Embed Size (px)
Citation preview

JOURNAL OF BONE AND MINERAL RESEARCH Volume 6. Number 9. 1991 Mary Ann Liebert. Inc.. Publishers
Role of Colony-Stimulating Factors in Osteoclast Development
NAOYUKI TAKAHASHI, NOBUYUKI UDAGAWA, TAKUHIKO AKATSU, HIROFUMI TANAKA, MANABU SHIONOME, and TATSUO SUDA
ABSTRACT
Effects of various colony-stimulating factors (CSFs) [interleukin3 (IL-3), granulocyte-macrophage CSF (GM-CSF), macrophage CSF (M-CSF), and granulocyte CSF (G-CSF)] on osteoclast-like cell formation were examined in two different culture systems: the one-step mouse marrow culture system and the two-step coculture system of mouse primary osteoblastic cells with the bone marrow cells collected from the colonies that formed in the methylcellulose in the presence of the CSFs. In the one-step mouse marrow cultures, none of the CSFs stimulated the formation of tartrate-resistant acid phosphatase (TRAP, a marker enzyme of osteoc1asts)-positive multinucleated cells (MNCs). Furthermore, the CSFs other than G-CSF inhibited in a dose-dependent manner the TRAP-positive MNC formation induced by la-25-dihydroxyvitamin D, [ la,25- (OH),D,]. In contrast, when marrow cells were first cultured in semisolid methylcellulose in the presence of a CSF and the recovered marrow cells from the semisolid cultures were subsequently cocultured with pri- mary osteoblastic cells in the presence of lcy,25-(0H),D3, numerous TRAP-positive MNCs were formed. [12sI]salmon calcitonin specifically bound to TRAP-positive cells formed in this two-step culture system. Over 90% of the TRAP-positive mononuclear cells and MNCs accumulated ['251]calcitonin. M-CSF was the most potent in inducing TRAP-positive MNCs, followed by GM-CSF, IL-3, and G-CSF in that order. N o TRAP-positive cells were formed in the absence of either osteoblastic cells or la,25-(OH),D3. These results clearly indicate that the CSFs, especially M-CSF, stimulate the growth of osteoclast progenitors, which dif- ferentiate into osteoclast-like cells in response to lc~,25-(OH)~D, in the presence of osteoblastic cells.
INTRODUCTION
STEOCLASTS, the principal cells responsible for bone re- 0 sorption, are multinucleated cells formed by fusion of mononuclear precursors, which are believed to originate from hematopoietic stem cells.".21 The growth factors that regulate the replication and differentiation of hematopoie- tic progenitor cells are well defined and have been termed colony-stimulating factors (CSFS).'~..'~ Four CSFs have been identified: interleukin 3 (IL-3), granulocyte-macro- phage CSF (GM-CSF), macrophage CSF (M-CSF), and granulocyte CSF (G-CSF). IL-3 supports the growth and differentiation of several blood cell lineages. GM-CSF stimulates the production of both macrophages and gran- ulocytes; M-CSF and G-CSF preferentially support the
growth and differentiation of either lineage. Although some of the CSFs apparently regulate the proliferation and differentiation of osteoclast p r o g e n i t o r ~ , ~ ~ - ' ~ ' it is still not known how CSFs are involved in osteoclast development.
We developed two different culture systems to study osteoclast-like cell differentiation: the mouse marrow cul- ture and the coculture system of primary osteo- blast-rich cell populations with spleen cells.' 1 4 ) Hormones that modulate bone-resorbing activity, such as la,25-dihy- droxyvitamin D, [la,25-(OH),D3],'" parathyroid hor- mane, 113-151 and prostaglandin E2'l4 I b 1 all stimulated the formation of osteoclast-like multinucleated cells (MNCs) in the two culture systems. The osteoclast-like MNCs formed in both systems satisfied most of the requirements for definition as osteoclasts. In this study, the previous co-
Department of Biochemistry, School of Dentistry, Showa University, Tokyo, Japan.
977

978 TAKAHASHI ET AL.
culture system was modified to determine the effects of Culture systems for osteoclast-like cell formation CSFs on osteoclast development. We report here that all the CSFs (IL-3, GM-CSF, M-CSF, and G-CSF), especially M-CSF, stimulate the growth of osteoclast progenitors, which differentiate into osteoclast-like MNCs in response to 1a,25-(OH),D3 in the presence of osteoblastic cells.
MATERIALS AND METHODS Materials
Mouse 1L-3, mouse GM-CSF, human M-CSF, and hu- man G-CSF, all recombinant materials, were obtained from Genzyme Co. (Boston, MA). The unit for 11-3 was defined by the manufacturer as the concentration required for half-maximal stimulation of [3H]thymidine incorpora- tion by FDC-P2 cells. The units for GM-CSF, M-CSF, and G-CSF were defined as the concentrations required to pro- duce a single cell colony in soft agar from 7.5 x lo4 bone marrow cells in a standard assay. la,25-(OH),D3 was pur- chased from Philips-Duphar (Amsterdam, The Nether- lands). Salmon calcitonin was kindly supplied by Dr. T. Saito, Teikoku Hormone Manufacturing (Kawasaki, Japan). lzsI-labeled salmon calcitonin was kindly provided by Dr. T.J. Martin, St. Vincent’s Institute of Medical Re- search (Melbourne, Australia). a-Minimum essential me- dium (a-MEM) was obtained from Flow Co. (McLean, VA) and fetal calf serum (FCS) from GlBCO (Grand Is- land, NY). Culture plates and dishes were from Corning (Corning, NY). All other chemicals used in this study were of analytic grade.
Methylcellulose culture
Bone marrow mononuclear cells were obtained from tibiae of 7- to 9-week-old male mice (ddy strain) was previ- ously described.[”) The marrow cells (lo5 cells per ml) were suspended in a-MEM containing 0.88% methylcellu- lose, 100 pM 2-mercaptoethanol, and 10% FCS and cul- tured in 35 mm plastic dishes (2 ml per dish) in the pres- ence of 50 units/ml of IL-3, GM-CSF, M-CSF, or G-CSF at 37°C in a humidified atmosphere of 5% CO, in air. After culture for 6 days, the culture plates were washed thoroughly with a-MEM to recover all the cells grown in the methylcellulose. The cells were then washed three times with a-MEM and resuspended in a-MEM with 10% FCS. In some experiments, each colony was isolated individually with an Eppendorf pipette from M-CSF-treated semisolid cultures, and the cells were washed and resuspended in a-MEM with 10% FCS. To identify monocyte-macro- phages and granulocytes, cytocentrifuge preparations of each marrow cell fraction were fixed and stained for non- specific esterase (NSE, the same enzyme as a-naphthyl ace- tate esterase) and naphthol AS-D chloroacetate esterase (ASE)‘”) using commercially available kits (Sigma). Monocyte-macrophages and granulocytes were positively stained for NSE and ASE, respectively. Cytocentrifuge preparations were also stained for tartrate-resistant acid phosphatase (TRAP) in the presence of 50 nM sodium tar- trate as described previously.c13’
One-Step Mouse Marrow Culture System: Freshly iso- lated whole marrow cells were cultured in a-MEM supple- mented with 10% FCS at 1.5 x lo6 cells per ml in 24-well plates (0.5 ml per well). Cultures were fed every 3 or 4 days by replacing 0.4 ml old medium with fresh medium. Non- adherent cells removed with old medium were not recov- ered, since they were supposed to contain no osteoclast progenitors, judging from the results shown in Fig. 2. In- creasing concentrations of CSFs were added at the begin- ning of the cultures and at each time of medium change. Some of the cultures were treated simultaneously with 10 nM 1a,25-(OH),D3. In other experiments, marrow cells were treated with 50 units/ml of a CSF for the first 4 days. Old medium was completely replaced with fresh medium. Nonadherent cells present in old medium were collected by centrifugation and added to the same wells. The cultures were then treated with 10 nM la,25-(OH),D3 for the subse- quent 4 days. Osteoclast-like cell formation was evaluated after culturing for 8 days.
Two-Step Coculture System: Osteoblastic cells were iso- lated from newborn mouse calvariae and cultured in a- MEM supplemented with 10% FCS as described previ- o ~ s I y . ( l * ~ The osteoblastic cells were subcultured once with trypsin-EDTA, resuspended in a-MEM with 10% FCS, plated at 1 x lo4 cells per well in 24-well plates, and cul- tured for 24 h. Various numbers of freshly isolated mar- row cells or marrow cells recovered from the semisolid methylcellulose cultures were then added to the culture of osteoblastic cells. They were cocultured in the presence or absence of 10 nM la,25-(OH),D1. CSFs (50 units) were added to some cocultures. Osteoclast-like cell formation was evaluated after culturing for 8 days.
Determination of osteoclast characteristics
Cells were fixed and stained for TRAP, and the TRAP- positive cells containing three or more nuclei were counted as osteoclast-like MNCs as described previously.‘I”) Calci- tonin-dependent cAMP production was determined as pre- viously described.“*’ Cells were incubated for 15 minutes with 10 nM salmon calcitonin in the presence of 1 mM iso- butylmethylxanthine. The amount of cAMP in the cell lay- ers was determined using a Yamasa cAMP assay kit (Ya- masa Shoyu Co., Chiba, Japan). Expression of calcitonin receptors was also assessed by autoradiography using [12sI]salmon calcitonin as described After incubation of the cells with 0.2 nM [1z51]salmon calcitonin for 1 h at 22”C, the cells were fixed, stained for TRAP, and processed for autoradiography. Nonspecific binding of [1*51]calcitonin was assessed in the presence of an excess (300 nM) of unlabeled salmon calcitonin.
The results were expressed as the mean + SEM (stan- dard error of the mean) of quadruplicate cultures. Signifi- cance of the differences was determined using Student’s I-test.
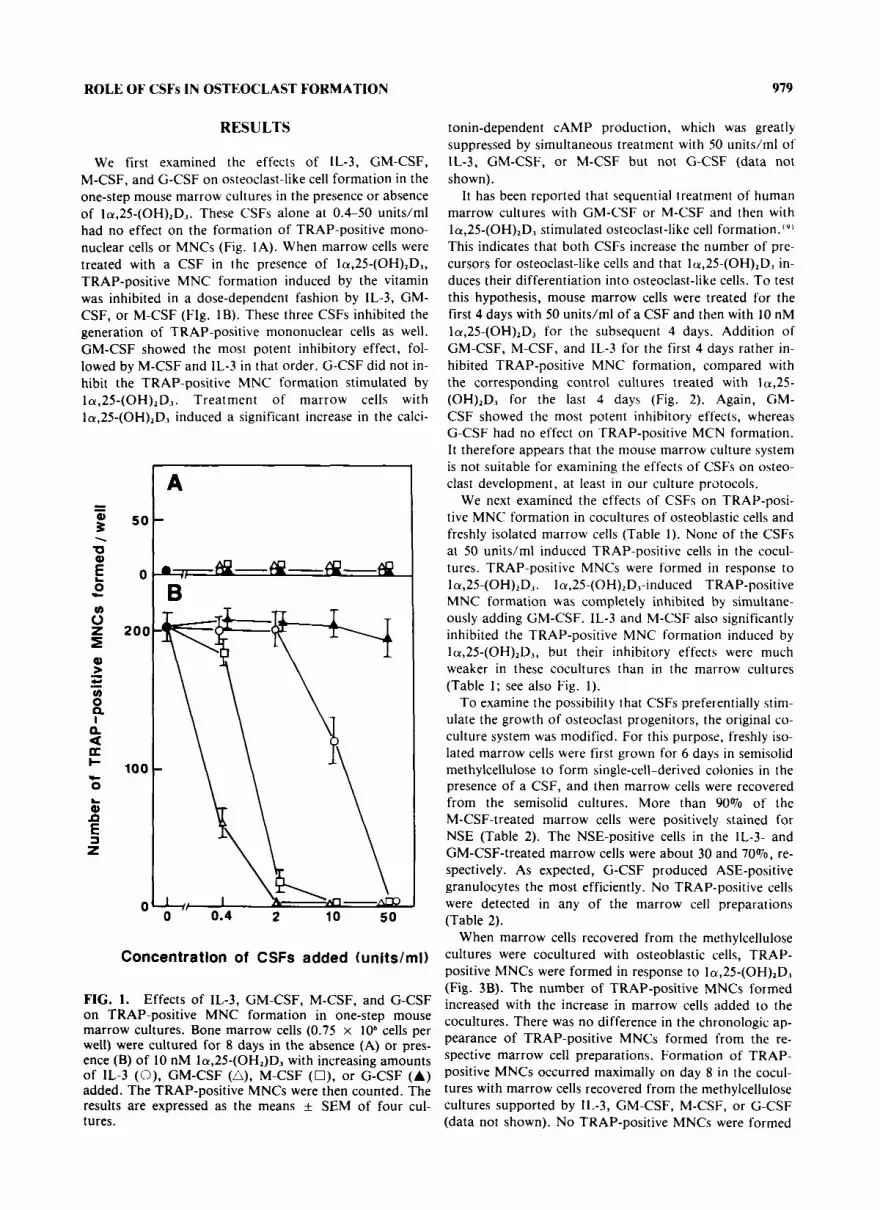
ROLE OF CSFs IN OSTEOCLAST FORMATION 979
RESULTS
We first examined the effects of IL-3, GM-CSF, M-CSF, and G-CSF on osteoclast-like cell formation in the one-step mouse marrow cultures in the presence or absence of la,25-(OH),D1. These CSFs alone at 0.4-50 units/ml had no effect on the formation of TRAP-positive mono- nuclear cells or MNCs (Fig. 1A). When marrow cells were treated with a CSF in the presence of la,25-(OH),D3, TRAP-positive MNC formation induced by the vitamin was inhibited in a dose-dependent fashion by IL-3, GM- CSF, or M-CSF (Flg. 1B). These three CSFs inhibited the generation of TRAP-positive mononuclear cells as well. GM-CSF showed the most potent inhibitory effect, fol- lowed by M-CSF and IL-3 in that order. G-CSF did not in- hibit the TRAP-positive MNC formation stimulated by la,25-(OH),D1. Treatment of marrow cells with la,25-(OH),D3 induced a significant increase in the calci-
0 0.4 2 10 50
Concentration of CSFs added (units/mi)
FIG. 1. Effects of IL-3, GM-CSF, M-CSF, and G-CSF on TRAP-positive MNC formation in one-step mouse marrow cultures. Bone marrow cells (0.75 x lo6 cells per well) were cultured for 8 days in the absence (A) or pres- ence (B) of 10 nM la,25-(OH2)D3 with increasing amounts of 1L-3 (0), GM-CSF (A), M-CSF (O), or G-CSF (A) added. The TRAP-positive MNCs were then counted. The results are expressed as the means k SEM of four cul- tures.
tonin-dependent CAMP production, which was greatly suppressed by simultaneous treatment with SO units/ml of IL-3, GM-CSF, or M-CSF but not G-CSF (data not shown).
It has been reported that sequential treatment of human marrow cultures with GM-CSF or M-CSF and then with la,25-(OH),D3 stimulated osteoclast-like cell formation.''] This indicates that both CSFs increase the number of pre- cursors for osteoclast-like cells and that la,25-(OH),D,5 in- duces their differentiation into osteoclast-like cells. To test this hypothesis, mouse marrow cells were treated for the first 4 days with 50 units/ml of a CSF and then with 10 nM la,25-(OH),D3 for the subsequent 4 days. Addition of GM-CSF, M-CSF, and IL-3 for the first 4 days rather in- hibited TRAP-positive MNC formation, compared with the corresponding control cultures treated with 1u,25- (OH),D, for the last 4 days (Fig. 2). Again, GM- CSF showed the most potent inhibitory effects, whereas G-CSF had no effect on TRAP-positive MCN formation. I t therefore appears that the mouse marrow culture system is not suitable for examining the effects of CSFs on osteo- clast development, at least in our culture protocols.
We next examined the effects of CSFs on TRAP-posi- tive MNC formation in cocultures of osteoblastic cells and freshly isolated marrow cells (Table 1). None of the CSFs at 50 units/ml induced TRAP-positive cells in the cocul- tures. TRAP-positive MNCs were formed in response to la,25-(OH),D3. la,25-(OH),D,l-induced TRAP-positive MNC formation was completely inhibited by simultane- ously adding GM-CSF. IL-3 and M-CSF also significantly inhibited the TRAP-positive MNC formation induced by la,25-(OH),D3, but their inhibitory effects were much weaker in these cocultures than in the marrow cultures (Table 1; see also Fig. I ) .
To examine the possibility that CSFs preferentially stim- ulate the growth of osteoclast progenitors, the original co- culture system was modified. For this purpose, freshly iso- lated marrow cells were first grown for 6 days in semisolid methylcellulose to form single-cell-derived colonies in the presence of a CSF, and then marrow cells were recovered from the semisolid cultures. More than 90% of the M-CSF-treated marrow cells were positively stained for NSE (Table 2). The NSE-positive cells in the IL-3- and GM-CSF-treated marrow cells were about 30 and 70%, re- spectively. As expected, G-CSF produced ASE-positive granulocytes the most efficiently. No TRAP-positive cells were detected in any of the marrow cell preparations (Table 2).
When marrow cells recovered from the methylcellulose cultures were cocultured with osteoblastic cells, TRAP- positive MNCs were formed in response to la,25-(0H),D, (Fig. 3B). The number of TRAP-positive MNCs formed increased with the increase in marrow cells added to the cocultures. There was no difference in the chronologic ap- pearance of TRAP-positive MNCs formed from the re- spective marrow cell preparations. Formation of TRAP- positive MNCs occurred maximally on day 8 in the cocul- tures with marrow cells recovered from the methylcellulose cultures supported by IL-3, GM-CSF, M-CSF, or G-CSF (data not shown). No TRAP-positive MNCs were formed

980 TAKAHASHI ET AL.
T
1 - 4 days vehicle vehicle IL-3 GM-CSF M-CSF G-CSF
vehicle 1,25D3 1,25D3 1,25D3 1,25D3 1925D3 5 - 8 days -- - - - Treat men t
FIG. 2. TRAP-positive MNC formation in one-step mouse marrow cultures sequentially treated with CSFs and la,25- (OH),D,. Bone marrow cells were cultured for the first 4 days with 50 units/ml of IL-3, GM-CSF, M-CSF, or G-CSF. The culture medium was then completely replaced with fresh medium containing 10 nM la,25-(OH),D., ( 1 ,25D3). Nonad- herent cells removed with old medium were collected and added to the same wells. After culturing for additional 4 days, TRAP-positive MNCs were counted. The results are expressed as the means f SEM of four cultures. *Significantly dif- ferent from the cultures treated with la,25-(OH),D, for the last 4 days, p < 0.01.
in the cocultures treated without la,25-(0H),D3 (Fig. 3A). When CSF-treated marrow cells ( 102-104 cells per well) were cultured in the absence of osteoblastic cells, no TRAP-positive cells were formed even in the presence of la,25-(OH),D3 (data not shown).
Autoradiographic studies using ['~51]calcitoinin showed that the labeled calcitonin was bound exclusively to the TRAP-positive mononuclear cells and to the MNCs gener- ated in two-step cocultures of osteoblastic cells with each CSF-treated marrow cell population (Fig. 4). More than 90% of TRAP-positive MNCs and mononuclear cells ac- cumulated dense grains as a result of the specific binding of labeled calcitonin. Accumulation of the grains was com- pletely eliminated by simultaneously added an excess of unlabeled salmon calcitonin (data not shown).
The number of TRAP-positive MNCs formed from 200 marrow cells in the two-step coculture system was esti- mated from the linear plots of TRAP-positive MNC for-
mation against the number of CSF-treated marrow cells added. Figure 5 shows that M-CSF was the most effective in inducing TRAP-positive MNCs, followed by GM-CSF, IL-3, and G-CSF in that order. The marrow cell prepara- tion recovered from the M-CSF-treated semisolid cultures contained about 40 times more precursor cells for TRAP- positive MNCs than freshly isolated marrow cells. This suggests that osteoclast precursors are the cells of the monocyte-macrophage lineage, the growth of which is stimulated by CSFs, especially by M-CSF.
To further examine the hypothesis, single-cell-derived colonies were carefully isolated with an Eppendorf pipette from the methylcellulose cultures supported by M-CSF and cocultured with osteoblastic cells using the two-step coculture system (Table 3). Numerous TRAP-positive MNCs were formed in the cocultures in the presence of la,25-(OH),D3. Calcitonin-dependent CAMP was also pro- duced in the la,25-(OH),D3-treated cocultures. Neither

ROLE OF CSFs IN OSTEOCLAST FORMATION 981
TRAP-positive MNC formation nor significant production of calcitonin-dependent CAMP occurred in the absence of la,25-(OH),D3 (Table 3).
DISCUSSION
It is well established that osteoclasts are derived from hematopoietic stem cells, but the role of CSFs in osteoclast development is controversial. This is due to the obscurity of the origin of osteoclasts and also to the lack of suitable
TABLE 1. EFFECTS OF IL-3, GM-CSF, M-CSF, AND G-CSF ON TRAP-POSITIVE MNC FORMATION IN
COCULTURES OF OSTEOBLASTIC CELLS A N D BONE MARROW C E L L S ~
TRAP-positive MNC formation (number of
TRAP-positive MNC per well), la,ZS-(OH),D,
(nM) Treat men t (50 units/ml) 0 I0
Vehicle IL-3 GM-CSF M-CSF G-CSF
0 275 + 18 0 221 * 14b 0 0 0 211 * l l b 0 285 f 6
aBone marrow cells (lo4 cells per well) were cocultured with osteoblastic cells in the presence or absence of 10 nM ln,25-(OH),D1. IL-3, GM-CSF, M-CSF, G-CSF, or vehicle was added at the beginning of culture and at each time of medium change. Nonadherent cells removed with old me- dium were not recovered. After culture for 8 days, TRAP- positive MNCs were counted. The results are expressed as the means f SEM of four cultures.
bsignificantly different from vehicle treated with la ,25- (OH),D, alone, p < 0.05.
systems to examine the effects of CSFs on osteoclast devel- opment. Lorenzo et al.(*) reported that neither IL-3 nor GM-CSF stimulated osteoclastic bone resorption in organ cultures of fetal mouse long bones. MacDonald et al.‘91 demonstrated that both GM-CSF and M-CSF stimulated osteoclast-like MNC formation in baboon long-term mar- mow cultures. The conflicting results of the effect of IL-3 on osteoclast-like cell formation were also reported in mouse marrow cultures. Barton and Mayer“’) reported that IL-3 stimulated the formation of osteoclast-like MNCs in mouse marrow cultures. Hattersley and Cham- bers‘”) also demonstrated that calcitonin receptor-positive cells were increased by the treatment of mouse marrow cells with IL-3. In contrast, Shinar et al.‘12’ reported that IL-3, GM-CSF, and M-CSF did not stimulate TRAP-posi- tive MNC formation but strongly inhibited the TRAP-pos- itive MNC formation induced by Ia,25-(OH),D,.
In the present study, we examined the effects of CSFs on osteoclast development in two different culture systems. In the one-step marrow cultures, none of the CSFs (IL-3, GM-CSF, M-CSF, and G-CSF) stimulated TRAP-positive MNC formation (Fig. 1). The CSFs other than G-CSF dose dependently inhibited TRAP-positive MNC forma- tion induced by la,25-(OH),D1. Although IL.-3, GM-CSF, and M-CSF all stimulated formation of TRAP-negative MNCs in the marrow cultures, autoradiographic studies in- dicated that the TRAP-negative MNCs were negative for calcitonin receptors (data not shown). These results are in good agreement with those of Shinar et al.‘”’ TRAP-posi- tive MNC formation induced by parathyroid hormone and prostaglandin E, was similarly inhibited by IL.-3, GM-CSF, and M-CSF in the one-step mouse marrow cultures (un- published observation). Furthermore, sequential treatment of the cultures with CSFs for the first 4 days and then with la,25-(OH),D3 for the subsequent 4 days failed to stimu- late TRAP-positive MNC formation (Fig. 2). These results show that the one-step mouse marrow culture system is not suitable for examining CSF effects on osteoclast develop- ment.
TABLE 2. EFFECTS OF IL-3, GM-CSF, M-CSF, AND G-CSF ON THE NUMBER OF COLONIES FORMED IN THE SEMISOLID METHYLCELLULOSE CULTURES OF MOUSE MARROW CELLS AND ON
THE PERCENTAGES OF THE TOTAL MARROW CELLS RECOVERED FROM THE SEMISOLID CULTURES THAT WERE NSE-, ASE-, OR TRAP-POSITIVE C E L L S ~
Cytokine treatment Number of colonies NSE’ ASE’ TRAP+ (SO units/ml) formed per lo5 cells cells (Yo) cells (Yo) cells (Yo)
1L-3 6 8 + 4 30.7 12.8 0 GM-CSF 135 + 12 71.9 6.9 0 M-CSF 8 7 + 8 91.5 1 .O 0 G-CSF 5 7 + 3 10.5 47.2 0
aBone marrow cells (lo5 cells per ml) were cultured in semisolid methylcellulose in the presence of IL-3, CM-CSF, M-CSF, or G-CSF. After culture for 6 days, the number of colonies that consisted of 40 or more cells were scored under an inverted phase microscope. The marrow cells were then recov- ered from the semisolid cultures, cytocentrifuged, and stained for NSE, ASE, and TRAP. The results are expressed as the means i SEM of four cultures for colony formation and also as the percentages of the total marrow cells scored that were NSE-, ASE-, or TRAP-positive (+) cells. More than lo00 cells were scored.

982 TAKAHASHI ET AL.
500 -
400 -
I
5 0 0 1000
Number of bone marrow cells co-cultured with l o 4 osteoblastic cells
FIG. 3. TRAP-positive MNC formation in two-step co- cultures of osteoblastic cells and freshly isolated marrow cells or marrow cells recovered from the semisolid methyl- cellulose cultures supported by IL-3, GM-CSF, M-CSF, or G-CSF. In the two-step cocultures, marrow cells were first cultured in methylcellulose for 6 days with 50 units/ml of IL-3 (0), GM-CSF (A), M-CSF (0), or G-CSF (A). The increased numbers of marrow cells recovered from the methylcellulose cultures or freshly isolated marrow cells ( 0 ) were then cocultured for 8 days with osteoblastic cells in the absence (A) or presence (B) of 10 nM 1a-25- (OH),D,. The TRAP-positive MNCs were then counted. The results are expressed as the means k SEM of four cul- tures.
Using the two-step coculture system of osteoblastic cells and marrow cells recovered from the semisolid methylcel- lulose cultures supported by CSFs, we found that all the CSFs stimulated the growth of osteoclast progenitors. Of the CSFs examined, M-CSF was the most potent in induc- ing osteoclast-like TRAP-positive MNCs, followed by GM-CSF, IL-3, and G-CSF in that order (Figs. 3 and 5 ) . ['zsII]Calcitonin specifically bound to TRAP-positive cells appearing in the cocultures. Our previous s t ~ d i e s ( ' ~ . ' ~ ) and those of o t h e r ~ ( ' ~ . ' ~ . ~ ~ ' indicated ' that the expression of cal-
fication of osteoclast differentiation in vitro. In this exper- iment, over 90% of the TRAP-positive mononuclear cells and MNCs accumulated ['zsl]calcitonin, indicating that those cells satisfy one of the most important criteria of osteoclasts.
The number of TRAP-positive MNCs formed in the two-step cocultures was clearly related to the number of NSE-positive cells formed in methylcellulose cultures (Table 2 and Fig. 5 ) . This suggests that the precursor cells of osteoclasts arise from NSE-positive cells of the mono- cyte-macrophage family and that M-CSF stimulates the de- velopment of their progenitor cells. This hypothesis is sup- ported by the finding that the cells recovered from the colonies, which were carefully isolated from the M-CSF- treated methylcellulose cultures, differentiated into osteo- clast-like cells in the two-step cocultures (Table 3). To de- termine whether a specific subpopulation in marrow cells treated with M-CSF differentiates into TRAP-positive cells, marrow cells were labeled with ('H)thymidine in methylcellulose cultures supported by M-CSF and then co- cultured with osteoblastic cells in the presence of la,25- (OH),D,. This attempt was unsuccessful, because colony formation in methylcellulose supported by M-CSF was strikingly inhibited by adding more than 0.05 pCi/ml of [3H]thymidine (specific activity, 80 Ci/mmol). At present, we cannot exclude the possibility that TRAP-positive MNCs are derived from the cells of colonies other than macrophage colonies, both of which are formed in re- sponse to M-CSF, although this is less likely.
We previously reported that the marrow-derived stromal cell line, ST2, could be substituted for primary osteoblastic cells in inducing osteoclast-like cells in cocultures with spleen cells in the presence of la,25-(OH),D, and dexa- methasone.(2') More recently, we found that not only mouse spleen cells but also mouse alveolar macrophages differentiate into TRAP-positive MNCs when they are co- cultured with ST2 cells in the presence of both hormones.("' The alveolar macrophage-derived TRAP- positive MNCs had calcitonin receptors and formed re- sorption lacunae on dentine slices.(zz1 Similarly, we found that mouse alveolar macrophages differentiated into TRAP-positive MNCs to the same extent as the M-CSF- treated marrow cells in the two-step cocultures: when 100 alveolar macrophages were cocultured for 8 days with pri- mary osteoblastic cells, many TRAP-positive MNCs (125
12 per well, mean f SEM of four cultures) were formed in response to la,25-(OH),D,. These results indi- cate that NSE-positive mature macrophages grow and dif- ferentiate into osteoclast-like MNCs in the presence of osteoblastic cells.
The importance of M-CSF in osteoclast development has been suggested by the experiments in osteopetrotic (op /op) mice.(23-zb' Wiktor-Jedrzejczak et al.(2J' first re- ported that the defect in op/op mice may be due to the failure of hematopoietic stromal cells that release sufficient amounts of M-CSF. Very recently, Yoshida et al.lzb) found a single base pair insertion in the coding region of the M-CSF gene in op/op mice. This mutation causes the failure in the production of functional M-CSF.'*b' We
citonin receptors is the most reliable marker for the identi- found that osteoblastic cells obtained from calvariae of

ROLE OF CSFs IN OSTEOCLAST FORMATION 983
t . *
FIG. 4. Autoradiographs of [1251]calcitonin binding in two-step cocultures of CSF-treated marrow cells and osteoblastic cells. Marrow cells isolated from the methylcellulose cultures supported by 50 units/ml of IL-3 (A), GM-CSF (B), M-CSF (C), or G-CSF (D) were cocultured for 8 days with osteoblastic cells in the presence of 10 nM lu,25-(OH),D.,. The cells were then incubated with 0.2 nM [1251]salmon calcitonin, stained for TRAP, and processed for autoradiography ( x 170). Note that dense grains due to the [1'51]calcitonin binding appear only on TRAP-positive cells.
op/op mouse d o not support osteoclast differentiation in cocultures with normal mouse spleen cells.'*') We also found that when a small number of mononuclear cells ob- tained from hematopoietic tissues were cultured on the ST2 cell layers, they formed single cell-derived colonies.'") All the colonies consisted of NSE-positive cells, and the TRAP-positive cells appeared in the colonies in response to la,25-(OH),D, and dexamethasone. I t has been re- ported that ST2 cells produce M-CSF spontaneously but d o not produce IL-3 and GM-CSF.l*') These findings indi- cate that the factor produced by ST2 cells and responsible for NSE-positive colony formation is M-CSF. Thus, M-CSF appears to be an important factor for the growth of osteoclast progenitors and a deficiency of M-CSF re- sults in osteopetrotic disorders in bone.
Why CSFs (GM-CSF, M-CSF, and IL-3) strongly inhib- ited osteoclast development in the one-step mouse marrow cultures is not known. The inhibition of osteoclast-like cell formation by M-CSF and IL-3 was much weaker when Ia,25-(OH),D, and the CSFs were simultaneously added to the coculture system (Table I ) . We previously reported that osteoclast-like marrow-derived stromal cells may be
involved in the osteoclast differentiation from its progeni- tors in mouse marrow cultures."" Treatment of marrow cells with IL-3, GM-CSF, or M-CSF increased the number of mononuclear phagocytes, but the growth of osteoblastic stromal cells appeared to be inhibited by the CSFs under microscopic observation. I t is therefore likely that the in- crease in the number of mononuclear phagocytes relative to that of osteoblastic stromal cells suppresses osteoclast differentiation. Alternatively, CSFs other than G-CSF may cause the precursor cycling without permitting their differ- entiation. I t is also possible that CSFs that stimulate differ- entiation of immature phagocytes into mature mac- rophages may directly inhibit their differentiation into osteoclasts, which is strictly regulated by osteoblastic cells. GM-CSF may be such a factor, because i t strikingly inhib- ited TRAP-positive MNC formation in cocultures of osteoblastic cells and freshly isolated marrow cells.
In conclusion, CSFs (IL-3, GM-CSF, hl-CSF, and G-CSF), especially M-CSF, are essential for the growth of osteoclast progenitors, but the CSFs other than G-CSF in- hibit their differentiation into osteoclast-like MNCs in- duced by la,25-(OH),D3 in the presence of osteoblastic

984 TAKAHASHI ET AL.
200
i o a
n 1
TABLE 3. EFFECTS OF Ia,2S-(OH)zD3 ON TRAP-POSITIVE MNC FORMATION AND CALCITONIN-DEPENDENT CAMP PRODUCTION IN TWO-STEP COCULTURES OF OSTEOBLASTIC
CELLS AND MARROW CELLS COLLECTED FROM THE COLONIES THAT WERE CAREFULLY ISOLATED FROM THE M-CSF-TREATED METHYLCELLULOSE CULTURES~
Production of CAMP (pmol per well)
TRAP-positive MNC formation Calcitonin (nM) (number of TRAP-positive
Treatment MNCs per well) 0 10
Vehicle 0 Ia,25-(OH)ZDJ 225 + 25
2.6 k 0.2 3.1 k 0.3 3.1 f 0.2 14.4 * 1.1
aBone marrow cells were first cultured for 6 days in semisolid methylcellulose in the presence of 50 unitslml of M-CSF. Each colony was individually isolated from the cultures with an Eppendorf pi- pette and pooled. The isolated marrow cells (200 cells per well) were then cocultured for 8 days with osteoblastic cells in the presence or absence of 10 nM la,25-(OH),D,. Some cultures were then fixed and stained for TRAP, and the number of TRAP-positive MNCs was counted. The remaining cul- tures were incubated with or without 10 nM salmon calcitonin for 15 minutes, and the amount of cAMP in the cell layer was determined. The results are expressed as the means f SEM of four cul- tures.
T
T
L " IL-3 GM-CSF M-CSI F G-CSF Fresh
BM
FIG. 5. Estimation of the number of TRAP-positive MNCs formed from 200 marrow cells in two-step cocul- tures with primary osteoblastic cells. The increasing num- bers of freshly isolated bone marrow cells (fresh BM) or the marrow cells recovered from semisolid methylcellulose cultures supported by 50 units/ml of IL-3, GM-CSF, M-CSF, or G-CSF were cocultured with osteoblastic cells (lo4 cells per well) in the presence of 10 nM la,25- (OH),D,. The number of TRAP-positive MNCs formed from 200 marrow cells of each preparation was estimated from linear plots between the number of TRAP-positive MNCs formed and the number of marrow cells cocultured with osteoblastic cells as shown in Fig. 3. The results are expressed as the means & SEM of three independent ex- periments consisting of different preparations of marrow cells and osteoblastic cells.
cells. Further studies are needed to elucidate t..: mecha- nism of the action of CSFs and the role of osteoblastic cells in regulating osteoclast differentiation and other mye- loid cell differentiation.
ACKNOWLEDGMENTS
We are grateful to Dr. K. Motoyoshi, National Defense Medical College (Saitama, Japan), for his helpful discus- sion of the role of CSFs. This work was supported by Grants-in-Aid (0148043 and 01870078) from the Ministry of Science, Education and Culture of Japan.
REFERENCES
1. Mundy GR, Roodman G D 1987 Osteoclast ontogeny and function. Bone Miner Res 9209-281.
2. Nijweide PJ , Burger EH. Feyen JHM 1986 Cells of bone: Proliferation, differentiation, and hormonal regulation. Physiol Rev 66:855-886.
3. Nicola NA, Vadas M 1987 Hemopoietic colony stimulating factors. lmmunol Today 5:76-81.
4. Sieff C 1987 Hemopoietic growth factors. J Clin Invest 79:
5 . Burger EH, Van der Meer JWM, Nijweide PJ 1984 Osteo- clast formation from mononuclear phagocytes: Role of bone-forming cells. J Cell Biol 99:190-1906.
6. Scheven BAA, Visser JWM, Nijweide PJ 1986 In vitro osteo- clast generation from different bone marrow fractions, in- cluding a highly enriched haematopoietic stem cell popula- tion. Nature 321:79-81.
7. Kurihara N, Suda T, Miura Y, Nakauchi H, Kodama H, Hiura K, Hakeda Y, Kumegawa M 1989 Generation of osteoclasts from hematopoietic progenitor cells. Blood 74: 1295-1302.
8. Lorenzo JA, Sousa SL, Fonseca JM, Hock JM, Medlock ES 1987 Colony-stimulating factors regulate the development of multinucleated osteoclasts from recently replicated cells in vitro. J Clin Invest 80:160-164.
1549- 1557.

ROLE OF CSFs IN OSTEOCLAST FORMATION 985
9. MacDonald BR, Mundy GR, Clark S. Wang EA, Kuehl TJ , Stanley ER, Roodman G D 1986 Effects of recombinant CSF- GM and highly purified CSF-1 on the formation of multinu- cleated cells with osteoclast characteristics in long-term bone marrow cultures. J Bone Miner Res 1:227-233.
10. Barton BE, Mayer R 1989 IL-3 induces differentiation of bone marrow precursor cells to osteoclast-like cells. J Immu- no1 143:3211-3216.
11. Hattersley G, Chambers TJ 1990 Effects of interleukin 3 and of granulocyte-macrophage and macrophages colony stimu- lating factors on osteoclast differentiation from mouse hemopoietic tissue. J Cell Physiol 142:201-209.
12. Shinar DM, Sato M, Rodan GA 1990 The effect of hemopoi- etic growth factors on the generation of osteoclast-like cells in mouse bone marrow cultures. Endocrinology 126:
13. Takahashi N. Yamana H. Yoshiki S, Roodman GD, Mundy GR, Jones SJ. Boyde A, Suda T 1988 Osteoclast-like cell for- mation and its regulation by osteotropic hormones in mouse marrow cultures. Endocrinology 122:1373-1382.
14. Takahashi N, Akatsu T. Udagawa N, Sasaki T, Yamaguchi A, Moseley JM, Martin TJ, Suda T 1988 Osteoblastic cells are involved in osteoclast formation. Endocrinology 123: 2600-2602.
IS. Akatsu T, Takahashi N. Udagawa N, Sato K , Nagata N, Moseley JM, Martin TJ , Suda T 1989 Parathyroid hormone (PTH)-related protein is a potent stimulator of osteoclast- like rnultinucleated cell formation to the same extent as PTH in mouse marrow cultures. Endocrinology 12920-27.
16. Akatsu T, Takahashi N , Debari K , Morita 1, Murota S, Na- gata N, Takatani 0. Suda T 1989 Prostaglandins promotes osteoclastlike cell formation by a mechanism involving cyclic adenosine 3’,5’-monophosphate in mouse bone marrow cul- tures. J Bone Miner Res 4:29-35.
17. Li CY, Lam KW. Yam LT 1973 Esterases in human leuko- cytes. J Histochem Cytochem 21:l-12.
IS. Takahashi N, Akatsu T. Sasaki T, Nicholson GC. Moseley JM, Martin TZJ, Suda T 1988 Induction of calcitonin recep- tors by la,25-dihydroxyvitamin D, in osteoclast-like multi- nucleated cells formed from mouse bone marrow cells. Endo- crinology 123:1504-1510.
19. Hattersley G, Chambers TJ 1989 Calcitonin receptors as markers for osteoclastic differentiation: Correlation between generation of bone-resorptive cells and cells that express cal- citonin receptors in mouse bone marrow cultures. Endocri- nology 125: 1606- 161 2.
20. Nicholson GC, Moseley JM, Sexton PM, Mendelsohn FAO, Martin TJ 1986 Abundant calcitonin receptors in isolated rat
1728-1735.
osteoclasts. Biochemical and autoradiographis characteriza- tion. J Clin Invest 78:355-360.
21. Udagawa N, Takahashi N, Akatsu T, Sasaki T, Yamaguchi A, Kodama H , Martin TJ , Suda T 1989 The bone marrow- derived stromal cell lines MC3T3-G2/PA6 and ST2 support osteoclast-like cell differentiation in cocultures with mouse spleen cells. Endocrinology 125:1805-1813.
22. Udagawa N, Takahashi N, Akatsu T, Tanaka H, Sasaki T, Nishihara T, Koga T, Martin TJ , Suda T 1990 The origin of osteoclasts: Mature monocytes-macrophages are capable of differentiating into osteoclasts under a suitable microenvi- ronment prepared by bone marrow-derived stromal cells. Proc Natl Acad Sci USA 87:7260-7264.
23. Wiktor-Jedrzejczak W , Ahmed A, Szczylik C, Skelly RR 1982 Hematological characterization of congenital osteope- trosis in op/op mouse. Possible mechanism for abnormal macrophage differentiation. J Exp Med 156:1516-1527.
24. Felix R. Cecchini MG, Hofstetter W, Elford PR, Stutzer A, Fleisch H 1990 Impairment of macrophage colony-stimu- lating factor production and lack of resident bone marrow macrophages in the osteopetrotic op/op mouse. J Bone Miner Res 5:781-789.
25. Wiktor-Jedrzejczak W, Bartocci A, Ferrante Jr AW, Ahmed-Ansari A, Sell KW, Pollard JW, Stanley ER 1990 Total absence of colony-stimulating factor 1 in the macro- phage-deficient osteopetrotic (op/op) mouse. Proc Natl Acad Sci USA 87:4828-4832.
26. Yoshida H, Hayashi S. Kunisada T, Ogawa M, Nishikawa S, Okamura H, Sudo T, Shultz LD, Nishikawa S 1990 The mu- rine mutation osteopetrosis is in the coding region of the macrophage colony stimulating factor gene. Nature 345:442- 444.
27. Takahashi N. Udagawa N, Akatsu T, Tanaka H , Isogai Y, Suda T 1991 Deficiency of osteoclasts in osteopetrotic mice is due to a defect in the local microenvironment provided by osteoblastic cells. Endocrinology 128:1792-1796.
Address reprint requests to: D r . Tatsuo Suda
Department of Biochemistry School of Dentistry
Sho wa University 1-5-8 Hatanodai
Shinagawa-ku, Tokyo 142, Japan
Received for publication August 21, 1990; in revised form April 2, 1991; acdepted April 2, 1991.





























