Embed Size (px)
Citation preview

Mar Biol (2010) 157:2511–2519
DOI 10.1007/s00227-010-1515-4ORIGINAL PAPER
Seasonal contrasts in abundance and reproductive parameters of Penilia avirostris (Cladocera, Ctenopoda) in a coastal subtropical area
Leonardo K. Miyashita · Mayza Pompeu · Salvador A. Gaeta · Rubens M. Lopes
Received: 24 April 2010 / Accepted: 6 July 2010 / Published online: 17 July 2010© Springer-Verlag 2010
Abstract We studied the population dynamics and thereproductive biology of Penilia avirostris during threeconsecutive years on the inner shelf oV Ubatuba, Brazil.Penilia avirostris individuals and its eggs and embryoswere counted, measured, and classiWed into stages. Thespecies occurred throughout the studied period, in a widetemperature range (14.8–28.2°C). Cladoceran densitieswere usually higher (>2,000 ind m¡3) in warm seasons,when the water column was stratiWed as a consequence ofbottom intrusions of the cold- and nutrient-rich SouthAtlantic Central Water. Juveniles, non-reproducing females,and parthenogenic females were the dominant develop-mental stages. Males and gamogenic females were rareand only occurred when females reached peak abundances.This suggests that in tropical and subtropical coastal seasgamogenesis in P. avirostris is not as common as intemperate seas, but may play a signiWcant role in thedensity-dependent control of the population precedingunfavourable periods.
Introduction
Copepods usually comprise 55–95% of metazooplanktonabundance in marine pelagic systems (Longhurst 1985), but
cladocerans may attain high densities in coastal and estua-rine environments, surpassing copepods as major grazers atcertain periods of the year (Grahame 1976). Cladoceranscan feed on a wide range of particle sizes (Atienza et al.2006a, 2007), from bacteria to large protozoans and smallmetazoans, contributing to the microbial food web (Turneret al. 1988; Lipej et al. 1997) and serving as food for uppertrophic levels (Liang and Vega-Pérez 1995; Nip et al.2003).
Among the currently accepted 620 species of cladocer-ans (Forró et al. 2008), only eight are truly marine (Onbé1977). They are arranged in two orders (Ctenopoda andOnychopoda) and Wve genera (Penilia, Evadne, Pseud-evadne, Podon, and Pleopis; Onbé 1999). The single repre-sentative of the order Ctenopoda in the oceans, Peniliaavirostris, is usually the most abundant and widely distrib-uted species, occurring in estuarine and neritic, temperate,subtropical, and tropical waters (Grahame 1976; Rocha1982; Yoo and Kim 1987; Onbé and Ikeda 1995; Tanget al. 1995; Marazzo and Valentin 2001; Rose et al. 2004;Atienza et al. 2008).
The cladoceran life cycle is unique among planktoniccrustaceans. As a rule, cladocerans reproduce asexually byparthenogenesis, at certain times they switch their repro-ductive mode to gamogenesis (sexual phase) and produceresting eggs (Onbé 1985; EgloV et al. 1997). These varyinglife cycle traits confer a high reproductive potential (Onbé1977), which allows a fast, population wide response toboth favourable and unfavourable environmental condi-tions (EgloV et al. 1997). The main trigger that induces thetransition from asexual to sexual reproduction is stillunknown (Onbé 1999); however, it is believed that a com-bination of factors (e.g. temperature, salinity, abundance,and photoperiod) is involved (Gyllström and Hansson2004).
Communicated by X. Irigoien.
L. K. Miyashita (&) · M. Pompeu · S. A. Gaeta · R. M. LopesDepartment of Biological Oceanography, Oceanographic Institute, University of São Paulo, Praça do OceanográWco 191, Cidade Universitária, São Paulo 05508-120, Brazile-mail: [email protected]
123

2512 Mar Biol (2010) 157:2511–2519
The ecological success of P. avirostris in coastal watersis attributed to its short developmental time and to parthe-nogenic reproduction (Atienza et al. 2007). It has been sug-gested that in warmer waters, gamogenesis is not essentialto maintain population stability throughout the year (DellaCroce and Venugopal 1973; Tang et al. 1995), but addi-tional information is needed in regard to P. avirostris repro-ductive biology in subtropical and tropical waters to answerthis question.
We studied the population dynamics and reproductivecharacteristics of P. avirostris in a subtropical coastal areato evaluate the main factors controlling its temporal vari-ability in abundance and reproductive mode from partheno-genesis to gamogenesis.
Methods
The temporal distribution and reproductive biology ofP. avirostris were studied in the Southeast coast of Brazil,for three consecutive years (July 2005 through June 2008)by monthly sampling (except September 2005) at a Wxedneritic station (23°36�S–44°58�W) located on the 40-m iso-bath and »12 nautical miles oV Ubatuba, state of São Paulo(Fig. 1). Zooplankton samples were collected during day-time (between 10:00 and 14:00 h) by vertical hauls of asingle opening–closing net (200-�m) with a 0.2 m2 moutharea, usually from 1 to 2 m above the seabed to the bottomof the thermocline or to the mid-depth and from this pointto the surface (except from May 2006 through September2006, when only one monthly sample covering the entirewater column was collected). Samples were preserved in4% buVered formaldehyde immediately after the tows.
Temperature and salinity data were obtained by verticalCTD casts. Water was collected at Wve depths (0, 5, 10, 25 and38 m) with Niskin bottles, and 296 ml were Wltered throughWhatman-GF/F Wlters for chlorophyll a (chl a) analysis byXuorometry (Welschmeyer 1994).
At least 160 individuals per sample were randomlysorted, counted with a stereomicroscope and classiWed intostages. These stages were juveniles (<525 �m), non-repro-ducing females, parthenogenic females (carrying embryos),gamogenic females (with resting eggs), and males. All thesamples were completely examined for gamogenic females,whereas males were inspected in smaller aliquots (up to 1/8of the total sample) because they are not as easily detectedas gamogenic females, which make the inspection of malesa much more laborious process. Embryonic developmentwas further divided into four diVerent stages based on eas-ily distinguished external characteristics of the embryos,according to the description of Della Croce and Bettanin(1965) as applied by Wong et al. (2004) and Atienza et al.(2008). Embryos were classiWed as follows:
Stage 1: starts with the appearance of ellipsoidal par-thenogenic eggs; at the end of this stage the head andthe second antennae are visible;Stage 2: begins with the appearance of the Wrst anten-nae; the second antennae starts to bifurcate, and at leastthe Wrst thoracic segment is observable;Stage 3: corresponds to embryos with the secondantennae fully developed and all thoracic segmentsbeing noticeable; andStage 4: represents embryos similar to adults, with car-apace, thoracic appendages, and eye fully developed.
Body length (BL, from the tip of the head to the base of thecaudal setae) from a maximum of 100 specimens per stageper sample (excepting juveniles and non-reproducingfemales, which were counted as a single group) was mea-sured using the ZoopBiom software and digitizing system(RoV and Hopcroft 1986). In each sample, resting eggs(length and width) and embryos (total length) from atleast 15 randomly sorted females per stage were carefullydissected, measured (only one embryo per female), andcounted. Only resting eggs on late stages (greyish, ovoidegg; Della Croce and Bettanin 1969) were measured. Totalabundance (D; individuals m¡3) was determined with theequation: D = N F¡1 V¡1, where N is number of organismscounted in the sample aliquot; F is the fraction size of thealiquot; and V is the volume of water Wltered by the plank-ton net.
The abundance, brood size (BS), BL, temperature, salin-ity, and chl a data were previously log(x + 1) transformedand then correlated through the Spearman’s rank correla-tion (rs); spatial and temporal diVerences were analysedwith the U test of Mann–Whitney (� = 0.05).Fig. 1 Sampling station oV Ubatuba, Brazil
123

Mar Biol (2010) 157:2511–2519 2513
Results
Hydrography and chlorophyll a
We observed two seasonal periods in the study area relatedto stratiWcation processes and the vertical distribution ofthermohaline properties. From mid-autumn to early spring(generally from May through September), the water columnwas well mixed, with only a slight vertical temperature gra-dient (not higher than 3°C; seasonal range of 19.3–25.0°C;Fig. 2) and salinity in the range of 31.1–36.0. The dominantwater mass during this period was the Coastal Water (CW;temperature >20°C and salinity <35). Conversely, the watercolumn was stratiWed from mid-spring to early autumn(generally from October through April), with temperaturevarying between 14.8 and 28.2°C (vertical gradient of up to12°C; Fig. 2) and salinity ranging from 33.5 to 35.7. Duringthe stratiWed period the South Atlantic Central Water(SACW; temperature <20°C and salinity <36.4) dominatedbelow the thermocline and CW above. The intensity andduration of SACW intrusions showed a clear seasonal pat-tern, but had interannual variations (Table 1).
The chl a vertical proWle was inXuenced by the distribu-tion pattern of water masses (Table 1). When only CW waspresent, chl a distribution was more vertically homoge-neous, with values usually lower than 1.3 mg m¡3, exceptin May and June 2008, when peak values ranging from 1.7to 2.5 mg m¡3 occurred (Fig. 3). During stratiWed periods,chl a concentrations were usually in the range of
Fig. 2 Multi-annual tempera-ture vertical proWles on a Wxed station oV Ubatuba, Brazil, from July 2005 through June 2008
Table 1 Seasonal variability in water mass distribution and the corre-sponding range of temperature and chlorophyll a (Chl a) oV Ubatuba,Brazil
CW Coastal Water, SACW South Atlantic Central Water
Water mass Period Temperature(°C)
Chl a(mg m¡3)
CW Jul 2005–Nov 2005 19.5–22.3 0.53–1.23
May 2006–Sept 2006 19.9–25.0 0.31–1.00
May 2007–Aug 2007 19.3–22.2 0.29–1.31
May 2008–Jun 2008 21.2–23.7 0.69–2.49
CW + SACW Dec 2005–Apr 2006 14.9–27.3 0.31–2.12
Oct 2006–Apr 2007 16.4–28.2 0.25–12.38
Sept 2007–Apr 2008 14.8–27.1 0.21–5.42
123

2514 Mar Biol (2010) 157:2511–2519
0.5–1.0 mg m¡3 in the upper layer (0–10 m; mean § SD =0.62 § 0.35 mg m¡3, N = 60), whereas values in the rangeof 1.5–2.5 mg m¡3 (mean § SD = 1.9 § 2.1 mg m¡3, N = 40)were the rule below 25 m depth.
Temporal variability of Penilia avirostris
Penilia avirostris occurred throughout the studied period(Fig. 3), under the entire observed range of temperature(14.8–28.2°C) and salinity (31.1–36.0). Highest abun-dances were usually found from mid-spring to early autumnabove the thermocline, but some winter maxima (August2005, June 2006, and June 2008) also occurred under well-mixed conditions (Fig. 3). The approximate period whenthe population started to increase varied from year to year(Fig. 3) and usually occurred 1 month following SACWintrusions. In 70% of the cases, abundances above 2,000ind m¡3 were observed when SACW was present. A sig-niWcant positive correlation between temperature and abun-dance was observed (Table 2); but not between abundanceand chl a (Table 2). In contrast, considering only the strati-Wed period (under SACW inXuence) a negative correlation(Spearman’s rank correlation, rs = ¡0.38, N = 49, P < 0.01)between abundance and chl a was detected, whereas nocorrelation occurred if only samples collected under CWinXuence are considered (rs = 0.37, N = 26, P > 0.05).
Reproductive biology
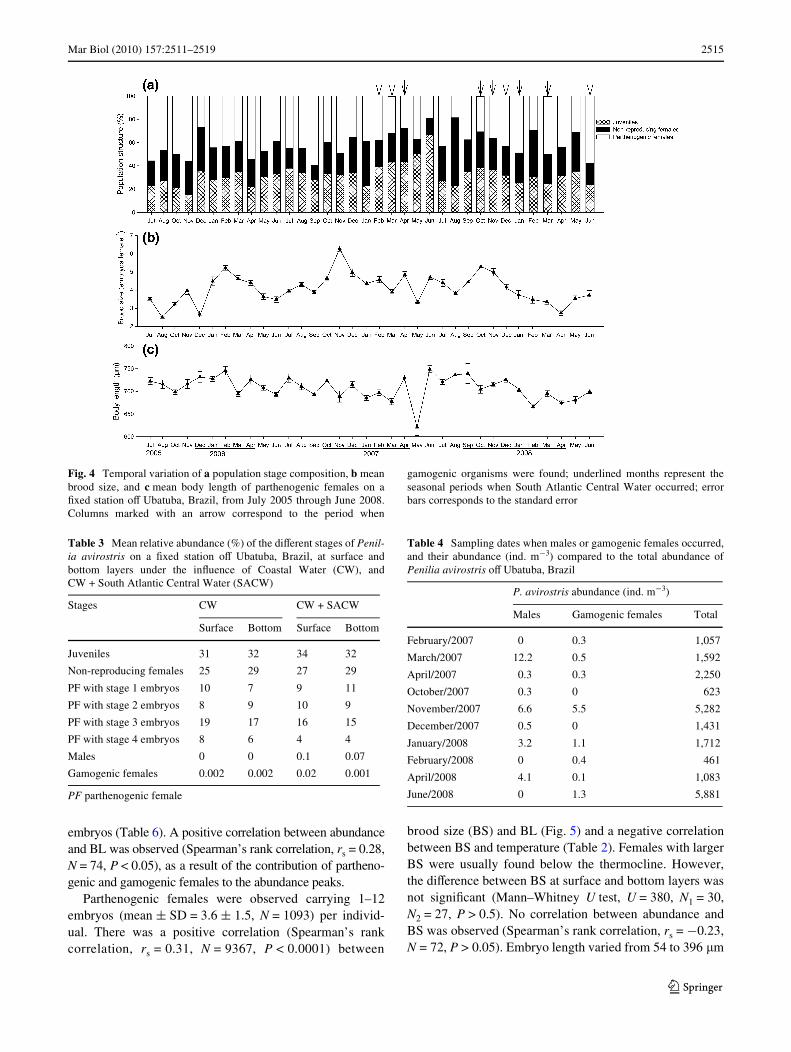
Juveniles, non-reproducing females, and parthenogenicfemales dominated throughout the study period (Fig. 4;Table 3). Body length (BL) varied between 304.5 and1,009.2 �m. Gamogenic individuals occurred at low densities(0–1.3% of the population), only in 2007 and 2008, generallyunder SACW inXuence and associated with high abundancesof the dominant stages (Fig. 4; Table 4). Gamogenic femaleswere found carrying one (78% of females) or two (22% offemales) resting eggs; females bearing two eggs wereobserved only during one abundance peak (>8,000 ind m¡3),which occurred in November 2007. Gamogenic females hadthe highest BL mean value, followed by parthenogenicfemales (Table 5). Parthenogenic females carrying late-stageembryos were usually larger than females bearing early-stage
Fig. 3 Seasonal and multi-annual distribution of a chlorophyll a (chl a), b temperature, and c Penilia avirostris abundance on a Wxed station oV Ubatuba, Brazil, from July 2005 through June 2008. A single plankton tow covering the entire water column was collected from May 2006 through September 2006; the corresponding data are plotted as upper layer abundances. Underlined months represent the period when South Atlantic Central Water occurred
Table 2 Spearman’s rank correlation between abundance, brood sizeand body length of Penilia avirostris, and temperature, chlorophylla (Chl a) and salinity on a Wxed station oV Ubatuba, Brazil
* SigniWcant at P < 0.05; ** P < 0.01; *** P < 0.001
Parameter Temperature Chl a Salinity
Abundance 0.35** ¡0.14 0.21
Brood size ¡0.39*** 0.15 0.24*
Body length ¡0.12 0.01 ¡0.21
123
Mar Biol (2010) 157:2511–2519 2515
embryos (Table 6). A positive correlation between abundanceand BL was observed (Spearman’s rank correlation, rs = 0.28,N = 74, P < 0.05), as a result of the contribution of partheno-genic and gamogenic females to the abundance peaks.
Parthenogenic females were observed carrying 1–12embryos (mean § SD = 3.6 § 1.5, N = 1093) per individ-ual. There was a positive correlation (Spearman’s rankcorrelation, rs = 0.31, N = 9367, P < 0.0001) between
brood size (BS) and BL (Fig. 5) and a negative correlationbetween BS and temperature (Table 2). Females with largerBS were usually found below the thermocline. However,the diVerence between BS at surface and bottom layers wasnot signiWcant (Mann–Whitney U test, U = 380, N1 = 30,N2 = 27, P > 0.5). No correlation between abundance andBS was observed (Spearman’s rank correlation, rs = ¡0.23,N = 72, P > 0.05). Embryo length varied from 54 to 396 �m
Table 3 Mean relative abundance (%) of the diVerent stages of Penil-ia avirostris on a Wxed station oV Ubatuba, Brazil, at surface andbottom layers under the inXuence of Coastal Water (CW), andCW + South Atlantic Central Water (SACW)
PF parthenogenic female
Stages CW CW + SACW
Surface Bottom Surface Bottom
Juveniles 31 32 34 32
Non-reproducing females 25 29 27 29
PF with stage 1 embryos 10 7 9 11
PF with stage 2 embryos 8 9 10 9
PF with stage 3 embryos 19 17 16 15
PF with stage 4 embryos 8 6 4 4
Males 0 0 0.1 0.07
Gamogenic females 0.002 0.002 0.02 0.001
Fig. 4 Temporal variation of a population stage composition, b meanbrood size, and c mean body length of parthenogenic females on aWxed station oV Ubatuba, Brazil, from July 2005 through June 2008.Columns marked with an arrow correspond to the period when
gamogenic organisms were found; underlined months represent theseasonal periods when South Atlantic Central Water occurred; errorbars corresponds to the standard error
Table 4 Sampling dates when males or gamogenic females occurred,and their abundance (ind. m¡3) compared to the total abundance ofPenilia avirostris oV Ubatuba, Brazil
P. avirostris abundance (ind. m¡3)
Males Gamogenic females Total
February/2007 0 0.3 1,057
March/2007 12.2 0.5 1,592
April/2007 0.3 0.3 2,250
October/2007 0.3 0 623
November/2007 6.6 5.5 5,282
December/2007 0.5 0 1,431
January/2008 3.2 1.1 1,712
February/2008 0 0.4 461
April/2008 4.1 0.1 1,083
June/2008 0 1.3 5,881
123

2516 Mar Biol (2010) 157:2511–2519
according to the developmental stage (Table 5). Femalescarrying stage 3 embryos were more abundant, followed bystages 1, 2, and 4 (Table 3).
Discussion
Population dynamics
In tropical and subtropical regions P. avirostris seems tooccur the entire year (Della Croce and Venugopal 1973;
Tang et al. 1995), as observed here. The maintenance of theP. avirostris population oV Ubatuba was a likely result ofthe persistence of high temperatures at the surface layers(>19°C) during the study period. Conversely, in temperateregions the life cycle of P. avirostris begins in springthrough parthenogenic females originating from restingeggs (Onbé 1985; EgloV et al. 1997). The populationincreases rapidly because of parthenogenic reproductionand embryos having short developmental times (EgloVet al. 1997; Atienza et al. 2007), leading to high abundances(Yoo and Kim 1987; Onbé and Ikeda 1995; Atienza et al.2008). The abundance maxima are preceded or accompa-nied by the appearance of gamogenic organisms (DellaCroce and Venugopal 1973; Onbé 1977; Marazzo and Val-entin 2003a), as observed in the present study. After restingegg production by gamogenic females, the population usu-ally disappears from the water column and returns only inthe next warm season (Yoo and Kim 1987; Onbé and Ikeda1995; EgloV et al. 1997; Atienza et al. 2008).
Temperature was the principal factor inXuencingP. avirostris temporal distribution oV Ubatuba, as describedby other works (Onbé 1977; Marazzo and Valentin 2001,2003b). Both in tropical and subtropical regions (Rocha1982; Tang et al. 1995; Marazzo and Valentin 2001) and intemperate areas (Onbé 1977; Yoo and Kim 1987; Onbé and
Table 5 Mean and standard deviation (SD) of body length (�m) of thediVerent stages of Penilia avirostris under the inXuence of CoastalWater (CW), and Coastal Water + South Atlantic Central Water(SACW) oV Ubatuba, Brazil
N number of organisms measured
Stages CW CW + SACW
Mean SD N Mean SD N
Juveniles 455.1 42.5 1,084 450.9 44.2 2,514
Non-reproducing females 598.4 56.1 879 593.1 53.3 1,783
Parthenogenic females 709.0 61.4 2,735 704.4 66.0 5,238
Gamogenic females 770.4 – 1 799.6 48.6 61
Males – – 0 554.4 44.1 40
Table 6 Penilia avirostris—total length of embryonary stages and resting eggs, body length of parthenogenic and gamogenic females, and num-ber of eggs or embryos per female of each embryonary stage on a Wxed station oV Ubatuba, Brazil
N number of organisms measured
Embryonary stage Total length (�m) Body length (�m) Eggs or embryos
Mean SD N Mean SD N Mean SD N
Stage 1 85.0 13.1 1,225 693.1 66.4 2,150 3.7 1.6 2,150
Stage 2 168.4 21.3 1,139 702.4 64.4 2,327 3.5 1.5 2,327
Stage 3 223.3 24.2 1,702 716.8 61.8 3,442 3.6 1.5 3,442
Stage 4 305.1 30.9 1,093 722.8 61.7 1,448 3.6 1.5 1,448
Resting egg 236.6 21.4 75 799.1 48.3 62 1.2 0.4 62
Fig. 5 Brood size (mean § standard deviation; embryos per female) of Penilia avirostris oV Ubatuba, Brazil, separated by size classes (�m). Values in parentheses are the number (n) of organisms ana-lysed in each size class. The only individual larger than 1,000 �m found in our samples had a brood size of 11, which is not depicted in the Wgure
123

Mar Biol (2010) 157:2511–2519 2517
Ikeda 1995; Atienza et al. 2008), P. avirostris reaches highabundances during the warm season when the water col-umn is stratiWed, generally associated with upwelled water(PaVenhöfer et al. 1984). Nonetheless, this species mayoccur in a wide temperature range. In the Bay of Chinhae,South Korea, Yoo and Kim (1987) found P. avirostrisbetween 9 and 28°C, whereas in coastal waters of China itwas found in temperatures ranging from 16 to 32°C (Tanget al. 1995). Temperatures above 22°C seem to be ideal forthe development of P. avirostris (Onbé 1977). However,the higher abundances of P. avirostris observed in thewarmer months may not be related only to temperature,but also to the intensity of the water column stratiWcation(Atienza et al. 2008). According to Atienza et al. (2008), inthe Catalan Sea, the relatively lower abundance of longchain-forming diatoms and the relatively higher abun-dance of nanoXagellates during stratiWed periods favourP. avirostris.
Penilia avirostris occurs mainly at surface layers (Rocha1982; Onbé and Ikeda 1995), as observed in the presentwork. In addition to temperature, the vertical distribution ofP. avirostris is probably controlled by reproductive factors,food availability, and predators (Wong et al. 2004). Parthe-nogenic females carrying advanced embryos (stage 4)occur mainly during the night, as a strategy to avoid visualpredators (Mullin and Onbé 1992; Wong et al. 2004, 2008).Females carrying advanced embryos are larger and there-fore easier to detect by planktivorous Wsh and chaetognaths,the most common P. avirostris predators in tropical andsubtropical environments (Tang et al. 1995; Nip et al. 2003;Rose et al. 2004), including the Southwest Atlantic (Liangand Vega-Pérez 1995; Marazzo and Valentin 2003a).Moreover, stage 4 embryos carry an eye pigment, renderingthe female even more visible to predators (Zaret 1972). Weobserved that females carrying stage 4 embryos were lessabundant in our samples, which is in agreement with ourdaytime sampling scheme.
Penilia avirostris feeding selectivity appears to berelated to food concentration and cell size (Turner et al.1988; Atienza et al. 2006a). This species feeds on a largerange of food particles, usually varying from 2.5 to 100 �m(Katechakis and Stibor 2004), comprising cyanobacteria,pico- and nanoXagellates, ciliates, dinoXagellates, and dia-toms (Turner et al. 1988; Lipej et al. 1997; Katechakis andStibor 2004; Atienza et al. 2006a, 2007). In the presentwork, when the water column was stratiWed because ofSACW intrusions, low chlorophyll values were observedabove the thermocline; nevertheless, high cladoceran abun-dances were found. During this season, P. avirostris proba-bly fed on dinoXagellates and diatoms, which are part ofP. avirostris diet (Atienza et al. 2006b) and usually occuras dominant phytoplankters at the subsurface chlorophyllmaximum in the study area during well-stratiWed conditions
(Aidar et al. 1993). On the other hand, when the water col-umn was well mixed, P. avirostris diet was likely restrictedto nanoplankton and picophytoXagellates, which usuallyaccount for more than 80% of the total phytoplankton abun-dance in the area when CW dominates, especially in win-tertime (Saldanha-Corrêa and Gianesella 2004). Thus, thetrophic role of P. avirostris probably changes with the pre-vailing water masses and the composition of the microbialcommunity.
Reproductive biology
The brood size (1–12; mean of 3.6) observed here was sim-ilar to those reported in other coastal oceans, such as in theGulf of Mexico and the Inland Sea of Japan [mean valuesof 3.1 and 3.25, respectively; Mullin and Onbé (1992)],southern China [1–14 embryos, mean of 4.4; Tang et al.(1995)], and Guanabara Bay [1–11 embryos, mean of 4.35;Marazzo and Valentin (2004)]. The greatest value regis-tered so far, to the best of our knowledge, comes from theNorthwest Atlantic (17 embryos; Della Croce 1966).Regarding body length spectra, Tang et al. (1995) foundspecimens with 360–1,040 �m and Yoo and Kim (1987)from 320 to 1,020 �m, i.e. values slightly larger than thosereported here (305–1,009 �m).
We observed a positive correlation between body lengthof parthenogenic females and brood size, as described inprevious studies (Della Croce 1966; Della Croce andVenugopal 1973; Atienza et al. 2008). On the other hand,Marazzo and Valentin (2003a) found a positive relationshiponly in organisms carrying a maximum of 6 embryos,whereas Tang et al. (1995) and Marazzo and Valentin(2004) did not Wnd any correlation. Brood size is usuallylarger during the initial growth phases of the population(Della Croce and Bettanin 1965; EgloV et al. 1997), but inthe present study no association with growth phase wasnoticed. Atienza et al. (2008) observed a negative correla-tion between brood size and the brood development stage,probably because of embryo abortion (EgloV et al. 1997);we did not observe such relationship.
We found gamogenic females of P. avirostris carryingone or two ovoid resting eggs, as previously described(Della Croce 1966; Onbé 1977; EgloV et al. 1997). In theInland Sea of Japan, Onbé (1985) found resting eggsbetween 210 and 290 �m large and from 140 to 200 �mwide, similar to our results (mean § SD = 237 § 21 �mand 154 § 20 �m, respectively). On the other hand, in theNorthwest Atlantic Ocean, Della Croce and Bettanin (1969)found larger eggs, with major axis ranging from 279 to356 �m, which may be related to lower temperatures andhigh food availability. Gamogenic specimens are generallylarger than parthenogenic individuals (Della Croce 1966;Atienza et al. 2008), whereas males are smaller than
123

2518 Mar Biol (2010) 157:2511–2519
females (EgloV et al. 1997). The organisms from theUbatuba area followed the same pattern.
Gamogenic specimens will likely comprise a low per-centage of the population in unpolluted coastal seas, whereenvironmental conditions are more stable than in inlandwaters (Hairston and Cáceres 1996). Nonetheless, in thepresent study, gamogenic organisms constituted an evenlower percentage of the population (1%) compared to othercoastal seas, as the Catalan Sea (1–20%; Atienza et al.2008). A combination of factors is necessary to stimulatesexual reproduction in freshwater cladocerans (Kleivenet al. 1992), including temperature, quantity and quality offood, overcrowding, photoperiod, and predation (Gyllströmand Hansson 2004). The appearance of males might alsobe stimulated by the release of hormones by females, asshowed by Olmstead and LeBlanc (2002) for Daphniamagna. When exposed to crowding chemicals, Daphniapulex shows signiWcant reduction on clearance rates, as if itwas food-limited (Lürling et al. 2003). The combination ofthese two stimuli may trigger the appearance of gamogenicindividuals during peaks of abundance. However, Marazzoand Valentin (2003a) believe that the occurrence of sexualindividuals is not caused by unfavourable environmentalconditions. They argue that males are always present at lowand constant proportion in the population. Thus, femalescarrying resting eggs tend to appear when female abun-dance increases, as the encounter probability between mat-ing individuals is proportional to the number of males andfemales (Marazzo and Valentin 2003a). Peak abundances(up to »9,000 ind m¡3) observed oV Ubatuba are not char-acteristic of overcrowding events, which are in the range ofhundreds of individuals per litre (Lürling et al. 2003; Smithet al. 2009); therefore, the hypothesis of Marazzo andValentin (2003a) seems more realistic in the present case.
The low densities of gamogenic organisms found in ourstudy suggest that in subtropical and tropical regions, gam-ogenesis is not as crucial to the life cycle of P. avirostris asin temperate regions. However, sexual reproduction mayhave an essential role to ensure population maintenance,either during events leading to a decrease in the populationsize (e.g. parasitism and predation) or when the populationreaches a certain upper abundance level. On the other hand,sexual reproduction and sediment egg banks may providegenetic variability, contributing to both short-term adapta-tion and long-term evolution in cladoceran populations(Frey 1982; Hairston and Fox 2009).
Acknowledgments We thank M. Melo Júnior, M.L. Zani-Teixeira,G.Q. Oliveira and L.C. Laurindo (IOUSP) for their assistance, E.L.Harvey (URIGSO) for revising the English on an earlier version of themanuscript, and the three anonymous reviewers for constructive com-ments. L.K.M. thanks C.E.F. Rocha (IBUSP) for the donation of all hisarticles concerning marine cladocerans. This research is a contributionof the international network ANTARES (www.antares.ws) and was
supported by Fundação de Amparo à Pesquisa do Estado de São Paulo(FAPESP #2007/56931-1) and Conselho Nacional de Desenvolvimen-to CientíWco e Tecnológico (CNPq #306266/2007-5) fellowships toL.K.M. and R.M.L, respectively.
References
Aidar E, Gaeta SA, Gianesella-Galvão SMF, Kutner MBB, Teixeira C(1993) Ecossistema costeiro subtropical: nutrientes dissolvidos,Wtoplâncton e cloroWla-a e suas relações com as condições ocean-ográWcas na região de Ubatuba, SP. Publção Esp Inst Oceanogr SPaulo 10:9–43
Atienza D, Saiz E, Calbet A (2006a) Feeding ecology of the marinecladoceran Penilia avirostris: natural diet, prey selectivity anddaily ration. Mar Ecol Prog Ser 315:211–220
Atienza D, Calbet A, Saiz E, Alcaraz M, Trepat I (2006b) Trophic im-pact, metabolism, and biogeochemical role of the marine cladoc-eran Penilia avirostris and the co-dominant copepod Oithonanana in NW Mediterranean coastal waters. Mar Biol 150:221–235
Atienza D, Calbet A, Saiz E, Lopes RM (2007) Ecological success ofthe cladoceran Penilia avirostris in the marine environment: feed-ing performance, gross growth eYciencies and life history. MarBiol 151:1385–1396
Atienza D, Saiz E, Skovgaard A, Trepat I, Calbet A (2008) Life historyand population dynamics of the marine cladoceran Penilia aviros-tris (Branchiopoda: Cladocera) in the Catalan Sea (NW Mediter-ranean). J Plankton Res 30:345–357
Della Croce N (1966) Observations on the marine cladoceran Peniliaavirostris in northwestern Atlantic waters. US Bur Sport FishWildlife Tech Pap 3:1–13
Della Croce N, Bettanin S (1965) Sviluppo embrionale della formapartenogenetica di Penilia avirostris Dana. Cah Biol Mar 6:269–275
Della Croce N, Bettanin S (1969) Formazione delle uova durevoli inPenilia avirostris Dana. Cah Biol Mar 10:95–102
Della Croce N, Venugopal P (1973) Penilia avirostris Dana in theIndian Ocean (Cladocera). Int Revue Ges Hydrobiol 58:713–721
EgloV DA, FofonoV PW, Onbé T (1997) Reproductive biology ofmarine cladocerans. Adv Mar Biol 31:79–168
Forró L, Korovchinsky N, Kotov AA, Petrusek A (2008) Global diver-sity of cladocerans (Cladocera; Crustacea) in freshwater. Hydro-biologia 595:177–184
Frey DG (1982) Contrasting strategies of gamogenesis in northern andsouthern populations of Cladocera. Ecology 63:223–241
Grahame J (1976) Zooplankton of a tropical harbour: the numbers,composition, and response to physical factors of zooplankton inKingston Harbour, Jamaica. J Exp Mar Biol Ecol 25:219–237
Gyllström M, Hansson LA (2004) Dormancy in freshwater zooplank-ton: induction, termination and the importance of benthic-pelagiccoupling. Aquat Sci 66:274–295
Hairston NG Jr, Cáceres CA (1996) Distribution of crustacean diapa-uses: micro and macroevolutionary pattern and process. Hydrobi-ologia 320:27–44
Hairston NG Jr, Fox JA (2009) Egg banks. In: Likens GE (ed) Ency-clopedia of inland waters, vol 3. Elsevier, Oxford, pp 659–666
Katechakis A, Stibor H (2004) Feeding selectivities on the marinecladocerans Penilia avirostris, Podon intermedius and Evadnenordmanni. Mar Biol 145:529–539
Kleiven OT, Larsson P, Hobaek A (1992) Sexual reproduction inDaphnia magna requires three stimuli. Oikos 65:197–206
Liang TH, Vega-Pérez LA (1995) Studies on chaetognaths oV Ubatubaregion, Brazil. II. Feeding habits. Bolm Inst Oceanogr S Paulo43:27–40
123

Mar Biol (2010) 157:2511–2519 2519
Lipej L, Mozetic P, Turk V, Malej A (1997) The trophic role ofthe marine cladoceran Penilia avirostris in the Gulf of Trieste.Hydrobiologia 360:197–203
Longhurst AR (1985) The structure and evolution of plankton commu-nities. Progr Oceanogr 15:1–35
Lürling M, Roozen F, Van Donk E, Goser B (2003) Response of Daph-nia to substances released from crowded congeners and conspe-ciWcs. J Plankton Res 25:967–978
Marazzo A, Valentin JL (2001) Spatial and temporal variations ofPenilia avirostris and Evadne tergestina (Crustacea, Branchiopoda)in a tropical bay, Brazil. Hydrobiologia 445:133–139
Marazzo A, Valentin JL (2003a) Penilia avirostris (Crustacea, Cteno-poda) in a tropical bay: variations in density and aspects of repro-duction. Acta Oecol 24:S251–S257
Marazzo A, Valentin JL (2003b) Population dynamics of Peniliaavirostris (Dana, 1852) (Cladocera) in a tropical bay. Crustaceana76:803–817
Marazzo A, Valentin JL (2004) Reproductive aspects of marine cla-docerans Penilia avirostris and Pseudevadne tergestina (Crusta-cea, Branchiopoda) in the outer part of Guanabara Bay, Brazil.Braz J Biol 64:543–549
Mullin MM, Onbé T (1992) Diel reproduction and vertical distribu-tions of the marine cladocerans, Evadne tergestina and Peniliaavirostris, in contrasting coastal environments. J Plankton Res14:41–59
Nip THM, Ho WY, Wong CK (2003) Feeding ecology of larval andjuvenile black seabream (Acanthopagrus schlegeli) and Japaneseseaperch (Lateolabrax japonicus) in Tolo Harbour, Hong Kong.Environ Biol Fish 66:197–209
Olmstead AW, LeBlanc GA (2002) Juvenoid hormone methyl farne-soate is a sex determinant in the crustacean Daphnia magna.J Exp Zool 293:736–739
Onbé T (1977) The biology of marine cladocerans in a warm temperatewater. In: Proc symp warm water zoopl spec publ natn inst ocea-nogr. UNESCO, Goa, pp 383–398
Onbé T (1985) Seasonal Xuctuations in the abundance of populationsof marine cladocerans and their resting eggs in the Inland Sea ofJapan. Mar Biol 87:83–88
Onbé T (1999) Ctenopoda and Onychopoda (= Cladocera). In: Boltov-skoy D (ed) South Atlantic zooplankton. Backhuys Publishers,Leiden, pp 797–813
Onbé T, Ikeda T (1995) Marine cladocerans in Toyama Bay, southernJapan: seasonal occurrence and day-night vertical distributions.J Plankton Res 17:595–609
PaVenhöfer GA, Wester BT, Nicholas WD (1984) Zooplankton abun-dance in relation to state and type of intrusions onto the southeast-ern United States shelf during summer. J Mar Res 42:995–1017
Rocha CEF (1982) Distribution of the marine cladocerans (Crustacea,Branchiopoda) oV Santos, Brazil. Bol Zool Univ S Paulo 7:155–169
RoV JC, Hopcroft RR (1986) High precision microcomputer basedmeasuring system for ecological research. Can J Fish Aquat Sci43:2044–2048
Rose K, RoV JC, Hopcroft RR (2004) Production of Penilia avirostrisin Kingston Harbour, Jamaica. J Plankton Res 26:605–615
Saldanha-Corrêa FMP, Gianesella SMF (2004) A microcosm approachon the potential eVects of the vertical mixing of water masses overthe primary productivity and phytoplankton biomass in the south-ern Brazilian coastal region. Braz J Oceanogr 52:167–182
Smith AS, Acharya K, Jack J (2009) Overcrowding, food and phospho-rus limitation eVects on ephippia production and populationdynamics in the invasive species Daphnia lumholtzi. Hydrobiolo-gia 618:47–56
Tang K, Chen QC, Wong CK (1995) Distribution and biologyof marine cladocerans in the coastal waters of southern China.Hydrobiologia 307:99–107
Turner JT, Tester PA, Ferguson RL (1988) The marine cladoceranPenilia avirostris and the ‘microbial loop’ of pelagic food webs.Limnol Oceanogr 33:245–255
Welschmeyer NA (1994) Fluorometric analysis of chlorophyll a in thepresence of chlorophyll b and pheopigments. Limnol Oceanogr39:1985–1992
Wong CK, Ji C, Nip THM (2004) Diel cycle in the percentage abun-dance of parthenogenetic females with embryos of diVerentdevelopmental stages in four species of marine cladocerans.J Plankton Res 26:1095–1103
Wong CK, Vivian CYL, Chan A (2008) Diel cycles of reproductionand vertical migration in the marine cladocerans Pseudevadnetergestina and Penilia avirostris. J Plankton Res 30:65–73
Yoo KI, Kim SW (1987) Seasonal distribution of marine cladoceransin Chinhae Bay, Korea. J Oceanol Soc Kor 22:80–86
Zaret TM (1972) Predators, invisible prey, and nature of polymorphismin Cladocera (Class Crustacea). Limnol Oceanogr 17:171–184
123





























