Embed Size (px)
Citation preview

Ž .Brain Research 823 1999 11–23
Research report
Serotonin-immunoreactive neurones in the visual system of the prayingmantis: an immunohistochemical, confocal laser scanning and electron
microscopic study
Gerd Leitinger a,b,), Maria Anna Pabst a, Karl Kral b
a Institut fur Histologie und Embryologie, Karl Franzens-UniÕersitat Graz, Harrachgasse 21, A-8010 Graz, Austria¨ ¨b Institut fur Zoologie, Karl Franzens-UniÕersitat Graz, UniÕersitatsplatz 2, A-8010 Graz, Austria¨ ¨ ¨
Accepted 8 December 1998
Abstract
Ž .The distribution, number, and morphology of serotonin-immunoreactive 5-HTi neurones in the optic lobe of the praying mantisTenodera sinensis were studied using conventional microscopy and confocal laser scanning microscopy. Five or six 5-HTi neuronesconnect the lobula complex with the medulla, and at least 50 5-HTi neurones appear to be confined to the medulla. In addition, a fewlarge 5-HTi processes from the protocerebrum supply the lobula complex, and two large 5-HTi processes from the protocerebrum ramifyin the medulla and lamina, where they show wide field arborisations. In order to provide a basis for understanding the action of serotoninin the lamina, the ultrastructure of its 5-HTi terminals was examined by conventional and immunohistochemical electron microscopy. The5-HTi profiles were filled with dense core vesicles and made synapses. Output synapses from 5-HTi profiles outnumbered inputs by about3 to 1. The terminals of the 5-HTi neurones were in close contact with cells of various types, including large monopolar cells, but closeapposition to photoreceptor terminals was rare, and no synapses were found between 5-HTi terminals and photoreceptor terminals.q 1999 Elsevier Science B.V. All rights reserved.
Keywords: Immunohistochemistry; Serotonin; Insect; Praying mantis; Optic lobe; Lamina
1. Introduction
ŽThe biogenic monoamine serotonin 5-hydroxytryp-.tamine, 5-HT is known to be released by neurones to
Ž w x.modulate neuronal signalling processes e.g., Refs. 9,37Ž w x.and insect behaviour e.g., Ref. 8 . Insects show intense
Ž5-HT-immunoreactivity within the optic lobes e.g., Refs.w x.20,32 . Anatomical, physiological and pharmacologicalfindings suggest that serotonin influences the diurnalrhythm of the sensitivity of insect visual neuronesw x6,14,16,42,58,59 .
The praying mantis visual system is frequently studiedas a model for insect vision and visually guided behaviourw x22,23,25,26,29,30,40,41,45–48,60 . The serotonin contentof the praying mantis optic lobe has been quantified by
w xHPLC measurements 13 , but so far there has been noaccount of the morphology of the serotonin-
) Corresponding author. Department of Neurobiology, Medical School,University of Newcastle upon Tyne, Newcastle upon Tyne, NE2 4HH,UK. E-mail: [email protected]
immunoreactive neurones in the praying mantis optic lobe.Our aim was to provide a basis for understanding serotoninaction in the praying mantis optic lobe. Thus, we usedimmunocytochemical staining for serotonin to show thepathways, number and distribution of serotonin-immunoreactive neurones in the praying mantis optic lobe.Furthermore, we used electron microscopic immunocyto-chemical methods to examine the first optic neuropile, thelamina, where the photoreceptor terminals are in synapticcontact with the large monopolar cells, the first orderinterneurones. The purpose of this electron microscopicanalysis was to obtain data about the connectivity patternbetween 5-HTi neurones and other neurones in the lamina.
2. Materials and methods
2.1. Animals
Thirty-two mantids used for the histological analysesŽ .were of the species Tenodera sinensis Saussure . They
0006-8993r99r$ - see front matter q 1999 Elsevier Science B.V. All rights reserved.Ž .PII: S0006-8993 98 01339-0

( )G. Leitinger et al.rBrain Research 823 1999 11–2312
w xwere raised under the conditions described in Ref. 40 . Allthe specimen were prepared on the third day after moultingbetween 1000 and 1100 in order to obtain a consistent timein the diurnal rhythm. The insects were cooled on ice forabout 8 min before the preparation started.
2.2. Serotonin immunohistochemistry
2.2.1. Immunohistochemistry on paraffin embedded sec-tions
For paraffin embedding, the animals were anaesthetisedwith CO and half heads were cut under fixative. The half2
heads were fixed in Bouin fixative for 3 h, placed in 80%alcohol, further dehydrated and embedded in HistosecŽ .Merck . Sections were cut at 8 mm thickness, mounted onprecoated slides and deparaffinised. Peroxidase labellingwas performed using a ‘labelled streptavidin biotin kit’Ž .Dako , which consists of blocking medium, a biotinylatedsecondary antibody and a streptavidin–biotin–horse radishperoxidase complex. Sections were preincubated withblocking medium from the kit, and afterwards, a rabbit
Žpolyclonal antiserum raised against serotonin 0601, Im-.munotech was applied at a dilution of 1:100 for 2 h at
378C, followed by incubations in biotinylated secondaryantibody and in the streptavidin–biotin–horse radish per-oxidase complex. Visualisation of the immunoreaction wasbrought about by applying 1 mgrml diaminobenzidine and
Ž .0.02% hydrogen peroxide DABrH O in 0.013 M PBS2 2
at pH 7.2 for 7 min. The sections were then rinsed indistilled water, dehydrated and mounted in gum damar.
2.2.2. Immunofluorescent stainingImmunofluorescence studies of serotonin localisation
were performed on vibratome sections. The brain and opticŽlobe of mantids were dissected under ice-cold fixative 2%
paraformaldehyde, 0.1% glutaraldehyde and 0.2% picric.acid in 0.1 M phosphate buffer at pH 7.4 and left in the
fixative for 3 h at room temperature. After rinsing in 0.1 Mphosphate buffer, the specimens were embedded in 14%gelatin, quickly cooled to 48C and sectioned at 50 mm
Ž .thickness on a vibratome Leica Vt 1000 cooled with ice.The sections were immunostained with a free floating
w xprocedure 53 . Incubations were performed in a solutionŽ .of 0.8% bovine serum albumin BSA , 0.1% gelatin and 2
mM sodium azide in PBS containing 0.1% Triton X-100Ž .PBS-Tx at pH 7.4. For rinsing, PBS-Tx was used for3=15 min after the incubation steps. The vibratome sec-tions were bathed in 0.05 M glycine in PBS for 10 min andpreincubated in 5% normal goat serum for 45 min. The
Žpolyclonal rabbit antiserum raised against serotonin 20080,.Inc Star was applied overnight at 48C at a dilution of
1:10 000. After rinsing, the vibratome sections were incu-bated in a goat-anti-rabbit secondary antibody conjugated
Ž .to Cy3 Jackson Immunoresearch at 1:400 dilution for 2 hŽ .and mounted on precoated slides SuperFrostPlus; Menzel .
After drying in a dark room for 6 h to achieve adherenceto the slides, the sections were rehydrated in distilled waterfor 15 min, dehydrated in an ethanol series and cleared inmethyl salicylate. Then they were mounted in MowiolŽ . ŽHoechst containing 4% diazabicyclooctane DABCO;
.Sigma .Immunofluorescently stained neurones were examined
Žin a Leica TCS NT confocal microscope excitation wave-length: 568 nm; KrAr laser; detection with a longpass 590
.nm filter . Optical sections were recorded at intervals of1–1.5 mm and projected to one plane in red pseudocolour.Counts of 5-HTi cell bodies were repeated on five individ-ual mantids. Since it is difficult to obtain an objective
w xthreshold criterion for positive staining 38 , the number ofcell bodies counted varied from specimen to specimen.Measurements of the maximum diameter of somata weretaken using Quantimed Image Analysis software and re-peated on two individual mantids. Consecutive vibratomesections of one specimen were aligned in Adobe Photo-shop and the outlines of single neurones were traced.
2.2.3. Immunohistochemistry for epoxy resin embeddingFor embedding in epoxy resin, the same dissection,
fixation, and sectioning procedures as for immunofluo-rescent staining were followed, except that vibratome sec-tion thickness was 70 mm. Free floating vibratome sec-tions were immunolabelled and then embedded in TAAB
Ž .epoxy resin ‘pre-embedding labelling’ . Some sectionswere stained for serotonin with the peroxidase method, andothers with an immunogold method.
2.3. StreptaÕidin–biotin–peroxidase method
The vibratome sections were permeabilised in an ethanolŽ .series 10, 30, 50, 30 and 10% ethanol in PBS pH 7.4 and
preincubated for 10 min in 0.05 M glycine in PBS pH 7.4and for 30 min in 5% normal swine serum diluted in bufferŽPBS pH 7.4 containing 0.8% BSA, 0.1% fish gelatin, and
.2 mM sodium azide . This was followed by incubationover night at 48C in polyclonal rabbit antiserum against
Ž .serotonin Inc Star diluted 1:6000 in buffer, rinses inŽ .buffer, biotinylated swine anti-rabbit antibody Dako di-
luted at 1:500 in buffer for 2 h, rinses in buffer, strepta-Žvidin–biotin–horse radish peroxidase complex Dako; from
.the LSAB Kit, diluted 1:5 in PBS pH 7.4 for 2 h, a rinsein PBS pH 7.4; DABrH O in PBS pH 7.4 for 5 min, and2 2
a last rinse in PBS pH 7.4.
2.4. Immunogold procedure
After permeabilisation as above, the vibratome sectionswere preincubated in 0.05 M glycine in PBS pH 7.4 and in5% normal goat serum in the same buffer. Afterwards, the
Ž .polyclonal rabbit antiserum against serotonin Inc Stardiluted 1:6000 in buffer was applied over night at 48C, the

( )G. Leitinger et al.rBrain Research 823 1999 11–23 13
sections were rinsed, and the immunogold-silver stainingwas performed as follows. Incubation with 1 nm gold
Žconjugated goat-anti-rabbit secondary antibody AuroProbe.One GAR, Amersham at 1:50 or 1:200 dilution in buffer
for 4 h at room temperature; rinse in 150 mM NaNO ; and3Ž .a silver enhancement step IntensE M, Amersham , with
one part of a 33% gum arabic solution added to two partsw xof the silver enhancement solution 54 . The development
lasted 30 to 45 min and the sections were rinsed in 150mM NaNO after the silver enhancement step. For the3
subsequent gold toning reaction, we adapted a protocolw x w x28 using 0.05% tetrachloroauric III acid in 150 mM ac-etate buffer, pH 5.6 for 7 min and with a rinse in 150 mMNaNO after the gold toning step.3
After both immunolabelling methods, the vibratomesections were post fixed in 2% glutaraldehyde, 2% para-formaldehyde, 0.2% picric acid for 10 min, rinsed in 0.1 Mphosphate buffer pH 7.4, and post fixed in 1% OsO for4
30 min. They were then dehydrated and embedded inTAAB epoxy resin. Semi- and ultrathin sections were cuton a Reichert Ultracut ultramicrotome. The semithin sec-tions were viewed under a Zeiss Axiophot and photo-graphed, while the ultrathin sections were collected oncopper grids, stained in uranyl acetate and lead citrate andexamined in a Zeiss EM 902 transmission electron micro-scope.
Electron micrographs of the lamina of five differentanimals, stained with the immunogold procedure, wereused to identify the spatial relationship between the 5-HTicells and neighbouring neurones, to perform measurementsof vesicle diameters and to count the incidence of synapses.For all these measurements, 100 immunoreactive profileswere randomly chosen from single cross sections of thelamina. Diameters of vesicles were measured on micro-graphs transferred to Quantimed Image Analysis software,and the size distribution of vesicles was further analysedusing GraphPad software.
2.5. Controls
For each different approach, negative controls weremade on some sections by incubating in buffer instead ofthe primary antiserum. On other sections, the secondaryantiserum was omitted. All of the controls remained un-stained. Furthermore, the antiserum against serotonin byImmunotech at a dilution of 1:400 was preadsorbed in 1.5
Ž .mM serotonin Sigma overnight at 48C and then appliedŽ w x.to paraffin sections see Ref. 57 . This resulted in total
suppression of the staining.
3. Results
3.1. Morphology of the optic lobe
To our knowledge, this is the first account of thestructure of the praying mantis optic lobe. An overview of
the lamina, medulla and lobula complex neuropiles isgiven in Fig. 1A. The axons of the eight photoreceptors of
Ž .each ommatidium R1–8 form bundles with neighbouringR-axons as they leave the compound eye and pass thedistal layer of the lamina, the first optic neuropile.
Due to similarities in morphology between locust andpraying mantis eyes and optic lobes, there is no reason toassume differences in the pathway of R1–8 from thelocust, where R1–8 project into the underlying laminaneuropile and six short R-axons of each ommatidium end
Žw xin a corresponding cartridge 31 ; cartridges named afterw x. Ž .4 . The remaining locust R-axons long axons traverse to
w xthe outer layer of the medulla neuropile 31 .In the praying mantis, each cartridge consists of the
R-axons of one ommatidium surrounding at least threeŽaxons of large monopolar cells LMCs, first order in-
.terneurones . At the proximal base of the lamina, thecartridges disintegrate where LMC-axons and R-axons tra-
Ž .verse to the medulla in the external optic chiasm Fig. 3D .In the medulla, a column corresponds to each laminacartridge. The medulla neuropile is divided into two parts,the outer and the inner neuropile, by a ‘serpentine layer’Ž w x.as in the fly, 55 . The outer neuropile is close to theexternal optic chiasm. Fibre tracts leave the inner medullaand traverse to the lobula complex in the internal opticchiasm. The lobula complex in the praying mantis consistsof four distinct parts, named lobula 1 to 4, from posteriorto anterior in relation to the body axis.
3.2. Light microscopical immunohistochemistry for sero-tonin
All three optic neuropile regions are invaded by sero-Žtonin-immunoreactive 5-hydroxytryptamine-immuno-
.reactive, 5-HTi processes. Intertwining 5-HTi ramifica-tions form a fine network in the lobula complex, and they
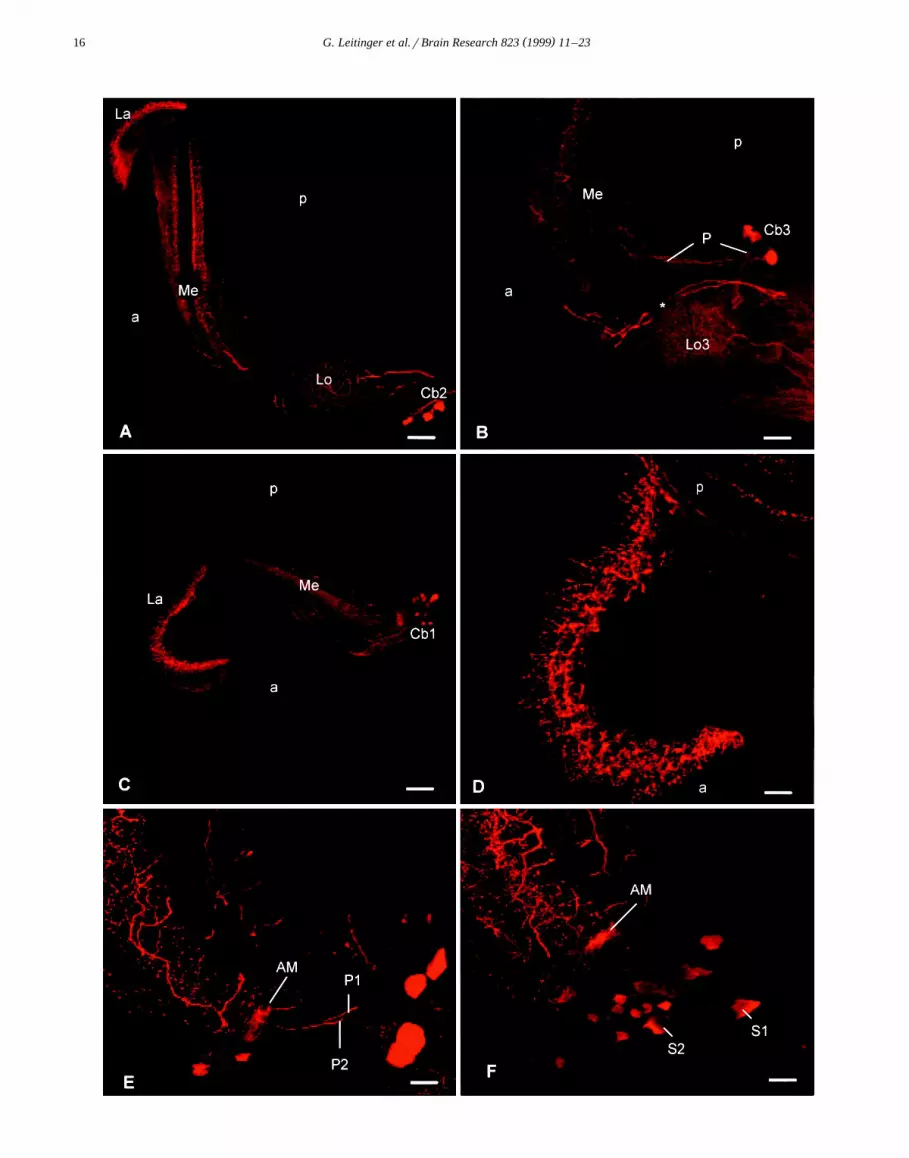
Ž .can be detected in five layers in the medulla Fig. 1A ;three layers in the outer medulla are more strongly stainedthan the two remaining layers and their extent can bedetected on Fig. 3B. In the lamina, varicose 5-HTi pro-
Ž .cesses form one single proximal layer Fig. 3C,D . The5-HTi processes clearly surround each set of LMC-axons
Ž .in each cartridge Fig. 3D .Three groups of 5-HTi cell bodies can be distinguished
on the basis of their location and the course of theirprimary neurites. Fig. 1 gives an overview of all 5-HTisomata observed in the optic lobe of the praying mantisand of their position in relation to processes of 5-HTineurones. The first group contains five to six large, stronglyimmunoreactive somata, named Cb3, situated in the poste-rior proximal side of the lobula complex. Their primary
Ž .neurites are directed towards the lobula 3 Fig. 2B . Theneurites send small branches into lobula 2, and they appearto turn and send processes into the lobula 1 and then
Ž .invade the inner medulla Fig. 1A . Fig. 1B shows a

( )G. Leitinger et al.rBrain Research 823 1999 11–2314
reconstruction of one neurone of the Cb3 group fromconsecutive vibratome sections.
In the second group, 10–15 cell bodies were situated inŽ .the anterior lobula complex Fig. 3A , with their primary
Ž .neurites leading in the direction of the midbrain Fig. 2A .We named this group of cell bodies Cb2.
A third group of at least 10 anterior somata was namedŽ .Cb1 Fig. 3B . The primary neurites of Cb1 somata enter
the medulla by its anterior side and ramify in the medullaŽ .neuropile Fig. 2C . The primary neurite of at least one
ovoid soma with a diameter of 20 mm directly leads to theanterior edge of the medulla and ramifies in an area
Ž .identified as the accessory medulla Fig. 2E,F . In additionto the larger cells in group three, there are approximately50 smaller 5-HTi somata whose primary neurites cannot be
Ž .detected. Fig. 2C . They extend alongside the anteriormedulla. The immunoreaction in these cells is of variableintensity. The number and diameter of all three groups ofsomata are listed in Table 1.
We distinguished two types of large neurites that con-nect the protocerebrum to the optic lobe. However, due tosuperposition of other 5-HTi processes, we have not identi-fied the site of their somata. The first type of neurites
consists of several 5-HTi processes of large diameter thatenter the lobula complex from the protocerebrum. Theyramify into a fine network of 5-HTi fibres in the lobula 2
Ž .and 3 Fig. 2A . The second type shows two large, stronglyimmunoreactive processes passing from the protocerebrum
Žbetween lobula 2 and 3 towards the anterior medulla Fig..2B and the lamina. These neurites invade the medulla
from the anterior, and they appear to ramify in three mainlayers in the medulla. Branches derived from the sameneurites were found running along the anterior medullaand through the first optic chiasm toward the posterior
Ž .lamina Fig. 2D . From there, these neurites send arborisa-tions toward the anterior side of the lamina and furtherarborise into multiple fibres that run along each lamina
Ž .cartridge Fig. 2D . Fig. 1C shows a reconstruction of themain branches of one of these neurites from consecutivevibratome sections.
In summary, the lobula mainly appears to be suppliedby large processes from the protocerebrum and by pro-cesses derived from Cb3 neurones. The medulla, on the
Ž .other hand, is invaded by three types of neurones: 1processes of Cb1 invading the medulla from its anterior
Ž .side; 2 processes that pass between lobula 2 and 3,
Ž . Ž .Fig. 1. A Schematic drawing of the serotonin-immunoreactive 5-HTi cells. Large immunostained processes enter the optic lobe from the protocerebrumand send ramifications to the lobula complex and toward the medulla and lamina. Within the optic lobe, three groups of somata can be distinguished: Cb1,in the anterior proximal medulla, with its primary neurites directed toward the medulla, Cb2, in the anterior lobula complex, with its primary neuritesdirected toward the protocerebrum, and Cb3, in the posterior lobula complex, with its primary neurites directed towards the lobula complex and to the innermedulla. Varicose processes of 5-HTi neurones can be detected in all three optic neuropile regions; they form a fine meshwork in the lobula complex. The
Ž .medulla exhibits three main 5-HTi layers and two layers of weaker immunoreactivity. The lamina exhibits a single 5-HTi layer. B Reconstruction of theŽ .main branches of a Cb3 neurone as seen on consecutive vibratome sections. C Reconstruction of the main branches of one of the two processes
Ž .innervating the medulla and lamina as seen on consecutive vibratome sections. For clarity, some of the branches have been omitted. A–C Ce, compoundeye; Ch, external optic chiasm; iMe, inner medulla; La, lamina; Lo1 to 4, lobula 1 to 4; oMe, outer medulla; Pc, protocerebrum; a, anterior; p, posterior.Scale bars: 50 mm.

( )G. Leitinger et al.rBrain Research 823 1999 11–23 15
Ž .Fig. 1 continued .
( )G. Leitinger et al.rBrain Research 823 1999 11–2316

( )G. Leitinger et al.rBrain Research 823 1999 11–23 17
Ž . Ž .Fig. 3. A–C Interference contrast pictures of paraffin sections labelled with peroxidase for serotonin. A Horizontal sections of cell bodies of Cb2Ž .arrowhead . The 5-HTi somata are situated in the anterior lobula cortex close to other cell bodies which do not react with the antiserotonin-antibody.
Ž . Ž . Ž .Arrow: 5-HTi process entering the lobula complex Lo . Scale bar: 35 mm. B Horizontal section of a 5-HTi soma belonging to Cb1 arrow , in theŽ .anterior medulla cortex. Some other cell bodies in the vicinity show a weak reaction with the antiserum arrowheads . Me Medulla. The extent of the
Ž . Ž .5-HTi-projections can be detected in three layers numbered 1, 2 and 3 . Scale bar: 35 mm. C Sagittal paraffin section. 5-HTi processes form extensiveŽ . Ž . Ž .ramifications in one layer arrow of the proximal lamina La . This layer starts at the border between lamina and external optic chiasm Ch . Ce
Ž .Compound eye with dark screening pigment. Scale bar: 70 mm. D Semithin cross section of the proximal lamina made from a vibratome section whichŽ .was immunostained and embedded in epoxy resin. The lamina cartridges Ca can clearly be discerned; the cartridges disintegrate at the border between the
Ž . Ž . Žlamina La and the external optic chiasm Ch . Just distal to the external optic chiasm, 5-HTi processes intermingle between the LMC-axons lucent.profiles of each lamina cartridge. Scale bar: 16 mm.
apparently derived from Cb3 entering from the posteriorŽ .side; and 3 two processes, derived from the protocere-
brum, entering between lobula 2 and 3, also invading themedulla from the anterior side. The single 5-HTi layer inthe proximal lamina seems to be derived from branches ofthe latter two processes.
3.3. Fine structure of 5-HTi neurones in the proximallamina
To test whether the proximal lamina can be considereda major output region of the serotonin-immunoreactiveneurones, the ultrastructural characteristics of this region
Ž .Fig. 2. Confocal laser scanning micrographs of 5-HTi processes labelled with Cy3 conjugated secondary antibody. A–C Projections of 32 optical sectionsŽ . Ž . Ž . Ž .through horizontal vibratome sections. A Overview over the immunoreactivity in the lamina La , the medulla Me and the lobula complex Lo . A
Ž .group of 5-HTi somata is situated in the anterior lobula complex Cb2 . Their primary neurites are directed toward the protocerebrum to the right of theŽ .section. Other processes that appear to be derived from the protocerebrum supply the lobula complex. a, Anterior; p, posterior. Scale bar: 50 mm. B
Ž . Ž . Ž .Group of 5-HTi somata in the posterior lobula complex Cb3 . Processes P derived from Cb3 somata lead toward the lobula 3 Lo 3 and toward theŽ . Ž . Ž .posterior medulla Me . Two processes asterisk , derived from the protocerebrum, are directed toward the anterior medulla. Scale bar: 40 mm. C Group
Ž . Ž . Ž . Ž .of cell bodies in the anterior medulla Cb1 and ramifications in the lamina La and medulla Me . a, anterior; p, posterior. Scale bar: 50 mm. D SingleŽ .optical section of a horizontal section through the lamina. On its posterior edge p , axons exhibiting 5-HT immunoreactivity enter the lamina neuropile.
Ž .They turn to run toward the anterior a and show ramifications perpendicular to the posterior–anterior axis. Note the varicosities exhibited by theseŽ .ramifications. Scale bar: 20 mm. E,F Projections of consecutive vibratome sections of the anterior medulla. The somata situated at the anterior medulla
Ž .belong to Cb1. Two processes, P1 and P2, are directed toward the accessory medulla AM . At least P2 enters the accessory medulla. P1 and P2 appear toŽ .be derived from the somata S1 and S2 which can be seen on F . Scale bar: 20 mm.

( )G. Leitinger et al.rBrain Research 823 1999 11–2318
Table 1Number and diameter of 5-HTi somata situated in the optic lobe
5-HTi Number Mean diameter Diameter rangeŽ . Ž .somata "S.D. n mm
Ž . Ž .Cb1 )50 24.7"5.0 20 19–35 larger somataŽ . Ž .13.6"2.0 26 11–18 smaller somataŽ . Ž .Cb2 10–15 27.1"3.6 7 24–33 larger somataŽ . Ž .13.7"1.7 7 12–16 smaller somataŽ .Cb3 5–6 25.1"2.9 11 21–29
were examined closely with respect to the morphology of5-HTi terminals and their relation to surrounding neurones.
Ž .In the lamina neuropile, photoreceptor R- terminals are
readily identified in electron micrographs by their densecytoplasm; LMC-axons, on the other hand, can be distin-guished from other axons by their larger diameter and their
Ž .weak electron density Fig. 4A,B . Other, unidentifiedprofiles are small and exhibit an electron lucent cytoplasmŽ . Ž w x.Fig. 4A e.g., Ref. 27 . In the proximal layer of thelamina neuropile, profiles filled with vesicles intermingledbetween the R-terminals and LMC-axons of each laminacartridge. This can be seen on ultrathin cross sections of
Ž .the proximal quarter of the lamina neuropile Fig. 4B .Some of these vesicle rich profiles were stained with the
Ž .antiserum to serotonin, while others were not Fig. 4A,B .R-terminals, LMC-axons and the position of intervening
5-HTi profiles could be identified clearly on low power
Ž .Fig. 4. Electron micrographs showing the location of 5-HT-immunoreactivity in the proximal lamina. AB Note the profiles containing multiple vesiclesŽ . Ž .situated in the vicinity of the large monopolar cell axons LMC . Some of these profiles react with the antiserum to serotonin arrows , while others remain
Ž . Ž .unstained arrowheads . A Peroxidase method; horizontal overview. Asterisks: small, electron lucent profiles; R: photoreceptor terminals. Magnification,Ž .=8000. Scale bar: 1.1 mm. B Immunogold method; cross section of a cartridge at low magnification. Asterisk: immunostained process. Magnification,Ž .=11 000. Scale bar: 1.1 mm. C Immunogold stained profile at high magnification. Arrow: small vesicle containing a dot-like structure. Magnification,
=44 000. Scale bar: 0.25 mm.
( )G. Leitinger et al.rBrain Research 823 1999 11–23 19
electron micrographs from sections stained with peroxi-Ž .dase Fig. 4A . Two disadvantages of the peroxidase
method were that the stain only penetrated 5 to 10 mm intothe tissue, and that diaminobenzidine was precipitatedclose to membranes, obscuring details of synapses. Onimmunogold labelled sections through the proximal lam-ina, vesicles and membranous structures could be resolved.Gold particles stained the outer 20 to 30 mm of vibratomesections, although staining density was lower at the centrethan at the periphery of the sections. Since the sections
were gold toned after the silver enhancement step, we wereable to apply a post fixation with 1% OsO for a satisfac-4
Žtory preservation of membrane structure Fig. 4B,C and.Fig. 5 .
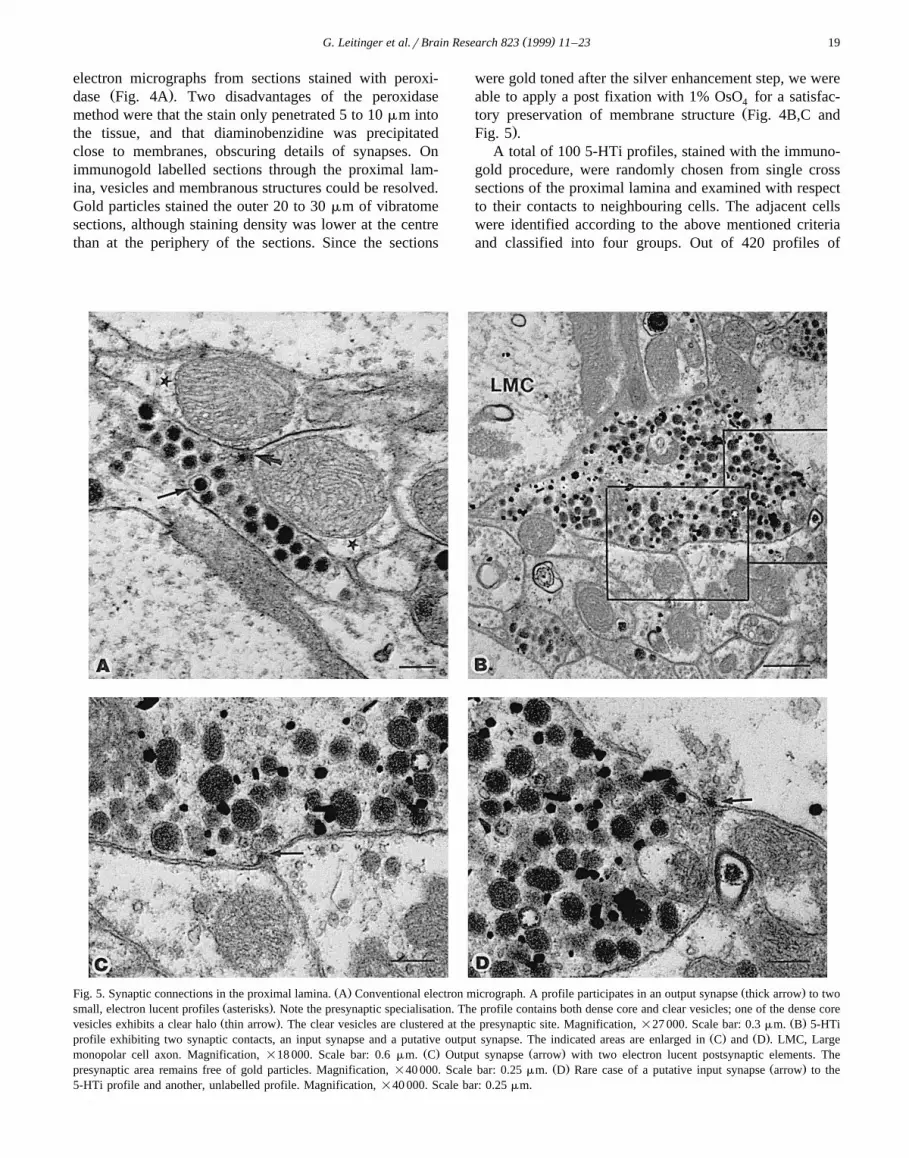
A total of 100 5-HTi profiles, stained with the immuno-gold procedure, were randomly chosen from single crosssections of the proximal lamina and examined with respectto their contacts to neighbouring cells. The adjacent cellswere identified according to the above mentioned criteriaand classified into four groups. Out of 420 profiles of
Ž . Ž .Fig. 5. Synaptic connections in the proximal lamina. A Conventional electron micrograph. A profile participates in an output synapse thick arrow to twoŽ .small, electron lucent profiles asterisks . Note the presynaptic specialisation. The profile contains both dense core and clear vesicles; one of the dense coreŽ . Ž .vesicles exhibits a clear halo thin arrow . The clear vesicles are clustered at the presynaptic site. Magnification, =27 000. Scale bar: 0.3 mm. B 5-HTi
Ž . Ž .profile exhibiting two synaptic contacts, an input synapse and a putative output synapse. The indicated areas are enlarged in C and D . LMC, LargeŽ . Ž .monopolar cell axon. Magnification, =18 000. Scale bar: 0.6 mm. C Output synapse arrow with two electron lucent postsynaptic elements. The
Ž . Ž .presynaptic area remains free of gold particles. Magnification, =40 000. Scale bar: 0.25 mm. D Rare case of a putative input synapse arrow to the5-HTi profile and another, unlabelled profile. Magnification, =40 000. Scale bar: 0.25 mm.

( )G. Leitinger et al.rBrain Research 823 1999 11–2320
neurones that were situated directly adjacent to 5-HTiprofiles, most had an electron lucent cytoplasm. Of these,
Ž .the majority 367 were classified as small, electron lucentprofiles, whereas 33 profiles were identified as LMC-axons.It is likely that the number of LMC-axons is underesti-mated here, since smaller sections of LMC-axons cannotbe distinguished from other small, electron lucent profiles.Fourteen adjacent profiles were filled with both dense coreand clear vesicles but did not stain for 5-HT, whereas onlysix profiles were identified as belonging to R cells. In nocase were two adjacent 5-HTi profiles detected.
Within the 5-HTi profiles and the other vesicle rich, butunstained profiles of similar position, at least four differenttypes of vesicle could be distinguished by their size andcontent. The two most common types were large, dense
Ž . Žcore granular vesicles and much smaller, clear agranu-. Ž .lar vesicles Fig. 4C and Fig. 5 . The latter were often
clustered in a separate part of the profile. For both celltypes—stained and unstained—the diameters of dense coreand clear vesicles were measured on 10 different profilesŽ .Fig. 6 . Both types of cell exhibited a closely similarcomposition of vesicles. In both immunostained and un-stained profiles, a few of the large dense core vesicles
Ž .exhibited a clear halo Fig. 5A and a dense dot was seenŽ .in the centre of some of the small vesicles Fig. 4C .
3.4. Spatial relationship between identified profiles
Conventional ultrathin sections and immunogold la-belled ultrathin sections of the proximal lamina werescreened for synaptic connections between identified neu-rones. Synapses with presynaptic electron dense materialin shape of a triangle or trapeze in cross section werecommon and most of them appeared to be dyadic—with
Ž .two postsynaptic elements Fig. 5A,B . These synapses
Fig. 6. Diameters of vesicles in 5-HTi profiles of the lamina and innonstaining terminals with a similar composition of vesicles. The graphshows mean values plus standard error of means: dense core vesicles instaining terminals, 129 nm, ns179, in non-staining terminals, 124 nm,ns159, clear vesicles in staining terminals, 46 nm, ns199, in non-stain-ing terminals, 50 nm, ns60.
were further characterised by a presynaptic cluster contain-ing small vesicles and an accumulation of electron dense
Žmaterial close to the postsynaptic cell membranes Fig..5C,D . Large dense core vesicles were not clustered at the
presynaptic site, which indicates they are non-synaptic.After identifying adjacent cells to the 100 5-HTi pro-
files referred to earlier, their membranes were screened forsynaptic contacts to those adjacent profiles. Nineteensynapses between 5-HTi profiles and adjacent profilescould clearly be identified; five of these were input
Ž .synapses onto 5-HTi profiles Fig. 5D , whereas the major-Ž . Ž .ity 14 were output synapses Fig. 5C from 5-HTi pro-
files. The other profiles that participated in these synapticconnections all exhibited an electron lucent cytoplasm:four of them were identified by their large diameter asLMC-axons, whereas the remaining profiles were classi-fied as small, electron lucent profiles. On two of the outputsynapses onto small electron lucent profiles, only onepostsynaptic profile could be detected on the section. Allthe other synapses were clearly dyadic. In no case was asynaptic connection between a 5-HTi profile and an R-axondetected.
4. Discussion
4.1. Localisation of 5-HT in the optic lobe
The antibody against serotonin binds to a variety ofneurones in the praying mantis optic lobe. Some of them
Žappear to be confined to a certain neuropile e.g., neurites.of Cb1 somata , whereas others seem to link different
Ž .neuropile areas with each other neurites of Cb2 somataor link the protocerebrum to the optic lobe. The two largeaxonal processes that enter the optic lobe from the proto-cerebrum and send varicose arborisations into the medullaand lamina form such links. Their branches in the laminaare filled with vesicles and make predominantly outputsynapses. Therefore, these neurones appear to be centrifu-gal. Although the location of their cell bodies could not betraced, these cells may be similar to the large bilateral
w xLBO5HT cells of flies 35 , and to the unilateral protocere-w xbral 5-HTi cells in the sphinx moth 21 . Additional 5-HTi
elements from the protocerebrum invade the praying man-tis lobula complex. The overall pattern of 5-HT immuno-reactivity is similar as in other insect species, but there aresignificant variations in number and morphology of the
Žneurones between insect species reviewed in Refs.w x.19,21,32,33 .
A major aim of the present study was to examine the5-HTi localisation in the lamina of the praying mantis. Ourexamination has shown that varicose 5-HTi processesdensely innervate a single layer in the proximal lamina,apparently all derived from two large neurites. This is incontrast to the closely related cockroaches, where many

( )G. Leitinger et al.rBrain Research 823 1999 11–23 21
small 5-HTi neurones with somata near the outer opticw xchiasm arborise in the lamina 3,36 .
4.2. Ultrastructure of the 5-HTi neurones in the lamina
The present ultrastructural analysis of 5-HTi varicositiesin the lamina of the praying mantis gives insight intopossible interactions between 5-HTi neurones and laminaneurones. Although 5-HTi varicosities are situated next toLMC axons, most of them are separated from photorecep-tor axons by at least one cell layer.
Most synaptic structures found on the 5-HTi terminalsseemed to be output synapses, with two processes post-synaptic to one 5-HTi process. The postsynaptic elementsare electron lucent and could belong to LMC-dendrites orto other, unidentified neurones in the lamina. Outputsynapses in the lamina outnumbered inputs by about 3:1,which indicates that the lamina is predominately an outputregion of the 5-HTi neurones. As in the praying mantis,output synapses to second order interneurones from 5-HTi
w xterminals are also found in ants and honey bees 34 .Moreover, synaptic contacts between 5-HTi terminals andR-terminals could be found neither in the praying mantis,nor in ants and honeybees.
In contrast to hymenoptera and orthopteroids, dipteraexhibit no synaptic connections on their 5-HTi terminalssituated distal of the lamina synaptic layer. Glial processessurround these terminals and the only other neuronal ele-ments present in this zone are R-axons and LMC-somataw x34 . Nevertheless, the 5-HTi terminals in the lamina of thepraying mantis, ants, honey bees and blowflies share twofeatures. Terminals of similar morphology as the 5-HTiterminals accompany them, but these terminals do not stainfor serotonin, whereby they also contain large, dense core
w xand small, clear vesicles 34 . An analysis of mean vesiclediameter in this study showed that there is little differencebetween terminals that stained and terminals that did notstain for 5-HT. Immunostaining for serotonin is the onlymorphological criterion so far to differentiate betweenthem at the electron microscope level.
Co-storage of dense core vesicles and clear vesicles wasalso detected in 5-HTi terminals in other parts of the
w xnervous tissue of various insects 7,49,56 . The occurrenceof various vesicle types in one cell can be interpreted as ahint for the storage of more than one neuroactive sub-stance. In moths, for example, the peptides FMRF amideand SCP were found colocalised within centrifugal 5-HTiB
w xoptic lobe neurones 20 . Whereas peptides are confined tow xthe large dense core vesicles 18 , both clear vesicles and
dense core vesicles may contain low molecular weightw xtransmitters such as serotonin 15,62,64 .
The 5-HTi terminals in the lamina of the praying mantisare densely packed with dense core vesicles that seem to
Ž w xbe ready for release at non-synaptic sites see Refs. 5,51 ;w x.non-synaptic release reviewed in Ref. 15 . Non-synaptic
release of a substance allows it to diffuse over time
through the extracellular space to its target receptors. Thiskind of interaction was defined as ‘volume transmission’w x1 . However, parts of the membranes of the 5-HTi termi-nals exhibit synaptic specialisations with clear vesicles forrapid and localised release, termed ‘wiring transmission’w x63 . Our ultrastructural studies indicate that, in the mantislamina, a single 5-HTi terminal can participate in bothtypes of transmission. Low frequency or lack of outputsynapses on 5-HTi terminals has also been reported in
w xother insects and crayfish 34,49,50,56 .
4.3. Possible function of serotonin in the optic lobe
The function of 5-HTi neurones of the optic lobe ofinsects has been associated with a diurnal modulation ofcompound eye sensitivity in species as diverse as cricketsw x w x w x w x59 , butterflies 24 , flies 42 and locusts 6 . In particular,the cells reported to be influenced by application of 5-HTwere either photoreceptor cells via modulation of potas-
w x w xsium channels 6,16 or optic lobe interneurones 42,59 .Glial cells have also been discussed as a primary target ofserotonin that would in turn regulate the activity of sur-
w xrounding neurones 42 .Mantis compound eyes exhibit pronounced diurnal
changes, the most conspicuous features of which arew xscreening pigment migration 52 and a change in rhabdom
w xdiameter 23 . The ommatidia operate during the day andw xat night, but with different angular sensitivity 44 . This
has been attributed to both adaptation processes and to aw xdiurnal rhythm 23 , but little is known about the mecha-
nisms that elicit such diurnal modifications.Our analysis of the lamina may shed some light on
which neurones could be targets of 5-HTi neurones in thepraying mantis. Since 5-HTi terminals are so rarely foundin direct apposition to photoreceptor axons, it seems un-likely that the latter are their main targets in this species.However, a diffusion of serotonin to the photoreceptor cellaxons after its release into the extracellular cleft cannot be
Ž .excluded see above . On the other hand, LMC-axons orother interneurones were frequently seen adjacent to 5-HTiterminals and were in synaptic contact with the latter. Theyare therefore much more likely candidates for being influ-enced by the 5-HTi neurones.
At least one medulla 5-HTi neurone clearly arborises inthe accessory medulla; this small neuropile has been sug-gested to play a role in co-ordinating circadian informationw x39,43,61 ; and it could thus provide information on thestate of circadian rhythm. However, we have not detectedconnections between the main processes targeting the lam-ina and the accessory medulla. If these processes belong todistinct neurones, they must have another way of beingsupplied with diurnal information. Extraocular photorecep-tors remain one possible, yet speculative, explanation.Such photoreceptors have been postulated for a number of
w xinsect species 2,12,17 and located at several diffusew xlocations in ants and beetles 10,11 .

( )G. Leitinger et al.rBrain Research 823 1999 11–2322
Acknowledgements
ŽWe thank Prof. Dr. G. Dohr Head of the Department of.Histology and Embryology for the interest in this work,
R. Schmied, A. Blaschitz, and E. Schoninkle for their¨Žexpert technical assistance, and W. Salvenmoser Depart-
.ment of Zoology, University of Innsbruck for method-ological introduction. We are also grateful to Dr. P.J.
Ž .Simmons Newcastle University for proof reading a draftŽof the manuscript and Dr. F.C. Rind Newcastle Univer-
.sity for discussions. This investigation was supported byGrant No. P9510-Bio to K. Kral from the Austrian ScienceFoundation.
References
w x1 L.F. Agnati, K. Fuxe, M. Zoli, I. Zini, G. Toffano, F. Ferraguti, Acorrelation analysis of the regional distribution of central enkephalinand b-endorphin immunoreactive terminals and of opiate receptorsin adult and old male rats. Evidence for the existence of two maintypes of communication in the central nervous system: the volumetransmission and the wiring transmission, Acta Physiol. Scand. 128Ž .1986 201–207.
w x2 R.A. Baines, J.P. Bacon, Pharmacological analysis of the cholinergicinput to the locust VLPI neuron from an extraocular photoreceptor
Ž .system, J. Neurophysiol. 72 1994 2864–2874.w x3 C.A. Bishop, M. O’Shea, Serotonin immunoreactive neurons in the
Ž .central nervous system of an insect Periplaneta americana , J.Ž .Neurobiol. 14 1983 251–269.
w x4 C.B. Boschek, On the fine structure of the peripheral retina andlamina ganglionaris of the fly, Musca domestica, Z. Zellforsch.
Ž .Mikrosk. Anat. 118 1971 369–409.w x5 J. Cuadras, Non-synaptic release from dense-cored vesicles occurs at
Ž .all terminal types in crayfish neuropile, Brain Res. 477 1989332–335.
w x6 M.F. Cuttle, W. Hevers, S.B. Laughlin, R.C. Hardie, Diurnal modu-lation of photoreceptor potassium conductance in the locust, J.
Ž .Comp. Physiol. A 176 1995 307–316.w x7 K. Elekes, R. Hustert, M. Geffard, Serotonin-immunoreactive and
dopamine-immunoreactive neurones in the terminal ganglion of thecricket, Acheta domestica: light and electron microscopic immuno-
Ž .cytochemistry, Cell Tissue Res. 250 1987 167–180.w x8 J. Erber, P. Kloppenburg, A. Scheidler, Neuromodulation by sero-
tonin and octopamine in the honeybee: behaviour, neuroanatomy andŽ .electrophysiology, Experientia 49 1993 1073–1083.
w x9 P.D. Evans, Biogenic amines in the insect nervous system, Adv.Ž .Insect Physiol. 15 1980 317–373.
w x10 F. Felisberti, D.F. Ventura, Cerebral extraocular photoreceptors inŽ .ants, Tissue Cell 28 1995 25–30.
w x11 F. Felisberti, D.F. Ventura, H. Hertel, Cerebral extraocular photore-Ž .ceptors in beetles, Comp. Biochem. Physiol. A 118 1997 1353–
1357.w x12 G. Fleissner, G. Fleissner, B. Frisch, A new type of putative
nonvisual photoreceptors in the optic lobe of beetles, Cell TissueŽ .Res. 273 1993 435–445.
w x13 M. Germ, Dopamine, N-acetyldopamine and serotonin concentra-tions in the visual system of praying mantis during postembryonic
Ž .development, Comp. Biochem. Physiol. A 116 1997 379–386.w x14 M. Germ, K. Kral, Influence of visual deprivation on levels of
dopamine and serotonin in the visual system of house crickets,Ž .Acheta domesticus, J. Insect Physiol. 41 1995 57–63.
w x15 D.W. Golding, A pattern confirmed and refined-synaptic, nonsynap-Ž .tic and parasynaptic exocytosis, Bio. Essays 16 1994 503–508.
w x16 W. Hevers, R.C. Hardie, Serotonin modulates the voltage depen-dence of delayed rectifier and shaker potassium channels in
Ž .Drosophila photoreceptors, Neuron 14 1995 845–856.w x17 A. Hofbauer, E. Buchner, Does Drosophila have 7 eyes, Naturwis-
Ž .senschaften 76 1989 335–336.w x18 T. Hokfelt, Neuropeptides in perspective—the last ten years, Neuron¨
Ž .7 1991 867–879.w x19 U. Homberg, Distribution of neurotransmitters in the insect brain,
Ž .Prog. Zool. 40 1994 1–88.w x20 U. Homberg, J.G. Hildebrand, Serotonin immunoreactivity in the
optic lobes of the sphinx moth Manduca sexta and colocalizationwith FMRFamide and SCPB immunoreactivity, J. Comp. Neurol.
Ž .288 1989 243–253.w x21 U. Homberg, J.G. Hildebrand, Serotonin-immunoreactive neurons in
the median protocerebrum and suboesophageal ganglion of theŽ .sphinx moth Manduca sexta, Cell Tissue Res. 258 1989 1–24.
w x22 G.A. Horridge, P. Duelli, Anatomy of the regional differences in theŽ .eye of the mantis Ciulfina, J. Exp. Biol. 80 1979 165–190.
w x23 G.A. Horridge, C. Duniec, L. Marcelja, A 24-hour cycle in singleŽ .locust and mantis photoreceptors, J. Exp. Biol. 91 1981 307–322.
w x24 T. Ichikawa, Light suppresses the activity of serotonin-immunoreactive neurons in the optic lobe of the swallowtail butter-
Ž .fly, Neurosci. Lett. 172 1994 115–118.w x25 W. Kirmse, Visual position information controlling smooth tracking
Ž .in the praying mantis, J. Exp. Biol. 119 1985 365–367.w x26 A. Kock, A.K. Jakobs, K. Kral, Visual prey discrimination in¨
monocular and binocular praying mantis Tenodera sinensis duringŽ .postembryonic development, J. Insect Physiol. 39 1993 485–491.
w x27 K. Kral, I.A. Meinertzhagen, Anatomical plasticity of synapses inŽ .the lamina of the optic lobe of the fly, Phil. T. Roy. Soc. Lond. B
Ž .323 1989 155–183.w x28 G. Laube, J. Roeper, J.C. Pitt, S. Sewing, U. Kistner, C.C. Garner,
O. Pongs, R.W. Veh, Ultrastructural localization of Shaker-relatedpotassium channel subunits and synapse-associated protein 90 toseptate-like junctions in rat cerebellar Pinceaux, Mol. Brain Res. 42Ž .1996 51–61.
w x29 H. Maldodano, J.C. Barros Pita, A fovea in the praying mantis eye:´Ž .I. Estimation of the catching distance, Z. Vergl. Physiol. 65 1970
58–78.w x30 U. Mathis, S. Eschbach, S. Rossel, Functional binocular vision is not
dependant on visual experience in the praying mantis, Vis. Neurosci.Ž .9 1992 199–203.
w x31 I.A. Meinertzhagen, The organization of perpendicular fibre path-Ž .ways in the insect optic lobe, Phil. Trans. Roy. Soc. Lond. B 274
Ž .1976 555–596.w x32 D.R. Nassel, Serotonin and serotonin-immunoreactive neurons in the¨
Ž .nervous system of insects, Prog. Neurobiol. 30 1988 1–85.w x33 D.R. Nassel, Neurotransmitters and neuromodulators in the insect¨
Ž .visual system, Prog. Neurobiol. 37 1991 179–254.w x34 D.R. Nassel, E.P. Meyer, N. Klemm, Mapping and ultrastructure of¨
serotonin-immunoreactive neurons in the optic lobe of three insectŽ .species, J. Comp. Neurol. 323 1985 190–204.
w x35 D.R. Nassel, L. Ohlsson, P. Sivasubramanian, Postembryonic differ-¨entiation of serotonin-immunoreactive neurons in fleshfly optic lobesdeveloping in situ or cultured in vivo without eye discs, J. Comp.
Ž .Neurol. 255 1987 327–340.w x36 D.R. Nassel, S. Shiga, E.M. Wikstrand, K.R. Rao, Pigment-disper-¨
sing hormone-immunoreactive neurons and their relation to seroton-ergic neurons in the blowfly and cockroach visual system, Cell
Ž .Tissue Res. 266 1991 511–523.w x37 N.N. Osborne, M. Hamon, Neuronal Serotonin, Wiley, New York,
1988.w x38 S.R. Ott, M. Burrows, Nitric oxide synthase in the thoracic ganglia
of the locust: distribution in the neuropiles and morphology ofŽ .neurones, J. Comp. Neurol. 395 1998 217–230.

( )G. Leitinger et al.rBrain Research 823 1999 11–23 23
w x39 B. Petri, M. Stengl, S. Wurden, U. Homberg, Immunocytochemical¨characterization of the accessory medulla in the cockroach Leu-
Ž .cophaea maderae, Cell Tissue Res. 282 1995 3–19.w x40 M. Poteser, K. Kral, Visual distance discrimination between station-
ary targets in praying mantis: an index of the use of motion parallax,Ž .J. Exp. Biol. 198 1995 2127–2137.
w x41 F.R. Prete, T. McLean, Responses to moving small field stimuli byŽ .the praying mantis, Brain Behav. Evol. 47 1996 42–54.
w x42 E. Pyza, I.A. Meinertzhagen, Neurotransmitters regulate rhythmicsize changes amongst cells in the fly’s optic lobe, J. Comp. Physiol.
Ž .A. 178 1996 33–45.w x43 T. Reischig, M. Stengl, Morphology and pigment-dispersing hor-
mone immunocytochemistry of the accessory medulla, the presump-tive circadian pacemaker of the cockroach Leucophaea maderae: a
Ž .light and electron microscopic study, Cell Tissue Res. 285 1996305–319.
w x44 S. Rossel, Regional differences in photoreceptor performance in theŽ .eye of the praying mantis, J. Comp. Physiol. 131 1979 95–112.
w x Ž .45 S. Rossel, Binocular stereopsis in an insect, Nature 302 1983821–822.
w x46 S. Rossel, Binocular spatial localization in the praying mantis, J.Ž .Exp. Biol. 120 1986 265–281.
w x47 S. Rossel, Spatial vision in the praying mantis—Is distance impli-Ž .cated in size detection, J. Comp. Physiol. A 169 1991 101–108.
w x48 S. Rossel, U. Mathis, T. Collett, Vertical disparity and binocularŽ .vision in the praying mantis, Vis. Neurosci. 8 1992 165–170.
w x49 I. Salecker, P. Distler, Serotonin-immunoreactive neurons in theantennal lobes of the American cockroach Periplaneta americana:light- and electron microscopic observations, Histochemistry 94Ž .1990 463–473.
w x50 R.E. Sandeman, A.H.D. Watson, D.C. Sandeman, Ultrastructure ofthe synaptic terminals of the dorsal giant serotonin-IR neuron anddeutocerebral commissure interneurons in the accessory and olfac-
Ž .tory lobes of the crayfish, J. Comp. Neurol. 361 1995 617–632.w x51 F.W. Schurmann, R. Sandeman, D. Sandeman, Dense core vesicles¨
and non-synaptic exocytosis in the central body of the crayfish brain,Ž .Cell Tissue Res. 265 1991 493–501.
w x52 D.G. Stavenga, Pseudopupils of compound eyes, in: H. AutrumŽ .Ed. , Handbook of Sensory Physiology, Vol. VIIr6A, Springer,Berlin, 1979, pp. 357–439.
w x53 L.A. Sternberger, Immunocytochemistry, Wiley, New York, 1979.w x54 Y.D. Stierhof, B.M. Humbel, H. Schwarz, Suitability of different
silver enhancement methods applied to 1 nm colloidal gold particles:an immunoelectron microscopic study, J. Electron Microsc. Techn.
Ž .17 1991 336–343.w x55 N.J. Strausfeld, Atlas of an Insect Brain, Springer, Berlin, 1976.w x56 X.J. Sun, L.P. Tolbert, J.G. Hildebrand, Ramification pattern and
ultrastructural characteristics of the serotonin-immunoreactive neu-ron in the antennal lobe of the moth Manduca sexta: a laserscanning confocal and electron microscopic study, J. Comp. Neurol.
Ž .338 1993 5–16.w x57 K.S.J. Thompson, M.P. Zeidler, J.P. Bacon, Comparative anatomy
of serotonin-like immunoreactive neurons in isopods: putative homo-Ž .logues in several species, J. Comp. Neurol. 347 1994 553–569.
w x58 K. Tomioka, Y. Chiba, Characterization of an optic lobe circadianpacemaker by in situ and in vitro recording of neural activity in the
Ž .cricket, Gryllus bimaculatus, J. Comp. Physiol. A 171 1992 1–7.w x59 K. Tomioka, M. Ikeda, T. Nagao, S. Tamotsu, Involvement of
serotonin in the circadian rhythm of an insect visual system, Natur-Ž .wissenschaften 80 1993 137–139.
w x60 F. Walcher, K. Kral, Visual deprivation and distance estimation inŽ .the praying mantis larva, Physiol. Entomol. 19 1994 230–240.
w x61 S. Wurden, U. Homberg, Immunocytochemical mapping of sero-¨tonin and neuropeptides in the accessory medulla of the locust,
Ž .Schistocerca gregaria, J. Comp. Neurol. 362 1995 305–319.w x62 H. Zimmermann, Synaptic Transmission. Cellular and Molecular
Basis, Thieme, Stuttgart, New YorkrOxford University Press, NewYork, 1993.
w x63 M. Zoli, L.F. Agnati, Wiring and volume transmission in the centralnervous system: the concept of closed and open synapses, Prog.
Ž .Neurobiol. 49 1996 363–380.w x64 G.K.H. Zupanc, Peptidergic transmission—from morphological cor-
Ž .relates to functional implications, Micron 27 1996 35–91.





























