have been documentedin previous studies [2029ndash313975] we could not predict from our greenhouse studiesthat any such outbreak would occur Egg laying was unaffected by deltamethrin in ourchoice and no-choice experiments and nymph survival and development both declinedon deltamethrin-treated rice in the survival bioassay (Table 1) The massive increases inplanthopper densities on deltamethrin-treated plants in our study seem to result froma heightened attraction of adult BPH and WBPH to the plants [49] particularly sincenumbers were so low on all other plants including both treated and non-treated plantsHowever once on the plants planthoppers also apparently had enhanced fitness Cuonget al (1997) [31] observed higher population densities of BPH on deltamethrin-treatedrice in Vietnam albeit with lower-level increases on resistant varieties Furthermore in agreenhouse study Azzam et al (2009) [20] found that deltamethrin reduced pre-ovipositiontimes increased the duration of oviposition and increased egg hatchability and femalelongevity Nanthakumar et al (2012) [43] further showed that deltamethrin increased BPHfeeding efficiency on resistant and susceptible rice plants as determined through honeydewproduction and population size In studies with WBPH Salim and Heinrichs (1987) [36]showed deltamethrin to increase settling (antixenosis) feeding and population growthand Suri and Singh (2011) [28] showed deltamethrin to increase WBPH reproductive ratesand nymph survival In our study the observed resurgence of planthoppers was similaron both varieties indicating as was previously noted by Joshi et al (1992) [41] thatresurgence-causing pyrethroids are antagonistic to resistance in IR62
Though not statistically significant cartap hydrochloride also increased BPH numberson IR62 but not on IR64 in our screenhouse study This indicates that the product mayalso be antagonistic to the hostrsquos resistance mechanisms The stronger effect on IR62 maybe related to the differences between IR62 biomass growth rates and anatomy (ie alarger plant with higher tillering) compared to IR64 (Table 3) Once resistance is no longereffective because of chemically-enhanced BPH fitness these plant characteristics which
Crops 2021 1 179
have been related to observed preferences in BPH and other sap-sucking insects [349]could further accelerate the build-up of BPH populations on cartap hydrochloride-treatedplants To further test this idea experiments could be conducted using near-isogenic ricelines that differ mainly in the presence or absence of tagged resistance genes [76] Suchtests would avoid confounding differences in plant anatomy between test varieties IR62resistance to BPH has been well studied [4] The variety is also resistant to WBPH atolder plant stages [54] Therefore in our study deltamethrin was also antagonistic tothis WBPH-resistance (Table 2) However BPH feeding also facilitates WBPH populationgrowth [5477] and there may have been added indirect plant-mediated effects [77] thataugmented WBPH populations on both varieties in our experiment IR62 is also resistant toGLH [45] In our experiments GLH densities also increased on deltamethrin-treated plantsbut the effect was only apparent on IR64 Therefore GLH-resistance in IR62 apparentlybuffered against deltamethrin-associated increases in GLH on the variety as was alsonoted from field studies in Vietnam [31] Deltamethrin is currently not recommended foruse in rice [202878] however the product continues to be marketed in rice-dominatedregions throughout Asia [131479] Although cypermethrin has been associated with BPHoutbreaks and was consequently removed for use on rice in Thailand and Vietnam [62]we did not observe cypermethrin-induced resurgence in our screenhouse experimentNevertheless cypermethrin has been associated with resurgence of BPH in South Asia [61]and we have noted stimulatory effects of cypermethrin on other rice herbivores in thePhilippines (Horgan unpublished data)
43 Phytotoxicity
Phytotoxicity is relatively common following applications of insecticides [182850]Phytotoxic effects may change a plantrsquos physiology without consequences for crop yieldsHowever such changes to plant physiology could have indirect effects on herbivores Forexample deltamethrin methyl parathion and quinalphos produce increases in total sugarsand reducing sugars as well as increasing the protein contents of the rice leaf sheath andleaf blade [28] Several insecticides also reduce plant phenols in ricemdashwhich increases therice plantrsquos susceptibility to BPH and other insects [27] In our screenhouse experiment wenoted several changes to rice anatomy and yields Importantly we noted that cypermethrinand fipronil both reduced rice yields (Table 3) This was not due to indirect herbivoreeffects because planthopper and leafhopper densities were consistently low on rice treatedwith these insecticides The effects therefore probably resulted from direct phytotoxiceffects on the plants Furthermore these effects were most pronounced on treated IR62plants Other minor effects on grain production included increased proportions of unfilledgrain in cartap hydrochloride-treated plants and delays in grain maturation on cartaphydrochloride-treated and cypermethrin-treated plants (Table 3) Despite the high densitiesof BPH and WBPH on deltamethrin-treated plants yields were not significantly lower thanin control non-treated plants Both IR62 and IR64 have been noted for their high toleranceto BPH damage particularly under high nitrogenous fertilizer applications [3456] as usedin our screenhouse experiment Therefore where plants have high tolerance to damageapplying insecticides could also result in profitability losses on rice farms and in the caseof insecticide-induced pest population increases tolerance might buffer against subsequentyield losses There are few reports of the phytotoxicity of insecticides to rice carbofuran istoxic to rice at high concentrations but may also enhance the growth of rice plants throughinteractions with nitrogen-fixing bacteria in the rhizosphere [80] Future studies couldmore carefully assess field-grown plants for possible phytotoxic effects particularly ifinsecticides have the potential to directly reduce yields
44 Management implications
Global trends suggest that resistance in rice has failed to reduce insecticide appli-cations [2] This may be due to aggressive marketing by agrochemical companies andstrategies to increase pesticide sales such as promoting prophylactic applications [16] Be-
Crops 2021 1 180
cause we selected our test insecticides (except the pyrethroids) without any a priori criteriaour results suggest that many currently used insecticides probably have some negativeeffects on yield either directly through phytotoxic effects or indirectly by inducing pestresurgence Furthermore our results indicate that even where pests reach high densitiesas on deltamethrin-treated plants in the screenhouse these densities do not necessarilylead to significant yield reductions
Among the products we used buprofezin although it had little effect on oviposi-tion was effective in reducing nymph survival Furthermore there were only minorapparent phytotoxic effects on the rice plants Buprofezin has a low toxicity to naturalenemies in rice fields [58] and is therefore the most compatible of the insecticides withstrategies to increase rice field biodiversity and restore ecosystem services [32ndash348182]Thiamethoxam-chlorantraniliprole was also effective in reducing both oviposition andnymph survival and had no apparent toxic effects on rice plants However thiamethoxam-chlorantraniliprole and other neonicotinoids although moderately toxic can have severeeffects on beneficial hymenopteran parasitoids [83] although they have apparently lowtoxicity to predatory mirid bugs [84] The remaining insecticides either directly reducedyields or were associated with higher BPH biomass or densities in our experiments
We noted few synergies with HPR although nymph biomass andor survival waslower in cartap hydrochloride and carbofuran-treated IR62 plants compared to correspond-ing IR64 plants Meanwhile cartap hydrochloride and deltamethrin were antagonisticto resistance in IR62 in the screenhouse experiment Among the strongest effects of HPRwas to act as a buffer against GLH population build-up on deltamethrin-treated IR62 andagainst carbofuran-induced increases in BPH nymph biomass in the survival bioassayTherefore HPR may play a role in reducing the possibilities of secondary outbreaks ofBPH and other insects The combined use of HPR and targeted insecticides against BPHcreates redundancy in the rice field and counters measures to manage insecticide resistanceand virulence by unnecessarily exposing plant protection products to evolving pest pop-ulations HPR (and host plant tolerance) can be effective against target pests as revealedin previous studies [4] and could be incorporated into other pest management practicesparticularly agroecological practices such as ecological engineering with which it is highlycompatible [818586]
5 Conclusions
Based on our results and the results of previous studies HPR and the applicationof chemical insecticides against BPH rarely produce synergistic effects Antagonisticeffects where chemicals caused increases in BPH fitness or populations on resistant ricewere more apparent in our study In particular deltamethrin caused an outbreak ofBPH in our screenhouse study Nevertheless HPR can buffer against the insecticide-induced enhancement of herbivore fitness (hormesis) and potentially reduce possibilities ofsecondary outbreaks as was suggested from results with cartap hydrochloride in our studyBased on our results we suggest that carbofuran cartap hydrochloride cypermethrindeltamethrin and fipronil should be avoided in rice particularly in the Philippines (wherethis research was conducted) due to their resurgence-causing potential or because of directreductions to rice yields
Author Contributions Conceptualization FGH methodology FGH AP-C and MLPA formalanalysis FGH and AP-C investigation FGH AP-C and MLPA data curation FGHAP-C and MLPA writingmdashoriginal draft preparation FGH and AP-C writingmdashreview andediting FGH AP-C and MLPA visualization FGH supervision FGH and MLPA projectadministration FGH funding acquisition FGH All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by the Bill and Melinda Gates Foundation (Cereal SystemsInitiative for South Asia [CSISA] OPP52303) and the Global Rice Science Platform (GRiSP) underthe directorship of Achim Dobermann AP-C was partially funded by The Spanish Governmentthrough Becas Faro International
Crops 2021 1 181
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data presented in this study are available on reasonable requestfrom the corresponding author
Acknowledgments The authors thank Fame Ramal Carmencita Bernal Alberto Naredo ReyeulQuintana Vincent Virtudes Marol Recide Jenyrose Geronda and Ellen Genil for assistance duringthis study The contributions of Rodante Abas (sadly deceased) during three decades of entomologicalresearch at IRRI are gratefully acknowledged Rice lines were supplied through the Genebank of IRRI
Conflicts of Interest The authors declare no conflict of interest
References1 Bottrell DG Schoenly KG Resurrecting the ghost of green revolutions past The brown planthopper as a recurring threat to
high-yielding rice production in tropical Asia J Asia-Pac Entomol 2012 15 122ndash140 [CrossRef]2 Horgan FG Integrated pest management for sustainable rice cultivation A holistic approach In Achieving Sustainable Cultivation
of Rice Cultivation Pest and Disease Management Sasaki T Ed Burleigh Dodds Scientific Cambridge UK 2017 pp 309ndash3423 Horgan FG de Freitas TFS Crisol-Martiacutenez E Mundaca EA Bernal CC Nitrogenous fertilizer reduces resistance but
enhances tolerance to the brown planthopper in fast-growing moderately resistant rice Insects 2021 12 989 [CrossRef] [PubMed]4 Horgan FG Pentildealver Cruz A Bernal CC Ramal AF Almazan MLP Wilby A Resistance and tolerance to the brown
planthopper Nilaparvata lugens (Staringl) in rice infested at different growth stages across a gradient of nitrogen applicationsField Crops Res 2018 217 53ndash65 [CrossRef]
5 Hereward JP Cai X Matias AMA Walter GH Xu C Wang Y Migration dynamics of an important rice pest The brownplanthopper (Nilaparvata lugens) across AsiamdashInsights from population genomics Evol Appl 2020 13 2449ndash2459 [CrossRef]
6 Hu C Guo J Fu X Huang Y Gao X Wu K Seasonal migration pattern of Nilaparvata lugens (Hemiptera Delphacidae) overthe Bohai Sea in northern China J Econ Entomol 2018 111 2129ndash2135 [CrossRef]
7 Ma J Wang Y-C Hu Y-Y Lu M-H Wan G-J Chen F-J Liu W-C Zhai B-P Hu G Brown planthopper Nilaparvatalugens was concentrated at the rear of the typhoon Soudelor in eastern China in August 2015 Insect Sci 2018 25 916ndash926[CrossRef]
8 Fujita D Kohli A Horgan FG Rice resistance to planthoppers and leafhoppers Crit Rev Plant Sci 2013 32 162ndash191[CrossRef]
9 Fujii T Yoshida K Kobayashi T Myint KKM Yasui H Sanada-Morimura S Matsumura M Long-term virulencemonitoring of differential cultivars in Japanrsquos immigrant populations of Nilaparvata lugens (Hemiptera Delphacidae) in 2001ndash2009Appl Entomol Zool 2021 56 407ndash418 [CrossRef]
10 Myint KKM Yasui H Takagi M Matsumura M Virulence of long-term laboratory populations of the brown planthopperNilaparvata lugens (Stal) and whitebacked planthopper Sogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) on rice differentialvarieties Appl Entomol Zool 2009 44 149ndash153 [CrossRef]
11 Horgan FG Ramal AF Bentur JS Kumar R Bhanu KV Sarao PS Iswanto EH Van Chien H Phyu MHBernal CC et al Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia againstresistant rice varieties Crop Prot 2015 78 222ndash231 [CrossRef]
12 Horgan FG Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers Crop Prot2018 110 21ndash33 [CrossRef]
13 Uddin ABMA Islam KS Jahan M Ara A Afrin S Farmerrsquos perception about resurgence of brown planthopper NilaparvataLugens (Staringl) in Bangladesh Bangladesh Rice J 2020 23 21ndash33 [CrossRef]
14 Triwidodo H Brown planthoppers infestations and insecticides use pattern in Java Indonesia AGRIVITA J Agric Sci 202042 320ndash330 [CrossRef]
15 Veres A Wyckhuys KA Kiss J Toacuteth F Burgio G Pons X Avilla C Vidal S Razinger J Bazok R et al An update of theWorldwide Integrated Assessment (WIA) on systemic pesticides Part 4 Alternatives in major cropping systems Environ SciPollut Res 2020 27 29867ndash29899 [CrossRef]
16 Bakker L van der Werf W Tittonell P Wyckhuys KA Bianchi FJ Neonicotinoids in global agriculture Evidence for a newpesticide treadmill Ecol Soc 2020 25 26 [CrossRef]
17 Wu J-C Xu J-X Yuan S-Z Liu J-L Jiang Y-H Xu J-F Pesticide-induced susceptibility of rice to brown planthopperNilaparvata lugens Entomol Exp Appl 2001 100 119ndash126 [CrossRef]
18 Wu J-C Qiu H-M Yang G-Q Liu J-L Liu G-J Wilkens RM Effective duration of pesticide-induced susceptibility of riceto brown planthopper (Nilaparvata lugens Staringl Homoptera Delphacidae) and physiological and biochemical changes in riceplants following pesticide application Int J Pest Manag 2004 50 55ndash62 [CrossRef]
19 Wang A-H Wu J-C Yu Y-S Liu J-L Yue J-F Wang MY Selective insecticide-induced stimulation on fecundity andbiochemical changes in Tryporyza incertulas (Lepidoptera Pyralidae) J Econ Entomol 2005 98 1144ndash1149 [CrossRef] [PubMed]
Crops 2021 1 182
20 Azzam S Wang F Wu J-C Shen J Wang L-P Yang G-Q Guo Y-R Comparisons of stimulatory effects of a seriesof concentrations of four insecticides on reproduction in the rice brown planthopper Nilaparvata lugens Staringl (HomopteraDelphacidae) Int J Pest Manag 2009 55 347ndash358 [CrossRef]
21 Ge L-Q Hu J-H Wu J-C Yang G-Q Gu H Insecticide-induced changes in protein RNA and DNA contents in ovary andfat body of female Nilaparvata lugens (Hemiptera Delphacidae) J Econ Entomol 2009 102 1506ndash1514 [CrossRef]
22 Ling S Zhang J Hu L Zhang R Effect of fipronil on the reproduction feeding and relative fitness of brown planthopperNilaparvata lugens Appl Entomol Zool 2009 44 543ndash548 [CrossRef]
23 Ge L-Q Wang L-P Zhao K-F Wu J-C Huang L-J Mating pair combinations of insecticide-treated male and femaleNilaparvata lugens Staringl (Hemiptera Delphacidae) planthoppers influence protein content in the male accessory glands (MAGs)and vitellin content in both fat bodies and ovaries of adult females Pestic Biochem Physiol 2010 98 279ndash288 [CrossRef]
24 Ge L-Q Wu J-C Zhao K-F Chen Y Yang G-Q Induction of Nlvg and suppression of Nljhe gene expression in Nilaparvatalugens (Staringl) (Hemiptera Delphacidae) adult females and males exposed to two insecticides Pestic Biochem Physiol 201098 269ndash278 [CrossRef]
25 Hu J-H Wu J-C Yin J-L Gu H-N Physiology of insecticide-induced stimulation of reproduction in the rice brownplanthopper (Nilaparvata lugens (Staringl)) Dynamics of protein in fat body and ovary Int J Pest Manag 2010 56 23ndash30 [CrossRef]
26 Wang L-P Shen J Ge L-Q Wu J-C Yang G-Q Jahn GC Insecticide-induced increase in the protein content of maleaccessory glands and its effect on the fecundity of females in the brown planthopper Nilaparvata lugens Staringl (HemipteraDelphacidae) Crop Prot 2010 29 1280ndash1285 [CrossRef]
27 Wu J Ge L Liu F Song Q Stanley D Pesticide-induced planthopper population resurgence in rice cropping systems AnnuRev Entomol 2020 65 409ndash429 [CrossRef]
28 Suri KS Singh G Insecticide-induced resurgence of the whitebacked planthopper Sogatella furcifera (Horvath) (HemipteraDelphacidae) on rice varieties with different levels of resistance Crop Prot 2011 30 118ndash124 [CrossRef]
29 Fabellar LT Heinrichs EA Toxicity of insecticides to predators of rice brown planthoppers Nilaparvata lugens (Staringl) (HomopteraDelphacidae) Environ Entomol 1984 13 832ndash837 [CrossRef]
30 Fabellar LT Heinrichs EA Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators Crop Prot1986 5 254ndash258 [CrossRef]
31 Cuong NL Ben PT Phuong LT Chau LM Cohen MB Effect of host plant resistance and insecticide on brown planthopperNilaparvata lugens (Staringl) and predator population development in the Mekong Delta Vietnam Crop Prot 1997 16 707ndash715[CrossRef]
32 Preetha G Stanley J Suresh S Samiyappan R Risk assessment of insecticides used in rice on mirid bug Cyrtorhinuslividipennis Reuter the important predator of brown planthopper Nilaparvata lugens (Stal) Chemosphere 2010 80 498ndash503[CrossRef] [PubMed]
33 Tanaka K Endo S Kazano H Toxicity of insecticides to predators of rice planthoppers Spiders the mirid bug and the dryinidwasp Appl Entomol Zool 2000 35 177ndash187 [CrossRef]
34 Zhang X Xu Q Lu W Liu F Sublethal effects of four synthetic insecticides on the generalist predator Cyrtorhinus lividipennisJ Pest Sci 2015 88 383ndash392 [CrossRef]
35 Settle WH Ariawan H Astuti ET Cahyana W Hakim AL Hindayana D Lestari AS Managing tropical rice peststhrough conservation of generalist natural enemies and alternative prey Ecology 1996 77 1975ndash1988 [CrossRef]
36 Salim M Heinrichs EA Insecticide-induced changes in the levels of resistance of rice cultivars to the whitebacked planthopperSogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) Crop Prot 1987 6 28ndash32 [CrossRef]
37 Yin J-L Xu H-W Wu J-C Hu J-H Yang G-Q Cultivar and insecticide applications affect the physiological development ofthe brown planthopper Nilaparvata lugens (Staringl) (Hemiptera Delphacidae) Environ Entomol 2014 37 206ndash212 [CrossRef]
38 Cheng C Chang W Studies on varietal resistance to the brown planthopper in Taiwan In Brown Planthopper Threat toRice Production in Asia International Rice Research Institute Ed International Rice Research Institute Manila Philippines1979 pp 251ndash271
39 Reissig WH Heinrichs EA Valencia SL Insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on ricevarieties with different levels of resistance Environ Entomol 1982 11 165ndash168 [CrossRef]
40 Heinrichs EA Fabellar LT Basilio RP Wen T-C Medrano F Susceptibility of rice planthoppers Nilaparvata lugens andSogatella furcifera (Homoptera Delphacidae) to insecticides as influenced by level of resistance in the host plant Environ Entomol1984 13 455ndash458 [CrossRef]
41 Joshi R Shepard B Kenmore P Lydia R Insecticide-induced resurgence of brown planthopper (BPH) on IR62 Int Rice ResNewsl 1992 17 9ndash10
42 Zhu Z-R Cheng J Jiang M-X Zhang X-X Complex influence of rice variety fertilization timing and insecticide onpopulation dynamics of Sogatella furcifera (Horvath) Nilaparvata lugens (Staringl) (Homoptera Delphacidae) and their natural enemiesin rice in Hangzhou China J Pest Sci 2004 77 65ndash74 [CrossRef]
43 Nanthakumar M Jhansi Lakshmi V Shashi Bhushan V Balachandran SM Mohan M Decrease of rice plant resistance andinduction of hormesis and carboxylesterase titre in brown planthopper Nilaparvata lugens (Staringl) by xenobiotics Pestic BiochemPhysiol 2012 102 146ndash152 [CrossRef]
Crops 2021 1 183
44 Yao H-W Jiang C-Y Ye G-Y Cheng J-A Studies on cultivar-induced changes in insecticide susceptibility and its relatedenzyme activities of the white-backed planthopper Sogatella furcifera (Horvath)(Homopter Delphacidae) Insect Sci 2002 9 9ndash15
45 Horgan FG Srinivasan TS Bentur JS Kumar R Bhanu KV Sarao PS Van Chien H Almazan MLP Bernal CCRamal AF et al Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers Implicationsfor rice breeding and gene deployment Agronomy 2017 7 62 [CrossRef]
46 Jairin J Phengrat K Teangdeerith S Vanavichit A Toojinda T Mapping of a broad-spectrum brown planthopper resistancegene Bph3 on rice chromosome 6 Mol Breed 2007 19 35ndash44 [CrossRef]
47 Ren J Gao F Wu X Lu X Zeng L Lv J Su X Luo H Ren G Bph32 a novel gene encoding an unknown SCRdomain-containing protein confers resistance against the brown planthopper in rice Sci Rep 2016 6 37645 [CrossRef]
48 Nguyen CD Zheng S-H Sanada-Morimura S Matsumura M Yasui H Fujita D Substitution mapping and characterizationof brown planthopper resistance genes from indica rice variety lsquoPTB33prime (Oryza sativa L) Breed Sci 2021 in press [CrossRef]
49 Chelliah S Heinrichs EA Factors affecting insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on RiceEnviron Entomol 1980 9 773ndash777 [CrossRef]
50 Moore MT Kroumlger R Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growthArch Environ Contam Toxicol 2010 59 574ndash581 [CrossRef]
51 Kim J Hwang I Han M Jang B Effect of insecticide and fungicide on phytotoxicity of herbicide in rice Korean J Weed Sci1986 6 67ndash75
52 Horgan FG Garcia CPF Haverkort F de Jong PW Ferrater JB Changes in insecticide resistance and host range performanceof planthoppers artificially selected to feed on resistant rice Crop Prot 2020 127 104963 [CrossRef] [PubMed]
53 Khush GS Virk PS IR Varieties and Their Impact International Rice Research Institute Manila Philippines 200554 Srinivasan TS Almazan MLP Bernal CC Ramal AF Subbarayalu MK Horgan FG Interactions between nymphs of
Nilaparvata lugens and Sogatella furcifera (Hemiptera Delphacidae) on resistant and susceptible rice varieties Appl Entomol Zool2016 51 81ndash90 [CrossRef]
55 Pentildealver Cruz A Arida A Heong KL Horgan FG Aspects of brown planthopper adaptation to resistant rice varieties withthe Bph3 gene Entomol Exp Appl 2011 141 245ndash257 [CrossRef]
56 Alam SN Cohen MB Detection and analysis of QTLs for resistance to the brown planthopper Nilaparvata lugens in adoubled-haploid rice population Theor Appl Genet 1998 97 1370ndash1379 [CrossRef]
57 Mackill DJ Khush GS IR64 A high-quality and high-yielding mega variety Rice 2018 11 18 [CrossRef] [PubMed]58 Heinrichs EA Basilio RP Valencia SL Buprofezin a selective insecticide for the management of rice planthoppers (Ho-
moptera Delphacidae) and leafhoppers (Homoptera Cicadellidae) Environ Entomol 1984 13 515ndash521 [CrossRef]59 Satapathy MK Anjaneyulu A Use of cypermethrin a synthetic pyrethroid in the control of rice tungro virus disease and its
vector Trop Pest Manag 1984 30 170ndash178 [CrossRef]60 Vorley W Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown
planthopper (BPH) in Kedah Malaysia Int Rice Res Newsl 1987 10 19ndash2061 Uddin A Islam KS Jahan M Ara A Khan MAI Factor influencing the resurgence of brown planthopper in Bangladesh
SAARC J Agric 2020 18 117ndash128 [CrossRef]62 Escalada MM Luecha M Heong KL Social impacts of planthopper outbreaks Case study from Thailand In Rice Planthoppers
Ecology Management Socio Economics and Policy Heong KL Cheng J Escalada MM Eds Zhejiang University PressHangzhou China 2015 pp 209ndash226
63 Zhang CP He HM Yu JZ Hu XQ Zhu YH Wang Q Residues of carbosulfan and its metabolites carbofuran and3-hydroxy carbofuran in rice field ecosystem in China J Environ Sci Health 2016 51 351ndash357 [CrossRef] [PubMed]
64 Samanta S Barman M Samanta NRA Bio-efficacy trials of carbofuran 3 CG against insect pests of rice J Entomol ZoolStud 2020 8 1529ndash1533
65 Donley N The USA lags behind other agricultural nations in banning harmful pesticides Environ Health 2019 18 44 [CrossRef][PubMed]
66 Adams A Gore J Musser F Cook D Catchot A Walker T Dobbins C Efficacy of selected insecticides applied to hybridrice seed J Econ Entomol 2015 109 200ndash206 [CrossRef]
67 Kumar R Singh B Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) Field Bull Environ Contam Toxicol2013 90 482ndash488 [CrossRef]
68 Dhaka S Prajapati C Singh D Singh R Field evaluation of insecticides and bio-pesticides against rice leaf folder CnaphalocrocisMedinalis (Guenee) Ann Plant Prot Sci 2011 19 324ndash326
69 Matsumura M Takeuchi H Satoh M Sanada-Morimura S Otuka A Watanabe T Van Thanh D Species-specific insecticideresistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in East and South-eastAsia Pest Manag Sci 2008 64 1115ndash1121 [CrossRef] [PubMed]
70 Tang J Li J Shao Y Yang B Liu Z Fipronil resistance in the whitebacked planthopper (Sogatella furcifera) Possible resistancemechanisms and cross-resistance Pest Manag Sci 2010 66 121ndash125 [CrossRef]
71 Horgan FG Arida A Ardestani G Almazan MLP Temperature-dependent oviposition and nymph performance revealdistinct thermal niches of coexisting planthoppers with similar thresholds for development PLoS ONE 2020 15 e0235506[CrossRef] [PubMed]
Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]

Crops 2021 1 171
rice production systems where farmers apply insecticides ge two times [14] however inour experiment we did not mix insecticides
Rice herbivores including BPH WBPH and GLH occurred at low densities in thescreenhouse Therefore at the opposite end of the screenhouse (separated by a distanceof 20 m) a bay with gt50 DAS TN1 rice plants was infested with high densities of BPH(approx 200 BPH plantminus1) This acted as a potential source of planthoppers to attackthe treated IR62 and IR64 plants in our experiment Natural enemies were largely absentfrom the screenhouse because of the insect-proof cladding (1 mm times 1 mm mesh bore)Microparasitoids (Oligosita spp and Anagrus spp) that could pass through the screenoccasionally attacked planthopper eggs in the screenhouse (estimated 5ndash10 of eggsparasitized unpublished data)
The IR62 and IR64 plants were continuously monitored to assess grain maturationWhen about 85 of the rice grain was mature any arthropods on the plants were collectedusing a vacuum sampler and stored in plastic vials The plants were then destructivelyharvested by carefully pulling the plant from the soil to maintain intact roots The plantroots were washed under running water and each plant was placed inside a paper bagThe plants and arthropods were then dried in a forced draft oven at 60 C for 7 days andweighed Arthropods were separated by species and counted The plants were separatedinto roots shoots and panicles and the numbers of reproductive and vegetative tillerswere recorded Rice grain was removed from the panicles and separated as filled andnon-filled grain The dried grain was counted and weighed
27 Data Analyses
The numbers of egg clusters and eggs laid in rice plants during the multichoice testwere analyzed using univariate general linear models (GLM) on ranked (within replicate)data Separate analyses were conducted for IR62 and IR64 plants Data from the binarychoice bioassays were analyzed using paired t-tests The numbers of egg clusters and eggslaid in treated and non-treated plants during the oviposition bioassay were compared usingunivariate GLM with plant biomass initially included as a covariate but later removedbecause it had no significant effect Cluster and egg numbers were ranked within blocksbefore analyses Average cluster size was estimated by dividing the number of eggs in eachplant by the number of clusters Cluster size was analyzed using univariate GLM removingthe block effect Similarly the proportions of surviving nymphs and nymph biomass fromthe survival bioassay were analyzed using univariate GLMs initially including plantbiomass as a covariate Data from the survival bioassay were ranked before analysesToo few nymphs survived under some treatments to examine development rates or thesex of surviving adults We estimated the combined effects of HPR and each insecticidelsquoxrsquo as -((fitness on untreated IR64 minus fitness on x-treated IR62)fitness on untreated IR64)To estimate the effect of the insecticide alone we removed the effect on HPR which weestimated as fitness on untreated IR62 minus fitness on untreated IR64 Fitness was assessedfrom the oviposition bioassay as the numbers of eggs laid and from the survival bioassayas nymph biomass Where nymph mortality on insecticide-treated IR64 plants approached100 HPR effects were regarded as insignificant
We also examined combined and insecticide effects from the screenhouse data basedon densities of BPH and WBPH (see below) Effects were calculated using insecticide andHPR combinations for each replicated block (N = 5) Estimated effects were comparedacross treatments using univariate GLMs Because three herbivores (BPH WBPH andGLH) occurred in our screenhouse plots we used a split-plot multivariate GLM to analyzeherbivore densities Total planthopper and leafhopper biomass and density and plantgrowth parameters (tiller number biomass yield grain number proportion of grainunfilled and grain size) were analyzed using split-plot univariate GLMs The covariateplant biomass was initially included in analyses of the effects on herbivores but had noeffect and was subsequently removed Similarly the covariate herbivore biomass wasinitially included in analyses of plant growth parameters but had no effect and was
Crops 2021 1 172
subsequently removed The combined effects of insecticide and HPR and the separateeffects of each insecticide (calculated as indicated above) on BPH and WBPH were alsoanalyzed using univariate GLM
Post-hoc Tukey tests were applied to all significant treatment factors Residuals wereplotted following all parametric analyses and were found to be normal and homogenous
3 Results31 Effects of HPR and Insecticides on BPH Preferences
Treatments affected the number of clusters and eggs that were laid in IR62 plants(ranked clusters and eggs F732 = 4738 p lt 0001) and in IR64 plants (clusters F732 = 6229p lt 0001 eggs F732 = 7757 p lt 0001) (Figure 1AB) Fewer eggs were laid on IR64 plantstreated with the pyrethroids (cypermethrin and deltamethrin) fipronil and thiamethoxam-chlorantraniliprole compared to plants treated with buprofezin carbofuran and cartaphydrochloride (Figure 1B) Cypermethrin-treated IR62 plants also had fewer eggs thanbuprofezin-treated IR62 plants (Figure 1A) Although BPH females tended to lay fewereggs in many of the insecticide-treated rice plants in the binary choice experiments therewere no statistically significant differences between the numbers of eggs laid in treated-IR62(t ranges from minus0746 to 3186 p gt 005) or treated-IR64 (t ranges from minus0108 to 2789p gt 005) compared to corresponding non-treated plants (Figure 1CD)
Crops 2021 1 FOR PEER REVIEW 7
herbivore densities Total planthopper and leafhopper biomass and density and plant growth parameters (tiller number biomass yield grain number proportion of grain un-filled and grain size) were analyzed using split-plot univariate GLMs The covariate plant biomass was initially included in analyses of the effects on herbivores but had no effect and was subsequently removed Similarly the covariate herbivore biomass was initially included in analyses of plant growth parameters but had no effect and was subsequently removed The combined effects of insecticide and HPR and the separate effects of each insecticide (calculated as indicated above) on BPH and WBPH were also analyzed using univariate GLM
Post-hoc Tukey tests were applied to all significant treatment factors Residuals were plotted following all parametric analyses and were found to be normal and homogenous
3 Results 31 Effects of HPR and Insecticides on BPH Preferences
Treatments affected the number of clusters and eggs that were laid in IR62 plants (ranked clusters and eggs F732 = 4738 p lt 0001) and in IR64 plants (clusters F732 = 6229 p lt 0001 eggs F732 = 7757 p lt 0001) (Figure 1AB) Fewer eggs were laid on IR64 plants treated with the pyrethroids (cypermethrin and deltamethrin) fipronil and thiameth-oxam-chlorantraniliprole compared to plants treated with buprofezin carbofuran and cartap hydrochloride (Figure 1B) Cypermethrin-treated IR62 plants also had fewer eggs than buprofezin-treated IR62 plants (Figure 1A) Although BPH females tended to lay fewer eggs in many of the insecticide-treated rice plants in the binary choice experiments there were no statistically significant differences between the numbers of eggs laid in treated-IR62 (t ranges from minus0746 to 3186 p gt 005) or treated-IR64 (t ranges from minus0108 to 2789 p gt 005) compared to corresponding non-treated plants (Figure 1CD)
Figure 1 Results from multichoice (AB) and binary choice (CD) bioassays conducted in the green-house Bioassays were conducted separately for IR62 (AC) and IR64 (BD) Plants were treated with one of seven insecticides (A to G) as identified in the legend Control plants were sprayed with water Error bars are presented (N = 5 for (A) and (B) N = 3 for (C) and (D)) Lowercase letters (andashc) indicate homogenous treatment groups (Tukey p lt 005)
A B C D E F G0
100
200
300
400
Treatment
A B C D E F G
Non treatedTreated
A B C D E F G Control
Num
ber o
f egg
s pe
r pla
nt
0
100
200
300
400
A B C D E F G Control
IR62 IR64
A
C
B
D
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliproleH Control
a abab
c
abc
abc bc bc
c
aaa
ab
ab
bb b
Figure 1 Results from multichoice (AB) and binary choice (CD) bioassays conducted in thegreenhouse Bioassays were conducted separately for IR62 (AC) and IR64 (BD) Plants were treatedwith one of seven insecticides (A to G) as identified in the legend Control plants were sprayed withwater Error bars are presented (N = 5 for (AB) N = 3 for (CD)) Lowercase letters (andashc) indicatehomogenous treatment groups (Tukey p lt 005)
32 Effects of HPR and Insecticides on BPH Fitness
More eggs were laid on IR64 plants than on IR62 plants but BPH produced similarnumbers of egg clusters on both plants The lower numbers of eggs in IR62 were due tosmaller clusters on the variety (Table 1) The numbers of eggs and clusters were affectedby insecticide treatment with fewer eggs and clusters on cypermethrin-treated fipronil-treated and thiamethoxam-chlorantraniliprole-treated plants than on plants under the
Crops 2021 1 173
remaining treatments (Table 1) The treatment-effects were not affected by host variety (ieinteractions were non-significant Table 1) Insecticide treatments had no effect on clustersize in the bioassay (Table 1) There was no significant effect of combined insecticide andHPR (F628 = 0449 p = 0840) on observed changes in egg laying during the bioassay butthe estimated insecticide effect was greater for thiamethoxam-chlorantraniliprole fiproniland cypermethrin than for buprofezin cartap hydrochloride and carbofuran (F628 = 3140p = 0018 Figure 2A)
Table 1 Results from greenhouse oviposition and survival tests with IR62 (resistant) and IR64 (susceptible) each treatedwith one of seven insecticides Numbers are means plusmn SEM (N = 5)
Treatments Egg Clusters(Number Plantminus1) 12
Eggs(Number Plantminus1) 12
Cluster Size(Number of Eggs) 1
Nymph Survival(Proportions) 12
Nymph Biomass (mgDry Weight Plantminus1) 12
IR62Buprofezin 1240 plusmn 140 b 5040 plusmn 904 b 393 plusmn 036 000 plusmn 000 a 000 plusmn 000 a
Carbofuran 1300 plusmn 089 b 5380 plusmn 421 b 417 plusmn 026 064 plusmn 064 d 154 plusmn 154 d
Cartaphydrochloride 1360 plusmn 075 b 5200 plusmn 486 b 382 plusmn 029 028 plusmn 028 b 151 plusmn 151 b
Cypermethrin 620 plusmn 169 a 2000 plusmn 608 a 316 plusmn 023 002 plusmn 002 a 002 plusmn 002 a
Deltamethrin 1360 plusmn 160 b 4340 plusmn 566 b 338 plusmn 069 040 plusmn 040 c 120 plusmn 120 c
Fipronil 520 plusmn 146 a 2240 plusmn 735 a 393 plusmn 064 000 plusmn 000 a 000 plusmn 000 a
Thiamethoxam-chlorantraniliprole 380 plusmn 086 a 1700 plusmn 435 a 447 plusmn 032 000 plusmn 000 a 000 plusmn 000 a
Control 1260 plusmn 103 b 4900 plusmn 716 b 388 plusmn 054 080 plusmn 080 d 292 plusmn 292 d
IR64Buprofezin 2100 plusmn 164 10940 plusmn 1346 556 plusmn 117 000 plusmn 000 000 plusmn 000Carbofuran 1820 plusmn 404 9880 plusmn 2939 524 plusmn 074 086 plusmn 086 420 plusmn 420
Cartaphydrochloride 2140 plusmn 619 11460 plusmn 3141 549 plusmn 064 010 plusmn 010 045 plusmn 045
Cypermethrin 380 plusmn 233 1980 plusmn 1155 550 plusmn 058 000 plusmn 000 000 plusmn 000Deltamethrin 1560 plusmn 209 7500 plusmn 1581 473 plusmn 069 040 plusmn 040 201 plusmn 201
Fipronil 380 plusmn 086 2100 plusmn 552 546 plusmn 080 000 plusmn 000 000 plusmn 000Thiamethoxam-chlorantraniliprole 260 plusmn 075 800 plusmn 212 373 plusmn 133 000 plusmn 000 000 plusmn 000
Control 1740 plusmn 364 7520 plusmn 2344 426 plusmn 073 086 plusmn 086 360 plusmn 360F-treatment 3 19898 19898 0541 ns 133811 108451
F-variety 3 2596 ns 7607 7471 1080 ns 0008 ns
F-interaction 3 1840 ns 2034 ns 0610 ns 4902 9496 1 Lowercase letters (andashd) indicate homogenous treatment groups (for IR62 and IR64 combined) 2 Data were ranked within blocks beforeanalysis 3 DF = 164 for varieties 764 for treatments and interactions ns = p gt 005 = p le 001 = p le 0001
Nymph survival and biomass were not affected by rice variety (Table 1) Buprofezinfipronil thiamethoxam-chlorantraniliprole and cypermethrin reduced survival to 0 in thebioassay and all insecticides except carbofuran resulted in lower survival and biomass thanon untreated plants (Table 1) There were significant interactions because of higher nymphmortality and a resulting lower biomass on carbofuran-treated IR62 compared to untreatedIR62 but lower mortality and a higher biomass of nymphs on carbofuran-treated IR64than on untreated IR64 (Table 1) There was no significant effect of insecticide treatment(F628 = 0946 p = 0479) on observed changes in nymph biomass during the bioassaybut the estimated combined effects of pesticide and HPR was greater for thiamethoxam-chlorantraniliprole fipronil cypermethrin and buprofezin than for cartap hydrochlorideand carbofuran (F628 = 5699 p = 0001) (Figure 2B)
Crops 2021 1 174
Crops 2021 1 FOR PEER REVIEW 9
Figure 2 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) egg laying and (B) nymph biomass in greenhouse bioas-says Effects are indicated as proportional changes in fitness parameters compared to untreated IR64 Negative numbers indicate reduced fitness Standard errors are indicated (N = 5) Lowercase letters (ab) indicated homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 1)
Nymph survival and biomass were not affected by rice variety (Table 1) Buprofezin fipronil thiamethoxam-chlorantraniliprole and cypermethrin reduced survival to 0 in the bioassay and all insecticides except carbofuran resulted in lower survival and biomass than on untreated plants (Table 1) There were significant interactions because of higher nymph mortality and a resulting lower biomass on carbofuran-treated IR62 compared to untreated IR62 but lower mortality and a higher biomass of nymphs on carbofuran-treated IR64 than on untreated IR64 (Table 1) There was no significant effect of insecticide treatment (F628 = 0946 p = 0479) on observed changes in nymph biomass during the bio-assay but the estimated combined effects of pesticide and HPR was greater for thiameth-oxam-chlorantraniliprole fipronil cypermethrin and buprofezin than for cartap hydro-chloride and carbofuran (F628 = 5699 p = 0001) (Figure 2B)
33 Effects of HPR and Insecticides on Planthoppers and Leafhoppers Plants in the screenhouse were colonized by BPH WBPH and GLH Densities for
each species were similar on IR62 and IR64 (Wilkrsquos λ = 0994 p = 0952) There was a sig-nificant treatment effect on all three species (Wilkrsquos λ = 0586 p = 0049) because of high densities on deltamethrin-treated plants (Table 2) The numbers and biomass of all three species combined were also greater on deltamethrin-treated plants than on the remaining treatments (Table 2) Host resistance reduced GLH densities under the deltamethrin treat-ment but had no apparent effect on densities under the other treatments (Table 2) There were significant combined effects of HPR and insecticide on the numbers of BPH (F624 = 2686 p = 0039) and WBPH (F624 = 2482 p = 0052) this was almost exclusively due to the resurgence effect of deltamethrin on the planthoppers (BPH F624 = 2686 p = 0039 WBPH F624 = 2482 p = 0052) (Figure 3)
Δ eg
gs(P
ropo
rtion
)
-10
-08
-06
-04
-02
00
02
04
06
Treatment
A B C D E F G
Δ N
ymph
bio
mas
s(P
ropo
rtion
)
-10
-08
-06
-04
-02
00
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
ab
a a
b
a
b b
a a a a a a a a
b b bb
bb
Figure 2 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone(P grey bars) on (A) egg laying and (B) nymph biomass in greenhouse bioassays Effects are indicated as proportionalchanges in fitness parameters compared to untreated IR64 Negative numbers indicate reduced fitness Standard errors areindicated (N = 5) Lowercase letters (ab) indicated homogenous treatment groups based on Tukey pairwise comparisons(p lt 005) (see also Table 1)
33 Effects of HPR and Insecticides on Planthoppers and Leafhoppers
Plants in the screenhouse were colonized by BPH WBPH and GLH Densities foreach species were similar on IR62 and IR64 (Wilkrsquos λ = 0994 p = 0952) There was asignificant treatment effect on all three species (Wilkrsquos λ = 0586 p = 0049) because of highdensities on deltamethrin-treated plants (Table 2) The numbers and biomass of all threespecies combined were also greater on deltamethrin-treated plants than on the remainingtreatments (Table 2) Host resistance reduced GLH densities under the deltamethrintreatment but had no apparent effect on densities under the other treatments (Table 2)There were significant combined effects of HPR and insecticide on the numbers of BPH(F624 = 2686 p = 0039) and WBPH (F624 = 2482 p = 0052) this was almost exclusively dueto the resurgence effect of deltamethrin on the planthoppers (BPH F624 = 2686 p = 0039WBPH F624 = 2482 p = 0052) (Figure 3)
Crops 2021 1 175
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants underseven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density(Number Plantminus1) 1
WBPH Density(Number Plantminus1) 1
GLH Density(Number Plantminus1) 1
Density ofPlanthoppers and
Leafhoppers(Number Plantminus1) 1
Biomass ofPlanthoppers and
Leafhoppers(mg Dry Weight) 1
IR62Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a
Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartaphydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a
Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a
Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a
Thiamethoxam-chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a
IR64Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartaphydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123
Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns
F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Crops 2021 1 FOR PEER REVIEW 10
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants under seven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density (Num-
ber Plantminus1) 1 WBPH Density
(Number Plantminus1) 1 GLH Density
(Number Plantminus1) 1
Density of Planthoppers and Leafhoppers (Number
Plantminus1) 1
Biomass of Planthoppers and Leafhoppers (mg Dry
Weight) 1 IR62
Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartap hydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a Thiamethoxam-
chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a IR64
Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132 Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartap hydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123 Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113 Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052 Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087 F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns 1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatments and interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in the screenhouse Effects are indicated as proportional changes in planthopper densities compared to untreated IR64 Negative numbers indicate reduced densities positive numbers are increased den-sities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
Δ BP
H(N
umbe
r pla
nt-1
)
0
5
10
15
20
Treatment
A B C D E F G
Δ W
BPH
(Num
ber p
lant
-1)
0
20
30
40
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
a a
aa a a
a a a
a
a a
b
b
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimatedpesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in thescreenhouse Effects are indicated as proportional changes in planthopper densities compared to
Crops 2021 1 176
untreated IR64 Negative numbers indicate reduced densities positive numbers are increaseddensities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenoustreatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
34 Direct and Indirect Effects of Insecticides on Rice Development
IR62 plants had more tillers and attained a greater biomass than IR64 plants in thescreenhouse (Table 3) IR62 grain matured faster and plants were harvested earlier than theIR64 plants (Table 3) IR62 plants also produced more grain (number of grain) and hadhigher yields than IR64 but had smaller grain and more unfilled grain than IR64 (Table 3)
Table 3 Rice growth parameters for IR62 (resistant) and IR64 (susceptible) under seven insecticide treatments and forcontrol non-treated plants in a screenhouse environment Numbers are means plusmn SEM
Treatment Tiller Number 1 Plant Biomass(g Dry Weight) 1
Time to Harvest(Days) 1
Weightof Filled Grain(g Dry Weight) 1
Number ofFilled Grain
(1000s) 1
Unfilled Grain(Proportion) 1
1000 GrainWeight 1
IR62Buprofezin 2660 plusmn 206 ab 4643 plusmn 385 ab 7990 plusmn 168 ab 5776 plusmn 683 ab 361plusmn 061 ab 011 plusmn 003 a 2008 plusmn 042 ab
Carbofuran 2900 plusmn 106 ab 5546 plusmn 357 b 7720 plusmn 080 ab 7448 plusmn 691 b 446plusmn 061 ab 011 plusmn 001 a 2103 plusmn 054 b
Cartaphydrochloride 2820 plusmn 327 ab 4896 plusmn 470 ab 7920 plusmn 153 b 6430 plusmn 712 ab 415plusmn 064 ab 012 plusmn 001 b 2023 plusmn 065 a
Cypermethrin 2870 plusmn 301 a 4957 plusmn 562 a 7760 plusmn 103 b 6032 plusmn 822 a 410plusmn 067 ab 014 plusmn 002 a 1965 plusmn 076 a
Deltamethrin 2620 plusmn 345 a 4592 plusmn 588 a 7840 plusmn 108 ab 5698 plusmn 832 ab 379plusmn 047 ab 012 plusmn 001 a 2030 plusmn 063 ab
Fipronil 2380 plusmn 176 ab 4607 plusmn 657 ab 7700 plusmn 179 ab 5266 plusmn 390 a 318 plusmn 032 a 013 plusmn 002 a 2128 plusmn 047 b
Thiamethoxam-chlorantraniliprole 2650 plusmn 250 b 4547 plusmn 429 b 7720 plusmn 102 a 5777 plusmn 688 ab 408 plusmn 061 b 013 plusmn 002 a 2019 plusmn 067 ab
Control 3120 plusmn 247 b 5401 plusmn 555 b 7720 plusmn 080 ab 7045 plusmn 520 b 396plusmn 036 ab 012 plusmn 002 a 2076 plusmn 050 ab
IR64Buprofezin 2340 plusmn 250 5064 plusmn 747 8350 plusmn 124 5080 plusmn 952 270 plusmn 054 008 plusmn 001 2277 plusmn 039Carbofuran 2040 plusmn 097 4591 plusmn 141 8540 plusmn 073 4951 plusmn 312 263 plusmn 023 006 plusmn 000 2295 plusmn 036
Cartaphydrochloride 2120 plusmn 075 4336 plusmn 285 8520 plusmn 140 4273 plusmn 579 238 plusmn 026 015 plusmn 007 2050 plusmn 234
Cypermethrin 1990 plusmn 089 3968 plusmn 272 8710 plusmn 135 3920 plusmn 552 250 plusmn 028 008 plusmn 002 2171 plusmn 070Deltamethrin 1980 plusmn 181 4325 plusmn 357 8460 plusmn 068 4710 plusmn 496 277 plusmn 024 008 plusmn 001 2314 plusmn 012
Fipronil 2120 plusmn 174 4697 plusmn 311 8450 plusmn 089 4888 plusmn 392 274 plusmn 027 008 plusmn 001 2247 plusmn 042Thiamethoxam-chlorantraniliprole 2520 plusmn 287 5404 plusmn 465 8380 plusmn 196 5594 plusmn 546 347 plusmn 053 010 plusmn 001 2265 plusmn 072
Control 2160 plusmn 076 4678 plusmn 342 8600 plusmn 096 4940 plusmn 543 267 plusmn 029 008 plusmn 001 2283 plusmn 049F-treatment 4100 ns 3410 2253 ns 5150 2555 3923 4924
F-variety 168232 7665 547500 104356 118242 32308 106597 F-interaction 5980 6113 4824 5760 2921 2769 2552
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Insecticides affected plant growth directly and indirectly (through increased planthop-per densities) Plants treated with cypermethrin and deltamethrin had fewer tillers had alower biomass and under cypermethrin had lower yields than untreated plants (Table 3)Plants treated with fipronil also had reduced yields due to lower grain numbers comparedto untreated plants (Table 3) Although yields were not affected treatment with cartaphydrochloride increased the proportion of unfilled grain compared to untreated plantsof both varieties Cartap hydrochloride and cypermethrin also caused a delay in grainmaturation on IR62 and IR64 respectively relative to thiamethoxam-chlorantraniliproletreated plants (Table 3)
There were several significant interactions tiller numbers on both plants were similarunder buprofezin fipronil and thiamethoxam-chlorantraniliprole treatments but wereotherwise higher on IR62 (Table 3) biomass was lower on IR64 under carbofuran andcypermethrin treatments but was otherwise similar on both plants (Table 3) buprofezindelayed grain maturation on IR62 but maturation was otherwise faster on IR62 comparedto IR64 (Table 3) yields under buprofezin fipronil and thiamethoxam-chlorantraniliprolewere similar for IR62 and IR64 but otherwise yields were higher on IR62 (Table 3) grain
Crops 2021 1 177
numbers were higher on IR62 than IR64 except under fipronil treatments (Table 3) theproportions of grain unfilled were higher on IR62 but were similar between varieties underthe fipronil treatment (Table 3) 1000-grain weight was higher in IR64 but similar on bothvarieties when treated with cartap hydrochloride (Table 3)
4 Discussion
Our results using a highly resistant rice variety and some commonly used insecticidesindicate a range of possible effects on planthoppers and rice plants by combining these twomanagement practices
41 Synergies
The most predictable effect of combining HPR and insecticides is a synergy in reduc-ing BPH numbers This is suggested because planthoppers are weakened by the hostrsquosresistance and should therefore be more susceptible to the toxic effects of insecticides [40]Using spray towers over potted plants Heinrichs et al (1984) [40] examined the combinedeffects of a range of insecticides (that included carbofuran) with resistance by comparingBPH mortality on six resistant and two susceptible rice varieties Their results indicatedthat mortality was greatest when insecticides were applied to BPH on resistant varietiesThe effects were similar when the study was repeated with WBPH for three insecticides(including carbofuran) on the resistant variety N22 (Wbph1 gene) [40] In our study hostresistance reduced egg laying and nymph biomass on non-treated IR62 compared to non-treated IR64 Egg laying was also reduced on plants treated with cypermethrin fipronilor thiamethoxam-chlorantraniliprole (Table 1) Based on the results of our choice andno-choice oviposition bioassays cypermethrin acted mainly as a deterrent of gravid fe-males whereas fipronil and thiamethoxam-chlorantraniliprole directly reduced egg layingDespite similar BPH responses to both the insecticides and resistant host there were no ap-parent synergies between any of these insecticides and resistance in reducing egg numbersin the oviposition bioassay In contrast to these three insecticides our bioassays indicatedthat buprofezin carbofuran and cartap hydrochloride can lead to higher numbers of eggson treated plants compared to untreated plants (Table 1) These effects were not statisticallysignificant and may be exaggerated because bioassays were conducted over relativelyshort time scales For example planthoppers will sometimes respond to stresses such ashigh temperatures by increasing oviposition rates [7172] Furthermore the stimulation ofegg-laying by these chemicals was not apparent in our screenhouse experiment (Table 2)Nevertheless our results do indicate that host resistance reduced the magnitude of anystimulatory effect on oviposition egg numbers on buprofezin-treated carbofuran-treatedand cartap hydrochloride-treated IR64 in the no-choice bioassay were twice as high as onthe corresponding IR62 plants In cases such as these the resistance of IR62 functions as abuffer or lsquoinsurancersquo against possible undesired pesticide effects [1238]
Although buprofezin appeared to stimulate oviposition the growth hormone had astrong impact on nymph development leading to zero survival of exposed nymphs in ourgreenhouse bioassay Cypermethrin fipronil and thiamethoxam-chlorantraniliprole alsoreduced nymph survival to zero in the greenhouse bioassay (Table 1) Where insecticideshave such high efficiencies any fitness losses to BPH due to host resistance then becomeredundant Unlike buprofezin carbofuran also stimulated nymph biomass on IR64 in thesurvival bioassay (Table 1) however nymph biomass was lower on carbofuran-treated IR62than on non-treated IR62 indicating that the hostrsquos resistance buffered against undesiredstimulatory effects on BPH and may have functioned synergistically to augment mortalityand slow BPH growth Such buffering allows insecticides to be used against other pestswithout inducing secondary outbreaks of non-target pests In our screenhouse experimentherbivore densities were generally low on both IR62 and IR64 for most treatments (exceptdeltamethrin) and were below threshold densities for insecticide applicationsmdashwhichare normally about 25ndash50 planthoppers per hill [1373] Under such low densities it wasnot possible to observe the impact of host resistance on planthopper population build-
Crops 2021 1 178
up There was some indication of higher densities on cartap hydrochloride-treated IR62in the screenhouse which may be related to the stimulatory effects of the product onegg laying and the low mortality of nymphs on cartap hydrochloride-treated plants asobserved in the greenhouse bioassays (Table 1) Field studies from the Philippines haveindicated that IR62 effectively reduces BPH densities during wet and dry seasons [4]However farmers are rarely aware of which anti-herbivore resistance their rice varietiespossess and frequently apply pesticides regardless of resistance thereby reducing theirfarm profits [3841] For example Joshi et al (1992) [41] reported widespread outbreaks ofBPH on IR62 in Mindanao Philippines after applications of decamethrin (a pyrethroid) tothe resistant variety Where such resurgence-causing insecticides are applied resistanceseems to be often incapable of reducing the stimulatory effects of the insecticide and itloses its capacity to buffer against outbreaks
42 Antagonisms
Several insecticides are associated with increasing densities of planthoppers andwith resurgence outbreaks in farmersrsquo fields [27293041496174] Indeed patches oflsquohopperburnrsquo (ie dead rice plants resulting from planthopper damage) are often anindicator of high insecticide use in tropical Asia [214] In many cases these resurgence-causing insecticides were once effective in reducing planthopper densities but over timepopulations developed resistance against the products The resulting low toxicity canthen have a stimulatory effect on planthopper feeding and development [2123242743]This may be through hormesis which is a direct response by the planthopper to low-levelexposure to the toxic chemical or it may result from physiological changes to the rice plantsthat ultimately benefit the planthopper [1849] In our screenhouse study we induced aresurgence outbreak of BPH WBPH and to a lesser extent GLH on deltamethrin-treatedrice plants (Table 2) The combined densities of all three herbivores increased almost10 times on the treated plants Furthermore herbivore biomass increased by between fourand six times Although the resurgence effects of deltamethrin have been documentedin previous studies [2029ndash313975] we could not predict from our greenhouse studiesthat any such outbreak would occur Egg laying was unaffected by deltamethrin in ourchoice and no-choice experiments and nymph survival and development both declinedon deltamethrin-treated rice in the survival bioassay (Table 1) The massive increases inplanthopper densities on deltamethrin-treated plants in our study seem to result froma heightened attraction of adult BPH and WBPH to the plants [49] particularly sincenumbers were so low on all other plants including both treated and non-treated plantsHowever once on the plants planthoppers also apparently had enhanced fitness Cuonget al (1997) [31] observed higher population densities of BPH on deltamethrin-treatedrice in Vietnam albeit with lower-level increases on resistant varieties Furthermore in agreenhouse study Azzam et al (2009) [20] found that deltamethrin reduced pre-ovipositiontimes increased the duration of oviposition and increased egg hatchability and femalelongevity Nanthakumar et al (2012) [43] further showed that deltamethrin increased BPHfeeding efficiency on resistant and susceptible rice plants as determined through honeydewproduction and population size In studies with WBPH Salim and Heinrichs (1987) [36]showed deltamethrin to increase settling (antixenosis) feeding and population growthand Suri and Singh (2011) [28] showed deltamethrin to increase WBPH reproductive ratesand nymph survival In our study the observed resurgence of planthoppers was similaron both varieties indicating as was previously noted by Joshi et al (1992) [41] thatresurgence-causing pyrethroids are antagonistic to resistance in IR62
Though not statistically significant cartap hydrochloride also increased BPH numberson IR62 but not on IR64 in our screenhouse study This indicates that the product mayalso be antagonistic to the hostrsquos resistance mechanisms The stronger effect on IR62 maybe related to the differences between IR62 biomass growth rates and anatomy (ie alarger plant with higher tillering) compared to IR64 (Table 3) Once resistance is no longereffective because of chemically-enhanced BPH fitness these plant characteristics which
Crops 2021 1 179
have been related to observed preferences in BPH and other sap-sucking insects [349]could further accelerate the build-up of BPH populations on cartap hydrochloride-treatedplants To further test this idea experiments could be conducted using near-isogenic ricelines that differ mainly in the presence or absence of tagged resistance genes [76] Suchtests would avoid confounding differences in plant anatomy between test varieties IR62resistance to BPH has been well studied [4] The variety is also resistant to WBPH atolder plant stages [54] Therefore in our study deltamethrin was also antagonistic tothis WBPH-resistance (Table 2) However BPH feeding also facilitates WBPH populationgrowth [5477] and there may have been added indirect plant-mediated effects [77] thataugmented WBPH populations on both varieties in our experiment IR62 is also resistant toGLH [45] In our experiments GLH densities also increased on deltamethrin-treated plantsbut the effect was only apparent on IR64 Therefore GLH-resistance in IR62 apparentlybuffered against deltamethrin-associated increases in GLH on the variety as was alsonoted from field studies in Vietnam [31] Deltamethrin is currently not recommended foruse in rice [202878] however the product continues to be marketed in rice-dominatedregions throughout Asia [131479] Although cypermethrin has been associated with BPHoutbreaks and was consequently removed for use on rice in Thailand and Vietnam [62]we did not observe cypermethrin-induced resurgence in our screenhouse experimentNevertheless cypermethrin has been associated with resurgence of BPH in South Asia [61]and we have noted stimulatory effects of cypermethrin on other rice herbivores in thePhilippines (Horgan unpublished data)
43 Phytotoxicity
Phytotoxicity is relatively common following applications of insecticides [182850]Phytotoxic effects may change a plantrsquos physiology without consequences for crop yieldsHowever such changes to plant physiology could have indirect effects on herbivores Forexample deltamethrin methyl parathion and quinalphos produce increases in total sugarsand reducing sugars as well as increasing the protein contents of the rice leaf sheath andleaf blade [28] Several insecticides also reduce plant phenols in ricemdashwhich increases therice plantrsquos susceptibility to BPH and other insects [27] In our screenhouse experiment wenoted several changes to rice anatomy and yields Importantly we noted that cypermethrinand fipronil both reduced rice yields (Table 3) This was not due to indirect herbivoreeffects because planthopper and leafhopper densities were consistently low on rice treatedwith these insecticides The effects therefore probably resulted from direct phytotoxiceffects on the plants Furthermore these effects were most pronounced on treated IR62plants Other minor effects on grain production included increased proportions of unfilledgrain in cartap hydrochloride-treated plants and delays in grain maturation on cartaphydrochloride-treated and cypermethrin-treated plants (Table 3) Despite the high densitiesof BPH and WBPH on deltamethrin-treated plants yields were not significantly lower thanin control non-treated plants Both IR62 and IR64 have been noted for their high toleranceto BPH damage particularly under high nitrogenous fertilizer applications [3456] as usedin our screenhouse experiment Therefore where plants have high tolerance to damageapplying insecticides could also result in profitability losses on rice farms and in the caseof insecticide-induced pest population increases tolerance might buffer against subsequentyield losses There are few reports of the phytotoxicity of insecticides to rice carbofuran istoxic to rice at high concentrations but may also enhance the growth of rice plants throughinteractions with nitrogen-fixing bacteria in the rhizosphere [80] Future studies couldmore carefully assess field-grown plants for possible phytotoxic effects particularly ifinsecticides have the potential to directly reduce yields
44 Management implications
Global trends suggest that resistance in rice has failed to reduce insecticide appli-cations [2] This may be due to aggressive marketing by agrochemical companies andstrategies to increase pesticide sales such as promoting prophylactic applications [16] Be-
Crops 2021 1 180
cause we selected our test insecticides (except the pyrethroids) without any a priori criteriaour results suggest that many currently used insecticides probably have some negativeeffects on yield either directly through phytotoxic effects or indirectly by inducing pestresurgence Furthermore our results indicate that even where pests reach high densitiesas on deltamethrin-treated plants in the screenhouse these densities do not necessarilylead to significant yield reductions
Among the products we used buprofezin although it had little effect on oviposi-tion was effective in reducing nymph survival Furthermore there were only minorapparent phytotoxic effects on the rice plants Buprofezin has a low toxicity to naturalenemies in rice fields [58] and is therefore the most compatible of the insecticides withstrategies to increase rice field biodiversity and restore ecosystem services [32ndash348182]Thiamethoxam-chlorantraniliprole was also effective in reducing both oviposition andnymph survival and had no apparent toxic effects on rice plants However thiamethoxam-chlorantraniliprole and other neonicotinoids although moderately toxic can have severeeffects on beneficial hymenopteran parasitoids [83] although they have apparently lowtoxicity to predatory mirid bugs [84] The remaining insecticides either directly reducedyields or were associated with higher BPH biomass or densities in our experiments
We noted few synergies with HPR although nymph biomass andor survival waslower in cartap hydrochloride and carbofuran-treated IR62 plants compared to correspond-ing IR64 plants Meanwhile cartap hydrochloride and deltamethrin were antagonisticto resistance in IR62 in the screenhouse experiment Among the strongest effects of HPRwas to act as a buffer against GLH population build-up on deltamethrin-treated IR62 andagainst carbofuran-induced increases in BPH nymph biomass in the survival bioassayTherefore HPR may play a role in reducing the possibilities of secondary outbreaks ofBPH and other insects The combined use of HPR and targeted insecticides against BPHcreates redundancy in the rice field and counters measures to manage insecticide resistanceand virulence by unnecessarily exposing plant protection products to evolving pest pop-ulations HPR (and host plant tolerance) can be effective against target pests as revealedin previous studies [4] and could be incorporated into other pest management practicesparticularly agroecological practices such as ecological engineering with which it is highlycompatible [818586]
5 Conclusions
Based on our results and the results of previous studies HPR and the applicationof chemical insecticides against BPH rarely produce synergistic effects Antagonisticeffects where chemicals caused increases in BPH fitness or populations on resistant ricewere more apparent in our study In particular deltamethrin caused an outbreak ofBPH in our screenhouse study Nevertheless HPR can buffer against the insecticide-induced enhancement of herbivore fitness (hormesis) and potentially reduce possibilities ofsecondary outbreaks as was suggested from results with cartap hydrochloride in our studyBased on our results we suggest that carbofuran cartap hydrochloride cypermethrindeltamethrin and fipronil should be avoided in rice particularly in the Philippines (wherethis research was conducted) due to their resurgence-causing potential or because of directreductions to rice yields
Author Contributions Conceptualization FGH methodology FGH AP-C and MLPA formalanalysis FGH and AP-C investigation FGH AP-C and MLPA data curation FGHAP-C and MLPA writingmdashoriginal draft preparation FGH and AP-C writingmdashreview andediting FGH AP-C and MLPA visualization FGH supervision FGH and MLPA projectadministration FGH funding acquisition FGH All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by the Bill and Melinda Gates Foundation (Cereal SystemsInitiative for South Asia [CSISA] OPP52303) and the Global Rice Science Platform (GRiSP) underthe directorship of Achim Dobermann AP-C was partially funded by The Spanish Governmentthrough Becas Faro International
Crops 2021 1 181
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data presented in this study are available on reasonable requestfrom the corresponding author
Acknowledgments The authors thank Fame Ramal Carmencita Bernal Alberto Naredo ReyeulQuintana Vincent Virtudes Marol Recide Jenyrose Geronda and Ellen Genil for assistance duringthis study The contributions of Rodante Abas (sadly deceased) during three decades of entomologicalresearch at IRRI are gratefully acknowledged Rice lines were supplied through the Genebank of IRRI
Conflicts of Interest The authors declare no conflict of interest
References1 Bottrell DG Schoenly KG Resurrecting the ghost of green revolutions past The brown planthopper as a recurring threat to
high-yielding rice production in tropical Asia J Asia-Pac Entomol 2012 15 122ndash140 [CrossRef]2 Horgan FG Integrated pest management for sustainable rice cultivation A holistic approach In Achieving Sustainable Cultivation
of Rice Cultivation Pest and Disease Management Sasaki T Ed Burleigh Dodds Scientific Cambridge UK 2017 pp 309ndash3423 Horgan FG de Freitas TFS Crisol-Martiacutenez E Mundaca EA Bernal CC Nitrogenous fertilizer reduces resistance but
enhances tolerance to the brown planthopper in fast-growing moderately resistant rice Insects 2021 12 989 [CrossRef] [PubMed]4 Horgan FG Pentildealver Cruz A Bernal CC Ramal AF Almazan MLP Wilby A Resistance and tolerance to the brown
planthopper Nilaparvata lugens (Staringl) in rice infested at different growth stages across a gradient of nitrogen applicationsField Crops Res 2018 217 53ndash65 [CrossRef]
5 Hereward JP Cai X Matias AMA Walter GH Xu C Wang Y Migration dynamics of an important rice pest The brownplanthopper (Nilaparvata lugens) across AsiamdashInsights from population genomics Evol Appl 2020 13 2449ndash2459 [CrossRef]
6 Hu C Guo J Fu X Huang Y Gao X Wu K Seasonal migration pattern of Nilaparvata lugens (Hemiptera Delphacidae) overthe Bohai Sea in northern China J Econ Entomol 2018 111 2129ndash2135 [CrossRef]
7 Ma J Wang Y-C Hu Y-Y Lu M-H Wan G-J Chen F-J Liu W-C Zhai B-P Hu G Brown planthopper Nilaparvatalugens was concentrated at the rear of the typhoon Soudelor in eastern China in August 2015 Insect Sci 2018 25 916ndash926[CrossRef]
8 Fujita D Kohli A Horgan FG Rice resistance to planthoppers and leafhoppers Crit Rev Plant Sci 2013 32 162ndash191[CrossRef]
9 Fujii T Yoshida K Kobayashi T Myint KKM Yasui H Sanada-Morimura S Matsumura M Long-term virulencemonitoring of differential cultivars in Japanrsquos immigrant populations of Nilaparvata lugens (Hemiptera Delphacidae) in 2001ndash2009Appl Entomol Zool 2021 56 407ndash418 [CrossRef]
10 Myint KKM Yasui H Takagi M Matsumura M Virulence of long-term laboratory populations of the brown planthopperNilaparvata lugens (Stal) and whitebacked planthopper Sogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) on rice differentialvarieties Appl Entomol Zool 2009 44 149ndash153 [CrossRef]
11 Horgan FG Ramal AF Bentur JS Kumar R Bhanu KV Sarao PS Iswanto EH Van Chien H Phyu MHBernal CC et al Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia againstresistant rice varieties Crop Prot 2015 78 222ndash231 [CrossRef]
12 Horgan FG Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers Crop Prot2018 110 21ndash33 [CrossRef]
13 Uddin ABMA Islam KS Jahan M Ara A Afrin S Farmerrsquos perception about resurgence of brown planthopper NilaparvataLugens (Staringl) in Bangladesh Bangladesh Rice J 2020 23 21ndash33 [CrossRef]
14 Triwidodo H Brown planthoppers infestations and insecticides use pattern in Java Indonesia AGRIVITA J Agric Sci 202042 320ndash330 [CrossRef]
15 Veres A Wyckhuys KA Kiss J Toacuteth F Burgio G Pons X Avilla C Vidal S Razinger J Bazok R et al An update of theWorldwide Integrated Assessment (WIA) on systemic pesticides Part 4 Alternatives in major cropping systems Environ SciPollut Res 2020 27 29867ndash29899 [CrossRef]
16 Bakker L van der Werf W Tittonell P Wyckhuys KA Bianchi FJ Neonicotinoids in global agriculture Evidence for a newpesticide treadmill Ecol Soc 2020 25 26 [CrossRef]
17 Wu J-C Xu J-X Yuan S-Z Liu J-L Jiang Y-H Xu J-F Pesticide-induced susceptibility of rice to brown planthopperNilaparvata lugens Entomol Exp Appl 2001 100 119ndash126 [CrossRef]
18 Wu J-C Qiu H-M Yang G-Q Liu J-L Liu G-J Wilkens RM Effective duration of pesticide-induced susceptibility of riceto brown planthopper (Nilaparvata lugens Staringl Homoptera Delphacidae) and physiological and biochemical changes in riceplants following pesticide application Int J Pest Manag 2004 50 55ndash62 [CrossRef]
19 Wang A-H Wu J-C Yu Y-S Liu J-L Yue J-F Wang MY Selective insecticide-induced stimulation on fecundity andbiochemical changes in Tryporyza incertulas (Lepidoptera Pyralidae) J Econ Entomol 2005 98 1144ndash1149 [CrossRef] [PubMed]
Crops 2021 1 182
20 Azzam S Wang F Wu J-C Shen J Wang L-P Yang G-Q Guo Y-R Comparisons of stimulatory effects of a seriesof concentrations of four insecticides on reproduction in the rice brown planthopper Nilaparvata lugens Staringl (HomopteraDelphacidae) Int J Pest Manag 2009 55 347ndash358 [CrossRef]
21 Ge L-Q Hu J-H Wu J-C Yang G-Q Gu H Insecticide-induced changes in protein RNA and DNA contents in ovary andfat body of female Nilaparvata lugens (Hemiptera Delphacidae) J Econ Entomol 2009 102 1506ndash1514 [CrossRef]
22 Ling S Zhang J Hu L Zhang R Effect of fipronil on the reproduction feeding and relative fitness of brown planthopperNilaparvata lugens Appl Entomol Zool 2009 44 543ndash548 [CrossRef]
23 Ge L-Q Wang L-P Zhao K-F Wu J-C Huang L-J Mating pair combinations of insecticide-treated male and femaleNilaparvata lugens Staringl (Hemiptera Delphacidae) planthoppers influence protein content in the male accessory glands (MAGs)and vitellin content in both fat bodies and ovaries of adult females Pestic Biochem Physiol 2010 98 279ndash288 [CrossRef]
24 Ge L-Q Wu J-C Zhao K-F Chen Y Yang G-Q Induction of Nlvg and suppression of Nljhe gene expression in Nilaparvatalugens (Staringl) (Hemiptera Delphacidae) adult females and males exposed to two insecticides Pestic Biochem Physiol 201098 269ndash278 [CrossRef]
25 Hu J-H Wu J-C Yin J-L Gu H-N Physiology of insecticide-induced stimulation of reproduction in the rice brownplanthopper (Nilaparvata lugens (Staringl)) Dynamics of protein in fat body and ovary Int J Pest Manag 2010 56 23ndash30 [CrossRef]
26 Wang L-P Shen J Ge L-Q Wu J-C Yang G-Q Jahn GC Insecticide-induced increase in the protein content of maleaccessory glands and its effect on the fecundity of females in the brown planthopper Nilaparvata lugens Staringl (HemipteraDelphacidae) Crop Prot 2010 29 1280ndash1285 [CrossRef]
27 Wu J Ge L Liu F Song Q Stanley D Pesticide-induced planthopper population resurgence in rice cropping systems AnnuRev Entomol 2020 65 409ndash429 [CrossRef]
28 Suri KS Singh G Insecticide-induced resurgence of the whitebacked planthopper Sogatella furcifera (Horvath) (HemipteraDelphacidae) on rice varieties with different levels of resistance Crop Prot 2011 30 118ndash124 [CrossRef]
29 Fabellar LT Heinrichs EA Toxicity of insecticides to predators of rice brown planthoppers Nilaparvata lugens (Staringl) (HomopteraDelphacidae) Environ Entomol 1984 13 832ndash837 [CrossRef]
30 Fabellar LT Heinrichs EA Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators Crop Prot1986 5 254ndash258 [CrossRef]
31 Cuong NL Ben PT Phuong LT Chau LM Cohen MB Effect of host plant resistance and insecticide on brown planthopperNilaparvata lugens (Staringl) and predator population development in the Mekong Delta Vietnam Crop Prot 1997 16 707ndash715[CrossRef]
32 Preetha G Stanley J Suresh S Samiyappan R Risk assessment of insecticides used in rice on mirid bug Cyrtorhinuslividipennis Reuter the important predator of brown planthopper Nilaparvata lugens (Stal) Chemosphere 2010 80 498ndash503[CrossRef] [PubMed]
33 Tanaka K Endo S Kazano H Toxicity of insecticides to predators of rice planthoppers Spiders the mirid bug and the dryinidwasp Appl Entomol Zool 2000 35 177ndash187 [CrossRef]
34 Zhang X Xu Q Lu W Liu F Sublethal effects of four synthetic insecticides on the generalist predator Cyrtorhinus lividipennisJ Pest Sci 2015 88 383ndash392 [CrossRef]
35 Settle WH Ariawan H Astuti ET Cahyana W Hakim AL Hindayana D Lestari AS Managing tropical rice peststhrough conservation of generalist natural enemies and alternative prey Ecology 1996 77 1975ndash1988 [CrossRef]
36 Salim M Heinrichs EA Insecticide-induced changes in the levels of resistance of rice cultivars to the whitebacked planthopperSogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) Crop Prot 1987 6 28ndash32 [CrossRef]
37 Yin J-L Xu H-W Wu J-C Hu J-H Yang G-Q Cultivar and insecticide applications affect the physiological development ofthe brown planthopper Nilaparvata lugens (Staringl) (Hemiptera Delphacidae) Environ Entomol 2014 37 206ndash212 [CrossRef]
38 Cheng C Chang W Studies on varietal resistance to the brown planthopper in Taiwan In Brown Planthopper Threat toRice Production in Asia International Rice Research Institute Ed International Rice Research Institute Manila Philippines1979 pp 251ndash271
39 Reissig WH Heinrichs EA Valencia SL Insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on ricevarieties with different levels of resistance Environ Entomol 1982 11 165ndash168 [CrossRef]
40 Heinrichs EA Fabellar LT Basilio RP Wen T-C Medrano F Susceptibility of rice planthoppers Nilaparvata lugens andSogatella furcifera (Homoptera Delphacidae) to insecticides as influenced by level of resistance in the host plant Environ Entomol1984 13 455ndash458 [CrossRef]
41 Joshi R Shepard B Kenmore P Lydia R Insecticide-induced resurgence of brown planthopper (BPH) on IR62 Int Rice ResNewsl 1992 17 9ndash10
42 Zhu Z-R Cheng J Jiang M-X Zhang X-X Complex influence of rice variety fertilization timing and insecticide onpopulation dynamics of Sogatella furcifera (Horvath) Nilaparvata lugens (Staringl) (Homoptera Delphacidae) and their natural enemiesin rice in Hangzhou China J Pest Sci 2004 77 65ndash74 [CrossRef]
43 Nanthakumar M Jhansi Lakshmi V Shashi Bhushan V Balachandran SM Mohan M Decrease of rice plant resistance andinduction of hormesis and carboxylesterase titre in brown planthopper Nilaparvata lugens (Staringl) by xenobiotics Pestic BiochemPhysiol 2012 102 146ndash152 [CrossRef]
Crops 2021 1 183
44 Yao H-W Jiang C-Y Ye G-Y Cheng J-A Studies on cultivar-induced changes in insecticide susceptibility and its relatedenzyme activities of the white-backed planthopper Sogatella furcifera (Horvath)(Homopter Delphacidae) Insect Sci 2002 9 9ndash15
45 Horgan FG Srinivasan TS Bentur JS Kumar R Bhanu KV Sarao PS Van Chien H Almazan MLP Bernal CCRamal AF et al Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers Implicationsfor rice breeding and gene deployment Agronomy 2017 7 62 [CrossRef]
46 Jairin J Phengrat K Teangdeerith S Vanavichit A Toojinda T Mapping of a broad-spectrum brown planthopper resistancegene Bph3 on rice chromosome 6 Mol Breed 2007 19 35ndash44 [CrossRef]
47 Ren J Gao F Wu X Lu X Zeng L Lv J Su X Luo H Ren G Bph32 a novel gene encoding an unknown SCRdomain-containing protein confers resistance against the brown planthopper in rice Sci Rep 2016 6 37645 [CrossRef]
48 Nguyen CD Zheng S-H Sanada-Morimura S Matsumura M Yasui H Fujita D Substitution mapping and characterizationof brown planthopper resistance genes from indica rice variety lsquoPTB33prime (Oryza sativa L) Breed Sci 2021 in press [CrossRef]
49 Chelliah S Heinrichs EA Factors affecting insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on RiceEnviron Entomol 1980 9 773ndash777 [CrossRef]
50 Moore MT Kroumlger R Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growthArch Environ Contam Toxicol 2010 59 574ndash581 [CrossRef]
51 Kim J Hwang I Han M Jang B Effect of insecticide and fungicide on phytotoxicity of herbicide in rice Korean J Weed Sci1986 6 67ndash75
52 Horgan FG Garcia CPF Haverkort F de Jong PW Ferrater JB Changes in insecticide resistance and host range performanceof planthoppers artificially selected to feed on resistant rice Crop Prot 2020 127 104963 [CrossRef] [PubMed]
53 Khush GS Virk PS IR Varieties and Their Impact International Rice Research Institute Manila Philippines 200554 Srinivasan TS Almazan MLP Bernal CC Ramal AF Subbarayalu MK Horgan FG Interactions between nymphs of
Nilaparvata lugens and Sogatella furcifera (Hemiptera Delphacidae) on resistant and susceptible rice varieties Appl Entomol Zool2016 51 81ndash90 [CrossRef]
55 Pentildealver Cruz A Arida A Heong KL Horgan FG Aspects of brown planthopper adaptation to resistant rice varieties withthe Bph3 gene Entomol Exp Appl 2011 141 245ndash257 [CrossRef]
56 Alam SN Cohen MB Detection and analysis of QTLs for resistance to the brown planthopper Nilaparvata lugens in adoubled-haploid rice population Theor Appl Genet 1998 97 1370ndash1379 [CrossRef]
57 Mackill DJ Khush GS IR64 A high-quality and high-yielding mega variety Rice 2018 11 18 [CrossRef] [PubMed]58 Heinrichs EA Basilio RP Valencia SL Buprofezin a selective insecticide for the management of rice planthoppers (Ho-
moptera Delphacidae) and leafhoppers (Homoptera Cicadellidae) Environ Entomol 1984 13 515ndash521 [CrossRef]59 Satapathy MK Anjaneyulu A Use of cypermethrin a synthetic pyrethroid in the control of rice tungro virus disease and its
vector Trop Pest Manag 1984 30 170ndash178 [CrossRef]60 Vorley W Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown
planthopper (BPH) in Kedah Malaysia Int Rice Res Newsl 1987 10 19ndash2061 Uddin A Islam KS Jahan M Ara A Khan MAI Factor influencing the resurgence of brown planthopper in Bangladesh
SAARC J Agric 2020 18 117ndash128 [CrossRef]62 Escalada MM Luecha M Heong KL Social impacts of planthopper outbreaks Case study from Thailand In Rice Planthoppers
Ecology Management Socio Economics and Policy Heong KL Cheng J Escalada MM Eds Zhejiang University PressHangzhou China 2015 pp 209ndash226
63 Zhang CP He HM Yu JZ Hu XQ Zhu YH Wang Q Residues of carbosulfan and its metabolites carbofuran and3-hydroxy carbofuran in rice field ecosystem in China J Environ Sci Health 2016 51 351ndash357 [CrossRef] [PubMed]
64 Samanta S Barman M Samanta NRA Bio-efficacy trials of carbofuran 3 CG against insect pests of rice J Entomol ZoolStud 2020 8 1529ndash1533
65 Donley N The USA lags behind other agricultural nations in banning harmful pesticides Environ Health 2019 18 44 [CrossRef][PubMed]
66 Adams A Gore J Musser F Cook D Catchot A Walker T Dobbins C Efficacy of selected insecticides applied to hybridrice seed J Econ Entomol 2015 109 200ndash206 [CrossRef]
67 Kumar R Singh B Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) Field Bull Environ Contam Toxicol2013 90 482ndash488 [CrossRef]
68 Dhaka S Prajapati C Singh D Singh R Field evaluation of insecticides and bio-pesticides against rice leaf folder CnaphalocrocisMedinalis (Guenee) Ann Plant Prot Sci 2011 19 324ndash326
69 Matsumura M Takeuchi H Satoh M Sanada-Morimura S Otuka A Watanabe T Van Thanh D Species-specific insecticideresistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in East and South-eastAsia Pest Manag Sci 2008 64 1115ndash1121 [CrossRef] [PubMed]
70 Tang J Li J Shao Y Yang B Liu Z Fipronil resistance in the whitebacked planthopper (Sogatella furcifera) Possible resistancemechanisms and cross-resistance Pest Manag Sci 2010 66 121ndash125 [CrossRef]
71 Horgan FG Arida A Ardestani G Almazan MLP Temperature-dependent oviposition and nymph performance revealdistinct thermal niches of coexisting planthoppers with similar thresholds for development PLoS ONE 2020 15 e0235506[CrossRef] [PubMed]
Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]
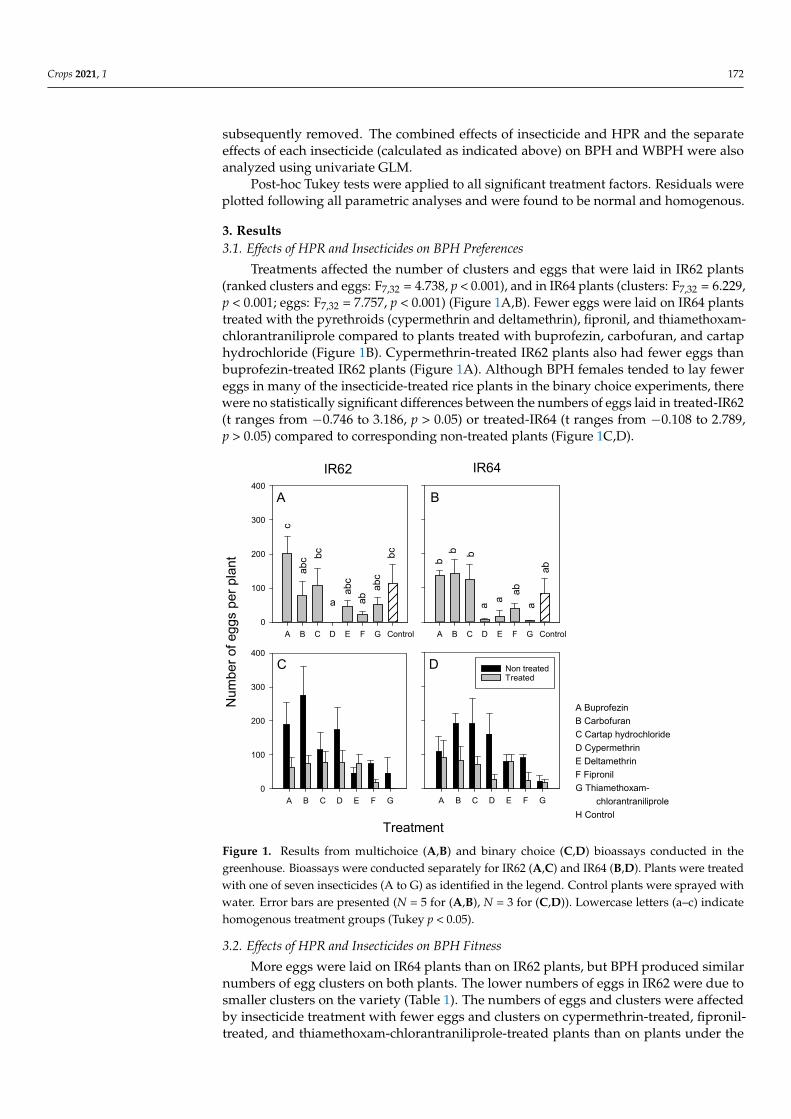
Crops 2021 1 172
subsequently removed The combined effects of insecticide and HPR and the separateeffects of each insecticide (calculated as indicated above) on BPH and WBPH were alsoanalyzed using univariate GLM
Post-hoc Tukey tests were applied to all significant treatment factors Residuals wereplotted following all parametric analyses and were found to be normal and homogenous
3 Results31 Effects of HPR and Insecticides on BPH Preferences
Treatments affected the number of clusters and eggs that were laid in IR62 plants(ranked clusters and eggs F732 = 4738 p lt 0001) and in IR64 plants (clusters F732 = 6229p lt 0001 eggs F732 = 7757 p lt 0001) (Figure 1AB) Fewer eggs were laid on IR64 plantstreated with the pyrethroids (cypermethrin and deltamethrin) fipronil and thiamethoxam-chlorantraniliprole compared to plants treated with buprofezin carbofuran and cartaphydrochloride (Figure 1B) Cypermethrin-treated IR62 plants also had fewer eggs thanbuprofezin-treated IR62 plants (Figure 1A) Although BPH females tended to lay fewereggs in many of the insecticide-treated rice plants in the binary choice experiments therewere no statistically significant differences between the numbers of eggs laid in treated-IR62(t ranges from minus0746 to 3186 p gt 005) or treated-IR64 (t ranges from minus0108 to 2789p gt 005) compared to corresponding non-treated plants (Figure 1CD)
Crops 2021 1 FOR PEER REVIEW 7
herbivore densities Total planthopper and leafhopper biomass and density and plant growth parameters (tiller number biomass yield grain number proportion of grain un-filled and grain size) were analyzed using split-plot univariate GLMs The covariate plant biomass was initially included in analyses of the effects on herbivores but had no effect and was subsequently removed Similarly the covariate herbivore biomass was initially included in analyses of plant growth parameters but had no effect and was subsequently removed The combined effects of insecticide and HPR and the separate effects of each insecticide (calculated as indicated above) on BPH and WBPH were also analyzed using univariate GLM
Post-hoc Tukey tests were applied to all significant treatment factors Residuals were plotted following all parametric analyses and were found to be normal and homogenous
3 Results 31 Effects of HPR and Insecticides on BPH Preferences
Treatments affected the number of clusters and eggs that were laid in IR62 plants (ranked clusters and eggs F732 = 4738 p lt 0001) and in IR64 plants (clusters F732 = 6229 p lt 0001 eggs F732 = 7757 p lt 0001) (Figure 1AB) Fewer eggs were laid on IR64 plants treated with the pyrethroids (cypermethrin and deltamethrin) fipronil and thiameth-oxam-chlorantraniliprole compared to plants treated with buprofezin carbofuran and cartap hydrochloride (Figure 1B) Cypermethrin-treated IR62 plants also had fewer eggs than buprofezin-treated IR62 plants (Figure 1A) Although BPH females tended to lay fewer eggs in many of the insecticide-treated rice plants in the binary choice experiments there were no statistically significant differences between the numbers of eggs laid in treated-IR62 (t ranges from minus0746 to 3186 p gt 005) or treated-IR64 (t ranges from minus0108 to 2789 p gt 005) compared to corresponding non-treated plants (Figure 1CD)
Figure 1 Results from multichoice (AB) and binary choice (CD) bioassays conducted in the green-house Bioassays were conducted separately for IR62 (AC) and IR64 (BD) Plants were treated with one of seven insecticides (A to G) as identified in the legend Control plants were sprayed with water Error bars are presented (N = 5 for (A) and (B) N = 3 for (C) and (D)) Lowercase letters (andashc) indicate homogenous treatment groups (Tukey p lt 005)
A B C D E F G0
100
200
300
400
Treatment
A B C D E F G
Non treatedTreated
A B C D E F G Control
Num
ber o
f egg
s pe
r pla
nt
0
100
200
300
400
A B C D E F G Control
IR62 IR64
A
C
B
D
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliproleH Control
a abab
c
abc
abc bc bc
c
aaa
ab
ab
bb b
Figure 1 Results from multichoice (AB) and binary choice (CD) bioassays conducted in thegreenhouse Bioassays were conducted separately for IR62 (AC) and IR64 (BD) Plants were treatedwith one of seven insecticides (A to G) as identified in the legend Control plants were sprayed withwater Error bars are presented (N = 5 for (AB) N = 3 for (CD)) Lowercase letters (andashc) indicatehomogenous treatment groups (Tukey p lt 005)
32 Effects of HPR and Insecticides on BPH Fitness
More eggs were laid on IR64 plants than on IR62 plants but BPH produced similarnumbers of egg clusters on both plants The lower numbers of eggs in IR62 were due tosmaller clusters on the variety (Table 1) The numbers of eggs and clusters were affectedby insecticide treatment with fewer eggs and clusters on cypermethrin-treated fipronil-treated and thiamethoxam-chlorantraniliprole-treated plants than on plants under the
Crops 2021 1 173
remaining treatments (Table 1) The treatment-effects were not affected by host variety (ieinteractions were non-significant Table 1) Insecticide treatments had no effect on clustersize in the bioassay (Table 1) There was no significant effect of combined insecticide andHPR (F628 = 0449 p = 0840) on observed changes in egg laying during the bioassay butthe estimated insecticide effect was greater for thiamethoxam-chlorantraniliprole fiproniland cypermethrin than for buprofezin cartap hydrochloride and carbofuran (F628 = 3140p = 0018 Figure 2A)
Table 1 Results from greenhouse oviposition and survival tests with IR62 (resistant) and IR64 (susceptible) each treatedwith one of seven insecticides Numbers are means plusmn SEM (N = 5)
Treatments Egg Clusters(Number Plantminus1) 12
Eggs(Number Plantminus1) 12
Cluster Size(Number of Eggs) 1
Nymph Survival(Proportions) 12
Nymph Biomass (mgDry Weight Plantminus1) 12
IR62Buprofezin 1240 plusmn 140 b 5040 plusmn 904 b 393 plusmn 036 000 plusmn 000 a 000 plusmn 000 a
Carbofuran 1300 plusmn 089 b 5380 plusmn 421 b 417 plusmn 026 064 plusmn 064 d 154 plusmn 154 d
Cartaphydrochloride 1360 plusmn 075 b 5200 plusmn 486 b 382 plusmn 029 028 plusmn 028 b 151 plusmn 151 b
Cypermethrin 620 plusmn 169 a 2000 plusmn 608 a 316 plusmn 023 002 plusmn 002 a 002 plusmn 002 a
Deltamethrin 1360 plusmn 160 b 4340 plusmn 566 b 338 plusmn 069 040 plusmn 040 c 120 plusmn 120 c
Fipronil 520 plusmn 146 a 2240 plusmn 735 a 393 plusmn 064 000 plusmn 000 a 000 plusmn 000 a
Thiamethoxam-chlorantraniliprole 380 plusmn 086 a 1700 plusmn 435 a 447 plusmn 032 000 plusmn 000 a 000 plusmn 000 a
Control 1260 plusmn 103 b 4900 plusmn 716 b 388 plusmn 054 080 plusmn 080 d 292 plusmn 292 d
IR64Buprofezin 2100 plusmn 164 10940 plusmn 1346 556 plusmn 117 000 plusmn 000 000 plusmn 000Carbofuran 1820 plusmn 404 9880 plusmn 2939 524 plusmn 074 086 plusmn 086 420 plusmn 420
Cartaphydrochloride 2140 plusmn 619 11460 plusmn 3141 549 plusmn 064 010 plusmn 010 045 plusmn 045
Cypermethrin 380 plusmn 233 1980 plusmn 1155 550 plusmn 058 000 plusmn 000 000 plusmn 000Deltamethrin 1560 plusmn 209 7500 plusmn 1581 473 plusmn 069 040 plusmn 040 201 plusmn 201
Fipronil 380 plusmn 086 2100 plusmn 552 546 plusmn 080 000 plusmn 000 000 plusmn 000Thiamethoxam-chlorantraniliprole 260 plusmn 075 800 plusmn 212 373 plusmn 133 000 plusmn 000 000 plusmn 000
Control 1740 plusmn 364 7520 plusmn 2344 426 plusmn 073 086 plusmn 086 360 plusmn 360F-treatment 3 19898 19898 0541 ns 133811 108451
F-variety 3 2596 ns 7607 7471 1080 ns 0008 ns
F-interaction 3 1840 ns 2034 ns 0610 ns 4902 9496 1 Lowercase letters (andashd) indicate homogenous treatment groups (for IR62 and IR64 combined) 2 Data were ranked within blocks beforeanalysis 3 DF = 164 for varieties 764 for treatments and interactions ns = p gt 005 = p le 001 = p le 0001
Nymph survival and biomass were not affected by rice variety (Table 1) Buprofezinfipronil thiamethoxam-chlorantraniliprole and cypermethrin reduced survival to 0 in thebioassay and all insecticides except carbofuran resulted in lower survival and biomass thanon untreated plants (Table 1) There were significant interactions because of higher nymphmortality and a resulting lower biomass on carbofuran-treated IR62 compared to untreatedIR62 but lower mortality and a higher biomass of nymphs on carbofuran-treated IR64than on untreated IR64 (Table 1) There was no significant effect of insecticide treatment(F628 = 0946 p = 0479) on observed changes in nymph biomass during the bioassaybut the estimated combined effects of pesticide and HPR was greater for thiamethoxam-chlorantraniliprole fipronil cypermethrin and buprofezin than for cartap hydrochlorideand carbofuran (F628 = 5699 p = 0001) (Figure 2B)
Crops 2021 1 174
Crops 2021 1 FOR PEER REVIEW 9
Figure 2 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) egg laying and (B) nymph biomass in greenhouse bioas-says Effects are indicated as proportional changes in fitness parameters compared to untreated IR64 Negative numbers indicate reduced fitness Standard errors are indicated (N = 5) Lowercase letters (ab) indicated homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 1)
Nymph survival and biomass were not affected by rice variety (Table 1) Buprofezin fipronil thiamethoxam-chlorantraniliprole and cypermethrin reduced survival to 0 in the bioassay and all insecticides except carbofuran resulted in lower survival and biomass than on untreated plants (Table 1) There were significant interactions because of higher nymph mortality and a resulting lower biomass on carbofuran-treated IR62 compared to untreated IR62 but lower mortality and a higher biomass of nymphs on carbofuran-treated IR64 than on untreated IR64 (Table 1) There was no significant effect of insecticide treatment (F628 = 0946 p = 0479) on observed changes in nymph biomass during the bio-assay but the estimated combined effects of pesticide and HPR was greater for thiameth-oxam-chlorantraniliprole fipronil cypermethrin and buprofezin than for cartap hydro-chloride and carbofuran (F628 = 5699 p = 0001) (Figure 2B)
33 Effects of HPR and Insecticides on Planthoppers and Leafhoppers Plants in the screenhouse were colonized by BPH WBPH and GLH Densities for
each species were similar on IR62 and IR64 (Wilkrsquos λ = 0994 p = 0952) There was a sig-nificant treatment effect on all three species (Wilkrsquos λ = 0586 p = 0049) because of high densities on deltamethrin-treated plants (Table 2) The numbers and biomass of all three species combined were also greater on deltamethrin-treated plants than on the remaining treatments (Table 2) Host resistance reduced GLH densities under the deltamethrin treat-ment but had no apparent effect on densities under the other treatments (Table 2) There were significant combined effects of HPR and insecticide on the numbers of BPH (F624 = 2686 p = 0039) and WBPH (F624 = 2482 p = 0052) this was almost exclusively due to the resurgence effect of deltamethrin on the planthoppers (BPH F624 = 2686 p = 0039 WBPH F624 = 2482 p = 0052) (Figure 3)
Δ eg
gs(P
ropo
rtion
)
-10
-08
-06
-04
-02
00
02
04
06
Treatment
A B C D E F G
Δ N
ymph
bio
mas
s(P
ropo
rtion
)
-10
-08
-06
-04
-02
00
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
ab
a a
b
a
b b
a a a a a a a a
b b bb
bb
Figure 2 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone(P grey bars) on (A) egg laying and (B) nymph biomass in greenhouse bioassays Effects are indicated as proportionalchanges in fitness parameters compared to untreated IR64 Negative numbers indicate reduced fitness Standard errors areindicated (N = 5) Lowercase letters (ab) indicated homogenous treatment groups based on Tukey pairwise comparisons(p lt 005) (see also Table 1)
33 Effects of HPR and Insecticides on Planthoppers and Leafhoppers
Plants in the screenhouse were colonized by BPH WBPH and GLH Densities foreach species were similar on IR62 and IR64 (Wilkrsquos λ = 0994 p = 0952) There was asignificant treatment effect on all three species (Wilkrsquos λ = 0586 p = 0049) because of highdensities on deltamethrin-treated plants (Table 2) The numbers and biomass of all threespecies combined were also greater on deltamethrin-treated plants than on the remainingtreatments (Table 2) Host resistance reduced GLH densities under the deltamethrintreatment but had no apparent effect on densities under the other treatments (Table 2)There were significant combined effects of HPR and insecticide on the numbers of BPH(F624 = 2686 p = 0039) and WBPH (F624 = 2482 p = 0052) this was almost exclusively dueto the resurgence effect of deltamethrin on the planthoppers (BPH F624 = 2686 p = 0039WBPH F624 = 2482 p = 0052) (Figure 3)
Crops 2021 1 175
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants underseven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density(Number Plantminus1) 1
WBPH Density(Number Plantminus1) 1
GLH Density(Number Plantminus1) 1
Density ofPlanthoppers and
Leafhoppers(Number Plantminus1) 1
Biomass ofPlanthoppers and
Leafhoppers(mg Dry Weight) 1
IR62Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a
Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartaphydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a
Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a
Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a
Thiamethoxam-chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a
IR64Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartaphydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123
Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns
F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Crops 2021 1 FOR PEER REVIEW 10
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants under seven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density (Num-
ber Plantminus1) 1 WBPH Density
(Number Plantminus1) 1 GLH Density
(Number Plantminus1) 1
Density of Planthoppers and Leafhoppers (Number
Plantminus1) 1
Biomass of Planthoppers and Leafhoppers (mg Dry
Weight) 1 IR62
Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartap hydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a Thiamethoxam-
chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a IR64
Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132 Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartap hydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123 Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113 Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052 Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087 F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns 1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatments and interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in the screenhouse Effects are indicated as proportional changes in planthopper densities compared to untreated IR64 Negative numbers indicate reduced densities positive numbers are increased den-sities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
Δ BP
H(N
umbe
r pla
nt-1
)
0
5
10
15
20
Treatment
A B C D E F G
Δ W
BPH
(Num
ber p
lant
-1)
0
20
30
40
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
a a
aa a a
a a a
a
a a
b
b
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimatedpesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in thescreenhouse Effects are indicated as proportional changes in planthopper densities compared to
Crops 2021 1 176
untreated IR64 Negative numbers indicate reduced densities positive numbers are increaseddensities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenoustreatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
34 Direct and Indirect Effects of Insecticides on Rice Development
IR62 plants had more tillers and attained a greater biomass than IR64 plants in thescreenhouse (Table 3) IR62 grain matured faster and plants were harvested earlier than theIR64 plants (Table 3) IR62 plants also produced more grain (number of grain) and hadhigher yields than IR64 but had smaller grain and more unfilled grain than IR64 (Table 3)
Table 3 Rice growth parameters for IR62 (resistant) and IR64 (susceptible) under seven insecticide treatments and forcontrol non-treated plants in a screenhouse environment Numbers are means plusmn SEM
Treatment Tiller Number 1 Plant Biomass(g Dry Weight) 1
Time to Harvest(Days) 1
Weightof Filled Grain(g Dry Weight) 1
Number ofFilled Grain
(1000s) 1
Unfilled Grain(Proportion) 1
1000 GrainWeight 1
IR62Buprofezin 2660 plusmn 206 ab 4643 plusmn 385 ab 7990 plusmn 168 ab 5776 plusmn 683 ab 361plusmn 061 ab 011 plusmn 003 a 2008 plusmn 042 ab
Carbofuran 2900 plusmn 106 ab 5546 plusmn 357 b 7720 plusmn 080 ab 7448 plusmn 691 b 446plusmn 061 ab 011 plusmn 001 a 2103 plusmn 054 b
Cartaphydrochloride 2820 plusmn 327 ab 4896 plusmn 470 ab 7920 plusmn 153 b 6430 plusmn 712 ab 415plusmn 064 ab 012 plusmn 001 b 2023 plusmn 065 a
Cypermethrin 2870 plusmn 301 a 4957 plusmn 562 a 7760 plusmn 103 b 6032 plusmn 822 a 410plusmn 067 ab 014 plusmn 002 a 1965 plusmn 076 a
Deltamethrin 2620 plusmn 345 a 4592 plusmn 588 a 7840 plusmn 108 ab 5698 plusmn 832 ab 379plusmn 047 ab 012 plusmn 001 a 2030 plusmn 063 ab
Fipronil 2380 plusmn 176 ab 4607 plusmn 657 ab 7700 plusmn 179 ab 5266 plusmn 390 a 318 plusmn 032 a 013 plusmn 002 a 2128 plusmn 047 b
Thiamethoxam-chlorantraniliprole 2650 plusmn 250 b 4547 plusmn 429 b 7720 plusmn 102 a 5777 plusmn 688 ab 408 plusmn 061 b 013 plusmn 002 a 2019 plusmn 067 ab
Control 3120 plusmn 247 b 5401 plusmn 555 b 7720 plusmn 080 ab 7045 plusmn 520 b 396plusmn 036 ab 012 plusmn 002 a 2076 plusmn 050 ab
IR64Buprofezin 2340 plusmn 250 5064 plusmn 747 8350 plusmn 124 5080 plusmn 952 270 plusmn 054 008 plusmn 001 2277 plusmn 039Carbofuran 2040 plusmn 097 4591 plusmn 141 8540 plusmn 073 4951 plusmn 312 263 plusmn 023 006 plusmn 000 2295 plusmn 036
Cartaphydrochloride 2120 plusmn 075 4336 plusmn 285 8520 plusmn 140 4273 plusmn 579 238 plusmn 026 015 plusmn 007 2050 plusmn 234
Cypermethrin 1990 plusmn 089 3968 plusmn 272 8710 plusmn 135 3920 plusmn 552 250 plusmn 028 008 plusmn 002 2171 plusmn 070Deltamethrin 1980 plusmn 181 4325 plusmn 357 8460 plusmn 068 4710 plusmn 496 277 plusmn 024 008 plusmn 001 2314 plusmn 012
Fipronil 2120 plusmn 174 4697 plusmn 311 8450 plusmn 089 4888 plusmn 392 274 plusmn 027 008 plusmn 001 2247 plusmn 042Thiamethoxam-chlorantraniliprole 2520 plusmn 287 5404 plusmn 465 8380 plusmn 196 5594 plusmn 546 347 plusmn 053 010 plusmn 001 2265 plusmn 072
Control 2160 plusmn 076 4678 plusmn 342 8600 plusmn 096 4940 plusmn 543 267 plusmn 029 008 plusmn 001 2283 plusmn 049F-treatment 4100 ns 3410 2253 ns 5150 2555 3923 4924
F-variety 168232 7665 547500 104356 118242 32308 106597 F-interaction 5980 6113 4824 5760 2921 2769 2552
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Insecticides affected plant growth directly and indirectly (through increased planthop-per densities) Plants treated with cypermethrin and deltamethrin had fewer tillers had alower biomass and under cypermethrin had lower yields than untreated plants (Table 3)Plants treated with fipronil also had reduced yields due to lower grain numbers comparedto untreated plants (Table 3) Although yields were not affected treatment with cartaphydrochloride increased the proportion of unfilled grain compared to untreated plantsof both varieties Cartap hydrochloride and cypermethrin also caused a delay in grainmaturation on IR62 and IR64 respectively relative to thiamethoxam-chlorantraniliproletreated plants (Table 3)
There were several significant interactions tiller numbers on both plants were similarunder buprofezin fipronil and thiamethoxam-chlorantraniliprole treatments but wereotherwise higher on IR62 (Table 3) biomass was lower on IR64 under carbofuran andcypermethrin treatments but was otherwise similar on both plants (Table 3) buprofezindelayed grain maturation on IR62 but maturation was otherwise faster on IR62 comparedto IR64 (Table 3) yields under buprofezin fipronil and thiamethoxam-chlorantraniliprolewere similar for IR62 and IR64 but otherwise yields were higher on IR62 (Table 3) grain
Crops 2021 1 177
numbers were higher on IR62 than IR64 except under fipronil treatments (Table 3) theproportions of grain unfilled were higher on IR62 but were similar between varieties underthe fipronil treatment (Table 3) 1000-grain weight was higher in IR64 but similar on bothvarieties when treated with cartap hydrochloride (Table 3)
4 Discussion
Our results using a highly resistant rice variety and some commonly used insecticidesindicate a range of possible effects on planthoppers and rice plants by combining these twomanagement practices
41 Synergies
The most predictable effect of combining HPR and insecticides is a synergy in reduc-ing BPH numbers This is suggested because planthoppers are weakened by the hostrsquosresistance and should therefore be more susceptible to the toxic effects of insecticides [40]Using spray towers over potted plants Heinrichs et al (1984) [40] examined the combinedeffects of a range of insecticides (that included carbofuran) with resistance by comparingBPH mortality on six resistant and two susceptible rice varieties Their results indicatedthat mortality was greatest when insecticides were applied to BPH on resistant varietiesThe effects were similar when the study was repeated with WBPH for three insecticides(including carbofuran) on the resistant variety N22 (Wbph1 gene) [40] In our study hostresistance reduced egg laying and nymph biomass on non-treated IR62 compared to non-treated IR64 Egg laying was also reduced on plants treated with cypermethrin fipronilor thiamethoxam-chlorantraniliprole (Table 1) Based on the results of our choice andno-choice oviposition bioassays cypermethrin acted mainly as a deterrent of gravid fe-males whereas fipronil and thiamethoxam-chlorantraniliprole directly reduced egg layingDespite similar BPH responses to both the insecticides and resistant host there were no ap-parent synergies between any of these insecticides and resistance in reducing egg numbersin the oviposition bioassay In contrast to these three insecticides our bioassays indicatedthat buprofezin carbofuran and cartap hydrochloride can lead to higher numbers of eggson treated plants compared to untreated plants (Table 1) These effects were not statisticallysignificant and may be exaggerated because bioassays were conducted over relativelyshort time scales For example planthoppers will sometimes respond to stresses such ashigh temperatures by increasing oviposition rates [7172] Furthermore the stimulation ofegg-laying by these chemicals was not apparent in our screenhouse experiment (Table 2)Nevertheless our results do indicate that host resistance reduced the magnitude of anystimulatory effect on oviposition egg numbers on buprofezin-treated carbofuran-treatedand cartap hydrochloride-treated IR64 in the no-choice bioassay were twice as high as onthe corresponding IR62 plants In cases such as these the resistance of IR62 functions as abuffer or lsquoinsurancersquo against possible undesired pesticide effects [1238]
Although buprofezin appeared to stimulate oviposition the growth hormone had astrong impact on nymph development leading to zero survival of exposed nymphs in ourgreenhouse bioassay Cypermethrin fipronil and thiamethoxam-chlorantraniliprole alsoreduced nymph survival to zero in the greenhouse bioassay (Table 1) Where insecticideshave such high efficiencies any fitness losses to BPH due to host resistance then becomeredundant Unlike buprofezin carbofuran also stimulated nymph biomass on IR64 in thesurvival bioassay (Table 1) however nymph biomass was lower on carbofuran-treated IR62than on non-treated IR62 indicating that the hostrsquos resistance buffered against undesiredstimulatory effects on BPH and may have functioned synergistically to augment mortalityand slow BPH growth Such buffering allows insecticides to be used against other pestswithout inducing secondary outbreaks of non-target pests In our screenhouse experimentherbivore densities were generally low on both IR62 and IR64 for most treatments (exceptdeltamethrin) and were below threshold densities for insecticide applicationsmdashwhichare normally about 25ndash50 planthoppers per hill [1373] Under such low densities it wasnot possible to observe the impact of host resistance on planthopper population build-
Crops 2021 1 178
up There was some indication of higher densities on cartap hydrochloride-treated IR62in the screenhouse which may be related to the stimulatory effects of the product onegg laying and the low mortality of nymphs on cartap hydrochloride-treated plants asobserved in the greenhouse bioassays (Table 1) Field studies from the Philippines haveindicated that IR62 effectively reduces BPH densities during wet and dry seasons [4]However farmers are rarely aware of which anti-herbivore resistance their rice varietiespossess and frequently apply pesticides regardless of resistance thereby reducing theirfarm profits [3841] For example Joshi et al (1992) [41] reported widespread outbreaks ofBPH on IR62 in Mindanao Philippines after applications of decamethrin (a pyrethroid) tothe resistant variety Where such resurgence-causing insecticides are applied resistanceseems to be often incapable of reducing the stimulatory effects of the insecticide and itloses its capacity to buffer against outbreaks
42 Antagonisms
Several insecticides are associated with increasing densities of planthoppers andwith resurgence outbreaks in farmersrsquo fields [27293041496174] Indeed patches oflsquohopperburnrsquo (ie dead rice plants resulting from planthopper damage) are often anindicator of high insecticide use in tropical Asia [214] In many cases these resurgence-causing insecticides were once effective in reducing planthopper densities but over timepopulations developed resistance against the products The resulting low toxicity canthen have a stimulatory effect on planthopper feeding and development [2123242743]This may be through hormesis which is a direct response by the planthopper to low-levelexposure to the toxic chemical or it may result from physiological changes to the rice plantsthat ultimately benefit the planthopper [1849] In our screenhouse study we induced aresurgence outbreak of BPH WBPH and to a lesser extent GLH on deltamethrin-treatedrice plants (Table 2) The combined densities of all three herbivores increased almost10 times on the treated plants Furthermore herbivore biomass increased by between fourand six times Although the resurgence effects of deltamethrin have been documentedin previous studies [2029ndash313975] we could not predict from our greenhouse studiesthat any such outbreak would occur Egg laying was unaffected by deltamethrin in ourchoice and no-choice experiments and nymph survival and development both declinedon deltamethrin-treated rice in the survival bioassay (Table 1) The massive increases inplanthopper densities on deltamethrin-treated plants in our study seem to result froma heightened attraction of adult BPH and WBPH to the plants [49] particularly sincenumbers were so low on all other plants including both treated and non-treated plantsHowever once on the plants planthoppers also apparently had enhanced fitness Cuonget al (1997) [31] observed higher population densities of BPH on deltamethrin-treatedrice in Vietnam albeit with lower-level increases on resistant varieties Furthermore in agreenhouse study Azzam et al (2009) [20] found that deltamethrin reduced pre-ovipositiontimes increased the duration of oviposition and increased egg hatchability and femalelongevity Nanthakumar et al (2012) [43] further showed that deltamethrin increased BPHfeeding efficiency on resistant and susceptible rice plants as determined through honeydewproduction and population size In studies with WBPH Salim and Heinrichs (1987) [36]showed deltamethrin to increase settling (antixenosis) feeding and population growthand Suri and Singh (2011) [28] showed deltamethrin to increase WBPH reproductive ratesand nymph survival In our study the observed resurgence of planthoppers was similaron both varieties indicating as was previously noted by Joshi et al (1992) [41] thatresurgence-causing pyrethroids are antagonistic to resistance in IR62
Though not statistically significant cartap hydrochloride also increased BPH numberson IR62 but not on IR64 in our screenhouse study This indicates that the product mayalso be antagonistic to the hostrsquos resistance mechanisms The stronger effect on IR62 maybe related to the differences between IR62 biomass growth rates and anatomy (ie alarger plant with higher tillering) compared to IR64 (Table 3) Once resistance is no longereffective because of chemically-enhanced BPH fitness these plant characteristics which
Crops 2021 1 179
have been related to observed preferences in BPH and other sap-sucking insects [349]could further accelerate the build-up of BPH populations on cartap hydrochloride-treatedplants To further test this idea experiments could be conducted using near-isogenic ricelines that differ mainly in the presence or absence of tagged resistance genes [76] Suchtests would avoid confounding differences in plant anatomy between test varieties IR62resistance to BPH has been well studied [4] The variety is also resistant to WBPH atolder plant stages [54] Therefore in our study deltamethrin was also antagonistic tothis WBPH-resistance (Table 2) However BPH feeding also facilitates WBPH populationgrowth [5477] and there may have been added indirect plant-mediated effects [77] thataugmented WBPH populations on both varieties in our experiment IR62 is also resistant toGLH [45] In our experiments GLH densities also increased on deltamethrin-treated plantsbut the effect was only apparent on IR64 Therefore GLH-resistance in IR62 apparentlybuffered against deltamethrin-associated increases in GLH on the variety as was alsonoted from field studies in Vietnam [31] Deltamethrin is currently not recommended foruse in rice [202878] however the product continues to be marketed in rice-dominatedregions throughout Asia [131479] Although cypermethrin has been associated with BPHoutbreaks and was consequently removed for use on rice in Thailand and Vietnam [62]we did not observe cypermethrin-induced resurgence in our screenhouse experimentNevertheless cypermethrin has been associated with resurgence of BPH in South Asia [61]and we have noted stimulatory effects of cypermethrin on other rice herbivores in thePhilippines (Horgan unpublished data)
43 Phytotoxicity
Phytotoxicity is relatively common following applications of insecticides [182850]Phytotoxic effects may change a plantrsquos physiology without consequences for crop yieldsHowever such changes to plant physiology could have indirect effects on herbivores Forexample deltamethrin methyl parathion and quinalphos produce increases in total sugarsand reducing sugars as well as increasing the protein contents of the rice leaf sheath andleaf blade [28] Several insecticides also reduce plant phenols in ricemdashwhich increases therice plantrsquos susceptibility to BPH and other insects [27] In our screenhouse experiment wenoted several changes to rice anatomy and yields Importantly we noted that cypermethrinand fipronil both reduced rice yields (Table 3) This was not due to indirect herbivoreeffects because planthopper and leafhopper densities were consistently low on rice treatedwith these insecticides The effects therefore probably resulted from direct phytotoxiceffects on the plants Furthermore these effects were most pronounced on treated IR62plants Other minor effects on grain production included increased proportions of unfilledgrain in cartap hydrochloride-treated plants and delays in grain maturation on cartaphydrochloride-treated and cypermethrin-treated plants (Table 3) Despite the high densitiesof BPH and WBPH on deltamethrin-treated plants yields were not significantly lower thanin control non-treated plants Both IR62 and IR64 have been noted for their high toleranceto BPH damage particularly under high nitrogenous fertilizer applications [3456] as usedin our screenhouse experiment Therefore where plants have high tolerance to damageapplying insecticides could also result in profitability losses on rice farms and in the caseof insecticide-induced pest population increases tolerance might buffer against subsequentyield losses There are few reports of the phytotoxicity of insecticides to rice carbofuran istoxic to rice at high concentrations but may also enhance the growth of rice plants throughinteractions with nitrogen-fixing bacteria in the rhizosphere [80] Future studies couldmore carefully assess field-grown plants for possible phytotoxic effects particularly ifinsecticides have the potential to directly reduce yields
44 Management implications
Global trends suggest that resistance in rice has failed to reduce insecticide appli-cations [2] This may be due to aggressive marketing by agrochemical companies andstrategies to increase pesticide sales such as promoting prophylactic applications [16] Be-
Crops 2021 1 180
cause we selected our test insecticides (except the pyrethroids) without any a priori criteriaour results suggest that many currently used insecticides probably have some negativeeffects on yield either directly through phytotoxic effects or indirectly by inducing pestresurgence Furthermore our results indicate that even where pests reach high densitiesas on deltamethrin-treated plants in the screenhouse these densities do not necessarilylead to significant yield reductions
Among the products we used buprofezin although it had little effect on oviposi-tion was effective in reducing nymph survival Furthermore there were only minorapparent phytotoxic effects on the rice plants Buprofezin has a low toxicity to naturalenemies in rice fields [58] and is therefore the most compatible of the insecticides withstrategies to increase rice field biodiversity and restore ecosystem services [32ndash348182]Thiamethoxam-chlorantraniliprole was also effective in reducing both oviposition andnymph survival and had no apparent toxic effects on rice plants However thiamethoxam-chlorantraniliprole and other neonicotinoids although moderately toxic can have severeeffects on beneficial hymenopteran parasitoids [83] although they have apparently lowtoxicity to predatory mirid bugs [84] The remaining insecticides either directly reducedyields or were associated with higher BPH biomass or densities in our experiments
We noted few synergies with HPR although nymph biomass andor survival waslower in cartap hydrochloride and carbofuran-treated IR62 plants compared to correspond-ing IR64 plants Meanwhile cartap hydrochloride and deltamethrin were antagonisticto resistance in IR62 in the screenhouse experiment Among the strongest effects of HPRwas to act as a buffer against GLH population build-up on deltamethrin-treated IR62 andagainst carbofuran-induced increases in BPH nymph biomass in the survival bioassayTherefore HPR may play a role in reducing the possibilities of secondary outbreaks ofBPH and other insects The combined use of HPR and targeted insecticides against BPHcreates redundancy in the rice field and counters measures to manage insecticide resistanceand virulence by unnecessarily exposing plant protection products to evolving pest pop-ulations HPR (and host plant tolerance) can be effective against target pests as revealedin previous studies [4] and could be incorporated into other pest management practicesparticularly agroecological practices such as ecological engineering with which it is highlycompatible [818586]
5 Conclusions
Based on our results and the results of previous studies HPR and the applicationof chemical insecticides against BPH rarely produce synergistic effects Antagonisticeffects where chemicals caused increases in BPH fitness or populations on resistant ricewere more apparent in our study In particular deltamethrin caused an outbreak ofBPH in our screenhouse study Nevertheless HPR can buffer against the insecticide-induced enhancement of herbivore fitness (hormesis) and potentially reduce possibilities ofsecondary outbreaks as was suggested from results with cartap hydrochloride in our studyBased on our results we suggest that carbofuran cartap hydrochloride cypermethrindeltamethrin and fipronil should be avoided in rice particularly in the Philippines (wherethis research was conducted) due to their resurgence-causing potential or because of directreductions to rice yields
Author Contributions Conceptualization FGH methodology FGH AP-C and MLPA formalanalysis FGH and AP-C investigation FGH AP-C and MLPA data curation FGHAP-C and MLPA writingmdashoriginal draft preparation FGH and AP-C writingmdashreview andediting FGH AP-C and MLPA visualization FGH supervision FGH and MLPA projectadministration FGH funding acquisition FGH All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by the Bill and Melinda Gates Foundation (Cereal SystemsInitiative for South Asia [CSISA] OPP52303) and the Global Rice Science Platform (GRiSP) underthe directorship of Achim Dobermann AP-C was partially funded by The Spanish Governmentthrough Becas Faro International
Crops 2021 1 181
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data presented in this study are available on reasonable requestfrom the corresponding author
Acknowledgments The authors thank Fame Ramal Carmencita Bernal Alberto Naredo ReyeulQuintana Vincent Virtudes Marol Recide Jenyrose Geronda and Ellen Genil for assistance duringthis study The contributions of Rodante Abas (sadly deceased) during three decades of entomologicalresearch at IRRI are gratefully acknowledged Rice lines were supplied through the Genebank of IRRI
Conflicts of Interest The authors declare no conflict of interest
References1 Bottrell DG Schoenly KG Resurrecting the ghost of green revolutions past The brown planthopper as a recurring threat to
high-yielding rice production in tropical Asia J Asia-Pac Entomol 2012 15 122ndash140 [CrossRef]2 Horgan FG Integrated pest management for sustainable rice cultivation A holistic approach In Achieving Sustainable Cultivation
of Rice Cultivation Pest and Disease Management Sasaki T Ed Burleigh Dodds Scientific Cambridge UK 2017 pp 309ndash3423 Horgan FG de Freitas TFS Crisol-Martiacutenez E Mundaca EA Bernal CC Nitrogenous fertilizer reduces resistance but
enhances tolerance to the brown planthopper in fast-growing moderately resistant rice Insects 2021 12 989 [CrossRef] [PubMed]4 Horgan FG Pentildealver Cruz A Bernal CC Ramal AF Almazan MLP Wilby A Resistance and tolerance to the brown
planthopper Nilaparvata lugens (Staringl) in rice infested at different growth stages across a gradient of nitrogen applicationsField Crops Res 2018 217 53ndash65 [CrossRef]
5 Hereward JP Cai X Matias AMA Walter GH Xu C Wang Y Migration dynamics of an important rice pest The brownplanthopper (Nilaparvata lugens) across AsiamdashInsights from population genomics Evol Appl 2020 13 2449ndash2459 [CrossRef]
6 Hu C Guo J Fu X Huang Y Gao X Wu K Seasonal migration pattern of Nilaparvata lugens (Hemiptera Delphacidae) overthe Bohai Sea in northern China J Econ Entomol 2018 111 2129ndash2135 [CrossRef]
7 Ma J Wang Y-C Hu Y-Y Lu M-H Wan G-J Chen F-J Liu W-C Zhai B-P Hu G Brown planthopper Nilaparvatalugens was concentrated at the rear of the typhoon Soudelor in eastern China in August 2015 Insect Sci 2018 25 916ndash926[CrossRef]
8 Fujita D Kohli A Horgan FG Rice resistance to planthoppers and leafhoppers Crit Rev Plant Sci 2013 32 162ndash191[CrossRef]
9 Fujii T Yoshida K Kobayashi T Myint KKM Yasui H Sanada-Morimura S Matsumura M Long-term virulencemonitoring of differential cultivars in Japanrsquos immigrant populations of Nilaparvata lugens (Hemiptera Delphacidae) in 2001ndash2009Appl Entomol Zool 2021 56 407ndash418 [CrossRef]
10 Myint KKM Yasui H Takagi M Matsumura M Virulence of long-term laboratory populations of the brown planthopperNilaparvata lugens (Stal) and whitebacked planthopper Sogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) on rice differentialvarieties Appl Entomol Zool 2009 44 149ndash153 [CrossRef]
11 Horgan FG Ramal AF Bentur JS Kumar R Bhanu KV Sarao PS Iswanto EH Van Chien H Phyu MHBernal CC et al Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia againstresistant rice varieties Crop Prot 2015 78 222ndash231 [CrossRef]
12 Horgan FG Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers Crop Prot2018 110 21ndash33 [CrossRef]
13 Uddin ABMA Islam KS Jahan M Ara A Afrin S Farmerrsquos perception about resurgence of brown planthopper NilaparvataLugens (Staringl) in Bangladesh Bangladesh Rice J 2020 23 21ndash33 [CrossRef]
14 Triwidodo H Brown planthoppers infestations and insecticides use pattern in Java Indonesia AGRIVITA J Agric Sci 202042 320ndash330 [CrossRef]
15 Veres A Wyckhuys KA Kiss J Toacuteth F Burgio G Pons X Avilla C Vidal S Razinger J Bazok R et al An update of theWorldwide Integrated Assessment (WIA) on systemic pesticides Part 4 Alternatives in major cropping systems Environ SciPollut Res 2020 27 29867ndash29899 [CrossRef]
16 Bakker L van der Werf W Tittonell P Wyckhuys KA Bianchi FJ Neonicotinoids in global agriculture Evidence for a newpesticide treadmill Ecol Soc 2020 25 26 [CrossRef]
17 Wu J-C Xu J-X Yuan S-Z Liu J-L Jiang Y-H Xu J-F Pesticide-induced susceptibility of rice to brown planthopperNilaparvata lugens Entomol Exp Appl 2001 100 119ndash126 [CrossRef]
18 Wu J-C Qiu H-M Yang G-Q Liu J-L Liu G-J Wilkens RM Effective duration of pesticide-induced susceptibility of riceto brown planthopper (Nilaparvata lugens Staringl Homoptera Delphacidae) and physiological and biochemical changes in riceplants following pesticide application Int J Pest Manag 2004 50 55ndash62 [CrossRef]
19 Wang A-H Wu J-C Yu Y-S Liu J-L Yue J-F Wang MY Selective insecticide-induced stimulation on fecundity andbiochemical changes in Tryporyza incertulas (Lepidoptera Pyralidae) J Econ Entomol 2005 98 1144ndash1149 [CrossRef] [PubMed]
Crops 2021 1 182
20 Azzam S Wang F Wu J-C Shen J Wang L-P Yang G-Q Guo Y-R Comparisons of stimulatory effects of a seriesof concentrations of four insecticides on reproduction in the rice brown planthopper Nilaparvata lugens Staringl (HomopteraDelphacidae) Int J Pest Manag 2009 55 347ndash358 [CrossRef]
21 Ge L-Q Hu J-H Wu J-C Yang G-Q Gu H Insecticide-induced changes in protein RNA and DNA contents in ovary andfat body of female Nilaparvata lugens (Hemiptera Delphacidae) J Econ Entomol 2009 102 1506ndash1514 [CrossRef]
22 Ling S Zhang J Hu L Zhang R Effect of fipronil on the reproduction feeding and relative fitness of brown planthopperNilaparvata lugens Appl Entomol Zool 2009 44 543ndash548 [CrossRef]
23 Ge L-Q Wang L-P Zhao K-F Wu J-C Huang L-J Mating pair combinations of insecticide-treated male and femaleNilaparvata lugens Staringl (Hemiptera Delphacidae) planthoppers influence protein content in the male accessory glands (MAGs)and vitellin content in both fat bodies and ovaries of adult females Pestic Biochem Physiol 2010 98 279ndash288 [CrossRef]
24 Ge L-Q Wu J-C Zhao K-F Chen Y Yang G-Q Induction of Nlvg and suppression of Nljhe gene expression in Nilaparvatalugens (Staringl) (Hemiptera Delphacidae) adult females and males exposed to two insecticides Pestic Biochem Physiol 201098 269ndash278 [CrossRef]
25 Hu J-H Wu J-C Yin J-L Gu H-N Physiology of insecticide-induced stimulation of reproduction in the rice brownplanthopper (Nilaparvata lugens (Staringl)) Dynamics of protein in fat body and ovary Int J Pest Manag 2010 56 23ndash30 [CrossRef]
26 Wang L-P Shen J Ge L-Q Wu J-C Yang G-Q Jahn GC Insecticide-induced increase in the protein content of maleaccessory glands and its effect on the fecundity of females in the brown planthopper Nilaparvata lugens Staringl (HemipteraDelphacidae) Crop Prot 2010 29 1280ndash1285 [CrossRef]
27 Wu J Ge L Liu F Song Q Stanley D Pesticide-induced planthopper population resurgence in rice cropping systems AnnuRev Entomol 2020 65 409ndash429 [CrossRef]
28 Suri KS Singh G Insecticide-induced resurgence of the whitebacked planthopper Sogatella furcifera (Horvath) (HemipteraDelphacidae) on rice varieties with different levels of resistance Crop Prot 2011 30 118ndash124 [CrossRef]
29 Fabellar LT Heinrichs EA Toxicity of insecticides to predators of rice brown planthoppers Nilaparvata lugens (Staringl) (HomopteraDelphacidae) Environ Entomol 1984 13 832ndash837 [CrossRef]
30 Fabellar LT Heinrichs EA Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators Crop Prot1986 5 254ndash258 [CrossRef]
31 Cuong NL Ben PT Phuong LT Chau LM Cohen MB Effect of host plant resistance and insecticide on brown planthopperNilaparvata lugens (Staringl) and predator population development in the Mekong Delta Vietnam Crop Prot 1997 16 707ndash715[CrossRef]
32 Preetha G Stanley J Suresh S Samiyappan R Risk assessment of insecticides used in rice on mirid bug Cyrtorhinuslividipennis Reuter the important predator of brown planthopper Nilaparvata lugens (Stal) Chemosphere 2010 80 498ndash503[CrossRef] [PubMed]
33 Tanaka K Endo S Kazano H Toxicity of insecticides to predators of rice planthoppers Spiders the mirid bug and the dryinidwasp Appl Entomol Zool 2000 35 177ndash187 [CrossRef]
34 Zhang X Xu Q Lu W Liu F Sublethal effects of four synthetic insecticides on the generalist predator Cyrtorhinus lividipennisJ Pest Sci 2015 88 383ndash392 [CrossRef]
35 Settle WH Ariawan H Astuti ET Cahyana W Hakim AL Hindayana D Lestari AS Managing tropical rice peststhrough conservation of generalist natural enemies and alternative prey Ecology 1996 77 1975ndash1988 [CrossRef]
36 Salim M Heinrichs EA Insecticide-induced changes in the levels of resistance of rice cultivars to the whitebacked planthopperSogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) Crop Prot 1987 6 28ndash32 [CrossRef]
37 Yin J-L Xu H-W Wu J-C Hu J-H Yang G-Q Cultivar and insecticide applications affect the physiological development ofthe brown planthopper Nilaparvata lugens (Staringl) (Hemiptera Delphacidae) Environ Entomol 2014 37 206ndash212 [CrossRef]
38 Cheng C Chang W Studies on varietal resistance to the brown planthopper in Taiwan In Brown Planthopper Threat toRice Production in Asia International Rice Research Institute Ed International Rice Research Institute Manila Philippines1979 pp 251ndash271
39 Reissig WH Heinrichs EA Valencia SL Insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on ricevarieties with different levels of resistance Environ Entomol 1982 11 165ndash168 [CrossRef]
40 Heinrichs EA Fabellar LT Basilio RP Wen T-C Medrano F Susceptibility of rice planthoppers Nilaparvata lugens andSogatella furcifera (Homoptera Delphacidae) to insecticides as influenced by level of resistance in the host plant Environ Entomol1984 13 455ndash458 [CrossRef]
41 Joshi R Shepard B Kenmore P Lydia R Insecticide-induced resurgence of brown planthopper (BPH) on IR62 Int Rice ResNewsl 1992 17 9ndash10
42 Zhu Z-R Cheng J Jiang M-X Zhang X-X Complex influence of rice variety fertilization timing and insecticide onpopulation dynamics of Sogatella furcifera (Horvath) Nilaparvata lugens (Staringl) (Homoptera Delphacidae) and their natural enemiesin rice in Hangzhou China J Pest Sci 2004 77 65ndash74 [CrossRef]
43 Nanthakumar M Jhansi Lakshmi V Shashi Bhushan V Balachandran SM Mohan M Decrease of rice plant resistance andinduction of hormesis and carboxylesterase titre in brown planthopper Nilaparvata lugens (Staringl) by xenobiotics Pestic BiochemPhysiol 2012 102 146ndash152 [CrossRef]
Crops 2021 1 183
44 Yao H-W Jiang C-Y Ye G-Y Cheng J-A Studies on cultivar-induced changes in insecticide susceptibility and its relatedenzyme activities of the white-backed planthopper Sogatella furcifera (Horvath)(Homopter Delphacidae) Insect Sci 2002 9 9ndash15
45 Horgan FG Srinivasan TS Bentur JS Kumar R Bhanu KV Sarao PS Van Chien H Almazan MLP Bernal CCRamal AF et al Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers Implicationsfor rice breeding and gene deployment Agronomy 2017 7 62 [CrossRef]
46 Jairin J Phengrat K Teangdeerith S Vanavichit A Toojinda T Mapping of a broad-spectrum brown planthopper resistancegene Bph3 on rice chromosome 6 Mol Breed 2007 19 35ndash44 [CrossRef]
47 Ren J Gao F Wu X Lu X Zeng L Lv J Su X Luo H Ren G Bph32 a novel gene encoding an unknown SCRdomain-containing protein confers resistance against the brown planthopper in rice Sci Rep 2016 6 37645 [CrossRef]
48 Nguyen CD Zheng S-H Sanada-Morimura S Matsumura M Yasui H Fujita D Substitution mapping and characterizationof brown planthopper resistance genes from indica rice variety lsquoPTB33prime (Oryza sativa L) Breed Sci 2021 in press [CrossRef]
49 Chelliah S Heinrichs EA Factors affecting insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on RiceEnviron Entomol 1980 9 773ndash777 [CrossRef]
50 Moore MT Kroumlger R Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growthArch Environ Contam Toxicol 2010 59 574ndash581 [CrossRef]
51 Kim J Hwang I Han M Jang B Effect of insecticide and fungicide on phytotoxicity of herbicide in rice Korean J Weed Sci1986 6 67ndash75
52 Horgan FG Garcia CPF Haverkort F de Jong PW Ferrater JB Changes in insecticide resistance and host range performanceof planthoppers artificially selected to feed on resistant rice Crop Prot 2020 127 104963 [CrossRef] [PubMed]
53 Khush GS Virk PS IR Varieties and Their Impact International Rice Research Institute Manila Philippines 200554 Srinivasan TS Almazan MLP Bernal CC Ramal AF Subbarayalu MK Horgan FG Interactions between nymphs of
Nilaparvata lugens and Sogatella furcifera (Hemiptera Delphacidae) on resistant and susceptible rice varieties Appl Entomol Zool2016 51 81ndash90 [CrossRef]
55 Pentildealver Cruz A Arida A Heong KL Horgan FG Aspects of brown planthopper adaptation to resistant rice varieties withthe Bph3 gene Entomol Exp Appl 2011 141 245ndash257 [CrossRef]
56 Alam SN Cohen MB Detection and analysis of QTLs for resistance to the brown planthopper Nilaparvata lugens in adoubled-haploid rice population Theor Appl Genet 1998 97 1370ndash1379 [CrossRef]
57 Mackill DJ Khush GS IR64 A high-quality and high-yielding mega variety Rice 2018 11 18 [CrossRef] [PubMed]58 Heinrichs EA Basilio RP Valencia SL Buprofezin a selective insecticide for the management of rice planthoppers (Ho-
moptera Delphacidae) and leafhoppers (Homoptera Cicadellidae) Environ Entomol 1984 13 515ndash521 [CrossRef]59 Satapathy MK Anjaneyulu A Use of cypermethrin a synthetic pyrethroid in the control of rice tungro virus disease and its
vector Trop Pest Manag 1984 30 170ndash178 [CrossRef]60 Vorley W Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown
planthopper (BPH) in Kedah Malaysia Int Rice Res Newsl 1987 10 19ndash2061 Uddin A Islam KS Jahan M Ara A Khan MAI Factor influencing the resurgence of brown planthopper in Bangladesh
SAARC J Agric 2020 18 117ndash128 [CrossRef]62 Escalada MM Luecha M Heong KL Social impacts of planthopper outbreaks Case study from Thailand In Rice Planthoppers
Ecology Management Socio Economics and Policy Heong KL Cheng J Escalada MM Eds Zhejiang University PressHangzhou China 2015 pp 209ndash226
63 Zhang CP He HM Yu JZ Hu XQ Zhu YH Wang Q Residues of carbosulfan and its metabolites carbofuran and3-hydroxy carbofuran in rice field ecosystem in China J Environ Sci Health 2016 51 351ndash357 [CrossRef] [PubMed]
64 Samanta S Barman M Samanta NRA Bio-efficacy trials of carbofuran 3 CG against insect pests of rice J Entomol ZoolStud 2020 8 1529ndash1533
65 Donley N The USA lags behind other agricultural nations in banning harmful pesticides Environ Health 2019 18 44 [CrossRef][PubMed]
66 Adams A Gore J Musser F Cook D Catchot A Walker T Dobbins C Efficacy of selected insecticides applied to hybridrice seed J Econ Entomol 2015 109 200ndash206 [CrossRef]
67 Kumar R Singh B Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) Field Bull Environ Contam Toxicol2013 90 482ndash488 [CrossRef]
68 Dhaka S Prajapati C Singh D Singh R Field evaluation of insecticides and bio-pesticides against rice leaf folder CnaphalocrocisMedinalis (Guenee) Ann Plant Prot Sci 2011 19 324ndash326
69 Matsumura M Takeuchi H Satoh M Sanada-Morimura S Otuka A Watanabe T Van Thanh D Species-specific insecticideresistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in East and South-eastAsia Pest Manag Sci 2008 64 1115ndash1121 [CrossRef] [PubMed]
70 Tang J Li J Shao Y Yang B Liu Z Fipronil resistance in the whitebacked planthopper (Sogatella furcifera) Possible resistancemechanisms and cross-resistance Pest Manag Sci 2010 66 121ndash125 [CrossRef]
71 Horgan FG Arida A Ardestani G Almazan MLP Temperature-dependent oviposition and nymph performance revealdistinct thermal niches of coexisting planthoppers with similar thresholds for development PLoS ONE 2020 15 e0235506[CrossRef] [PubMed]
Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]
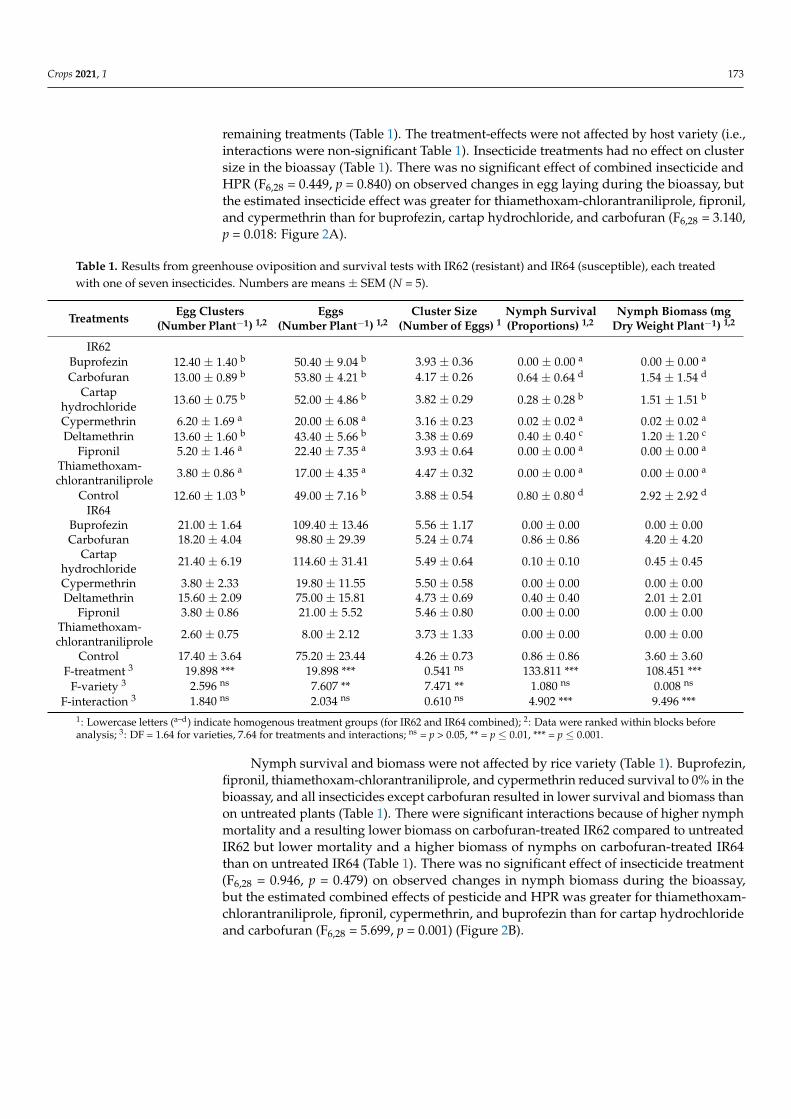
Crops 2021 1 173
remaining treatments (Table 1) The treatment-effects were not affected by host variety (ieinteractions were non-significant Table 1) Insecticide treatments had no effect on clustersize in the bioassay (Table 1) There was no significant effect of combined insecticide andHPR (F628 = 0449 p = 0840) on observed changes in egg laying during the bioassay butthe estimated insecticide effect was greater for thiamethoxam-chlorantraniliprole fiproniland cypermethrin than for buprofezin cartap hydrochloride and carbofuran (F628 = 3140p = 0018 Figure 2A)
Table 1 Results from greenhouse oviposition and survival tests with IR62 (resistant) and IR64 (susceptible) each treatedwith one of seven insecticides Numbers are means plusmn SEM (N = 5)
Treatments Egg Clusters(Number Plantminus1) 12
Eggs(Number Plantminus1) 12
Cluster Size(Number of Eggs) 1
Nymph Survival(Proportions) 12
Nymph Biomass (mgDry Weight Plantminus1) 12
IR62Buprofezin 1240 plusmn 140 b 5040 plusmn 904 b 393 plusmn 036 000 plusmn 000 a 000 plusmn 000 a
Carbofuran 1300 plusmn 089 b 5380 plusmn 421 b 417 plusmn 026 064 plusmn 064 d 154 plusmn 154 d
Cartaphydrochloride 1360 plusmn 075 b 5200 plusmn 486 b 382 plusmn 029 028 plusmn 028 b 151 plusmn 151 b
Cypermethrin 620 plusmn 169 a 2000 plusmn 608 a 316 plusmn 023 002 plusmn 002 a 002 plusmn 002 a
Deltamethrin 1360 plusmn 160 b 4340 plusmn 566 b 338 plusmn 069 040 plusmn 040 c 120 plusmn 120 c
Fipronil 520 plusmn 146 a 2240 plusmn 735 a 393 plusmn 064 000 plusmn 000 a 000 plusmn 000 a
Thiamethoxam-chlorantraniliprole 380 plusmn 086 a 1700 plusmn 435 a 447 plusmn 032 000 plusmn 000 a 000 plusmn 000 a
Control 1260 plusmn 103 b 4900 plusmn 716 b 388 plusmn 054 080 plusmn 080 d 292 plusmn 292 d
IR64Buprofezin 2100 plusmn 164 10940 plusmn 1346 556 plusmn 117 000 plusmn 000 000 plusmn 000Carbofuran 1820 plusmn 404 9880 plusmn 2939 524 plusmn 074 086 plusmn 086 420 plusmn 420
Cartaphydrochloride 2140 plusmn 619 11460 plusmn 3141 549 plusmn 064 010 plusmn 010 045 plusmn 045
Cypermethrin 380 plusmn 233 1980 plusmn 1155 550 plusmn 058 000 plusmn 000 000 plusmn 000Deltamethrin 1560 plusmn 209 7500 plusmn 1581 473 plusmn 069 040 plusmn 040 201 plusmn 201
Fipronil 380 plusmn 086 2100 plusmn 552 546 plusmn 080 000 plusmn 000 000 plusmn 000Thiamethoxam-chlorantraniliprole 260 plusmn 075 800 plusmn 212 373 plusmn 133 000 plusmn 000 000 plusmn 000
Control 1740 plusmn 364 7520 plusmn 2344 426 plusmn 073 086 plusmn 086 360 plusmn 360F-treatment 3 19898 19898 0541 ns 133811 108451
F-variety 3 2596 ns 7607 7471 1080 ns 0008 ns
F-interaction 3 1840 ns 2034 ns 0610 ns 4902 9496 1 Lowercase letters (andashd) indicate homogenous treatment groups (for IR62 and IR64 combined) 2 Data were ranked within blocks beforeanalysis 3 DF = 164 for varieties 764 for treatments and interactions ns = p gt 005 = p le 001 = p le 0001
Nymph survival and biomass were not affected by rice variety (Table 1) Buprofezinfipronil thiamethoxam-chlorantraniliprole and cypermethrin reduced survival to 0 in thebioassay and all insecticides except carbofuran resulted in lower survival and biomass thanon untreated plants (Table 1) There were significant interactions because of higher nymphmortality and a resulting lower biomass on carbofuran-treated IR62 compared to untreatedIR62 but lower mortality and a higher biomass of nymphs on carbofuran-treated IR64than on untreated IR64 (Table 1) There was no significant effect of insecticide treatment(F628 = 0946 p = 0479) on observed changes in nymph biomass during the bioassaybut the estimated combined effects of pesticide and HPR was greater for thiamethoxam-chlorantraniliprole fipronil cypermethrin and buprofezin than for cartap hydrochlorideand carbofuran (F628 = 5699 p = 0001) (Figure 2B)
Crops 2021 1 174
Crops 2021 1 FOR PEER REVIEW 9
Figure 2 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) egg laying and (B) nymph biomass in greenhouse bioas-says Effects are indicated as proportional changes in fitness parameters compared to untreated IR64 Negative numbers indicate reduced fitness Standard errors are indicated (N = 5) Lowercase letters (ab) indicated homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 1)
Nymph survival and biomass were not affected by rice variety (Table 1) Buprofezin fipronil thiamethoxam-chlorantraniliprole and cypermethrin reduced survival to 0 in the bioassay and all insecticides except carbofuran resulted in lower survival and biomass than on untreated plants (Table 1) There were significant interactions because of higher nymph mortality and a resulting lower biomass on carbofuran-treated IR62 compared to untreated IR62 but lower mortality and a higher biomass of nymphs on carbofuran-treated IR64 than on untreated IR64 (Table 1) There was no significant effect of insecticide treatment (F628 = 0946 p = 0479) on observed changes in nymph biomass during the bio-assay but the estimated combined effects of pesticide and HPR was greater for thiameth-oxam-chlorantraniliprole fipronil cypermethrin and buprofezin than for cartap hydro-chloride and carbofuran (F628 = 5699 p = 0001) (Figure 2B)
33 Effects of HPR and Insecticides on Planthoppers and Leafhoppers Plants in the screenhouse were colonized by BPH WBPH and GLH Densities for
each species were similar on IR62 and IR64 (Wilkrsquos λ = 0994 p = 0952) There was a sig-nificant treatment effect on all three species (Wilkrsquos λ = 0586 p = 0049) because of high densities on deltamethrin-treated plants (Table 2) The numbers and biomass of all three species combined were also greater on deltamethrin-treated plants than on the remaining treatments (Table 2) Host resistance reduced GLH densities under the deltamethrin treat-ment but had no apparent effect on densities under the other treatments (Table 2) There were significant combined effects of HPR and insecticide on the numbers of BPH (F624 = 2686 p = 0039) and WBPH (F624 = 2482 p = 0052) this was almost exclusively due to the resurgence effect of deltamethrin on the planthoppers (BPH F624 = 2686 p = 0039 WBPH F624 = 2482 p = 0052) (Figure 3)
Δ eg
gs(P
ropo
rtion
)
-10
-08
-06
-04
-02
00
02
04
06
Treatment
A B C D E F G
Δ N
ymph
bio
mas
s(P
ropo
rtion
)
-10
-08
-06
-04
-02
00
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
ab
a a
b
a
b b
a a a a a a a a
b b bb
bb
Figure 2 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone(P grey bars) on (A) egg laying and (B) nymph biomass in greenhouse bioassays Effects are indicated as proportionalchanges in fitness parameters compared to untreated IR64 Negative numbers indicate reduced fitness Standard errors areindicated (N = 5) Lowercase letters (ab) indicated homogenous treatment groups based on Tukey pairwise comparisons(p lt 005) (see also Table 1)
33 Effects of HPR and Insecticides on Planthoppers and Leafhoppers
Plants in the screenhouse were colonized by BPH WBPH and GLH Densities foreach species were similar on IR62 and IR64 (Wilkrsquos λ = 0994 p = 0952) There was asignificant treatment effect on all three species (Wilkrsquos λ = 0586 p = 0049) because of highdensities on deltamethrin-treated plants (Table 2) The numbers and biomass of all threespecies combined were also greater on deltamethrin-treated plants than on the remainingtreatments (Table 2) Host resistance reduced GLH densities under the deltamethrintreatment but had no apparent effect on densities under the other treatments (Table 2)There were significant combined effects of HPR and insecticide on the numbers of BPH(F624 = 2686 p = 0039) and WBPH (F624 = 2482 p = 0052) this was almost exclusively dueto the resurgence effect of deltamethrin on the planthoppers (BPH F624 = 2686 p = 0039WBPH F624 = 2482 p = 0052) (Figure 3)
Crops 2021 1 175
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants underseven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density(Number Plantminus1) 1
WBPH Density(Number Plantminus1) 1
GLH Density(Number Plantminus1) 1
Density ofPlanthoppers and
Leafhoppers(Number Plantminus1) 1
Biomass ofPlanthoppers and
Leafhoppers(mg Dry Weight) 1
IR62Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a
Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartaphydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a
Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a
Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a
Thiamethoxam-chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a
IR64Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartaphydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123
Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns
F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Crops 2021 1 FOR PEER REVIEW 10
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants under seven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density (Num-
ber Plantminus1) 1 WBPH Density
(Number Plantminus1) 1 GLH Density
(Number Plantminus1) 1
Density of Planthoppers and Leafhoppers (Number
Plantminus1) 1
Biomass of Planthoppers and Leafhoppers (mg Dry
Weight) 1 IR62
Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartap hydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a Thiamethoxam-
chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a IR64
Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132 Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartap hydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123 Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113 Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052 Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087 F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns 1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatments and interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in the screenhouse Effects are indicated as proportional changes in planthopper densities compared to untreated IR64 Negative numbers indicate reduced densities positive numbers are increased den-sities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
Δ BP
H(N
umbe
r pla
nt-1
)
0
5
10
15
20
Treatment
A B C D E F G
Δ W
BPH
(Num
ber p
lant
-1)
0
20
30
40
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
a a
aa a a
a a a
a
a a
b
b
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimatedpesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in thescreenhouse Effects are indicated as proportional changes in planthopper densities compared to
Crops 2021 1 176
untreated IR64 Negative numbers indicate reduced densities positive numbers are increaseddensities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenoustreatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
34 Direct and Indirect Effects of Insecticides on Rice Development
IR62 plants had more tillers and attained a greater biomass than IR64 plants in thescreenhouse (Table 3) IR62 grain matured faster and plants were harvested earlier than theIR64 plants (Table 3) IR62 plants also produced more grain (number of grain) and hadhigher yields than IR64 but had smaller grain and more unfilled grain than IR64 (Table 3)
Table 3 Rice growth parameters for IR62 (resistant) and IR64 (susceptible) under seven insecticide treatments and forcontrol non-treated plants in a screenhouse environment Numbers are means plusmn SEM
Treatment Tiller Number 1 Plant Biomass(g Dry Weight) 1
Time to Harvest(Days) 1
Weightof Filled Grain(g Dry Weight) 1
Number ofFilled Grain
(1000s) 1
Unfilled Grain(Proportion) 1
1000 GrainWeight 1
IR62Buprofezin 2660 plusmn 206 ab 4643 plusmn 385 ab 7990 plusmn 168 ab 5776 plusmn 683 ab 361plusmn 061 ab 011 plusmn 003 a 2008 plusmn 042 ab
Carbofuran 2900 plusmn 106 ab 5546 plusmn 357 b 7720 plusmn 080 ab 7448 plusmn 691 b 446plusmn 061 ab 011 plusmn 001 a 2103 plusmn 054 b
Cartaphydrochloride 2820 plusmn 327 ab 4896 plusmn 470 ab 7920 plusmn 153 b 6430 plusmn 712 ab 415plusmn 064 ab 012 plusmn 001 b 2023 plusmn 065 a
Cypermethrin 2870 plusmn 301 a 4957 plusmn 562 a 7760 plusmn 103 b 6032 plusmn 822 a 410plusmn 067 ab 014 plusmn 002 a 1965 plusmn 076 a
Deltamethrin 2620 plusmn 345 a 4592 plusmn 588 a 7840 plusmn 108 ab 5698 plusmn 832 ab 379plusmn 047 ab 012 plusmn 001 a 2030 plusmn 063 ab
Fipronil 2380 plusmn 176 ab 4607 plusmn 657 ab 7700 plusmn 179 ab 5266 plusmn 390 a 318 plusmn 032 a 013 plusmn 002 a 2128 plusmn 047 b
Thiamethoxam-chlorantraniliprole 2650 plusmn 250 b 4547 plusmn 429 b 7720 plusmn 102 a 5777 plusmn 688 ab 408 plusmn 061 b 013 plusmn 002 a 2019 plusmn 067 ab
Control 3120 plusmn 247 b 5401 plusmn 555 b 7720 plusmn 080 ab 7045 plusmn 520 b 396plusmn 036 ab 012 plusmn 002 a 2076 plusmn 050 ab
IR64Buprofezin 2340 plusmn 250 5064 plusmn 747 8350 plusmn 124 5080 plusmn 952 270 plusmn 054 008 plusmn 001 2277 plusmn 039Carbofuran 2040 plusmn 097 4591 plusmn 141 8540 plusmn 073 4951 plusmn 312 263 plusmn 023 006 plusmn 000 2295 plusmn 036
Cartaphydrochloride 2120 plusmn 075 4336 plusmn 285 8520 plusmn 140 4273 plusmn 579 238 plusmn 026 015 plusmn 007 2050 plusmn 234
Cypermethrin 1990 plusmn 089 3968 plusmn 272 8710 plusmn 135 3920 plusmn 552 250 plusmn 028 008 plusmn 002 2171 plusmn 070Deltamethrin 1980 plusmn 181 4325 plusmn 357 8460 plusmn 068 4710 plusmn 496 277 plusmn 024 008 plusmn 001 2314 plusmn 012
Fipronil 2120 plusmn 174 4697 plusmn 311 8450 plusmn 089 4888 plusmn 392 274 plusmn 027 008 plusmn 001 2247 plusmn 042Thiamethoxam-chlorantraniliprole 2520 plusmn 287 5404 plusmn 465 8380 plusmn 196 5594 plusmn 546 347 plusmn 053 010 plusmn 001 2265 plusmn 072
Control 2160 plusmn 076 4678 plusmn 342 8600 plusmn 096 4940 plusmn 543 267 plusmn 029 008 plusmn 001 2283 plusmn 049F-treatment 4100 ns 3410 2253 ns 5150 2555 3923 4924
F-variety 168232 7665 547500 104356 118242 32308 106597 F-interaction 5980 6113 4824 5760 2921 2769 2552
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Insecticides affected plant growth directly and indirectly (through increased planthop-per densities) Plants treated with cypermethrin and deltamethrin had fewer tillers had alower biomass and under cypermethrin had lower yields than untreated plants (Table 3)Plants treated with fipronil also had reduced yields due to lower grain numbers comparedto untreated plants (Table 3) Although yields were not affected treatment with cartaphydrochloride increased the proportion of unfilled grain compared to untreated plantsof both varieties Cartap hydrochloride and cypermethrin also caused a delay in grainmaturation on IR62 and IR64 respectively relative to thiamethoxam-chlorantraniliproletreated plants (Table 3)
There were several significant interactions tiller numbers on both plants were similarunder buprofezin fipronil and thiamethoxam-chlorantraniliprole treatments but wereotherwise higher on IR62 (Table 3) biomass was lower on IR64 under carbofuran andcypermethrin treatments but was otherwise similar on both plants (Table 3) buprofezindelayed grain maturation on IR62 but maturation was otherwise faster on IR62 comparedto IR64 (Table 3) yields under buprofezin fipronil and thiamethoxam-chlorantraniliprolewere similar for IR62 and IR64 but otherwise yields were higher on IR62 (Table 3) grain
Crops 2021 1 177
numbers were higher on IR62 than IR64 except under fipronil treatments (Table 3) theproportions of grain unfilled were higher on IR62 but were similar between varieties underthe fipronil treatment (Table 3) 1000-grain weight was higher in IR64 but similar on bothvarieties when treated with cartap hydrochloride (Table 3)
4 Discussion
Our results using a highly resistant rice variety and some commonly used insecticidesindicate a range of possible effects on planthoppers and rice plants by combining these twomanagement practices
41 Synergies
The most predictable effect of combining HPR and insecticides is a synergy in reduc-ing BPH numbers This is suggested because planthoppers are weakened by the hostrsquosresistance and should therefore be more susceptible to the toxic effects of insecticides [40]Using spray towers over potted plants Heinrichs et al (1984) [40] examined the combinedeffects of a range of insecticides (that included carbofuran) with resistance by comparingBPH mortality on six resistant and two susceptible rice varieties Their results indicatedthat mortality was greatest when insecticides were applied to BPH on resistant varietiesThe effects were similar when the study was repeated with WBPH for three insecticides(including carbofuran) on the resistant variety N22 (Wbph1 gene) [40] In our study hostresistance reduced egg laying and nymph biomass on non-treated IR62 compared to non-treated IR64 Egg laying was also reduced on plants treated with cypermethrin fipronilor thiamethoxam-chlorantraniliprole (Table 1) Based on the results of our choice andno-choice oviposition bioassays cypermethrin acted mainly as a deterrent of gravid fe-males whereas fipronil and thiamethoxam-chlorantraniliprole directly reduced egg layingDespite similar BPH responses to both the insecticides and resistant host there were no ap-parent synergies between any of these insecticides and resistance in reducing egg numbersin the oviposition bioassay In contrast to these three insecticides our bioassays indicatedthat buprofezin carbofuran and cartap hydrochloride can lead to higher numbers of eggson treated plants compared to untreated plants (Table 1) These effects were not statisticallysignificant and may be exaggerated because bioassays were conducted over relativelyshort time scales For example planthoppers will sometimes respond to stresses such ashigh temperatures by increasing oviposition rates [7172] Furthermore the stimulation ofegg-laying by these chemicals was not apparent in our screenhouse experiment (Table 2)Nevertheless our results do indicate that host resistance reduced the magnitude of anystimulatory effect on oviposition egg numbers on buprofezin-treated carbofuran-treatedand cartap hydrochloride-treated IR64 in the no-choice bioassay were twice as high as onthe corresponding IR62 plants In cases such as these the resistance of IR62 functions as abuffer or lsquoinsurancersquo against possible undesired pesticide effects [1238]
Although buprofezin appeared to stimulate oviposition the growth hormone had astrong impact on nymph development leading to zero survival of exposed nymphs in ourgreenhouse bioassay Cypermethrin fipronil and thiamethoxam-chlorantraniliprole alsoreduced nymph survival to zero in the greenhouse bioassay (Table 1) Where insecticideshave such high efficiencies any fitness losses to BPH due to host resistance then becomeredundant Unlike buprofezin carbofuran also stimulated nymph biomass on IR64 in thesurvival bioassay (Table 1) however nymph biomass was lower on carbofuran-treated IR62than on non-treated IR62 indicating that the hostrsquos resistance buffered against undesiredstimulatory effects on BPH and may have functioned synergistically to augment mortalityand slow BPH growth Such buffering allows insecticides to be used against other pestswithout inducing secondary outbreaks of non-target pests In our screenhouse experimentherbivore densities were generally low on both IR62 and IR64 for most treatments (exceptdeltamethrin) and were below threshold densities for insecticide applicationsmdashwhichare normally about 25ndash50 planthoppers per hill [1373] Under such low densities it wasnot possible to observe the impact of host resistance on planthopper population build-
Crops 2021 1 178
up There was some indication of higher densities on cartap hydrochloride-treated IR62in the screenhouse which may be related to the stimulatory effects of the product onegg laying and the low mortality of nymphs on cartap hydrochloride-treated plants asobserved in the greenhouse bioassays (Table 1) Field studies from the Philippines haveindicated that IR62 effectively reduces BPH densities during wet and dry seasons [4]However farmers are rarely aware of which anti-herbivore resistance their rice varietiespossess and frequently apply pesticides regardless of resistance thereby reducing theirfarm profits [3841] For example Joshi et al (1992) [41] reported widespread outbreaks ofBPH on IR62 in Mindanao Philippines after applications of decamethrin (a pyrethroid) tothe resistant variety Where such resurgence-causing insecticides are applied resistanceseems to be often incapable of reducing the stimulatory effects of the insecticide and itloses its capacity to buffer against outbreaks
42 Antagonisms
Several insecticides are associated with increasing densities of planthoppers andwith resurgence outbreaks in farmersrsquo fields [27293041496174] Indeed patches oflsquohopperburnrsquo (ie dead rice plants resulting from planthopper damage) are often anindicator of high insecticide use in tropical Asia [214] In many cases these resurgence-causing insecticides were once effective in reducing planthopper densities but over timepopulations developed resistance against the products The resulting low toxicity canthen have a stimulatory effect on planthopper feeding and development [2123242743]This may be through hormesis which is a direct response by the planthopper to low-levelexposure to the toxic chemical or it may result from physiological changes to the rice plantsthat ultimately benefit the planthopper [1849] In our screenhouse study we induced aresurgence outbreak of BPH WBPH and to a lesser extent GLH on deltamethrin-treatedrice plants (Table 2) The combined densities of all three herbivores increased almost10 times on the treated plants Furthermore herbivore biomass increased by between fourand six times Although the resurgence effects of deltamethrin have been documentedin previous studies [2029ndash313975] we could not predict from our greenhouse studiesthat any such outbreak would occur Egg laying was unaffected by deltamethrin in ourchoice and no-choice experiments and nymph survival and development both declinedon deltamethrin-treated rice in the survival bioassay (Table 1) The massive increases inplanthopper densities on deltamethrin-treated plants in our study seem to result froma heightened attraction of adult BPH and WBPH to the plants [49] particularly sincenumbers were so low on all other plants including both treated and non-treated plantsHowever once on the plants planthoppers also apparently had enhanced fitness Cuonget al (1997) [31] observed higher population densities of BPH on deltamethrin-treatedrice in Vietnam albeit with lower-level increases on resistant varieties Furthermore in agreenhouse study Azzam et al (2009) [20] found that deltamethrin reduced pre-ovipositiontimes increased the duration of oviposition and increased egg hatchability and femalelongevity Nanthakumar et al (2012) [43] further showed that deltamethrin increased BPHfeeding efficiency on resistant and susceptible rice plants as determined through honeydewproduction and population size In studies with WBPH Salim and Heinrichs (1987) [36]showed deltamethrin to increase settling (antixenosis) feeding and population growthand Suri and Singh (2011) [28] showed deltamethrin to increase WBPH reproductive ratesand nymph survival In our study the observed resurgence of planthoppers was similaron both varieties indicating as was previously noted by Joshi et al (1992) [41] thatresurgence-causing pyrethroids are antagonistic to resistance in IR62
Though not statistically significant cartap hydrochloride also increased BPH numberson IR62 but not on IR64 in our screenhouse study This indicates that the product mayalso be antagonistic to the hostrsquos resistance mechanisms The stronger effect on IR62 maybe related to the differences between IR62 biomass growth rates and anatomy (ie alarger plant with higher tillering) compared to IR64 (Table 3) Once resistance is no longereffective because of chemically-enhanced BPH fitness these plant characteristics which
Crops 2021 1 179
have been related to observed preferences in BPH and other sap-sucking insects [349]could further accelerate the build-up of BPH populations on cartap hydrochloride-treatedplants To further test this idea experiments could be conducted using near-isogenic ricelines that differ mainly in the presence or absence of tagged resistance genes [76] Suchtests would avoid confounding differences in plant anatomy between test varieties IR62resistance to BPH has been well studied [4] The variety is also resistant to WBPH atolder plant stages [54] Therefore in our study deltamethrin was also antagonistic tothis WBPH-resistance (Table 2) However BPH feeding also facilitates WBPH populationgrowth [5477] and there may have been added indirect plant-mediated effects [77] thataugmented WBPH populations on both varieties in our experiment IR62 is also resistant toGLH [45] In our experiments GLH densities also increased on deltamethrin-treated plantsbut the effect was only apparent on IR64 Therefore GLH-resistance in IR62 apparentlybuffered against deltamethrin-associated increases in GLH on the variety as was alsonoted from field studies in Vietnam [31] Deltamethrin is currently not recommended foruse in rice [202878] however the product continues to be marketed in rice-dominatedregions throughout Asia [131479] Although cypermethrin has been associated with BPHoutbreaks and was consequently removed for use on rice in Thailand and Vietnam [62]we did not observe cypermethrin-induced resurgence in our screenhouse experimentNevertheless cypermethrin has been associated with resurgence of BPH in South Asia [61]and we have noted stimulatory effects of cypermethrin on other rice herbivores in thePhilippines (Horgan unpublished data)
43 Phytotoxicity
Phytotoxicity is relatively common following applications of insecticides [182850]Phytotoxic effects may change a plantrsquos physiology without consequences for crop yieldsHowever such changes to plant physiology could have indirect effects on herbivores Forexample deltamethrin methyl parathion and quinalphos produce increases in total sugarsand reducing sugars as well as increasing the protein contents of the rice leaf sheath andleaf blade [28] Several insecticides also reduce plant phenols in ricemdashwhich increases therice plantrsquos susceptibility to BPH and other insects [27] In our screenhouse experiment wenoted several changes to rice anatomy and yields Importantly we noted that cypermethrinand fipronil both reduced rice yields (Table 3) This was not due to indirect herbivoreeffects because planthopper and leafhopper densities were consistently low on rice treatedwith these insecticides The effects therefore probably resulted from direct phytotoxiceffects on the plants Furthermore these effects were most pronounced on treated IR62plants Other minor effects on grain production included increased proportions of unfilledgrain in cartap hydrochloride-treated plants and delays in grain maturation on cartaphydrochloride-treated and cypermethrin-treated plants (Table 3) Despite the high densitiesof BPH and WBPH on deltamethrin-treated plants yields were not significantly lower thanin control non-treated plants Both IR62 and IR64 have been noted for their high toleranceto BPH damage particularly under high nitrogenous fertilizer applications [3456] as usedin our screenhouse experiment Therefore where plants have high tolerance to damageapplying insecticides could also result in profitability losses on rice farms and in the caseof insecticide-induced pest population increases tolerance might buffer against subsequentyield losses There are few reports of the phytotoxicity of insecticides to rice carbofuran istoxic to rice at high concentrations but may also enhance the growth of rice plants throughinteractions with nitrogen-fixing bacteria in the rhizosphere [80] Future studies couldmore carefully assess field-grown plants for possible phytotoxic effects particularly ifinsecticides have the potential to directly reduce yields
44 Management implications
Global trends suggest that resistance in rice has failed to reduce insecticide appli-cations [2] This may be due to aggressive marketing by agrochemical companies andstrategies to increase pesticide sales such as promoting prophylactic applications [16] Be-
Crops 2021 1 180
cause we selected our test insecticides (except the pyrethroids) without any a priori criteriaour results suggest that many currently used insecticides probably have some negativeeffects on yield either directly through phytotoxic effects or indirectly by inducing pestresurgence Furthermore our results indicate that even where pests reach high densitiesas on deltamethrin-treated plants in the screenhouse these densities do not necessarilylead to significant yield reductions
Among the products we used buprofezin although it had little effect on oviposi-tion was effective in reducing nymph survival Furthermore there were only minorapparent phytotoxic effects on the rice plants Buprofezin has a low toxicity to naturalenemies in rice fields [58] and is therefore the most compatible of the insecticides withstrategies to increase rice field biodiversity and restore ecosystem services [32ndash348182]Thiamethoxam-chlorantraniliprole was also effective in reducing both oviposition andnymph survival and had no apparent toxic effects on rice plants However thiamethoxam-chlorantraniliprole and other neonicotinoids although moderately toxic can have severeeffects on beneficial hymenopteran parasitoids [83] although they have apparently lowtoxicity to predatory mirid bugs [84] The remaining insecticides either directly reducedyields or were associated with higher BPH biomass or densities in our experiments
We noted few synergies with HPR although nymph biomass andor survival waslower in cartap hydrochloride and carbofuran-treated IR62 plants compared to correspond-ing IR64 plants Meanwhile cartap hydrochloride and deltamethrin were antagonisticto resistance in IR62 in the screenhouse experiment Among the strongest effects of HPRwas to act as a buffer against GLH population build-up on deltamethrin-treated IR62 andagainst carbofuran-induced increases in BPH nymph biomass in the survival bioassayTherefore HPR may play a role in reducing the possibilities of secondary outbreaks ofBPH and other insects The combined use of HPR and targeted insecticides against BPHcreates redundancy in the rice field and counters measures to manage insecticide resistanceand virulence by unnecessarily exposing plant protection products to evolving pest pop-ulations HPR (and host plant tolerance) can be effective against target pests as revealedin previous studies [4] and could be incorporated into other pest management practicesparticularly agroecological practices such as ecological engineering with which it is highlycompatible [818586]
5 Conclusions
Based on our results and the results of previous studies HPR and the applicationof chemical insecticides against BPH rarely produce synergistic effects Antagonisticeffects where chemicals caused increases in BPH fitness or populations on resistant ricewere more apparent in our study In particular deltamethrin caused an outbreak ofBPH in our screenhouse study Nevertheless HPR can buffer against the insecticide-induced enhancement of herbivore fitness (hormesis) and potentially reduce possibilities ofsecondary outbreaks as was suggested from results with cartap hydrochloride in our studyBased on our results we suggest that carbofuran cartap hydrochloride cypermethrindeltamethrin and fipronil should be avoided in rice particularly in the Philippines (wherethis research was conducted) due to their resurgence-causing potential or because of directreductions to rice yields
Author Contributions Conceptualization FGH methodology FGH AP-C and MLPA formalanalysis FGH and AP-C investigation FGH AP-C and MLPA data curation FGHAP-C and MLPA writingmdashoriginal draft preparation FGH and AP-C writingmdashreview andediting FGH AP-C and MLPA visualization FGH supervision FGH and MLPA projectadministration FGH funding acquisition FGH All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by the Bill and Melinda Gates Foundation (Cereal SystemsInitiative for South Asia [CSISA] OPP52303) and the Global Rice Science Platform (GRiSP) underthe directorship of Achim Dobermann AP-C was partially funded by The Spanish Governmentthrough Becas Faro International
Crops 2021 1 181
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data presented in this study are available on reasonable requestfrom the corresponding author
Acknowledgments The authors thank Fame Ramal Carmencita Bernal Alberto Naredo ReyeulQuintana Vincent Virtudes Marol Recide Jenyrose Geronda and Ellen Genil for assistance duringthis study The contributions of Rodante Abas (sadly deceased) during three decades of entomologicalresearch at IRRI are gratefully acknowledged Rice lines were supplied through the Genebank of IRRI
Conflicts of Interest The authors declare no conflict of interest
References1 Bottrell DG Schoenly KG Resurrecting the ghost of green revolutions past The brown planthopper as a recurring threat to
high-yielding rice production in tropical Asia J Asia-Pac Entomol 2012 15 122ndash140 [CrossRef]2 Horgan FG Integrated pest management for sustainable rice cultivation A holistic approach In Achieving Sustainable Cultivation
of Rice Cultivation Pest and Disease Management Sasaki T Ed Burleigh Dodds Scientific Cambridge UK 2017 pp 309ndash3423 Horgan FG de Freitas TFS Crisol-Martiacutenez E Mundaca EA Bernal CC Nitrogenous fertilizer reduces resistance but
enhances tolerance to the brown planthopper in fast-growing moderately resistant rice Insects 2021 12 989 [CrossRef] [PubMed]4 Horgan FG Pentildealver Cruz A Bernal CC Ramal AF Almazan MLP Wilby A Resistance and tolerance to the brown
planthopper Nilaparvata lugens (Staringl) in rice infested at different growth stages across a gradient of nitrogen applicationsField Crops Res 2018 217 53ndash65 [CrossRef]
5 Hereward JP Cai X Matias AMA Walter GH Xu C Wang Y Migration dynamics of an important rice pest The brownplanthopper (Nilaparvata lugens) across AsiamdashInsights from population genomics Evol Appl 2020 13 2449ndash2459 [CrossRef]
6 Hu C Guo J Fu X Huang Y Gao X Wu K Seasonal migration pattern of Nilaparvata lugens (Hemiptera Delphacidae) overthe Bohai Sea in northern China J Econ Entomol 2018 111 2129ndash2135 [CrossRef]
7 Ma J Wang Y-C Hu Y-Y Lu M-H Wan G-J Chen F-J Liu W-C Zhai B-P Hu G Brown planthopper Nilaparvatalugens was concentrated at the rear of the typhoon Soudelor in eastern China in August 2015 Insect Sci 2018 25 916ndash926[CrossRef]
8 Fujita D Kohli A Horgan FG Rice resistance to planthoppers and leafhoppers Crit Rev Plant Sci 2013 32 162ndash191[CrossRef]
9 Fujii T Yoshida K Kobayashi T Myint KKM Yasui H Sanada-Morimura S Matsumura M Long-term virulencemonitoring of differential cultivars in Japanrsquos immigrant populations of Nilaparvata lugens (Hemiptera Delphacidae) in 2001ndash2009Appl Entomol Zool 2021 56 407ndash418 [CrossRef]
10 Myint KKM Yasui H Takagi M Matsumura M Virulence of long-term laboratory populations of the brown planthopperNilaparvata lugens (Stal) and whitebacked planthopper Sogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) on rice differentialvarieties Appl Entomol Zool 2009 44 149ndash153 [CrossRef]
11 Horgan FG Ramal AF Bentur JS Kumar R Bhanu KV Sarao PS Iswanto EH Van Chien H Phyu MHBernal CC et al Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia againstresistant rice varieties Crop Prot 2015 78 222ndash231 [CrossRef]
12 Horgan FG Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers Crop Prot2018 110 21ndash33 [CrossRef]
13 Uddin ABMA Islam KS Jahan M Ara A Afrin S Farmerrsquos perception about resurgence of brown planthopper NilaparvataLugens (Staringl) in Bangladesh Bangladesh Rice J 2020 23 21ndash33 [CrossRef]
14 Triwidodo H Brown planthoppers infestations and insecticides use pattern in Java Indonesia AGRIVITA J Agric Sci 202042 320ndash330 [CrossRef]
15 Veres A Wyckhuys KA Kiss J Toacuteth F Burgio G Pons X Avilla C Vidal S Razinger J Bazok R et al An update of theWorldwide Integrated Assessment (WIA) on systemic pesticides Part 4 Alternatives in major cropping systems Environ SciPollut Res 2020 27 29867ndash29899 [CrossRef]
16 Bakker L van der Werf W Tittonell P Wyckhuys KA Bianchi FJ Neonicotinoids in global agriculture Evidence for a newpesticide treadmill Ecol Soc 2020 25 26 [CrossRef]
17 Wu J-C Xu J-X Yuan S-Z Liu J-L Jiang Y-H Xu J-F Pesticide-induced susceptibility of rice to brown planthopperNilaparvata lugens Entomol Exp Appl 2001 100 119ndash126 [CrossRef]
18 Wu J-C Qiu H-M Yang G-Q Liu J-L Liu G-J Wilkens RM Effective duration of pesticide-induced susceptibility of riceto brown planthopper (Nilaparvata lugens Staringl Homoptera Delphacidae) and physiological and biochemical changes in riceplants following pesticide application Int J Pest Manag 2004 50 55ndash62 [CrossRef]
19 Wang A-H Wu J-C Yu Y-S Liu J-L Yue J-F Wang MY Selective insecticide-induced stimulation on fecundity andbiochemical changes in Tryporyza incertulas (Lepidoptera Pyralidae) J Econ Entomol 2005 98 1144ndash1149 [CrossRef] [PubMed]
Crops 2021 1 182
20 Azzam S Wang F Wu J-C Shen J Wang L-P Yang G-Q Guo Y-R Comparisons of stimulatory effects of a seriesof concentrations of four insecticides on reproduction in the rice brown planthopper Nilaparvata lugens Staringl (HomopteraDelphacidae) Int J Pest Manag 2009 55 347ndash358 [CrossRef]
21 Ge L-Q Hu J-H Wu J-C Yang G-Q Gu H Insecticide-induced changes in protein RNA and DNA contents in ovary andfat body of female Nilaparvata lugens (Hemiptera Delphacidae) J Econ Entomol 2009 102 1506ndash1514 [CrossRef]
22 Ling S Zhang J Hu L Zhang R Effect of fipronil on the reproduction feeding and relative fitness of brown planthopperNilaparvata lugens Appl Entomol Zool 2009 44 543ndash548 [CrossRef]
23 Ge L-Q Wang L-P Zhao K-F Wu J-C Huang L-J Mating pair combinations of insecticide-treated male and femaleNilaparvata lugens Staringl (Hemiptera Delphacidae) planthoppers influence protein content in the male accessory glands (MAGs)and vitellin content in both fat bodies and ovaries of adult females Pestic Biochem Physiol 2010 98 279ndash288 [CrossRef]
24 Ge L-Q Wu J-C Zhao K-F Chen Y Yang G-Q Induction of Nlvg and suppression of Nljhe gene expression in Nilaparvatalugens (Staringl) (Hemiptera Delphacidae) adult females and males exposed to two insecticides Pestic Biochem Physiol 201098 269ndash278 [CrossRef]
25 Hu J-H Wu J-C Yin J-L Gu H-N Physiology of insecticide-induced stimulation of reproduction in the rice brownplanthopper (Nilaparvata lugens (Staringl)) Dynamics of protein in fat body and ovary Int J Pest Manag 2010 56 23ndash30 [CrossRef]
26 Wang L-P Shen J Ge L-Q Wu J-C Yang G-Q Jahn GC Insecticide-induced increase in the protein content of maleaccessory glands and its effect on the fecundity of females in the brown planthopper Nilaparvata lugens Staringl (HemipteraDelphacidae) Crop Prot 2010 29 1280ndash1285 [CrossRef]
27 Wu J Ge L Liu F Song Q Stanley D Pesticide-induced planthopper population resurgence in rice cropping systems AnnuRev Entomol 2020 65 409ndash429 [CrossRef]
28 Suri KS Singh G Insecticide-induced resurgence of the whitebacked planthopper Sogatella furcifera (Horvath) (HemipteraDelphacidae) on rice varieties with different levels of resistance Crop Prot 2011 30 118ndash124 [CrossRef]
29 Fabellar LT Heinrichs EA Toxicity of insecticides to predators of rice brown planthoppers Nilaparvata lugens (Staringl) (HomopteraDelphacidae) Environ Entomol 1984 13 832ndash837 [CrossRef]
30 Fabellar LT Heinrichs EA Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators Crop Prot1986 5 254ndash258 [CrossRef]
31 Cuong NL Ben PT Phuong LT Chau LM Cohen MB Effect of host plant resistance and insecticide on brown planthopperNilaparvata lugens (Staringl) and predator population development in the Mekong Delta Vietnam Crop Prot 1997 16 707ndash715[CrossRef]
32 Preetha G Stanley J Suresh S Samiyappan R Risk assessment of insecticides used in rice on mirid bug Cyrtorhinuslividipennis Reuter the important predator of brown planthopper Nilaparvata lugens (Stal) Chemosphere 2010 80 498ndash503[CrossRef] [PubMed]
33 Tanaka K Endo S Kazano H Toxicity of insecticides to predators of rice planthoppers Spiders the mirid bug and the dryinidwasp Appl Entomol Zool 2000 35 177ndash187 [CrossRef]
34 Zhang X Xu Q Lu W Liu F Sublethal effects of four synthetic insecticides on the generalist predator Cyrtorhinus lividipennisJ Pest Sci 2015 88 383ndash392 [CrossRef]
35 Settle WH Ariawan H Astuti ET Cahyana W Hakim AL Hindayana D Lestari AS Managing tropical rice peststhrough conservation of generalist natural enemies and alternative prey Ecology 1996 77 1975ndash1988 [CrossRef]
36 Salim M Heinrichs EA Insecticide-induced changes in the levels of resistance of rice cultivars to the whitebacked planthopperSogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) Crop Prot 1987 6 28ndash32 [CrossRef]
37 Yin J-L Xu H-W Wu J-C Hu J-H Yang G-Q Cultivar and insecticide applications affect the physiological development ofthe brown planthopper Nilaparvata lugens (Staringl) (Hemiptera Delphacidae) Environ Entomol 2014 37 206ndash212 [CrossRef]
38 Cheng C Chang W Studies on varietal resistance to the brown planthopper in Taiwan In Brown Planthopper Threat toRice Production in Asia International Rice Research Institute Ed International Rice Research Institute Manila Philippines1979 pp 251ndash271
39 Reissig WH Heinrichs EA Valencia SL Insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on ricevarieties with different levels of resistance Environ Entomol 1982 11 165ndash168 [CrossRef]
40 Heinrichs EA Fabellar LT Basilio RP Wen T-C Medrano F Susceptibility of rice planthoppers Nilaparvata lugens andSogatella furcifera (Homoptera Delphacidae) to insecticides as influenced by level of resistance in the host plant Environ Entomol1984 13 455ndash458 [CrossRef]
41 Joshi R Shepard B Kenmore P Lydia R Insecticide-induced resurgence of brown planthopper (BPH) on IR62 Int Rice ResNewsl 1992 17 9ndash10
42 Zhu Z-R Cheng J Jiang M-X Zhang X-X Complex influence of rice variety fertilization timing and insecticide onpopulation dynamics of Sogatella furcifera (Horvath) Nilaparvata lugens (Staringl) (Homoptera Delphacidae) and their natural enemiesin rice in Hangzhou China J Pest Sci 2004 77 65ndash74 [CrossRef]
43 Nanthakumar M Jhansi Lakshmi V Shashi Bhushan V Balachandran SM Mohan M Decrease of rice plant resistance andinduction of hormesis and carboxylesterase titre in brown planthopper Nilaparvata lugens (Staringl) by xenobiotics Pestic BiochemPhysiol 2012 102 146ndash152 [CrossRef]
Crops 2021 1 183
44 Yao H-W Jiang C-Y Ye G-Y Cheng J-A Studies on cultivar-induced changes in insecticide susceptibility and its relatedenzyme activities of the white-backed planthopper Sogatella furcifera (Horvath)(Homopter Delphacidae) Insect Sci 2002 9 9ndash15
45 Horgan FG Srinivasan TS Bentur JS Kumar R Bhanu KV Sarao PS Van Chien H Almazan MLP Bernal CCRamal AF et al Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers Implicationsfor rice breeding and gene deployment Agronomy 2017 7 62 [CrossRef]
46 Jairin J Phengrat K Teangdeerith S Vanavichit A Toojinda T Mapping of a broad-spectrum brown planthopper resistancegene Bph3 on rice chromosome 6 Mol Breed 2007 19 35ndash44 [CrossRef]
47 Ren J Gao F Wu X Lu X Zeng L Lv J Su X Luo H Ren G Bph32 a novel gene encoding an unknown SCRdomain-containing protein confers resistance against the brown planthopper in rice Sci Rep 2016 6 37645 [CrossRef]
48 Nguyen CD Zheng S-H Sanada-Morimura S Matsumura M Yasui H Fujita D Substitution mapping and characterizationof brown planthopper resistance genes from indica rice variety lsquoPTB33prime (Oryza sativa L) Breed Sci 2021 in press [CrossRef]
49 Chelliah S Heinrichs EA Factors affecting insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on RiceEnviron Entomol 1980 9 773ndash777 [CrossRef]
50 Moore MT Kroumlger R Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growthArch Environ Contam Toxicol 2010 59 574ndash581 [CrossRef]
51 Kim J Hwang I Han M Jang B Effect of insecticide and fungicide on phytotoxicity of herbicide in rice Korean J Weed Sci1986 6 67ndash75
52 Horgan FG Garcia CPF Haverkort F de Jong PW Ferrater JB Changes in insecticide resistance and host range performanceof planthoppers artificially selected to feed on resistant rice Crop Prot 2020 127 104963 [CrossRef] [PubMed]
53 Khush GS Virk PS IR Varieties and Their Impact International Rice Research Institute Manila Philippines 200554 Srinivasan TS Almazan MLP Bernal CC Ramal AF Subbarayalu MK Horgan FG Interactions between nymphs of
Nilaparvata lugens and Sogatella furcifera (Hemiptera Delphacidae) on resistant and susceptible rice varieties Appl Entomol Zool2016 51 81ndash90 [CrossRef]
55 Pentildealver Cruz A Arida A Heong KL Horgan FG Aspects of brown planthopper adaptation to resistant rice varieties withthe Bph3 gene Entomol Exp Appl 2011 141 245ndash257 [CrossRef]
56 Alam SN Cohen MB Detection and analysis of QTLs for resistance to the brown planthopper Nilaparvata lugens in adoubled-haploid rice population Theor Appl Genet 1998 97 1370ndash1379 [CrossRef]
57 Mackill DJ Khush GS IR64 A high-quality and high-yielding mega variety Rice 2018 11 18 [CrossRef] [PubMed]58 Heinrichs EA Basilio RP Valencia SL Buprofezin a selective insecticide for the management of rice planthoppers (Ho-
moptera Delphacidae) and leafhoppers (Homoptera Cicadellidae) Environ Entomol 1984 13 515ndash521 [CrossRef]59 Satapathy MK Anjaneyulu A Use of cypermethrin a synthetic pyrethroid in the control of rice tungro virus disease and its
vector Trop Pest Manag 1984 30 170ndash178 [CrossRef]60 Vorley W Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown
planthopper (BPH) in Kedah Malaysia Int Rice Res Newsl 1987 10 19ndash2061 Uddin A Islam KS Jahan M Ara A Khan MAI Factor influencing the resurgence of brown planthopper in Bangladesh
SAARC J Agric 2020 18 117ndash128 [CrossRef]62 Escalada MM Luecha M Heong KL Social impacts of planthopper outbreaks Case study from Thailand In Rice Planthoppers
Ecology Management Socio Economics and Policy Heong KL Cheng J Escalada MM Eds Zhejiang University PressHangzhou China 2015 pp 209ndash226
63 Zhang CP He HM Yu JZ Hu XQ Zhu YH Wang Q Residues of carbosulfan and its metabolites carbofuran and3-hydroxy carbofuran in rice field ecosystem in China J Environ Sci Health 2016 51 351ndash357 [CrossRef] [PubMed]
64 Samanta S Barman M Samanta NRA Bio-efficacy trials of carbofuran 3 CG against insect pests of rice J Entomol ZoolStud 2020 8 1529ndash1533
65 Donley N The USA lags behind other agricultural nations in banning harmful pesticides Environ Health 2019 18 44 [CrossRef][PubMed]
66 Adams A Gore J Musser F Cook D Catchot A Walker T Dobbins C Efficacy of selected insecticides applied to hybridrice seed J Econ Entomol 2015 109 200ndash206 [CrossRef]
67 Kumar R Singh B Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) Field Bull Environ Contam Toxicol2013 90 482ndash488 [CrossRef]
68 Dhaka S Prajapati C Singh D Singh R Field evaluation of insecticides and bio-pesticides against rice leaf folder CnaphalocrocisMedinalis (Guenee) Ann Plant Prot Sci 2011 19 324ndash326
69 Matsumura M Takeuchi H Satoh M Sanada-Morimura S Otuka A Watanabe T Van Thanh D Species-specific insecticideresistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in East and South-eastAsia Pest Manag Sci 2008 64 1115ndash1121 [CrossRef] [PubMed]
70 Tang J Li J Shao Y Yang B Liu Z Fipronil resistance in the whitebacked planthopper (Sogatella furcifera) Possible resistancemechanisms and cross-resistance Pest Manag Sci 2010 66 121ndash125 [CrossRef]
71 Horgan FG Arida A Ardestani G Almazan MLP Temperature-dependent oviposition and nymph performance revealdistinct thermal niches of coexisting planthoppers with similar thresholds for development PLoS ONE 2020 15 e0235506[CrossRef] [PubMed]
Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]
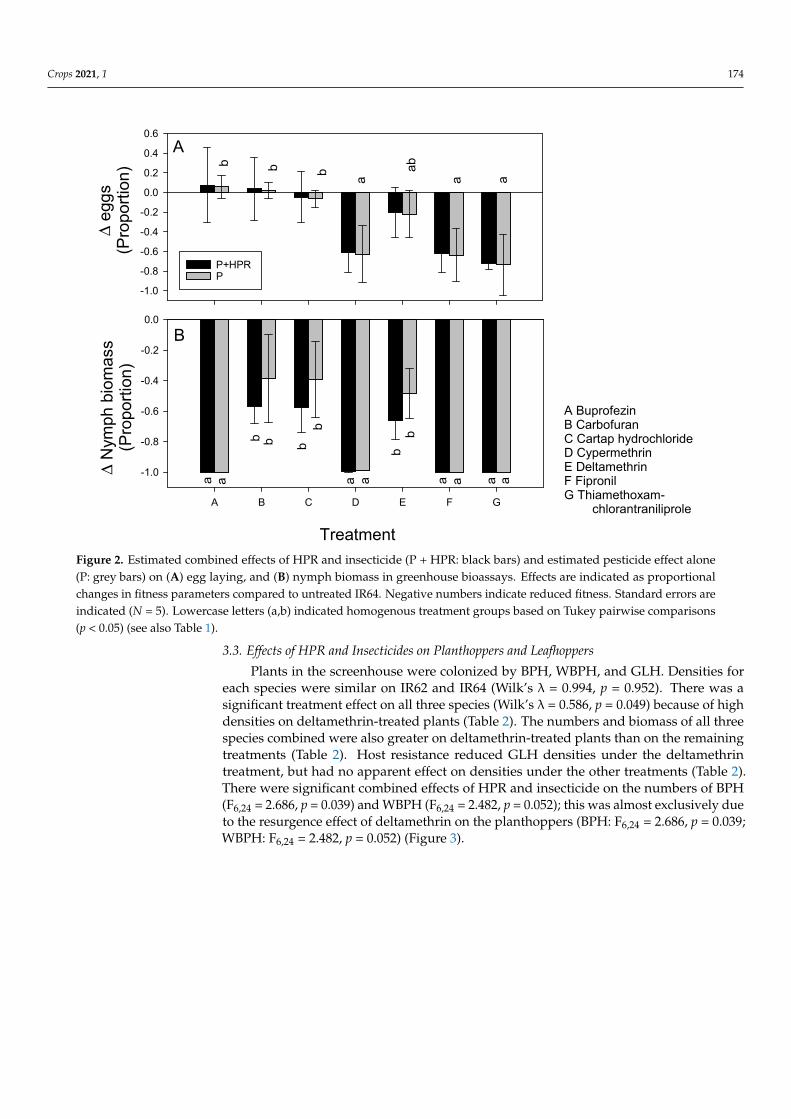
Crops 2021 1 174
Crops 2021 1 FOR PEER REVIEW 9
Figure 2 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) egg laying and (B) nymph biomass in greenhouse bioas-says Effects are indicated as proportional changes in fitness parameters compared to untreated IR64 Negative numbers indicate reduced fitness Standard errors are indicated (N = 5) Lowercase letters (ab) indicated homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 1)
Nymph survival and biomass were not affected by rice variety (Table 1) Buprofezin fipronil thiamethoxam-chlorantraniliprole and cypermethrin reduced survival to 0 in the bioassay and all insecticides except carbofuran resulted in lower survival and biomass than on untreated plants (Table 1) There were significant interactions because of higher nymph mortality and a resulting lower biomass on carbofuran-treated IR62 compared to untreated IR62 but lower mortality and a higher biomass of nymphs on carbofuran-treated IR64 than on untreated IR64 (Table 1) There was no significant effect of insecticide treatment (F628 = 0946 p = 0479) on observed changes in nymph biomass during the bio-assay but the estimated combined effects of pesticide and HPR was greater for thiameth-oxam-chlorantraniliprole fipronil cypermethrin and buprofezin than for cartap hydro-chloride and carbofuran (F628 = 5699 p = 0001) (Figure 2B)
33 Effects of HPR and Insecticides on Planthoppers and Leafhoppers Plants in the screenhouse were colonized by BPH WBPH and GLH Densities for
each species were similar on IR62 and IR64 (Wilkrsquos λ = 0994 p = 0952) There was a sig-nificant treatment effect on all three species (Wilkrsquos λ = 0586 p = 0049) because of high densities on deltamethrin-treated plants (Table 2) The numbers and biomass of all three species combined were also greater on deltamethrin-treated plants than on the remaining treatments (Table 2) Host resistance reduced GLH densities under the deltamethrin treat-ment but had no apparent effect on densities under the other treatments (Table 2) There were significant combined effects of HPR and insecticide on the numbers of BPH (F624 = 2686 p = 0039) and WBPH (F624 = 2482 p = 0052) this was almost exclusively due to the resurgence effect of deltamethrin on the planthoppers (BPH F624 = 2686 p = 0039 WBPH F624 = 2482 p = 0052) (Figure 3)
Δ eg
gs(P
ropo
rtion
)
-10
-08
-06
-04
-02
00
02
04
06
Treatment
A B C D E F G
Δ N
ymph
bio
mas
s(P
ropo
rtion
)
-10
-08
-06
-04
-02
00
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
ab
a a
b
a
b b
a a a a a a a a
b b bb
bb
Figure 2 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone(P grey bars) on (A) egg laying and (B) nymph biomass in greenhouse bioassays Effects are indicated as proportionalchanges in fitness parameters compared to untreated IR64 Negative numbers indicate reduced fitness Standard errors areindicated (N = 5) Lowercase letters (ab) indicated homogenous treatment groups based on Tukey pairwise comparisons(p lt 005) (see also Table 1)
33 Effects of HPR and Insecticides on Planthoppers and Leafhoppers
Plants in the screenhouse were colonized by BPH WBPH and GLH Densities foreach species were similar on IR62 and IR64 (Wilkrsquos λ = 0994 p = 0952) There was asignificant treatment effect on all three species (Wilkrsquos λ = 0586 p = 0049) because of highdensities on deltamethrin-treated plants (Table 2) The numbers and biomass of all threespecies combined were also greater on deltamethrin-treated plants than on the remainingtreatments (Table 2) Host resistance reduced GLH densities under the deltamethrintreatment but had no apparent effect on densities under the other treatments (Table 2)There were significant combined effects of HPR and insecticide on the numbers of BPH(F624 = 2686 p = 0039) and WBPH (F624 = 2482 p = 0052) this was almost exclusively dueto the resurgence effect of deltamethrin on the planthoppers (BPH F624 = 2686 p = 0039WBPH F624 = 2482 p = 0052) (Figure 3)
Crops 2021 1 175
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants underseven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density(Number Plantminus1) 1
WBPH Density(Number Plantminus1) 1
GLH Density(Number Plantminus1) 1
Density ofPlanthoppers and
Leafhoppers(Number Plantminus1) 1
Biomass ofPlanthoppers and
Leafhoppers(mg Dry Weight) 1
IR62Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a
Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartaphydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a
Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a
Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a
Thiamethoxam-chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a
IR64Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartaphydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123
Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns
F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Crops 2021 1 FOR PEER REVIEW 10
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants under seven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density (Num-
ber Plantminus1) 1 WBPH Density
(Number Plantminus1) 1 GLH Density
(Number Plantminus1) 1
Density of Planthoppers and Leafhoppers (Number
Plantminus1) 1
Biomass of Planthoppers and Leafhoppers (mg Dry
Weight) 1 IR62
Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartap hydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a Thiamethoxam-
chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a IR64
Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132 Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartap hydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123 Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113 Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052 Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087 F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns 1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatments and interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in the screenhouse Effects are indicated as proportional changes in planthopper densities compared to untreated IR64 Negative numbers indicate reduced densities positive numbers are increased den-sities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
Δ BP
H(N
umbe
r pla
nt-1
)
0
5
10
15
20
Treatment
A B C D E F G
Δ W
BPH
(Num
ber p
lant
-1)
0
20
30
40
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
a a
aa a a
a a a
a
a a
b
b
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimatedpesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in thescreenhouse Effects are indicated as proportional changes in planthopper densities compared to
Crops 2021 1 176
untreated IR64 Negative numbers indicate reduced densities positive numbers are increaseddensities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenoustreatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
34 Direct and Indirect Effects of Insecticides on Rice Development
IR62 plants had more tillers and attained a greater biomass than IR64 plants in thescreenhouse (Table 3) IR62 grain matured faster and plants were harvested earlier than theIR64 plants (Table 3) IR62 plants also produced more grain (number of grain) and hadhigher yields than IR64 but had smaller grain and more unfilled grain than IR64 (Table 3)
Table 3 Rice growth parameters for IR62 (resistant) and IR64 (susceptible) under seven insecticide treatments and forcontrol non-treated plants in a screenhouse environment Numbers are means plusmn SEM
Treatment Tiller Number 1 Plant Biomass(g Dry Weight) 1
Time to Harvest(Days) 1
Weightof Filled Grain(g Dry Weight) 1
Number ofFilled Grain
(1000s) 1
Unfilled Grain(Proportion) 1
1000 GrainWeight 1
IR62Buprofezin 2660 plusmn 206 ab 4643 plusmn 385 ab 7990 plusmn 168 ab 5776 plusmn 683 ab 361plusmn 061 ab 011 plusmn 003 a 2008 plusmn 042 ab
Carbofuran 2900 plusmn 106 ab 5546 plusmn 357 b 7720 plusmn 080 ab 7448 plusmn 691 b 446plusmn 061 ab 011 plusmn 001 a 2103 plusmn 054 b
Cartaphydrochloride 2820 plusmn 327 ab 4896 plusmn 470 ab 7920 plusmn 153 b 6430 plusmn 712 ab 415plusmn 064 ab 012 plusmn 001 b 2023 plusmn 065 a
Cypermethrin 2870 plusmn 301 a 4957 plusmn 562 a 7760 plusmn 103 b 6032 plusmn 822 a 410plusmn 067 ab 014 plusmn 002 a 1965 plusmn 076 a
Deltamethrin 2620 plusmn 345 a 4592 plusmn 588 a 7840 plusmn 108 ab 5698 plusmn 832 ab 379plusmn 047 ab 012 plusmn 001 a 2030 plusmn 063 ab
Fipronil 2380 plusmn 176 ab 4607 plusmn 657 ab 7700 plusmn 179 ab 5266 plusmn 390 a 318 plusmn 032 a 013 plusmn 002 a 2128 plusmn 047 b
Thiamethoxam-chlorantraniliprole 2650 plusmn 250 b 4547 plusmn 429 b 7720 plusmn 102 a 5777 plusmn 688 ab 408 plusmn 061 b 013 plusmn 002 a 2019 plusmn 067 ab
Control 3120 plusmn 247 b 5401 plusmn 555 b 7720 plusmn 080 ab 7045 plusmn 520 b 396plusmn 036 ab 012 plusmn 002 a 2076 plusmn 050 ab
IR64Buprofezin 2340 plusmn 250 5064 plusmn 747 8350 plusmn 124 5080 plusmn 952 270 plusmn 054 008 plusmn 001 2277 plusmn 039Carbofuran 2040 plusmn 097 4591 plusmn 141 8540 plusmn 073 4951 plusmn 312 263 plusmn 023 006 plusmn 000 2295 plusmn 036
Cartaphydrochloride 2120 plusmn 075 4336 plusmn 285 8520 plusmn 140 4273 plusmn 579 238 plusmn 026 015 plusmn 007 2050 plusmn 234
Cypermethrin 1990 plusmn 089 3968 plusmn 272 8710 plusmn 135 3920 plusmn 552 250 plusmn 028 008 plusmn 002 2171 plusmn 070Deltamethrin 1980 plusmn 181 4325 plusmn 357 8460 plusmn 068 4710 plusmn 496 277 plusmn 024 008 plusmn 001 2314 plusmn 012
Fipronil 2120 plusmn 174 4697 plusmn 311 8450 plusmn 089 4888 plusmn 392 274 plusmn 027 008 plusmn 001 2247 plusmn 042Thiamethoxam-chlorantraniliprole 2520 plusmn 287 5404 plusmn 465 8380 plusmn 196 5594 plusmn 546 347 plusmn 053 010 plusmn 001 2265 plusmn 072
Control 2160 plusmn 076 4678 plusmn 342 8600 plusmn 096 4940 plusmn 543 267 plusmn 029 008 plusmn 001 2283 plusmn 049F-treatment 4100 ns 3410 2253 ns 5150 2555 3923 4924
F-variety 168232 7665 547500 104356 118242 32308 106597 F-interaction 5980 6113 4824 5760 2921 2769 2552
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Insecticides affected plant growth directly and indirectly (through increased planthop-per densities) Plants treated with cypermethrin and deltamethrin had fewer tillers had alower biomass and under cypermethrin had lower yields than untreated plants (Table 3)Plants treated with fipronil also had reduced yields due to lower grain numbers comparedto untreated plants (Table 3) Although yields were not affected treatment with cartaphydrochloride increased the proportion of unfilled grain compared to untreated plantsof both varieties Cartap hydrochloride and cypermethrin also caused a delay in grainmaturation on IR62 and IR64 respectively relative to thiamethoxam-chlorantraniliproletreated plants (Table 3)
There were several significant interactions tiller numbers on both plants were similarunder buprofezin fipronil and thiamethoxam-chlorantraniliprole treatments but wereotherwise higher on IR62 (Table 3) biomass was lower on IR64 under carbofuran andcypermethrin treatments but was otherwise similar on both plants (Table 3) buprofezindelayed grain maturation on IR62 but maturation was otherwise faster on IR62 comparedto IR64 (Table 3) yields under buprofezin fipronil and thiamethoxam-chlorantraniliprolewere similar for IR62 and IR64 but otherwise yields were higher on IR62 (Table 3) grain
Crops 2021 1 177
numbers were higher on IR62 than IR64 except under fipronil treatments (Table 3) theproportions of grain unfilled were higher on IR62 but were similar between varieties underthe fipronil treatment (Table 3) 1000-grain weight was higher in IR64 but similar on bothvarieties when treated with cartap hydrochloride (Table 3)
4 Discussion
Our results using a highly resistant rice variety and some commonly used insecticidesindicate a range of possible effects on planthoppers and rice plants by combining these twomanagement practices
41 Synergies
The most predictable effect of combining HPR and insecticides is a synergy in reduc-ing BPH numbers This is suggested because planthoppers are weakened by the hostrsquosresistance and should therefore be more susceptible to the toxic effects of insecticides [40]Using spray towers over potted plants Heinrichs et al (1984) [40] examined the combinedeffects of a range of insecticides (that included carbofuran) with resistance by comparingBPH mortality on six resistant and two susceptible rice varieties Their results indicatedthat mortality was greatest when insecticides were applied to BPH on resistant varietiesThe effects were similar when the study was repeated with WBPH for three insecticides(including carbofuran) on the resistant variety N22 (Wbph1 gene) [40] In our study hostresistance reduced egg laying and nymph biomass on non-treated IR62 compared to non-treated IR64 Egg laying was also reduced on plants treated with cypermethrin fipronilor thiamethoxam-chlorantraniliprole (Table 1) Based on the results of our choice andno-choice oviposition bioassays cypermethrin acted mainly as a deterrent of gravid fe-males whereas fipronil and thiamethoxam-chlorantraniliprole directly reduced egg layingDespite similar BPH responses to both the insecticides and resistant host there were no ap-parent synergies between any of these insecticides and resistance in reducing egg numbersin the oviposition bioassay In contrast to these three insecticides our bioassays indicatedthat buprofezin carbofuran and cartap hydrochloride can lead to higher numbers of eggson treated plants compared to untreated plants (Table 1) These effects were not statisticallysignificant and may be exaggerated because bioassays were conducted over relativelyshort time scales For example planthoppers will sometimes respond to stresses such ashigh temperatures by increasing oviposition rates [7172] Furthermore the stimulation ofegg-laying by these chemicals was not apparent in our screenhouse experiment (Table 2)Nevertheless our results do indicate that host resistance reduced the magnitude of anystimulatory effect on oviposition egg numbers on buprofezin-treated carbofuran-treatedand cartap hydrochloride-treated IR64 in the no-choice bioassay were twice as high as onthe corresponding IR62 plants In cases such as these the resistance of IR62 functions as abuffer or lsquoinsurancersquo against possible undesired pesticide effects [1238]
Although buprofezin appeared to stimulate oviposition the growth hormone had astrong impact on nymph development leading to zero survival of exposed nymphs in ourgreenhouse bioassay Cypermethrin fipronil and thiamethoxam-chlorantraniliprole alsoreduced nymph survival to zero in the greenhouse bioassay (Table 1) Where insecticideshave such high efficiencies any fitness losses to BPH due to host resistance then becomeredundant Unlike buprofezin carbofuran also stimulated nymph biomass on IR64 in thesurvival bioassay (Table 1) however nymph biomass was lower on carbofuran-treated IR62than on non-treated IR62 indicating that the hostrsquos resistance buffered against undesiredstimulatory effects on BPH and may have functioned synergistically to augment mortalityand slow BPH growth Such buffering allows insecticides to be used against other pestswithout inducing secondary outbreaks of non-target pests In our screenhouse experimentherbivore densities were generally low on both IR62 and IR64 for most treatments (exceptdeltamethrin) and were below threshold densities for insecticide applicationsmdashwhichare normally about 25ndash50 planthoppers per hill [1373] Under such low densities it wasnot possible to observe the impact of host resistance on planthopper population build-
Crops 2021 1 178
up There was some indication of higher densities on cartap hydrochloride-treated IR62in the screenhouse which may be related to the stimulatory effects of the product onegg laying and the low mortality of nymphs on cartap hydrochloride-treated plants asobserved in the greenhouse bioassays (Table 1) Field studies from the Philippines haveindicated that IR62 effectively reduces BPH densities during wet and dry seasons [4]However farmers are rarely aware of which anti-herbivore resistance their rice varietiespossess and frequently apply pesticides regardless of resistance thereby reducing theirfarm profits [3841] For example Joshi et al (1992) [41] reported widespread outbreaks ofBPH on IR62 in Mindanao Philippines after applications of decamethrin (a pyrethroid) tothe resistant variety Where such resurgence-causing insecticides are applied resistanceseems to be often incapable of reducing the stimulatory effects of the insecticide and itloses its capacity to buffer against outbreaks
42 Antagonisms
Several insecticides are associated with increasing densities of planthoppers andwith resurgence outbreaks in farmersrsquo fields [27293041496174] Indeed patches oflsquohopperburnrsquo (ie dead rice plants resulting from planthopper damage) are often anindicator of high insecticide use in tropical Asia [214] In many cases these resurgence-causing insecticides were once effective in reducing planthopper densities but over timepopulations developed resistance against the products The resulting low toxicity canthen have a stimulatory effect on planthopper feeding and development [2123242743]This may be through hormesis which is a direct response by the planthopper to low-levelexposure to the toxic chemical or it may result from physiological changes to the rice plantsthat ultimately benefit the planthopper [1849] In our screenhouse study we induced aresurgence outbreak of BPH WBPH and to a lesser extent GLH on deltamethrin-treatedrice plants (Table 2) The combined densities of all three herbivores increased almost10 times on the treated plants Furthermore herbivore biomass increased by between fourand six times Although the resurgence effects of deltamethrin have been documentedin previous studies [2029ndash313975] we could not predict from our greenhouse studiesthat any such outbreak would occur Egg laying was unaffected by deltamethrin in ourchoice and no-choice experiments and nymph survival and development both declinedon deltamethrin-treated rice in the survival bioassay (Table 1) The massive increases inplanthopper densities on deltamethrin-treated plants in our study seem to result froma heightened attraction of adult BPH and WBPH to the plants [49] particularly sincenumbers were so low on all other plants including both treated and non-treated plantsHowever once on the plants planthoppers also apparently had enhanced fitness Cuonget al (1997) [31] observed higher population densities of BPH on deltamethrin-treatedrice in Vietnam albeit with lower-level increases on resistant varieties Furthermore in agreenhouse study Azzam et al (2009) [20] found that deltamethrin reduced pre-ovipositiontimes increased the duration of oviposition and increased egg hatchability and femalelongevity Nanthakumar et al (2012) [43] further showed that deltamethrin increased BPHfeeding efficiency on resistant and susceptible rice plants as determined through honeydewproduction and population size In studies with WBPH Salim and Heinrichs (1987) [36]showed deltamethrin to increase settling (antixenosis) feeding and population growthand Suri and Singh (2011) [28] showed deltamethrin to increase WBPH reproductive ratesand nymph survival In our study the observed resurgence of planthoppers was similaron both varieties indicating as was previously noted by Joshi et al (1992) [41] thatresurgence-causing pyrethroids are antagonistic to resistance in IR62
Though not statistically significant cartap hydrochloride also increased BPH numberson IR62 but not on IR64 in our screenhouse study This indicates that the product mayalso be antagonistic to the hostrsquos resistance mechanisms The stronger effect on IR62 maybe related to the differences between IR62 biomass growth rates and anatomy (ie alarger plant with higher tillering) compared to IR64 (Table 3) Once resistance is no longereffective because of chemically-enhanced BPH fitness these plant characteristics which
Crops 2021 1 179
have been related to observed preferences in BPH and other sap-sucking insects [349]could further accelerate the build-up of BPH populations on cartap hydrochloride-treatedplants To further test this idea experiments could be conducted using near-isogenic ricelines that differ mainly in the presence or absence of tagged resistance genes [76] Suchtests would avoid confounding differences in plant anatomy between test varieties IR62resistance to BPH has been well studied [4] The variety is also resistant to WBPH atolder plant stages [54] Therefore in our study deltamethrin was also antagonistic tothis WBPH-resistance (Table 2) However BPH feeding also facilitates WBPH populationgrowth [5477] and there may have been added indirect plant-mediated effects [77] thataugmented WBPH populations on both varieties in our experiment IR62 is also resistant toGLH [45] In our experiments GLH densities also increased on deltamethrin-treated plantsbut the effect was only apparent on IR64 Therefore GLH-resistance in IR62 apparentlybuffered against deltamethrin-associated increases in GLH on the variety as was alsonoted from field studies in Vietnam [31] Deltamethrin is currently not recommended foruse in rice [202878] however the product continues to be marketed in rice-dominatedregions throughout Asia [131479] Although cypermethrin has been associated with BPHoutbreaks and was consequently removed for use on rice in Thailand and Vietnam [62]we did not observe cypermethrin-induced resurgence in our screenhouse experimentNevertheless cypermethrin has been associated with resurgence of BPH in South Asia [61]and we have noted stimulatory effects of cypermethrin on other rice herbivores in thePhilippines (Horgan unpublished data)
43 Phytotoxicity
Phytotoxicity is relatively common following applications of insecticides [182850]Phytotoxic effects may change a plantrsquos physiology without consequences for crop yieldsHowever such changes to plant physiology could have indirect effects on herbivores Forexample deltamethrin methyl parathion and quinalphos produce increases in total sugarsand reducing sugars as well as increasing the protein contents of the rice leaf sheath andleaf blade [28] Several insecticides also reduce plant phenols in ricemdashwhich increases therice plantrsquos susceptibility to BPH and other insects [27] In our screenhouse experiment wenoted several changes to rice anatomy and yields Importantly we noted that cypermethrinand fipronil both reduced rice yields (Table 3) This was not due to indirect herbivoreeffects because planthopper and leafhopper densities were consistently low on rice treatedwith these insecticides The effects therefore probably resulted from direct phytotoxiceffects on the plants Furthermore these effects were most pronounced on treated IR62plants Other minor effects on grain production included increased proportions of unfilledgrain in cartap hydrochloride-treated plants and delays in grain maturation on cartaphydrochloride-treated and cypermethrin-treated plants (Table 3) Despite the high densitiesof BPH and WBPH on deltamethrin-treated plants yields were not significantly lower thanin control non-treated plants Both IR62 and IR64 have been noted for their high toleranceto BPH damage particularly under high nitrogenous fertilizer applications [3456] as usedin our screenhouse experiment Therefore where plants have high tolerance to damageapplying insecticides could also result in profitability losses on rice farms and in the caseof insecticide-induced pest population increases tolerance might buffer against subsequentyield losses There are few reports of the phytotoxicity of insecticides to rice carbofuran istoxic to rice at high concentrations but may also enhance the growth of rice plants throughinteractions with nitrogen-fixing bacteria in the rhizosphere [80] Future studies couldmore carefully assess field-grown plants for possible phytotoxic effects particularly ifinsecticides have the potential to directly reduce yields
44 Management implications
Global trends suggest that resistance in rice has failed to reduce insecticide appli-cations [2] This may be due to aggressive marketing by agrochemical companies andstrategies to increase pesticide sales such as promoting prophylactic applications [16] Be-
Crops 2021 1 180
cause we selected our test insecticides (except the pyrethroids) without any a priori criteriaour results suggest that many currently used insecticides probably have some negativeeffects on yield either directly through phytotoxic effects or indirectly by inducing pestresurgence Furthermore our results indicate that even where pests reach high densitiesas on deltamethrin-treated plants in the screenhouse these densities do not necessarilylead to significant yield reductions
Among the products we used buprofezin although it had little effect on oviposi-tion was effective in reducing nymph survival Furthermore there were only minorapparent phytotoxic effects on the rice plants Buprofezin has a low toxicity to naturalenemies in rice fields [58] and is therefore the most compatible of the insecticides withstrategies to increase rice field biodiversity and restore ecosystem services [32ndash348182]Thiamethoxam-chlorantraniliprole was also effective in reducing both oviposition andnymph survival and had no apparent toxic effects on rice plants However thiamethoxam-chlorantraniliprole and other neonicotinoids although moderately toxic can have severeeffects on beneficial hymenopteran parasitoids [83] although they have apparently lowtoxicity to predatory mirid bugs [84] The remaining insecticides either directly reducedyields or were associated with higher BPH biomass or densities in our experiments
We noted few synergies with HPR although nymph biomass andor survival waslower in cartap hydrochloride and carbofuran-treated IR62 plants compared to correspond-ing IR64 plants Meanwhile cartap hydrochloride and deltamethrin were antagonisticto resistance in IR62 in the screenhouse experiment Among the strongest effects of HPRwas to act as a buffer against GLH population build-up on deltamethrin-treated IR62 andagainst carbofuran-induced increases in BPH nymph biomass in the survival bioassayTherefore HPR may play a role in reducing the possibilities of secondary outbreaks ofBPH and other insects The combined use of HPR and targeted insecticides against BPHcreates redundancy in the rice field and counters measures to manage insecticide resistanceand virulence by unnecessarily exposing plant protection products to evolving pest pop-ulations HPR (and host plant tolerance) can be effective against target pests as revealedin previous studies [4] and could be incorporated into other pest management practicesparticularly agroecological practices such as ecological engineering with which it is highlycompatible [818586]
5 Conclusions
Based on our results and the results of previous studies HPR and the applicationof chemical insecticides against BPH rarely produce synergistic effects Antagonisticeffects where chemicals caused increases in BPH fitness or populations on resistant ricewere more apparent in our study In particular deltamethrin caused an outbreak ofBPH in our screenhouse study Nevertheless HPR can buffer against the insecticide-induced enhancement of herbivore fitness (hormesis) and potentially reduce possibilities ofsecondary outbreaks as was suggested from results with cartap hydrochloride in our studyBased on our results we suggest that carbofuran cartap hydrochloride cypermethrindeltamethrin and fipronil should be avoided in rice particularly in the Philippines (wherethis research was conducted) due to their resurgence-causing potential or because of directreductions to rice yields
Author Contributions Conceptualization FGH methodology FGH AP-C and MLPA formalanalysis FGH and AP-C investigation FGH AP-C and MLPA data curation FGHAP-C and MLPA writingmdashoriginal draft preparation FGH and AP-C writingmdashreview andediting FGH AP-C and MLPA visualization FGH supervision FGH and MLPA projectadministration FGH funding acquisition FGH All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by the Bill and Melinda Gates Foundation (Cereal SystemsInitiative for South Asia [CSISA] OPP52303) and the Global Rice Science Platform (GRiSP) underthe directorship of Achim Dobermann AP-C was partially funded by The Spanish Governmentthrough Becas Faro International
Crops 2021 1 181
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data presented in this study are available on reasonable requestfrom the corresponding author
Acknowledgments The authors thank Fame Ramal Carmencita Bernal Alberto Naredo ReyeulQuintana Vincent Virtudes Marol Recide Jenyrose Geronda and Ellen Genil for assistance duringthis study The contributions of Rodante Abas (sadly deceased) during three decades of entomologicalresearch at IRRI are gratefully acknowledged Rice lines were supplied through the Genebank of IRRI
Conflicts of Interest The authors declare no conflict of interest
References1 Bottrell DG Schoenly KG Resurrecting the ghost of green revolutions past The brown planthopper as a recurring threat to
high-yielding rice production in tropical Asia J Asia-Pac Entomol 2012 15 122ndash140 [CrossRef]2 Horgan FG Integrated pest management for sustainable rice cultivation A holistic approach In Achieving Sustainable Cultivation
of Rice Cultivation Pest and Disease Management Sasaki T Ed Burleigh Dodds Scientific Cambridge UK 2017 pp 309ndash3423 Horgan FG de Freitas TFS Crisol-Martiacutenez E Mundaca EA Bernal CC Nitrogenous fertilizer reduces resistance but
enhances tolerance to the brown planthopper in fast-growing moderately resistant rice Insects 2021 12 989 [CrossRef] [PubMed]4 Horgan FG Pentildealver Cruz A Bernal CC Ramal AF Almazan MLP Wilby A Resistance and tolerance to the brown
planthopper Nilaparvata lugens (Staringl) in rice infested at different growth stages across a gradient of nitrogen applicationsField Crops Res 2018 217 53ndash65 [CrossRef]
5 Hereward JP Cai X Matias AMA Walter GH Xu C Wang Y Migration dynamics of an important rice pest The brownplanthopper (Nilaparvata lugens) across AsiamdashInsights from population genomics Evol Appl 2020 13 2449ndash2459 [CrossRef]
6 Hu C Guo J Fu X Huang Y Gao X Wu K Seasonal migration pattern of Nilaparvata lugens (Hemiptera Delphacidae) overthe Bohai Sea in northern China J Econ Entomol 2018 111 2129ndash2135 [CrossRef]
7 Ma J Wang Y-C Hu Y-Y Lu M-H Wan G-J Chen F-J Liu W-C Zhai B-P Hu G Brown planthopper Nilaparvatalugens was concentrated at the rear of the typhoon Soudelor in eastern China in August 2015 Insect Sci 2018 25 916ndash926[CrossRef]
8 Fujita D Kohli A Horgan FG Rice resistance to planthoppers and leafhoppers Crit Rev Plant Sci 2013 32 162ndash191[CrossRef]
9 Fujii T Yoshida K Kobayashi T Myint KKM Yasui H Sanada-Morimura S Matsumura M Long-term virulencemonitoring of differential cultivars in Japanrsquos immigrant populations of Nilaparvata lugens (Hemiptera Delphacidae) in 2001ndash2009Appl Entomol Zool 2021 56 407ndash418 [CrossRef]
10 Myint KKM Yasui H Takagi M Matsumura M Virulence of long-term laboratory populations of the brown planthopperNilaparvata lugens (Stal) and whitebacked planthopper Sogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) on rice differentialvarieties Appl Entomol Zool 2009 44 149ndash153 [CrossRef]
11 Horgan FG Ramal AF Bentur JS Kumar R Bhanu KV Sarao PS Iswanto EH Van Chien H Phyu MHBernal CC et al Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia againstresistant rice varieties Crop Prot 2015 78 222ndash231 [CrossRef]
12 Horgan FG Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers Crop Prot2018 110 21ndash33 [CrossRef]
13 Uddin ABMA Islam KS Jahan M Ara A Afrin S Farmerrsquos perception about resurgence of brown planthopper NilaparvataLugens (Staringl) in Bangladesh Bangladesh Rice J 2020 23 21ndash33 [CrossRef]
14 Triwidodo H Brown planthoppers infestations and insecticides use pattern in Java Indonesia AGRIVITA J Agric Sci 202042 320ndash330 [CrossRef]
15 Veres A Wyckhuys KA Kiss J Toacuteth F Burgio G Pons X Avilla C Vidal S Razinger J Bazok R et al An update of theWorldwide Integrated Assessment (WIA) on systemic pesticides Part 4 Alternatives in major cropping systems Environ SciPollut Res 2020 27 29867ndash29899 [CrossRef]
16 Bakker L van der Werf W Tittonell P Wyckhuys KA Bianchi FJ Neonicotinoids in global agriculture Evidence for a newpesticide treadmill Ecol Soc 2020 25 26 [CrossRef]
17 Wu J-C Xu J-X Yuan S-Z Liu J-L Jiang Y-H Xu J-F Pesticide-induced susceptibility of rice to brown planthopperNilaparvata lugens Entomol Exp Appl 2001 100 119ndash126 [CrossRef]
18 Wu J-C Qiu H-M Yang G-Q Liu J-L Liu G-J Wilkens RM Effective duration of pesticide-induced susceptibility of riceto brown planthopper (Nilaparvata lugens Staringl Homoptera Delphacidae) and physiological and biochemical changes in riceplants following pesticide application Int J Pest Manag 2004 50 55ndash62 [CrossRef]
19 Wang A-H Wu J-C Yu Y-S Liu J-L Yue J-F Wang MY Selective insecticide-induced stimulation on fecundity andbiochemical changes in Tryporyza incertulas (Lepidoptera Pyralidae) J Econ Entomol 2005 98 1144ndash1149 [CrossRef] [PubMed]
Crops 2021 1 182
20 Azzam S Wang F Wu J-C Shen J Wang L-P Yang G-Q Guo Y-R Comparisons of stimulatory effects of a seriesof concentrations of four insecticides on reproduction in the rice brown planthopper Nilaparvata lugens Staringl (HomopteraDelphacidae) Int J Pest Manag 2009 55 347ndash358 [CrossRef]
21 Ge L-Q Hu J-H Wu J-C Yang G-Q Gu H Insecticide-induced changes in protein RNA and DNA contents in ovary andfat body of female Nilaparvata lugens (Hemiptera Delphacidae) J Econ Entomol 2009 102 1506ndash1514 [CrossRef]
22 Ling S Zhang J Hu L Zhang R Effect of fipronil on the reproduction feeding and relative fitness of brown planthopperNilaparvata lugens Appl Entomol Zool 2009 44 543ndash548 [CrossRef]
23 Ge L-Q Wang L-P Zhao K-F Wu J-C Huang L-J Mating pair combinations of insecticide-treated male and femaleNilaparvata lugens Staringl (Hemiptera Delphacidae) planthoppers influence protein content in the male accessory glands (MAGs)and vitellin content in both fat bodies and ovaries of adult females Pestic Biochem Physiol 2010 98 279ndash288 [CrossRef]
24 Ge L-Q Wu J-C Zhao K-F Chen Y Yang G-Q Induction of Nlvg and suppression of Nljhe gene expression in Nilaparvatalugens (Staringl) (Hemiptera Delphacidae) adult females and males exposed to two insecticides Pestic Biochem Physiol 201098 269ndash278 [CrossRef]
25 Hu J-H Wu J-C Yin J-L Gu H-N Physiology of insecticide-induced stimulation of reproduction in the rice brownplanthopper (Nilaparvata lugens (Staringl)) Dynamics of protein in fat body and ovary Int J Pest Manag 2010 56 23ndash30 [CrossRef]
26 Wang L-P Shen J Ge L-Q Wu J-C Yang G-Q Jahn GC Insecticide-induced increase in the protein content of maleaccessory glands and its effect on the fecundity of females in the brown planthopper Nilaparvata lugens Staringl (HemipteraDelphacidae) Crop Prot 2010 29 1280ndash1285 [CrossRef]
27 Wu J Ge L Liu F Song Q Stanley D Pesticide-induced planthopper population resurgence in rice cropping systems AnnuRev Entomol 2020 65 409ndash429 [CrossRef]
28 Suri KS Singh G Insecticide-induced resurgence of the whitebacked planthopper Sogatella furcifera (Horvath) (HemipteraDelphacidae) on rice varieties with different levels of resistance Crop Prot 2011 30 118ndash124 [CrossRef]
29 Fabellar LT Heinrichs EA Toxicity of insecticides to predators of rice brown planthoppers Nilaparvata lugens (Staringl) (HomopteraDelphacidae) Environ Entomol 1984 13 832ndash837 [CrossRef]
30 Fabellar LT Heinrichs EA Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators Crop Prot1986 5 254ndash258 [CrossRef]
31 Cuong NL Ben PT Phuong LT Chau LM Cohen MB Effect of host plant resistance and insecticide on brown planthopperNilaparvata lugens (Staringl) and predator population development in the Mekong Delta Vietnam Crop Prot 1997 16 707ndash715[CrossRef]
32 Preetha G Stanley J Suresh S Samiyappan R Risk assessment of insecticides used in rice on mirid bug Cyrtorhinuslividipennis Reuter the important predator of brown planthopper Nilaparvata lugens (Stal) Chemosphere 2010 80 498ndash503[CrossRef] [PubMed]
33 Tanaka K Endo S Kazano H Toxicity of insecticides to predators of rice planthoppers Spiders the mirid bug and the dryinidwasp Appl Entomol Zool 2000 35 177ndash187 [CrossRef]
34 Zhang X Xu Q Lu W Liu F Sublethal effects of four synthetic insecticides on the generalist predator Cyrtorhinus lividipennisJ Pest Sci 2015 88 383ndash392 [CrossRef]
35 Settle WH Ariawan H Astuti ET Cahyana W Hakim AL Hindayana D Lestari AS Managing tropical rice peststhrough conservation of generalist natural enemies and alternative prey Ecology 1996 77 1975ndash1988 [CrossRef]
36 Salim M Heinrichs EA Insecticide-induced changes in the levels of resistance of rice cultivars to the whitebacked planthopperSogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) Crop Prot 1987 6 28ndash32 [CrossRef]
37 Yin J-L Xu H-W Wu J-C Hu J-H Yang G-Q Cultivar and insecticide applications affect the physiological development ofthe brown planthopper Nilaparvata lugens (Staringl) (Hemiptera Delphacidae) Environ Entomol 2014 37 206ndash212 [CrossRef]
38 Cheng C Chang W Studies on varietal resistance to the brown planthopper in Taiwan In Brown Planthopper Threat toRice Production in Asia International Rice Research Institute Ed International Rice Research Institute Manila Philippines1979 pp 251ndash271
39 Reissig WH Heinrichs EA Valencia SL Insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on ricevarieties with different levels of resistance Environ Entomol 1982 11 165ndash168 [CrossRef]
40 Heinrichs EA Fabellar LT Basilio RP Wen T-C Medrano F Susceptibility of rice planthoppers Nilaparvata lugens andSogatella furcifera (Homoptera Delphacidae) to insecticides as influenced by level of resistance in the host plant Environ Entomol1984 13 455ndash458 [CrossRef]
41 Joshi R Shepard B Kenmore P Lydia R Insecticide-induced resurgence of brown planthopper (BPH) on IR62 Int Rice ResNewsl 1992 17 9ndash10
42 Zhu Z-R Cheng J Jiang M-X Zhang X-X Complex influence of rice variety fertilization timing and insecticide onpopulation dynamics of Sogatella furcifera (Horvath) Nilaparvata lugens (Staringl) (Homoptera Delphacidae) and their natural enemiesin rice in Hangzhou China J Pest Sci 2004 77 65ndash74 [CrossRef]
43 Nanthakumar M Jhansi Lakshmi V Shashi Bhushan V Balachandran SM Mohan M Decrease of rice plant resistance andinduction of hormesis and carboxylesterase titre in brown planthopper Nilaparvata lugens (Staringl) by xenobiotics Pestic BiochemPhysiol 2012 102 146ndash152 [CrossRef]
Crops 2021 1 183
44 Yao H-W Jiang C-Y Ye G-Y Cheng J-A Studies on cultivar-induced changes in insecticide susceptibility and its relatedenzyme activities of the white-backed planthopper Sogatella furcifera (Horvath)(Homopter Delphacidae) Insect Sci 2002 9 9ndash15
45 Horgan FG Srinivasan TS Bentur JS Kumar R Bhanu KV Sarao PS Van Chien H Almazan MLP Bernal CCRamal AF et al Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers Implicationsfor rice breeding and gene deployment Agronomy 2017 7 62 [CrossRef]
46 Jairin J Phengrat K Teangdeerith S Vanavichit A Toojinda T Mapping of a broad-spectrum brown planthopper resistancegene Bph3 on rice chromosome 6 Mol Breed 2007 19 35ndash44 [CrossRef]
47 Ren J Gao F Wu X Lu X Zeng L Lv J Su X Luo H Ren G Bph32 a novel gene encoding an unknown SCRdomain-containing protein confers resistance against the brown planthopper in rice Sci Rep 2016 6 37645 [CrossRef]
48 Nguyen CD Zheng S-H Sanada-Morimura S Matsumura M Yasui H Fujita D Substitution mapping and characterizationof brown planthopper resistance genes from indica rice variety lsquoPTB33prime (Oryza sativa L) Breed Sci 2021 in press [CrossRef]
49 Chelliah S Heinrichs EA Factors affecting insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on RiceEnviron Entomol 1980 9 773ndash777 [CrossRef]
50 Moore MT Kroumlger R Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growthArch Environ Contam Toxicol 2010 59 574ndash581 [CrossRef]
51 Kim J Hwang I Han M Jang B Effect of insecticide and fungicide on phytotoxicity of herbicide in rice Korean J Weed Sci1986 6 67ndash75
52 Horgan FG Garcia CPF Haverkort F de Jong PW Ferrater JB Changes in insecticide resistance and host range performanceof planthoppers artificially selected to feed on resistant rice Crop Prot 2020 127 104963 [CrossRef] [PubMed]
53 Khush GS Virk PS IR Varieties and Their Impact International Rice Research Institute Manila Philippines 200554 Srinivasan TS Almazan MLP Bernal CC Ramal AF Subbarayalu MK Horgan FG Interactions between nymphs of
Nilaparvata lugens and Sogatella furcifera (Hemiptera Delphacidae) on resistant and susceptible rice varieties Appl Entomol Zool2016 51 81ndash90 [CrossRef]
55 Pentildealver Cruz A Arida A Heong KL Horgan FG Aspects of brown planthopper adaptation to resistant rice varieties withthe Bph3 gene Entomol Exp Appl 2011 141 245ndash257 [CrossRef]
56 Alam SN Cohen MB Detection and analysis of QTLs for resistance to the brown planthopper Nilaparvata lugens in adoubled-haploid rice population Theor Appl Genet 1998 97 1370ndash1379 [CrossRef]
57 Mackill DJ Khush GS IR64 A high-quality and high-yielding mega variety Rice 2018 11 18 [CrossRef] [PubMed]58 Heinrichs EA Basilio RP Valencia SL Buprofezin a selective insecticide for the management of rice planthoppers (Ho-
moptera Delphacidae) and leafhoppers (Homoptera Cicadellidae) Environ Entomol 1984 13 515ndash521 [CrossRef]59 Satapathy MK Anjaneyulu A Use of cypermethrin a synthetic pyrethroid in the control of rice tungro virus disease and its
vector Trop Pest Manag 1984 30 170ndash178 [CrossRef]60 Vorley W Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown
planthopper (BPH) in Kedah Malaysia Int Rice Res Newsl 1987 10 19ndash2061 Uddin A Islam KS Jahan M Ara A Khan MAI Factor influencing the resurgence of brown planthopper in Bangladesh
SAARC J Agric 2020 18 117ndash128 [CrossRef]62 Escalada MM Luecha M Heong KL Social impacts of planthopper outbreaks Case study from Thailand In Rice Planthoppers
Ecology Management Socio Economics and Policy Heong KL Cheng J Escalada MM Eds Zhejiang University PressHangzhou China 2015 pp 209ndash226
63 Zhang CP He HM Yu JZ Hu XQ Zhu YH Wang Q Residues of carbosulfan and its metabolites carbofuran and3-hydroxy carbofuran in rice field ecosystem in China J Environ Sci Health 2016 51 351ndash357 [CrossRef] [PubMed]
64 Samanta S Barman M Samanta NRA Bio-efficacy trials of carbofuran 3 CG against insect pests of rice J Entomol ZoolStud 2020 8 1529ndash1533
65 Donley N The USA lags behind other agricultural nations in banning harmful pesticides Environ Health 2019 18 44 [CrossRef][PubMed]
66 Adams A Gore J Musser F Cook D Catchot A Walker T Dobbins C Efficacy of selected insecticides applied to hybridrice seed J Econ Entomol 2015 109 200ndash206 [CrossRef]
67 Kumar R Singh B Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) Field Bull Environ Contam Toxicol2013 90 482ndash488 [CrossRef]
68 Dhaka S Prajapati C Singh D Singh R Field evaluation of insecticides and bio-pesticides against rice leaf folder CnaphalocrocisMedinalis (Guenee) Ann Plant Prot Sci 2011 19 324ndash326
69 Matsumura M Takeuchi H Satoh M Sanada-Morimura S Otuka A Watanabe T Van Thanh D Species-specific insecticideresistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in East and South-eastAsia Pest Manag Sci 2008 64 1115ndash1121 [CrossRef] [PubMed]
70 Tang J Li J Shao Y Yang B Liu Z Fipronil resistance in the whitebacked planthopper (Sogatella furcifera) Possible resistancemechanisms and cross-resistance Pest Manag Sci 2010 66 121ndash125 [CrossRef]
71 Horgan FG Arida A Ardestani G Almazan MLP Temperature-dependent oviposition and nymph performance revealdistinct thermal niches of coexisting planthoppers with similar thresholds for development PLoS ONE 2020 15 e0235506[CrossRef] [PubMed]
Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]
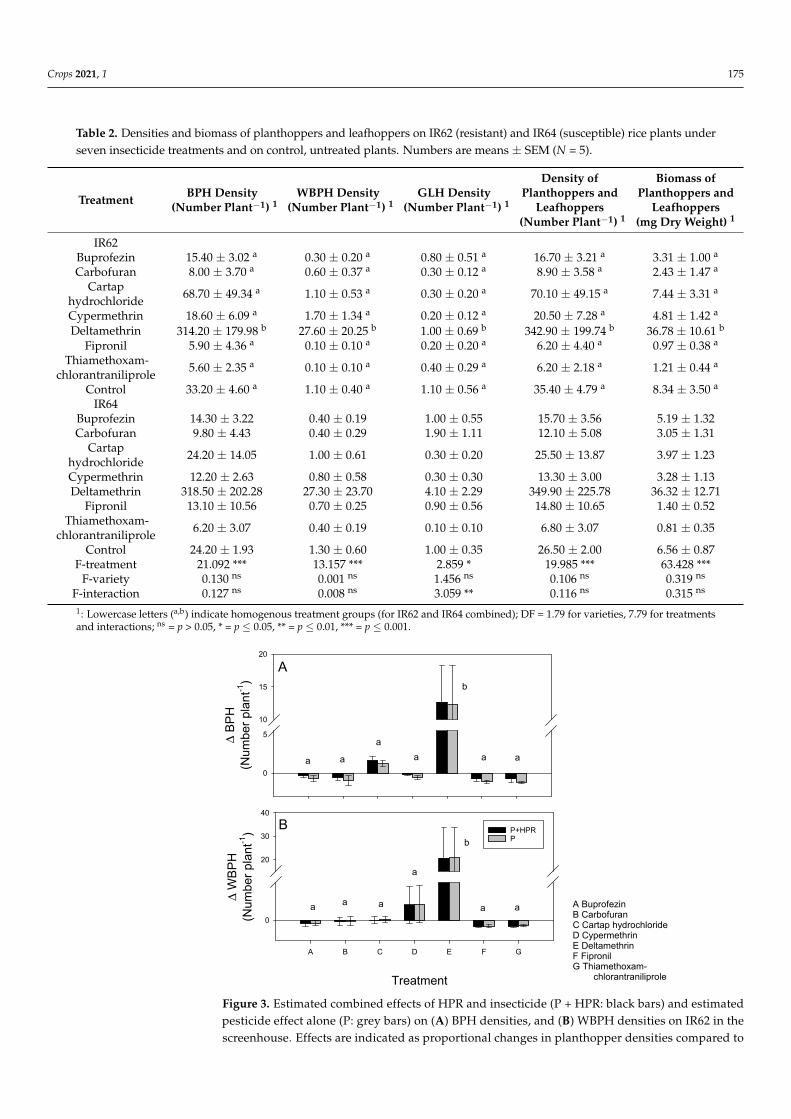
Crops 2021 1 175
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants underseven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density(Number Plantminus1) 1
WBPH Density(Number Plantminus1) 1
GLH Density(Number Plantminus1) 1
Density ofPlanthoppers and
Leafhoppers(Number Plantminus1) 1
Biomass ofPlanthoppers and
Leafhoppers(mg Dry Weight) 1
IR62Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a
Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartaphydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a
Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a
Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a
Thiamethoxam-chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a
IR64Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartaphydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123
Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns
F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Crops 2021 1 FOR PEER REVIEW 10
Table 2 Densities and biomass of planthoppers and leafhoppers on IR62 (resistant) and IR64 (susceptible) rice plants under seven insecticide treatments and on control untreated plants Numbers are means plusmn SEM (N = 5)
Treatment BPH Density (Num-
ber Plantminus1) 1 WBPH Density
(Number Plantminus1) 1 GLH Density
(Number Plantminus1) 1
Density of Planthoppers and Leafhoppers (Number
Plantminus1) 1
Biomass of Planthoppers and Leafhoppers (mg Dry
Weight) 1 IR62
Buprofezin 1540 plusmn 302 a 030 plusmn 020 a 080 plusmn 051 a 1670 plusmn 321 a 331 plusmn 100 a Carbofuran 800 plusmn 370 a 060 plusmn 037 a 030 plusmn 012 a 890 plusmn 358 a 243 plusmn 147 a
Cartap hydrochloride 6870 plusmn 4934 a 110 plusmn 053 a 030 plusmn 020 a 7010 plusmn 4915 a 744 plusmn 331 a Cypermethrin 1860 plusmn 609 a 170 plusmn 134 a 020 plusmn 012 a 2050 plusmn 728 a 481 plusmn 142 a Deltamethrin 31420 plusmn 17998 b 2760 plusmn 2025 b 100 plusmn 069 b 34290 plusmn 19974 b 3678 plusmn 1061 b
Fipronil 590 plusmn 436 a 010 plusmn 010 a 020 plusmn 020 a 620 plusmn 440 a 097 plusmn 038 a Thiamethoxam-
chlorantraniliprole 560 plusmn 235 a 010 plusmn 010 a 040 plusmn 029 a 620 plusmn 218 a 121 plusmn 044 a
Control 3320 plusmn 460 a 110 plusmn 040 a 110 plusmn 056 a 3540 plusmn 479 a 834 plusmn 350 a IR64
Buprofezin 1430 plusmn 322 040 plusmn 019 100 plusmn 055 1570 plusmn 356 519 plusmn 132 Carbofuran 980 plusmn 443 040 plusmn 029 190 plusmn 111 1210 plusmn 508 305 plusmn 131
Cartap hydrochloride 2420 plusmn 1405 100 plusmn 061 030 plusmn 020 2550 plusmn 1387 397 plusmn 123 Cypermethrin 1220 plusmn 263 080 plusmn 058 030 plusmn 030 1330 plusmn 300 328 plusmn 113 Deltamethrin 31850 plusmn 20228 2730 plusmn 2370 410 plusmn 229 34990 plusmn 22578 3632 plusmn 1271
Fipronil 1310 plusmn 1056 070 plusmn 025 090 plusmn 056 1480 plusmn 1065 140 plusmn 052 Thiamethoxam-
chlorantraniliprole 620 plusmn 307 040 plusmn 019 010 plusmn 010 680 plusmn 307 081 plusmn 035
Control 2420 plusmn 193 130 plusmn 060 100 plusmn 035 2650 plusmn 200 656 plusmn 087 F-treatment 21092 13157 2859 19985 63428
F-variety 0130 ns 0001 ns 1456 ns 0106 ns 0319 ns F-interaction 0127 ns 0008 ns 3059 0116 ns 0315 ns 1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatments and interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimated pesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in the screenhouse Effects are indicated as proportional changes in planthopper densities compared to untreated IR64 Negative numbers indicate reduced densities positive numbers are increased den-sities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenous treatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
Δ BP
H(N
umbe
r pla
nt-1
)
0
5
10
15
20
Treatment
A B C D E F G
Δ W
BPH
(Num
ber p
lant
-1)
0
20
30
40
P+HPR P
A BuprofezinB CarbofuranC Cartap hydrochlorideD CypermethrinE DeltamethrinF FipronilG Thiamethoxam- chlorantraniliprole
A
B
a a
aa a a
a a a
a
a a
b
b
Figure 3 Estimated combined effects of HPR and insecticide (P + HPR black bars) and estimatedpesticide effect alone (P grey bars) on (A) BPH densities and (B) WBPH densities on IR62 in thescreenhouse Effects are indicated as proportional changes in planthopper densities compared to
Crops 2021 1 176
untreated IR64 Negative numbers indicate reduced densities positive numbers are increaseddensities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenoustreatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
34 Direct and Indirect Effects of Insecticides on Rice Development
IR62 plants had more tillers and attained a greater biomass than IR64 plants in thescreenhouse (Table 3) IR62 grain matured faster and plants were harvested earlier than theIR64 plants (Table 3) IR62 plants also produced more grain (number of grain) and hadhigher yields than IR64 but had smaller grain and more unfilled grain than IR64 (Table 3)
Table 3 Rice growth parameters for IR62 (resistant) and IR64 (susceptible) under seven insecticide treatments and forcontrol non-treated plants in a screenhouse environment Numbers are means plusmn SEM
Treatment Tiller Number 1 Plant Biomass(g Dry Weight) 1
Time to Harvest(Days) 1
Weightof Filled Grain(g Dry Weight) 1
Number ofFilled Grain
(1000s) 1
Unfilled Grain(Proportion) 1
1000 GrainWeight 1
IR62Buprofezin 2660 plusmn 206 ab 4643 plusmn 385 ab 7990 plusmn 168 ab 5776 plusmn 683 ab 361plusmn 061 ab 011 plusmn 003 a 2008 plusmn 042 ab
Carbofuran 2900 plusmn 106 ab 5546 plusmn 357 b 7720 plusmn 080 ab 7448 plusmn 691 b 446plusmn 061 ab 011 plusmn 001 a 2103 plusmn 054 b
Cartaphydrochloride 2820 plusmn 327 ab 4896 plusmn 470 ab 7920 plusmn 153 b 6430 plusmn 712 ab 415plusmn 064 ab 012 plusmn 001 b 2023 plusmn 065 a
Cypermethrin 2870 plusmn 301 a 4957 plusmn 562 a 7760 plusmn 103 b 6032 plusmn 822 a 410plusmn 067 ab 014 plusmn 002 a 1965 plusmn 076 a
Deltamethrin 2620 plusmn 345 a 4592 plusmn 588 a 7840 plusmn 108 ab 5698 plusmn 832 ab 379plusmn 047 ab 012 plusmn 001 a 2030 plusmn 063 ab
Fipronil 2380 plusmn 176 ab 4607 plusmn 657 ab 7700 plusmn 179 ab 5266 plusmn 390 a 318 plusmn 032 a 013 plusmn 002 a 2128 plusmn 047 b
Thiamethoxam-chlorantraniliprole 2650 plusmn 250 b 4547 plusmn 429 b 7720 plusmn 102 a 5777 plusmn 688 ab 408 plusmn 061 b 013 plusmn 002 a 2019 plusmn 067 ab
Control 3120 plusmn 247 b 5401 plusmn 555 b 7720 plusmn 080 ab 7045 plusmn 520 b 396plusmn 036 ab 012 plusmn 002 a 2076 plusmn 050 ab
IR64Buprofezin 2340 plusmn 250 5064 plusmn 747 8350 plusmn 124 5080 plusmn 952 270 plusmn 054 008 plusmn 001 2277 plusmn 039Carbofuran 2040 plusmn 097 4591 plusmn 141 8540 plusmn 073 4951 plusmn 312 263 plusmn 023 006 plusmn 000 2295 plusmn 036
Cartaphydrochloride 2120 plusmn 075 4336 plusmn 285 8520 plusmn 140 4273 plusmn 579 238 plusmn 026 015 plusmn 007 2050 plusmn 234
Cypermethrin 1990 plusmn 089 3968 plusmn 272 8710 plusmn 135 3920 plusmn 552 250 plusmn 028 008 plusmn 002 2171 plusmn 070Deltamethrin 1980 plusmn 181 4325 plusmn 357 8460 plusmn 068 4710 plusmn 496 277 plusmn 024 008 plusmn 001 2314 plusmn 012
Fipronil 2120 plusmn 174 4697 plusmn 311 8450 plusmn 089 4888 plusmn 392 274 plusmn 027 008 plusmn 001 2247 plusmn 042Thiamethoxam-chlorantraniliprole 2520 plusmn 287 5404 plusmn 465 8380 plusmn 196 5594 plusmn 546 347 plusmn 053 010 plusmn 001 2265 plusmn 072
Control 2160 plusmn 076 4678 plusmn 342 8600 plusmn 096 4940 plusmn 543 267 plusmn 029 008 plusmn 001 2283 plusmn 049F-treatment 4100 ns 3410 2253 ns 5150 2555 3923 4924
F-variety 168232 7665 547500 104356 118242 32308 106597 F-interaction 5980 6113 4824 5760 2921 2769 2552
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Insecticides affected plant growth directly and indirectly (through increased planthop-per densities) Plants treated with cypermethrin and deltamethrin had fewer tillers had alower biomass and under cypermethrin had lower yields than untreated plants (Table 3)Plants treated with fipronil also had reduced yields due to lower grain numbers comparedto untreated plants (Table 3) Although yields were not affected treatment with cartaphydrochloride increased the proportion of unfilled grain compared to untreated plantsof both varieties Cartap hydrochloride and cypermethrin also caused a delay in grainmaturation on IR62 and IR64 respectively relative to thiamethoxam-chlorantraniliproletreated plants (Table 3)
There were several significant interactions tiller numbers on both plants were similarunder buprofezin fipronil and thiamethoxam-chlorantraniliprole treatments but wereotherwise higher on IR62 (Table 3) biomass was lower on IR64 under carbofuran andcypermethrin treatments but was otherwise similar on both plants (Table 3) buprofezindelayed grain maturation on IR62 but maturation was otherwise faster on IR62 comparedto IR64 (Table 3) yields under buprofezin fipronil and thiamethoxam-chlorantraniliprolewere similar for IR62 and IR64 but otherwise yields were higher on IR62 (Table 3) grain
Crops 2021 1 177
numbers were higher on IR62 than IR64 except under fipronil treatments (Table 3) theproportions of grain unfilled were higher on IR62 but were similar between varieties underthe fipronil treatment (Table 3) 1000-grain weight was higher in IR64 but similar on bothvarieties when treated with cartap hydrochloride (Table 3)
4 Discussion
Our results using a highly resistant rice variety and some commonly used insecticidesindicate a range of possible effects on planthoppers and rice plants by combining these twomanagement practices
41 Synergies
The most predictable effect of combining HPR and insecticides is a synergy in reduc-ing BPH numbers This is suggested because planthoppers are weakened by the hostrsquosresistance and should therefore be more susceptible to the toxic effects of insecticides [40]Using spray towers over potted plants Heinrichs et al (1984) [40] examined the combinedeffects of a range of insecticides (that included carbofuran) with resistance by comparingBPH mortality on six resistant and two susceptible rice varieties Their results indicatedthat mortality was greatest when insecticides were applied to BPH on resistant varietiesThe effects were similar when the study was repeated with WBPH for three insecticides(including carbofuran) on the resistant variety N22 (Wbph1 gene) [40] In our study hostresistance reduced egg laying and nymph biomass on non-treated IR62 compared to non-treated IR64 Egg laying was also reduced on plants treated with cypermethrin fipronilor thiamethoxam-chlorantraniliprole (Table 1) Based on the results of our choice andno-choice oviposition bioassays cypermethrin acted mainly as a deterrent of gravid fe-males whereas fipronil and thiamethoxam-chlorantraniliprole directly reduced egg layingDespite similar BPH responses to both the insecticides and resistant host there were no ap-parent synergies between any of these insecticides and resistance in reducing egg numbersin the oviposition bioassay In contrast to these three insecticides our bioassays indicatedthat buprofezin carbofuran and cartap hydrochloride can lead to higher numbers of eggson treated plants compared to untreated plants (Table 1) These effects were not statisticallysignificant and may be exaggerated because bioassays were conducted over relativelyshort time scales For example planthoppers will sometimes respond to stresses such ashigh temperatures by increasing oviposition rates [7172] Furthermore the stimulation ofegg-laying by these chemicals was not apparent in our screenhouse experiment (Table 2)Nevertheless our results do indicate that host resistance reduced the magnitude of anystimulatory effect on oviposition egg numbers on buprofezin-treated carbofuran-treatedand cartap hydrochloride-treated IR64 in the no-choice bioassay were twice as high as onthe corresponding IR62 plants In cases such as these the resistance of IR62 functions as abuffer or lsquoinsurancersquo against possible undesired pesticide effects [1238]
Although buprofezin appeared to stimulate oviposition the growth hormone had astrong impact on nymph development leading to zero survival of exposed nymphs in ourgreenhouse bioassay Cypermethrin fipronil and thiamethoxam-chlorantraniliprole alsoreduced nymph survival to zero in the greenhouse bioassay (Table 1) Where insecticideshave such high efficiencies any fitness losses to BPH due to host resistance then becomeredundant Unlike buprofezin carbofuran also stimulated nymph biomass on IR64 in thesurvival bioassay (Table 1) however nymph biomass was lower on carbofuran-treated IR62than on non-treated IR62 indicating that the hostrsquos resistance buffered against undesiredstimulatory effects on BPH and may have functioned synergistically to augment mortalityand slow BPH growth Such buffering allows insecticides to be used against other pestswithout inducing secondary outbreaks of non-target pests In our screenhouse experimentherbivore densities were generally low on both IR62 and IR64 for most treatments (exceptdeltamethrin) and were below threshold densities for insecticide applicationsmdashwhichare normally about 25ndash50 planthoppers per hill [1373] Under such low densities it wasnot possible to observe the impact of host resistance on planthopper population build-
Crops 2021 1 178
up There was some indication of higher densities on cartap hydrochloride-treated IR62in the screenhouse which may be related to the stimulatory effects of the product onegg laying and the low mortality of nymphs on cartap hydrochloride-treated plants asobserved in the greenhouse bioassays (Table 1) Field studies from the Philippines haveindicated that IR62 effectively reduces BPH densities during wet and dry seasons [4]However farmers are rarely aware of which anti-herbivore resistance their rice varietiespossess and frequently apply pesticides regardless of resistance thereby reducing theirfarm profits [3841] For example Joshi et al (1992) [41] reported widespread outbreaks ofBPH on IR62 in Mindanao Philippines after applications of decamethrin (a pyrethroid) tothe resistant variety Where such resurgence-causing insecticides are applied resistanceseems to be often incapable of reducing the stimulatory effects of the insecticide and itloses its capacity to buffer against outbreaks
42 Antagonisms
Several insecticides are associated with increasing densities of planthoppers andwith resurgence outbreaks in farmersrsquo fields [27293041496174] Indeed patches oflsquohopperburnrsquo (ie dead rice plants resulting from planthopper damage) are often anindicator of high insecticide use in tropical Asia [214] In many cases these resurgence-causing insecticides were once effective in reducing planthopper densities but over timepopulations developed resistance against the products The resulting low toxicity canthen have a stimulatory effect on planthopper feeding and development [2123242743]This may be through hormesis which is a direct response by the planthopper to low-levelexposure to the toxic chemical or it may result from physiological changes to the rice plantsthat ultimately benefit the planthopper [1849] In our screenhouse study we induced aresurgence outbreak of BPH WBPH and to a lesser extent GLH on deltamethrin-treatedrice plants (Table 2) The combined densities of all three herbivores increased almost10 times on the treated plants Furthermore herbivore biomass increased by between fourand six times Although the resurgence effects of deltamethrin have been documentedin previous studies [2029ndash313975] we could not predict from our greenhouse studiesthat any such outbreak would occur Egg laying was unaffected by deltamethrin in ourchoice and no-choice experiments and nymph survival and development both declinedon deltamethrin-treated rice in the survival bioassay (Table 1) The massive increases inplanthopper densities on deltamethrin-treated plants in our study seem to result froma heightened attraction of adult BPH and WBPH to the plants [49] particularly sincenumbers were so low on all other plants including both treated and non-treated plantsHowever once on the plants planthoppers also apparently had enhanced fitness Cuonget al (1997) [31] observed higher population densities of BPH on deltamethrin-treatedrice in Vietnam albeit with lower-level increases on resistant varieties Furthermore in agreenhouse study Azzam et al (2009) [20] found that deltamethrin reduced pre-ovipositiontimes increased the duration of oviposition and increased egg hatchability and femalelongevity Nanthakumar et al (2012) [43] further showed that deltamethrin increased BPHfeeding efficiency on resistant and susceptible rice plants as determined through honeydewproduction and population size In studies with WBPH Salim and Heinrichs (1987) [36]showed deltamethrin to increase settling (antixenosis) feeding and population growthand Suri and Singh (2011) [28] showed deltamethrin to increase WBPH reproductive ratesand nymph survival In our study the observed resurgence of planthoppers was similaron both varieties indicating as was previously noted by Joshi et al (1992) [41] thatresurgence-causing pyrethroids are antagonistic to resistance in IR62
Though not statistically significant cartap hydrochloride also increased BPH numberson IR62 but not on IR64 in our screenhouse study This indicates that the product mayalso be antagonistic to the hostrsquos resistance mechanisms The stronger effect on IR62 maybe related to the differences between IR62 biomass growth rates and anatomy (ie alarger plant with higher tillering) compared to IR64 (Table 3) Once resistance is no longereffective because of chemically-enhanced BPH fitness these plant characteristics which
Crops 2021 1 179
have been related to observed preferences in BPH and other sap-sucking insects [349]could further accelerate the build-up of BPH populations on cartap hydrochloride-treatedplants To further test this idea experiments could be conducted using near-isogenic ricelines that differ mainly in the presence or absence of tagged resistance genes [76] Suchtests would avoid confounding differences in plant anatomy between test varieties IR62resistance to BPH has been well studied [4] The variety is also resistant to WBPH atolder plant stages [54] Therefore in our study deltamethrin was also antagonistic tothis WBPH-resistance (Table 2) However BPH feeding also facilitates WBPH populationgrowth [5477] and there may have been added indirect plant-mediated effects [77] thataugmented WBPH populations on both varieties in our experiment IR62 is also resistant toGLH [45] In our experiments GLH densities also increased on deltamethrin-treated plantsbut the effect was only apparent on IR64 Therefore GLH-resistance in IR62 apparentlybuffered against deltamethrin-associated increases in GLH on the variety as was alsonoted from field studies in Vietnam [31] Deltamethrin is currently not recommended foruse in rice [202878] however the product continues to be marketed in rice-dominatedregions throughout Asia [131479] Although cypermethrin has been associated with BPHoutbreaks and was consequently removed for use on rice in Thailand and Vietnam [62]we did not observe cypermethrin-induced resurgence in our screenhouse experimentNevertheless cypermethrin has been associated with resurgence of BPH in South Asia [61]and we have noted stimulatory effects of cypermethrin on other rice herbivores in thePhilippines (Horgan unpublished data)
43 Phytotoxicity
Phytotoxicity is relatively common following applications of insecticides [182850]Phytotoxic effects may change a plantrsquos physiology without consequences for crop yieldsHowever such changes to plant physiology could have indirect effects on herbivores Forexample deltamethrin methyl parathion and quinalphos produce increases in total sugarsand reducing sugars as well as increasing the protein contents of the rice leaf sheath andleaf blade [28] Several insecticides also reduce plant phenols in ricemdashwhich increases therice plantrsquos susceptibility to BPH and other insects [27] In our screenhouse experiment wenoted several changes to rice anatomy and yields Importantly we noted that cypermethrinand fipronil both reduced rice yields (Table 3) This was not due to indirect herbivoreeffects because planthopper and leafhopper densities were consistently low on rice treatedwith these insecticides The effects therefore probably resulted from direct phytotoxiceffects on the plants Furthermore these effects were most pronounced on treated IR62plants Other minor effects on grain production included increased proportions of unfilledgrain in cartap hydrochloride-treated plants and delays in grain maturation on cartaphydrochloride-treated and cypermethrin-treated plants (Table 3) Despite the high densitiesof BPH and WBPH on deltamethrin-treated plants yields were not significantly lower thanin control non-treated plants Both IR62 and IR64 have been noted for their high toleranceto BPH damage particularly under high nitrogenous fertilizer applications [3456] as usedin our screenhouse experiment Therefore where plants have high tolerance to damageapplying insecticides could also result in profitability losses on rice farms and in the caseof insecticide-induced pest population increases tolerance might buffer against subsequentyield losses There are few reports of the phytotoxicity of insecticides to rice carbofuran istoxic to rice at high concentrations but may also enhance the growth of rice plants throughinteractions with nitrogen-fixing bacteria in the rhizosphere [80] Future studies couldmore carefully assess field-grown plants for possible phytotoxic effects particularly ifinsecticides have the potential to directly reduce yields
44 Management implications
Global trends suggest that resistance in rice has failed to reduce insecticide appli-cations [2] This may be due to aggressive marketing by agrochemical companies andstrategies to increase pesticide sales such as promoting prophylactic applications [16] Be-
Crops 2021 1 180
cause we selected our test insecticides (except the pyrethroids) without any a priori criteriaour results suggest that many currently used insecticides probably have some negativeeffects on yield either directly through phytotoxic effects or indirectly by inducing pestresurgence Furthermore our results indicate that even where pests reach high densitiesas on deltamethrin-treated plants in the screenhouse these densities do not necessarilylead to significant yield reductions
Among the products we used buprofezin although it had little effect on oviposi-tion was effective in reducing nymph survival Furthermore there were only minorapparent phytotoxic effects on the rice plants Buprofezin has a low toxicity to naturalenemies in rice fields [58] and is therefore the most compatible of the insecticides withstrategies to increase rice field biodiversity and restore ecosystem services [32ndash348182]Thiamethoxam-chlorantraniliprole was also effective in reducing both oviposition andnymph survival and had no apparent toxic effects on rice plants However thiamethoxam-chlorantraniliprole and other neonicotinoids although moderately toxic can have severeeffects on beneficial hymenopteran parasitoids [83] although they have apparently lowtoxicity to predatory mirid bugs [84] The remaining insecticides either directly reducedyields or were associated with higher BPH biomass or densities in our experiments
We noted few synergies with HPR although nymph biomass andor survival waslower in cartap hydrochloride and carbofuran-treated IR62 plants compared to correspond-ing IR64 plants Meanwhile cartap hydrochloride and deltamethrin were antagonisticto resistance in IR62 in the screenhouse experiment Among the strongest effects of HPRwas to act as a buffer against GLH population build-up on deltamethrin-treated IR62 andagainst carbofuran-induced increases in BPH nymph biomass in the survival bioassayTherefore HPR may play a role in reducing the possibilities of secondary outbreaks ofBPH and other insects The combined use of HPR and targeted insecticides against BPHcreates redundancy in the rice field and counters measures to manage insecticide resistanceand virulence by unnecessarily exposing plant protection products to evolving pest pop-ulations HPR (and host plant tolerance) can be effective against target pests as revealedin previous studies [4] and could be incorporated into other pest management practicesparticularly agroecological practices such as ecological engineering with which it is highlycompatible [818586]
5 Conclusions
Based on our results and the results of previous studies HPR and the applicationof chemical insecticides against BPH rarely produce synergistic effects Antagonisticeffects where chemicals caused increases in BPH fitness or populations on resistant ricewere more apparent in our study In particular deltamethrin caused an outbreak ofBPH in our screenhouse study Nevertheless HPR can buffer against the insecticide-induced enhancement of herbivore fitness (hormesis) and potentially reduce possibilities ofsecondary outbreaks as was suggested from results with cartap hydrochloride in our studyBased on our results we suggest that carbofuran cartap hydrochloride cypermethrindeltamethrin and fipronil should be avoided in rice particularly in the Philippines (wherethis research was conducted) due to their resurgence-causing potential or because of directreductions to rice yields
Author Contributions Conceptualization FGH methodology FGH AP-C and MLPA formalanalysis FGH and AP-C investigation FGH AP-C and MLPA data curation FGHAP-C and MLPA writingmdashoriginal draft preparation FGH and AP-C writingmdashreview andediting FGH AP-C and MLPA visualization FGH supervision FGH and MLPA projectadministration FGH funding acquisition FGH All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by the Bill and Melinda Gates Foundation (Cereal SystemsInitiative for South Asia [CSISA] OPP52303) and the Global Rice Science Platform (GRiSP) underthe directorship of Achim Dobermann AP-C was partially funded by The Spanish Governmentthrough Becas Faro International
Crops 2021 1 181
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data presented in this study are available on reasonable requestfrom the corresponding author
Acknowledgments The authors thank Fame Ramal Carmencita Bernal Alberto Naredo ReyeulQuintana Vincent Virtudes Marol Recide Jenyrose Geronda and Ellen Genil for assistance duringthis study The contributions of Rodante Abas (sadly deceased) during three decades of entomologicalresearch at IRRI are gratefully acknowledged Rice lines were supplied through the Genebank of IRRI
Conflicts of Interest The authors declare no conflict of interest
References1 Bottrell DG Schoenly KG Resurrecting the ghost of green revolutions past The brown planthopper as a recurring threat to
high-yielding rice production in tropical Asia J Asia-Pac Entomol 2012 15 122ndash140 [CrossRef]2 Horgan FG Integrated pest management for sustainable rice cultivation A holistic approach In Achieving Sustainable Cultivation
of Rice Cultivation Pest and Disease Management Sasaki T Ed Burleigh Dodds Scientific Cambridge UK 2017 pp 309ndash3423 Horgan FG de Freitas TFS Crisol-Martiacutenez E Mundaca EA Bernal CC Nitrogenous fertilizer reduces resistance but
enhances tolerance to the brown planthopper in fast-growing moderately resistant rice Insects 2021 12 989 [CrossRef] [PubMed]4 Horgan FG Pentildealver Cruz A Bernal CC Ramal AF Almazan MLP Wilby A Resistance and tolerance to the brown
planthopper Nilaparvata lugens (Staringl) in rice infested at different growth stages across a gradient of nitrogen applicationsField Crops Res 2018 217 53ndash65 [CrossRef]
5 Hereward JP Cai X Matias AMA Walter GH Xu C Wang Y Migration dynamics of an important rice pest The brownplanthopper (Nilaparvata lugens) across AsiamdashInsights from population genomics Evol Appl 2020 13 2449ndash2459 [CrossRef]
6 Hu C Guo J Fu X Huang Y Gao X Wu K Seasonal migration pattern of Nilaparvata lugens (Hemiptera Delphacidae) overthe Bohai Sea in northern China J Econ Entomol 2018 111 2129ndash2135 [CrossRef]
7 Ma J Wang Y-C Hu Y-Y Lu M-H Wan G-J Chen F-J Liu W-C Zhai B-P Hu G Brown planthopper Nilaparvatalugens was concentrated at the rear of the typhoon Soudelor in eastern China in August 2015 Insect Sci 2018 25 916ndash926[CrossRef]
8 Fujita D Kohli A Horgan FG Rice resistance to planthoppers and leafhoppers Crit Rev Plant Sci 2013 32 162ndash191[CrossRef]
9 Fujii T Yoshida K Kobayashi T Myint KKM Yasui H Sanada-Morimura S Matsumura M Long-term virulencemonitoring of differential cultivars in Japanrsquos immigrant populations of Nilaparvata lugens (Hemiptera Delphacidae) in 2001ndash2009Appl Entomol Zool 2021 56 407ndash418 [CrossRef]
10 Myint KKM Yasui H Takagi M Matsumura M Virulence of long-term laboratory populations of the brown planthopperNilaparvata lugens (Stal) and whitebacked planthopper Sogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) on rice differentialvarieties Appl Entomol Zool 2009 44 149ndash153 [CrossRef]
11 Horgan FG Ramal AF Bentur JS Kumar R Bhanu KV Sarao PS Iswanto EH Van Chien H Phyu MHBernal CC et al Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia againstresistant rice varieties Crop Prot 2015 78 222ndash231 [CrossRef]
12 Horgan FG Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers Crop Prot2018 110 21ndash33 [CrossRef]
13 Uddin ABMA Islam KS Jahan M Ara A Afrin S Farmerrsquos perception about resurgence of brown planthopper NilaparvataLugens (Staringl) in Bangladesh Bangladesh Rice J 2020 23 21ndash33 [CrossRef]
14 Triwidodo H Brown planthoppers infestations and insecticides use pattern in Java Indonesia AGRIVITA J Agric Sci 202042 320ndash330 [CrossRef]
15 Veres A Wyckhuys KA Kiss J Toacuteth F Burgio G Pons X Avilla C Vidal S Razinger J Bazok R et al An update of theWorldwide Integrated Assessment (WIA) on systemic pesticides Part 4 Alternatives in major cropping systems Environ SciPollut Res 2020 27 29867ndash29899 [CrossRef]
16 Bakker L van der Werf W Tittonell P Wyckhuys KA Bianchi FJ Neonicotinoids in global agriculture Evidence for a newpesticide treadmill Ecol Soc 2020 25 26 [CrossRef]
17 Wu J-C Xu J-X Yuan S-Z Liu J-L Jiang Y-H Xu J-F Pesticide-induced susceptibility of rice to brown planthopperNilaparvata lugens Entomol Exp Appl 2001 100 119ndash126 [CrossRef]
18 Wu J-C Qiu H-M Yang G-Q Liu J-L Liu G-J Wilkens RM Effective duration of pesticide-induced susceptibility of riceto brown planthopper (Nilaparvata lugens Staringl Homoptera Delphacidae) and physiological and biochemical changes in riceplants following pesticide application Int J Pest Manag 2004 50 55ndash62 [CrossRef]
19 Wang A-H Wu J-C Yu Y-S Liu J-L Yue J-F Wang MY Selective insecticide-induced stimulation on fecundity andbiochemical changes in Tryporyza incertulas (Lepidoptera Pyralidae) J Econ Entomol 2005 98 1144ndash1149 [CrossRef] [PubMed]
Crops 2021 1 182
20 Azzam S Wang F Wu J-C Shen J Wang L-P Yang G-Q Guo Y-R Comparisons of stimulatory effects of a seriesof concentrations of four insecticides on reproduction in the rice brown planthopper Nilaparvata lugens Staringl (HomopteraDelphacidae) Int J Pest Manag 2009 55 347ndash358 [CrossRef]
21 Ge L-Q Hu J-H Wu J-C Yang G-Q Gu H Insecticide-induced changes in protein RNA and DNA contents in ovary andfat body of female Nilaparvata lugens (Hemiptera Delphacidae) J Econ Entomol 2009 102 1506ndash1514 [CrossRef]
22 Ling S Zhang J Hu L Zhang R Effect of fipronil on the reproduction feeding and relative fitness of brown planthopperNilaparvata lugens Appl Entomol Zool 2009 44 543ndash548 [CrossRef]
23 Ge L-Q Wang L-P Zhao K-F Wu J-C Huang L-J Mating pair combinations of insecticide-treated male and femaleNilaparvata lugens Staringl (Hemiptera Delphacidae) planthoppers influence protein content in the male accessory glands (MAGs)and vitellin content in both fat bodies and ovaries of adult females Pestic Biochem Physiol 2010 98 279ndash288 [CrossRef]
24 Ge L-Q Wu J-C Zhao K-F Chen Y Yang G-Q Induction of Nlvg and suppression of Nljhe gene expression in Nilaparvatalugens (Staringl) (Hemiptera Delphacidae) adult females and males exposed to two insecticides Pestic Biochem Physiol 201098 269ndash278 [CrossRef]
25 Hu J-H Wu J-C Yin J-L Gu H-N Physiology of insecticide-induced stimulation of reproduction in the rice brownplanthopper (Nilaparvata lugens (Staringl)) Dynamics of protein in fat body and ovary Int J Pest Manag 2010 56 23ndash30 [CrossRef]
26 Wang L-P Shen J Ge L-Q Wu J-C Yang G-Q Jahn GC Insecticide-induced increase in the protein content of maleaccessory glands and its effect on the fecundity of females in the brown planthopper Nilaparvata lugens Staringl (HemipteraDelphacidae) Crop Prot 2010 29 1280ndash1285 [CrossRef]
27 Wu J Ge L Liu F Song Q Stanley D Pesticide-induced planthopper population resurgence in rice cropping systems AnnuRev Entomol 2020 65 409ndash429 [CrossRef]
28 Suri KS Singh G Insecticide-induced resurgence of the whitebacked planthopper Sogatella furcifera (Horvath) (HemipteraDelphacidae) on rice varieties with different levels of resistance Crop Prot 2011 30 118ndash124 [CrossRef]
29 Fabellar LT Heinrichs EA Toxicity of insecticides to predators of rice brown planthoppers Nilaparvata lugens (Staringl) (HomopteraDelphacidae) Environ Entomol 1984 13 832ndash837 [CrossRef]
30 Fabellar LT Heinrichs EA Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators Crop Prot1986 5 254ndash258 [CrossRef]
31 Cuong NL Ben PT Phuong LT Chau LM Cohen MB Effect of host plant resistance and insecticide on brown planthopperNilaparvata lugens (Staringl) and predator population development in the Mekong Delta Vietnam Crop Prot 1997 16 707ndash715[CrossRef]
32 Preetha G Stanley J Suresh S Samiyappan R Risk assessment of insecticides used in rice on mirid bug Cyrtorhinuslividipennis Reuter the important predator of brown planthopper Nilaparvata lugens (Stal) Chemosphere 2010 80 498ndash503[CrossRef] [PubMed]
33 Tanaka K Endo S Kazano H Toxicity of insecticides to predators of rice planthoppers Spiders the mirid bug and the dryinidwasp Appl Entomol Zool 2000 35 177ndash187 [CrossRef]
34 Zhang X Xu Q Lu W Liu F Sublethal effects of four synthetic insecticides on the generalist predator Cyrtorhinus lividipennisJ Pest Sci 2015 88 383ndash392 [CrossRef]
35 Settle WH Ariawan H Astuti ET Cahyana W Hakim AL Hindayana D Lestari AS Managing tropical rice peststhrough conservation of generalist natural enemies and alternative prey Ecology 1996 77 1975ndash1988 [CrossRef]
36 Salim M Heinrichs EA Insecticide-induced changes in the levels of resistance of rice cultivars to the whitebacked planthopperSogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) Crop Prot 1987 6 28ndash32 [CrossRef]
37 Yin J-L Xu H-W Wu J-C Hu J-H Yang G-Q Cultivar and insecticide applications affect the physiological development ofthe brown planthopper Nilaparvata lugens (Staringl) (Hemiptera Delphacidae) Environ Entomol 2014 37 206ndash212 [CrossRef]
38 Cheng C Chang W Studies on varietal resistance to the brown planthopper in Taiwan In Brown Planthopper Threat toRice Production in Asia International Rice Research Institute Ed International Rice Research Institute Manila Philippines1979 pp 251ndash271
39 Reissig WH Heinrichs EA Valencia SL Insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on ricevarieties with different levels of resistance Environ Entomol 1982 11 165ndash168 [CrossRef]
40 Heinrichs EA Fabellar LT Basilio RP Wen T-C Medrano F Susceptibility of rice planthoppers Nilaparvata lugens andSogatella furcifera (Homoptera Delphacidae) to insecticides as influenced by level of resistance in the host plant Environ Entomol1984 13 455ndash458 [CrossRef]
41 Joshi R Shepard B Kenmore P Lydia R Insecticide-induced resurgence of brown planthopper (BPH) on IR62 Int Rice ResNewsl 1992 17 9ndash10
42 Zhu Z-R Cheng J Jiang M-X Zhang X-X Complex influence of rice variety fertilization timing and insecticide onpopulation dynamics of Sogatella furcifera (Horvath) Nilaparvata lugens (Staringl) (Homoptera Delphacidae) and their natural enemiesin rice in Hangzhou China J Pest Sci 2004 77 65ndash74 [CrossRef]
43 Nanthakumar M Jhansi Lakshmi V Shashi Bhushan V Balachandran SM Mohan M Decrease of rice plant resistance andinduction of hormesis and carboxylesterase titre in brown planthopper Nilaparvata lugens (Staringl) by xenobiotics Pestic BiochemPhysiol 2012 102 146ndash152 [CrossRef]
Crops 2021 1 183
44 Yao H-W Jiang C-Y Ye G-Y Cheng J-A Studies on cultivar-induced changes in insecticide susceptibility and its relatedenzyme activities of the white-backed planthopper Sogatella furcifera (Horvath)(Homopter Delphacidae) Insect Sci 2002 9 9ndash15
45 Horgan FG Srinivasan TS Bentur JS Kumar R Bhanu KV Sarao PS Van Chien H Almazan MLP Bernal CCRamal AF et al Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers Implicationsfor rice breeding and gene deployment Agronomy 2017 7 62 [CrossRef]
46 Jairin J Phengrat K Teangdeerith S Vanavichit A Toojinda T Mapping of a broad-spectrum brown planthopper resistancegene Bph3 on rice chromosome 6 Mol Breed 2007 19 35ndash44 [CrossRef]
47 Ren J Gao F Wu X Lu X Zeng L Lv J Su X Luo H Ren G Bph32 a novel gene encoding an unknown SCRdomain-containing protein confers resistance against the brown planthopper in rice Sci Rep 2016 6 37645 [CrossRef]
48 Nguyen CD Zheng S-H Sanada-Morimura S Matsumura M Yasui H Fujita D Substitution mapping and characterizationof brown planthopper resistance genes from indica rice variety lsquoPTB33prime (Oryza sativa L) Breed Sci 2021 in press [CrossRef]
49 Chelliah S Heinrichs EA Factors affecting insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on RiceEnviron Entomol 1980 9 773ndash777 [CrossRef]
50 Moore MT Kroumlger R Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growthArch Environ Contam Toxicol 2010 59 574ndash581 [CrossRef]
51 Kim J Hwang I Han M Jang B Effect of insecticide and fungicide on phytotoxicity of herbicide in rice Korean J Weed Sci1986 6 67ndash75
52 Horgan FG Garcia CPF Haverkort F de Jong PW Ferrater JB Changes in insecticide resistance and host range performanceof planthoppers artificially selected to feed on resistant rice Crop Prot 2020 127 104963 [CrossRef] [PubMed]
53 Khush GS Virk PS IR Varieties and Their Impact International Rice Research Institute Manila Philippines 200554 Srinivasan TS Almazan MLP Bernal CC Ramal AF Subbarayalu MK Horgan FG Interactions between nymphs of
Nilaparvata lugens and Sogatella furcifera (Hemiptera Delphacidae) on resistant and susceptible rice varieties Appl Entomol Zool2016 51 81ndash90 [CrossRef]
55 Pentildealver Cruz A Arida A Heong KL Horgan FG Aspects of brown planthopper adaptation to resistant rice varieties withthe Bph3 gene Entomol Exp Appl 2011 141 245ndash257 [CrossRef]
56 Alam SN Cohen MB Detection and analysis of QTLs for resistance to the brown planthopper Nilaparvata lugens in adoubled-haploid rice population Theor Appl Genet 1998 97 1370ndash1379 [CrossRef]
57 Mackill DJ Khush GS IR64 A high-quality and high-yielding mega variety Rice 2018 11 18 [CrossRef] [PubMed]58 Heinrichs EA Basilio RP Valencia SL Buprofezin a selective insecticide for the management of rice planthoppers (Ho-
moptera Delphacidae) and leafhoppers (Homoptera Cicadellidae) Environ Entomol 1984 13 515ndash521 [CrossRef]59 Satapathy MK Anjaneyulu A Use of cypermethrin a synthetic pyrethroid in the control of rice tungro virus disease and its
vector Trop Pest Manag 1984 30 170ndash178 [CrossRef]60 Vorley W Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown
planthopper (BPH) in Kedah Malaysia Int Rice Res Newsl 1987 10 19ndash2061 Uddin A Islam KS Jahan M Ara A Khan MAI Factor influencing the resurgence of brown planthopper in Bangladesh
SAARC J Agric 2020 18 117ndash128 [CrossRef]62 Escalada MM Luecha M Heong KL Social impacts of planthopper outbreaks Case study from Thailand In Rice Planthoppers
Ecology Management Socio Economics and Policy Heong KL Cheng J Escalada MM Eds Zhejiang University PressHangzhou China 2015 pp 209ndash226
63 Zhang CP He HM Yu JZ Hu XQ Zhu YH Wang Q Residues of carbosulfan and its metabolites carbofuran and3-hydroxy carbofuran in rice field ecosystem in China J Environ Sci Health 2016 51 351ndash357 [CrossRef] [PubMed]
64 Samanta S Barman M Samanta NRA Bio-efficacy trials of carbofuran 3 CG against insect pests of rice J Entomol ZoolStud 2020 8 1529ndash1533
65 Donley N The USA lags behind other agricultural nations in banning harmful pesticides Environ Health 2019 18 44 [CrossRef][PubMed]
66 Adams A Gore J Musser F Cook D Catchot A Walker T Dobbins C Efficacy of selected insecticides applied to hybridrice seed J Econ Entomol 2015 109 200ndash206 [CrossRef]
67 Kumar R Singh B Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) Field Bull Environ Contam Toxicol2013 90 482ndash488 [CrossRef]
68 Dhaka S Prajapati C Singh D Singh R Field evaluation of insecticides and bio-pesticides against rice leaf folder CnaphalocrocisMedinalis (Guenee) Ann Plant Prot Sci 2011 19 324ndash326
69 Matsumura M Takeuchi H Satoh M Sanada-Morimura S Otuka A Watanabe T Van Thanh D Species-specific insecticideresistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in East and South-eastAsia Pest Manag Sci 2008 64 1115ndash1121 [CrossRef] [PubMed]
70 Tang J Li J Shao Y Yang B Liu Z Fipronil resistance in the whitebacked planthopper (Sogatella furcifera) Possible resistancemechanisms and cross-resistance Pest Manag Sci 2010 66 121ndash125 [CrossRef]
71 Horgan FG Arida A Ardestani G Almazan MLP Temperature-dependent oviposition and nymph performance revealdistinct thermal niches of coexisting planthoppers with similar thresholds for development PLoS ONE 2020 15 e0235506[CrossRef] [PubMed]
Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]
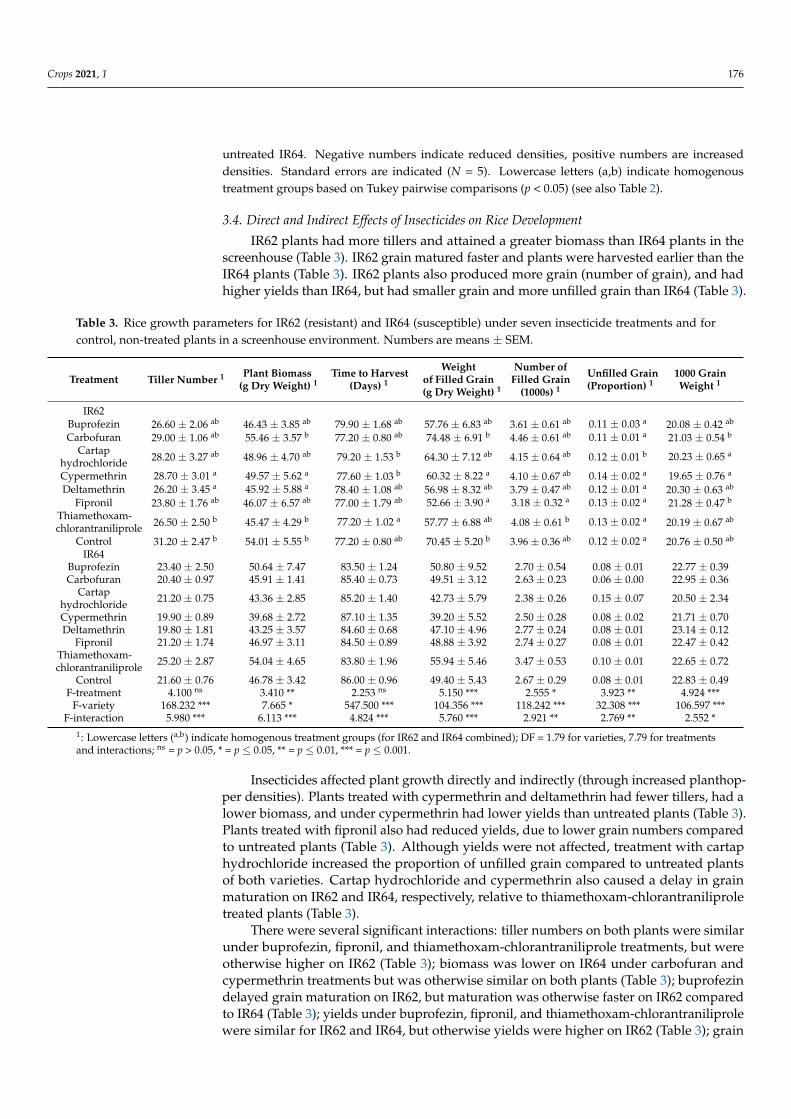
Crops 2021 1 176
untreated IR64 Negative numbers indicate reduced densities positive numbers are increaseddensities Standard errors are indicated (N = 5) Lowercase letters (ab) indicate homogenoustreatment groups based on Tukey pairwise comparisons (p lt 005) (see also Table 2)
34 Direct and Indirect Effects of Insecticides on Rice Development
IR62 plants had more tillers and attained a greater biomass than IR64 plants in thescreenhouse (Table 3) IR62 grain matured faster and plants were harvested earlier than theIR64 plants (Table 3) IR62 plants also produced more grain (number of grain) and hadhigher yields than IR64 but had smaller grain and more unfilled grain than IR64 (Table 3)
Table 3 Rice growth parameters for IR62 (resistant) and IR64 (susceptible) under seven insecticide treatments and forcontrol non-treated plants in a screenhouse environment Numbers are means plusmn SEM
Treatment Tiller Number 1 Plant Biomass(g Dry Weight) 1
Time to Harvest(Days) 1
Weightof Filled Grain(g Dry Weight) 1
Number ofFilled Grain
(1000s) 1
Unfilled Grain(Proportion) 1
1000 GrainWeight 1
IR62Buprofezin 2660 plusmn 206 ab 4643 plusmn 385 ab 7990 plusmn 168 ab 5776 plusmn 683 ab 361plusmn 061 ab 011 plusmn 003 a 2008 plusmn 042 ab
Carbofuran 2900 plusmn 106 ab 5546 plusmn 357 b 7720 plusmn 080 ab 7448 plusmn 691 b 446plusmn 061 ab 011 plusmn 001 a 2103 plusmn 054 b
Cartaphydrochloride 2820 plusmn 327 ab 4896 plusmn 470 ab 7920 plusmn 153 b 6430 plusmn 712 ab 415plusmn 064 ab 012 plusmn 001 b 2023 plusmn 065 a
Cypermethrin 2870 plusmn 301 a 4957 plusmn 562 a 7760 plusmn 103 b 6032 plusmn 822 a 410plusmn 067 ab 014 plusmn 002 a 1965 plusmn 076 a
Deltamethrin 2620 plusmn 345 a 4592 plusmn 588 a 7840 plusmn 108 ab 5698 plusmn 832 ab 379plusmn 047 ab 012 plusmn 001 a 2030 plusmn 063 ab
Fipronil 2380 plusmn 176 ab 4607 plusmn 657 ab 7700 plusmn 179 ab 5266 plusmn 390 a 318 plusmn 032 a 013 plusmn 002 a 2128 plusmn 047 b
Thiamethoxam-chlorantraniliprole 2650 plusmn 250 b 4547 plusmn 429 b 7720 plusmn 102 a 5777 plusmn 688 ab 408 plusmn 061 b 013 plusmn 002 a 2019 plusmn 067 ab
Control 3120 plusmn 247 b 5401 plusmn 555 b 7720 plusmn 080 ab 7045 plusmn 520 b 396plusmn 036 ab 012 plusmn 002 a 2076 plusmn 050 ab
IR64Buprofezin 2340 plusmn 250 5064 plusmn 747 8350 plusmn 124 5080 plusmn 952 270 plusmn 054 008 plusmn 001 2277 plusmn 039Carbofuran 2040 plusmn 097 4591 plusmn 141 8540 plusmn 073 4951 plusmn 312 263 plusmn 023 006 plusmn 000 2295 plusmn 036
Cartaphydrochloride 2120 plusmn 075 4336 plusmn 285 8520 plusmn 140 4273 plusmn 579 238 plusmn 026 015 plusmn 007 2050 plusmn 234
Cypermethrin 1990 plusmn 089 3968 plusmn 272 8710 plusmn 135 3920 plusmn 552 250 plusmn 028 008 plusmn 002 2171 plusmn 070Deltamethrin 1980 plusmn 181 4325 plusmn 357 8460 plusmn 068 4710 plusmn 496 277 plusmn 024 008 plusmn 001 2314 plusmn 012
Fipronil 2120 plusmn 174 4697 plusmn 311 8450 plusmn 089 4888 plusmn 392 274 plusmn 027 008 plusmn 001 2247 plusmn 042Thiamethoxam-chlorantraniliprole 2520 plusmn 287 5404 plusmn 465 8380 plusmn 196 5594 plusmn 546 347 plusmn 053 010 plusmn 001 2265 plusmn 072
Control 2160 plusmn 076 4678 plusmn 342 8600 plusmn 096 4940 plusmn 543 267 plusmn 029 008 plusmn 001 2283 plusmn 049F-treatment 4100 ns 3410 2253 ns 5150 2555 3923 4924
F-variety 168232 7665 547500 104356 118242 32308 106597 F-interaction 5980 6113 4824 5760 2921 2769 2552
1 Lowercase letters (ab) indicate homogenous treatment groups (for IR62 and IR64 combined) DF = 179 for varieties 779 for treatmentsand interactions ns = p gt 005 = p le 005 = p le 001 = p le 0001
Insecticides affected plant growth directly and indirectly (through increased planthop-per densities) Plants treated with cypermethrin and deltamethrin had fewer tillers had alower biomass and under cypermethrin had lower yields than untreated plants (Table 3)Plants treated with fipronil also had reduced yields due to lower grain numbers comparedto untreated plants (Table 3) Although yields were not affected treatment with cartaphydrochloride increased the proportion of unfilled grain compared to untreated plantsof both varieties Cartap hydrochloride and cypermethrin also caused a delay in grainmaturation on IR62 and IR64 respectively relative to thiamethoxam-chlorantraniliproletreated plants (Table 3)
There were several significant interactions tiller numbers on both plants were similarunder buprofezin fipronil and thiamethoxam-chlorantraniliprole treatments but wereotherwise higher on IR62 (Table 3) biomass was lower on IR64 under carbofuran andcypermethrin treatments but was otherwise similar on both plants (Table 3) buprofezindelayed grain maturation on IR62 but maturation was otherwise faster on IR62 comparedto IR64 (Table 3) yields under buprofezin fipronil and thiamethoxam-chlorantraniliprolewere similar for IR62 and IR64 but otherwise yields were higher on IR62 (Table 3) grain
Crops 2021 1 177
numbers were higher on IR62 than IR64 except under fipronil treatments (Table 3) theproportions of grain unfilled were higher on IR62 but were similar between varieties underthe fipronil treatment (Table 3) 1000-grain weight was higher in IR64 but similar on bothvarieties when treated with cartap hydrochloride (Table 3)
4 Discussion
Our results using a highly resistant rice variety and some commonly used insecticidesindicate a range of possible effects on planthoppers and rice plants by combining these twomanagement practices
41 Synergies
The most predictable effect of combining HPR and insecticides is a synergy in reduc-ing BPH numbers This is suggested because planthoppers are weakened by the hostrsquosresistance and should therefore be more susceptible to the toxic effects of insecticides [40]Using spray towers over potted plants Heinrichs et al (1984) [40] examined the combinedeffects of a range of insecticides (that included carbofuran) with resistance by comparingBPH mortality on six resistant and two susceptible rice varieties Their results indicatedthat mortality was greatest when insecticides were applied to BPH on resistant varietiesThe effects were similar when the study was repeated with WBPH for three insecticides(including carbofuran) on the resistant variety N22 (Wbph1 gene) [40] In our study hostresistance reduced egg laying and nymph biomass on non-treated IR62 compared to non-treated IR64 Egg laying was also reduced on plants treated with cypermethrin fipronilor thiamethoxam-chlorantraniliprole (Table 1) Based on the results of our choice andno-choice oviposition bioassays cypermethrin acted mainly as a deterrent of gravid fe-males whereas fipronil and thiamethoxam-chlorantraniliprole directly reduced egg layingDespite similar BPH responses to both the insecticides and resistant host there were no ap-parent synergies between any of these insecticides and resistance in reducing egg numbersin the oviposition bioassay In contrast to these three insecticides our bioassays indicatedthat buprofezin carbofuran and cartap hydrochloride can lead to higher numbers of eggson treated plants compared to untreated plants (Table 1) These effects were not statisticallysignificant and may be exaggerated because bioassays were conducted over relativelyshort time scales For example planthoppers will sometimes respond to stresses such ashigh temperatures by increasing oviposition rates [7172] Furthermore the stimulation ofegg-laying by these chemicals was not apparent in our screenhouse experiment (Table 2)Nevertheless our results do indicate that host resistance reduced the magnitude of anystimulatory effect on oviposition egg numbers on buprofezin-treated carbofuran-treatedand cartap hydrochloride-treated IR64 in the no-choice bioassay were twice as high as onthe corresponding IR62 plants In cases such as these the resistance of IR62 functions as abuffer or lsquoinsurancersquo against possible undesired pesticide effects [1238]
Although buprofezin appeared to stimulate oviposition the growth hormone had astrong impact on nymph development leading to zero survival of exposed nymphs in ourgreenhouse bioassay Cypermethrin fipronil and thiamethoxam-chlorantraniliprole alsoreduced nymph survival to zero in the greenhouse bioassay (Table 1) Where insecticideshave such high efficiencies any fitness losses to BPH due to host resistance then becomeredundant Unlike buprofezin carbofuran also stimulated nymph biomass on IR64 in thesurvival bioassay (Table 1) however nymph biomass was lower on carbofuran-treated IR62than on non-treated IR62 indicating that the hostrsquos resistance buffered against undesiredstimulatory effects on BPH and may have functioned synergistically to augment mortalityand slow BPH growth Such buffering allows insecticides to be used against other pestswithout inducing secondary outbreaks of non-target pests In our screenhouse experimentherbivore densities were generally low on both IR62 and IR64 for most treatments (exceptdeltamethrin) and were below threshold densities for insecticide applicationsmdashwhichare normally about 25ndash50 planthoppers per hill [1373] Under such low densities it wasnot possible to observe the impact of host resistance on planthopper population build-
Crops 2021 1 178
up There was some indication of higher densities on cartap hydrochloride-treated IR62in the screenhouse which may be related to the stimulatory effects of the product onegg laying and the low mortality of nymphs on cartap hydrochloride-treated plants asobserved in the greenhouse bioassays (Table 1) Field studies from the Philippines haveindicated that IR62 effectively reduces BPH densities during wet and dry seasons [4]However farmers are rarely aware of which anti-herbivore resistance their rice varietiespossess and frequently apply pesticides regardless of resistance thereby reducing theirfarm profits [3841] For example Joshi et al (1992) [41] reported widespread outbreaks ofBPH on IR62 in Mindanao Philippines after applications of decamethrin (a pyrethroid) tothe resistant variety Where such resurgence-causing insecticides are applied resistanceseems to be often incapable of reducing the stimulatory effects of the insecticide and itloses its capacity to buffer against outbreaks
42 Antagonisms
Several insecticides are associated with increasing densities of planthoppers andwith resurgence outbreaks in farmersrsquo fields [27293041496174] Indeed patches oflsquohopperburnrsquo (ie dead rice plants resulting from planthopper damage) are often anindicator of high insecticide use in tropical Asia [214] In many cases these resurgence-causing insecticides were once effective in reducing planthopper densities but over timepopulations developed resistance against the products The resulting low toxicity canthen have a stimulatory effect on planthopper feeding and development [2123242743]This may be through hormesis which is a direct response by the planthopper to low-levelexposure to the toxic chemical or it may result from physiological changes to the rice plantsthat ultimately benefit the planthopper [1849] In our screenhouse study we induced aresurgence outbreak of BPH WBPH and to a lesser extent GLH on deltamethrin-treatedrice plants (Table 2) The combined densities of all three herbivores increased almost10 times on the treated plants Furthermore herbivore biomass increased by between fourand six times Although the resurgence effects of deltamethrin have been documentedin previous studies [2029ndash313975] we could not predict from our greenhouse studiesthat any such outbreak would occur Egg laying was unaffected by deltamethrin in ourchoice and no-choice experiments and nymph survival and development both declinedon deltamethrin-treated rice in the survival bioassay (Table 1) The massive increases inplanthopper densities on deltamethrin-treated plants in our study seem to result froma heightened attraction of adult BPH and WBPH to the plants [49] particularly sincenumbers were so low on all other plants including both treated and non-treated plantsHowever once on the plants planthoppers also apparently had enhanced fitness Cuonget al (1997) [31] observed higher population densities of BPH on deltamethrin-treatedrice in Vietnam albeit with lower-level increases on resistant varieties Furthermore in agreenhouse study Azzam et al (2009) [20] found that deltamethrin reduced pre-ovipositiontimes increased the duration of oviposition and increased egg hatchability and femalelongevity Nanthakumar et al (2012) [43] further showed that deltamethrin increased BPHfeeding efficiency on resistant and susceptible rice plants as determined through honeydewproduction and population size In studies with WBPH Salim and Heinrichs (1987) [36]showed deltamethrin to increase settling (antixenosis) feeding and population growthand Suri and Singh (2011) [28] showed deltamethrin to increase WBPH reproductive ratesand nymph survival In our study the observed resurgence of planthoppers was similaron both varieties indicating as was previously noted by Joshi et al (1992) [41] thatresurgence-causing pyrethroids are antagonistic to resistance in IR62
Though not statistically significant cartap hydrochloride also increased BPH numberson IR62 but not on IR64 in our screenhouse study This indicates that the product mayalso be antagonistic to the hostrsquos resistance mechanisms The stronger effect on IR62 maybe related to the differences between IR62 biomass growth rates and anatomy (ie alarger plant with higher tillering) compared to IR64 (Table 3) Once resistance is no longereffective because of chemically-enhanced BPH fitness these plant characteristics which
Crops 2021 1 179
have been related to observed preferences in BPH and other sap-sucking insects [349]could further accelerate the build-up of BPH populations on cartap hydrochloride-treatedplants To further test this idea experiments could be conducted using near-isogenic ricelines that differ mainly in the presence or absence of tagged resistance genes [76] Suchtests would avoid confounding differences in plant anatomy between test varieties IR62resistance to BPH has been well studied [4] The variety is also resistant to WBPH atolder plant stages [54] Therefore in our study deltamethrin was also antagonistic tothis WBPH-resistance (Table 2) However BPH feeding also facilitates WBPH populationgrowth [5477] and there may have been added indirect plant-mediated effects [77] thataugmented WBPH populations on both varieties in our experiment IR62 is also resistant toGLH [45] In our experiments GLH densities also increased on deltamethrin-treated plantsbut the effect was only apparent on IR64 Therefore GLH-resistance in IR62 apparentlybuffered against deltamethrin-associated increases in GLH on the variety as was alsonoted from field studies in Vietnam [31] Deltamethrin is currently not recommended foruse in rice [202878] however the product continues to be marketed in rice-dominatedregions throughout Asia [131479] Although cypermethrin has been associated with BPHoutbreaks and was consequently removed for use on rice in Thailand and Vietnam [62]we did not observe cypermethrin-induced resurgence in our screenhouse experimentNevertheless cypermethrin has been associated with resurgence of BPH in South Asia [61]and we have noted stimulatory effects of cypermethrin on other rice herbivores in thePhilippines (Horgan unpublished data)
43 Phytotoxicity
Phytotoxicity is relatively common following applications of insecticides [182850]Phytotoxic effects may change a plantrsquos physiology without consequences for crop yieldsHowever such changes to plant physiology could have indirect effects on herbivores Forexample deltamethrin methyl parathion and quinalphos produce increases in total sugarsand reducing sugars as well as increasing the protein contents of the rice leaf sheath andleaf blade [28] Several insecticides also reduce plant phenols in ricemdashwhich increases therice plantrsquos susceptibility to BPH and other insects [27] In our screenhouse experiment wenoted several changes to rice anatomy and yields Importantly we noted that cypermethrinand fipronil both reduced rice yields (Table 3) This was not due to indirect herbivoreeffects because planthopper and leafhopper densities were consistently low on rice treatedwith these insecticides The effects therefore probably resulted from direct phytotoxiceffects on the plants Furthermore these effects were most pronounced on treated IR62plants Other minor effects on grain production included increased proportions of unfilledgrain in cartap hydrochloride-treated plants and delays in grain maturation on cartaphydrochloride-treated and cypermethrin-treated plants (Table 3) Despite the high densitiesof BPH and WBPH on deltamethrin-treated plants yields were not significantly lower thanin control non-treated plants Both IR62 and IR64 have been noted for their high toleranceto BPH damage particularly under high nitrogenous fertilizer applications [3456] as usedin our screenhouse experiment Therefore where plants have high tolerance to damageapplying insecticides could also result in profitability losses on rice farms and in the caseof insecticide-induced pest population increases tolerance might buffer against subsequentyield losses There are few reports of the phytotoxicity of insecticides to rice carbofuran istoxic to rice at high concentrations but may also enhance the growth of rice plants throughinteractions with nitrogen-fixing bacteria in the rhizosphere [80] Future studies couldmore carefully assess field-grown plants for possible phytotoxic effects particularly ifinsecticides have the potential to directly reduce yields
44 Management implications
Global trends suggest that resistance in rice has failed to reduce insecticide appli-cations [2] This may be due to aggressive marketing by agrochemical companies andstrategies to increase pesticide sales such as promoting prophylactic applications [16] Be-
Crops 2021 1 180
cause we selected our test insecticides (except the pyrethroids) without any a priori criteriaour results suggest that many currently used insecticides probably have some negativeeffects on yield either directly through phytotoxic effects or indirectly by inducing pestresurgence Furthermore our results indicate that even where pests reach high densitiesas on deltamethrin-treated plants in the screenhouse these densities do not necessarilylead to significant yield reductions
Among the products we used buprofezin although it had little effect on oviposi-tion was effective in reducing nymph survival Furthermore there were only minorapparent phytotoxic effects on the rice plants Buprofezin has a low toxicity to naturalenemies in rice fields [58] and is therefore the most compatible of the insecticides withstrategies to increase rice field biodiversity and restore ecosystem services [32ndash348182]Thiamethoxam-chlorantraniliprole was also effective in reducing both oviposition andnymph survival and had no apparent toxic effects on rice plants However thiamethoxam-chlorantraniliprole and other neonicotinoids although moderately toxic can have severeeffects on beneficial hymenopteran parasitoids [83] although they have apparently lowtoxicity to predatory mirid bugs [84] The remaining insecticides either directly reducedyields or were associated with higher BPH biomass or densities in our experiments
We noted few synergies with HPR although nymph biomass andor survival waslower in cartap hydrochloride and carbofuran-treated IR62 plants compared to correspond-ing IR64 plants Meanwhile cartap hydrochloride and deltamethrin were antagonisticto resistance in IR62 in the screenhouse experiment Among the strongest effects of HPRwas to act as a buffer against GLH population build-up on deltamethrin-treated IR62 andagainst carbofuran-induced increases in BPH nymph biomass in the survival bioassayTherefore HPR may play a role in reducing the possibilities of secondary outbreaks ofBPH and other insects The combined use of HPR and targeted insecticides against BPHcreates redundancy in the rice field and counters measures to manage insecticide resistanceand virulence by unnecessarily exposing plant protection products to evolving pest pop-ulations HPR (and host plant tolerance) can be effective against target pests as revealedin previous studies [4] and could be incorporated into other pest management practicesparticularly agroecological practices such as ecological engineering with which it is highlycompatible [818586]
5 Conclusions
Based on our results and the results of previous studies HPR and the applicationof chemical insecticides against BPH rarely produce synergistic effects Antagonisticeffects where chemicals caused increases in BPH fitness or populations on resistant ricewere more apparent in our study In particular deltamethrin caused an outbreak ofBPH in our screenhouse study Nevertheless HPR can buffer against the insecticide-induced enhancement of herbivore fitness (hormesis) and potentially reduce possibilities ofsecondary outbreaks as was suggested from results with cartap hydrochloride in our studyBased on our results we suggest that carbofuran cartap hydrochloride cypermethrindeltamethrin and fipronil should be avoided in rice particularly in the Philippines (wherethis research was conducted) due to their resurgence-causing potential or because of directreductions to rice yields
Author Contributions Conceptualization FGH methodology FGH AP-C and MLPA formalanalysis FGH and AP-C investigation FGH AP-C and MLPA data curation FGHAP-C and MLPA writingmdashoriginal draft preparation FGH and AP-C writingmdashreview andediting FGH AP-C and MLPA visualization FGH supervision FGH and MLPA projectadministration FGH funding acquisition FGH All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by the Bill and Melinda Gates Foundation (Cereal SystemsInitiative for South Asia [CSISA] OPP52303) and the Global Rice Science Platform (GRiSP) underthe directorship of Achim Dobermann AP-C was partially funded by The Spanish Governmentthrough Becas Faro International
Crops 2021 1 181
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data presented in this study are available on reasonable requestfrom the corresponding author
Acknowledgments The authors thank Fame Ramal Carmencita Bernal Alberto Naredo ReyeulQuintana Vincent Virtudes Marol Recide Jenyrose Geronda and Ellen Genil for assistance duringthis study The contributions of Rodante Abas (sadly deceased) during three decades of entomologicalresearch at IRRI are gratefully acknowledged Rice lines were supplied through the Genebank of IRRI
Conflicts of Interest The authors declare no conflict of interest
References1 Bottrell DG Schoenly KG Resurrecting the ghost of green revolutions past The brown planthopper as a recurring threat to
high-yielding rice production in tropical Asia J Asia-Pac Entomol 2012 15 122ndash140 [CrossRef]2 Horgan FG Integrated pest management for sustainable rice cultivation A holistic approach In Achieving Sustainable Cultivation
of Rice Cultivation Pest and Disease Management Sasaki T Ed Burleigh Dodds Scientific Cambridge UK 2017 pp 309ndash3423 Horgan FG de Freitas TFS Crisol-Martiacutenez E Mundaca EA Bernal CC Nitrogenous fertilizer reduces resistance but
enhances tolerance to the brown planthopper in fast-growing moderately resistant rice Insects 2021 12 989 [CrossRef] [PubMed]4 Horgan FG Pentildealver Cruz A Bernal CC Ramal AF Almazan MLP Wilby A Resistance and tolerance to the brown
planthopper Nilaparvata lugens (Staringl) in rice infested at different growth stages across a gradient of nitrogen applicationsField Crops Res 2018 217 53ndash65 [CrossRef]
5 Hereward JP Cai X Matias AMA Walter GH Xu C Wang Y Migration dynamics of an important rice pest The brownplanthopper (Nilaparvata lugens) across AsiamdashInsights from population genomics Evol Appl 2020 13 2449ndash2459 [CrossRef]
6 Hu C Guo J Fu X Huang Y Gao X Wu K Seasonal migration pattern of Nilaparvata lugens (Hemiptera Delphacidae) overthe Bohai Sea in northern China J Econ Entomol 2018 111 2129ndash2135 [CrossRef]
7 Ma J Wang Y-C Hu Y-Y Lu M-H Wan G-J Chen F-J Liu W-C Zhai B-P Hu G Brown planthopper Nilaparvatalugens was concentrated at the rear of the typhoon Soudelor in eastern China in August 2015 Insect Sci 2018 25 916ndash926[CrossRef]
8 Fujita D Kohli A Horgan FG Rice resistance to planthoppers and leafhoppers Crit Rev Plant Sci 2013 32 162ndash191[CrossRef]
9 Fujii T Yoshida K Kobayashi T Myint KKM Yasui H Sanada-Morimura S Matsumura M Long-term virulencemonitoring of differential cultivars in Japanrsquos immigrant populations of Nilaparvata lugens (Hemiptera Delphacidae) in 2001ndash2009Appl Entomol Zool 2021 56 407ndash418 [CrossRef]
10 Myint KKM Yasui H Takagi M Matsumura M Virulence of long-term laboratory populations of the brown planthopperNilaparvata lugens (Stal) and whitebacked planthopper Sogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) on rice differentialvarieties Appl Entomol Zool 2009 44 149ndash153 [CrossRef]
11 Horgan FG Ramal AF Bentur JS Kumar R Bhanu KV Sarao PS Iswanto EH Van Chien H Phyu MHBernal CC et al Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia againstresistant rice varieties Crop Prot 2015 78 222ndash231 [CrossRef]
12 Horgan FG Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers Crop Prot2018 110 21ndash33 [CrossRef]
13 Uddin ABMA Islam KS Jahan M Ara A Afrin S Farmerrsquos perception about resurgence of brown planthopper NilaparvataLugens (Staringl) in Bangladesh Bangladesh Rice J 2020 23 21ndash33 [CrossRef]
14 Triwidodo H Brown planthoppers infestations and insecticides use pattern in Java Indonesia AGRIVITA J Agric Sci 202042 320ndash330 [CrossRef]
15 Veres A Wyckhuys KA Kiss J Toacuteth F Burgio G Pons X Avilla C Vidal S Razinger J Bazok R et al An update of theWorldwide Integrated Assessment (WIA) on systemic pesticides Part 4 Alternatives in major cropping systems Environ SciPollut Res 2020 27 29867ndash29899 [CrossRef]
16 Bakker L van der Werf W Tittonell P Wyckhuys KA Bianchi FJ Neonicotinoids in global agriculture Evidence for a newpesticide treadmill Ecol Soc 2020 25 26 [CrossRef]
17 Wu J-C Xu J-X Yuan S-Z Liu J-L Jiang Y-H Xu J-F Pesticide-induced susceptibility of rice to brown planthopperNilaparvata lugens Entomol Exp Appl 2001 100 119ndash126 [CrossRef]
18 Wu J-C Qiu H-M Yang G-Q Liu J-L Liu G-J Wilkens RM Effective duration of pesticide-induced susceptibility of riceto brown planthopper (Nilaparvata lugens Staringl Homoptera Delphacidae) and physiological and biochemical changes in riceplants following pesticide application Int J Pest Manag 2004 50 55ndash62 [CrossRef]
19 Wang A-H Wu J-C Yu Y-S Liu J-L Yue J-F Wang MY Selective insecticide-induced stimulation on fecundity andbiochemical changes in Tryporyza incertulas (Lepidoptera Pyralidae) J Econ Entomol 2005 98 1144ndash1149 [CrossRef] [PubMed]
Crops 2021 1 182
20 Azzam S Wang F Wu J-C Shen J Wang L-P Yang G-Q Guo Y-R Comparisons of stimulatory effects of a seriesof concentrations of four insecticides on reproduction in the rice brown planthopper Nilaparvata lugens Staringl (HomopteraDelphacidae) Int J Pest Manag 2009 55 347ndash358 [CrossRef]
21 Ge L-Q Hu J-H Wu J-C Yang G-Q Gu H Insecticide-induced changes in protein RNA and DNA contents in ovary andfat body of female Nilaparvata lugens (Hemiptera Delphacidae) J Econ Entomol 2009 102 1506ndash1514 [CrossRef]
22 Ling S Zhang J Hu L Zhang R Effect of fipronil on the reproduction feeding and relative fitness of brown planthopperNilaparvata lugens Appl Entomol Zool 2009 44 543ndash548 [CrossRef]
23 Ge L-Q Wang L-P Zhao K-F Wu J-C Huang L-J Mating pair combinations of insecticide-treated male and femaleNilaparvata lugens Staringl (Hemiptera Delphacidae) planthoppers influence protein content in the male accessory glands (MAGs)and vitellin content in both fat bodies and ovaries of adult females Pestic Biochem Physiol 2010 98 279ndash288 [CrossRef]
24 Ge L-Q Wu J-C Zhao K-F Chen Y Yang G-Q Induction of Nlvg and suppression of Nljhe gene expression in Nilaparvatalugens (Staringl) (Hemiptera Delphacidae) adult females and males exposed to two insecticides Pestic Biochem Physiol 201098 269ndash278 [CrossRef]
25 Hu J-H Wu J-C Yin J-L Gu H-N Physiology of insecticide-induced stimulation of reproduction in the rice brownplanthopper (Nilaparvata lugens (Staringl)) Dynamics of protein in fat body and ovary Int J Pest Manag 2010 56 23ndash30 [CrossRef]
26 Wang L-P Shen J Ge L-Q Wu J-C Yang G-Q Jahn GC Insecticide-induced increase in the protein content of maleaccessory glands and its effect on the fecundity of females in the brown planthopper Nilaparvata lugens Staringl (HemipteraDelphacidae) Crop Prot 2010 29 1280ndash1285 [CrossRef]
27 Wu J Ge L Liu F Song Q Stanley D Pesticide-induced planthopper population resurgence in rice cropping systems AnnuRev Entomol 2020 65 409ndash429 [CrossRef]
28 Suri KS Singh G Insecticide-induced resurgence of the whitebacked planthopper Sogatella furcifera (Horvath) (HemipteraDelphacidae) on rice varieties with different levels of resistance Crop Prot 2011 30 118ndash124 [CrossRef]
29 Fabellar LT Heinrichs EA Toxicity of insecticides to predators of rice brown planthoppers Nilaparvata lugens (Staringl) (HomopteraDelphacidae) Environ Entomol 1984 13 832ndash837 [CrossRef]
30 Fabellar LT Heinrichs EA Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators Crop Prot1986 5 254ndash258 [CrossRef]
31 Cuong NL Ben PT Phuong LT Chau LM Cohen MB Effect of host plant resistance and insecticide on brown planthopperNilaparvata lugens (Staringl) and predator population development in the Mekong Delta Vietnam Crop Prot 1997 16 707ndash715[CrossRef]
32 Preetha G Stanley J Suresh S Samiyappan R Risk assessment of insecticides used in rice on mirid bug Cyrtorhinuslividipennis Reuter the important predator of brown planthopper Nilaparvata lugens (Stal) Chemosphere 2010 80 498ndash503[CrossRef] [PubMed]
33 Tanaka K Endo S Kazano H Toxicity of insecticides to predators of rice planthoppers Spiders the mirid bug and the dryinidwasp Appl Entomol Zool 2000 35 177ndash187 [CrossRef]
34 Zhang X Xu Q Lu W Liu F Sublethal effects of four synthetic insecticides on the generalist predator Cyrtorhinus lividipennisJ Pest Sci 2015 88 383ndash392 [CrossRef]
35 Settle WH Ariawan H Astuti ET Cahyana W Hakim AL Hindayana D Lestari AS Managing tropical rice peststhrough conservation of generalist natural enemies and alternative prey Ecology 1996 77 1975ndash1988 [CrossRef]
36 Salim M Heinrichs EA Insecticide-induced changes in the levels of resistance of rice cultivars to the whitebacked planthopperSogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) Crop Prot 1987 6 28ndash32 [CrossRef]
37 Yin J-L Xu H-W Wu J-C Hu J-H Yang G-Q Cultivar and insecticide applications affect the physiological development ofthe brown planthopper Nilaparvata lugens (Staringl) (Hemiptera Delphacidae) Environ Entomol 2014 37 206ndash212 [CrossRef]
38 Cheng C Chang W Studies on varietal resistance to the brown planthopper in Taiwan In Brown Planthopper Threat toRice Production in Asia International Rice Research Institute Ed International Rice Research Institute Manila Philippines1979 pp 251ndash271
39 Reissig WH Heinrichs EA Valencia SL Insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on ricevarieties with different levels of resistance Environ Entomol 1982 11 165ndash168 [CrossRef]
40 Heinrichs EA Fabellar LT Basilio RP Wen T-C Medrano F Susceptibility of rice planthoppers Nilaparvata lugens andSogatella furcifera (Homoptera Delphacidae) to insecticides as influenced by level of resistance in the host plant Environ Entomol1984 13 455ndash458 [CrossRef]
41 Joshi R Shepard B Kenmore P Lydia R Insecticide-induced resurgence of brown planthopper (BPH) on IR62 Int Rice ResNewsl 1992 17 9ndash10
42 Zhu Z-R Cheng J Jiang M-X Zhang X-X Complex influence of rice variety fertilization timing and insecticide onpopulation dynamics of Sogatella furcifera (Horvath) Nilaparvata lugens (Staringl) (Homoptera Delphacidae) and their natural enemiesin rice in Hangzhou China J Pest Sci 2004 77 65ndash74 [CrossRef]
43 Nanthakumar M Jhansi Lakshmi V Shashi Bhushan V Balachandran SM Mohan M Decrease of rice plant resistance andinduction of hormesis and carboxylesterase titre in brown planthopper Nilaparvata lugens (Staringl) by xenobiotics Pestic BiochemPhysiol 2012 102 146ndash152 [CrossRef]
Crops 2021 1 183
44 Yao H-W Jiang C-Y Ye G-Y Cheng J-A Studies on cultivar-induced changes in insecticide susceptibility and its relatedenzyme activities of the white-backed planthopper Sogatella furcifera (Horvath)(Homopter Delphacidae) Insect Sci 2002 9 9ndash15
45 Horgan FG Srinivasan TS Bentur JS Kumar R Bhanu KV Sarao PS Van Chien H Almazan MLP Bernal CCRamal AF et al Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers Implicationsfor rice breeding and gene deployment Agronomy 2017 7 62 [CrossRef]
46 Jairin J Phengrat K Teangdeerith S Vanavichit A Toojinda T Mapping of a broad-spectrum brown planthopper resistancegene Bph3 on rice chromosome 6 Mol Breed 2007 19 35ndash44 [CrossRef]
47 Ren J Gao F Wu X Lu X Zeng L Lv J Su X Luo H Ren G Bph32 a novel gene encoding an unknown SCRdomain-containing protein confers resistance against the brown planthopper in rice Sci Rep 2016 6 37645 [CrossRef]
48 Nguyen CD Zheng S-H Sanada-Morimura S Matsumura M Yasui H Fujita D Substitution mapping and characterizationof brown planthopper resistance genes from indica rice variety lsquoPTB33prime (Oryza sativa L) Breed Sci 2021 in press [CrossRef]
49 Chelliah S Heinrichs EA Factors affecting insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on RiceEnviron Entomol 1980 9 773ndash777 [CrossRef]
50 Moore MT Kroumlger R Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growthArch Environ Contam Toxicol 2010 59 574ndash581 [CrossRef]
51 Kim J Hwang I Han M Jang B Effect of insecticide and fungicide on phytotoxicity of herbicide in rice Korean J Weed Sci1986 6 67ndash75
52 Horgan FG Garcia CPF Haverkort F de Jong PW Ferrater JB Changes in insecticide resistance and host range performanceof planthoppers artificially selected to feed on resistant rice Crop Prot 2020 127 104963 [CrossRef] [PubMed]
53 Khush GS Virk PS IR Varieties and Their Impact International Rice Research Institute Manila Philippines 200554 Srinivasan TS Almazan MLP Bernal CC Ramal AF Subbarayalu MK Horgan FG Interactions between nymphs of
Nilaparvata lugens and Sogatella furcifera (Hemiptera Delphacidae) on resistant and susceptible rice varieties Appl Entomol Zool2016 51 81ndash90 [CrossRef]
55 Pentildealver Cruz A Arida A Heong KL Horgan FG Aspects of brown planthopper adaptation to resistant rice varieties withthe Bph3 gene Entomol Exp Appl 2011 141 245ndash257 [CrossRef]
56 Alam SN Cohen MB Detection and analysis of QTLs for resistance to the brown planthopper Nilaparvata lugens in adoubled-haploid rice population Theor Appl Genet 1998 97 1370ndash1379 [CrossRef]
57 Mackill DJ Khush GS IR64 A high-quality and high-yielding mega variety Rice 2018 11 18 [CrossRef] [PubMed]58 Heinrichs EA Basilio RP Valencia SL Buprofezin a selective insecticide for the management of rice planthoppers (Ho-
moptera Delphacidae) and leafhoppers (Homoptera Cicadellidae) Environ Entomol 1984 13 515ndash521 [CrossRef]59 Satapathy MK Anjaneyulu A Use of cypermethrin a synthetic pyrethroid in the control of rice tungro virus disease and its
vector Trop Pest Manag 1984 30 170ndash178 [CrossRef]60 Vorley W Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown
planthopper (BPH) in Kedah Malaysia Int Rice Res Newsl 1987 10 19ndash2061 Uddin A Islam KS Jahan M Ara A Khan MAI Factor influencing the resurgence of brown planthopper in Bangladesh
SAARC J Agric 2020 18 117ndash128 [CrossRef]62 Escalada MM Luecha M Heong KL Social impacts of planthopper outbreaks Case study from Thailand In Rice Planthoppers
Ecology Management Socio Economics and Policy Heong KL Cheng J Escalada MM Eds Zhejiang University PressHangzhou China 2015 pp 209ndash226
63 Zhang CP He HM Yu JZ Hu XQ Zhu YH Wang Q Residues of carbosulfan and its metabolites carbofuran and3-hydroxy carbofuran in rice field ecosystem in China J Environ Sci Health 2016 51 351ndash357 [CrossRef] [PubMed]
64 Samanta S Barman M Samanta NRA Bio-efficacy trials of carbofuran 3 CG against insect pests of rice J Entomol ZoolStud 2020 8 1529ndash1533
65 Donley N The USA lags behind other agricultural nations in banning harmful pesticides Environ Health 2019 18 44 [CrossRef][PubMed]
66 Adams A Gore J Musser F Cook D Catchot A Walker T Dobbins C Efficacy of selected insecticides applied to hybridrice seed J Econ Entomol 2015 109 200ndash206 [CrossRef]
67 Kumar R Singh B Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) Field Bull Environ Contam Toxicol2013 90 482ndash488 [CrossRef]
68 Dhaka S Prajapati C Singh D Singh R Field evaluation of insecticides and bio-pesticides against rice leaf folder CnaphalocrocisMedinalis (Guenee) Ann Plant Prot Sci 2011 19 324ndash326
69 Matsumura M Takeuchi H Satoh M Sanada-Morimura S Otuka A Watanabe T Van Thanh D Species-specific insecticideresistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in East and South-eastAsia Pest Manag Sci 2008 64 1115ndash1121 [CrossRef] [PubMed]
70 Tang J Li J Shao Y Yang B Liu Z Fipronil resistance in the whitebacked planthopper (Sogatella furcifera) Possible resistancemechanisms and cross-resistance Pest Manag Sci 2010 66 121ndash125 [CrossRef]
71 Horgan FG Arida A Ardestani G Almazan MLP Temperature-dependent oviposition and nymph performance revealdistinct thermal niches of coexisting planthoppers with similar thresholds for development PLoS ONE 2020 15 e0235506[CrossRef] [PubMed]
Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]

Crops 2021 1 177
numbers were higher on IR62 than IR64 except under fipronil treatments (Table 3) theproportions of grain unfilled were higher on IR62 but were similar between varieties underthe fipronil treatment (Table 3) 1000-grain weight was higher in IR64 but similar on bothvarieties when treated with cartap hydrochloride (Table 3)
4 Discussion
Our results using a highly resistant rice variety and some commonly used insecticidesindicate a range of possible effects on planthoppers and rice plants by combining these twomanagement practices
41 Synergies
The most predictable effect of combining HPR and insecticides is a synergy in reduc-ing BPH numbers This is suggested because planthoppers are weakened by the hostrsquosresistance and should therefore be more susceptible to the toxic effects of insecticides [40]Using spray towers over potted plants Heinrichs et al (1984) [40] examined the combinedeffects of a range of insecticides (that included carbofuran) with resistance by comparingBPH mortality on six resistant and two susceptible rice varieties Their results indicatedthat mortality was greatest when insecticides were applied to BPH on resistant varietiesThe effects were similar when the study was repeated with WBPH for three insecticides(including carbofuran) on the resistant variety N22 (Wbph1 gene) [40] In our study hostresistance reduced egg laying and nymph biomass on non-treated IR62 compared to non-treated IR64 Egg laying was also reduced on plants treated with cypermethrin fipronilor thiamethoxam-chlorantraniliprole (Table 1) Based on the results of our choice andno-choice oviposition bioassays cypermethrin acted mainly as a deterrent of gravid fe-males whereas fipronil and thiamethoxam-chlorantraniliprole directly reduced egg layingDespite similar BPH responses to both the insecticides and resistant host there were no ap-parent synergies between any of these insecticides and resistance in reducing egg numbersin the oviposition bioassay In contrast to these three insecticides our bioassays indicatedthat buprofezin carbofuran and cartap hydrochloride can lead to higher numbers of eggson treated plants compared to untreated plants (Table 1) These effects were not statisticallysignificant and may be exaggerated because bioassays were conducted over relativelyshort time scales For example planthoppers will sometimes respond to stresses such ashigh temperatures by increasing oviposition rates [7172] Furthermore the stimulation ofegg-laying by these chemicals was not apparent in our screenhouse experiment (Table 2)Nevertheless our results do indicate that host resistance reduced the magnitude of anystimulatory effect on oviposition egg numbers on buprofezin-treated carbofuran-treatedand cartap hydrochloride-treated IR64 in the no-choice bioassay were twice as high as onthe corresponding IR62 plants In cases such as these the resistance of IR62 functions as abuffer or lsquoinsurancersquo against possible undesired pesticide effects [1238]
Although buprofezin appeared to stimulate oviposition the growth hormone had astrong impact on nymph development leading to zero survival of exposed nymphs in ourgreenhouse bioassay Cypermethrin fipronil and thiamethoxam-chlorantraniliprole alsoreduced nymph survival to zero in the greenhouse bioassay (Table 1) Where insecticideshave such high efficiencies any fitness losses to BPH due to host resistance then becomeredundant Unlike buprofezin carbofuran also stimulated nymph biomass on IR64 in thesurvival bioassay (Table 1) however nymph biomass was lower on carbofuran-treated IR62than on non-treated IR62 indicating that the hostrsquos resistance buffered against undesiredstimulatory effects on BPH and may have functioned synergistically to augment mortalityand slow BPH growth Such buffering allows insecticides to be used against other pestswithout inducing secondary outbreaks of non-target pests In our screenhouse experimentherbivore densities were generally low on both IR62 and IR64 for most treatments (exceptdeltamethrin) and were below threshold densities for insecticide applicationsmdashwhichare normally about 25ndash50 planthoppers per hill [1373] Under such low densities it wasnot possible to observe the impact of host resistance on planthopper population build-
Crops 2021 1 178
up There was some indication of higher densities on cartap hydrochloride-treated IR62in the screenhouse which may be related to the stimulatory effects of the product onegg laying and the low mortality of nymphs on cartap hydrochloride-treated plants asobserved in the greenhouse bioassays (Table 1) Field studies from the Philippines haveindicated that IR62 effectively reduces BPH densities during wet and dry seasons [4]However farmers are rarely aware of which anti-herbivore resistance their rice varietiespossess and frequently apply pesticides regardless of resistance thereby reducing theirfarm profits [3841] For example Joshi et al (1992) [41] reported widespread outbreaks ofBPH on IR62 in Mindanao Philippines after applications of decamethrin (a pyrethroid) tothe resistant variety Where such resurgence-causing insecticides are applied resistanceseems to be often incapable of reducing the stimulatory effects of the insecticide and itloses its capacity to buffer against outbreaks
42 Antagonisms
Several insecticides are associated with increasing densities of planthoppers andwith resurgence outbreaks in farmersrsquo fields [27293041496174] Indeed patches oflsquohopperburnrsquo (ie dead rice plants resulting from planthopper damage) are often anindicator of high insecticide use in tropical Asia [214] In many cases these resurgence-causing insecticides were once effective in reducing planthopper densities but over timepopulations developed resistance against the products The resulting low toxicity canthen have a stimulatory effect on planthopper feeding and development [2123242743]This may be through hormesis which is a direct response by the planthopper to low-levelexposure to the toxic chemical or it may result from physiological changes to the rice plantsthat ultimately benefit the planthopper [1849] In our screenhouse study we induced aresurgence outbreak of BPH WBPH and to a lesser extent GLH on deltamethrin-treatedrice plants (Table 2) The combined densities of all three herbivores increased almost10 times on the treated plants Furthermore herbivore biomass increased by between fourand six times Although the resurgence effects of deltamethrin have been documentedin previous studies [2029ndash313975] we could not predict from our greenhouse studiesthat any such outbreak would occur Egg laying was unaffected by deltamethrin in ourchoice and no-choice experiments and nymph survival and development both declinedon deltamethrin-treated rice in the survival bioassay (Table 1) The massive increases inplanthopper densities on deltamethrin-treated plants in our study seem to result froma heightened attraction of adult BPH and WBPH to the plants [49] particularly sincenumbers were so low on all other plants including both treated and non-treated plantsHowever once on the plants planthoppers also apparently had enhanced fitness Cuonget al (1997) [31] observed higher population densities of BPH on deltamethrin-treatedrice in Vietnam albeit with lower-level increases on resistant varieties Furthermore in agreenhouse study Azzam et al (2009) [20] found that deltamethrin reduced pre-ovipositiontimes increased the duration of oviposition and increased egg hatchability and femalelongevity Nanthakumar et al (2012) [43] further showed that deltamethrin increased BPHfeeding efficiency on resistant and susceptible rice plants as determined through honeydewproduction and population size In studies with WBPH Salim and Heinrichs (1987) [36]showed deltamethrin to increase settling (antixenosis) feeding and population growthand Suri and Singh (2011) [28] showed deltamethrin to increase WBPH reproductive ratesand nymph survival In our study the observed resurgence of planthoppers was similaron both varieties indicating as was previously noted by Joshi et al (1992) [41] thatresurgence-causing pyrethroids are antagonistic to resistance in IR62
Though not statistically significant cartap hydrochloride also increased BPH numberson IR62 but not on IR64 in our screenhouse study This indicates that the product mayalso be antagonistic to the hostrsquos resistance mechanisms The stronger effect on IR62 maybe related to the differences between IR62 biomass growth rates and anatomy (ie alarger plant with higher tillering) compared to IR64 (Table 3) Once resistance is no longereffective because of chemically-enhanced BPH fitness these plant characteristics which
Crops 2021 1 179
have been related to observed preferences in BPH and other sap-sucking insects [349]could further accelerate the build-up of BPH populations on cartap hydrochloride-treatedplants To further test this idea experiments could be conducted using near-isogenic ricelines that differ mainly in the presence or absence of tagged resistance genes [76] Suchtests would avoid confounding differences in plant anatomy between test varieties IR62resistance to BPH has been well studied [4] The variety is also resistant to WBPH atolder plant stages [54] Therefore in our study deltamethrin was also antagonistic tothis WBPH-resistance (Table 2) However BPH feeding also facilitates WBPH populationgrowth [5477] and there may have been added indirect plant-mediated effects [77] thataugmented WBPH populations on both varieties in our experiment IR62 is also resistant toGLH [45] In our experiments GLH densities also increased on deltamethrin-treated plantsbut the effect was only apparent on IR64 Therefore GLH-resistance in IR62 apparentlybuffered against deltamethrin-associated increases in GLH on the variety as was alsonoted from field studies in Vietnam [31] Deltamethrin is currently not recommended foruse in rice [202878] however the product continues to be marketed in rice-dominatedregions throughout Asia [131479] Although cypermethrin has been associated with BPHoutbreaks and was consequently removed for use on rice in Thailand and Vietnam [62]we did not observe cypermethrin-induced resurgence in our screenhouse experimentNevertheless cypermethrin has been associated with resurgence of BPH in South Asia [61]and we have noted stimulatory effects of cypermethrin on other rice herbivores in thePhilippines (Horgan unpublished data)
43 Phytotoxicity
Phytotoxicity is relatively common following applications of insecticides [182850]Phytotoxic effects may change a plantrsquos physiology without consequences for crop yieldsHowever such changes to plant physiology could have indirect effects on herbivores Forexample deltamethrin methyl parathion and quinalphos produce increases in total sugarsand reducing sugars as well as increasing the protein contents of the rice leaf sheath andleaf blade [28] Several insecticides also reduce plant phenols in ricemdashwhich increases therice plantrsquos susceptibility to BPH and other insects [27] In our screenhouse experiment wenoted several changes to rice anatomy and yields Importantly we noted that cypermethrinand fipronil both reduced rice yields (Table 3) This was not due to indirect herbivoreeffects because planthopper and leafhopper densities were consistently low on rice treatedwith these insecticides The effects therefore probably resulted from direct phytotoxiceffects on the plants Furthermore these effects were most pronounced on treated IR62plants Other minor effects on grain production included increased proportions of unfilledgrain in cartap hydrochloride-treated plants and delays in grain maturation on cartaphydrochloride-treated and cypermethrin-treated plants (Table 3) Despite the high densitiesof BPH and WBPH on deltamethrin-treated plants yields were not significantly lower thanin control non-treated plants Both IR62 and IR64 have been noted for their high toleranceto BPH damage particularly under high nitrogenous fertilizer applications [3456] as usedin our screenhouse experiment Therefore where plants have high tolerance to damageapplying insecticides could also result in profitability losses on rice farms and in the caseof insecticide-induced pest population increases tolerance might buffer against subsequentyield losses There are few reports of the phytotoxicity of insecticides to rice carbofuran istoxic to rice at high concentrations but may also enhance the growth of rice plants throughinteractions with nitrogen-fixing bacteria in the rhizosphere [80] Future studies couldmore carefully assess field-grown plants for possible phytotoxic effects particularly ifinsecticides have the potential to directly reduce yields
44 Management implications
Global trends suggest that resistance in rice has failed to reduce insecticide appli-cations [2] This may be due to aggressive marketing by agrochemical companies andstrategies to increase pesticide sales such as promoting prophylactic applications [16] Be-
Crops 2021 1 180
cause we selected our test insecticides (except the pyrethroids) without any a priori criteriaour results suggest that many currently used insecticides probably have some negativeeffects on yield either directly through phytotoxic effects or indirectly by inducing pestresurgence Furthermore our results indicate that even where pests reach high densitiesas on deltamethrin-treated plants in the screenhouse these densities do not necessarilylead to significant yield reductions
Among the products we used buprofezin although it had little effect on oviposi-tion was effective in reducing nymph survival Furthermore there were only minorapparent phytotoxic effects on the rice plants Buprofezin has a low toxicity to naturalenemies in rice fields [58] and is therefore the most compatible of the insecticides withstrategies to increase rice field biodiversity and restore ecosystem services [32ndash348182]Thiamethoxam-chlorantraniliprole was also effective in reducing both oviposition andnymph survival and had no apparent toxic effects on rice plants However thiamethoxam-chlorantraniliprole and other neonicotinoids although moderately toxic can have severeeffects on beneficial hymenopteran parasitoids [83] although they have apparently lowtoxicity to predatory mirid bugs [84] The remaining insecticides either directly reducedyields or were associated with higher BPH biomass or densities in our experiments
We noted few synergies with HPR although nymph biomass andor survival waslower in cartap hydrochloride and carbofuran-treated IR62 plants compared to correspond-ing IR64 plants Meanwhile cartap hydrochloride and deltamethrin were antagonisticto resistance in IR62 in the screenhouse experiment Among the strongest effects of HPRwas to act as a buffer against GLH population build-up on deltamethrin-treated IR62 andagainst carbofuran-induced increases in BPH nymph biomass in the survival bioassayTherefore HPR may play a role in reducing the possibilities of secondary outbreaks ofBPH and other insects The combined use of HPR and targeted insecticides against BPHcreates redundancy in the rice field and counters measures to manage insecticide resistanceand virulence by unnecessarily exposing plant protection products to evolving pest pop-ulations HPR (and host plant tolerance) can be effective against target pests as revealedin previous studies [4] and could be incorporated into other pest management practicesparticularly agroecological practices such as ecological engineering with which it is highlycompatible [818586]
5 Conclusions
Based on our results and the results of previous studies HPR and the applicationof chemical insecticides against BPH rarely produce synergistic effects Antagonisticeffects where chemicals caused increases in BPH fitness or populations on resistant ricewere more apparent in our study In particular deltamethrin caused an outbreak ofBPH in our screenhouse study Nevertheless HPR can buffer against the insecticide-induced enhancement of herbivore fitness (hormesis) and potentially reduce possibilities ofsecondary outbreaks as was suggested from results with cartap hydrochloride in our studyBased on our results we suggest that carbofuran cartap hydrochloride cypermethrindeltamethrin and fipronil should be avoided in rice particularly in the Philippines (wherethis research was conducted) due to their resurgence-causing potential or because of directreductions to rice yields
Author Contributions Conceptualization FGH methodology FGH AP-C and MLPA formalanalysis FGH and AP-C investigation FGH AP-C and MLPA data curation FGHAP-C and MLPA writingmdashoriginal draft preparation FGH and AP-C writingmdashreview andediting FGH AP-C and MLPA visualization FGH supervision FGH and MLPA projectadministration FGH funding acquisition FGH All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by the Bill and Melinda Gates Foundation (Cereal SystemsInitiative for South Asia [CSISA] OPP52303) and the Global Rice Science Platform (GRiSP) underthe directorship of Achim Dobermann AP-C was partially funded by The Spanish Governmentthrough Becas Faro International
Crops 2021 1 181
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data presented in this study are available on reasonable requestfrom the corresponding author
Acknowledgments The authors thank Fame Ramal Carmencita Bernal Alberto Naredo ReyeulQuintana Vincent Virtudes Marol Recide Jenyrose Geronda and Ellen Genil for assistance duringthis study The contributions of Rodante Abas (sadly deceased) during three decades of entomologicalresearch at IRRI are gratefully acknowledged Rice lines were supplied through the Genebank of IRRI
Conflicts of Interest The authors declare no conflict of interest
References1 Bottrell DG Schoenly KG Resurrecting the ghost of green revolutions past The brown planthopper as a recurring threat to
high-yielding rice production in tropical Asia J Asia-Pac Entomol 2012 15 122ndash140 [CrossRef]2 Horgan FG Integrated pest management for sustainable rice cultivation A holistic approach In Achieving Sustainable Cultivation
of Rice Cultivation Pest and Disease Management Sasaki T Ed Burleigh Dodds Scientific Cambridge UK 2017 pp 309ndash3423 Horgan FG de Freitas TFS Crisol-Martiacutenez E Mundaca EA Bernal CC Nitrogenous fertilizer reduces resistance but
enhances tolerance to the brown planthopper in fast-growing moderately resistant rice Insects 2021 12 989 [CrossRef] [PubMed]4 Horgan FG Pentildealver Cruz A Bernal CC Ramal AF Almazan MLP Wilby A Resistance and tolerance to the brown
planthopper Nilaparvata lugens (Staringl) in rice infested at different growth stages across a gradient of nitrogen applicationsField Crops Res 2018 217 53ndash65 [CrossRef]
5 Hereward JP Cai X Matias AMA Walter GH Xu C Wang Y Migration dynamics of an important rice pest The brownplanthopper (Nilaparvata lugens) across AsiamdashInsights from population genomics Evol Appl 2020 13 2449ndash2459 [CrossRef]
6 Hu C Guo J Fu X Huang Y Gao X Wu K Seasonal migration pattern of Nilaparvata lugens (Hemiptera Delphacidae) overthe Bohai Sea in northern China J Econ Entomol 2018 111 2129ndash2135 [CrossRef]
7 Ma J Wang Y-C Hu Y-Y Lu M-H Wan G-J Chen F-J Liu W-C Zhai B-P Hu G Brown planthopper Nilaparvatalugens was concentrated at the rear of the typhoon Soudelor in eastern China in August 2015 Insect Sci 2018 25 916ndash926[CrossRef]
8 Fujita D Kohli A Horgan FG Rice resistance to planthoppers and leafhoppers Crit Rev Plant Sci 2013 32 162ndash191[CrossRef]
9 Fujii T Yoshida K Kobayashi T Myint KKM Yasui H Sanada-Morimura S Matsumura M Long-term virulencemonitoring of differential cultivars in Japanrsquos immigrant populations of Nilaparvata lugens (Hemiptera Delphacidae) in 2001ndash2009Appl Entomol Zool 2021 56 407ndash418 [CrossRef]
10 Myint KKM Yasui H Takagi M Matsumura M Virulence of long-term laboratory populations of the brown planthopperNilaparvata lugens (Stal) and whitebacked planthopper Sogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) on rice differentialvarieties Appl Entomol Zool 2009 44 149ndash153 [CrossRef]
11 Horgan FG Ramal AF Bentur JS Kumar R Bhanu KV Sarao PS Iswanto EH Van Chien H Phyu MHBernal CC et al Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia againstresistant rice varieties Crop Prot 2015 78 222ndash231 [CrossRef]
12 Horgan FG Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers Crop Prot2018 110 21ndash33 [CrossRef]
13 Uddin ABMA Islam KS Jahan M Ara A Afrin S Farmerrsquos perception about resurgence of brown planthopper NilaparvataLugens (Staringl) in Bangladesh Bangladesh Rice J 2020 23 21ndash33 [CrossRef]
14 Triwidodo H Brown planthoppers infestations and insecticides use pattern in Java Indonesia AGRIVITA J Agric Sci 202042 320ndash330 [CrossRef]
15 Veres A Wyckhuys KA Kiss J Toacuteth F Burgio G Pons X Avilla C Vidal S Razinger J Bazok R et al An update of theWorldwide Integrated Assessment (WIA) on systemic pesticides Part 4 Alternatives in major cropping systems Environ SciPollut Res 2020 27 29867ndash29899 [CrossRef]
16 Bakker L van der Werf W Tittonell P Wyckhuys KA Bianchi FJ Neonicotinoids in global agriculture Evidence for a newpesticide treadmill Ecol Soc 2020 25 26 [CrossRef]
17 Wu J-C Xu J-X Yuan S-Z Liu J-L Jiang Y-H Xu J-F Pesticide-induced susceptibility of rice to brown planthopperNilaparvata lugens Entomol Exp Appl 2001 100 119ndash126 [CrossRef]
18 Wu J-C Qiu H-M Yang G-Q Liu J-L Liu G-J Wilkens RM Effective duration of pesticide-induced susceptibility of riceto brown planthopper (Nilaparvata lugens Staringl Homoptera Delphacidae) and physiological and biochemical changes in riceplants following pesticide application Int J Pest Manag 2004 50 55ndash62 [CrossRef]
19 Wang A-H Wu J-C Yu Y-S Liu J-L Yue J-F Wang MY Selective insecticide-induced stimulation on fecundity andbiochemical changes in Tryporyza incertulas (Lepidoptera Pyralidae) J Econ Entomol 2005 98 1144ndash1149 [CrossRef] [PubMed]
Crops 2021 1 182
20 Azzam S Wang F Wu J-C Shen J Wang L-P Yang G-Q Guo Y-R Comparisons of stimulatory effects of a seriesof concentrations of four insecticides on reproduction in the rice brown planthopper Nilaparvata lugens Staringl (HomopteraDelphacidae) Int J Pest Manag 2009 55 347ndash358 [CrossRef]
21 Ge L-Q Hu J-H Wu J-C Yang G-Q Gu H Insecticide-induced changes in protein RNA and DNA contents in ovary andfat body of female Nilaparvata lugens (Hemiptera Delphacidae) J Econ Entomol 2009 102 1506ndash1514 [CrossRef]
22 Ling S Zhang J Hu L Zhang R Effect of fipronil on the reproduction feeding and relative fitness of brown planthopperNilaparvata lugens Appl Entomol Zool 2009 44 543ndash548 [CrossRef]
23 Ge L-Q Wang L-P Zhao K-F Wu J-C Huang L-J Mating pair combinations of insecticide-treated male and femaleNilaparvata lugens Staringl (Hemiptera Delphacidae) planthoppers influence protein content in the male accessory glands (MAGs)and vitellin content in both fat bodies and ovaries of adult females Pestic Biochem Physiol 2010 98 279ndash288 [CrossRef]
24 Ge L-Q Wu J-C Zhao K-F Chen Y Yang G-Q Induction of Nlvg and suppression of Nljhe gene expression in Nilaparvatalugens (Staringl) (Hemiptera Delphacidae) adult females and males exposed to two insecticides Pestic Biochem Physiol 201098 269ndash278 [CrossRef]
25 Hu J-H Wu J-C Yin J-L Gu H-N Physiology of insecticide-induced stimulation of reproduction in the rice brownplanthopper (Nilaparvata lugens (Staringl)) Dynamics of protein in fat body and ovary Int J Pest Manag 2010 56 23ndash30 [CrossRef]
26 Wang L-P Shen J Ge L-Q Wu J-C Yang G-Q Jahn GC Insecticide-induced increase in the protein content of maleaccessory glands and its effect on the fecundity of females in the brown planthopper Nilaparvata lugens Staringl (HemipteraDelphacidae) Crop Prot 2010 29 1280ndash1285 [CrossRef]
27 Wu J Ge L Liu F Song Q Stanley D Pesticide-induced planthopper population resurgence in rice cropping systems AnnuRev Entomol 2020 65 409ndash429 [CrossRef]
28 Suri KS Singh G Insecticide-induced resurgence of the whitebacked planthopper Sogatella furcifera (Horvath) (HemipteraDelphacidae) on rice varieties with different levels of resistance Crop Prot 2011 30 118ndash124 [CrossRef]
29 Fabellar LT Heinrichs EA Toxicity of insecticides to predators of rice brown planthoppers Nilaparvata lugens (Staringl) (HomopteraDelphacidae) Environ Entomol 1984 13 832ndash837 [CrossRef]
30 Fabellar LT Heinrichs EA Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators Crop Prot1986 5 254ndash258 [CrossRef]
31 Cuong NL Ben PT Phuong LT Chau LM Cohen MB Effect of host plant resistance and insecticide on brown planthopperNilaparvata lugens (Staringl) and predator population development in the Mekong Delta Vietnam Crop Prot 1997 16 707ndash715[CrossRef]
32 Preetha G Stanley J Suresh S Samiyappan R Risk assessment of insecticides used in rice on mirid bug Cyrtorhinuslividipennis Reuter the important predator of brown planthopper Nilaparvata lugens (Stal) Chemosphere 2010 80 498ndash503[CrossRef] [PubMed]
33 Tanaka K Endo S Kazano H Toxicity of insecticides to predators of rice planthoppers Spiders the mirid bug and the dryinidwasp Appl Entomol Zool 2000 35 177ndash187 [CrossRef]
34 Zhang X Xu Q Lu W Liu F Sublethal effects of four synthetic insecticides on the generalist predator Cyrtorhinus lividipennisJ Pest Sci 2015 88 383ndash392 [CrossRef]
35 Settle WH Ariawan H Astuti ET Cahyana W Hakim AL Hindayana D Lestari AS Managing tropical rice peststhrough conservation of generalist natural enemies and alternative prey Ecology 1996 77 1975ndash1988 [CrossRef]
36 Salim M Heinrichs EA Insecticide-induced changes in the levels of resistance of rice cultivars to the whitebacked planthopperSogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) Crop Prot 1987 6 28ndash32 [CrossRef]
37 Yin J-L Xu H-W Wu J-C Hu J-H Yang G-Q Cultivar and insecticide applications affect the physiological development ofthe brown planthopper Nilaparvata lugens (Staringl) (Hemiptera Delphacidae) Environ Entomol 2014 37 206ndash212 [CrossRef]
38 Cheng C Chang W Studies on varietal resistance to the brown planthopper in Taiwan In Brown Planthopper Threat toRice Production in Asia International Rice Research Institute Ed International Rice Research Institute Manila Philippines1979 pp 251ndash271
39 Reissig WH Heinrichs EA Valencia SL Insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on ricevarieties with different levels of resistance Environ Entomol 1982 11 165ndash168 [CrossRef]
40 Heinrichs EA Fabellar LT Basilio RP Wen T-C Medrano F Susceptibility of rice planthoppers Nilaparvata lugens andSogatella furcifera (Homoptera Delphacidae) to insecticides as influenced by level of resistance in the host plant Environ Entomol1984 13 455ndash458 [CrossRef]
41 Joshi R Shepard B Kenmore P Lydia R Insecticide-induced resurgence of brown planthopper (BPH) on IR62 Int Rice ResNewsl 1992 17 9ndash10
42 Zhu Z-R Cheng J Jiang M-X Zhang X-X Complex influence of rice variety fertilization timing and insecticide onpopulation dynamics of Sogatella furcifera (Horvath) Nilaparvata lugens (Staringl) (Homoptera Delphacidae) and their natural enemiesin rice in Hangzhou China J Pest Sci 2004 77 65ndash74 [CrossRef]
43 Nanthakumar M Jhansi Lakshmi V Shashi Bhushan V Balachandran SM Mohan M Decrease of rice plant resistance andinduction of hormesis and carboxylesterase titre in brown planthopper Nilaparvata lugens (Staringl) by xenobiotics Pestic BiochemPhysiol 2012 102 146ndash152 [CrossRef]
Crops 2021 1 183
44 Yao H-W Jiang C-Y Ye G-Y Cheng J-A Studies on cultivar-induced changes in insecticide susceptibility and its relatedenzyme activities of the white-backed planthopper Sogatella furcifera (Horvath)(Homopter Delphacidae) Insect Sci 2002 9 9ndash15
45 Horgan FG Srinivasan TS Bentur JS Kumar R Bhanu KV Sarao PS Van Chien H Almazan MLP Bernal CCRamal AF et al Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers Implicationsfor rice breeding and gene deployment Agronomy 2017 7 62 [CrossRef]
46 Jairin J Phengrat K Teangdeerith S Vanavichit A Toojinda T Mapping of a broad-spectrum brown planthopper resistancegene Bph3 on rice chromosome 6 Mol Breed 2007 19 35ndash44 [CrossRef]
47 Ren J Gao F Wu X Lu X Zeng L Lv J Su X Luo H Ren G Bph32 a novel gene encoding an unknown SCRdomain-containing protein confers resistance against the brown planthopper in rice Sci Rep 2016 6 37645 [CrossRef]
48 Nguyen CD Zheng S-H Sanada-Morimura S Matsumura M Yasui H Fujita D Substitution mapping and characterizationof brown planthopper resistance genes from indica rice variety lsquoPTB33prime (Oryza sativa L) Breed Sci 2021 in press [CrossRef]
49 Chelliah S Heinrichs EA Factors affecting insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on RiceEnviron Entomol 1980 9 773ndash777 [CrossRef]
50 Moore MT Kroumlger R Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growthArch Environ Contam Toxicol 2010 59 574ndash581 [CrossRef]
51 Kim J Hwang I Han M Jang B Effect of insecticide and fungicide on phytotoxicity of herbicide in rice Korean J Weed Sci1986 6 67ndash75
52 Horgan FG Garcia CPF Haverkort F de Jong PW Ferrater JB Changes in insecticide resistance and host range performanceof planthoppers artificially selected to feed on resistant rice Crop Prot 2020 127 104963 [CrossRef] [PubMed]
53 Khush GS Virk PS IR Varieties and Their Impact International Rice Research Institute Manila Philippines 200554 Srinivasan TS Almazan MLP Bernal CC Ramal AF Subbarayalu MK Horgan FG Interactions between nymphs of
Nilaparvata lugens and Sogatella furcifera (Hemiptera Delphacidae) on resistant and susceptible rice varieties Appl Entomol Zool2016 51 81ndash90 [CrossRef]
55 Pentildealver Cruz A Arida A Heong KL Horgan FG Aspects of brown planthopper adaptation to resistant rice varieties withthe Bph3 gene Entomol Exp Appl 2011 141 245ndash257 [CrossRef]
56 Alam SN Cohen MB Detection and analysis of QTLs for resistance to the brown planthopper Nilaparvata lugens in adoubled-haploid rice population Theor Appl Genet 1998 97 1370ndash1379 [CrossRef]
57 Mackill DJ Khush GS IR64 A high-quality and high-yielding mega variety Rice 2018 11 18 [CrossRef] [PubMed]58 Heinrichs EA Basilio RP Valencia SL Buprofezin a selective insecticide for the management of rice planthoppers (Ho-
moptera Delphacidae) and leafhoppers (Homoptera Cicadellidae) Environ Entomol 1984 13 515ndash521 [CrossRef]59 Satapathy MK Anjaneyulu A Use of cypermethrin a synthetic pyrethroid in the control of rice tungro virus disease and its
vector Trop Pest Manag 1984 30 170ndash178 [CrossRef]60 Vorley W Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown
planthopper (BPH) in Kedah Malaysia Int Rice Res Newsl 1987 10 19ndash2061 Uddin A Islam KS Jahan M Ara A Khan MAI Factor influencing the resurgence of brown planthopper in Bangladesh
SAARC J Agric 2020 18 117ndash128 [CrossRef]62 Escalada MM Luecha M Heong KL Social impacts of planthopper outbreaks Case study from Thailand In Rice Planthoppers
Ecology Management Socio Economics and Policy Heong KL Cheng J Escalada MM Eds Zhejiang University PressHangzhou China 2015 pp 209ndash226
63 Zhang CP He HM Yu JZ Hu XQ Zhu YH Wang Q Residues of carbosulfan and its metabolites carbofuran and3-hydroxy carbofuran in rice field ecosystem in China J Environ Sci Health 2016 51 351ndash357 [CrossRef] [PubMed]
64 Samanta S Barman M Samanta NRA Bio-efficacy trials of carbofuran 3 CG against insect pests of rice J Entomol ZoolStud 2020 8 1529ndash1533
65 Donley N The USA lags behind other agricultural nations in banning harmful pesticides Environ Health 2019 18 44 [CrossRef][PubMed]
66 Adams A Gore J Musser F Cook D Catchot A Walker T Dobbins C Efficacy of selected insecticides applied to hybridrice seed J Econ Entomol 2015 109 200ndash206 [CrossRef]
67 Kumar R Singh B Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) Field Bull Environ Contam Toxicol2013 90 482ndash488 [CrossRef]
68 Dhaka S Prajapati C Singh D Singh R Field evaluation of insecticides and bio-pesticides against rice leaf folder CnaphalocrocisMedinalis (Guenee) Ann Plant Prot Sci 2011 19 324ndash326
69 Matsumura M Takeuchi H Satoh M Sanada-Morimura S Otuka A Watanabe T Van Thanh D Species-specific insecticideresistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in East and South-eastAsia Pest Manag Sci 2008 64 1115ndash1121 [CrossRef] [PubMed]
70 Tang J Li J Shao Y Yang B Liu Z Fipronil resistance in the whitebacked planthopper (Sogatella furcifera) Possible resistancemechanisms and cross-resistance Pest Manag Sci 2010 66 121ndash125 [CrossRef]
71 Horgan FG Arida A Ardestani G Almazan MLP Temperature-dependent oviposition and nymph performance revealdistinct thermal niches of coexisting planthoppers with similar thresholds for development PLoS ONE 2020 15 e0235506[CrossRef] [PubMed]
Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]

Crops 2021 1 178
up There was some indication of higher densities on cartap hydrochloride-treated IR62in the screenhouse which may be related to the stimulatory effects of the product onegg laying and the low mortality of nymphs on cartap hydrochloride-treated plants asobserved in the greenhouse bioassays (Table 1) Field studies from the Philippines haveindicated that IR62 effectively reduces BPH densities during wet and dry seasons [4]However farmers are rarely aware of which anti-herbivore resistance their rice varietiespossess and frequently apply pesticides regardless of resistance thereby reducing theirfarm profits [3841] For example Joshi et al (1992) [41] reported widespread outbreaks ofBPH on IR62 in Mindanao Philippines after applications of decamethrin (a pyrethroid) tothe resistant variety Where such resurgence-causing insecticides are applied resistanceseems to be often incapable of reducing the stimulatory effects of the insecticide and itloses its capacity to buffer against outbreaks
42 Antagonisms
Several insecticides are associated with increasing densities of planthoppers andwith resurgence outbreaks in farmersrsquo fields [27293041496174] Indeed patches oflsquohopperburnrsquo (ie dead rice plants resulting from planthopper damage) are often anindicator of high insecticide use in tropical Asia [214] In many cases these resurgence-causing insecticides were once effective in reducing planthopper densities but over timepopulations developed resistance against the products The resulting low toxicity canthen have a stimulatory effect on planthopper feeding and development [2123242743]This may be through hormesis which is a direct response by the planthopper to low-levelexposure to the toxic chemical or it may result from physiological changes to the rice plantsthat ultimately benefit the planthopper [1849] In our screenhouse study we induced aresurgence outbreak of BPH WBPH and to a lesser extent GLH on deltamethrin-treatedrice plants (Table 2) The combined densities of all three herbivores increased almost10 times on the treated plants Furthermore herbivore biomass increased by between fourand six times Although the resurgence effects of deltamethrin have been documentedin previous studies [2029ndash313975] we could not predict from our greenhouse studiesthat any such outbreak would occur Egg laying was unaffected by deltamethrin in ourchoice and no-choice experiments and nymph survival and development both declinedon deltamethrin-treated rice in the survival bioassay (Table 1) The massive increases inplanthopper densities on deltamethrin-treated plants in our study seem to result froma heightened attraction of adult BPH and WBPH to the plants [49] particularly sincenumbers were so low on all other plants including both treated and non-treated plantsHowever once on the plants planthoppers also apparently had enhanced fitness Cuonget al (1997) [31] observed higher population densities of BPH on deltamethrin-treatedrice in Vietnam albeit with lower-level increases on resistant varieties Furthermore in agreenhouse study Azzam et al (2009) [20] found that deltamethrin reduced pre-ovipositiontimes increased the duration of oviposition and increased egg hatchability and femalelongevity Nanthakumar et al (2012) [43] further showed that deltamethrin increased BPHfeeding efficiency on resistant and susceptible rice plants as determined through honeydewproduction and population size In studies with WBPH Salim and Heinrichs (1987) [36]showed deltamethrin to increase settling (antixenosis) feeding and population growthand Suri and Singh (2011) [28] showed deltamethrin to increase WBPH reproductive ratesand nymph survival In our study the observed resurgence of planthoppers was similaron both varieties indicating as was previously noted by Joshi et al (1992) [41] thatresurgence-causing pyrethroids are antagonistic to resistance in IR62
Though not statistically significant cartap hydrochloride also increased BPH numberson IR62 but not on IR64 in our screenhouse study This indicates that the product mayalso be antagonistic to the hostrsquos resistance mechanisms The stronger effect on IR62 maybe related to the differences between IR62 biomass growth rates and anatomy (ie alarger plant with higher tillering) compared to IR64 (Table 3) Once resistance is no longereffective because of chemically-enhanced BPH fitness these plant characteristics which
Crops 2021 1 179
have been related to observed preferences in BPH and other sap-sucking insects [349]could further accelerate the build-up of BPH populations on cartap hydrochloride-treatedplants To further test this idea experiments could be conducted using near-isogenic ricelines that differ mainly in the presence or absence of tagged resistance genes [76] Suchtests would avoid confounding differences in plant anatomy between test varieties IR62resistance to BPH has been well studied [4] The variety is also resistant to WBPH atolder plant stages [54] Therefore in our study deltamethrin was also antagonistic tothis WBPH-resistance (Table 2) However BPH feeding also facilitates WBPH populationgrowth [5477] and there may have been added indirect plant-mediated effects [77] thataugmented WBPH populations on both varieties in our experiment IR62 is also resistant toGLH [45] In our experiments GLH densities also increased on deltamethrin-treated plantsbut the effect was only apparent on IR64 Therefore GLH-resistance in IR62 apparentlybuffered against deltamethrin-associated increases in GLH on the variety as was alsonoted from field studies in Vietnam [31] Deltamethrin is currently not recommended foruse in rice [202878] however the product continues to be marketed in rice-dominatedregions throughout Asia [131479] Although cypermethrin has been associated with BPHoutbreaks and was consequently removed for use on rice in Thailand and Vietnam [62]we did not observe cypermethrin-induced resurgence in our screenhouse experimentNevertheless cypermethrin has been associated with resurgence of BPH in South Asia [61]and we have noted stimulatory effects of cypermethrin on other rice herbivores in thePhilippines (Horgan unpublished data)
43 Phytotoxicity
Phytotoxicity is relatively common following applications of insecticides [182850]Phytotoxic effects may change a plantrsquos physiology without consequences for crop yieldsHowever such changes to plant physiology could have indirect effects on herbivores Forexample deltamethrin methyl parathion and quinalphos produce increases in total sugarsand reducing sugars as well as increasing the protein contents of the rice leaf sheath andleaf blade [28] Several insecticides also reduce plant phenols in ricemdashwhich increases therice plantrsquos susceptibility to BPH and other insects [27] In our screenhouse experiment wenoted several changes to rice anatomy and yields Importantly we noted that cypermethrinand fipronil both reduced rice yields (Table 3) This was not due to indirect herbivoreeffects because planthopper and leafhopper densities were consistently low on rice treatedwith these insecticides The effects therefore probably resulted from direct phytotoxiceffects on the plants Furthermore these effects were most pronounced on treated IR62plants Other minor effects on grain production included increased proportions of unfilledgrain in cartap hydrochloride-treated plants and delays in grain maturation on cartaphydrochloride-treated and cypermethrin-treated plants (Table 3) Despite the high densitiesof BPH and WBPH on deltamethrin-treated plants yields were not significantly lower thanin control non-treated plants Both IR62 and IR64 have been noted for their high toleranceto BPH damage particularly under high nitrogenous fertilizer applications [3456] as usedin our screenhouse experiment Therefore where plants have high tolerance to damageapplying insecticides could also result in profitability losses on rice farms and in the caseof insecticide-induced pest population increases tolerance might buffer against subsequentyield losses There are few reports of the phytotoxicity of insecticides to rice carbofuran istoxic to rice at high concentrations but may also enhance the growth of rice plants throughinteractions with nitrogen-fixing bacteria in the rhizosphere [80] Future studies couldmore carefully assess field-grown plants for possible phytotoxic effects particularly ifinsecticides have the potential to directly reduce yields
44 Management implications
Global trends suggest that resistance in rice has failed to reduce insecticide appli-cations [2] This may be due to aggressive marketing by agrochemical companies andstrategies to increase pesticide sales such as promoting prophylactic applications [16] Be-
Crops 2021 1 180
cause we selected our test insecticides (except the pyrethroids) without any a priori criteriaour results suggest that many currently used insecticides probably have some negativeeffects on yield either directly through phytotoxic effects or indirectly by inducing pestresurgence Furthermore our results indicate that even where pests reach high densitiesas on deltamethrin-treated plants in the screenhouse these densities do not necessarilylead to significant yield reductions
Among the products we used buprofezin although it had little effect on oviposi-tion was effective in reducing nymph survival Furthermore there were only minorapparent phytotoxic effects on the rice plants Buprofezin has a low toxicity to naturalenemies in rice fields [58] and is therefore the most compatible of the insecticides withstrategies to increase rice field biodiversity and restore ecosystem services [32ndash348182]Thiamethoxam-chlorantraniliprole was also effective in reducing both oviposition andnymph survival and had no apparent toxic effects on rice plants However thiamethoxam-chlorantraniliprole and other neonicotinoids although moderately toxic can have severeeffects on beneficial hymenopteran parasitoids [83] although they have apparently lowtoxicity to predatory mirid bugs [84] The remaining insecticides either directly reducedyields or were associated with higher BPH biomass or densities in our experiments
We noted few synergies with HPR although nymph biomass andor survival waslower in cartap hydrochloride and carbofuran-treated IR62 plants compared to correspond-ing IR64 plants Meanwhile cartap hydrochloride and deltamethrin were antagonisticto resistance in IR62 in the screenhouse experiment Among the strongest effects of HPRwas to act as a buffer against GLH population build-up on deltamethrin-treated IR62 andagainst carbofuran-induced increases in BPH nymph biomass in the survival bioassayTherefore HPR may play a role in reducing the possibilities of secondary outbreaks ofBPH and other insects The combined use of HPR and targeted insecticides against BPHcreates redundancy in the rice field and counters measures to manage insecticide resistanceand virulence by unnecessarily exposing plant protection products to evolving pest pop-ulations HPR (and host plant tolerance) can be effective against target pests as revealedin previous studies [4] and could be incorporated into other pest management practicesparticularly agroecological practices such as ecological engineering with which it is highlycompatible [818586]
5 Conclusions
Based on our results and the results of previous studies HPR and the applicationof chemical insecticides against BPH rarely produce synergistic effects Antagonisticeffects where chemicals caused increases in BPH fitness or populations on resistant ricewere more apparent in our study In particular deltamethrin caused an outbreak ofBPH in our screenhouse study Nevertheless HPR can buffer against the insecticide-induced enhancement of herbivore fitness (hormesis) and potentially reduce possibilities ofsecondary outbreaks as was suggested from results with cartap hydrochloride in our studyBased on our results we suggest that carbofuran cartap hydrochloride cypermethrindeltamethrin and fipronil should be avoided in rice particularly in the Philippines (wherethis research was conducted) due to their resurgence-causing potential or because of directreductions to rice yields
Author Contributions Conceptualization FGH methodology FGH AP-C and MLPA formalanalysis FGH and AP-C investigation FGH AP-C and MLPA data curation FGHAP-C and MLPA writingmdashoriginal draft preparation FGH and AP-C writingmdashreview andediting FGH AP-C and MLPA visualization FGH supervision FGH and MLPA projectadministration FGH funding acquisition FGH All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by the Bill and Melinda Gates Foundation (Cereal SystemsInitiative for South Asia [CSISA] OPP52303) and the Global Rice Science Platform (GRiSP) underthe directorship of Achim Dobermann AP-C was partially funded by The Spanish Governmentthrough Becas Faro International
Crops 2021 1 181
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data presented in this study are available on reasonable requestfrom the corresponding author
Acknowledgments The authors thank Fame Ramal Carmencita Bernal Alberto Naredo ReyeulQuintana Vincent Virtudes Marol Recide Jenyrose Geronda and Ellen Genil for assistance duringthis study The contributions of Rodante Abas (sadly deceased) during three decades of entomologicalresearch at IRRI are gratefully acknowledged Rice lines were supplied through the Genebank of IRRI
Conflicts of Interest The authors declare no conflict of interest
References1 Bottrell DG Schoenly KG Resurrecting the ghost of green revolutions past The brown planthopper as a recurring threat to
high-yielding rice production in tropical Asia J Asia-Pac Entomol 2012 15 122ndash140 [CrossRef]2 Horgan FG Integrated pest management for sustainable rice cultivation A holistic approach In Achieving Sustainable Cultivation
of Rice Cultivation Pest and Disease Management Sasaki T Ed Burleigh Dodds Scientific Cambridge UK 2017 pp 309ndash3423 Horgan FG de Freitas TFS Crisol-Martiacutenez E Mundaca EA Bernal CC Nitrogenous fertilizer reduces resistance but
enhances tolerance to the brown planthopper in fast-growing moderately resistant rice Insects 2021 12 989 [CrossRef] [PubMed]4 Horgan FG Pentildealver Cruz A Bernal CC Ramal AF Almazan MLP Wilby A Resistance and tolerance to the brown
planthopper Nilaparvata lugens (Staringl) in rice infested at different growth stages across a gradient of nitrogen applicationsField Crops Res 2018 217 53ndash65 [CrossRef]
5 Hereward JP Cai X Matias AMA Walter GH Xu C Wang Y Migration dynamics of an important rice pest The brownplanthopper (Nilaparvata lugens) across AsiamdashInsights from population genomics Evol Appl 2020 13 2449ndash2459 [CrossRef]
6 Hu C Guo J Fu X Huang Y Gao X Wu K Seasonal migration pattern of Nilaparvata lugens (Hemiptera Delphacidae) overthe Bohai Sea in northern China J Econ Entomol 2018 111 2129ndash2135 [CrossRef]
7 Ma J Wang Y-C Hu Y-Y Lu M-H Wan G-J Chen F-J Liu W-C Zhai B-P Hu G Brown planthopper Nilaparvatalugens was concentrated at the rear of the typhoon Soudelor in eastern China in August 2015 Insect Sci 2018 25 916ndash926[CrossRef]
8 Fujita D Kohli A Horgan FG Rice resistance to planthoppers and leafhoppers Crit Rev Plant Sci 2013 32 162ndash191[CrossRef]
9 Fujii T Yoshida K Kobayashi T Myint KKM Yasui H Sanada-Morimura S Matsumura M Long-term virulencemonitoring of differential cultivars in Japanrsquos immigrant populations of Nilaparvata lugens (Hemiptera Delphacidae) in 2001ndash2009Appl Entomol Zool 2021 56 407ndash418 [CrossRef]
10 Myint KKM Yasui H Takagi M Matsumura M Virulence of long-term laboratory populations of the brown planthopperNilaparvata lugens (Stal) and whitebacked planthopper Sogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) on rice differentialvarieties Appl Entomol Zool 2009 44 149ndash153 [CrossRef]
11 Horgan FG Ramal AF Bentur JS Kumar R Bhanu KV Sarao PS Iswanto EH Van Chien H Phyu MHBernal CC et al Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia againstresistant rice varieties Crop Prot 2015 78 222ndash231 [CrossRef]
12 Horgan FG Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers Crop Prot2018 110 21ndash33 [CrossRef]
13 Uddin ABMA Islam KS Jahan M Ara A Afrin S Farmerrsquos perception about resurgence of brown planthopper NilaparvataLugens (Staringl) in Bangladesh Bangladesh Rice J 2020 23 21ndash33 [CrossRef]
14 Triwidodo H Brown planthoppers infestations and insecticides use pattern in Java Indonesia AGRIVITA J Agric Sci 202042 320ndash330 [CrossRef]
15 Veres A Wyckhuys KA Kiss J Toacuteth F Burgio G Pons X Avilla C Vidal S Razinger J Bazok R et al An update of theWorldwide Integrated Assessment (WIA) on systemic pesticides Part 4 Alternatives in major cropping systems Environ SciPollut Res 2020 27 29867ndash29899 [CrossRef]
16 Bakker L van der Werf W Tittonell P Wyckhuys KA Bianchi FJ Neonicotinoids in global agriculture Evidence for a newpesticide treadmill Ecol Soc 2020 25 26 [CrossRef]
17 Wu J-C Xu J-X Yuan S-Z Liu J-L Jiang Y-H Xu J-F Pesticide-induced susceptibility of rice to brown planthopperNilaparvata lugens Entomol Exp Appl 2001 100 119ndash126 [CrossRef]
18 Wu J-C Qiu H-M Yang G-Q Liu J-L Liu G-J Wilkens RM Effective duration of pesticide-induced susceptibility of riceto brown planthopper (Nilaparvata lugens Staringl Homoptera Delphacidae) and physiological and biochemical changes in riceplants following pesticide application Int J Pest Manag 2004 50 55ndash62 [CrossRef]
19 Wang A-H Wu J-C Yu Y-S Liu J-L Yue J-F Wang MY Selective insecticide-induced stimulation on fecundity andbiochemical changes in Tryporyza incertulas (Lepidoptera Pyralidae) J Econ Entomol 2005 98 1144ndash1149 [CrossRef] [PubMed]
Crops 2021 1 182
20 Azzam S Wang F Wu J-C Shen J Wang L-P Yang G-Q Guo Y-R Comparisons of stimulatory effects of a seriesof concentrations of four insecticides on reproduction in the rice brown planthopper Nilaparvata lugens Staringl (HomopteraDelphacidae) Int J Pest Manag 2009 55 347ndash358 [CrossRef]
21 Ge L-Q Hu J-H Wu J-C Yang G-Q Gu H Insecticide-induced changes in protein RNA and DNA contents in ovary andfat body of female Nilaparvata lugens (Hemiptera Delphacidae) J Econ Entomol 2009 102 1506ndash1514 [CrossRef]
22 Ling S Zhang J Hu L Zhang R Effect of fipronil on the reproduction feeding and relative fitness of brown planthopperNilaparvata lugens Appl Entomol Zool 2009 44 543ndash548 [CrossRef]
23 Ge L-Q Wang L-P Zhao K-F Wu J-C Huang L-J Mating pair combinations of insecticide-treated male and femaleNilaparvata lugens Staringl (Hemiptera Delphacidae) planthoppers influence protein content in the male accessory glands (MAGs)and vitellin content in both fat bodies and ovaries of adult females Pestic Biochem Physiol 2010 98 279ndash288 [CrossRef]
24 Ge L-Q Wu J-C Zhao K-F Chen Y Yang G-Q Induction of Nlvg and suppression of Nljhe gene expression in Nilaparvatalugens (Staringl) (Hemiptera Delphacidae) adult females and males exposed to two insecticides Pestic Biochem Physiol 201098 269ndash278 [CrossRef]
25 Hu J-H Wu J-C Yin J-L Gu H-N Physiology of insecticide-induced stimulation of reproduction in the rice brownplanthopper (Nilaparvata lugens (Staringl)) Dynamics of protein in fat body and ovary Int J Pest Manag 2010 56 23ndash30 [CrossRef]
26 Wang L-P Shen J Ge L-Q Wu J-C Yang G-Q Jahn GC Insecticide-induced increase in the protein content of maleaccessory glands and its effect on the fecundity of females in the brown planthopper Nilaparvata lugens Staringl (HemipteraDelphacidae) Crop Prot 2010 29 1280ndash1285 [CrossRef]
27 Wu J Ge L Liu F Song Q Stanley D Pesticide-induced planthopper population resurgence in rice cropping systems AnnuRev Entomol 2020 65 409ndash429 [CrossRef]
28 Suri KS Singh G Insecticide-induced resurgence of the whitebacked planthopper Sogatella furcifera (Horvath) (HemipteraDelphacidae) on rice varieties with different levels of resistance Crop Prot 2011 30 118ndash124 [CrossRef]
29 Fabellar LT Heinrichs EA Toxicity of insecticides to predators of rice brown planthoppers Nilaparvata lugens (Staringl) (HomopteraDelphacidae) Environ Entomol 1984 13 832ndash837 [CrossRef]
30 Fabellar LT Heinrichs EA Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators Crop Prot1986 5 254ndash258 [CrossRef]
31 Cuong NL Ben PT Phuong LT Chau LM Cohen MB Effect of host plant resistance and insecticide on brown planthopperNilaparvata lugens (Staringl) and predator population development in the Mekong Delta Vietnam Crop Prot 1997 16 707ndash715[CrossRef]
32 Preetha G Stanley J Suresh S Samiyappan R Risk assessment of insecticides used in rice on mirid bug Cyrtorhinuslividipennis Reuter the important predator of brown planthopper Nilaparvata lugens (Stal) Chemosphere 2010 80 498ndash503[CrossRef] [PubMed]
33 Tanaka K Endo S Kazano H Toxicity of insecticides to predators of rice planthoppers Spiders the mirid bug and the dryinidwasp Appl Entomol Zool 2000 35 177ndash187 [CrossRef]
34 Zhang X Xu Q Lu W Liu F Sublethal effects of four synthetic insecticides on the generalist predator Cyrtorhinus lividipennisJ Pest Sci 2015 88 383ndash392 [CrossRef]
35 Settle WH Ariawan H Astuti ET Cahyana W Hakim AL Hindayana D Lestari AS Managing tropical rice peststhrough conservation of generalist natural enemies and alternative prey Ecology 1996 77 1975ndash1988 [CrossRef]
36 Salim M Heinrichs EA Insecticide-induced changes in the levels of resistance of rice cultivars to the whitebacked planthopperSogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) Crop Prot 1987 6 28ndash32 [CrossRef]
37 Yin J-L Xu H-W Wu J-C Hu J-H Yang G-Q Cultivar and insecticide applications affect the physiological development ofthe brown planthopper Nilaparvata lugens (Staringl) (Hemiptera Delphacidae) Environ Entomol 2014 37 206ndash212 [CrossRef]
38 Cheng C Chang W Studies on varietal resistance to the brown planthopper in Taiwan In Brown Planthopper Threat toRice Production in Asia International Rice Research Institute Ed International Rice Research Institute Manila Philippines1979 pp 251ndash271
39 Reissig WH Heinrichs EA Valencia SL Insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on ricevarieties with different levels of resistance Environ Entomol 1982 11 165ndash168 [CrossRef]
40 Heinrichs EA Fabellar LT Basilio RP Wen T-C Medrano F Susceptibility of rice planthoppers Nilaparvata lugens andSogatella furcifera (Homoptera Delphacidae) to insecticides as influenced by level of resistance in the host plant Environ Entomol1984 13 455ndash458 [CrossRef]
41 Joshi R Shepard B Kenmore P Lydia R Insecticide-induced resurgence of brown planthopper (BPH) on IR62 Int Rice ResNewsl 1992 17 9ndash10
42 Zhu Z-R Cheng J Jiang M-X Zhang X-X Complex influence of rice variety fertilization timing and insecticide onpopulation dynamics of Sogatella furcifera (Horvath) Nilaparvata lugens (Staringl) (Homoptera Delphacidae) and their natural enemiesin rice in Hangzhou China J Pest Sci 2004 77 65ndash74 [CrossRef]
43 Nanthakumar M Jhansi Lakshmi V Shashi Bhushan V Balachandran SM Mohan M Decrease of rice plant resistance andinduction of hormesis and carboxylesterase titre in brown planthopper Nilaparvata lugens (Staringl) by xenobiotics Pestic BiochemPhysiol 2012 102 146ndash152 [CrossRef]
Crops 2021 1 183
44 Yao H-W Jiang C-Y Ye G-Y Cheng J-A Studies on cultivar-induced changes in insecticide susceptibility and its relatedenzyme activities of the white-backed planthopper Sogatella furcifera (Horvath)(Homopter Delphacidae) Insect Sci 2002 9 9ndash15
45 Horgan FG Srinivasan TS Bentur JS Kumar R Bhanu KV Sarao PS Van Chien H Almazan MLP Bernal CCRamal AF et al Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers Implicationsfor rice breeding and gene deployment Agronomy 2017 7 62 [CrossRef]
46 Jairin J Phengrat K Teangdeerith S Vanavichit A Toojinda T Mapping of a broad-spectrum brown planthopper resistancegene Bph3 on rice chromosome 6 Mol Breed 2007 19 35ndash44 [CrossRef]
47 Ren J Gao F Wu X Lu X Zeng L Lv J Su X Luo H Ren G Bph32 a novel gene encoding an unknown SCRdomain-containing protein confers resistance against the brown planthopper in rice Sci Rep 2016 6 37645 [CrossRef]
48 Nguyen CD Zheng S-H Sanada-Morimura S Matsumura M Yasui H Fujita D Substitution mapping and characterizationof brown planthopper resistance genes from indica rice variety lsquoPTB33prime (Oryza sativa L) Breed Sci 2021 in press [CrossRef]
49 Chelliah S Heinrichs EA Factors affecting insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on RiceEnviron Entomol 1980 9 773ndash777 [CrossRef]
50 Moore MT Kroumlger R Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growthArch Environ Contam Toxicol 2010 59 574ndash581 [CrossRef]
51 Kim J Hwang I Han M Jang B Effect of insecticide and fungicide on phytotoxicity of herbicide in rice Korean J Weed Sci1986 6 67ndash75
52 Horgan FG Garcia CPF Haverkort F de Jong PW Ferrater JB Changes in insecticide resistance and host range performanceof planthoppers artificially selected to feed on resistant rice Crop Prot 2020 127 104963 [CrossRef] [PubMed]
53 Khush GS Virk PS IR Varieties and Their Impact International Rice Research Institute Manila Philippines 200554 Srinivasan TS Almazan MLP Bernal CC Ramal AF Subbarayalu MK Horgan FG Interactions between nymphs of
Nilaparvata lugens and Sogatella furcifera (Hemiptera Delphacidae) on resistant and susceptible rice varieties Appl Entomol Zool2016 51 81ndash90 [CrossRef]
55 Pentildealver Cruz A Arida A Heong KL Horgan FG Aspects of brown planthopper adaptation to resistant rice varieties withthe Bph3 gene Entomol Exp Appl 2011 141 245ndash257 [CrossRef]
56 Alam SN Cohen MB Detection and analysis of QTLs for resistance to the brown planthopper Nilaparvata lugens in adoubled-haploid rice population Theor Appl Genet 1998 97 1370ndash1379 [CrossRef]
57 Mackill DJ Khush GS IR64 A high-quality and high-yielding mega variety Rice 2018 11 18 [CrossRef] [PubMed]58 Heinrichs EA Basilio RP Valencia SL Buprofezin a selective insecticide for the management of rice planthoppers (Ho-
moptera Delphacidae) and leafhoppers (Homoptera Cicadellidae) Environ Entomol 1984 13 515ndash521 [CrossRef]59 Satapathy MK Anjaneyulu A Use of cypermethrin a synthetic pyrethroid in the control of rice tungro virus disease and its
vector Trop Pest Manag 1984 30 170ndash178 [CrossRef]60 Vorley W Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown
planthopper (BPH) in Kedah Malaysia Int Rice Res Newsl 1987 10 19ndash2061 Uddin A Islam KS Jahan M Ara A Khan MAI Factor influencing the resurgence of brown planthopper in Bangladesh
SAARC J Agric 2020 18 117ndash128 [CrossRef]62 Escalada MM Luecha M Heong KL Social impacts of planthopper outbreaks Case study from Thailand In Rice Planthoppers
Ecology Management Socio Economics and Policy Heong KL Cheng J Escalada MM Eds Zhejiang University PressHangzhou China 2015 pp 209ndash226
63 Zhang CP He HM Yu JZ Hu XQ Zhu YH Wang Q Residues of carbosulfan and its metabolites carbofuran and3-hydroxy carbofuran in rice field ecosystem in China J Environ Sci Health 2016 51 351ndash357 [CrossRef] [PubMed]
64 Samanta S Barman M Samanta NRA Bio-efficacy trials of carbofuran 3 CG against insect pests of rice J Entomol ZoolStud 2020 8 1529ndash1533
65 Donley N The USA lags behind other agricultural nations in banning harmful pesticides Environ Health 2019 18 44 [CrossRef][PubMed]
66 Adams A Gore J Musser F Cook D Catchot A Walker T Dobbins C Efficacy of selected insecticides applied to hybridrice seed J Econ Entomol 2015 109 200ndash206 [CrossRef]
67 Kumar R Singh B Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) Field Bull Environ Contam Toxicol2013 90 482ndash488 [CrossRef]
68 Dhaka S Prajapati C Singh D Singh R Field evaluation of insecticides and bio-pesticides against rice leaf folder CnaphalocrocisMedinalis (Guenee) Ann Plant Prot Sci 2011 19 324ndash326
69 Matsumura M Takeuchi H Satoh M Sanada-Morimura S Otuka A Watanabe T Van Thanh D Species-specific insecticideresistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in East and South-eastAsia Pest Manag Sci 2008 64 1115ndash1121 [CrossRef] [PubMed]
70 Tang J Li J Shao Y Yang B Liu Z Fipronil resistance in the whitebacked planthopper (Sogatella furcifera) Possible resistancemechanisms and cross-resistance Pest Manag Sci 2010 66 121ndash125 [CrossRef]
71 Horgan FG Arida A Ardestani G Almazan MLP Temperature-dependent oviposition and nymph performance revealdistinct thermal niches of coexisting planthoppers with similar thresholds for development PLoS ONE 2020 15 e0235506[CrossRef] [PubMed]
Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]

Crops 2021 1 179
have been related to observed preferences in BPH and other sap-sucking insects [349]could further accelerate the build-up of BPH populations on cartap hydrochloride-treatedplants To further test this idea experiments could be conducted using near-isogenic ricelines that differ mainly in the presence or absence of tagged resistance genes [76] Suchtests would avoid confounding differences in plant anatomy between test varieties IR62resistance to BPH has been well studied [4] The variety is also resistant to WBPH atolder plant stages [54] Therefore in our study deltamethrin was also antagonistic tothis WBPH-resistance (Table 2) However BPH feeding also facilitates WBPH populationgrowth [5477] and there may have been added indirect plant-mediated effects [77] thataugmented WBPH populations on both varieties in our experiment IR62 is also resistant toGLH [45] In our experiments GLH densities also increased on deltamethrin-treated plantsbut the effect was only apparent on IR64 Therefore GLH-resistance in IR62 apparentlybuffered against deltamethrin-associated increases in GLH on the variety as was alsonoted from field studies in Vietnam [31] Deltamethrin is currently not recommended foruse in rice [202878] however the product continues to be marketed in rice-dominatedregions throughout Asia [131479] Although cypermethrin has been associated with BPHoutbreaks and was consequently removed for use on rice in Thailand and Vietnam [62]we did not observe cypermethrin-induced resurgence in our screenhouse experimentNevertheless cypermethrin has been associated with resurgence of BPH in South Asia [61]and we have noted stimulatory effects of cypermethrin on other rice herbivores in thePhilippines (Horgan unpublished data)
43 Phytotoxicity
Phytotoxicity is relatively common following applications of insecticides [182850]Phytotoxic effects may change a plantrsquos physiology without consequences for crop yieldsHowever such changes to plant physiology could have indirect effects on herbivores Forexample deltamethrin methyl parathion and quinalphos produce increases in total sugarsand reducing sugars as well as increasing the protein contents of the rice leaf sheath andleaf blade [28] Several insecticides also reduce plant phenols in ricemdashwhich increases therice plantrsquos susceptibility to BPH and other insects [27] In our screenhouse experiment wenoted several changes to rice anatomy and yields Importantly we noted that cypermethrinand fipronil both reduced rice yields (Table 3) This was not due to indirect herbivoreeffects because planthopper and leafhopper densities were consistently low on rice treatedwith these insecticides The effects therefore probably resulted from direct phytotoxiceffects on the plants Furthermore these effects were most pronounced on treated IR62plants Other minor effects on grain production included increased proportions of unfilledgrain in cartap hydrochloride-treated plants and delays in grain maturation on cartaphydrochloride-treated and cypermethrin-treated plants (Table 3) Despite the high densitiesof BPH and WBPH on deltamethrin-treated plants yields were not significantly lower thanin control non-treated plants Both IR62 and IR64 have been noted for their high toleranceto BPH damage particularly under high nitrogenous fertilizer applications [3456] as usedin our screenhouse experiment Therefore where plants have high tolerance to damageapplying insecticides could also result in profitability losses on rice farms and in the caseof insecticide-induced pest population increases tolerance might buffer against subsequentyield losses There are few reports of the phytotoxicity of insecticides to rice carbofuran istoxic to rice at high concentrations but may also enhance the growth of rice plants throughinteractions with nitrogen-fixing bacteria in the rhizosphere [80] Future studies couldmore carefully assess field-grown plants for possible phytotoxic effects particularly ifinsecticides have the potential to directly reduce yields
44 Management implications
Global trends suggest that resistance in rice has failed to reduce insecticide appli-cations [2] This may be due to aggressive marketing by agrochemical companies andstrategies to increase pesticide sales such as promoting prophylactic applications [16] Be-
Crops 2021 1 180
cause we selected our test insecticides (except the pyrethroids) without any a priori criteriaour results suggest that many currently used insecticides probably have some negativeeffects on yield either directly through phytotoxic effects or indirectly by inducing pestresurgence Furthermore our results indicate that even where pests reach high densitiesas on deltamethrin-treated plants in the screenhouse these densities do not necessarilylead to significant yield reductions
Among the products we used buprofezin although it had little effect on oviposi-tion was effective in reducing nymph survival Furthermore there were only minorapparent phytotoxic effects on the rice plants Buprofezin has a low toxicity to naturalenemies in rice fields [58] and is therefore the most compatible of the insecticides withstrategies to increase rice field biodiversity and restore ecosystem services [32ndash348182]Thiamethoxam-chlorantraniliprole was also effective in reducing both oviposition andnymph survival and had no apparent toxic effects on rice plants However thiamethoxam-chlorantraniliprole and other neonicotinoids although moderately toxic can have severeeffects on beneficial hymenopteran parasitoids [83] although they have apparently lowtoxicity to predatory mirid bugs [84] The remaining insecticides either directly reducedyields or were associated with higher BPH biomass or densities in our experiments
We noted few synergies with HPR although nymph biomass andor survival waslower in cartap hydrochloride and carbofuran-treated IR62 plants compared to correspond-ing IR64 plants Meanwhile cartap hydrochloride and deltamethrin were antagonisticto resistance in IR62 in the screenhouse experiment Among the strongest effects of HPRwas to act as a buffer against GLH population build-up on deltamethrin-treated IR62 andagainst carbofuran-induced increases in BPH nymph biomass in the survival bioassayTherefore HPR may play a role in reducing the possibilities of secondary outbreaks ofBPH and other insects The combined use of HPR and targeted insecticides against BPHcreates redundancy in the rice field and counters measures to manage insecticide resistanceand virulence by unnecessarily exposing plant protection products to evolving pest pop-ulations HPR (and host plant tolerance) can be effective against target pests as revealedin previous studies [4] and could be incorporated into other pest management practicesparticularly agroecological practices such as ecological engineering with which it is highlycompatible [818586]
5 Conclusions
Based on our results and the results of previous studies HPR and the applicationof chemical insecticides against BPH rarely produce synergistic effects Antagonisticeffects where chemicals caused increases in BPH fitness or populations on resistant ricewere more apparent in our study In particular deltamethrin caused an outbreak ofBPH in our screenhouse study Nevertheless HPR can buffer against the insecticide-induced enhancement of herbivore fitness (hormesis) and potentially reduce possibilities ofsecondary outbreaks as was suggested from results with cartap hydrochloride in our studyBased on our results we suggest that carbofuran cartap hydrochloride cypermethrindeltamethrin and fipronil should be avoided in rice particularly in the Philippines (wherethis research was conducted) due to their resurgence-causing potential or because of directreductions to rice yields
Author Contributions Conceptualization FGH methodology FGH AP-C and MLPA formalanalysis FGH and AP-C investigation FGH AP-C and MLPA data curation FGHAP-C and MLPA writingmdashoriginal draft preparation FGH and AP-C writingmdashreview andediting FGH AP-C and MLPA visualization FGH supervision FGH and MLPA projectadministration FGH funding acquisition FGH All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by the Bill and Melinda Gates Foundation (Cereal SystemsInitiative for South Asia [CSISA] OPP52303) and the Global Rice Science Platform (GRiSP) underthe directorship of Achim Dobermann AP-C was partially funded by The Spanish Governmentthrough Becas Faro International
Crops 2021 1 181
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data presented in this study are available on reasonable requestfrom the corresponding author
Acknowledgments The authors thank Fame Ramal Carmencita Bernal Alberto Naredo ReyeulQuintana Vincent Virtudes Marol Recide Jenyrose Geronda and Ellen Genil for assistance duringthis study The contributions of Rodante Abas (sadly deceased) during three decades of entomologicalresearch at IRRI are gratefully acknowledged Rice lines were supplied through the Genebank of IRRI
Conflicts of Interest The authors declare no conflict of interest
References1 Bottrell DG Schoenly KG Resurrecting the ghost of green revolutions past The brown planthopper as a recurring threat to
high-yielding rice production in tropical Asia J Asia-Pac Entomol 2012 15 122ndash140 [CrossRef]2 Horgan FG Integrated pest management for sustainable rice cultivation A holistic approach In Achieving Sustainable Cultivation
of Rice Cultivation Pest and Disease Management Sasaki T Ed Burleigh Dodds Scientific Cambridge UK 2017 pp 309ndash3423 Horgan FG de Freitas TFS Crisol-Martiacutenez E Mundaca EA Bernal CC Nitrogenous fertilizer reduces resistance but
enhances tolerance to the brown planthopper in fast-growing moderately resistant rice Insects 2021 12 989 [CrossRef] [PubMed]4 Horgan FG Pentildealver Cruz A Bernal CC Ramal AF Almazan MLP Wilby A Resistance and tolerance to the brown
planthopper Nilaparvata lugens (Staringl) in rice infested at different growth stages across a gradient of nitrogen applicationsField Crops Res 2018 217 53ndash65 [CrossRef]
5 Hereward JP Cai X Matias AMA Walter GH Xu C Wang Y Migration dynamics of an important rice pest The brownplanthopper (Nilaparvata lugens) across AsiamdashInsights from population genomics Evol Appl 2020 13 2449ndash2459 [CrossRef]
6 Hu C Guo J Fu X Huang Y Gao X Wu K Seasonal migration pattern of Nilaparvata lugens (Hemiptera Delphacidae) overthe Bohai Sea in northern China J Econ Entomol 2018 111 2129ndash2135 [CrossRef]
7 Ma J Wang Y-C Hu Y-Y Lu M-H Wan G-J Chen F-J Liu W-C Zhai B-P Hu G Brown planthopper Nilaparvatalugens was concentrated at the rear of the typhoon Soudelor in eastern China in August 2015 Insect Sci 2018 25 916ndash926[CrossRef]
8 Fujita D Kohli A Horgan FG Rice resistance to planthoppers and leafhoppers Crit Rev Plant Sci 2013 32 162ndash191[CrossRef]
9 Fujii T Yoshida K Kobayashi T Myint KKM Yasui H Sanada-Morimura S Matsumura M Long-term virulencemonitoring of differential cultivars in Japanrsquos immigrant populations of Nilaparvata lugens (Hemiptera Delphacidae) in 2001ndash2009Appl Entomol Zool 2021 56 407ndash418 [CrossRef]
10 Myint KKM Yasui H Takagi M Matsumura M Virulence of long-term laboratory populations of the brown planthopperNilaparvata lugens (Stal) and whitebacked planthopper Sogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) on rice differentialvarieties Appl Entomol Zool 2009 44 149ndash153 [CrossRef]
11 Horgan FG Ramal AF Bentur JS Kumar R Bhanu KV Sarao PS Iswanto EH Van Chien H Phyu MHBernal CC et al Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia againstresistant rice varieties Crop Prot 2015 78 222ndash231 [CrossRef]
12 Horgan FG Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers Crop Prot2018 110 21ndash33 [CrossRef]
13 Uddin ABMA Islam KS Jahan M Ara A Afrin S Farmerrsquos perception about resurgence of brown planthopper NilaparvataLugens (Staringl) in Bangladesh Bangladesh Rice J 2020 23 21ndash33 [CrossRef]
14 Triwidodo H Brown planthoppers infestations and insecticides use pattern in Java Indonesia AGRIVITA J Agric Sci 202042 320ndash330 [CrossRef]
15 Veres A Wyckhuys KA Kiss J Toacuteth F Burgio G Pons X Avilla C Vidal S Razinger J Bazok R et al An update of theWorldwide Integrated Assessment (WIA) on systemic pesticides Part 4 Alternatives in major cropping systems Environ SciPollut Res 2020 27 29867ndash29899 [CrossRef]
16 Bakker L van der Werf W Tittonell P Wyckhuys KA Bianchi FJ Neonicotinoids in global agriculture Evidence for a newpesticide treadmill Ecol Soc 2020 25 26 [CrossRef]
17 Wu J-C Xu J-X Yuan S-Z Liu J-L Jiang Y-H Xu J-F Pesticide-induced susceptibility of rice to brown planthopperNilaparvata lugens Entomol Exp Appl 2001 100 119ndash126 [CrossRef]
18 Wu J-C Qiu H-M Yang G-Q Liu J-L Liu G-J Wilkens RM Effective duration of pesticide-induced susceptibility of riceto brown planthopper (Nilaparvata lugens Staringl Homoptera Delphacidae) and physiological and biochemical changes in riceplants following pesticide application Int J Pest Manag 2004 50 55ndash62 [CrossRef]
19 Wang A-H Wu J-C Yu Y-S Liu J-L Yue J-F Wang MY Selective insecticide-induced stimulation on fecundity andbiochemical changes in Tryporyza incertulas (Lepidoptera Pyralidae) J Econ Entomol 2005 98 1144ndash1149 [CrossRef] [PubMed]
Crops 2021 1 182
20 Azzam S Wang F Wu J-C Shen J Wang L-P Yang G-Q Guo Y-R Comparisons of stimulatory effects of a seriesof concentrations of four insecticides on reproduction in the rice brown planthopper Nilaparvata lugens Staringl (HomopteraDelphacidae) Int J Pest Manag 2009 55 347ndash358 [CrossRef]
21 Ge L-Q Hu J-H Wu J-C Yang G-Q Gu H Insecticide-induced changes in protein RNA and DNA contents in ovary andfat body of female Nilaparvata lugens (Hemiptera Delphacidae) J Econ Entomol 2009 102 1506ndash1514 [CrossRef]
22 Ling S Zhang J Hu L Zhang R Effect of fipronil on the reproduction feeding and relative fitness of brown planthopperNilaparvata lugens Appl Entomol Zool 2009 44 543ndash548 [CrossRef]
23 Ge L-Q Wang L-P Zhao K-F Wu J-C Huang L-J Mating pair combinations of insecticide-treated male and femaleNilaparvata lugens Staringl (Hemiptera Delphacidae) planthoppers influence protein content in the male accessory glands (MAGs)and vitellin content in both fat bodies and ovaries of adult females Pestic Biochem Physiol 2010 98 279ndash288 [CrossRef]
24 Ge L-Q Wu J-C Zhao K-F Chen Y Yang G-Q Induction of Nlvg and suppression of Nljhe gene expression in Nilaparvatalugens (Staringl) (Hemiptera Delphacidae) adult females and males exposed to two insecticides Pestic Biochem Physiol 201098 269ndash278 [CrossRef]
25 Hu J-H Wu J-C Yin J-L Gu H-N Physiology of insecticide-induced stimulation of reproduction in the rice brownplanthopper (Nilaparvata lugens (Staringl)) Dynamics of protein in fat body and ovary Int J Pest Manag 2010 56 23ndash30 [CrossRef]
26 Wang L-P Shen J Ge L-Q Wu J-C Yang G-Q Jahn GC Insecticide-induced increase in the protein content of maleaccessory glands and its effect on the fecundity of females in the brown planthopper Nilaparvata lugens Staringl (HemipteraDelphacidae) Crop Prot 2010 29 1280ndash1285 [CrossRef]
27 Wu J Ge L Liu F Song Q Stanley D Pesticide-induced planthopper population resurgence in rice cropping systems AnnuRev Entomol 2020 65 409ndash429 [CrossRef]
28 Suri KS Singh G Insecticide-induced resurgence of the whitebacked planthopper Sogatella furcifera (Horvath) (HemipteraDelphacidae) on rice varieties with different levels of resistance Crop Prot 2011 30 118ndash124 [CrossRef]
29 Fabellar LT Heinrichs EA Toxicity of insecticides to predators of rice brown planthoppers Nilaparvata lugens (Staringl) (HomopteraDelphacidae) Environ Entomol 1984 13 832ndash837 [CrossRef]
30 Fabellar LT Heinrichs EA Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators Crop Prot1986 5 254ndash258 [CrossRef]
31 Cuong NL Ben PT Phuong LT Chau LM Cohen MB Effect of host plant resistance and insecticide on brown planthopperNilaparvata lugens (Staringl) and predator population development in the Mekong Delta Vietnam Crop Prot 1997 16 707ndash715[CrossRef]
32 Preetha G Stanley J Suresh S Samiyappan R Risk assessment of insecticides used in rice on mirid bug Cyrtorhinuslividipennis Reuter the important predator of brown planthopper Nilaparvata lugens (Stal) Chemosphere 2010 80 498ndash503[CrossRef] [PubMed]
33 Tanaka K Endo S Kazano H Toxicity of insecticides to predators of rice planthoppers Spiders the mirid bug and the dryinidwasp Appl Entomol Zool 2000 35 177ndash187 [CrossRef]
34 Zhang X Xu Q Lu W Liu F Sublethal effects of four synthetic insecticides on the generalist predator Cyrtorhinus lividipennisJ Pest Sci 2015 88 383ndash392 [CrossRef]
35 Settle WH Ariawan H Astuti ET Cahyana W Hakim AL Hindayana D Lestari AS Managing tropical rice peststhrough conservation of generalist natural enemies and alternative prey Ecology 1996 77 1975ndash1988 [CrossRef]
36 Salim M Heinrichs EA Insecticide-induced changes in the levels of resistance of rice cultivars to the whitebacked planthopperSogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) Crop Prot 1987 6 28ndash32 [CrossRef]
37 Yin J-L Xu H-W Wu J-C Hu J-H Yang G-Q Cultivar and insecticide applications affect the physiological development ofthe brown planthopper Nilaparvata lugens (Staringl) (Hemiptera Delphacidae) Environ Entomol 2014 37 206ndash212 [CrossRef]
38 Cheng C Chang W Studies on varietal resistance to the brown planthopper in Taiwan In Brown Planthopper Threat toRice Production in Asia International Rice Research Institute Ed International Rice Research Institute Manila Philippines1979 pp 251ndash271
39 Reissig WH Heinrichs EA Valencia SL Insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on ricevarieties with different levels of resistance Environ Entomol 1982 11 165ndash168 [CrossRef]
40 Heinrichs EA Fabellar LT Basilio RP Wen T-C Medrano F Susceptibility of rice planthoppers Nilaparvata lugens andSogatella furcifera (Homoptera Delphacidae) to insecticides as influenced by level of resistance in the host plant Environ Entomol1984 13 455ndash458 [CrossRef]
41 Joshi R Shepard B Kenmore P Lydia R Insecticide-induced resurgence of brown planthopper (BPH) on IR62 Int Rice ResNewsl 1992 17 9ndash10
42 Zhu Z-R Cheng J Jiang M-X Zhang X-X Complex influence of rice variety fertilization timing and insecticide onpopulation dynamics of Sogatella furcifera (Horvath) Nilaparvata lugens (Staringl) (Homoptera Delphacidae) and their natural enemiesin rice in Hangzhou China J Pest Sci 2004 77 65ndash74 [CrossRef]
43 Nanthakumar M Jhansi Lakshmi V Shashi Bhushan V Balachandran SM Mohan M Decrease of rice plant resistance andinduction of hormesis and carboxylesterase titre in brown planthopper Nilaparvata lugens (Staringl) by xenobiotics Pestic BiochemPhysiol 2012 102 146ndash152 [CrossRef]
Crops 2021 1 183
44 Yao H-W Jiang C-Y Ye G-Y Cheng J-A Studies on cultivar-induced changes in insecticide susceptibility and its relatedenzyme activities of the white-backed planthopper Sogatella furcifera (Horvath)(Homopter Delphacidae) Insect Sci 2002 9 9ndash15
45 Horgan FG Srinivasan TS Bentur JS Kumar R Bhanu KV Sarao PS Van Chien H Almazan MLP Bernal CCRamal AF et al Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers Implicationsfor rice breeding and gene deployment Agronomy 2017 7 62 [CrossRef]
46 Jairin J Phengrat K Teangdeerith S Vanavichit A Toojinda T Mapping of a broad-spectrum brown planthopper resistancegene Bph3 on rice chromosome 6 Mol Breed 2007 19 35ndash44 [CrossRef]
47 Ren J Gao F Wu X Lu X Zeng L Lv J Su X Luo H Ren G Bph32 a novel gene encoding an unknown SCRdomain-containing protein confers resistance against the brown planthopper in rice Sci Rep 2016 6 37645 [CrossRef]
48 Nguyen CD Zheng S-H Sanada-Morimura S Matsumura M Yasui H Fujita D Substitution mapping and characterizationof brown planthopper resistance genes from indica rice variety lsquoPTB33prime (Oryza sativa L) Breed Sci 2021 in press [CrossRef]
49 Chelliah S Heinrichs EA Factors affecting insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on RiceEnviron Entomol 1980 9 773ndash777 [CrossRef]
50 Moore MT Kroumlger R Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growthArch Environ Contam Toxicol 2010 59 574ndash581 [CrossRef]
51 Kim J Hwang I Han M Jang B Effect of insecticide and fungicide on phytotoxicity of herbicide in rice Korean J Weed Sci1986 6 67ndash75
52 Horgan FG Garcia CPF Haverkort F de Jong PW Ferrater JB Changes in insecticide resistance and host range performanceof planthoppers artificially selected to feed on resistant rice Crop Prot 2020 127 104963 [CrossRef] [PubMed]
53 Khush GS Virk PS IR Varieties and Their Impact International Rice Research Institute Manila Philippines 200554 Srinivasan TS Almazan MLP Bernal CC Ramal AF Subbarayalu MK Horgan FG Interactions between nymphs of
Nilaparvata lugens and Sogatella furcifera (Hemiptera Delphacidae) on resistant and susceptible rice varieties Appl Entomol Zool2016 51 81ndash90 [CrossRef]
55 Pentildealver Cruz A Arida A Heong KL Horgan FG Aspects of brown planthopper adaptation to resistant rice varieties withthe Bph3 gene Entomol Exp Appl 2011 141 245ndash257 [CrossRef]
56 Alam SN Cohen MB Detection and analysis of QTLs for resistance to the brown planthopper Nilaparvata lugens in adoubled-haploid rice population Theor Appl Genet 1998 97 1370ndash1379 [CrossRef]
57 Mackill DJ Khush GS IR64 A high-quality and high-yielding mega variety Rice 2018 11 18 [CrossRef] [PubMed]58 Heinrichs EA Basilio RP Valencia SL Buprofezin a selective insecticide for the management of rice planthoppers (Ho-
moptera Delphacidae) and leafhoppers (Homoptera Cicadellidae) Environ Entomol 1984 13 515ndash521 [CrossRef]59 Satapathy MK Anjaneyulu A Use of cypermethrin a synthetic pyrethroid in the control of rice tungro virus disease and its
vector Trop Pest Manag 1984 30 170ndash178 [CrossRef]60 Vorley W Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown
planthopper (BPH) in Kedah Malaysia Int Rice Res Newsl 1987 10 19ndash2061 Uddin A Islam KS Jahan M Ara A Khan MAI Factor influencing the resurgence of brown planthopper in Bangladesh
SAARC J Agric 2020 18 117ndash128 [CrossRef]62 Escalada MM Luecha M Heong KL Social impacts of planthopper outbreaks Case study from Thailand In Rice Planthoppers
Ecology Management Socio Economics and Policy Heong KL Cheng J Escalada MM Eds Zhejiang University PressHangzhou China 2015 pp 209ndash226
63 Zhang CP He HM Yu JZ Hu XQ Zhu YH Wang Q Residues of carbosulfan and its metabolites carbofuran and3-hydroxy carbofuran in rice field ecosystem in China J Environ Sci Health 2016 51 351ndash357 [CrossRef] [PubMed]
64 Samanta S Barman M Samanta NRA Bio-efficacy trials of carbofuran 3 CG against insect pests of rice J Entomol ZoolStud 2020 8 1529ndash1533
65 Donley N The USA lags behind other agricultural nations in banning harmful pesticides Environ Health 2019 18 44 [CrossRef][PubMed]
66 Adams A Gore J Musser F Cook D Catchot A Walker T Dobbins C Efficacy of selected insecticides applied to hybridrice seed J Econ Entomol 2015 109 200ndash206 [CrossRef]
67 Kumar R Singh B Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) Field Bull Environ Contam Toxicol2013 90 482ndash488 [CrossRef]
68 Dhaka S Prajapati C Singh D Singh R Field evaluation of insecticides and bio-pesticides against rice leaf folder CnaphalocrocisMedinalis (Guenee) Ann Plant Prot Sci 2011 19 324ndash326
69 Matsumura M Takeuchi H Satoh M Sanada-Morimura S Otuka A Watanabe T Van Thanh D Species-specific insecticideresistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in East and South-eastAsia Pest Manag Sci 2008 64 1115ndash1121 [CrossRef] [PubMed]
70 Tang J Li J Shao Y Yang B Liu Z Fipronil resistance in the whitebacked planthopper (Sogatella furcifera) Possible resistancemechanisms and cross-resistance Pest Manag Sci 2010 66 121ndash125 [CrossRef]
71 Horgan FG Arida A Ardestani G Almazan MLP Temperature-dependent oviposition and nymph performance revealdistinct thermal niches of coexisting planthoppers with similar thresholds for development PLoS ONE 2020 15 e0235506[CrossRef] [PubMed]
Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]

Crops 2021 1 180
cause we selected our test insecticides (except the pyrethroids) without any a priori criteriaour results suggest that many currently used insecticides probably have some negativeeffects on yield either directly through phytotoxic effects or indirectly by inducing pestresurgence Furthermore our results indicate that even where pests reach high densitiesas on deltamethrin-treated plants in the screenhouse these densities do not necessarilylead to significant yield reductions
Among the products we used buprofezin although it had little effect on oviposi-tion was effective in reducing nymph survival Furthermore there were only minorapparent phytotoxic effects on the rice plants Buprofezin has a low toxicity to naturalenemies in rice fields [58] and is therefore the most compatible of the insecticides withstrategies to increase rice field biodiversity and restore ecosystem services [32ndash348182]Thiamethoxam-chlorantraniliprole was also effective in reducing both oviposition andnymph survival and had no apparent toxic effects on rice plants However thiamethoxam-chlorantraniliprole and other neonicotinoids although moderately toxic can have severeeffects on beneficial hymenopteran parasitoids [83] although they have apparently lowtoxicity to predatory mirid bugs [84] The remaining insecticides either directly reducedyields or were associated with higher BPH biomass or densities in our experiments
We noted few synergies with HPR although nymph biomass andor survival waslower in cartap hydrochloride and carbofuran-treated IR62 plants compared to correspond-ing IR64 plants Meanwhile cartap hydrochloride and deltamethrin were antagonisticto resistance in IR62 in the screenhouse experiment Among the strongest effects of HPRwas to act as a buffer against GLH population build-up on deltamethrin-treated IR62 andagainst carbofuran-induced increases in BPH nymph biomass in the survival bioassayTherefore HPR may play a role in reducing the possibilities of secondary outbreaks ofBPH and other insects The combined use of HPR and targeted insecticides against BPHcreates redundancy in the rice field and counters measures to manage insecticide resistanceand virulence by unnecessarily exposing plant protection products to evolving pest pop-ulations HPR (and host plant tolerance) can be effective against target pests as revealedin previous studies [4] and could be incorporated into other pest management practicesparticularly agroecological practices such as ecological engineering with which it is highlycompatible [818586]
5 Conclusions
Based on our results and the results of previous studies HPR and the applicationof chemical insecticides against BPH rarely produce synergistic effects Antagonisticeffects where chemicals caused increases in BPH fitness or populations on resistant ricewere more apparent in our study In particular deltamethrin caused an outbreak ofBPH in our screenhouse study Nevertheless HPR can buffer against the insecticide-induced enhancement of herbivore fitness (hormesis) and potentially reduce possibilities ofsecondary outbreaks as was suggested from results with cartap hydrochloride in our studyBased on our results we suggest that carbofuran cartap hydrochloride cypermethrindeltamethrin and fipronil should be avoided in rice particularly in the Philippines (wherethis research was conducted) due to their resurgence-causing potential or because of directreductions to rice yields
Author Contributions Conceptualization FGH methodology FGH AP-C and MLPA formalanalysis FGH and AP-C investigation FGH AP-C and MLPA data curation FGHAP-C and MLPA writingmdashoriginal draft preparation FGH and AP-C writingmdashreview andediting FGH AP-C and MLPA visualization FGH supervision FGH and MLPA projectadministration FGH funding acquisition FGH All authors have read and agreed to the publishedversion of the manuscript
Funding This research was funded by the Bill and Melinda Gates Foundation (Cereal SystemsInitiative for South Asia [CSISA] OPP52303) and the Global Rice Science Platform (GRiSP) underthe directorship of Achim Dobermann AP-C was partially funded by The Spanish Governmentthrough Becas Faro International
Crops 2021 1 181
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data presented in this study are available on reasonable requestfrom the corresponding author
Acknowledgments The authors thank Fame Ramal Carmencita Bernal Alberto Naredo ReyeulQuintana Vincent Virtudes Marol Recide Jenyrose Geronda and Ellen Genil for assistance duringthis study The contributions of Rodante Abas (sadly deceased) during three decades of entomologicalresearch at IRRI are gratefully acknowledged Rice lines were supplied through the Genebank of IRRI
Conflicts of Interest The authors declare no conflict of interest
References1 Bottrell DG Schoenly KG Resurrecting the ghost of green revolutions past The brown planthopper as a recurring threat to
high-yielding rice production in tropical Asia J Asia-Pac Entomol 2012 15 122ndash140 [CrossRef]2 Horgan FG Integrated pest management for sustainable rice cultivation A holistic approach In Achieving Sustainable Cultivation
of Rice Cultivation Pest and Disease Management Sasaki T Ed Burleigh Dodds Scientific Cambridge UK 2017 pp 309ndash3423 Horgan FG de Freitas TFS Crisol-Martiacutenez E Mundaca EA Bernal CC Nitrogenous fertilizer reduces resistance but
enhances tolerance to the brown planthopper in fast-growing moderately resistant rice Insects 2021 12 989 [CrossRef] [PubMed]4 Horgan FG Pentildealver Cruz A Bernal CC Ramal AF Almazan MLP Wilby A Resistance and tolerance to the brown
planthopper Nilaparvata lugens (Staringl) in rice infested at different growth stages across a gradient of nitrogen applicationsField Crops Res 2018 217 53ndash65 [CrossRef]
5 Hereward JP Cai X Matias AMA Walter GH Xu C Wang Y Migration dynamics of an important rice pest The brownplanthopper (Nilaparvata lugens) across AsiamdashInsights from population genomics Evol Appl 2020 13 2449ndash2459 [CrossRef]
6 Hu C Guo J Fu X Huang Y Gao X Wu K Seasonal migration pattern of Nilaparvata lugens (Hemiptera Delphacidae) overthe Bohai Sea in northern China J Econ Entomol 2018 111 2129ndash2135 [CrossRef]
7 Ma J Wang Y-C Hu Y-Y Lu M-H Wan G-J Chen F-J Liu W-C Zhai B-P Hu G Brown planthopper Nilaparvatalugens was concentrated at the rear of the typhoon Soudelor in eastern China in August 2015 Insect Sci 2018 25 916ndash926[CrossRef]
8 Fujita D Kohli A Horgan FG Rice resistance to planthoppers and leafhoppers Crit Rev Plant Sci 2013 32 162ndash191[CrossRef]
9 Fujii T Yoshida K Kobayashi T Myint KKM Yasui H Sanada-Morimura S Matsumura M Long-term virulencemonitoring of differential cultivars in Japanrsquos immigrant populations of Nilaparvata lugens (Hemiptera Delphacidae) in 2001ndash2009Appl Entomol Zool 2021 56 407ndash418 [CrossRef]
10 Myint KKM Yasui H Takagi M Matsumura M Virulence of long-term laboratory populations of the brown planthopperNilaparvata lugens (Stal) and whitebacked planthopper Sogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) on rice differentialvarieties Appl Entomol Zool 2009 44 149ndash153 [CrossRef]
11 Horgan FG Ramal AF Bentur JS Kumar R Bhanu KV Sarao PS Iswanto EH Van Chien H Phyu MHBernal CC et al Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia againstresistant rice varieties Crop Prot 2015 78 222ndash231 [CrossRef]
12 Horgan FG Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers Crop Prot2018 110 21ndash33 [CrossRef]
13 Uddin ABMA Islam KS Jahan M Ara A Afrin S Farmerrsquos perception about resurgence of brown planthopper NilaparvataLugens (Staringl) in Bangladesh Bangladesh Rice J 2020 23 21ndash33 [CrossRef]
14 Triwidodo H Brown planthoppers infestations and insecticides use pattern in Java Indonesia AGRIVITA J Agric Sci 202042 320ndash330 [CrossRef]
15 Veres A Wyckhuys KA Kiss J Toacuteth F Burgio G Pons X Avilla C Vidal S Razinger J Bazok R et al An update of theWorldwide Integrated Assessment (WIA) on systemic pesticides Part 4 Alternatives in major cropping systems Environ SciPollut Res 2020 27 29867ndash29899 [CrossRef]
16 Bakker L van der Werf W Tittonell P Wyckhuys KA Bianchi FJ Neonicotinoids in global agriculture Evidence for a newpesticide treadmill Ecol Soc 2020 25 26 [CrossRef]
17 Wu J-C Xu J-X Yuan S-Z Liu J-L Jiang Y-H Xu J-F Pesticide-induced susceptibility of rice to brown planthopperNilaparvata lugens Entomol Exp Appl 2001 100 119ndash126 [CrossRef]
18 Wu J-C Qiu H-M Yang G-Q Liu J-L Liu G-J Wilkens RM Effective duration of pesticide-induced susceptibility of riceto brown planthopper (Nilaparvata lugens Staringl Homoptera Delphacidae) and physiological and biochemical changes in riceplants following pesticide application Int J Pest Manag 2004 50 55ndash62 [CrossRef]
19 Wang A-H Wu J-C Yu Y-S Liu J-L Yue J-F Wang MY Selective insecticide-induced stimulation on fecundity andbiochemical changes in Tryporyza incertulas (Lepidoptera Pyralidae) J Econ Entomol 2005 98 1144ndash1149 [CrossRef] [PubMed]
Crops 2021 1 182
20 Azzam S Wang F Wu J-C Shen J Wang L-P Yang G-Q Guo Y-R Comparisons of stimulatory effects of a seriesof concentrations of four insecticides on reproduction in the rice brown planthopper Nilaparvata lugens Staringl (HomopteraDelphacidae) Int J Pest Manag 2009 55 347ndash358 [CrossRef]
21 Ge L-Q Hu J-H Wu J-C Yang G-Q Gu H Insecticide-induced changes in protein RNA and DNA contents in ovary andfat body of female Nilaparvata lugens (Hemiptera Delphacidae) J Econ Entomol 2009 102 1506ndash1514 [CrossRef]
22 Ling S Zhang J Hu L Zhang R Effect of fipronil on the reproduction feeding and relative fitness of brown planthopperNilaparvata lugens Appl Entomol Zool 2009 44 543ndash548 [CrossRef]
23 Ge L-Q Wang L-P Zhao K-F Wu J-C Huang L-J Mating pair combinations of insecticide-treated male and femaleNilaparvata lugens Staringl (Hemiptera Delphacidae) planthoppers influence protein content in the male accessory glands (MAGs)and vitellin content in both fat bodies and ovaries of adult females Pestic Biochem Physiol 2010 98 279ndash288 [CrossRef]
24 Ge L-Q Wu J-C Zhao K-F Chen Y Yang G-Q Induction of Nlvg and suppression of Nljhe gene expression in Nilaparvatalugens (Staringl) (Hemiptera Delphacidae) adult females and males exposed to two insecticides Pestic Biochem Physiol 201098 269ndash278 [CrossRef]
25 Hu J-H Wu J-C Yin J-L Gu H-N Physiology of insecticide-induced stimulation of reproduction in the rice brownplanthopper (Nilaparvata lugens (Staringl)) Dynamics of protein in fat body and ovary Int J Pest Manag 2010 56 23ndash30 [CrossRef]
26 Wang L-P Shen J Ge L-Q Wu J-C Yang G-Q Jahn GC Insecticide-induced increase in the protein content of maleaccessory glands and its effect on the fecundity of females in the brown planthopper Nilaparvata lugens Staringl (HemipteraDelphacidae) Crop Prot 2010 29 1280ndash1285 [CrossRef]
27 Wu J Ge L Liu F Song Q Stanley D Pesticide-induced planthopper population resurgence in rice cropping systems AnnuRev Entomol 2020 65 409ndash429 [CrossRef]
28 Suri KS Singh G Insecticide-induced resurgence of the whitebacked planthopper Sogatella furcifera (Horvath) (HemipteraDelphacidae) on rice varieties with different levels of resistance Crop Prot 2011 30 118ndash124 [CrossRef]
29 Fabellar LT Heinrichs EA Toxicity of insecticides to predators of rice brown planthoppers Nilaparvata lugens (Staringl) (HomopteraDelphacidae) Environ Entomol 1984 13 832ndash837 [CrossRef]
30 Fabellar LT Heinrichs EA Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators Crop Prot1986 5 254ndash258 [CrossRef]
31 Cuong NL Ben PT Phuong LT Chau LM Cohen MB Effect of host plant resistance and insecticide on brown planthopperNilaparvata lugens (Staringl) and predator population development in the Mekong Delta Vietnam Crop Prot 1997 16 707ndash715[CrossRef]
32 Preetha G Stanley J Suresh S Samiyappan R Risk assessment of insecticides used in rice on mirid bug Cyrtorhinuslividipennis Reuter the important predator of brown planthopper Nilaparvata lugens (Stal) Chemosphere 2010 80 498ndash503[CrossRef] [PubMed]
33 Tanaka K Endo S Kazano H Toxicity of insecticides to predators of rice planthoppers Spiders the mirid bug and the dryinidwasp Appl Entomol Zool 2000 35 177ndash187 [CrossRef]
34 Zhang X Xu Q Lu W Liu F Sublethal effects of four synthetic insecticides on the generalist predator Cyrtorhinus lividipennisJ Pest Sci 2015 88 383ndash392 [CrossRef]
35 Settle WH Ariawan H Astuti ET Cahyana W Hakim AL Hindayana D Lestari AS Managing tropical rice peststhrough conservation of generalist natural enemies and alternative prey Ecology 1996 77 1975ndash1988 [CrossRef]
36 Salim M Heinrichs EA Insecticide-induced changes in the levels of resistance of rice cultivars to the whitebacked planthopperSogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) Crop Prot 1987 6 28ndash32 [CrossRef]
37 Yin J-L Xu H-W Wu J-C Hu J-H Yang G-Q Cultivar and insecticide applications affect the physiological development ofthe brown planthopper Nilaparvata lugens (Staringl) (Hemiptera Delphacidae) Environ Entomol 2014 37 206ndash212 [CrossRef]
38 Cheng C Chang W Studies on varietal resistance to the brown planthopper in Taiwan In Brown Planthopper Threat toRice Production in Asia International Rice Research Institute Ed International Rice Research Institute Manila Philippines1979 pp 251ndash271
39 Reissig WH Heinrichs EA Valencia SL Insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on ricevarieties with different levels of resistance Environ Entomol 1982 11 165ndash168 [CrossRef]
40 Heinrichs EA Fabellar LT Basilio RP Wen T-C Medrano F Susceptibility of rice planthoppers Nilaparvata lugens andSogatella furcifera (Homoptera Delphacidae) to insecticides as influenced by level of resistance in the host plant Environ Entomol1984 13 455ndash458 [CrossRef]
41 Joshi R Shepard B Kenmore P Lydia R Insecticide-induced resurgence of brown planthopper (BPH) on IR62 Int Rice ResNewsl 1992 17 9ndash10
42 Zhu Z-R Cheng J Jiang M-X Zhang X-X Complex influence of rice variety fertilization timing and insecticide onpopulation dynamics of Sogatella furcifera (Horvath) Nilaparvata lugens (Staringl) (Homoptera Delphacidae) and their natural enemiesin rice in Hangzhou China J Pest Sci 2004 77 65ndash74 [CrossRef]
43 Nanthakumar M Jhansi Lakshmi V Shashi Bhushan V Balachandran SM Mohan M Decrease of rice plant resistance andinduction of hormesis and carboxylesterase titre in brown planthopper Nilaparvata lugens (Staringl) by xenobiotics Pestic BiochemPhysiol 2012 102 146ndash152 [CrossRef]
Crops 2021 1 183
44 Yao H-W Jiang C-Y Ye G-Y Cheng J-A Studies on cultivar-induced changes in insecticide susceptibility and its relatedenzyme activities of the white-backed planthopper Sogatella furcifera (Horvath)(Homopter Delphacidae) Insect Sci 2002 9 9ndash15
45 Horgan FG Srinivasan TS Bentur JS Kumar R Bhanu KV Sarao PS Van Chien H Almazan MLP Bernal CCRamal AF et al Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers Implicationsfor rice breeding and gene deployment Agronomy 2017 7 62 [CrossRef]
46 Jairin J Phengrat K Teangdeerith S Vanavichit A Toojinda T Mapping of a broad-spectrum brown planthopper resistancegene Bph3 on rice chromosome 6 Mol Breed 2007 19 35ndash44 [CrossRef]
47 Ren J Gao F Wu X Lu X Zeng L Lv J Su X Luo H Ren G Bph32 a novel gene encoding an unknown SCRdomain-containing protein confers resistance against the brown planthopper in rice Sci Rep 2016 6 37645 [CrossRef]
48 Nguyen CD Zheng S-H Sanada-Morimura S Matsumura M Yasui H Fujita D Substitution mapping and characterizationof brown planthopper resistance genes from indica rice variety lsquoPTB33prime (Oryza sativa L) Breed Sci 2021 in press [CrossRef]
49 Chelliah S Heinrichs EA Factors affecting insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on RiceEnviron Entomol 1980 9 773ndash777 [CrossRef]
50 Moore MT Kroumlger R Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growthArch Environ Contam Toxicol 2010 59 574ndash581 [CrossRef]
51 Kim J Hwang I Han M Jang B Effect of insecticide and fungicide on phytotoxicity of herbicide in rice Korean J Weed Sci1986 6 67ndash75
52 Horgan FG Garcia CPF Haverkort F de Jong PW Ferrater JB Changes in insecticide resistance and host range performanceof planthoppers artificially selected to feed on resistant rice Crop Prot 2020 127 104963 [CrossRef] [PubMed]
53 Khush GS Virk PS IR Varieties and Their Impact International Rice Research Institute Manila Philippines 200554 Srinivasan TS Almazan MLP Bernal CC Ramal AF Subbarayalu MK Horgan FG Interactions between nymphs of
Nilaparvata lugens and Sogatella furcifera (Hemiptera Delphacidae) on resistant and susceptible rice varieties Appl Entomol Zool2016 51 81ndash90 [CrossRef]
55 Pentildealver Cruz A Arida A Heong KL Horgan FG Aspects of brown planthopper adaptation to resistant rice varieties withthe Bph3 gene Entomol Exp Appl 2011 141 245ndash257 [CrossRef]
56 Alam SN Cohen MB Detection and analysis of QTLs for resistance to the brown planthopper Nilaparvata lugens in adoubled-haploid rice population Theor Appl Genet 1998 97 1370ndash1379 [CrossRef]
57 Mackill DJ Khush GS IR64 A high-quality and high-yielding mega variety Rice 2018 11 18 [CrossRef] [PubMed]58 Heinrichs EA Basilio RP Valencia SL Buprofezin a selective insecticide for the management of rice planthoppers (Ho-
moptera Delphacidae) and leafhoppers (Homoptera Cicadellidae) Environ Entomol 1984 13 515ndash521 [CrossRef]59 Satapathy MK Anjaneyulu A Use of cypermethrin a synthetic pyrethroid in the control of rice tungro virus disease and its
vector Trop Pest Manag 1984 30 170ndash178 [CrossRef]60 Vorley W Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown
planthopper (BPH) in Kedah Malaysia Int Rice Res Newsl 1987 10 19ndash2061 Uddin A Islam KS Jahan M Ara A Khan MAI Factor influencing the resurgence of brown planthopper in Bangladesh
SAARC J Agric 2020 18 117ndash128 [CrossRef]62 Escalada MM Luecha M Heong KL Social impacts of planthopper outbreaks Case study from Thailand In Rice Planthoppers
Ecology Management Socio Economics and Policy Heong KL Cheng J Escalada MM Eds Zhejiang University PressHangzhou China 2015 pp 209ndash226
63 Zhang CP He HM Yu JZ Hu XQ Zhu YH Wang Q Residues of carbosulfan and its metabolites carbofuran and3-hydroxy carbofuran in rice field ecosystem in China J Environ Sci Health 2016 51 351ndash357 [CrossRef] [PubMed]
64 Samanta S Barman M Samanta NRA Bio-efficacy trials of carbofuran 3 CG against insect pests of rice J Entomol ZoolStud 2020 8 1529ndash1533
65 Donley N The USA lags behind other agricultural nations in banning harmful pesticides Environ Health 2019 18 44 [CrossRef][PubMed]
66 Adams A Gore J Musser F Cook D Catchot A Walker T Dobbins C Efficacy of selected insecticides applied to hybridrice seed J Econ Entomol 2015 109 200ndash206 [CrossRef]
67 Kumar R Singh B Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) Field Bull Environ Contam Toxicol2013 90 482ndash488 [CrossRef]
68 Dhaka S Prajapati C Singh D Singh R Field evaluation of insecticides and bio-pesticides against rice leaf folder CnaphalocrocisMedinalis (Guenee) Ann Plant Prot Sci 2011 19 324ndash326
69 Matsumura M Takeuchi H Satoh M Sanada-Morimura S Otuka A Watanabe T Van Thanh D Species-specific insecticideresistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in East and South-eastAsia Pest Manag Sci 2008 64 1115ndash1121 [CrossRef] [PubMed]
70 Tang J Li J Shao Y Yang B Liu Z Fipronil resistance in the whitebacked planthopper (Sogatella furcifera) Possible resistancemechanisms and cross-resistance Pest Manag Sci 2010 66 121ndash125 [CrossRef]
71 Horgan FG Arida A Ardestani G Almazan MLP Temperature-dependent oviposition and nymph performance revealdistinct thermal niches of coexisting planthoppers with similar thresholds for development PLoS ONE 2020 15 e0235506[CrossRef] [PubMed]
Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]

Crops 2021 1 181
Institutional Review Board Statement Not applicable
Informed Consent Statement Not applicable
Data Availability Statement The data presented in this study are available on reasonable requestfrom the corresponding author
Acknowledgments The authors thank Fame Ramal Carmencita Bernal Alberto Naredo ReyeulQuintana Vincent Virtudes Marol Recide Jenyrose Geronda and Ellen Genil for assistance duringthis study The contributions of Rodante Abas (sadly deceased) during three decades of entomologicalresearch at IRRI are gratefully acknowledged Rice lines were supplied through the Genebank of IRRI
Conflicts of Interest The authors declare no conflict of interest
References1 Bottrell DG Schoenly KG Resurrecting the ghost of green revolutions past The brown planthopper as a recurring threat to
high-yielding rice production in tropical Asia J Asia-Pac Entomol 2012 15 122ndash140 [CrossRef]2 Horgan FG Integrated pest management for sustainable rice cultivation A holistic approach In Achieving Sustainable Cultivation
of Rice Cultivation Pest and Disease Management Sasaki T Ed Burleigh Dodds Scientific Cambridge UK 2017 pp 309ndash3423 Horgan FG de Freitas TFS Crisol-Martiacutenez E Mundaca EA Bernal CC Nitrogenous fertilizer reduces resistance but
enhances tolerance to the brown planthopper in fast-growing moderately resistant rice Insects 2021 12 989 [CrossRef] [PubMed]4 Horgan FG Pentildealver Cruz A Bernal CC Ramal AF Almazan MLP Wilby A Resistance and tolerance to the brown
planthopper Nilaparvata lugens (Staringl) in rice infested at different growth stages across a gradient of nitrogen applicationsField Crops Res 2018 217 53ndash65 [CrossRef]
5 Hereward JP Cai X Matias AMA Walter GH Xu C Wang Y Migration dynamics of an important rice pest The brownplanthopper (Nilaparvata lugens) across AsiamdashInsights from population genomics Evol Appl 2020 13 2449ndash2459 [CrossRef]
6 Hu C Guo J Fu X Huang Y Gao X Wu K Seasonal migration pattern of Nilaparvata lugens (Hemiptera Delphacidae) overthe Bohai Sea in northern China J Econ Entomol 2018 111 2129ndash2135 [CrossRef]
7 Ma J Wang Y-C Hu Y-Y Lu M-H Wan G-J Chen F-J Liu W-C Zhai B-P Hu G Brown planthopper Nilaparvatalugens was concentrated at the rear of the typhoon Soudelor in eastern China in August 2015 Insect Sci 2018 25 916ndash926[CrossRef]
8 Fujita D Kohli A Horgan FG Rice resistance to planthoppers and leafhoppers Crit Rev Plant Sci 2013 32 162ndash191[CrossRef]
9 Fujii T Yoshida K Kobayashi T Myint KKM Yasui H Sanada-Morimura S Matsumura M Long-term virulencemonitoring of differential cultivars in Japanrsquos immigrant populations of Nilaparvata lugens (Hemiptera Delphacidae) in 2001ndash2009Appl Entomol Zool 2021 56 407ndash418 [CrossRef]
10 Myint KKM Yasui H Takagi M Matsumura M Virulence of long-term laboratory populations of the brown planthopperNilaparvata lugens (Stal) and whitebacked planthopper Sogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) on rice differentialvarieties Appl Entomol Zool 2009 44 149ndash153 [CrossRef]
11 Horgan FG Ramal AF Bentur JS Kumar R Bhanu KV Sarao PS Iswanto EH Van Chien H Phyu MHBernal CC et al Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia againstresistant rice varieties Crop Prot 2015 78 222ndash231 [CrossRef]
12 Horgan FG Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers Crop Prot2018 110 21ndash33 [CrossRef]
13 Uddin ABMA Islam KS Jahan M Ara A Afrin S Farmerrsquos perception about resurgence of brown planthopper NilaparvataLugens (Staringl) in Bangladesh Bangladesh Rice J 2020 23 21ndash33 [CrossRef]
14 Triwidodo H Brown planthoppers infestations and insecticides use pattern in Java Indonesia AGRIVITA J Agric Sci 202042 320ndash330 [CrossRef]
15 Veres A Wyckhuys KA Kiss J Toacuteth F Burgio G Pons X Avilla C Vidal S Razinger J Bazok R et al An update of theWorldwide Integrated Assessment (WIA) on systemic pesticides Part 4 Alternatives in major cropping systems Environ SciPollut Res 2020 27 29867ndash29899 [CrossRef]
16 Bakker L van der Werf W Tittonell P Wyckhuys KA Bianchi FJ Neonicotinoids in global agriculture Evidence for a newpesticide treadmill Ecol Soc 2020 25 26 [CrossRef]
17 Wu J-C Xu J-X Yuan S-Z Liu J-L Jiang Y-H Xu J-F Pesticide-induced susceptibility of rice to brown planthopperNilaparvata lugens Entomol Exp Appl 2001 100 119ndash126 [CrossRef]
18 Wu J-C Qiu H-M Yang G-Q Liu J-L Liu G-J Wilkens RM Effective duration of pesticide-induced susceptibility of riceto brown planthopper (Nilaparvata lugens Staringl Homoptera Delphacidae) and physiological and biochemical changes in riceplants following pesticide application Int J Pest Manag 2004 50 55ndash62 [CrossRef]
19 Wang A-H Wu J-C Yu Y-S Liu J-L Yue J-F Wang MY Selective insecticide-induced stimulation on fecundity andbiochemical changes in Tryporyza incertulas (Lepidoptera Pyralidae) J Econ Entomol 2005 98 1144ndash1149 [CrossRef] [PubMed]
Crops 2021 1 182
20 Azzam S Wang F Wu J-C Shen J Wang L-P Yang G-Q Guo Y-R Comparisons of stimulatory effects of a seriesof concentrations of four insecticides on reproduction in the rice brown planthopper Nilaparvata lugens Staringl (HomopteraDelphacidae) Int J Pest Manag 2009 55 347ndash358 [CrossRef]
21 Ge L-Q Hu J-H Wu J-C Yang G-Q Gu H Insecticide-induced changes in protein RNA and DNA contents in ovary andfat body of female Nilaparvata lugens (Hemiptera Delphacidae) J Econ Entomol 2009 102 1506ndash1514 [CrossRef]
22 Ling S Zhang J Hu L Zhang R Effect of fipronil on the reproduction feeding and relative fitness of brown planthopperNilaparvata lugens Appl Entomol Zool 2009 44 543ndash548 [CrossRef]
23 Ge L-Q Wang L-P Zhao K-F Wu J-C Huang L-J Mating pair combinations of insecticide-treated male and femaleNilaparvata lugens Staringl (Hemiptera Delphacidae) planthoppers influence protein content in the male accessory glands (MAGs)and vitellin content in both fat bodies and ovaries of adult females Pestic Biochem Physiol 2010 98 279ndash288 [CrossRef]
24 Ge L-Q Wu J-C Zhao K-F Chen Y Yang G-Q Induction of Nlvg and suppression of Nljhe gene expression in Nilaparvatalugens (Staringl) (Hemiptera Delphacidae) adult females and males exposed to two insecticides Pestic Biochem Physiol 201098 269ndash278 [CrossRef]
25 Hu J-H Wu J-C Yin J-L Gu H-N Physiology of insecticide-induced stimulation of reproduction in the rice brownplanthopper (Nilaparvata lugens (Staringl)) Dynamics of protein in fat body and ovary Int J Pest Manag 2010 56 23ndash30 [CrossRef]
26 Wang L-P Shen J Ge L-Q Wu J-C Yang G-Q Jahn GC Insecticide-induced increase in the protein content of maleaccessory glands and its effect on the fecundity of females in the brown planthopper Nilaparvata lugens Staringl (HemipteraDelphacidae) Crop Prot 2010 29 1280ndash1285 [CrossRef]
27 Wu J Ge L Liu F Song Q Stanley D Pesticide-induced planthopper population resurgence in rice cropping systems AnnuRev Entomol 2020 65 409ndash429 [CrossRef]
28 Suri KS Singh G Insecticide-induced resurgence of the whitebacked planthopper Sogatella furcifera (Horvath) (HemipteraDelphacidae) on rice varieties with different levels of resistance Crop Prot 2011 30 118ndash124 [CrossRef]
29 Fabellar LT Heinrichs EA Toxicity of insecticides to predators of rice brown planthoppers Nilaparvata lugens (Staringl) (HomopteraDelphacidae) Environ Entomol 1984 13 832ndash837 [CrossRef]
30 Fabellar LT Heinrichs EA Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators Crop Prot1986 5 254ndash258 [CrossRef]
31 Cuong NL Ben PT Phuong LT Chau LM Cohen MB Effect of host plant resistance and insecticide on brown planthopperNilaparvata lugens (Staringl) and predator population development in the Mekong Delta Vietnam Crop Prot 1997 16 707ndash715[CrossRef]
32 Preetha G Stanley J Suresh S Samiyappan R Risk assessment of insecticides used in rice on mirid bug Cyrtorhinuslividipennis Reuter the important predator of brown planthopper Nilaparvata lugens (Stal) Chemosphere 2010 80 498ndash503[CrossRef] [PubMed]
33 Tanaka K Endo S Kazano H Toxicity of insecticides to predators of rice planthoppers Spiders the mirid bug and the dryinidwasp Appl Entomol Zool 2000 35 177ndash187 [CrossRef]
34 Zhang X Xu Q Lu W Liu F Sublethal effects of four synthetic insecticides on the generalist predator Cyrtorhinus lividipennisJ Pest Sci 2015 88 383ndash392 [CrossRef]
35 Settle WH Ariawan H Astuti ET Cahyana W Hakim AL Hindayana D Lestari AS Managing tropical rice peststhrough conservation of generalist natural enemies and alternative prey Ecology 1996 77 1975ndash1988 [CrossRef]
36 Salim M Heinrichs EA Insecticide-induced changes in the levels of resistance of rice cultivars to the whitebacked planthopperSogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) Crop Prot 1987 6 28ndash32 [CrossRef]
37 Yin J-L Xu H-W Wu J-C Hu J-H Yang G-Q Cultivar and insecticide applications affect the physiological development ofthe brown planthopper Nilaparvata lugens (Staringl) (Hemiptera Delphacidae) Environ Entomol 2014 37 206ndash212 [CrossRef]
38 Cheng C Chang W Studies on varietal resistance to the brown planthopper in Taiwan In Brown Planthopper Threat toRice Production in Asia International Rice Research Institute Ed International Rice Research Institute Manila Philippines1979 pp 251ndash271
39 Reissig WH Heinrichs EA Valencia SL Insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on ricevarieties with different levels of resistance Environ Entomol 1982 11 165ndash168 [CrossRef]
40 Heinrichs EA Fabellar LT Basilio RP Wen T-C Medrano F Susceptibility of rice planthoppers Nilaparvata lugens andSogatella furcifera (Homoptera Delphacidae) to insecticides as influenced by level of resistance in the host plant Environ Entomol1984 13 455ndash458 [CrossRef]
41 Joshi R Shepard B Kenmore P Lydia R Insecticide-induced resurgence of brown planthopper (BPH) on IR62 Int Rice ResNewsl 1992 17 9ndash10
42 Zhu Z-R Cheng J Jiang M-X Zhang X-X Complex influence of rice variety fertilization timing and insecticide onpopulation dynamics of Sogatella furcifera (Horvath) Nilaparvata lugens (Staringl) (Homoptera Delphacidae) and their natural enemiesin rice in Hangzhou China J Pest Sci 2004 77 65ndash74 [CrossRef]
43 Nanthakumar M Jhansi Lakshmi V Shashi Bhushan V Balachandran SM Mohan M Decrease of rice plant resistance andinduction of hormesis and carboxylesterase titre in brown planthopper Nilaparvata lugens (Staringl) by xenobiotics Pestic BiochemPhysiol 2012 102 146ndash152 [CrossRef]
Crops 2021 1 183
44 Yao H-W Jiang C-Y Ye G-Y Cheng J-A Studies on cultivar-induced changes in insecticide susceptibility and its relatedenzyme activities of the white-backed planthopper Sogatella furcifera (Horvath)(Homopter Delphacidae) Insect Sci 2002 9 9ndash15
45 Horgan FG Srinivasan TS Bentur JS Kumar R Bhanu KV Sarao PS Van Chien H Almazan MLP Bernal CCRamal AF et al Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers Implicationsfor rice breeding and gene deployment Agronomy 2017 7 62 [CrossRef]
46 Jairin J Phengrat K Teangdeerith S Vanavichit A Toojinda T Mapping of a broad-spectrum brown planthopper resistancegene Bph3 on rice chromosome 6 Mol Breed 2007 19 35ndash44 [CrossRef]
47 Ren J Gao F Wu X Lu X Zeng L Lv J Su X Luo H Ren G Bph32 a novel gene encoding an unknown SCRdomain-containing protein confers resistance against the brown planthopper in rice Sci Rep 2016 6 37645 [CrossRef]
48 Nguyen CD Zheng S-H Sanada-Morimura S Matsumura M Yasui H Fujita D Substitution mapping and characterizationof brown planthopper resistance genes from indica rice variety lsquoPTB33prime (Oryza sativa L) Breed Sci 2021 in press [CrossRef]
49 Chelliah S Heinrichs EA Factors affecting insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on RiceEnviron Entomol 1980 9 773ndash777 [CrossRef]
50 Moore MT Kroumlger R Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growthArch Environ Contam Toxicol 2010 59 574ndash581 [CrossRef]
51 Kim J Hwang I Han M Jang B Effect of insecticide and fungicide on phytotoxicity of herbicide in rice Korean J Weed Sci1986 6 67ndash75
52 Horgan FG Garcia CPF Haverkort F de Jong PW Ferrater JB Changes in insecticide resistance and host range performanceof planthoppers artificially selected to feed on resistant rice Crop Prot 2020 127 104963 [CrossRef] [PubMed]
53 Khush GS Virk PS IR Varieties and Their Impact International Rice Research Institute Manila Philippines 200554 Srinivasan TS Almazan MLP Bernal CC Ramal AF Subbarayalu MK Horgan FG Interactions between nymphs of
Nilaparvata lugens and Sogatella furcifera (Hemiptera Delphacidae) on resistant and susceptible rice varieties Appl Entomol Zool2016 51 81ndash90 [CrossRef]
55 Pentildealver Cruz A Arida A Heong KL Horgan FG Aspects of brown planthopper adaptation to resistant rice varieties withthe Bph3 gene Entomol Exp Appl 2011 141 245ndash257 [CrossRef]
56 Alam SN Cohen MB Detection and analysis of QTLs for resistance to the brown planthopper Nilaparvata lugens in adoubled-haploid rice population Theor Appl Genet 1998 97 1370ndash1379 [CrossRef]
57 Mackill DJ Khush GS IR64 A high-quality and high-yielding mega variety Rice 2018 11 18 [CrossRef] [PubMed]58 Heinrichs EA Basilio RP Valencia SL Buprofezin a selective insecticide for the management of rice planthoppers (Ho-
moptera Delphacidae) and leafhoppers (Homoptera Cicadellidae) Environ Entomol 1984 13 515ndash521 [CrossRef]59 Satapathy MK Anjaneyulu A Use of cypermethrin a synthetic pyrethroid in the control of rice tungro virus disease and its
vector Trop Pest Manag 1984 30 170ndash178 [CrossRef]60 Vorley W Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown
planthopper (BPH) in Kedah Malaysia Int Rice Res Newsl 1987 10 19ndash2061 Uddin A Islam KS Jahan M Ara A Khan MAI Factor influencing the resurgence of brown planthopper in Bangladesh
SAARC J Agric 2020 18 117ndash128 [CrossRef]62 Escalada MM Luecha M Heong KL Social impacts of planthopper outbreaks Case study from Thailand In Rice Planthoppers
Ecology Management Socio Economics and Policy Heong KL Cheng J Escalada MM Eds Zhejiang University PressHangzhou China 2015 pp 209ndash226
63 Zhang CP He HM Yu JZ Hu XQ Zhu YH Wang Q Residues of carbosulfan and its metabolites carbofuran and3-hydroxy carbofuran in rice field ecosystem in China J Environ Sci Health 2016 51 351ndash357 [CrossRef] [PubMed]
64 Samanta S Barman M Samanta NRA Bio-efficacy trials of carbofuran 3 CG against insect pests of rice J Entomol ZoolStud 2020 8 1529ndash1533
65 Donley N The USA lags behind other agricultural nations in banning harmful pesticides Environ Health 2019 18 44 [CrossRef][PubMed]
66 Adams A Gore J Musser F Cook D Catchot A Walker T Dobbins C Efficacy of selected insecticides applied to hybridrice seed J Econ Entomol 2015 109 200ndash206 [CrossRef]
67 Kumar R Singh B Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) Field Bull Environ Contam Toxicol2013 90 482ndash488 [CrossRef]
68 Dhaka S Prajapati C Singh D Singh R Field evaluation of insecticides and bio-pesticides against rice leaf folder CnaphalocrocisMedinalis (Guenee) Ann Plant Prot Sci 2011 19 324ndash326
69 Matsumura M Takeuchi H Satoh M Sanada-Morimura S Otuka A Watanabe T Van Thanh D Species-specific insecticideresistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in East and South-eastAsia Pest Manag Sci 2008 64 1115ndash1121 [CrossRef] [PubMed]
70 Tang J Li J Shao Y Yang B Liu Z Fipronil resistance in the whitebacked planthopper (Sogatella furcifera) Possible resistancemechanisms and cross-resistance Pest Manag Sci 2010 66 121ndash125 [CrossRef]
71 Horgan FG Arida A Ardestani G Almazan MLP Temperature-dependent oviposition and nymph performance revealdistinct thermal niches of coexisting planthoppers with similar thresholds for development PLoS ONE 2020 15 e0235506[CrossRef] [PubMed]
Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]

Crops 2021 1 182
20 Azzam S Wang F Wu J-C Shen J Wang L-P Yang G-Q Guo Y-R Comparisons of stimulatory effects of a seriesof concentrations of four insecticides on reproduction in the rice brown planthopper Nilaparvata lugens Staringl (HomopteraDelphacidae) Int J Pest Manag 2009 55 347ndash358 [CrossRef]
21 Ge L-Q Hu J-H Wu J-C Yang G-Q Gu H Insecticide-induced changes in protein RNA and DNA contents in ovary andfat body of female Nilaparvata lugens (Hemiptera Delphacidae) J Econ Entomol 2009 102 1506ndash1514 [CrossRef]
22 Ling S Zhang J Hu L Zhang R Effect of fipronil on the reproduction feeding and relative fitness of brown planthopperNilaparvata lugens Appl Entomol Zool 2009 44 543ndash548 [CrossRef]
23 Ge L-Q Wang L-P Zhao K-F Wu J-C Huang L-J Mating pair combinations of insecticide-treated male and femaleNilaparvata lugens Staringl (Hemiptera Delphacidae) planthoppers influence protein content in the male accessory glands (MAGs)and vitellin content in both fat bodies and ovaries of adult females Pestic Biochem Physiol 2010 98 279ndash288 [CrossRef]
24 Ge L-Q Wu J-C Zhao K-F Chen Y Yang G-Q Induction of Nlvg and suppression of Nljhe gene expression in Nilaparvatalugens (Staringl) (Hemiptera Delphacidae) adult females and males exposed to two insecticides Pestic Biochem Physiol 201098 269ndash278 [CrossRef]
25 Hu J-H Wu J-C Yin J-L Gu H-N Physiology of insecticide-induced stimulation of reproduction in the rice brownplanthopper (Nilaparvata lugens (Staringl)) Dynamics of protein in fat body and ovary Int J Pest Manag 2010 56 23ndash30 [CrossRef]
26 Wang L-P Shen J Ge L-Q Wu J-C Yang G-Q Jahn GC Insecticide-induced increase in the protein content of maleaccessory glands and its effect on the fecundity of females in the brown planthopper Nilaparvata lugens Staringl (HemipteraDelphacidae) Crop Prot 2010 29 1280ndash1285 [CrossRef]
27 Wu J Ge L Liu F Song Q Stanley D Pesticide-induced planthopper population resurgence in rice cropping systems AnnuRev Entomol 2020 65 409ndash429 [CrossRef]
28 Suri KS Singh G Insecticide-induced resurgence of the whitebacked planthopper Sogatella furcifera (Horvath) (HemipteraDelphacidae) on rice varieties with different levels of resistance Crop Prot 2011 30 118ndash124 [CrossRef]
29 Fabellar LT Heinrichs EA Toxicity of insecticides to predators of rice brown planthoppers Nilaparvata lugens (Staringl) (HomopteraDelphacidae) Environ Entomol 1984 13 832ndash837 [CrossRef]
30 Fabellar LT Heinrichs EA Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators Crop Prot1986 5 254ndash258 [CrossRef]
31 Cuong NL Ben PT Phuong LT Chau LM Cohen MB Effect of host plant resistance and insecticide on brown planthopperNilaparvata lugens (Staringl) and predator population development in the Mekong Delta Vietnam Crop Prot 1997 16 707ndash715[CrossRef]
32 Preetha G Stanley J Suresh S Samiyappan R Risk assessment of insecticides used in rice on mirid bug Cyrtorhinuslividipennis Reuter the important predator of brown planthopper Nilaparvata lugens (Stal) Chemosphere 2010 80 498ndash503[CrossRef] [PubMed]
33 Tanaka K Endo S Kazano H Toxicity of insecticides to predators of rice planthoppers Spiders the mirid bug and the dryinidwasp Appl Entomol Zool 2000 35 177ndash187 [CrossRef]
34 Zhang X Xu Q Lu W Liu F Sublethal effects of four synthetic insecticides on the generalist predator Cyrtorhinus lividipennisJ Pest Sci 2015 88 383ndash392 [CrossRef]
35 Settle WH Ariawan H Astuti ET Cahyana W Hakim AL Hindayana D Lestari AS Managing tropical rice peststhrough conservation of generalist natural enemies and alternative prey Ecology 1996 77 1975ndash1988 [CrossRef]
36 Salim M Heinrichs EA Insecticide-induced changes in the levels of resistance of rice cultivars to the whitebacked planthopperSogatella furcifera (Horvaacuteth) (Homoptera Delphacidae) Crop Prot 1987 6 28ndash32 [CrossRef]
37 Yin J-L Xu H-W Wu J-C Hu J-H Yang G-Q Cultivar and insecticide applications affect the physiological development ofthe brown planthopper Nilaparvata lugens (Staringl) (Hemiptera Delphacidae) Environ Entomol 2014 37 206ndash212 [CrossRef]
38 Cheng C Chang W Studies on varietal resistance to the brown planthopper in Taiwan In Brown Planthopper Threat toRice Production in Asia International Rice Research Institute Ed International Rice Research Institute Manila Philippines1979 pp 251ndash271
39 Reissig WH Heinrichs EA Valencia SL Insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on ricevarieties with different levels of resistance Environ Entomol 1982 11 165ndash168 [CrossRef]
40 Heinrichs EA Fabellar LT Basilio RP Wen T-C Medrano F Susceptibility of rice planthoppers Nilaparvata lugens andSogatella furcifera (Homoptera Delphacidae) to insecticides as influenced by level of resistance in the host plant Environ Entomol1984 13 455ndash458 [CrossRef]
41 Joshi R Shepard B Kenmore P Lydia R Insecticide-induced resurgence of brown planthopper (BPH) on IR62 Int Rice ResNewsl 1992 17 9ndash10
42 Zhu Z-R Cheng J Jiang M-X Zhang X-X Complex influence of rice variety fertilization timing and insecticide onpopulation dynamics of Sogatella furcifera (Horvath) Nilaparvata lugens (Staringl) (Homoptera Delphacidae) and their natural enemiesin rice in Hangzhou China J Pest Sci 2004 77 65ndash74 [CrossRef]
43 Nanthakumar M Jhansi Lakshmi V Shashi Bhushan V Balachandran SM Mohan M Decrease of rice plant resistance andinduction of hormesis and carboxylesterase titre in brown planthopper Nilaparvata lugens (Staringl) by xenobiotics Pestic BiochemPhysiol 2012 102 146ndash152 [CrossRef]
Crops 2021 1 183
44 Yao H-W Jiang C-Y Ye G-Y Cheng J-A Studies on cultivar-induced changes in insecticide susceptibility and its relatedenzyme activities of the white-backed planthopper Sogatella furcifera (Horvath)(Homopter Delphacidae) Insect Sci 2002 9 9ndash15
45 Horgan FG Srinivasan TS Bentur JS Kumar R Bhanu KV Sarao PS Van Chien H Almazan MLP Bernal CCRamal AF et al Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers Implicationsfor rice breeding and gene deployment Agronomy 2017 7 62 [CrossRef]
46 Jairin J Phengrat K Teangdeerith S Vanavichit A Toojinda T Mapping of a broad-spectrum brown planthopper resistancegene Bph3 on rice chromosome 6 Mol Breed 2007 19 35ndash44 [CrossRef]
47 Ren J Gao F Wu X Lu X Zeng L Lv J Su X Luo H Ren G Bph32 a novel gene encoding an unknown SCRdomain-containing protein confers resistance against the brown planthopper in rice Sci Rep 2016 6 37645 [CrossRef]
48 Nguyen CD Zheng S-H Sanada-Morimura S Matsumura M Yasui H Fujita D Substitution mapping and characterizationof brown planthopper resistance genes from indica rice variety lsquoPTB33prime (Oryza sativa L) Breed Sci 2021 in press [CrossRef]
49 Chelliah S Heinrichs EA Factors affecting insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on RiceEnviron Entomol 1980 9 773ndash777 [CrossRef]
50 Moore MT Kroumlger R Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growthArch Environ Contam Toxicol 2010 59 574ndash581 [CrossRef]
51 Kim J Hwang I Han M Jang B Effect of insecticide and fungicide on phytotoxicity of herbicide in rice Korean J Weed Sci1986 6 67ndash75
52 Horgan FG Garcia CPF Haverkort F de Jong PW Ferrater JB Changes in insecticide resistance and host range performanceof planthoppers artificially selected to feed on resistant rice Crop Prot 2020 127 104963 [CrossRef] [PubMed]
53 Khush GS Virk PS IR Varieties and Their Impact International Rice Research Institute Manila Philippines 200554 Srinivasan TS Almazan MLP Bernal CC Ramal AF Subbarayalu MK Horgan FG Interactions between nymphs of
Nilaparvata lugens and Sogatella furcifera (Hemiptera Delphacidae) on resistant and susceptible rice varieties Appl Entomol Zool2016 51 81ndash90 [CrossRef]
55 Pentildealver Cruz A Arida A Heong KL Horgan FG Aspects of brown planthopper adaptation to resistant rice varieties withthe Bph3 gene Entomol Exp Appl 2011 141 245ndash257 [CrossRef]
56 Alam SN Cohen MB Detection and analysis of QTLs for resistance to the brown planthopper Nilaparvata lugens in adoubled-haploid rice population Theor Appl Genet 1998 97 1370ndash1379 [CrossRef]
57 Mackill DJ Khush GS IR64 A high-quality and high-yielding mega variety Rice 2018 11 18 [CrossRef] [PubMed]58 Heinrichs EA Basilio RP Valencia SL Buprofezin a selective insecticide for the management of rice planthoppers (Ho-
moptera Delphacidae) and leafhoppers (Homoptera Cicadellidae) Environ Entomol 1984 13 515ndash521 [CrossRef]59 Satapathy MK Anjaneyulu A Use of cypermethrin a synthetic pyrethroid in the control of rice tungro virus disease and its
vector Trop Pest Manag 1984 30 170ndash178 [CrossRef]60 Vorley W Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown
planthopper (BPH) in Kedah Malaysia Int Rice Res Newsl 1987 10 19ndash2061 Uddin A Islam KS Jahan M Ara A Khan MAI Factor influencing the resurgence of brown planthopper in Bangladesh
SAARC J Agric 2020 18 117ndash128 [CrossRef]62 Escalada MM Luecha M Heong KL Social impacts of planthopper outbreaks Case study from Thailand In Rice Planthoppers
Ecology Management Socio Economics and Policy Heong KL Cheng J Escalada MM Eds Zhejiang University PressHangzhou China 2015 pp 209ndash226
63 Zhang CP He HM Yu JZ Hu XQ Zhu YH Wang Q Residues of carbosulfan and its metabolites carbofuran and3-hydroxy carbofuran in rice field ecosystem in China J Environ Sci Health 2016 51 351ndash357 [CrossRef] [PubMed]
64 Samanta S Barman M Samanta NRA Bio-efficacy trials of carbofuran 3 CG against insect pests of rice J Entomol ZoolStud 2020 8 1529ndash1533
65 Donley N The USA lags behind other agricultural nations in banning harmful pesticides Environ Health 2019 18 44 [CrossRef][PubMed]
66 Adams A Gore J Musser F Cook D Catchot A Walker T Dobbins C Efficacy of selected insecticides applied to hybridrice seed J Econ Entomol 2015 109 200ndash206 [CrossRef]
67 Kumar R Singh B Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) Field Bull Environ Contam Toxicol2013 90 482ndash488 [CrossRef]
68 Dhaka S Prajapati C Singh D Singh R Field evaluation of insecticides and bio-pesticides against rice leaf folder CnaphalocrocisMedinalis (Guenee) Ann Plant Prot Sci 2011 19 324ndash326
69 Matsumura M Takeuchi H Satoh M Sanada-Morimura S Otuka A Watanabe T Van Thanh D Species-specific insecticideresistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in East and South-eastAsia Pest Manag Sci 2008 64 1115ndash1121 [CrossRef] [PubMed]
70 Tang J Li J Shao Y Yang B Liu Z Fipronil resistance in the whitebacked planthopper (Sogatella furcifera) Possible resistancemechanisms and cross-resistance Pest Manag Sci 2010 66 121ndash125 [CrossRef]
71 Horgan FG Arida A Ardestani G Almazan MLP Temperature-dependent oviposition and nymph performance revealdistinct thermal niches of coexisting planthoppers with similar thresholds for development PLoS ONE 2020 15 e0235506[CrossRef] [PubMed]
Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]

Crops 2021 1 183
44 Yao H-W Jiang C-Y Ye G-Y Cheng J-A Studies on cultivar-induced changes in insecticide susceptibility and its relatedenzyme activities of the white-backed planthopper Sogatella furcifera (Horvath)(Homopter Delphacidae) Insect Sci 2002 9 9ndash15
45 Horgan FG Srinivasan TS Bentur JS Kumar R Bhanu KV Sarao PS Van Chien H Almazan MLP Bernal CCRamal AF et al Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers Implicationsfor rice breeding and gene deployment Agronomy 2017 7 62 [CrossRef]
46 Jairin J Phengrat K Teangdeerith S Vanavichit A Toojinda T Mapping of a broad-spectrum brown planthopper resistancegene Bph3 on rice chromosome 6 Mol Breed 2007 19 35ndash44 [CrossRef]
47 Ren J Gao F Wu X Lu X Zeng L Lv J Su X Luo H Ren G Bph32 a novel gene encoding an unknown SCRdomain-containing protein confers resistance against the brown planthopper in rice Sci Rep 2016 6 37645 [CrossRef]
48 Nguyen CD Zheng S-H Sanada-Morimura S Matsumura M Yasui H Fujita D Substitution mapping and characterizationof brown planthopper resistance genes from indica rice variety lsquoPTB33prime (Oryza sativa L) Breed Sci 2021 in press [CrossRef]
49 Chelliah S Heinrichs EA Factors affecting insecticide-induced resurgence of the brown planthopper Nilaparvata lugens on RiceEnviron Entomol 1980 9 773ndash777 [CrossRef]
50 Moore MT Kroumlger R Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growthArch Environ Contam Toxicol 2010 59 574ndash581 [CrossRef]
51 Kim J Hwang I Han M Jang B Effect of insecticide and fungicide on phytotoxicity of herbicide in rice Korean J Weed Sci1986 6 67ndash75
52 Horgan FG Garcia CPF Haverkort F de Jong PW Ferrater JB Changes in insecticide resistance and host range performanceof planthoppers artificially selected to feed on resistant rice Crop Prot 2020 127 104963 [CrossRef] [PubMed]
53 Khush GS Virk PS IR Varieties and Their Impact International Rice Research Institute Manila Philippines 200554 Srinivasan TS Almazan MLP Bernal CC Ramal AF Subbarayalu MK Horgan FG Interactions between nymphs of
Nilaparvata lugens and Sogatella furcifera (Hemiptera Delphacidae) on resistant and susceptible rice varieties Appl Entomol Zool2016 51 81ndash90 [CrossRef]
55 Pentildealver Cruz A Arida A Heong KL Horgan FG Aspects of brown planthopper adaptation to resistant rice varieties withthe Bph3 gene Entomol Exp Appl 2011 141 245ndash257 [CrossRef]
56 Alam SN Cohen MB Detection and analysis of QTLs for resistance to the brown planthopper Nilaparvata lugens in adoubled-haploid rice population Theor Appl Genet 1998 97 1370ndash1379 [CrossRef]
57 Mackill DJ Khush GS IR64 A high-quality and high-yielding mega variety Rice 2018 11 18 [CrossRef] [PubMed]58 Heinrichs EA Basilio RP Valencia SL Buprofezin a selective insecticide for the management of rice planthoppers (Ho-
moptera Delphacidae) and leafhoppers (Homoptera Cicadellidae) Environ Entomol 1984 13 515ndash521 [CrossRef]59 Satapathy MK Anjaneyulu A Use of cypermethrin a synthetic pyrethroid in the control of rice tungro virus disease and its
vector Trop Pest Manag 1984 30 170ndash178 [CrossRef]60 Vorley W Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown
planthopper (BPH) in Kedah Malaysia Int Rice Res Newsl 1987 10 19ndash2061 Uddin A Islam KS Jahan M Ara A Khan MAI Factor influencing the resurgence of brown planthopper in Bangladesh
SAARC J Agric 2020 18 117ndash128 [CrossRef]62 Escalada MM Luecha M Heong KL Social impacts of planthopper outbreaks Case study from Thailand In Rice Planthoppers
Ecology Management Socio Economics and Policy Heong KL Cheng J Escalada MM Eds Zhejiang University PressHangzhou China 2015 pp 209ndash226
63 Zhang CP He HM Yu JZ Hu XQ Zhu YH Wang Q Residues of carbosulfan and its metabolites carbofuran and3-hydroxy carbofuran in rice field ecosystem in China J Environ Sci Health 2016 51 351ndash357 [CrossRef] [PubMed]
64 Samanta S Barman M Samanta NRA Bio-efficacy trials of carbofuran 3 CG against insect pests of rice J Entomol ZoolStud 2020 8 1529ndash1533
65 Donley N The USA lags behind other agricultural nations in banning harmful pesticides Environ Health 2019 18 44 [CrossRef][PubMed]
66 Adams A Gore J Musser F Cook D Catchot A Walker T Dobbins C Efficacy of selected insecticides applied to hybridrice seed J Econ Entomol 2015 109 200ndash206 [CrossRef]
67 Kumar R Singh B Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) Field Bull Environ Contam Toxicol2013 90 482ndash488 [CrossRef]
68 Dhaka S Prajapati C Singh D Singh R Field evaluation of insecticides and bio-pesticides against rice leaf folder CnaphalocrocisMedinalis (Guenee) Ann Plant Prot Sci 2011 19 324ndash326
69 Matsumura M Takeuchi H Satoh M Sanada-Morimura S Otuka A Watanabe T Van Thanh D Species-specific insecticideresistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in East and South-eastAsia Pest Manag Sci 2008 64 1115ndash1121 [CrossRef] [PubMed]
70 Tang J Li J Shao Y Yang B Liu Z Fipronil resistance in the whitebacked planthopper (Sogatella furcifera) Possible resistancemechanisms and cross-resistance Pest Manag Sci 2010 66 121ndash125 [CrossRef]
71 Horgan FG Arida A Ardestani G Almazan MLP Temperature-dependent oviposition and nymph performance revealdistinct thermal niches of coexisting planthoppers with similar thresholds for development PLoS ONE 2020 15 e0235506[CrossRef] [PubMed]
Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]

Crops 2021 1 184
72 Horgan FG Arida A Ardestani G Almazan MLP Intraspecific competition counters the effects of elevated and optimaltemperature on phloem-feeding insects in tropical and temperate rice PLoS ONE 2020 15 e0240130 [CrossRef]
73 Sogawa K Cheng C Economic thresholds nature of damage and losses caused by the brown planthopper In BrownPlanthopper Threat to Rice Production in Asia International Rice Research Institute Ed International Rice Research InstituteManila Philippines 1979 pp 125ndash142
74 Zhang Y Guo L Atlihan R Chi H Chu D Demographic analysis of progeny fitness and timing of resurgence of Laodelphaxstriatellus after insecticides exposure Entomol Gen 2019 39 221ndash230 [CrossRef]
75 Zhao K-F Shi Z-P Wu J-C Insecticide-induced enhancement of flight capacity of the brown planthopper Nilaparvata lugensStaringl (Hemiptera Delphacidae) Crop Prot 2011 30 476ndash482 [CrossRef]
76 Nguyen CD Verdeprado H Zita D Sanada-Morimura S Matsumura M Virk PS Brar DS Horgan FG Yasui H FujitaD The development and characterization of near-isogenic and pyramided lines carrying resistance genes to brown planthopperwith the genetic background of japonica rice (Oryza sativa L) Plants 2019 8 498 [CrossRef]
77 Horgan FG Arida A Ardestani G Almazan MLP Positive and negative interspecific interactions between coexisting riceplanthoppers neutralise the effects of elevated temperatures Funct Ecol 2021 35 181ndash192 [CrossRef] [PubMed]
78 Zhou C Yang H Yang H Wang Z Long G-Y Jin D-C Effects of sublethal concentrations of deltamethrin on fitness ofwhite-backed planthopper Sogatella furcifera (Horvaacuteth) Int J Pest Manag 2019 65 165ndash170 [CrossRef]
79 Horgan FG Kudavidanage EP Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystemsof southern Sri Lanka Environ Manag 2020 65 787ndash803 [CrossRef]
80 Kanungo PK Adhya TK Rajaranamohan Rao V Influence of repeated applications of carbofuran on nitrogenase activity andnitrogen-fixing bacteria associated with rhizosphere of tropical rice Chemosphere 1995 31 3249ndash3257 [CrossRef]
81 Horgan FG Crisol Martiacutenez E Stuart AM Bernal CC de Cima Martiacuten E Almazan MLP Ramal AF Effects of vegetationstrips fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystemInsects 2019 10 328 [CrossRef] [PubMed]
82 Widiarta IN Matsumura M Suzuki Y Nakasuji F Effects of sublethal doses of imidacloprid on the fecundity of greenleafhoppers Nephotettix spp (Hemiptera Cicadellidae) and their natural enemies Appl Entomol Zool 2001 36 501ndash507[CrossRef]
83 Preetha G Stanley J Suresh S Kuttalam S Samiyappan R Toxicity of selected insecticides to Trichogramma chilonis Assessingtheir safety in the rice ecosystem Phytoparasitica 2009 37 209ndash215 [CrossRef]
84 Lakshmi VJ Krishnaiah N Pasalu I Lingaiah T Krishnaiah K Safety of thiamethoxam to Cyrtorhinus lividipennis Reuter(Hemiptera Miridae) a predator of brown planthopper Nilaparvata lugens (Stal) in rice J Biol Control 2001 15 53ndash58
85 Horgan FG Ramal AF Villegas JM Jamoralin A Bernal CC Perez MO Pasang JM Naredo AI Almazan MLPEffects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields J ApplEntomol 2017 141 587ndash599 [CrossRef]
86 Fritz LL Heinrichs EA Machado V Andreis TF Pandolfo M de Salles SM de Oliviera JV Fiuza LM Impact oflambdacyhalothrin on arthropod natural enemy populations in irrigated rice fields in southern Brazil Int J Trop Insect Sci 201333 178ndash187 [CrossRef]





























