Embed Size (px)
Citation preview

iedsNDgbNbNwti(9iarfaacsimimnA
dP
gpm
Experimental Neurology 159, 451–458 (1999)Article ID exnr.1999.7191, available online at http://www.idealibrary.com on
Selective Agenesis of Mesencephalic Dopaminergic Neuronsin Nurr1-Deficient Mice
Wei-dong Le,* Orla M. Conneely,† Linglong Zou,* Yi He,* Odila Saucedo-Cardenas,† Joseph Jankovic,*Dennis R. Mosier,* and Stanley H. Appel*
*Department of Neurology and †Department of Cell Biology, Baylor College of Medicine, One Baylor Plaza, Houston, Texas 77030
Received February 22, 1999; accepted June 30, 1999
osDaebnmHDv
scmfsrtttcr(artDhtpnimct
sne(
Nurr1, a member of the nuclear receptor superfam-ly of transcription factors, has been found to bessential for the development of ventral midbrainopamine (DA)ergic neurons. To study the regionalelectivity and phenotypic specificity of regulation byurr1 of the genesis of DAergic neurons, we examinedAergic, serotonin (5-HT)ergic, norepinephrine (NE)er-ic, cholinergic, glutamate (GLU)ergic, and g-amino-utyric acid (GABA)ergic neurons in the brains ofurr1-deficient mice by immunohistochemistry andiochemistry. We demonstrated that in homozygousurr1-deficient mice (Nurr12/2), DAergic neuronsere totally absent in substantia nigra and ventral
egmental area, but preserved in other regions includ-ng diencephalon and hypothalamus, olfactory bulbOB). Levels of DA in Nurr12/2 mice were decreased by8% in striatum (Str) and 65% in OB. NEergic neuronsn locus ceruleus, 5-HTergic neurons in raphe nuclei,nd cholinergic neurons in basal forebrain and otheregions were not changed. A 30% reduction of NE wasound in the Str of Nurr12/2 mice. The levels of GLUnd GABA and the activity of choline acetyl transfer-se in the brains of Nurr12/2 mice were not signifi-antly altered. Our results demonstrate a selective andpecific deficit of DA and absence of DAergic neuronsn the mesencephalic structures of Nurr1-deficient
ice, which resembles the pattern similar to that seenn patients with Parkinson’s disease (PD). This model
ay contribute to our understanding of the mecha-isms influencing DAergic cell survival in PD. r 1999
cademic Press
Key Words: Nurr1; null mutant mice; tyrosine hy-roxylase; dopamine; mesencephalon; development;arkinson’s disease.
INTRODUCTION
Dopaminergic (DAergic) neurons in the ventral re-ion of the midbrain, which innervate the striatum,lay an important role in control of the voluntary
ovement, cognition, and emotive behaviors (8). Loss n451
f DAergic neurons in the midbrain occurs in Parkin-on’s disease (PD) (9), and disturbances of centralAergic transmission are implicated in schizophreniand other mental disorders (24). During embryogen-sis, induction of ventral DAergic neurons is influencedy interactions with the underlying mesoderm of theotochord and the floor plate, which lies at the ventralidline of the developing central nervous system (23).owever, the molecular signals that direct centralAergic neuron development and maintain their sur-ival are still poorly understood.Nurr1, a member of the orphan nuclear receptor gene
uperfamily (12), has been recently found to be aritical genetic factor regulating the development andaintenance of midbrain DAergic neurons and their
unctions (2, 19, 26). Our previous study has demon-trated that Nurr1 is highly expressed in severalegions including substantia nigra (SN), ventral tegmen-al area (VTA), olfactory bulb (OB), hippocampus (Hip),emporal cortex, subiculum, cerebellum, posterior hypo-halamus, and habenular nuclei (18). In the mesen-ephalon, the distribution of Nurr1 mRNA expressionesembles the distribution of DA-containing neurons18, 25). Using Nurr1 null mutant mice, Zetterstrom etl. (26) and Saucedo-Cardenas et al. (19) have recentlyeported that ablation of Nurr1 leads to absence ofyrosine hydroxylase (TH)-immunoreactive cells andA transmission in the midbrain. It is not known,owever, whether this absence of TH-positive cells inhe mesencephalon is due to the lack of DAergichenotype induction or due to actual loss of DAergiceurons. Furthermore, it is not clear whether the
mpact of Nurr1 on DAergic neurons is restricted to theidbrain or is more widespread and whether Nurr1
an influence the development of other neurotransmit-er systems in the brain.
The objective of this study is to investigate theelectivity of Nurr1 effects on the development of majoreurotransmitters including DA, serotonin (5-HT), nor-pinepherine (NE), and acetylcholine (Ach), glutamateGLU), and g-aminobutyric acid (GABA) in the central
ervous systems. Here we demonstrate that deficiency0014-4886/99 $30.00Copyright r 1999 by Academic Press
All rights of reproduction in any form reserved.

idtidm
N
wilugtornttbtmcEh
asmlmsNu
I
xnwbp0fitCibanmm
hl5agTEpConmamd(b(w
B
idBamue2dpw(aca
tmsnpf2aGszeottu
452 LE ET AL.
n Nurr1 selectively and specifically affects both theevelopment and the survival of DAergic neurons inhe SN and VTA regions. Our results warrant furthernvestigations to define the role of Nurr1 expression inisorders affecting predominantly DAergic neurons inesencephalic regions, such as PD.
MATERIALS AND METHODS
urr1-Deficient Mice
As previously described (19), Nurr1 gene disruptionas generated by insertion of neor gene PGKNEObpA
nto a unique BamHI restriction site in exon 3 that wasocated downstream from the initiator codon ATG butpstream of the DNA-binding domain in the Nurr1ene. The insertion of the neor gene into exon 3 divideshe 7.6-kb Nurr1 genomic fragment into 58 and 38 armsf Nurr1 homology that are 1.9 and 5.7 kb in size,espectively. The herpes simplex virus thymidine ki-ase gene was attached 58 to exon 3 and inserted withranscriptional orientation opposite to both the neo andhe Nurr1 genes. The target vector was then introducedy electroporation to embryonic stem cells, and theransmission of the Nurr1 mutation was generated byicroinjection into the blastocoel of 3.5-day-old blasto-
yst stage embryos derived from C57BL/6 females.mbryos were transferred unilateral into the uterineorn of pseudopregnant F1 foster mothers.The homozygous (Nurr12/2) mice were hypoactive
nd died within 1 day after birth due to an inability touckle milk (19, 26). The heterozygous (Nurr11/2)ice showed no overt phenotypic abnormality. DA
evels in the striatum of the adult Nurr11/2 mice (2–16onths old) were not significantly different from the
ame age wild-type (Nurr11/1) mice (14). Sinceurr12/2 cannot survive over 24 h after birth, we havesed only newborn mice in this study.
mmunohistochemical Studies
After anesthesia (42.8 mg/ml ketamine, 8.6 mg/mlylazine, and 1.4 mg/ml acepromazine; 10 µl/mouse),eonatal Nurr12/2, Nurr11/2, and Nurr11/1 miceere perfused through the heart with 2 ml phosphateuffer solution (PBS, pH 7.4) and then with 2 ml 4%araformaldehyde containing 0.05% glutaraldehyde and.2% picric acid. The brains were removed and post-xed in the same fixative, protected in 30% sucrose, andhen subsequently frozen in dry ice cooled acetone.oronal sections (20 µm) of the entire brains were cut
n a cryostat, blocked with 5% goat serum, and incu-ated for 24 h at 4°C with following specific polyclonalntibodies (ab): 1, TH ab (1:500) to detect DAergiceurons in mesencephalon, diencephalon, hypothala-us, OB (1), and other TH-containing cells in adrenal
edulla (AM) and sympathetic ganglia (SG); dopa-b- sydroxylase (DBH) ab (1:400) to detect NEergic cells inocus ceruleus (LC), and serotonin-1 ab (1:400) to detect-HTergic cells in the raphe nucleus; and cholinecetyltransferase (ChAT) ab (1:500) to detect choliner-ic neurons in the basal forebrain and other regions.H, DBH, and serotonin-1 ab were purchased fromugen International (Allendale, NJ), and ChAT ab wasurchased from Chemicon International (Temecula,A). Some of the sections in the mesencephalic regionf newborn mice were immunostained with anti-euronal nuclei ab (NeuN, 1:200; Chemicon) to deter-ine whether there was any neuronal loss in the SN
nd VTA of Nurr12/2 mice. After three washes with 10M PBS, the sections were incubated with a 1:200
ilution of peroxidase-labeled corresponding second abVector Laboratories, Burlingame, CA) for 1 h, followedy standard avidin–biotin immunostaining proceduresVector Laboratories, Burlingame, CA). The sectionsere then counterstained with methyl green.
iochemical Examination
The levels of DA, NE, 5-HT, and related compoundsn the striatum (Str), OB, AM, and retina (RE) wereetermined according to our previous method (13).riefly, the tissues of interest were dissected fromnesthetized neonatal mice on ice under a dissectingicroscope, washed in cold PBS, and extracted by
ltrasonication with 10% perchloric acid (1:10 v/v). Thextracts were clarified by centrifugation at 15,000g for0 min. The supernatants were filtered through acro-isc filters (mesh size 0.25 µm) and subjected to high-erformance liquid chromatography (HPLC) packedith a BAS P/N reversed-phase cartridge column
Phase-II ODS 3 3 100 3 3.2 mm). DA, NE, 5-HIAA,nd related compounds were detected by an electro-hemical detector (BAS, Inc., West Lafayette, Indiana)nd calibrated by a HP 3395 integrator.To measure the levels of GLU and GABA in the brain
issues of Nurr1-deficient mice, an improved detectionethod with HPLC was employed (21). Tissues dis-
ected from the Str, temporal cortex (TC), and Hip ofeonatal mice were extracted by sonication in 10%erchloric acid (1:10 v/v).After centrifugation at 12,000gor 20 min, the supernatants were adjusted to a pH of.2 and then placed in storage loops on an amino acidnalyzer’s injector (Dionex DX500; Sunnyvale, CA).lu, GABA, and other amino acids were loosely ad-
orbed to the ion-exchange resin (sulfonated divinylben-ene cross-linked polystrene beads) and differentiallyluted from the column with a series of lithium buffersf increasing pH and ionic strength and further posi-ioned by increasing the column temperature duringhe run. Ninhydrin, mixed continuously with the col-mn eluent, reacts in a highly specific manner with the
eparated amino acid present. An in-line colorimeter
mc
nauB(amaem9raR
S
twd1tniinDiwcctca(dolN
R
miviawt
naa5wmh(mOA5sN2sN
ivSmsosnaaNs(DhttrshDnaNrp
Ns1(dto
453NURR1 AND MESENCEPHALIC DOPAMINERGIC NEURONS
easures the adsorbance of the amino acid–ninhydrinomplex at two wavelengths, 570 and 440 nm.To measure the biochemical properties of cholinergic
eurons, we determined the activity of ChAT. ChATctivity in the tissues of Str, FC, and Hip was assayedsing Fonnum’s methods with some modification (5).riefly, the brain tissues were homogenized in 1:50
v/v) cold 10 mM PBS (pH 7.4) containing 1 mM EDTAnd centrifuged at 10,000g for 10 min. Twenty-fiveicroliters of supernatant was incubated with 75 µl of
ssay medium containing 0.2 mM [3H]acetyl-CoA (Am-rsham Co. Arlington Heights, IL; sp act 217 mCi/mol), 5 mm choline chloride, and 0.15 mM eserine in
6-well plate microwells at 37°C for 45 min. Theeaction was terminated by transferring it to cold PBSnd determined in Fonnum’s scintillation solution by aockbeta LKB counter.
RESULTS
elective Loss of DA Neurons in SN and VTA Regions
In Nurr1-deficient mice, there was a complete deple-ion of TH-positive neurons in SN and VTA regions,hile the immunoreactivity of TH-positive neurons iniencephalon and hypothalamus was not affected (Fig., Table 1). In parallel with the TH-positive cell loss,here was a decrease in the number of NeuN-positiveeuronal nuclei in the SN and VTA regions, which
ndicates the loss of mesencephalic TH immunoreactiv-ty (TH-IR) was most likely due to the loss of DAergiceurons (Fig. 2). Nurr1 deficiency did not affect theAergic neurons in OB (Fig. 1, Table 1) or TH contain-
ng cells in AM and SG (Table 1). There was a slightlyeaker TH-IR in the SN and VTA in Nurr11/2 mice
ompared with Nurr11/1 mice (Table 1). Biochemi-ally, HPLC analysis of Str DA levels confirmed thathe loss of DAergic neurons was associated with aomplete absence of DA in the Str of Nurr12/2 micend a significant decrease (39%) in Nurr11/2 miceTable 2). The DA levels in OB of Nurr12/2 miceemonstrated a 65% decrease (Table 2). However, inther DA-containing regions such as AM and RE, theevels of DA did not differ among the Nurr12/2,urr11/2, and Nurr11/1 mice (Table 2).
elative Sparing of Neurons withOther Biochemical Phenotypes
The expression of Nurr1 was widely distributed inesencephalon and rhombencephalon (18, 25), suggest-
ng that the Nurr1 may have regulatory effects onarious neurotransmitter systems. To study the broadnfluence of Nurr1 in CNS, we examined the immunore-ctivity of NEergic, 5-HTergic, and cholinergic neuronsith antibodies to DBH, serotonin-1, and ChAT, respec-
ively, in the whole brain, with a focus on LC, raphe i
ucleus, and basal forebrain regions. As shown in Fig. 1nd Table 1 the DBH-IR in Nurr12/2 mice was notltered in LC but was slightly decreased in SN. The-HT-IR in dorsal, medial, and central raphe nucleusas not changed (Fig. 1; Table 1). ChAT-IR in Nurr12/2ice was intact in the basal forebrain and nuclei of
ypoglossal nerve, facial nerve, and oculomotor nerveFig. 1). A biochemical assay of NE levels in Nurr12/2
ice showed a 30% reduction in the Str, but not in theB and AM (Table 2). The NE levels in the Str, OB, andM of Nurr11/2 mice were not affected (Table 2).-HIAA, a major metabolic product of 5-HT, was notignificantly different in the Str and OB amongurr12/2 mice, Nurr11/2, and Nurr11/1 mice (Table). The levels of GLU and GABA and activity of ChAT ineveral brain areas were not altered among theurr12/2, Nurr11/2, and Nurr11/1 mice (Table 2).
DISCUSSION
The development of midbrain DAergic neurons isnitiated at embryonic day 10 in the mouse in theentrolateral neural tube adjacent to the floor plate.everal genes have been found to regulate the develop-ent of mesencephalic DAergic neurons, including
onic hedgehog (SHH) and Nurr1 (19, 23). SHH is onef a family of vertebrate genes related to the Drosophilaegment polarity gene hedgehog, which is expressed inotochord and floor plate cells and can induce appropri-te ventral cell types in the basal forebrain, spinal cord,nd DAergic neuron phenotype in the midbrain (23).urr1 is a member of the orphan nuclear receptor gene
uperfamily, which starts to express from embryonicE) day 10.5, 1.5 day earlier than the appearance of theA phenotype in mouse midbrain. Our previous studyas demonstrated that Nurr1 is essential for the induc-ion of mesencephalic DAergic neurons (19), possiblyhrough its interaction with Ptx-3, a novel bicoid-elated homeobox gene (22). The expression of Ptx-3tarts at E11.5 in the developing mouse midbrain andighly overlaps with the appearance of mesencephalicAergic neurons (22). The number of Ptx3-expressingeurons is reduced in PD patients and these neuronsre absent from 6-OHDA-lesioned rats (22). Althoughurr1 is not required for the induction of Ptx-3, it is
equired for the maintenance of Ptx-3-positive DAergicrecursor cells (19).Our immunohistochemical examinations of theurr12/2 mice with TH and NeuN antibodies reveal a
elective loss of DAergic neurons in SN and VTA (Figs.b, 1d, 2b, 2d, and 2f) and their axon terminals in StrFig. 1f). Analysis of Str DA levels confirms that thisefect is associated with a total absence of the neuro-ransmitter at Str axon terminals (Table 2). Our previ-us study (19) has reported that during embryogenesis,
n the absence of Nurr1, neuroepithelial cells can adapt
DNtNNN(c
FIG. 1. TH, DBH, serotonin-1, and ChAT immunostaining in thAergic neurons in the retrorubral field (A8), substantia nigra (Aurr12/2 mouse (b), 350. (c and d) High-magnification photograph
erminals in the striatum of Nurr11/1 and Nurr12/2 mouse (e and furr12/2 mouse (h), 3100. TH-IR in the caudal hypothalamic periveurr11/1 mouse (i) and in Nurr12/2 mouse (j), 350. DBH-positiveurr12/2 mouse (l), 350. Serotonin1-positive 5-HTergic neurons in t
n), 350. ChAT-positive cholinergic neurons in the basal forebrain in
e brain slides of neonatal Nurr11/1 and Nurr12/2 mice. TH-positive9), and ventral tegmental area (A10) in Nurr11/1 mouse (a) and in
from left side of the figure (a and b), respectively, 3200. TH-positive), 3200. TH-IR in the olfactory bulb (A16) in Nurr11/1 mouse (g) and inntricular nucleus (A11), arcuate nucleus (A12), and zona incerta (A13) in
NEergic neurons in the locus ceruleus in Nurr11/1 mouse (k) and inhe dorsal raphe nuclei in Nurr11/1 mouse (m) and in Nurr12/2 mouseNurr11/1 mouse (o) and in Nurr12/2 mouse (p), 350. ChAT-positive
holinergic neurons in the ocular motor nerve nucleus in Nurr11/1 mouse (q) and in Nurr12/2 mouse (r), 350.
454

aagi
srT1aA1ePVsmwnc
itOdpfttb1SmsMinNNtn
dtdcmtDadPd(mdtetciNsLmabni
ttPgsa
D
N
5
C
ssDgN1i
455NURR1 AND MESENCEPHALIC DOPAMINERGIC NEURONS
normal ventral localization and neuronal phenotype,nd as development progresses, these midbrain DAer-ic precursor cells degenerate, resulting in an increasen apoptosis of the ventral midbrain neurons.
This present study with a larger number of animalamples confirms our previous study (19) and othereports (2, 26) that Nurr1 deficiency does not affect theH-IR in the hypothalamic and OB areas (Fig. 1, Table). We have also found that TH-IR in the diencephalicrea and in the peripheral tissues such as the SG andM in the Nurr12/2 mice are completely spared (Fig., Table 1). It is interesting to know that Nurr1 is alsoxpressed in those areas (18, 25). It is reported thattx-3 is highly coexpressed with Nurr1 in the SN andTA, but not in other areas (22). Therefore, furthertudies to define the impact of Ptx-3 on the induction ofesencephalic DAergic phenotype and its relationshipith Nurr1 will be critical for understanding the mecha-isms by which Nurr1 selectively regulates the mesen-
TABLE 1
Immunostaining in DAergic, NEergic, 5-HTergic,and Cholinergic Neurons in the Brains of Nurr12/2,
Nurr11/2, and Nurr11/1 Mice
Nurr12/2 Nurr11/2 Nurr11/1
Aergic neurons or TH-1R cellsRetrorubral field (A8) 2 11 111Substantia nigra (A9) 2 11 111Ventral tegmental area (A10) 2 11 111Caudal hypothalamic periven-
tricular nucleus (A11) 111 111 111Arcuate nucleus (A12) 111 111 111Zona incerta (A13) 111 111 111Rostral hypothalamic periven-
tricular nucleus (A14, A15) 111 111 111Olfactory bulb (A16) 111 111 111Adrenal medulla 111 111 111Sympathetic ganglia 111 111 111Eergic neuronsLocus ceruleus 111 111 111Substantia nigra 11 111 111
-HTergic neuronsRaphe dorsal nucleus 111 111 111Raphe central nucleus 111 111 111Raphe medial nucleus 111 111 111holinergic neuronsBasal forebrain 111 111 111Ocular motor nerve nucleus 111 111 111Facial nerve nucleus 111 111 111Hypoglossal nerve nucleus 111 111 111
Note. Six animals in each group have been analyzed. Coronalection (20 µm) were cut with cryostat. Antibodies of TH, DHB,erotonin-1, and ChAT have been employed to immunostain theAergic neurons, NEergic neurons, 5-HTergic neurons, and choliner-ic neurons respectively in the brains of Nurr12/2, Nurr11/2, andurr11/1 mice. Semiquantitative analysis has been represented11, 11, and 2 as no change, slight decrease, and total loss of
mmunoreactivity respectively compared to wild-type mice.
ephalic DAergic neuron development. In addition, our s
mmunohistochemical and biochemical analyses showhat the DA transmitter, not the DAergic neurons in theB, are significantly decreased (Fig. 1, Table 2). Theecrease in DA levels in the OB of Nurr12/2 micerobably results from the loss of efferent projectionrom mesencephalon–OB pathway (1). Furthermore,his study demonstrates that besides 5-HT and Ach,wo other neurotransmitters GLU and GABA in therain of Nurr12/2 mice are relatively preserved (Fig., Tables 1 and 2). A moderate decreased NE-IR in theN and reduced NE levels in the Str (Tables 1 and 2)ay suggest that NE neurotransmission in the nigro-
triatal system is affected in Nurr1-deficient mice.oreover, our present study, using NeuN immunostain-
ng, documents that most of the ventral midbraineurons in the SN and VTA are lost in newbornurr12/2 mice, which further support the notion thaturr1 is essential for both survival and final differentia-
ion of ventral mesencephalic late DAergic precursoreurons into a complete DAergic phenotype (19).The results of this study are of interest because,
espite some differences, there is significant overlap inhe alteration of neurotransmitter phenotypes in Nurr1-eficient mice and that seen in PD (8). In PD, signifi-ant loss of DAergic neurons in SN and VTA andarkedly decreased DA levels in striatal terminals are
he most characteristic neurotransmitter changes, whileAergic neurons in diencephalon and hypothalamusre less affected (4). About 70% of PD patients show OBysfunction and anosmia may precede other signs ofD (7). It is not clear whether OB dysfunction in PD isue to local OB periglomerular DAergic neuron losswhich has been found in PD but not in Nurr1-deficientice) or is due to the mesencephalon–OB terminal
egeneration. Besides DA, the biochemical changes inhe brains of patients with PD may affect to somextent other neurotransmitter systems. For instance,he levels of NE in the Str of PD patients are signifi-antly reduced, and these changes have also been seenn Nurr1-deficient mice in this study. In addition, LCEergic neurons are partially lost in PD patients, but
pared in Nurr12/2 mice. Ptx-3 is not expressed in theC (22), which may account for the difference. Otherajor neurotransmitters such as 5-HT, Ach, GABA,
nd neuropeptides are not consistently altered in therains of PD and it is not known whether changes ineurotransmitter systems other than DA are involved
n the symptoms or progression of the disease.The similarity of central neurotransmitter dysfunc-
ion between Nurr12/2 mice and PD patients raiseshe question of a possible role of Nurr1 expression inD or other neurodegenerative disorders affecting DAer-ic neurons. Our previous study showed that the ab-ence of Nurr1 in the mouse resulted in increasedpoptosis of ventral midbrain neurons (19), indicating a
urvival role of Nurr1 for the mesencephalic DAergic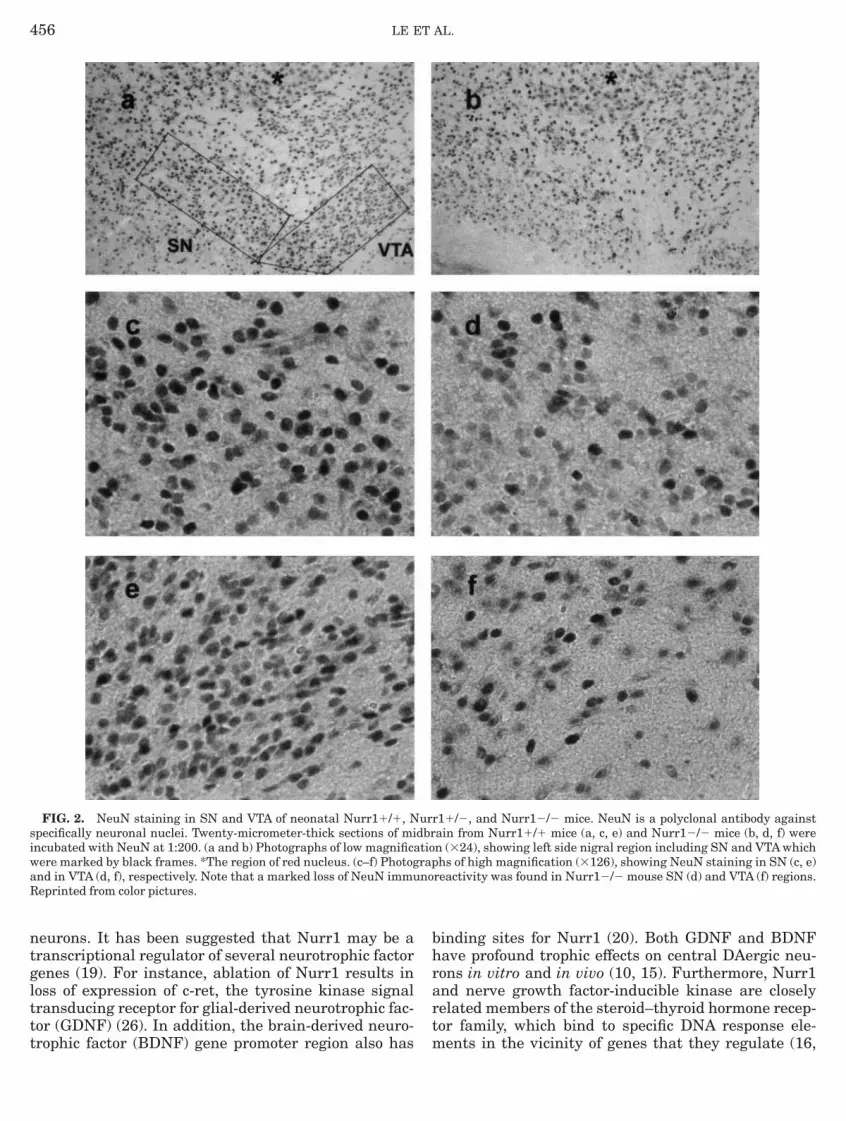
ntglttt
bhrart
siwaR
456 LE ET AL.
eurons. It has been suggested that Nurr1 may be aranscriptional regulator of several neurotrophic factorenes (19). For instance, ablation of Nurr1 results inoss of expression of c-ret, the tyrosine kinase signalransducing receptor for glial-derived neurotrophic fac-or (GDNF) (26). In addition, the brain-derived neuro-
FIG. 2. NeuN staining in SN and VTA of neonatal Nurr11/1, Npecifically neuronal nuclei. Twenty-micrometer-thick sections of mincubated with NeuN at 1:200. (a and b) Photographs of low magnificaere marked by black frames. *The region of red nucleus. (c–f) Photognd in VTA (d, f), respectively. Note that a marked loss of NeuN immueprinted from color pictures.
rophic factor (BDNF) gene promoter region also has m
inding sites for Nurr1 (20). Both GDNF and BDNFave profound trophic effects on central DAergic neu-ons in vitro and in vivo (10, 15). Furthermore, Nurr1nd nerve growth factor-inducible kinase are closelyelated members of the steroid–thyroid hormone recep-or family, which bind to specific DNA response ele-
r11/2, and Nurr12/2 mice. NeuN is a polyclonal antibody againstain from Nurr11/1 mice (a, c, e) and Nurr12/2 mice (b, d, f) weren (324), showing left side nigral region including SN and VTA whichhs of high magnification (3126), showing NeuN staining in SN (c, e)
reactivity was found in Nurr12/2 mouse SN (d) and VTA (f) regions.
urdbrtiorapno
ents in the vicinity of genes that they regulate (16,

1ens
rptsFdpqdNtstNtt
Tf5
1
1
1
1
1
1
1
1
D
N
5
G
G
C
iGsfC0S
457NURR1 AND MESENCEPHALIC DOPAMINERGIC NEURONS
7). Based on these results, it is reasonable to hypoth-size that changes in Nurr1 expression could influenceeuronal survival under a number of different circum-tances.Multiple factors such as genetic defects, aging, envi-
onmental toxins, and oxidative stress have been hy-othesized to influence DAergic neuronal susceptibilityo degeneration in PD (6, 11). In adult mice, we havehown that Nurr1 expression is reduced with age (14).urthermore, 1-methyl-4-phenyl-1,2,3,6-tetrahy-ropyridine, a neurotoxin which is believed to actrimarily via mitochondrial impairment and subse-uent oxidative injury (3, 11), produces greater nigralamage and delayed recovery of DA levels in the Str ofurr11/2 mice than in Nurr11/1 controls (14). Al-
hough the relationship between Nurr1 expression andusceptibility to DAergic neuronal loss in PD remainso be established, these data suggest that alteration ofurr1 expression may have effects in the adult and
herefore may be relevant to the pathogenesis or selec-
TABLE 2
The Levels of DA, NE, SHIAA, GLU, GABA,and ChAT Activity in the Different Tissues of Nurr11/1,
Nurr11/2, and Nurr12/2 Mice
Nurr11/1 (n) Nurr11/2 (n) Nurr12/2 (n)
AStr 664 6 71 9 408 6 50* 11 10 6 5*** 11OB 145 6 28 8 130 6 19 7 65 6 14** 10AM 2,950 6 318 4 3,100 6 456 5 3,245 6 527 5RE 52 6 10 4 47 6 5 4 55 6 9 4EStr 227 6 45 9 207 6 13 11 160 6 33* 11OB 126 6 49 8 113 6 35 7 96 6 25 10AM 14,587 6 2,780 4 16,850 6 3,250 4 17,150 6 2,130 4
-HIAAStr 412 6 21 9 438 6 23 11 399 6 45 11OB 385 6 68 8 401 6 52 7 353 6 70 10LUStr 2,838 6 264 4 2,650 6 330 4 2,552 6 292 4TC 2,205 6 272 4 2,110 6 248 4 1,985 6 250 4Hip 2,515 6 270 4 2,685 6 360 4 2,358 6 358 4ABAStr 814 6 130 4 776 6 136 4 677 6 135 4TC 666 6 102 4 779 6 126 4 738 6 162 4Hip 704 6 58 4 692 6 128 4 689 6 231 4
hATStr 8.9 6 0.8 4 8.1 6 0.9 4 8.0 6 1.4 4FC 4.8 6 0.3 4 4.7 6 0.4 4 4.2 6 0.7 4Hip 6.7 6 0.8 4 6.0 6 0.7 4 5.9 6 0.9 4
Note. The tissues of Str, OB, AM, RE, FC, and Hip were dissected ince from neonatal mice. The levels of DA, NE, 5-HIAA, GLU, andABA are represented as means 6 SEM of ng/g wet tissue. Str,
triatum; OB, olfactory bulb; AM, adrenal medullar; RE, retina; FC,rontal cortex; TC, temporal cortex; Hip, hippocampus. The values forhAT activity are represented as means 6 SEM of nmol/mg/h. * P ,.05, ** P , 0.01, and *** P , 0.001 compared with Nurr11/1 mice.tudent t test.
ive vulnerability of midbrain DAergic neurons in PD.
ACKNOWLEDGMENT
We are grateful to Mrs. Silvia Briones for her technical support.his work was supported by The Methodist Research Grants, a grant
rom the Houston Area Parkinson Society, and a grant from NIH (DK2429).
REFERENCES
1. Bjorklund, A., S. Kleinau, and M. Goldstein. 1984. Distribu-tional maps of tyrosine-hydroxylase-immunoreactivity neuronsin the rat brain. In Handbook of Chemical Neuroanatomy (A.Bjorklund and A. Hokfelt, Eds.), Part 2, pp. 55–122. Elsevier,Amsterdam.
2. Castillo, S. O., J. S. Baffi, M. Oalkovits, D. S. Goldstein, I. J.Kopin, J. Witta, M. A. Magnuson, and V. M. Nikodem. 1998.Dopamine biosynthesis is selectively abolished in substantianigra/ventral tegmental area but not in hypothalamic neuronsin mice with targeted disruption of the Nurr1 gene. Mol. Cell.Neurosci. 11: 36–46.
3. Fahn, S., and G. Cohen. 1992. The oxidative stress hypothesis inParkinson’s disease: Evidence supporting it. Ann. Neurol. 32:804–809.
4. Fearnley, J., and A. Lees. 1994. Pathology of Parkinson’sdisease. In Neurodegenerative Diseases (D. B. Calne, Ed.), pp.545–554. Saunders, Philadelphia.
5. Fonnum, F. 1975. A rapid radiochemical method for the determi-nation of choline acetyltransferase. J. Neurochem. 24: 407–409.
6. Gasser, T. 1998. Genetics of Parkinson’s disease. Ann Neurol44(Suppl. 1): S53–S57.
7. Hawkes, C. H., B. C. Shephard, and S. E. Daniel. 1997.Olfactory dysfunction in Parkinson’s disease. J. Neurol. Neuro-surg. Psychiatry 62: 436–446.
8. Hedera, P., and P. J. Whitehouse. 1994. Neurotransmitters inneurodegeneration. In Neurodegenerative Diseases (D. B. Calne,Ed.), pp. 97–117. Saunders, Philadelphia.
9. Jenner, P., and W. Olanow. 1998. Understanding cell death inParkinson’s disease. Ann. Neurol. 44:(Suppl 1): S72–S84.
0. Knusel, B., J. W. Winslow, A. Rosenthal, et al. 1991. Promotionof central cholinergic and dopaminergic neuron differentiationby brain-derived neurotrophic factor but not neurotrophin 3.Proc. Natl. Acad. Sci. USA 88: 961–965.
1. Langston, J. W. 1998. Epidemiology versus genetics in Parkin-son’s disease: progress in resolving an age-old debate. Ann.Neurol. 44(Suppl 1): S45–S52.
2. Law, S. W., O. M. Conneely, F. J. DeMayo, and B. W. O’Malley.1992. Identification of a new brain-specific transcription factor,Nurr1. Mol. Endocrinol. 16: 2129–2135.
3. Le, W. D., J. R. Bostwick, and S. H. Appel. 1992. Use of3H-GBR12935 to measure dopaminergic nerve terminal growthinto rat striatum. Dev. Brain Res. 67: 375–377.
4. Le, W. D., Y. He, O. M. Conneely, J. Jankovic, and S. H. Appel.1999. Reduced Nurr1 expression increases the vulnerability ofmesencephalic dopaminergic neurons to MPTP-induced injury.J. Neurochem. (in press).
5. Lin, L. F., D. H. Doherty, J. D. Lile, et al. 1993. GDNF: A glial cellline-derived neurotrophic factor for midbrain dopaminergicneurons. Science 260: 1130–1132.
6. Milbrandt, J. 1988. Nerve growth factor induces a gene homolo-gous to the glucocorticoid receptor gene. Neuron 1: 183–188.
7. Perlmann, T., and L. Jansson. 1995. A novel pathway forvitamin A signaling mediated by RXR heterodimerization with
NGFI-B and Nurr1. Genes Dev. 9: 769–782.
1
1
2
2
2
2
2
2
2
458 LE ET AL.
8. Saucedo-Cardenas, O., and O. M. Conneely. 1996. Comparativedistribution of Nurr1 and Nur77 nuclear receptors in the mousecentral nervous system. J. Mol. Neurosci. 7: 51–63.
9. Saucedo-Cardenas, O., J. D. Quintana-Hau, W. D. Le, M. Smidt,et al. 1998. Nurr1 is essential for the induction of the dopaminer-gic phenotype and the survival of ventral mesencephalic latedopaminergic precursor neurons. Proc. Natl. Acad. Sci. USA 95:4013–4018.
0. Shintani,A., Y. Ono, Y. Kaisho, and K. Igarashi. 1992. Character-ization of the 5-flanking region of the human brain-derivedneurotrophic factor gene. Biochem. Biophys. Res. Commun. 182:323–332.
1. Slocum, R. H., and J. G. Cummings. 1991. Amino acid analysisof physiological samples. In Techniques in Diagnostic HumanBiochemical Genetics (F. A. Hommes, Eds.), pp. 87–126. Wiley–
Liss, New York.2. Smidt, M. P., H. A. S. Schaick, C. Lanctot, et al. 1998. Ahomeodomain gene Ptx3 has highly restricted brain expressionin mesencephalic dopaminergic neurons. Proc. Natl. Acad. Sci.USA 94: 13305–13310.
3. Wang, M. Z., P. Jin, D. A. Bumcrot, et al. 1995. Induction ofdopaminergic neuron phenotype in the midbrain by sonic hedge-hog protein. Nature Med. 1: 1184–1188.
4. Wickelgren, I. 1997. Getting the brain’s attention. Science 278:35–37.
5. Zetterstrom, R. H., R. Williams, T. Perlmann, and L. Olson.1996. Cellular expression of the immediate early transcriptionfactors Nurr1 and NGFI-B suggests a gene regulatory role inseveral brain regions including the nigrostriatal dopaminesystem. Mol. Brain Res. 41: 111–120.
6. Zetterstrom, R. H., L. Solomin, L. Jansson, et al. 1997. Dopa-mine neuron agenesis in Nurr1-deficient mice. Science 276:
248–250.




























