Embed Size (px)
Citation preview

198
BRES 17223
Brain Research, 566 (1991) 198-207 (~) 1991 Elsevier Science Publishers B.V. All rights reserved. 0006.8993/911503.50
Selective expression of Jun proteins following axotomy and axonal transport block in peripheral nerves in the rat: evidence for a role
in the regeneration process
J.D. Leah 1, T. H e r d e g e n 2 a n d R. B r a v o 3
Z Schooi of Science, Grif~nth University, Brisbane (Australia), 211. Ph ysiologisches Institut, Universit~t Heidelberg, Heidelberg ( E R. G. ) and Department of Molecular Biology, Squibb Institute for Medical Research, Princeton (U.S.A.)
(Accepted 16 July 1991)
Key words: Jun protein; Axotomy; Axonal transport; Regeneration; Gene expression
Expression of the protein products of the immediate-early genes (IEGs), members of the los, ~an and krox families (Jun, Fos, and Krox, resp.) was investigated in the spinal cord and sensory ganglia (DRG) of normal rats; and following transection of, block of axonal transport in, or electrical stimulation of their peripheral axons. The nuclei of many moto- and DRG neurons showed a faint basal immunoreactivity (IR) for Jun proteins, but not for los or Krox proteins. There was a strong and selective induction of Jun-IR in moto- and DRG neurons after peripheral nerve transection or crush, or colchicine- or vinblastine-induced block of axonal transport. The Jun-IR induced by nerve transection disappeared after nerve regeneration. In contrast, Jun, Fos and Krox proteins were all induced transynaptically in spinal dorsal horn neurons following electrical stimulation of the C-fibers in the afferent nerves. Thus in differentiated neurons in vivo these lEG proteins can be expressed either independently or concomitantly depending on the type of stimulus.
INTRODUCTION
The immediate-early genes (IEGs) belonging to the los, jun and krox families are some of the earliest genes expressed following cellular stimulation 4, Their nuclear protein products have been demonstrated to bind to spe- cific DNA sequences and to control the expression of other genest'S't°'26'27'~2 '43'4'. The different members of the Ju~ (c-Jun, Jun B and Jan D) and Fos (c-Fos, Fos B, Fra-1 and Fra-2) families of proteins have been shown to form heterodimers whose binding activity and tran- scriptional activity is several-fold higher than the Jun proteins alone s'°'tt'ts'23'34-s6'st. In the adult nervous sys- tem these IEGs have been shown to have a basal ex- pression in certain groups of neurons; for example jan has a basal expression in spinal moto- and preganglionic neuro,~s, and c-los, c-/un and krox.24 all have a basal expression in many neurons in the cerebral cortex ta'tT' 18.5o Additionally, all 3 can be induced in central neu- rons following many types of physiological and pharma- cological stimulation t'ta'tT'ts,4°,4t. For example, they are induced in spinal neurons by transynaptic excitation fol- lowing noxious stimulation in the periphery, or by in- trathecal administration of the neuroactive peptide sub- stance pt8.2o.25. The exact roles of these genes in ma-
ture differentiated neurons remains to be clarified. They are known to control transcription of the enkephalin gene47; and generally they are likely to have a role in producing the persisting changes in neuronal functioning that often occur following prolonged afferent stimulation or axotomy t4'3°'32'48. Here we have u~ed immunoc) - tochemistry to investigate the production of the protein products of these IEOs in spinal and sensory ganglion neurons following transection of, or chemically induced block of axonal transport in their peripheral axons. The results suggest that Jun has a role in the regeneration process that follows peripheral nerve transection 'n.
MATERIALS AND METHODS
All experiments used made Sprague-Dawley rats (250-350 g) anesthetized with pentobarbital sodium, (Nembutal, 35 mg/kg, i.p). The sciatic or saphenous nerve were exposed by an incision in the mid-thigh and (a) a 1 mm segment was crushed with a pair of fine forceps, or (b) the nerve was ligated with 5-0 thread, transected and a 10 mm segment removed distally, or (c) a small cotton ball soaked in 10 mM colchicine or 0.3 mM vinblastine in Tyrode, or 1% capsaicin in vegetable oil, was applied to a 3 mm segment of the intact nerve for 15 min to block axonal transport. In other rats the ventral roots were ligated and transected after being exposed by a xm,ll sacral laminectomy. To retrogradely label axotomized neurons, the proximal end of the sciatic nerve was placed in a 2% solution of Fast blue (Iliing, K.G.) in distilled water for 20 rain im- mediately after transection. The nerve end was then washed exten-
Correspondence: J.D. Leah, School of Science, Griflith University, Nathan, Brisbane, Qld. 4111, Australia.

IVTP a-c-JUN IVTP a-c-FOS IVTP a-KROX-24 I II I / I I I i' II I
MW ~ MW ,L ,L MW ~'
199
45-
JUN
/
69-
46- ~ , ~
A B
6 9 - W ~:
m c
N ~ i ~ ¸ ~
Fig. 1. Specificity of the antibodies. A: in vitro translated proteins (IVTP) Jun B, c-Jun and Jun D immunoprecipitated with anti-c-Jun (a-c-Jun) antibody 636/5. B: in vitro translated c-Fos, Fos B and Fra-1 immunoprecipitated with anti-c-Fos (a-c-Fos) antibody 689/5. C: in vitro translated Krox-20 and Krox-24 immunoprecipitated with anti-Krox-24 (a-Krox-24) antibody 802/1.
sively with saline. After such treatments the wounds were sutured and the animals allowed to recover under observation. In a further set of experiments the exposed sciatic nerve was placed on a pair of platinum electrodes and stimulated wit 0.5 V, 0.05 ms pulses at 50 Hz for 10 min to activate the A/3-fibers, or at 30 V, 0.5 ms pulses at 5 Hz for 10 min to activate both the A- and C-fibers.
After various times (1 h to 15 months) the animals were rean- esthetized with Nembutal and perfused through the aorta with 100 ml phosphate-buffered saline (PBS, pH 7.4) followed by 150 ml 4% paraformaldehyde in 0.1 M phosphate buffer (PB, pH 7.4). The spinal cord and lumbar sensory ga:~glia were then removed, post. fixed overnight in the same fixative, cryoprotected in 30% s,crose
JUN and FOS in L/. spinal cord of rat Transection of sciatic nerve
2 hours 2/. hours /. days 20 days FOS
Fig. 2. Time course of the expression of Jun and Fos proteins in lumbar spinal neurons following sciatic nerve transection. Both Jun-lR (upper row) and Fos-IR (lower row) are induced in ipsilateral (left side of the cord) transynaptically excited dorsal horn neurons (dots) within 2 h of nerve transection. Most of these are in the medial laminae I/II (numbers, shaded area), where sciatic nerve nociceptive C.fiber afferents terminate. The lower boundary of lamina II is marked by the dashed line. A basal Jun-IR, but no basal Fos-IR is faintly present in some normal ventral horn motoneurons (circles at 2 h). The intensity of this Jun-lR, and the number of labelled motoneurons increases ipsilaterally 20 h after sciatic nerve transection and persists for more than 20 days (circles in left ventral horn 24 h-20 days).

2 0 0
in PB and 50 om sections cut on a cryostat. These were incubated as free floating sections for I h in a blocking solution of PBS con- taining 02% Triton X-100 (PBST) plus 3% normal goat serum; and then for at least 48 h at 4 °C in one of the primary antisera diluted in PBST containing 1% normal goat serum. The previously char- acterized antisera against the Jun, Krox and Fos proteins were used at dilutions of 1:4000 to 1:15,00G. A detailed characterization of these antisera will be described elsewhere (Kovary and Bravo, in preparation). Briefly, the rabbit anti-Jun antibody, Jun 636/5, was raised against amino acids 80-334 of the c-Jun protein. As deter- mined by immunoprecipttation analysis of in ~:ro translated pro- teins this antiserum recognized the c-Jun protein efficiently and also showed a strong cr&~.s-reactivity with both the Jun B and Jun D proteins (Fig. 1A). This antiserum thus recognizes all members of the Jun family. The rabbit anti-Fos antibody, Fos 689/5, was raised against the complete c-Fos molecule• lmmunoprecipitation analysis shows a strong reaction with c-Fos, no cross-reactivity with Fos B and a weak reaction with one of the Fos-related antigens, Fra-1 (Fig. IB). This antiserum thus recognizes both members of the Fos family• The Krox-24 antiserum, Krox 802/1, showed a strong reac- tion with the Krox-24 protein and a very weak cross-reactivity with the Krox-20 protein with which it shares some sequence homology (Fig. 1C).
After incubation in the primary antisera the sections were washed in 3 changes of PBST, incubated at room temperature in biotinated goat anti.rabbit antiserum for ! h, washed and then inc,~t.ed far 2 h in avidin-biotin-horseradish peroxidase complex (Vectastain, Vector Laboratories). After a final wash the sections were devel-
oped in 20 mg/100 mi diaminobenzidine and 0.01% hydrogen per- oxide in PBS containing 0.01% cobalt chloride and 0.01% nickel ammonium sulphate, the development of immunoreaoivity (IR) being observed under the microscope. In some instances the sec- ond antibody used was fluorescently labelled with Texas red. The sections were then air dried, mounted, and the stained cells exam- ined.
RESULTS
For all these studies we have used Jun, Fos and Krox
antisera that recognize c-Jun, Jun B and Jun D; c-Fos,
Fos B and F ra l ; and Krox-24 respectively. Fig. 1 shows
the immunoprecipi ta t ion data for each of these three
antisera. We first examined the basal expression of the IEGs in
the normal spinal cord and sensory ganglia. In the cord
of normal rats there was a weak but distinct basal nu-
clear staini~g for Jun, but not for Fos or Krox proteins,
in both somatic (Figs. 2 and 5b,d) and visceral motoneu-
rons and in sensory ganglion neurons (Fig. 6a,b). A de-
tailed description of the basal IEO-IR in the spinal cord
,! t
-~.. .__.~-. ~ . '..:
""l , ';~,~ ,i i
f f , '1
'= , I
, i
i *
. i
b
\ ' I
I t
/
1
~.. ,.~"--. --- .-~ ,~ • . o,'¢: ~ . , .~ .
I t I I
C
~ '
h,
\
I
o
' I ilml " I
I '
Fig. 3. Lumbar dorsal horn neurons with nuclei stained for (a) Jun, (b) Fos, and :c) Krox 2 h after sciatic nerve transection• Note that most neurons with stained nuclei are located in the medial superficial dorsal horn, with fewer in the deeper laminae of the dorsal horn• The dashed line indicates the border of the dorsal horn grey matter. Bar = 500 ~m.

(and brainstem) is presented elsewhere z't~a'tT'm. Ligation and transection of the sciatic r~crve ~.nduced,
within 1 h, intense .lun-IR, Fos-IR and Krox-IR in 60- 100 dorsal horn neuronal nuclei per 50 /~m section throughout that region of ipsilateral lumbar dorsal horn innervated by this nerve (Figs. 2 and 3). Labelled nuclei were only rarely seen in the contralateral dorsal horn and the basal Jun-IR in sensory ganglion and motoneu- rons was not altered. The number of these labelled nu- clei decreased markedly over the next several hours and then decreased more slowly over the next 3 weeks (Fig. 2).
This rapid transynaptic induction of the IEG proteins, and the subsequent slow decrease over the subsequent 3 weeks, is likely to have been caused by the initial bar- rage, and the subsequent decrease but persisting action potential discharge that is produced in peripheral nerves after transection ~3'~. In support of this proposal we found that pretreatment of the sciatic nerve with lidocain for 10 min proximal to the site of trm~section, to pre-
201
vent action potentials propagating to the spinal cord, completely abolished this induction of IEG-IR in the dorsal horn. A similar rapid appearance and distribution of these IEG proteins could be produced by direct elec- trical stimulation of the sciatic nerve Ab- plus C-fibers, but not A# fibers alone, as has been described previously ~aa'l~'m. Such brief electrical stimulation of the sciatic nerve only rarely produced any IEG-IR in the nuclei of motoneurones, and never in the nuclei of neu- rons of the stimulated sensory ganglia.
In contrast to the almost immediate induction of all IEOs in many dorsal horn neurons following activity in afferent nerves, there was a more slowly appearing and persisting induction of Jun, but not Fos or Krox proteins, in motoneurons and sensory ganglion neurons following peripheral axotomy (Figs. 2, 4b and 5a,c). Ten hours af- ter sciatic nerve transection intense Jun-IR, but neither Fos- nor Krox-IR began to appear in nuclei in the ipsi- lateral lumbar senso~y ganglia. After a further 10 h in- tense Jun-IR also appeared in motoneuron nuclei in the
8 b /
/ I
I / ,
I I
I I
- I
, @
I
\ \
",,"-,2.
I
D
e. O
I
I
I , . °
I
,oi • 4 ¸
' ' I , ( ~ . . o p
• ' , • " i , : , , . .
. . . . ,
° / . " ,
C ~,¢/°
/ / i
• i
\ \ \
\ / ~ . . . . _ _ - - - - - - --- .~" - - ' ~ , ' .
%', '
......... ~', ~,~
/ t
d , /
/ /
/ .
/ / '
I
f , ° " Y'
• . .
n : . ' ~ * ' - ~ , ~ ' '" I
I ' " " ' " ~ ' ' ~ " .~ . r , o .
i
Fig. 4. Jun, los and Krox immunostaining in spinal motoneurons following axotomy, a: drawing of a cross-section of the lumbar spinal cord indicating (dashed line) the region of the ventral horn shown in the following photographs, b: intense Jun-IR in ventral horn motoneurons 4 days after sciatic nerve transection. The ventral horn grey matter in this and the following figures is outlined with a dashed line. c,d: the ventral horn stained for Fos and Krox, 4 days after sciatic nerve transection, no staining of motoneurons or other ventral horn neurons is evident. Bar in d = 200 #m for b, c and d.

202
ipsilaterai ventral horn. Four days after transection or crush of the sciatic nerve this Jun-IR was maximally present in many ipsilateral spinal motoneurons (Fig. 2) and sensory ganglion neurons. This Jun-IR was not present in all these neurons because they do not all send their peripheral axons through the sciatic nerve. The in- duced Jun-IR was particularly intense and immediately distinguish~.ble from the faint basal Jun-IR present in these neurons in the normal rat (Figs. 5b,d and 6a,b). Transection of the ventral roots, which contain only mo- toneuron axons, produced intense Jun-IR only in mo- toneurons, whereas transection of the saphenous nerve, which contains sensory but not motoneuron axons, I~ro- duced Jun-IR only in sensory neurons. When the sciatic nerve was transected and also ligated to prevent regen- eration, the induced intense Jun-IR in motoneurons de- creased after 40-60 days when the axotomized motoneu- rons begin to degenerate. In contrast, the induced Jun-IR in this case persisted in most sensory ganglion neurons for up to 15 months after axotomy.
When axotomized neurons were retrogradely labelled
by applying Fast blue to the transected nerve, and im- munocytochemically processed with Texas red labelled second antibodies for detecting Jun expression 7 days later, the intense Jun-IR was seen to be induced prima- rily in those moto- and sensory neurons that had been axotomized (Fig. 7). Thus 88% of the sensory ganglion neurons whose perikarya were retrogradely labelled with Fast blue applied to their transected axons showed in- tense nuclear Jun-IR, whereas those not retrogradely la- belled with Fast blue showed only the usual faint basal nuclear Jun-IR. There were a small number of Fast blue- labelled (i.e. axotomized) neurons not strongly stained for Jun. These sensory neurons were most likely those having axons in both the sciatic nerve and another hind- limb nerve 3t, and so were not completely axotomized.
This persisting, intense Jun-IR induced by axotomy also appeared in motoneurons and sensory ganglion neu- rons when examined 4 days after the sciatic nerve was simply transected without ligation, or crushed and al- lowed to regenerate unimpeded. When these sciatic nerves were allowed to complete regeneration over 3
/ / , , . s /
/ ' I s ) •
I . " 0 I o • •
• • • ° q) ,
I °
I ' \ \ / ' \ \ / \ \ / \
\ , \ \ /
y .-..
a ,I// b /// I
I I
I , . /
/ , o
/ r /
/
C o • d l
0
Q
0
O
Q
f Fix. 5. Jun-IR in normal and axotomized spinal motoneurons, a: Jun-IR in ventral horn motoneuron nuclei 4 days after axotomy, b: Jun-IR in the normal ventral horn. c: higher magnification of axotomy-induced Jun-IR in spinal motoneurons, d: faint basal Jun-IR in the normal ventral horn showing that some nuclei (arrows) have a more intense basal immunoreactivity. Bar = 200 ~m in a, b and 40 ~m in c, d

203
8 ,A • . g .a , •
w 4 P
I
4,
d
, ~ t ~ ~ , ~Y O
• 4' ~ •
• q D ~" • • , e l l , O I ! 4
Fig. 6. Jun-IR in the nuclei of lumbar (I.4) sensory ganglion neurons, a: normal ganglion neurons: arrowheads show the faint basal Jun-IR present in the nuclei of many normal, non.axotomizcd neurons, b: intense Jun-lR in ganglion neurons following axotomy of ~he sciatic nerve. Arrows in the lower left indicate the basal Jun-IR seen in the nuclei of normal neurons whose peripheral axons do not course through the sciatic nerve and were not transected. Arrows and arrowheads in the center of the photograph point to large and small neurons, respectively, whoso nu©lei contain intense Jun-IR induced by axotomy (see also Fig. 7 ) c,d: Jun-IR in ganglion neurons after axonal transport block induced by colchicine and capsaicin treatment, respectively, of the sciatic nerve. Bar -- tOO ~m in a, 50 ~m in b and 200 ~m in c,d.
a • ~ Awl e o
@ • @
• • ,:
Fig. 7. Jun-IR in retrogradely labelled sensory ganglion neurons, a: Jun-IR in ganglion neuron nuclei 4 days after sciatic nerve transection, detected using Texas red immunofluorescence, b: the same field through different excitation and barrier filters showing those neurons ret- rogradely 18belled with Fast blue (and thus axotomized). Jun-IR is seen almost exclusively in the nuclei of the retrogradely neurons.
204
J .
a /
~ o o e
i •
\ \
\ \ "
%
." °-
Q . ~ .~ . . ,t . o _ , /
• o. ~ . " . ~ . • . f
• . . • / • . '
b
\
\
~ , "e
/
\ , \
\ • \
. ]
• ~ , . ; • /
: . • . . /
• /
/ • / /
C i /
/ ; /
I °
[ , "
I
\ . °.
' 4 ' ' , * / ' . dr . ~ • :
/ • . ., ~ . J
,. ,m
J f
d
\ \
\ -
\ \
\ \
\ , \
\ \ ,
• ' t i,," \
w . | !
• '
• t
¢t ,', • ]
/ ." Q J
\ /
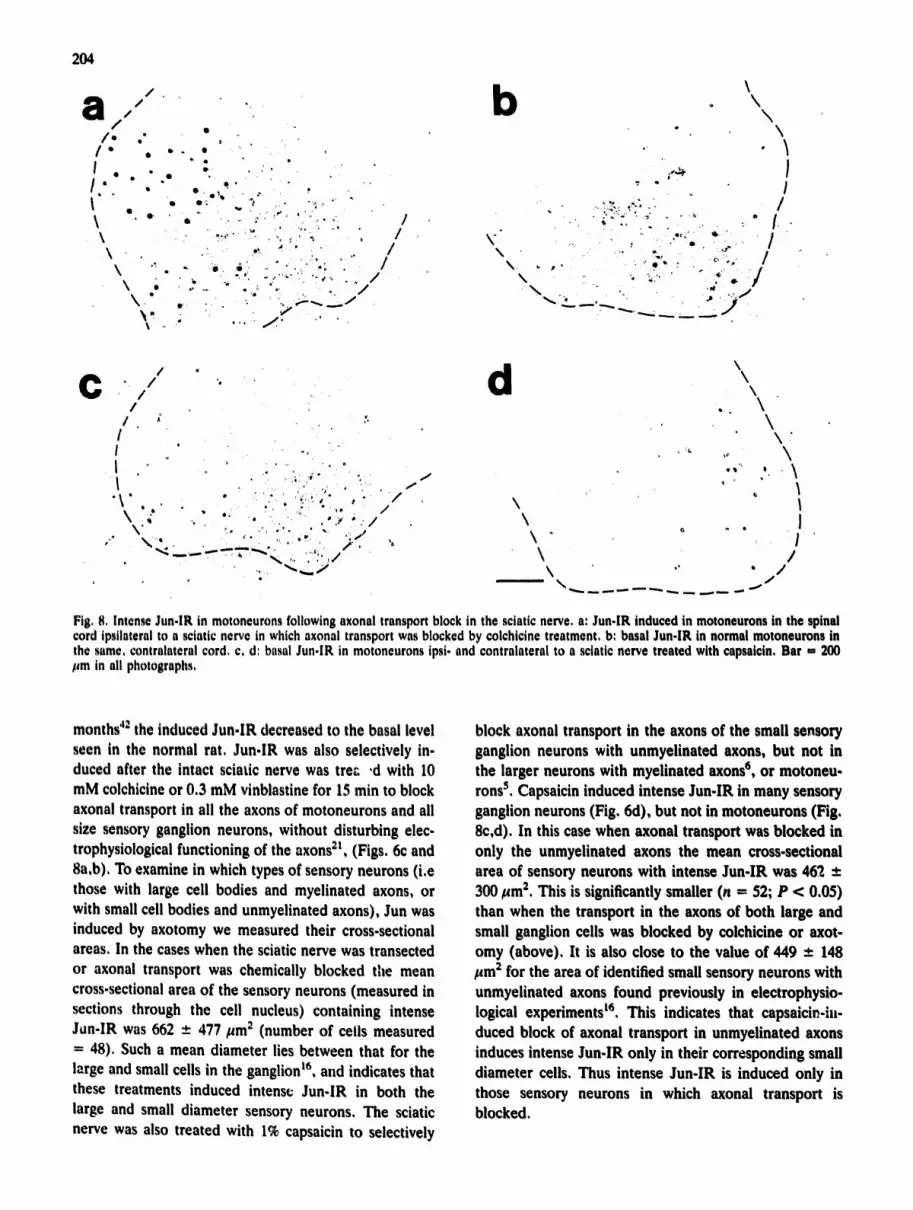
Fig. 8, Intense Jun-lR in motoneurons following axonal transport block in the sciatic nerve, a: Jun-lR induced in motoneurons in the spinal cord ipstlateral to a sciatic nerve in which axonal transport was blocked by colchicine treatment, b: basal Jun-lR in normal motoneurons in the same, contralateral cord. c. d: basal Jun-lR in motoneurons ipsi. and contralateral to a sciatic nerve treated with capsaicin, Bar - 200 lira in all photographs.
months 42 the induced Jun-lR decreased to the basal level seen in the normal rat. Jun-IR was also selectively in- duced after the intact sciatic nerve was trec ,d with 10 mM colchicine or 0.3 mM vinblastine for 15 min to block axonal transport in all the axons of motoneurons and all size sensory ganglion neurons, without disturbing elec- trophysiological functioning of the axons 21, (Figs, 6c and 8a,b), To examine in which types of sensory neurons (i.e those with large cell bodies and myelinated axons, or with small cell bodies and unmyelinated axons), Jun was induced by axotomy we measured their cross-sectional areas. In the cases when the sciatic nerve was transected or axonal transport was chemically blocked the mean cross-sectional area of the sensory neurons (measured in sections through the cell nucleus) containing intense Jun-IR was 662 - 477/~m 2 (number of cells measured - 48). Such a mean diameter lies between that for the large and small cells in the ganglion t6` and indicates that these treatments induced intense Jun-IR in both the large and small diameter sensory neurons, The sciatic nerve was also treated with 1% capsaicin to selectively
block axonal transport in the axons of the small sensory ganglion neurons with unmyelinated axons, but not in the larger neurons with myelinated axons ~, or motoneu- rons s. Capsaicin induced intense Jun-IR in many sensory ganglion neurons (Fig, 6d), but not in motoneurons (Fig, 8c,d), In this case when axonal transport was blocked in only the unmyeUnated axons the mean cross-sectional area of sensory neurons with intense Jun-IR was 462 +- 300/~m 2, This is significantly smaller (n = 52; P < 0.05) than when the transport in the axons of both large and small ganglion cells was blocked by colchicine or axot- omy (above), It is also close to the value of 449 _ 148 ~m 2 for the area of identified small sensory neurons with unmyelinated axons found previously in electrophysio- logical experiments t6. This indicates that capsaicin-iw duced block of axonal transport in unmyelinated axons induces intense Jun-lR only in their corresponding small diameter cells, Thus intense Jun-IR is induced only in those sensory neurons in which axonal transport is blocked,

DISCUSSION
Jun, Fos and Krox-24 proteins are all induced in the nuclei of many spinal neurons by direct pharmacological activation, and physiologically by stimulation of periph- eral afferent C-fibers 1s'2°'25. However, we show here for the first time that Jun proteins alone are induced in spi- nal and sensory ganglion neurons in vivo by peripheral axotomy, or block of transport in their peripheral axons. The retrograde labelling of axotomized neurons with Fast blue, and the selective block of transport in unmy- elinated axons using capsaicin both confirm that it is only in those neurons with peripheral axotomy or axonal transport block that the Jun-IR is induced. Neither the Fos nor Krox antisera stained the nuclei of axotomized neurons. It remains possible, however, that newly dis- covered members of these families, or new IEGs, will also be found to be induced in neuronal nuclei by axo- tomy or axonal transport block. We have yet to ascer- tain whether all, or only one or two members of the Jun family, i.e. c-Jun, Jun B and Jun D 19'3s'39 a re thus in- duced. Our preliminary results suggest that all the mem- bers of the Jun family can be induced in different types of neurons by axotomy. Such a result would contrast with findings that, in some cells in vitro, the different iuns can have distinct patterns of expression in response to growth factors zg, and that in the neuron-like PC-12 cells in vitro, growth factors activate c.fos, jun B and c.jun whereas membrane depolarization activates c.fos and ,iun B without c-jun 3.
Our results clearly show that the pattern of IEG ex- pression can be different when induced by occupation of membrane receptors and activation of second messenger systems by synaptic activity and by axotomy. Axotomy and block of axonal transport do not have entirely the same consequences for a neuron, but they do have in common their preventing the normal transport, via the axon, to the soma and nucleus of the compounds re- leased by the neurons' target tissues. Thus we propose that it is the absence of compound(s) normally retro- gradely transported from the peripheral tissues (skin, muscle) that signals the production of Jun in the axoto- mized sensory ganglion and motoneuronai nuclei. This is supported by the observation that when the sciatic nerve was crushed, or transected without ligation, and allowed to freely l'egenerate the induced Jun-IR reverted to normal basal levels after 3 months when the nerve reinnervates the hindlimb skin and muscle 42. The ap- pearance of Jun proteins in sensory f_anglion neurons 10 h after transection of the sciatic nerve, and subsequently in motoneurons after a further 10 h, together with the corresponding distances of approximately 40 mm and 80 mm indicates that the compound(s) are retrogradely
205
transported at a 4 mm/h, i.e. at the fast axonal transport rate. We have as yet no evidence for what the relevant compound(s) might be, but those with known trophic effects on neurons, such as nerve growth factor, are ob- ,,iou~,ly candidates. The increased production of Jun pro- teins during the course of the sciatic nerves' regenera- tion strongly suggests that they have some role in the neurochemical mechanisms of either degeneration or re- generation. Both moto- and sensory neurons undergo degeneration following axotomy. However, whereas the majority of large diameter sensory neurons with myeli- hated axons do not undergo degeneration following ax- otomy, 70% of small diameter sensory neurons with un- myelinated axons do degenerate 2z. Intense Jun-IR was found in both large and small diameter sensory neurons following axotomy. Thus the induced Jun-IR is found in both degenerating and non-degenerating sensory neu- rons, indicating that it is more likely to be involved in a regeneration process.
In vitro studies indicate that the Jun family of proteins can form heterodimers with the Fos family of proteins, and that these Jun-Fos heterodimers have an enhanced affinity for AP-1 recognition sites and transcription ac- tivity compared to Jun-Jun homodimers s'ls'a3'4s'sl. Krox-20 and Krox-24 are also co-regulated with c-Fos ~' a6,z7 and Krox-24 is co-expressed with Jan and Fos in central neurons 16a, and may act in concert with Jun and Fos in controlling gene expression. Here we have shown that in the nervous system in vivo Jun proteins can be induced independently of Fos and Krox proteins and thus might act alone as a transcription regulator, prob- ably by forming Jun-Jun dimers Is,
The full spectrum of genes whose expression is con- trolled by Jun, Fos and Krox proteins in the nervous system remains to be determined, although it has re- cently been shown that c-Jun and c-Fos control the syn- thesis of the opioid neuropeptide enkephalin in the hip- pocampus 47. Our present and previous experments show that Fos, Jun and Krox proteins are all induced in spinal neurones by transynaptic stimulation resulting from pe- ripheral noxious stimuli 16a'~7'ts'2s. It is possible that in this case they control the increased production of opioid neuropeptides which occurs in spinal neurons following prolonged peripheral noxious stimulation 37'49. Jun pro- teins alone are induced following axotomy and axonal transport block and most likely initiate the production of a different set of proteins. These could mediate the changes in electrical and neurochemical properties that occur in sensory neurons after axotomy 4s. Our results also indicate that Jun proteins have a role during nerve regeneration. Thus a possibility of considerable interest is that these proteins could also control the expression of those genes required for axonal regeneration 46.

206
Acknowledgements. We thank C. Weis, G. Overesch and S. Dav- ies for excellent assistance, and Frau Manasali for the diagrams. Supported by the DFG and NH&MRC.
REFERENCES
1 Angel, P., Allegretto, E.A., Okino, S.T,, Hattori, K., Boyle, W.J., Hunter, T. and Karin, M., Oncogene jun encodes a se- quence-specific trans-activator similar to AP-1, Nature, 332 (1988) 166-171.
2 Anton, E, Herdegen, T., Peppel, E and Leah, J.D., C-los like immunoreactivity in rat brainstem neurons following noxious chemical stimulation of the nasal mucosa, Neuroscience, 41 (1991) 629-641.
3 Bat'tel, D.E, Sheng, M., Lau, L.E and Greenberg, M.E., Growth factors and membrane depolarization activate distinct programs of early response gone expression: dissociation of fos and jun induction, Genes Dev., 3 (1989) 304-313.
4 Bravo, R., Growth factor inducible genes in fibroblasts. In A. Habenicht (Ed.), Growth Factors, Differentiation Factors and Cytokines, Springer, Berlin, 1990, pp. 324-343.
5 Buck, S.H. and Burks, T.F., The neuropharmacology of capsa- icin: review of some recent observations, Pharmacol. Rev., 38 (1986) 179-226.
6 Cameron, A.A., Leah, J.D. and Snow, P.J., The electrophysi- ological and morphological characteristics of feline dorsal root ganglion cells, Brain Research, 362 (198G) 1-6.
7 Chavrier, P., Janssen-Timmen, U., Mattei, M.G., Zerial, M., Bravo, R. and Charnay, P., Structure, chromosome location, and expression of the mouse zinc finger gene krox.24: multiple gone products and coregulation with the proto-oncogene c.fos, Mol. Cell Biol., 9 (1989) 787-797.
8 Chiu, R., Boyle, W.J., Meek, J., Smeal, T., Hunter, T. and Karin, M., The c-fos protein interacts with c.jun/AP.l to stim- ulate transcription of AP-I responsive genes, Cell, 54 (1988) 541-.552,
9 Chiu, R., Angel, P. and Karin, M., Jun B differs in its biolog. ical properties from, and is a negative regulator of c./un, Ceil, 59 (1989) 979-986,
10 Christy, B.A. and Nathans, D., DNA-binding site of the growth factor-inducible protein ZIF 268, Proc. Nati. Acad. Set. U.S.A.. 86 (1989) 8737-8741.
II Cohen, D.R., Ferreira, P.C.P,, Oentz, R,, Franza Jr., B.R, and Curran, T., The product of a/as-related gone, fra.l, binds co- operatively to the AP.I site with Jun: transcription factor AP-I is comprised of multiple protein complexes, Genes Day., 3 (1989) 173=184.
12 Draganow, M. and Robertson, H.A,, Localization and indue. tion of c-fos protein-like immunoreactive material in the nuclei of adair mammalian neurons. Brain Research, 440 (1988) 252- 260,
13 Devor, M., The pathophysiology and anatomy of damaged nerve, in P. Wall and R. Melzack (Eds,), A Textbook of Pain, Churchill-Livingstone, London, 1984, pp. 49-64.
14 Goelet, E, Castellucci, V.E, Schacher, S, and Kandel, E,R., The long and the short of long-term memory: a molecular framework, Nature, 322 (1986) 419-422,
15 Halazonetis, T.D., Georgeopoulos, K., Greenberg, M,E, and Leder, E, C-jun dimerizes with itself and with c-fos, forming complexes of different DNA binding affinities, Cell, 55 (1988) 917-924.
16 Harper, A.A, and Lawson, S,N., Conduction velocity is related to morphological cell type in rat dorsal root ganglion neurones, J. Physiol., 359 (1985) 31-46.
16a Herdegen, T., Kovary, K,, Leah, J,D, and Bravo, R,, Specific temporal and spatial distribution of JUN, FOS and KROX-24 proteins in spinal neurons following noxious transsynaptic stim- ulation, J. Camp, Neural., 312 (1991) in press.
17 Herdegen, T., Leah, J.D., Manisali, A,, Bravo, R, and Zim-
mermann, M., c-JUN-like immunoreactivity in the CNS of the adult rat: basal and transynaptically induced expression of an immediate early gene, Neuroscience, 41 (1991) 643-654.
18 Herdegea, T., Leah, J.D., Zimmermann, M. and Bravo, R., The KROX-24 protein, a new transcription regulating factor: expression in the rat central nervous system following afferent somatosensory stimulation, Neurosci. Lett., 120 (1990) 21-24.
19 Hirai, S.-I., Ryseck, R.-P., Mechta, F., Bravo, R. and Yaniv, M., Characterization of junD: a new member of the jan proto- oncogene family, EMBO J., 8 (1989) 1433-1439.
20 Hunt, S.P., Pini, A. and Evans, G., Induction of c-fos like pro- tein in spinal cord neurons following sensory stimulation, Na- ture, 328 (1987) 632-634.
21 Jackson, 1), and Diamond, J., Colchicine block of cholinesterase transport in rabbit sensory nerves without interference with the long-term viability of the axons, Brain Research, 130 (1977) 579-584.
22 Janig, W. and McLachlan, E., On the fate of sympathetic and sensory neurons projecting into a neuroma of the superficial peroneal nerve in the cat, J. Camp. Neural., 225 (1984) 302- 311.
23 Kouzarides, T. and Ziff, E., The role of the leucine zipper in the fos-jun interaction, Nature, 336 (1988) 632-634.
24 Kruijer, W., Cooper, J.A., Hunter, T. and Verma, I.M., Plate- let-cXerived growth factor induces rapid but transient expression of the c.fos gene and protein, Nature, 312 (1984) 711-716.
25 Leah, J.D., Herdegen, T. and Zimmermann, M., Physiological and pharmacological induction of fos in spinal dorsal horn neu- rones. In E Cervera, P.M, Hedly and O. Bennett (Eds.), Sen. sory ProcesSing in the Dorsal Horn of the Spinal Cord, Elsevier, Amsterdam, 1989, pp. 307-310,
26 Lemarie, P., Relevant, O., Bravo, R. and Charnay, P., Two mouse genes encoding potential transcription factors with iden- tical DNA-binding domains are activated by growth factors m cultured ceils, Proc, Natl. Acad. Sci, U,S.A,, 85 (1988) 4691- 4695.
27 Lemarie, P,, Vesque, C., Schmitt, J,, Stunnenberg, H., Reiner, F. and Charnay, P,, The serum-inducible mouse gone Krox.24 encodes a sequence specific transcriptional activator, Mol. Cell Biol., 10 (1988) 3456-3467,
28 Lombard, M,C, and Larabi, Y,, Electrophysiological study of cervical dorsal horn cells in partially deafferented rats, In J,J. Bonica and A. Iggo (Eds.), Advances in Pain Research and Therapy, Raven, New York, 1983, pp, 147-154.
29 Li, L., Hu, J.L, and Oisen, E,N,, Different members of the jan proto-oncogene family exhibit distinct patterns of expression in response to type beta transforming growth factor, J, Biol. Chem,, 265 (1990) 1556-1562.
30 Marx, J.L,, The los gene as a 'master switch', Science, 327 (1987) 854-857.
31 McMahon, S,B. and Wall, P,D,, Physiological evidence for branching of peripheral unmyelinated sensory afferent fibers in the rat, J, Cmnp, Neural,, 261 (1987) 130-136,
32 Morgan, J,l, and Curran, T,, Stimulus-transcription coupling in neurons: role of cellular immediate-early genes, Trends Neuro. sci,, 12 (1989) 459-462,
33 Muller, R,, Bravo, R,, Burckhard, J, and Curran, T., Indl, c- tion of c-los gone and protein by growth factors precedes acti. ration of c-myc, Nature, 312 (1984) 716-720,
34 Nakabeppu, Y,, Ryder, K, and Nathans, D., DNA binding ac- tivities of three murine jun proteins: stimulation by los, Cell, 55 (1988) 907-915.
35 Nishina, H,, Sate, H, Suzuki, T,, Sate, M. and Iba, H., Isola- tion and characterization of fra-2, an additional member of the Jos gone family, Prec. Natl. Acad. Sci. U.S.A., 87 (1990) 3619-

3623. 36 Rauscher, R.J., Sambucetti, L.C., Curran, T., Distel, R.J. and
Spiegelmann, B.M., Common DNA binding site for los protein complexes and transcription factor AP-1, Cell, 52 (1988) 471- 480.
37 Ruda, M.A., ladarola, M.J., Cohen, LV. and Young, W.S., In situ hybridization histochemistry and immunocytochemistry re- veal an increase in spinal dynorphin biosynthesis ":u a rat model of peripheral inflammation and hyperalgesia, Proc. Natl. Acad. Sci. U.S.A., 85 (1988) 622-626.
38 Ryder, K., Lau, L.F. and Nathans, D., A gene activated by growth factors is related to the oncogene v-jun, Proc. Natl. Acad. Sci. U.S.A., 85, 1487-1491.
39 Ryseck, ER., Hirai, S.I., Yauiv, M. and Bravo, R., Transcrip- tional activation of c-jun during Go/Gt transition in mouse fi- broblasts, Nature, 34 (1988) 535-537.
40 Saffen, D.W., Cole, A.J., Worley, EE, Christy, S.A., Rydel, K. and Baraban, J.M., Convulsant-induced increase in tran- scription factor messenger RNAs in the rat brain, Proc. Natl. Acad. Sci. U.S.A., 85 (1988) 7795-7799.
41 Sagar, S.M. and Sharp, ER., Light induces a Fos-like antig.en in retinal neurons, Mol. Brain Res., 7 (1990) 17-21.
42 Sanders, K.H. and Zimmermann, M., Mechanoreceptors in rat glabrous skin: redevelopment of function after nerve crush, Z Neurophysiol., 55 (1986) 644-659.
43 Sassone-Corsi, E, Der, C.J. and Verma, I.M., Ras-induced neuronal differentiation of PC-12 cells: possible involvement of los and jun, Mol. Cell Biol., 9 (1989) 3174-3183.
44 Schonthal, A., Herrlich, E, Rahmsdorf, J.H. and Ponta, H., Requirements for .los 8ene ex~re.~sion in the transcriptional ac-
207
tivation of collagenase by other oncogenes and phorbol esters, Cell, 54 (1988) 325-334.
45 Schuermann, M., Neuverg, M., Hunter, J.B., Jenuwein, T., Ryseck, R.P., Bravo, R. and Muller, R., The leucine repeat motif in los protein mediates complex formation with jun/AP-1 and is required for transformation, Cell, 56 (1989) 507-516.
46 Snipes, G.J., Costello, B., McGuire, C.B., Mayes, B.N., Bock, S.S., Norden, J.J. and Freeman, J.A., Regulation of specific neuronal and nonneuronal proteins during development and fol- lowing injury in the rat central nervous system. In F. Seil, E. Herbert and B.M. Carlson (Eds.), Progress in Brain Research, Vol. 71, Elsevier, Amsterdam, 1987, pp. 155-175.
47 Sonnenberg, .LL., Rauscher, F.J., Morga~'l, J.I. and Curran, T., Regulation of proenkephalin by fos and jun, Science, 246 (1989) 1622-1625.
48 Titmus, M.J. and Faber, D.S., Axotomy-induced alterations in the electrophysiological characteristics of neurons, Prog. Neu- robiol., 35 (1990) 1-51.
49 Weihe, E., Millan, M.J., Leibold, A., Nohr, D. and Herz, A., Co-localization of proenkephalin- and prodynorphin-derived opioid peptides in laminae IV/V spinal neurons revealed in ar- thritic rats, Neurosci. Leu., 85 (1988) 187-192.
50 Wilkinson, D.G., Bhatt, S., Ryseck, R.E and Bravo, R., Tis- sue-specific expression of c-jun and jun B during organogenesis in the mouse, Development, 106 (1989) 465-47129.
51 Zeriai, M., Toschi, L., Ryseck, R.E, Schuermann, M., Muller, R. and Bravo, R., The product of a novel growth factor acti- vated gene, los B, interacts with jun proteins enhancing their DNA binding activity, EMBO J., 8 (1989) 805-813.





























