Embed Size (px)
Citation preview

International Journal of Pancreatology, 2 (1987) 223-235 Elsevier
PAN 0078
223
Role of secretin and CCK in the stimulation of pancreatic secretion in conscious dogs. Effects of
atropine and somatostatin
Stanislaw J. Konturek, Jan. W. Konturek, Cornelis B. Lamers,
Janine Tasler and Jan Bilski
Institute qf Physiology, Academy o/ Medicine, Krakow, Poland, and Department qf Gastroenterology, University Hospital, Leiden, The Netherlands
(Received 4 August 1986; Accepted 10 February 1987)
Key words: Secretin; CCK; Gastrin; Pancreatic secretion; Somatostatin; Atropine
Summary
The role of gut hormones, such as secretin and CCK, in the stimulation of pan- creatic secretion by duodenal HC1 or oleate and by meat feeding has been studied in conscious dogs before and after pretreatment with atropine and somatostatin. Plasma hormones were measured by specific and sensitive radioimmunoassays. Duodenal perfusion with HC1 and oleate stimulated dose-dependently pancreatic HCO3 and protein secretion and raised plasma levels of secretin and CCK, respectively. Atropine reduced significantly both HCO3 and protein secretion but did not affect plasma secretin and CCK levels in these studies. Both exocrine pancreatic secretion and plasma secretin and CCK levels were suppressed by somatostatin. Meat feeding caused a marked pancreatic HCO3 and protein secretion accompanied by a signifi- cant increase in plasma secretin and CCK which seem to play an important role in the postprandial pancreatic stimulation. Both atropine and somatostatin reduced the pancreatic secretion induced by exogenous hormones but only somatostatin, but not atropine, significantly decreased plasma secretin and CCK responses to intestinal stimulants. We conclude that both atropine and somatostatin reduce the pancreatic responses to duodenal HC1 or oleate or to meat feeding but only somatostatin is capable of suppressing the release of secretin and CCK.
Correspondence to: Prof. Dr. S.J. Konturek, Institute of Physiology, 31-531 Krakow, ul, Grzegorzecka 16, Poland.
0169-4197/87/$03.50 �9 1987 Elsevier Science Publishers B.V. (Biomedical Division)

224
Introduction
It is generally accepted that the postprandial pancreatic secretion results from the interplay of the stimulatory and inhibitory factors of neural and hormonal nature [1]. Pavlov and his pupils [2,3] thought that the vagal reflex mechanism plays a major role in the pancreatic response to a meal. After the discovery of gut hormones such as secretin [4] and cholecystokinin [5], the concept of the hormonal regulation of pancreatric secretion has been postulated though Popielski [6] claimed that short nervous reflexes between the gut and the pancreas may also be involved in the stimu- lation of exocrine pancreas.
With the successful development of radioimmunoassay of gut hormonal peptides, the determination of the role of these hormones and their interaction with vagal- cholinergic component in the pancreatic stimulation became possible. The picture of the overall control of exocrine pancreas has been completed by the discovery of the peptides inhibiting pancreatic secretion such as somatostatin [7,8] and pancreatic polypeptide (PP) [9].
The contribution and the relative importance of the above neural and hormonal factors have been little studied. The purpose of this paper was (1) to evaluate the importance of the gut hormones, particularly secretin and CCK, in the pancreatic responses to duodenal perfusion with HC1 or oleate and to a meat feeding, and (2) to determine the role of the cholinergic component and somatostatin in the release and action of these hormones on exocrine pancreas.
Methods
Six mongrel dogs (16-22 kg) were prepared with a Thomas-type gastric fistula (GF) and a modified Herrera-type pancreatic fistula (PF) as described before [10]. At least 4-5 months were allowed for the full recovery from the surgery before the start of experiments. The same 6 dogs were used throughout the study. The dogs were fasted 18 h before each experiment, the GF was opened (except in tests with feeding) throughout the experiment to prevent gastric acid from entering the duo- de'num and releasing intestinal hormones.
Secretion from the PF was continuously collected in 15-rain portions. The samples of the juice were taken for determination of HCO3 and protein concentrations and outputs as described previously [10]. The rest of the juice was reinfused into the duodenum via the hollowed obturator of the PF.
All perfusates were prepared on the day of the experiment, using HCI solution and oleic acid (Sigma Chemical Co., St Louis, MO) made as sodium soap [11] in concentrations ranging from 6.5 to 200 mM. These solutions were prepared iso-os- motically (300 + 10 mOsm/kg) by adding NaC1 as needed and were instilled intra- duodenally through the hollow obturator of the cannula of the PF at a constant rate of 80-160 ml/h.

225
In tests with feeding, dogs were given a meat meal (ground beef 20 g/kg body weight). The GF was closed, while the PF was open and the pancreatic juice was collected, the samples were taken for HCO3 and protein determination and the rest was reinfused into the duodenum.
In tests with exogenous hormones, secretin and CCK (containing mainly CCK- 33) were highly purified natural hormonal preparations (purchased from Kabi Diag- nostica, Stockholm, Sweden), and used as fresh solutions containing 1% (w/v) canine albumin (Sigma Chemical Co., St. Louis, MO) to prevent adsorption of the peptides to plastic tubes. They were infused intravenously either in graded doses, doubled every 30 min during the same test day, or in a constant dose being administered for 180 min as a single dose given per day. Somatostatin (gift of Dr. A.V. Schally, New Orleans, LA) was pure synthetic cyclic somatostatin-14 used in a solution containing 1% canine albumin. It was infused intravenously at a constant dose (300 pmol/kg-h) during mid-hour of pancreatic stimulation by intravenous secretin or CCK, and duo- denal perfusion with HC1 or oleate and meat feeding. In some studies, atropine (Pol- fa, Poland) was given as an i.v. bolus of 25 #g/kg injected in the mid-hour of pan- creatic stimulation. In control tests, pancreatic responses to intravenous secretin or CCK and duodenal instillation of HC1 or oleate or meat feeding were determined for the time of the experiment.
Radioimmunoassay of plasma hormones
Blood samples (5 ml) were collected in chilled glass tubes containing 500 U of aprotonin (Novo Alle Bagsvard, Denmark) and 50 U ofheparin (Polfa, Poland). The plasma was separated within 15 rain, frozen and stored at -20~ until the radioim- munoassays of secretin, CCK, gastrin and PP. Blood samples were taken twice under basal conditions and at 15-30-rain intervals after the start of exogenous hormone infusion or duodenal instillation of HC1 or oleate or meat feeding.
Plasma secretin was extracted with ethanol and determined using the technique described by Schaffalitzky de Muckadell and Fahrenkrug [12]. Radioiodinated se- cretin (kindly provided by Dr. O.B. Schaffalitzky de Muckadell) was prepared from synthetic porcine secretin that was purified by ion exchange as described [12]. Anti- serum 5595-3 (gift of Dr. Schaffalitzky de Muckadell) was used. The detection limit of the assay was 0.5 pM. Intra-assay and inter-assay variations were about 9% and 14%, respectively.
Plasma CCK concentration was measured in plasma extract using antibody T-204 that binds to all carboxyl-terminal CCK peptides containing a sulfated thyrosine region as described [13,14]. The detection limit of the assay was 0.3 pM. The intra- assay and inter-assay variations were about 7% and 17%, respectively.
Plasma gastrin was determined using antibody 4560 (gift of Dr. J. Rehfeld, Co- penhagen, Denmark) and plasma PP was measured using antibody Lilly Lot 615- R110-146-10 (gift of Dr. R.E. Chance, Indianapolis, IN) as described before [15-17]. All samples of plasma hormones were measured in triplicate.

226
Calculations
Pancreatic secretory outputs and plasma hormone concentrations were expressed as means 4- SEM. In tests with gradually increasing doses of secretin or CCK or with increasing loads of duodenal HC1 or oleate, the increments in plasma secretin or CCK and in pancreatic secretory outputs from the baseline values were calculated and averaged to provide the incremental concentrations of plasma hormones and incremental secretory outputs for the experimental period. The significance of the difference between means was evaluated using Student's t test for paired values. Dif- ferences were considered significant if P was less than 0.05.
Results
In fasted dogs with GF and PF, the basal pancreatic HCO3 output averaged 0.04 4- 0.01 mmol/30 rain and the protein output averaged 140 4- 32 mg/30 min. Basal concentration of secretin was about 2.1 + 0.2 pM, basal CCK was 0.6 4- 0.2 pM and basal PP was 22 4- 4 pM in these tests.
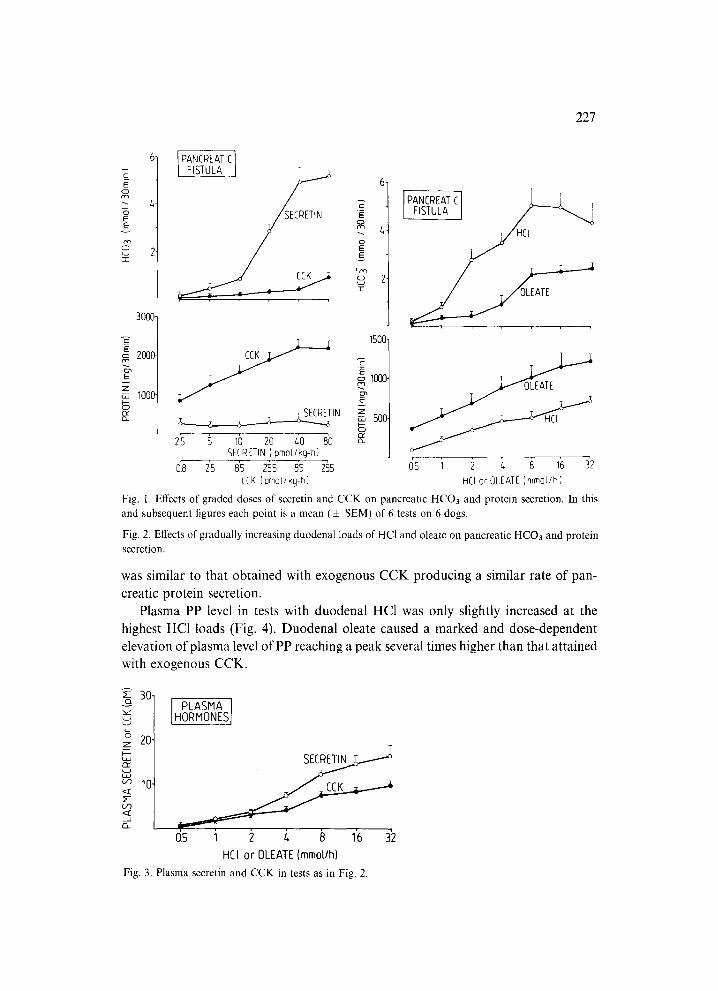
Secretin and CCK infused i.v. in graded doses induced a dose-dependent increase in pancreatic bicarbonate and protein secretion, respectively. The maximal HCO3 output was achieved by secretin infused at a dose of 80 pmol/kg-h and it averaged about 5.04 + 0.57 mmol/30 min. The maximal protein response was obtained by CCK infused at a dose of 85 pmol/kg-h and it averaged about 2200 4- 100 mg/30 min. Secretin failed to increase significantly the pancreatic protein secretion while CCK caused a small but significant elevation in HCO3 secretion observed only at the highest dose of the peptide (Fig. 1).
Duodenal perfusion with HC1 and sodium oleate in gradually increasing amounts resulted in a dose-dependent increment in both pancreatic HCO3 and protein secre- tion, duodenal HC1 being the relatively more potent stimulant of HCO3, whereas duodenal oleate the more potent stimulant of protein secretion (Fig. 2). The maximal HCO3 response was attained at 8 mmol HC1/h and amounted to about 5.10 + 0.72 mmol/30 min. This value was not significantly different from that obtained with exogenous secretin in these animals. The maximal protein response to duodenal oleate was attained at 16 mmol/h and it averaged about 1220 :t: 140 mg/30 min. This value amounted to about 60% of that obtained with exogenous CCK.
Plasma secretin in tests with duodenal HC1 and plasma CCK in experiments with duodenal oleate are shown on Fig. 3. Both duodenal HC1 and oleate raised dose- dependently plasma hormones, the highest concentrations of secretin and CCK being achieved at the highesl~ duodenal loads of HC1 and oleate, respectively. Plasma se- cretin level in tests with duodenal acidification was not significantly different from that recorded in tests with exogenous secretin producing a similar rate of pancreatic HCO3 secretion in these anirrvals. Plasma CCK level obtained with duodenal oleate
227
A
C
o
E~
'G
3000
C
2000
E v
Z ,T, 1000
(3-
PANCREATIC
SECRETIN E N 4
IOm
i Z 1500- CCK ~ -E
o 1000.
~ l N z ,T, 500. C~
25 5 10 20 L~0 80 o_ SECRETIN ( pmol/kg-h}
[ PAI%~'I~EL~AT I s
T OLEATE
0"8 25 8 S 255 85 255 05 1 2 L~ 8 16 32 CCK (pmoI/kg-h) HCi or OLEATE (mmol/h)
Fig. l. Effects of graded doses of secretin and CCK on pancreatic HCO3 and protein secretion. In this and subsequent figures each point is a mean (+ SEM) of 6 tests on 6 dogs.
Fig. 2. Effects of gradually increasing duodenal loads of HC1 and oleate on pancreatic HCO3 and protein secretion.
was similar to that obtained with exogenous CCK producing a similar rate of pan- creatic protein secretion.
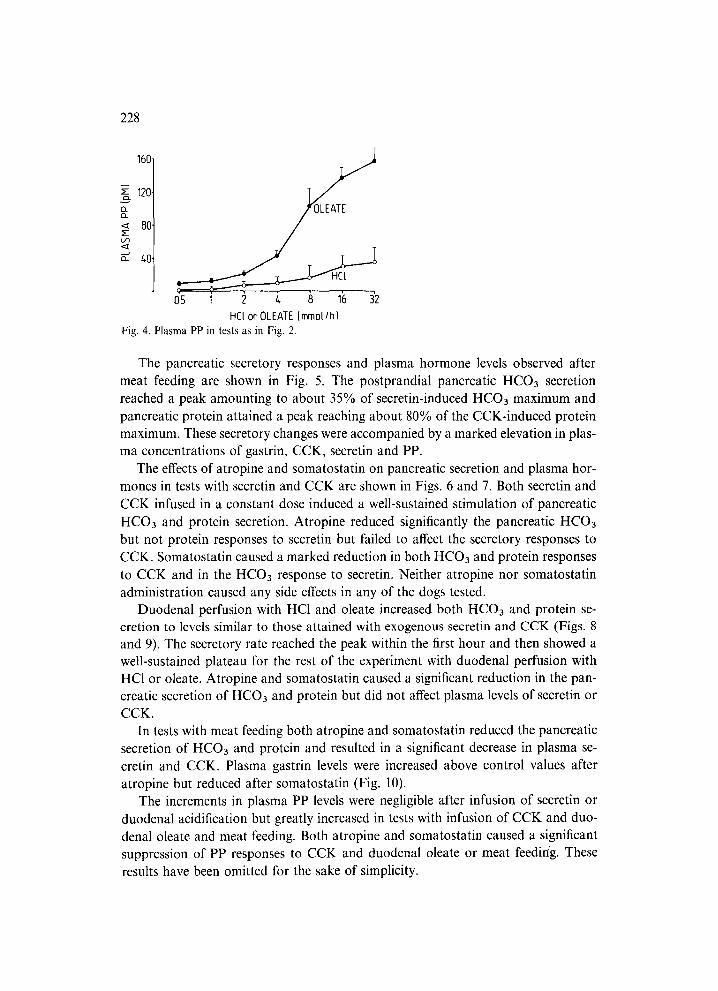
Plasma PP level in tests with duodenal HC1 was only slightly increased at the highest HCI loads (Fig. 4). Duodenal oleate caused a marked and dose-dependent elevation of plasma level of PP reaching a peak several times higher than that attained with exogenous CCK.
~-I 30 I
~ 20- 7
I ' - - I - u
t . J
m 10
PLASMA] HORMONES I
0.5 1 2 t+ 8 16 32
HC[ or OLEATE (mmol/h)
Fig. 3. Plasma secretin and CCK in tests as in Fig. 2.
228
160-
z 120"
13..
< 80- >--
s
OI 5 1 2 4 8 1~) 32
HCl or OLEAIE Immol/h) Fig. 4. Plasma PP in tests as in Fig. 2.
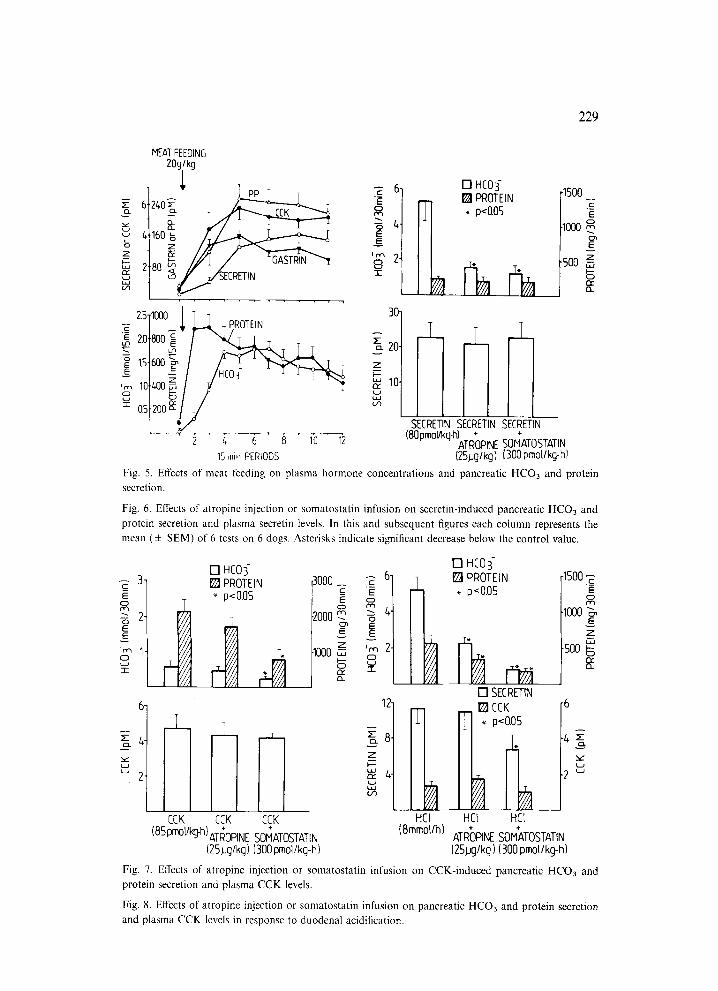
The pancreatic secretory responses and plasma hormone levels observed after meat feeding are shown in Fig. 5. The postprandial pancreatic HCO3 secretion reached a peak amounting to about 35% of secretin-induced HCO3 maximum and pancreatic protein attained a peak reaching about 80% of the CCK-induced protein maximum. These secretory changes were accompanied by a marked elevation in plas- ma concentrations of gastrin, CCK, secretin and PP.
The effects of atropine and somatostatin on pancreatic secretion and plasma hor- mones in tests with secretin and CCK are shown in Figs. 6 and 7. Both secretin and CCK infused in a constant dose induced a well-sustained stimulation of pancreatic HCO3 and protein secretion. Atropine reduced significantly the pancreatic HCO3 but not protein responses to secretin but failed to affect the secretory responses to CCK. Somatostatin caused a marked reduction in both HCO3 and protein responses to CCK and in the HCO3 response to secretin. Neither atropine nor somatostatin administration caused any side effects in any of the dogs tested.
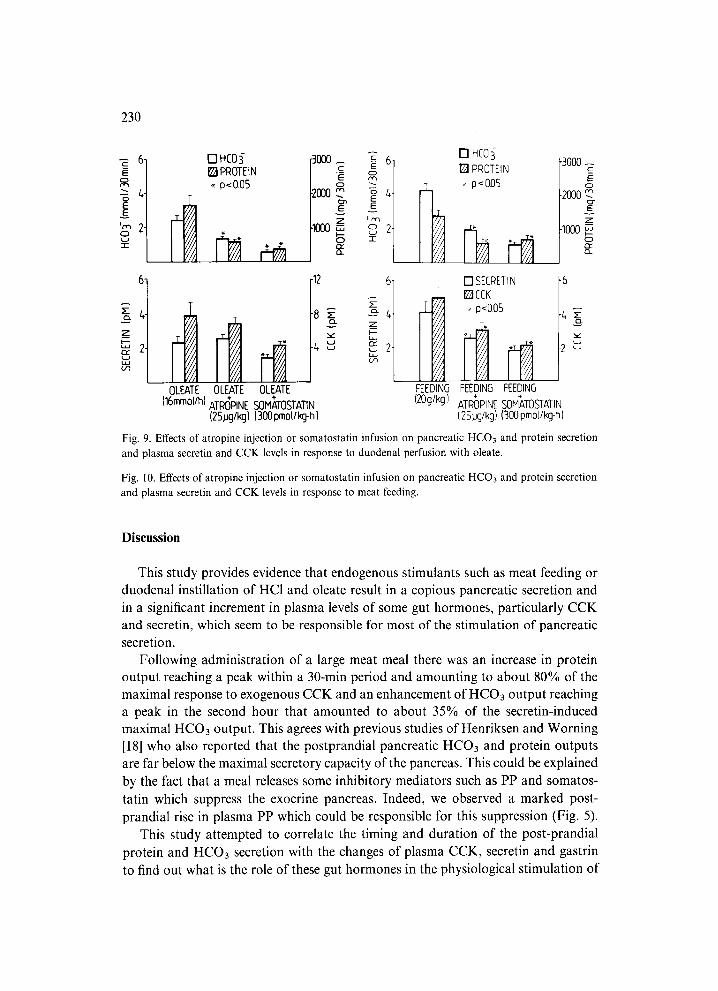
Duodenal perfusion with HC1 and oleate increased both HCO3 and protein se- cretion to levels similar to those attained with exogenous secretin and CCK (Figs. 8 and 9). The secretory rate reached the peak within the first hour and then showed a well-sustained plateau for the rest of the experiment with duodenal perfusion with HC1 or oleate. Atropine and somatostatin caused a significant reduction in the pan- creatic secretion of HCO3 and protein but did not affect plasma levels of secredn or CCK.
In tests with meat feeding both atropine and somatostatin reduced the pancreatic secretion of HCO3 and protein and resulted in a significant decrease in plasma se- cretin and CCK. Plasma gastrin levels were increased above control values after atropine but reduced after somatostatin (Fig. 10).
The increments in plasma PP levels were negligible after infusion of secretin or duodenal acidification but greatly increased in tests with infusion of CCK and duo- denal oleate and meat feeding. Both atropine and somatostatin caused a significant suppression of PP responses to CCK and duodenal oleate or meat feeding. These results have been omitted for the sake of simplicity.
229
MEAT FEEDING 20g/kg
E 6 240 "~
~ t <~ 7 " ~ ~' 4 160 ~ �9
_z ~ ' ~ 2- 2 80 r,r- - r -
,,ooo
i / "i ~| -~~ / 7
:c 0.5 m
2 z,, 6 8 10 12 15 rain PERIODS
[] HC03- [] PROTEIN * p<0.05
-1500 c
E
.1000 E
Z 500 m
PINE SOMATOSTATIN {25,ug/kgl (300 pmo[/kg-hl
Fig. 5. Effects of meat feeding on plasma hormone concentrations and pancreatic HCO3 and protein secretion.
Fig. 6. Effects of atropine injection or somatostatin infusion on secretin-induced pancreatic HCO3 and protein secretion and plasma secretin levels. In this and subsequent figures each column represents the mean ( + SEM) of 6 tests on 6 dogs. Asterisks indicate significant decrease below the control value.
- - 3- r-
E
o 2 E E
O
u
[ ] HCO]- [ ] PROTEIN "' p< 0,05
3000_ ~ 6. ._c E= E O
O ?000 m ~-- 4
o~
z '~, 2 I000 o
l T IIIT CCK CCK CCK
4"
(85 pm~ ATROPINE SOMATOSTATIN (25 LJg/kg) (300 pmol/kg-h)
12-
~ 8 - Z I I
54-
HEr (8retool/h)
[] HCOi [ ] PROTEIN
. p<0.05
[ ] SECRETIN I~CCK
T ~ p<0.05
HE1 HC[
AT ROPINE SOMATOSTATIN (25#g/kg) (300 pmol/kg-h)
-1500
-1000 ~ E
Z
-5oo S E
-6
-~, :E
2 ~
Fig. 7. Effects of atropine injection or somatostatin infusion on CCK-induced pancreatic HCO3 and protein secretion and plasma CCK levels.
Fig. 8. Effects of atropine injection or somatostatin infusion on pancreatic HCO3 and protein secretion and plasma CCK levels in response to duodenal acidification.
230
- - 6- c_ E RR -- t,- o E
I~ 2" o " r
z:z, . w
I - -
D HC03- [] PROTEIN . p<0.05
3000 _ .c_ E
.2ooo ~n E
z .lOOO
-12
OLEATE
4
OLEATE OLEATE (16mmol/h) ATR(3PINE SOM~,TOSTATIN
(2%g/kg) [300 pmol/kg-h)
Irv~ 02- -r-
6-
E
Z
~-~ 2"
FEEDING (20g/kg)
[] HC0 [] PROTDN
I~' p ( 0.05
3000 c__ E
o 2000
1000 n," O-
[ ] SECRETIN raCCK
p<0.05
FEEDING FEEDING ATR~)PINE SOMATOSTATIN (2.5ug/kg) (300 pmotlkg-h)
"6
-2
Fig. 9. Effects of atropine injection or somatostatin infusion on pancreatic HCO3 and protein secretion and plasma secretin and CCK levels in response to duodenal perfusion with oleate.
Fig. 10. Effects of atropine injection or somatostatin infusion on pancreatic HCO3 and protein secretion and plasma secretin and CCK levels in response to meat feeding.
DiscusSion
This study provides evidence that endogenous stimulants such as meat feeding or duodenal instillation of HC1 and oleate result in a copious pancreatic secretion and in a significant increment in plasma levels of some gut hormones, particularly CCK and secretin, which seem to be responsible for most of the stimulation of pancreatic secretion.
Following administration of a large meat meal there was an increase in protein output reaching a peak within a 30-min period and amounting to about 80% of the maximal response to exogenous CCK and an enhancement of HCO3 output reaching a peak in the second hour that amounted to about 35% of the secretin-induced maximal HCO3 output. This agrees with previous studies of Henriksen and Worning [18] who also reported that the postprandial pancreatic HCO3 and protein outputs are far below the maximal secretory capacity of the pancreas. This could be explained by the fact that a meal releases some inhibitory mediators such as PP and somatos-
tatin which suppress the exocrine pancreas. Indeed, we observed a marked post- prandial rise in plasma PP which could be responsible for this suppression (Fig. 5).
This study attempted to correlate the timing and duration of the post-prandial
protein and HCO3 secretion with the changes of plasma CCK, secretin and gastrin to find out what is the role of these gut hormones in the physiological stimulation of

231
exocrine pancreas. It was found that the initial increase in pancreatic protein secretion that occurred soon after feeding was unrelated to the plasma CCK or gastrin, which peaked in the 'second postprandial hour. Thus neither of these hormones could be responsible for the initial stimulation of the pancreas. It is likely that the vagal re- flexes activated during the combined cephalic and gastric phases are the major stimu- lants of the initial postprandial increase in pancreatic protein secretion. This is sup- ported by previous findings showing a stimulation of canine pancreatic protein se- cretion by the process of sham-feeding [17,19], and gastric distention [20].
Unlike the initial phase, the later postprandial phase of pancreatic secretion ap- pears to be driven mostly by gut hormones. It is evidenced by our results showing that the increments in plasma CCK in that later postprandial period was closely related to the pancreatic protein secretion (Fig. 5) and the rise in plasma CCK after a meal was similar to that observed after exogenous CCK (85 pmol/kg-h) producing a similar rate of pancreatic protein secretion (Figs. 5 and 7). Plasma gastrin also showed a marked rise but this may not be sufficient to stimulate the pancreatic protein to the level occurring postprandially [21]. Thus we have evidence that out of the gut peptides released by a meal, CCK seems to play a dominant role as a mediator of pancreatic secretion in the late postprandial period that corresponds to the intestinal phase of pancreatic secretion [22]. This conclusion disagrees, in part, with previous findings of Solomon and his colleagues [23,24] in dogs with denervated autotrans- planted pancreas. They postulated that the intestinal stimuli predominantly activate a neural pathway (an entero-pancreatic reflex) of pancreatic stimulation, but plasma hormones, particularly CCK, have not been measured in these studies.
The postprandial increase in HCO3 secretion was better correlated with plasma secretin levels than protein secretion with plasma CCK levels. Both HCO3 secretion and plasma secretin reached peaks in the later postprandial period and this probably corresponded to the delivery of acidified gastric contents to the duodenum. The over- all increment in plasma secretin after feeding was relatively low and by comparison with that achieved by exogenous secretin at a dose of 80 pmol/kg-h (Figs. 5 and 6) it could contribute to less than 50% of the pancreatic HCO3 secretion. The difference between the magnitude of the pancreatic HCO3 secretion in response to endogenous (meal-stimulated) and exogenous secretin could be explained by the fact that, in addition to secretin, the meat meal also releases CCK which interacts which secretin in the stimulation of pancreatic secretion. Such a potentiating interaction between secretin and CCK [18] as well as between the intestinal phase stimulants [25,26] for pancreatic HCO3 secretion was reported before.
It is generally accepted that the major physiological stimulant of the release of secretin and CCK is the delivery of gastric acid and the protein or fat digests into the duodenum. This is why we performed separate tests with duodenal perfusion of HC1 and oleate with the aim to correlate plasma increments in gut hormones with the pancreatic secretory outputs. These studies showed that the incremental pan- creatic HCO3 outputs induced by duodenal acid and exogenous secretin (Figs. 6 and

232
8) were accompanied by the increments in plasma secretin levels. However, the HCO3 response to endogenous secretin (released by duodenal acid) was much larger than that to exogenous secretin. This indicates that endogenous secretin is an important but not the only mediator of the pancreatic HCO3 response to duodenal acid and that other hormones, mainly CCK [27], could contribute to the augmentation of secretin effects on the HCO3 producing cells. This is supported by the fact that the pancreatic protein secretion in tests with duodenal acid reached higher values than these obtained with exogenous secretin producing an equal rate of HCO3 secretion.
Duodenal perfusion with oleate in these studies was a highly effective stimulant of both protein and HCO3 secretion. The maximal protein response to duodenal oleate was similar to that obtained with a meat meal but smaller than the maximal response to exogenous CCK. It was accompanied by a marked and dose-dependent increment in plasma CCK reaching (at various duodenal oleate loads) levels similar to those obtained with exogenous CCK producing an equal rate of pancreatic protein secretion (Figs. 2, 3 and 7). This indicates that the major mediator of the oleate- induced pancreatic protein secretion is endogenous CCK [22]. Duodenal oleate also caused a marked stimulation of pancreatic HCO3 secretion that was accompanied by a small but significant increase in plasma secretin. When compared with exogenous secretin producing similar increments in plasma secretin, oleate induced several times higher pancreatic HCO3 outputs. This could be explained by the fact that oleate in the gut is also a potent releaser of other stimulants of HCO3 secretion such as neu- rotensin [28] and possibly VIP [29]. The interaction of oleate-released secretin with CCK also could add to the stimulation of HCO3 secretion to a higher rate than that expected from the increments in plasma secretin. Our exocrine pancreatic secretory data obtained with oleate are in keeping with previous studies showing a potent stimulatory action of intestinal oleate on exocrine pancreas [11]. Our study demon- strates a close correlation of the secretory results with plasma hormones, particularly CCK and secretin, that has not been demonstrated previously. A small difference between the oleate-induced secretion and the CCK-induced maximal secretory ca- pacity of the pancreas could be explained by the inhibitory action of PP released in large amounts by oleate.
The major finding of this study is the demonstration that the blockade of mus- carinic receptors with atropine, and thus the elimination of the cholinergic compo- nent from the pancreatic stimulation, reduced the secretory rate of exocrine pancreas but virtually failed to affect plasma levels of gut hormones such as CCK or secretin. The only exception was the postprandial release of gut hormones but this probably reflected the reduced gastric acid secretion and/or delayed gastric emptying resulting in the decreased exposure of the intestinal mucosa to digestive products releasing these hormones. Our finding that following atropinization the pancreas was still capa- ble of secreting HCO3 and protein is also evidence for the importance of the hor- monal mechanism in the pancreatic stimulation. Since atropine did not affect the plasma CCK or secretin responses to intestinal stimulants, such as acid or oleate, but

233
caused a marked reduction in the pancreatic responses to these stimulants, it is reasonable to assume that atropine mainly interfered with the action of endogenously released gut hormones on the pancreatic secretory cells. The suppression by atropine of the pancreatic responses to low doses of secretin [32] and CCK [33] was recognized before and this has been attributed to removal of the potentiation between these two hormones or to the blockade of the potentiating effect of intrapancreatic acetylcho- line on CCK- or secretin-induced excitation of exocrine pancreas [1]. An alternative explanation could be that atropine interfered with the cholinergic vago-vagal reflexes from the intestine to the pancreas as suggested by Solomon et al. [23,24].
The inhibitory action of somatostatin on exocrine pancreatic secretion induced by exogenous hormones or endogenous stimulants is well documented [34]. Somatos- tatin was found to suppress competitively the secretin-induced HCO3 secretion and CCK-induced protein secretion [8] suggesting the competition for the common re- ceptor sites between somatostatin and these hormones. Both somatostatin-14 and somatostatin-28 were identified and isolated from the intestinal mucosa [35,36] and both these molecular forms of somatostatin were effective inhibitors of pancreatic secretion [37,38].
Although somatostatin is a potent inhibitor of exocrine pancreatic secretion, it has not been determined whether this peptide at physiological dose can influence the release of secretion or CCK, or exocrine pancreatic secretion induced by physio- logical doses of secretin or CCK or by an ordinary meal. Plasma somatostatin was not measured in this study, but based on previous reports on plasma somatostatin responses to sham-feeding [39] and/or a peptone meal [40], we selected a relatively small dose (300 pmol/kg-h) which is within the amounts of the peptide released under physiological conditions. This dose was used against a constant background stimu- lation with meat feeding, duodenal perfusion of acid or oleate solution and with exogenous hormones, secretin and CCK inducing the pancreatic secretory rate ob- tained with intestinal stimulants. The most sensitive to somatostatin inhibition was found to be postprandial pancreatic secretion possibly because of the suppression of the release and action of both secretin and CCK. The fall in the postprandial pan- creatic secretion could also be attributed to the suppression by somatostatin of gastric acid secretion [38,39] and to the delayed gastric emptying of the acidified meal to the duodenum. The latter possibility does not seem to be the major mechanism of the inhibitory action of somatostatin on the exocrine pancreas because this hormone reduced also the pancreatic secretory responses to intestinal perfusion with HC1 or oleate, the major postprandial stimulants of the pancreas. Thus the inhibitory action was accompanied by a significant reduction in plasma secretin and CCK levels.
Since somatostatin was an effective inhibitor of the pancreatic response to ex- ogenous secretin or CCK it is obvious that this peptide acts on the exocrine pancreas by two complementary mechanisms, that is by suppressing the release of gut hor- mones and by direct action on the pancreatic secretory cells to interfere with the stimulatory effect of these hormones. It must be mentioned, however, that somatos-

234
tatin is not inhibitory in the preparation of the isolated pancreatic acini [41] sug- gesting that its action on the intact pancreas results from an indirect effect on the gland perhaps by reducing the release of intrapancreatic acetylcholine [42] or de- creasing blood flow through the organ [38,43].
References
1 Solomon, T.E., Regulation of pancreatic secretion, Clin. Gastroenterol., 1984; 13: 657-678. 2 Pavlov, I.P., The Work of the Digestive Glands, 2nd edn., translated by H. Thompson, Griffin, Lon-
don, 1910. 3 Dolinski, T.L., Influence of acids on the secretion of the pancreatic gland, Arch. Sci. Biol., 1894; 3:
399~420. (In Russian.) 4 Bayliss, W.M. and Starling, E.H., The mechanism of pancreatic secretion, J. Physiol. (Lond.), 1902;
28: 325-353. 5 Harper, A.A. and Raper, H.S., Pancreozymin, a stimulant of the secretion of pancreatic enzymes in
extracts of the small intestine, J. Physiol. (Lond.), 1943; 102: 115-125. 6 Popielski, L., Ueber das peripherische reflextorische Nervenzentrum des Pancreas, Pflfigers Arch. Ges.
Physiol., 1902; 86: 215-221. 7 Boden, G., Sivitz, M.C., Owen, O.E., Essa-Konmar, N. and Landor, J.H., Somatostatin suppresses
secretin and pancreas exocrine secretion, Science, 1975; 90: 163-164. 8 Konturek, S.J., Tasler, J., Obtulowicz, W., Coy, D.H. and Schally, A.V., Effect of growth hormone-
releasing inhibiting hormone on hormones stimulating exocrine pancreatic secretion, J. Clin. Invest., 1976; 58: 1-6.
9 Lin, T.M., Evans, D.C., Chance, R.E. and Spray, G.F., Bovine pancreatic peptide: action on gastric and pancreatic secretion in dogs, Am. J. Physiol., 1977; 232:E311 E315.
10 Konturek, S.J., Pucher, A. and Radecki, T., Comparison of vasoactive intestinal peptide and secretin in stimulation of pancreatic secretion, J. Physiol. (Lond.), 1976; 255:497 509.
11 Meyer, J.H. and Jones, R.S., Canine pancreatic responses to intestinally perfused fat and products of fat digestion, Am. J. Physiol., 1974; 226:1178 1187.
12 Schaffalitzky de Muckadell, O.B. and Fahrenkrug, J., Radioimmunoassay of secretin in plasma, Scand. J. Clin. Lab. Invest., 1977; 37: 155-162.
13 Jansen, J.B.M.J. and Lamers, G.W., Radioimmunoassay of cholecystokinin in human tissue and plas- ma, Clin. Chim. Acta, 1983; 131: 305-316.
14 Jansen, J.B.M.J. and Lamers, G.W., Radioimmunoassay and evaluation of antibodies, J. Clin. Chem. Biochem., 1983; 21: 387-394.
15 Konturek, S.J., Tasler, J., Mikos, E., Jaeger, E. and Wunsch, E., Comparison of biological activity and disappearance rates of synthetic big gastrin, little gastrin and minigastrin in the dog, Hepato- Gastroenterology, 1980; 27: 3540.
16 Swierczek, J.S., Pawlik, W. and Konturek, S.J., Organ removal and disappearance half-time of syn- thetic pancreatic polypeptide, Digestion, 1982; 25: 197-200.
17 Swierczek, J.S., Konturek, S.J., Tasler, J., Jaworek, J. and Cieszkowski, M., Pancreatic polypeptide and vagal stimulation of gastric and pancreatic secretion in dogs, Hepato-Gastroenterology, 1981; 28: 206-209.
18 Henriksen, F.W. and Worning, H., External pancreatic response to food and its relation to the maximal secretory capacity in dogs, Gut, 1969; 10: 209-214.
19 Preshaw, R.M., Cooke, A.R. and Grossman, M.I., Sham feeding and pancreatic secretion in the dog, Gastroenterology, 1966: 50:171 178.
20 Vagne, M. and Grossman, M.[., Gastric and pancreatic secretion in response to gastric distention in dogs, Gastroenterology, 1969; 57: 300-310.

235
21 Konturek, S.J. and Solomon, T.C., Does gastric play a role in cephalic phase of pancreatic secretion?, Dig. Dis. Sci., 1984; 29: 44S.
22 Konturek, S.J., Tasler, J., Bilski, J., Jansen, J.B.M.J. and Lamers, C.B., Physiological role and local- ization of cholecystokinin release in dogs, Am. J. Physiol., 1985; 250: G391-G397.
23 Singer, M.V., Solomon, T.E. and Grossman, M.I., Effect of atropine on secretion from intact and transplanted pancreas in dog, Am. J. Physiol., 1980; 238:GI8 CJ22.
24 Singer, M.V., Solomon, T.E. and Grossman, M.I., Latency of pancreatic enzyme response to intrad- uodenal stimulants, Am. J. Physiol., 1981; 240:G376 G380.
25 Fink, A.S., Luxemburg, M. and Meyer, J.H., Regionally perfused fatty acids augment acid-induced canine pancreatic secretion, Am. J. Physiol., 1983; 245: G78-G84.
26 Fink, A.S., Miller, J.C., Jehn, D.W. and Meyer, J.H., Digests of protein augment acid-induced canine pancreatic secretion, Am. J. Physiol., 1982; 242:G634 G641.
27 Chen, Y.F., Chey, W.Y., Chang, T.M. and Lee, K.Y., Duodenal acidification releases cholecystokinin, Am. J. Physiol., 1985; 249:G29 G33.
28 Konturek, S.J., Jaworek, J., Cieszkowski, M., Pawlik, W., Kania, J. and Blom, S.R., Comparison of effects of neurotensin and fat on pancreatic stimulation in dogs, Am. J. Physiol., 1983; 244:G590 G598.
29 Schaffalitzky de Muckadell, O.B., Fahrenkrug, J., Holst, J.J. and Lauritsen, K.B., Release ofvasoactive intestinal polypeptide (VIP) by intraduodenal stimuli, Scand. J. Gastroenterol., 1977; 12: 793-799.
30 Chey, W.Y., Kim, M.S. and Lee, K.Y., Influence of vagus nerve on release and action of secretin in dog, J. Physiol. (Lond.), 1979; 293:435 446.
31 Niebel, W., Singer, V., Hanssen, L.E. and Goebell, H., Effect of atropine on pancreatic bicarbonate output and plasma immunoreactive secretin in response to intraduodenal stimulants, Scand. J. Gas- troenterol., 1983; 18: 803-808.
32 Singer, M.V., Niebel, W., Hoffmeister, D. and Goebell, H., Dose-response effects of atropine on pancreatic response to low doses of secretin, Regul. Peptides, 1983; 2 (Suppl.): SI08.
33 Konturek, S.J., Tasler, J. and Obtulowicz, W., Effect of atropine on pancreatic responses to endogen- ous and exogenous cholecystokinin, Am. J. Dig. Dis., 1972; 17:911 917.
34 Arnold, R. and Lankisch, P.G., Somatostatin and the gastrointestinal tract, Clin. Gastroenterol., 1980;
9:733 754. 35 Conlon, J.M., Zyznar, E., Vale, W. and Unger, R.H., Multiple forms of somatostatin-like immuno-
reactivity in canine pancreas, FEBS Lett., 1978; 94: 327-330. 36 Pradayrol, L., Jornvall, H., Mutt, V. and Ribet, A., N-Terminally extended somatostatin: the primary
structure of somatostatin-28, FEBS Lett., 1980; 109: 55-58. 37 Vaysse, N., Chayvialle, J.A., Pradayrol, L., Esteve, J.P., Susini, C., Lapuelle, J., Descos, F. and Ribet,
A., Somatostatin 28: comparison with somatostatin 14 for plasma kinetics and low-dose effect on the exocrine pancreas in dogs, Gastroenterology, 1981; 81: 700-706.
38 Konturek, S.J., Tasler, J., Jaworek, J., Pawlik, W., Walus, K., Schusdziarra, V., Meyers, C.A., Coy, D.H. and Schally, A.V., Gastrointestinal secretory, motor, circulatory and metabolic effects of pre- somatostatin, Proc. Natl. Acad. Sci. USA, 1981; 78:1967 1971.
39 Seal, A., Yamada, T., Debas, H., Hollinshead, J., Osadchey, B., Aponte, G. and Walsh, J., Somatos- tatin-14 and 28: clearance and potency on gastric function in dogs, Am. J. Physiol., 1982; 243: G97- G102.
40 De-Graef, J. and Woussen-Colle, M.C., Effect of sham feeding, bethanechol, and bombesin on so- matostatin release in dogs, Am. J. Physiol., 1985; 248: G1-G7.
41 Singh, M., Effect of somatostatin on amylase secretion from in vivo and in vitro rat pancreas, Dig.
Dis. Sci., 1986; 31: 506~512. 42 Guillemin, R., Somatostatin inhibits the release of acetylcholine induced electrically in the myenteric
plexus, Endocrinology, 1976; 99: 1653-1654. 43 Samnegard, H., Thulin, L., Adreen, M., Tyder, G., Hallberg, D. and Efendic, S., Circulatory effects
of somatostatin in anesthetized dog, Acta Chit. Scand., 1979; 145: 209-212.














![[The beginnings of modern biological psychiatry in Hungary: the atropine coma. A historical overview]](https://img.pdfslide.net/doc/110x75/634db87c7b55c75a34073438/the-beginnings-of-modern-biological-psychiatry-in-hungary-the-atropine-coma-a.jpg)














