Embed Size (px)
Citation preview

GLIA 16:351-358 (1996)
Selective Ablation of Astrocytes by Intracerebral Injections
of a-Arninoadipate ‘MOSHE KHURGEL,’ ALICE C. KOO,’ AND GWEN 0. IVY1”
Departments of ’Anatomy and Cell Biology, and ‘Psychology, Scarborough Campus, University of Toronto.
KEY WORDS Gliotoxin, Amygdala, Neurotoxicity, Microglia, Aminoadipic acid, Sloop, GFAP
ABSTRACT The efficacy and the specificity of the putative astrotoxin, a-aminoadi- pate, were examined in this study. The integrity of astrocytes was evaluated at several time points following a single injection of a-aminoadipate into amygdala of adult rats using immunohistochemistry. The density and the morphological appearance of neurons and the response of microglia were also examined. The injection of a-aminoadipate disrupted the astrocytic network in that region. There was a profound loss of glial fibrillary acidic protein-positive and S100P-positive astrocytes, normally present in the region, while vimentin immunohistochemistry revealed the presence of deformed cell processes, presumably astrocytic. The presence of reactive microglia a t the injection site was suggestive of an active degenerative process, while the normal neuronal density and appearance, as compared to controls, suggested that the damage was confined to astrocytes. The confirmed effectiveness and cellular specificity of a-aminoadipate in vivo makes it a potentially important experimental tool for attempting to decipher the functional significance of astrocytes. o 1996 WiIey-Liss, Inc.
INTRODUCTION
During the last decade, there has been a tremendous increase in the number of studies on astrocytes, proba- bly due to the unavoidable realization that these cells actively participate in all neural processes. A multitude of novel properties of astrocytes have been discovered, largely in in vitro preparations. The question of whether astrocytes in vivo share those properties has been a difficult one to address, in part because of the lack of experimental tools to manipulate the functions of these cells.
a-Aminoadipate (aAA), a homolog of glutamate, is an intermediate in metabolism of lysine and occurs natu- rally in the brain (Chang, 1978, 1982). aAA was pro- posed to be a selective toxin for astrocytes and related glial cells over two decades ago when Olney and col- leagues (1971) noticed the gliotoxic properties of ciAA during the neurotoxicity testing of a number of com- pounds. In their study, subcutaneous injections of the racemic mixture of aAA resulted in severe swelling of 0 1996 Wiley-Liss, Inc.
Miiller cells in the retina and degenerative changes in the non-neuronal cells in the arcuate region of hypothal- amus in infant mice. Further in vitro investigations into the properties of aAA confirmed that it has selective cytotoxic effects towards astrocytes, but not neurons, and that it is taken up by astrocytes before exerting its toxic effect on these cells (Huck et al., 1984a,b). How- ever, the selectivity and the effectiveness of aAA toxicity has been controversial. Olney et al. (1980) reported dif- ferences in toxicity following systemic administration of aAA to immature mice, where the L isomer (L-OLAA) appeared to be both neurotoxic and gliotoxic, while the racemic mixture (D,L-aAA) and the D isomer were only gliotoxic. Also, in vitro and in vivo studies on the effects
Received June 12, 1995; acccptcd December 6, 1995.
Address reprint requests to Dr. G.O. Ivy, Division of Life Sciences, Scarborough Campus, University of Toronto, 1265 Military trail, Scarborough. ON M1C 1A4, Canada.
*Present affiliation: Dr. M. Khurgel Dept. Neuroscience Univ. Virginia H.S.C. Box 5148, MR4 Charlottesville, VA 22908, U.S.A.

352 KJ3URGEL ET AL.
of aAA on the retinal cells suggested that there may be a reversible neuronal damage in addition to damage to Miiller cells following exposure to L-OLAA (Casper and Reif-Lehrer, 1983; Sugawara et al., 1990). In contrast, Takada and Hattori (1986) presented ultrastructural evidence of degenerative changes confined to astrocytes only, following injections of L-aAA into the striatum of adult rats, and no degenerative changes in any cell type following injections of D-aAA. Concurrently, Saffran and Crutcher (1987) were unable to replicate the glio- toxic effects of L-CXAA following injections into hippo- campus and striatum.
We have undertaken the following study in an at- tempt to independently test the effectiveness of aAA as a selective gliotoxin, since availability of such a com- pound would likely facilitate research on astrocytes. We report that intracerebral injections of L-aAA disrupt the astrocytic network in the vicinity of injection, and that the degenerative changes appear to be confined only to astrocytes. Preliminary results have been pre- sented (Khurgel and Ivy, 1994).
active as well as lectin-stained cells was compared be- tween the groups under the light microscope. In addi- tion, neuronal density in the area of injection was estimated by counting neuronal profiles within a lim- ited area in the 48 h p.i. vehicle and drug-injected rats, since the area devoid of astrocytes was maximal at that time. The image of a given section was projected using camera lucida. Neurons, which were identified by their morphological appearance in the Nissl-stained sections, were counted within a 100 x 100 pm2 field, which was placed at four semi-randomly chosen locations within 400 +m of the injection tract. Cell counts were per- formed independently by two experimenters. The mean number of neurons per unit area were pooled for each group. Also, the extent of the astrocytic lesions was estimated by measuring the width of the areas devoid of astrocytes (WD) in 3 to 5 GFAP-labeled sections per animal using a densitometer on camera lucida-pro- jected images. The maximal WD scores were pooled and means were obtained for each group. Numerical data were analyzed using analysis of variance, regression analysis and Student’s t-test.
MATERIALS AND METHODS RESULTS
Adult male Long Evans Hooded (LEH) and Sprague- Dawley (SD) rats, weighing 470-610 g, were stereotaxi- cally injected with 5 pl of artificial cerebrospinal fluid (ACSF) or phosphate-buffered saline (PBS) (0.85% NaCl in 0.1 M phosphate buffer) (n = 8), or L-aAA (Sigma Chemical Co., St. Louis, MO, 20 pg/p1 in ACSF or in PBS) (n = 13). The pH of all solutions was adjusted to 7.36. All surgeries were performed under ketaminel xylazine-induced anesthesia. The injections were aimed at the amygdaldpiriform cortex area and were executed intermittently, a t 1 pl/min with 1 min delays. The injec- tion needle was left in place for an additional 5 min, and was withdrawn over 2 min. Two unoperated ani- mals were used as “naive” controls. Rats were euthan- ized at 4 hr (n = 5) , 48 h r (n = 11) and 7 days (n = 5) post drug or vehicle injection (p.i.) by intracardial perfu- sion with 4% buffered paraformaldehyde, under deep anesthesia; the brains were removed, stored in 4% para- formaldehyde for 16-20 hr, and then stored for 16-20 h in 2% dimethylsulfoxide in 20% glycerol for cryopro- tection, and subsequently frozen in isopentane at -40°C. Frozen brains were sectioned coronally (20 and 30 pm). Every fourth section was processed for immuno- histochemistry with anti-glial fibrillary acidic protein (GFAP) (Boehringer Mannheim, Laval, Quebec; 1:200). Adjacent sections were immunolabeled with anti-vi- mentin (Vim) (Boehringer Mannheim, Laval; 1:200) or anti-SlOOP (Sigma, St. Louis Chemical Co., MO; 1:200). All antibodies were monoclonal; standard immunohis- tochemical procedures were used. Every eighth GFAP- immunostained section was counterstained with cresyl violet. Additional sections were stained for microglia using the lectin method of Streit (1990), or stained with cresyl violet.
The appearance of GFAP, Vim, and S100P-immunore-
Injections of the drug vehicle solutions, ACSF or PBS, alone resulted in a typical stab wound-like astrogliosis around the site of injection. In contrast, injections of aAA resulted in pronounced depletions of astrocytes within a large area around the site of injection.
The most prominent difference between the vehicle and drug-injected groups was observed a t 48 h r p.i. Injections of ACSF or PBS produced an increase in GFAP-immunoreactivity (GFAP-IR) around the needle tract and especially in the vicinity of the injection (Fig. 1A). The changes in GFAP+ astrocytes and the spatial limits of altered GFAP-IR were evident by a qualitative comparison between the “reactive” astrocytes in the vi- cinity of the vehicle injection and the immediately adja- cent cells (Fig. lA,B). A narrow area devoid of GFAP- IR could be seen at the injection site (Fig. 1A). The distribution of Sloop’ astrocytes in the vicinity of the injection was similar to that in the adjacent region (Fig. 1C) or in the homologous, non-injected contralateral area. In contrast, the striking absence of GFAP-IR within a wide area around the OLAA injection was appar- ent even a t a low magnification (Fig. 1D). Examination under higher magnification revealed a complete ab- sence of GFAP‘ and S l O O P ‘ astrocytes within that re- gion (Fig. 1E,F). The size of this “astrocyte-free zone” varied, with a range of 200 to 500 pm in either direction from the injection in most animals.
Examination of counterstained GFAP-labeled sec- tions and estimation of neuronal densities revealed a normal appearance and density of neurons in the astrocyte-free zone (Fig. 2A,B). The neuronal density scores obtained in the unoperated rats were combined with those obtained from vehicle-injected controls, since there was no significant difference between the two; the
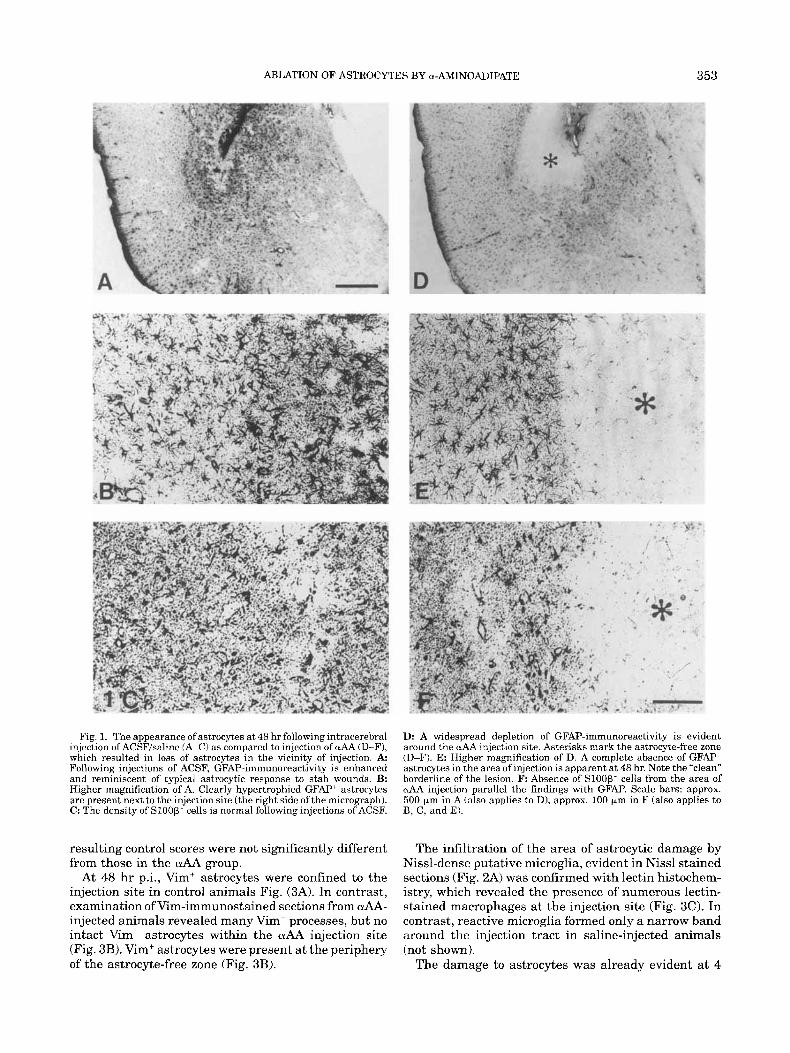
ABLATION OF ASTROCYTES BY a-AMINOADIPATE 353
Fig. 1. The appearance of astrocytes a t 48 hr following intracerebral injection of ACSF/saline (A-C) as compared to injection of uAA (D-FJ, which resulted in loss of astrocytes in the vicinity of injection. A Following injections of ACSF, GFAP-immunoreactivity is enhanced and reminiscent of typical astrocytic response to stab wounds. B: Higher magnification of A. Clearly hypertrophied GFAP+ astrocytes are present next to the injection site (the right side ofthe micrograph). C: The density of SlOOp’ cells is normal following injections of ACSF.
resulting control scores were not significantly different from those in the aAA group.
At 48 h r pi., Vim+ astrocytes were confined to the injection site in control animals Fig. (3A). In contrast, examination of Vim-immunostained sections from &A- injected animals revealed many Vim- processes, but no intact Vim+ astrocytes within the OlAA injection site (Fig. 3B). Vim+ astrocytes were present a t the periphery of the astrocyte-free zone (Fig. 3B).
Ik A widespread depletion of GFAP-immunoreactivity is evident around the wAA injection site. Asterisks mark the astrocyte-free zone (D-F). E: Higher magnification of D. A complete absence of GFAP astrocytes in the area of injection is apparent at 48 hr. Note the “clean” borderline of the lesion. F: Absence of Sloop- cells from the area of uAA injection parallel the findings with GFAI? Scale bars: approx. 500 pm in A (also applies to D), approx. 100 pm in F (also applies to B, C, and EL
The infiltration of the area of astrocytic damage by Nissl-dense putative microglia, evident in Nissl stained sections (Fig. 2A) was confirmed with lectin histochem- istry, which revealed the presence of numerous lectin- stained macrophages at the injection site (Fig. 3C). In contrast, reactive microglia formed only a narrow band around the injection tract in saline-injected animals (not shown).
The damage to astrocytes was already evident a t 4
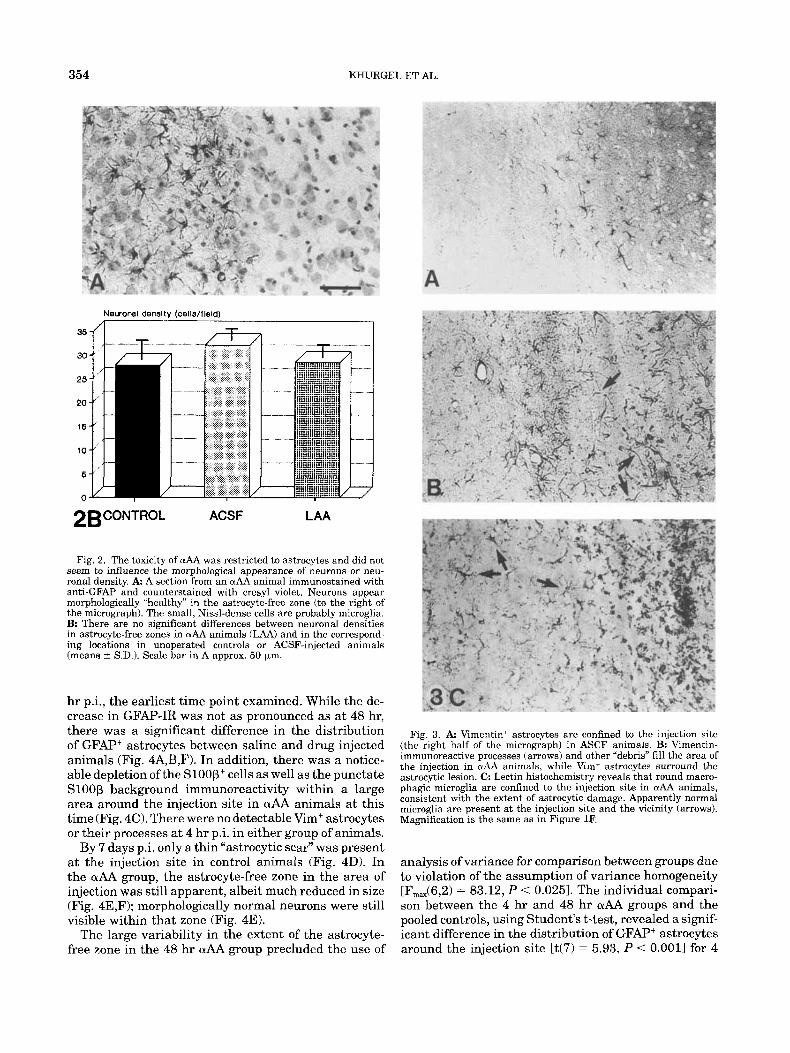
354
Neuronal densltv ICellS/fleldl
KHURGEL ET AL
36 4
30-
26 - 20 - 16 -‘
10 -
6 -
T 1
0
. . _.
2BCONTROL ACSF LAA
Fig. 2. The toxicity of uAA was restricted to astrocytes and did not seem to influence the morphological appearance of neurons or neu- ronal density. A A section from an wAA animal immunostaincd with anti-GFAF’ and counterstained with cresyl violet. Neurons appear morphologically “healthy” in the astrocytt-free zone (to the right of the micrograph). The small, Nissl-dense cells are probably microglia. B: There are no significant differences between neuronal densities in astrocyte-free zones in wAA animals (LAA) and in the correspond- ing locations in unoperated controls or ACSF-injected animals (means ? S.D.). Scale bar in A approx. 50 pm.
hr p.i., the earliest time point examined. While the de- crease in GFAP-IR was not as pronounced as a t 48 hr, there was a significant difference in the distribution of GFAF” astrocytes between saline and drug injected animals (Fig. 4A,B,F). In addition, there was a notice- able depletion of the Sloop+ cells as well as the punctate Sloop background immunoreactivity within a large area around the injection site in OlAA animals at this time (Fig. 4C). There were no detectable Vim+ astrocytes or their processes at 4 hr p i in either group of animals.
By 7 days p.i. only a thin “astrocytic scar” was present at the injection site in control animals (Fig. 4D). In the aAA group, the astrocyte-free zone in the area of injection was still apparent, albeit much reduced in size (Fig. 4E,F); morphologically normal neurons were still visible within that zone (Fig. 4E).
The large variability in the extent of the astrocyte- free zone in the 48 h r OLAA group precluded the use of
Fig. 3. A Vimentin’ astrocytes are confined to the injection site (the right half of the micrograph) in ASCF animals. B: Vimentin- immunoreactive processes (arrows) and other “debris” fill the area of the injection in OLtlA animals, w h i l e Vim+ astrocytes surround the astrocytic lesion. C: Lectin histochemistry reveals that round macro- phagic microglia are confined to the injection site in aAA animals, consistent with the extent of astrocytic damage. Apparently normal microglia are present at the injection site and the vicinity (arrows). Magnification is the same as in Figure 1F.
analysis of variance for comparison between groups due to violation of the assumption of variance homogeneity [F,,(6,2) = 83.12, P < 0.0251. The individual compari- son between the 4 hr and 48 hr aAA groups and the pooled controls, using Student’s t-test, revealed a signif- icant difference in the distribution of GFAP+ astrocytes around the injection site [t(7) = 5.93, P < 0.0011 for 4
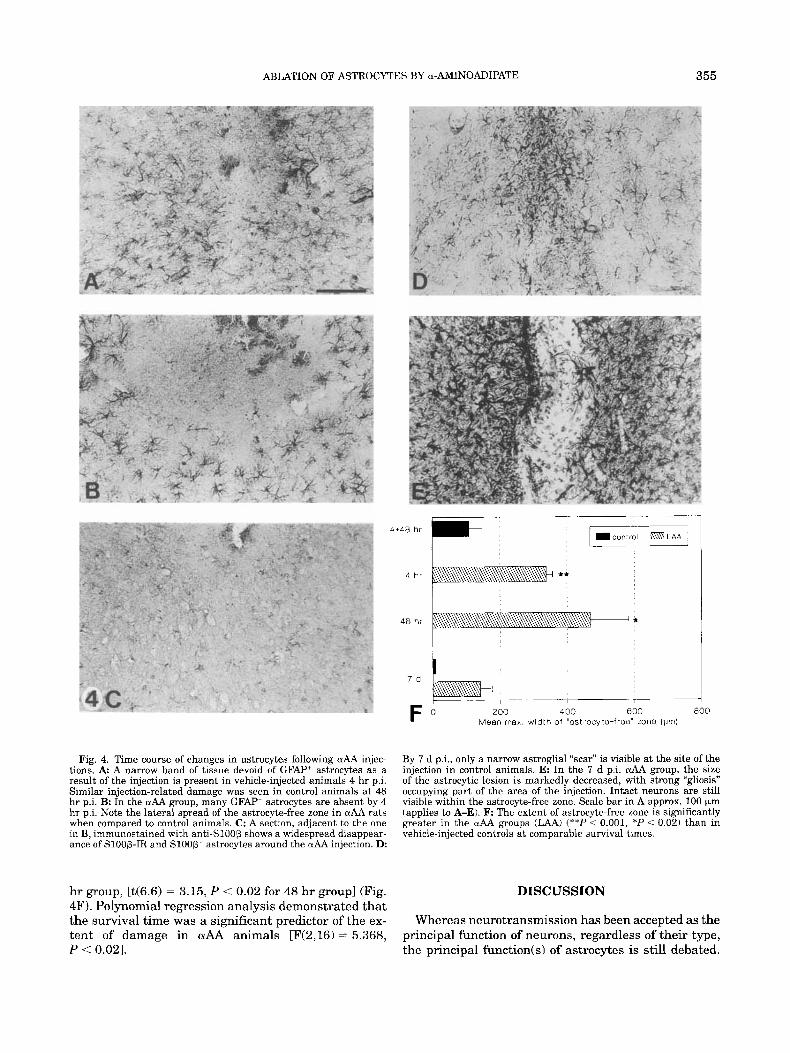
ABLATION OF ASTROCYTES BY a-AMINOADIPATE
Fig. 4. Time course of changes in astrocytes following uAA injec- tions. A A narrow band of tissue devoid of GFAP' astrocytes as a result of the injection is present in vehicle-injected animals 4 hr p i . Similar injection-related damage was seen in control animals at 48 hr p i . B: In the aAA group, many GFAP' astrocytes are absent by 4 hr p.i. Note the lateral spread of the astrocyte-free zone in aAA rats when compared to control animals. C: A section, adjacent to the one in B, immunostained with anti-Sl00P shows a widespread disappear- ance of S100B-IR and S l O O B + astrocytes around the & injection. D:
hr group, [t(6.6) = 3.15, P < 0.02 for 48 hr group] (Fig. 4F). Polynomial regression analysis demonstrated that the survival time was a significant predictor of the ex- tent of damage in aAA animals [F(2,16) = 5.368, P < 0.021.
4+48 hr I-- 4 hr
4 8 hr
355
200 4 00 600 800 Mean max. width of "astfocyte-free" zone (prn)
By 7 d p.i., only a narrow astroglial "scar" is visible at the site of the injection in control animals. E: In the 7 d p.i. crAA group, the size of the astrocytic lesion is markedly decreased, with strong "gliosis" occupying part of the area of the injection. Intact neurons are still visible within the astrocyte-free zone. Scale bar in A approx. 100 Fm (applies to A-E). F The extent of astrocyte-free zone is significantly greater in the aAA groups (JAA) ("*I' < 0.001, *P < 0.02) than in vehicle-injected controls at comparable survival times.
DISCUSSION
Whereas neurotransmission has been accepted as the principal function of neurons, regardless of their type, the principal function(s1 of astrocytes is still debated.

356 KHURGEL ET AL.
Many plausible roles have been assigned to these cells over the years. Some of the questions about the func- tional significance of astrocytes have been eloquently summarized by Somjen (1988). They include hypotheses about a purely structural role, induction and mainte- nance of blood-brain barrier, metabolism of neurotrans- mitters, maintenance of ionic equilibrium and repair following trauma. Some of the hypothesized roles have received strong experimental support and are consid- ered certain (reviewed in Barres, 1991). Others have been recently questioned (Holash et al., 19931, while many others remain hypothetical or controversial. Evi- dence of astrocyte heterogeneity suggests that different subclasses of astrocytes fulfill distinct roles, depending on their location within the CNS and the particular interactions with other neural elements.
The lack of experimental agents to selectivcly manip- ulate astrocytes has hindered somewhat the ability of researchers to explore the various possible roles of astrocytes, especially in in vivo preparations. In this study we present anatomical evidence that intracere- bra1 injections of aAA result in localized, selective abla- tion of astrocytes. This evidence supports the use of aAA as an astrocyte-specific toxin in probing the functional significance of astrocytes in mammalian CNS.
Olney et al. (1971) observed the apparently selective toxic effects of a group of amino acids, but especially D,L-aAA, on non-neuronal cells in the arcuate region of hypothalamus and in the retina when they examined the neurotoxic effects of a variety of systemically admin- istered compounds which were structurally related to monosodium glutamate. Follow-up in vitrn investiga- tions confirmed the gliotoxicity of aAA (Bridges et al, 1992); Huck et al., 1984a,b). Significantly, the in vivo study by Olney et al. (1971) was performed on very young animals, whose blood-brain barrier was still per- meable to the compounds in question, while the in vitro studies used astrocytes from neonatal animals, which raised the possibility of an age-specific effect. One study probed the in vivo toxicity of aAA in adult rats, but it was focused specifically on the Muller cells in the retina (Pedersen and Karlsen, 1979). Thus, until recently there has been no convincing demonstration of the effi- cacy of this compound in the adult mammalian brain.
Takada and Hattori (1986) showed that intrastriatal injections of L-aAA in adult rats result in rapid degener- ative changes in astrocytes, manifested as a loss of structural integrity a t the ultrastructural level. Already at 1 day p.i., swelling, accompanied by severe vacuoliza- tion and many membranous inclusions, was observed in soma and processes of astrocytes in the vicinity of the aAA injection (Takada and Hattori, 1986). Also, in a recent study Chang et al. (1993) found that repeated injections of aAA result in a decrease in the amount of GFAP in substantia nigra and locus coeruleus. On the other hand, Saffran and Crutcher (1987) failed to obtain evidence in support of gliotoxic properties of aAA. In their study, there were no observable changes in the morphology of GFAP' astrocytes a t 4 or 48 h r following the injection of L-aAA into hippocampus or striatum.
To avoid interpetational problems when comparisons
between various studies are made, we have used two different vehicle solutions (ACSF and PBS) and two different strains of rats (LEH and SD). There were no apparent differences in the anatomical results with re- spect to these variables. Also, while the present study focused on changes in the amygdaldpiriform area, a regional heterogeneity in sensitivity of astrocytes to OlAA is unlikely. This possibility has been rejected by Saffran and Crutcher (1987) and is not supported by our work in progress (M. Khurgel, unpublished observa- tions), or by other studies (Chang et al., 1993; McBean, 1990; Takada and Hattori, 1986).
We have used several astrocytic markers to monitor the viability of astrocytes following intracerebral injections of &. GFAP is a well-studied astrocyte-specific protein, which makes up the intermediate filaments (IFs) in cy- toskeleton of differentiated astrocytes (Eng, 1985). Since changes in cell shape are accompanied by reorganization of the cytoskeleton, GFAP-IR has been used extensively in studies on morphological changes in astrocytes in a va- riety of experimental conditions. An increase in GFAP- IR is usually associated with hypertrophy of astrocytes as part of their transformation in response to a variety of factors in vitro and in vivo. Under certain conditions, a decrease in GFAP content and GFAP-IR (Canady et al., 1990; Huck et al., 1984a) or a total abolition of GFAP- IR (Blaugrund et al., 1992; Schmidt-Kastner and Ingvar, 1994) have been observed and interpreted to represent an impairment or a depletion of astrocytes from the area under examination. Using electron microscopy, Blau- grund et al. (1992) confirmed that the absence of GFAP- IR following a crush injury to the optic nerve was due to the absence of astrocytes from the lesioned area. In pres- ent study we observed a small, but significant, depletion of GFAP' astrocytes already at 4 hr following aAA injec- tions (Fig. 4B). The toxicity of aAA was most apparent a t 48 h r p.i, whereby a wide area around the injection site was devoid of GFAP' cells in most animals (Fig. lD,E). In a few animals this effect was not as pronounced, which may be due to the inadvertent eflux of aAA during the withdrawal of the injection needle.
The second astroglial marker used in this study, Sloop, is a small calcium-binding protein which in CNS is synthesized primarily by astrocytes (Haan et al., 1982; Zimmer and Van Eldik, 1987). SlOOP has been hypothesized to be a gliotrophic and a neurotrophic factor and to play a role in development of the CNS and possibly in certain pathological conditions (Balazs and J~rgensen , 1993; Marshak, 1990). The relative den- sity of Sloop+ astrocytes in the area of vehicle injection appeared normal (Fig. 1C) and was similar to that in contralateral amygdala or in unoperated controls. In the UAA animals, S100P-IR disappeared very rapidly, by 4 hr p i . (compare Figs. 1C and 4C). This result is in agreement with an earlier study, where an intrastriatal injection of D,L-aAA caused a significant reduction in the levels of glutamine synthetase by 6 hr post injection (McBean, 1990). Since S l O O P is thought to be a secreted molecule, it is very likely that damage to the integrity of astrocytes results in loss of this protein. Therefore, the absence of S100P-IR in astrocytes within the area

ABLATION OF ASTROCYTES BY a-AMINOADIPATE 357
of aAA injection may indicate a functional impairment of astrocytes in that region. Interestingly, some GFAP’ astrocytes were still prominent in the same area in adjacent sections from aAA animals in the 4 h group (see Figs. 4B and 4C). However, by 48 h p.i., the loss of GFAP’ astrocytes in a wide area around the injection (Fig. 1E) paralleled the absence of Sloop- cells (Fig. 1F). The apparent delay between the functional impair- ment of astrocytes and the final elimination of GFAP is to be expected since IFs are thought to be the most stable intracellular components (Bershadsky and Vasi- liev, 1988). These results also confirm the in vitro obser- vation that “the ghosts” of irreversibly damaged astrocytes retained GFAP-IR for some period after expo- sure to aAA (Huck et al., 1984a).
Another interesting observation in our study was the presence of Vim+ processes in the area of astrocytic damage. Vim is the intermediate filament (IF) protein which is present in astrocytes early in development (Dahl et al., 1981) and is gradually replaced by GFAP as the main IF protein in astrocytes during maturation (Pixley and De Vellis, 1984; Stichel et al., 1991). Vim is usually undetectable in gray matter astrocytes of adult animals (see Khurgel et al., 1992), but it appears to be re-expressed in these cells following activation of astrocytes due to trauma (Calvo et al., 1991; Mikucki and Oblinger, 1991; Petito et al., 1990; Schiffer et al., 1986; Takamiya et al., 1988) or seizure activity (Khurgel et al., 1992, 1995). Therefore, the presence of Vim+ astrocytes a t the site of vehicle injection (Fig. 3A) re- flects the activation of these cells due to the trauma of acute injection. Similarly, the appearance of Vim-IR in what appear to be astrocytic processes in aAA animals may represent the accumulation of this protein in astrocytes as part ofthe response to local injury. Alterna- tively, it is possible that the breakdown of the astrocytic cytoskeleton unmasked the epitopes of constitutive Vim molecules or altered the GFAP epitopes. The latter expla- nation is supported by recent evidence that polyclonal antibodies to GFAP label what appear to be damaged astrocytes in the area of severe necrosis, while mono- clonal antibodies fail to label these cells (Schmidt-Kas- tner and Inguar, 1994). Regardless ofthe exact molecular basis for re-appearance of Vim-IR in the putative astrocytic processes within the lesion, it is important to note that there were no intact Vim’ astrocytes in that area (Fig. 3B). Thus, our combined immunohistochemi- cal results, coupled with the ultrastructural evidence by Takada and Hattori (1986), suggest that astrocytes are irreparably damaged following injections of aAA.
Our results also confirm the previous observations that neurons are not adversely affected by &. Huck et al. (1984a) did not detect any detrimental changes in neurons following addition of aAA to dissociated cere- bellar cultures; in fact, even the subsequent elimination of astrocytes did not affect the survival of neurons. Ta- kada and Hattori (1986) demonstrated the absence of degenerative changes in neurons adjacent to degenerat- ing astrocytes at the ultrastructural level. In the pres- ent study, the morphological appearance and the den- sity of neurons in the astrocyte-free zone were normal
(Fig. 2A,B). Also, in a pilot study, systemic administra- tion of L-aAA or D,L-& to adult rats did not result in any signs of neuronal degeneration in the brain, as assessed with the cupric silver degeneration stain (Khurgel, Switzer, and Ivy, unpublished observations). Thus, it appears that neuronal necrosis observed follow- ing the systemic administration of L-& by Olney et al. (1980) may have been due to the large dose of that compound or the fact that immature animals were used in their study. While it is conceivable that a small loss of neurons, due to either a direct action by the drug or the absence of functional astrocytes, has gone unde- tected in present study, the extent of such loss is clearly not significant. The surprising finding of neuronal via- bility in the absence of astrocytes at 48 hr, and even at 7 days p.i. presents an exciting opportunity for re- evaluation of the role of astrocytes in maintenance of the microenvironment in CNS and the dependence of neurons on neighboring astrocytes.
With respect to cell types other than neurons and astrocytes, only microglia were analyzed in our study. The presence of occasional lectin’ cells with morphology of ramified microglia within the area of astrocytic lesion (Fig. 3C) suggests that ciAA is not toxic toward these cells. Takada and Hattori (1986) also showed that a t the ultrastructural level oligodendrocytes, microglia, and endothelial cells were not adversely affected by aAA injections. On the other hand, the presence of numerous macrophage-like cells immediately in the area of aAA injection (Fig. 3C) is suggestive of an active phagocytic process, where astrocytes are likely the only cell type undergoing degenerative changes. In light of the above evidence and a growing awareness about the functional distinctiveness of different types of glial cells, it may bc more appropriate to refer to aAA as an astrotoxin.
The limitations of using aAAfor in vivo studies should be noted. The mechanism of toxicity of this compound is still unknown and deserves further study. Theoretically, aAA could be binding to an astrocyte-specific gluta- mate-like receptor; excess of aAA could then initiate a sequence of excitotoxic events, similar to the one hy- pothesized to cause neuronal degeneration (Choi, 1987). Also, aAA appears to cause an ablation of astrocytes, as opposed to a temporary “shut-down,” which precludes certain types of experiments. aAA apparently does not cross the blood-brain barrier in adult CNS, and there- fore requires invasive techniques such as intracerebral injections. In addition, the apparent re-population by astrocytes of the astrocyte-free zone, requires either repeated injections or continuous administration of aAA if long term absence of astrocytes is the goal. De- spite these limitations, it is hoped that use of aAA, or other putative gliotoxins, such as fluorocitrate (Paulsen et al., 19871, or 6-aminonicotinamide ( h u m and Ro- senstein, 1993; Politis, 1989) will advance the study of astrocytes and astrocyte-neuronal interactions.
In conclusion, we believe that our results present a convincing demonstration of the specificity and efficacy of LVAA with respect to selective ablation of astrocytes in the adult mammalian brain. The long term sequelae of local elimination of astrocytes remain to be deter-

358 KHURGEL ET AL.
mined. Of particular interest is the putative re-popula- tion of the astrocyte-free zone either by migrating or proliferating astrocytes. Certainly, the evaluation of neuronal function in the absence of astrocytes in vivo should provide invaluable information on the properties of both neurons and astrocytes. The potential use- fulness of (wAA in studying post-traumatic regeneration or in studies of gliomas also should be emphasized.
ACKNOWLEDGMENTS
We wish t o thank S.C. Barsoum for assistance with some aspects of this study, M.P. Murphy for help with statistical analysis, Dr. R.C. Switzer for collaboration in the pilot degeneration study, and Dr. T. Hattori for critical reading of the manuscript. This research was supported by an NSERC grant to G.I. and the Epilepsy Canada 1995 Canadian National Studentship Award to M.K.; A.C.K. was a recipient of a Summer Research Internship from the Department of Anatomy and Cell Biology, University of Toronto.
REFERENCES
Balazs, R. and Jwgensen, O.S. (1993) Down’s syndrome and 5-100 protein. In: Biology and Patho1og.y ofAstrocyte-Neuron Interactions. S . Fedoroff, B.H.J. Juurlink, and R. Doucette, cds. Plenum Press, New York, pp. 383394.
Barres, B.A. (1991) New roles for glia. J. Neurosci., 11:3685-3694. Bershadsky, A.D. and Vasiliev, J.M. (1988) Cytoskelrton. Plenum, New
York, 298 pp. Blaugrund, E., Duvdevani, R., Lavie, V., Solomon, A,, and Schwartz,
M. (1992) Disappearance of astrocytes and invasion of macrophages following crush injury of adult rodent optic nerves: Implications for regeneration. Exp. Neurol., 118:105-115.
Bridges, R.J., Hatalski, C.G., Shim, S.N., Cummings, B.J., Vijayan, V., Kundi, A., and Cotman, C.W. (1992) Gliotoxic actions ofexcitatory amino acids. Neurophurmacology, 31:899-907.
Calvo, J.L., Carbonell, A.L., and Boya, J. (1991) Co-expression of glial acidic protein and vimentin in reactive astrocytes following brain injury in rats. Bruin Res., 566:333-336.
Canady, K.S. Ali-Osman, F., and Rubel, E.W. (1990) Extracellular potassium influences DNA and protein synthesis and glial fibrillary acidic protein expression in cultured glial cells. Glia, 3:368-374.
Casper, D.S. and Reif-Lehrer, L. (1983) Effects of alpha-aminoadipate on the morphology of the isolated chick embryo retina. Inuest. Op- thalm. Visual Sci., 24~1480-1488.
Chang, Y.F. (1978) Lysine metabolism in the rat brain: The pipecolic acid-forming pathway. J. Neurochem., 30:347-354.
Chang, Y.F. (1982) Lysine metabolism in the human and the monkey: demonstration of pipecolic acid formation in the brain and other organs. Neurochem. Res., 7577-588.
Chang, F.W., Wang, S.D., Lu, K.T., and Lee, E.H.Y. (1993) Differential effects of gliotoxin and MPTP in the substantia nigra and the locus coeruleus in BALBic mice. Bruin Res. Bull., 31 :253-266.
Choi, D.W. (1987) Ionic dependence of glutamate neurotoxicity. J. Nerr- rosci., 7:369-379.
Dahl, D., Hueger, D.G., Bimami, A,, Weber, K., and Osborn, M. (1981) Vimentin, che 57 000 molecular weight protein of fibroblast fila- ments, is the major cytoskeletal component in immature glia. Eur J. Cell Hiol., 24:191-196.
Eng, L.F. (1985) Glial fibrillary acidic protein (GFAP): The major pro- tein of glial intermediate filaments in differentiated astrocytes. J. Neuroimmunol., 8:203-214.
Haan, E.A., Boss, B.D., and Cowan, W.M. (1982) Production and char- acterization of monoclonal antibodies against the “brain-specific” proteins 14-3-2 and S-100. Proc. Natl. Acad. Sci. U.S.A., 79:
Holash, J.A., Noden, D.M., and Stewart, P.A. (1993) Re-evaluating the 7585-7589.
role of astrocytes in blood-brain barrier induction. Deu. Dynamics, 197: 14-25.
Huck, S., Grass, F., and Hatten, M.E. (1984a) Gliotoxic effects of a- aminoadipic acid on monolayer cultures of dissociated postnatal mouse cerebellum. Neuroscience, 3:783-791.
Huck, S., Grass, F., and Hortnagl, H. (1984b) The glutamate analogue a-aminoadipic acid is taken up by astrocytes before exerting its gliotoxic effect in vitro. J. Neurosci., 4:2650-2657.
Khurgel, M., Racine, R.J., and Ivy, G.O. (1992). Kindling causes changes in the composition of the astrocytic cytoskeleton. Brain. Res., 592:338-342.
Khurgel, M. and Ivy, G.O. (1994) Evidence of a localized ablation of astrocytes by intracerebral injection of L-a-aminoadipate. Soc. Neurosci. Abstr , 20:1503.
Khurgel, M., Switzer, R.C., Teskey, G.C., Spiller, A.E., Racine, R.J., and Ivy, G.O. (1995) Activation of astrocytes during epileptogenesis in the absence o f neuronal degeneration. Neurobiol. Dis. 2:23-35.
Krum, J.M. and Rosenstein, J.M. (1993) Effect of astroglial degenera- tion on the blood-brain barrier to protein in neonatal rats. Dev. Brain Res., 74:41-50.
Marshak, D.R. (1990) SlO0D as a neurotrophic factor. Prog. Brain Res., 86:169-181.
McBean, G.J. (1990) Intrastriatal injection of IIL-alpha-aminoadiuate reduces kainate toxicity in vitro. Neuroscience, 34:225-234.
Mikucki, S.A. and Oblinger, M.M. (1991) Vimentin mRNA expression increases after corticospinal axotomy in the adult hamster. Metab. Brain Dis., 6:3349.
Olney, J.W., Oi Lan Ho, and Rhee, V. (1971) Cytotoxic effects of acidic and sulfur containing amino acids on the infant mouse central ner- vous system. Exp. Brain Res., 14:61-76.
Olney, J.W., de Gubareff, T., and Collins, J.F. (1980) Stereospecificity of the gliotoxic and anti-neurotoxic actions of alpha-aminoadipate. Neurosci. Lett., 19:277-282.
Paulsen, R.E., Contestabile, A,, Villani, L., and F. Fonnum (1987) An in vivo model for studyingfunction of brain tissue temporarily devoid of glial metabolism: The use of fluorocitrate. J. Neurochem., 48:1377-1385.
Pedersen, 0.9. and Lund Karlsen, R. (1979) Destruction of Miiller cells in the adult rat by intravitreal injections of D,L alpha-aminoad- ipic acid. An electron microscopic study. Exp. Eye Res., 14:569-575.
Petito, C.K., Morgello, S., Felix, J.C., and Lesser, M.L. (1990) The two patterns of reactive gliosis in postischemic rat brain. J. Cereb. BZood Flow Metub., 10:850-859.
Pixley, S.K.R. and De Vellis, J . (1984) Transition between immature radial glia and mature astrocytes studied with a monoclonal anti- body to vimentin. Deu. Brain Res., 15:201-209.
Politis, M.J. (1989) 6-Aminonicotinamide selectively causes necrosis in reactive astrocytes in vivo, J. Neurol. Sci., 92:71-79.
Saffran B. N. and Crutcher, K.A. (1987) Putative gliotoxin, a-aminoad- ipic acid, fails to kill hippocampal astrocytes in vivo. Neurosci. Lett., 81:215-220.
Schiffer, D., Giordana, M.T., Migheli, A,, Giaccone, G., Pezzotta, S . and Mauro, A. (1986) Glial fibrillary aci&c protein and vimentin in the experimental glial reaction of the rat brain. Brain Res., 374:llO-118.
Schmidt-Kastner, R. and Inguar, M. (1994) Loss of immunoreactivity for glial fibrillary acidic protein (GFAP) in astrocytes as a marker for profound tissue damage in substantia nigra and basal cortical areas after status epilepticus induced by pilocarpine in rat. Glia, 12:165-172.
Somjen, G.G. (1988) Nervenkitt: Notes on the history of the concept of neuroglia. Gliu, 1:2-9.
&ichel, C.C., Miiller, C.M., and Zilles, K. (1991) Distribution of glial acidic protein and vimentin immunoreactivity during rat visual cor- tex development. J. Neurocytol., 20:97-108.
Streit, W.J. (1990) An improved staining method for rat microglial cells using the lectin from Grifonia simplicifulia (GSA I-BJ. J. Histochem. Cytochem., 38:1683-1686.
Sugawara, K., Torigoe, K., Okoyama, S., Negishi, K., and Kato, S. (1990) Neurotoxic effects of L-a-aminoadipic acid on the carp retina: a long term observation. Neuroscience, 36:155-163.
Takada, M. and Hattori, T. (1986) Fine structural changes in the rat brain after local injections of gliotoxin, alpha-aminoadipic acid. Hist. Histopathol., 1:271-275.
Takamyia, Y., Kohsaka, S., Toya, S., Otani, M., and Tsukada, Y. (1988) Immunohistochemical studies on the proliferation of reactive astrocytes and the expression of cytoskeletal proteins following brain injury in rats. Deu. Bruin Res., 38:201-210.
Zimmer, D.B. and Van Eldik, L.J. (1987) Tissue distribution of rat S- 1 0 0 ~ and S-loop and S-100-binding proteins. Am. J. Physiol., 252:C285-C289.





























