Embed Size (px)
Citation preview

Shear Stress–Mediated Chromatin Remodeling ProvidesMolecular Basis for Flow-Dependent Regulation
of Gene ExpressionBarbara Illi, Simona Nanni, Alessandro Scopece, Antonella Farsetti, Paolo Biglioli,
Maurizio C. Capogrossi, Carlo Gaetano
Abstract—Shear stress (SS), the tangential component of hemodynamic forces, modulates the expression of several genesin endothelial cells. However, no information is available about its effect on chromatin structure, which plays a key rolein gene transcription. In this study, a link between SS and chromatin remodeling was established in human umbilicalvein endothelial cells (HUVECs). HUVECs were exposed to SS of 10 dyne/cm2 per second, in the presence or absenceof the histone deacetylase inhibitor trichostatin A, and assayed for histone H3 and histone H4 modifications. SS inducedhistone H3 serine phosphorylation at position 10 (S10) and lysine acetylation at position 14 (K14) but requiredtrichostatin A to induce H3 phosphoacetylation and H4 acetylation. The phosphatidylinositol 3-kinase inhibitorwortmannin and the mitogen-activated protein kinase inhibitor PD98059 decreased SS-dependent histone H3phosphorylation, without affecting its acetylation; the p38 inhibitor SB203580 reduced both H3 phosphorylation andacetylation, whereas the protein kinase A inhibitor PKI-tide reduced histone H3 acetylation. Remarkably, the abrogationof histone acetylation inhibited SS-dependent c-fos expression. SS also activated ribosomal S6 kinase-2 and mitogen-and stress-activated kinase-1 protein kinases and promoted the formation of a cAMP-responsive element–bindingprotein (CREB)/CREB-binding protein complex, providing the molecular basis for the increase in histone acetyltrans-ferase activity observed in HUVECs exposed to SS. Finally, the effect of SS on chromatin remodeling was examined.In HUVECs exposed to SS, chromatin within c-fos and c-jun promoters was specifically immunoprecipitated by anantibody against acetylated histone H3 on K14. These results indicate that SS induces posttransduction modificationsof histones; this is an early step toward the flow-dependent regulation of gene expression. (Circ Res. 2003;93:155-161.)
Key Words: gene expression � histone acetylation � histone phosphorylation � chromatin � endothelial cells
Shear stress (SS) modulates endothelial cell (EC) functionthrough mechanosensors, which activate intracellular sig-
naling pathways, leading to the transcription of specificgenes. Vascular endothelial growth factor (VEGF)receptor-2, focal adhesion kinase, and the integrins �v�1, �3,and �5 have been identified as important components of theSS mechanotransduction machinery.1–3 In response to SS, theintracellular protein kinases src and focal adhesion kinasebecome activated and interact with docking proteins grb-2,shc, and paxillin,4 stimulating downstream mitogen-activatedprotein kinases (MAPKs).5 MAPKs modulate the function oftranscription factors (TFs)6 and lead to the transcriptionalinduction of immediate-early genes.1,7 Activated TFs interactwith specific SS response elements, which are present in thepromoter region of many SS-responsive genes modulatingtheir expression.8 In fact, SS activates nuclear factor-�B,9,10
the cAMP-responsive element–binding protein (CREB)11 and
activator protein-1,9,12 regulating the expression of manyendothelial genes. Moreover, Smad proteins have been re-cently identified as a novel class of SS-dependent TFs, buttheir role in flow-regulated gene expression remains un-clear.13,14 Transcriptional regulation of gene expression isprimarily modulated by the accessibility of chromatin to TFs,and recent observations indicate that modifications in thechromatin structure are required for the expression of severalgenes.15 Chromatin structure depends on the activity ofhistone-modifying enzymes, including histone acetyltrans-ferases (HATs) and histone deacetylases (HDACs).15,16
HATs catalyze the addition of acetyl groups to specificlysines (K) present in the histone tails, disrupting nucleosomeinteractions and allowing the transcriptional machinery toaccess DNA. Yeast GCN5, mammalian CREB-binding pro-tein (CBP)/p300 and P/CAF, and some important componentof the basal transcription machinery, as well as the TBP-
Original received January 9, 2003; revision received May 29, 2003; accepted May 30, 2003.From Laboratorio di Biologia Vascolare e Terapia Genica (B.I.), Centro Cardiologico Fondazione “I. Monzino,” IRCCS, Milan; Centro Cardiologico
Fondazione “I. Monzino” (B.I., P.B.), IRCCS, Milan; Laboratorio di Oncogenesi Molecolare (S.N, A.F.), Istituto Regina Elena, Rome; Laboratorio diPatologia Vascolare (A.S., M.C.C., C.G.), Istituto Dermopatico dell’Immacolata, IRCCS, Rome; and Istituto di Neurobiologia e Medicina Molecolare(A.F.), CNR, Rome, Italy.
Correspondence to Carlo Gaetano, MD, Laboratorio di Patologia Vascolare, IDI-IRCCS, Via dei Monti di Creta 104, 00167 Rome, Italy. [email protected]
© 2003 American Heart Association, Inc.
Circulation Research is available at http://www.circresaha.org DOI: 10.1161/01.RES.0000080933.82105.29
155 by guest on November 16, 2015http://circres.ahajournals.org/Downloaded from by guest on November 16, 2015http://circres.ahajournals.org/Downloaded from by guest on November 16, 2015http://circres.ahajournals.org/Downloaded from by guest on November 16, 2015http://circres.ahajournals.org/Downloaded from by guest on November 16, 2015http://circres.ahajournals.org/Downloaded from by guest on November 16, 2015http://circres.ahajournals.org/Downloaded from by guest on November 16, 2015http://circres.ahajournals.org/Downloaded from by guest on November 16, 2015http://circres.ahajournals.org/Downloaded from

associated factor II 250, belong to this family and exhibitintrinsic HAT activity.15,16 Conversely, gene silencing isprovided by histone deacetylation and methylation. HDACsand histone methyltransferases, like the yeast suppressor ofvariegation [Su(var)] group,15 catalyze, respectively, the sub-straction of acetyl groups and the addition of methyl groupsto histones, switching DNA from a euchromatic to anetherochromatic state in which TFs do not recognize theircognate response elements on target promoters.16
Recently, histone phosphorylation has been identified as anadditional chromatin modification involved in the transcrip-tional control of gene expression. This activity is, at least inpart, dependent on the function of the ribosomal S6 kinase-2(RSK-2), a member of the pp90rsk family of kinases, which isknown to regulate gene expression in response to mito-gens,17,18 and on mitogen- and stress-activated kinase-1(MSK-1), making histones susceptible to hyperacetylation byHATs.19 Although a number of physiological and pharmaco-logical stimuli17,19,20 are known to regulate histone modifica-tions, no prior studies have examined the effect of mechanicalforces on chromatin remodeling. The present study showsthat SS induces histone H3 acetylation and phosphorylationand cooperates with the HDAC inhibitor trichostatin A (TSA)to enhance histone H3 phosphoacetylation and histone H4acetylation. These changes at the level of histones provide themolecular basis for those chromatin modifications requiredfor DNA unwinding and may play a fundamental role in theSS-dependent regulation of gene expression.
Materials and MethodsCell Culture, Treatments, and SS ExposureHuman umbilical vein ECs (HUVECs) were grown in endothelialgrowth medium-2 (EGM-2, Clonetics). TSA (32 nmol/L) and 10ng/mL of recombinant VEGF and fibroblast growth factor (FGF)-2or an equal amount of control solvent were added to completemedium, and the cells were immediately exposed to SS in acone-plate apparatus2 or kept in a static condition (ST). Wortmannin(50 nmol/L), PD98059 (50 �mol/L), SB203580 (5 �mol/L), andPKI-tide (5 �mol/L, a protein kinase A inhibitor) or equal amount ofcontrol solvent were added 30 minutes before SS exposure toendothelial basal medium-2 (EBM-2, Clonetics) supplemented with2% FBS after an overnight starvation of cells in EBM-2 supple-mented with 0.5% FBS.
Western BlotsTotal protein extracts were obtained by lysing cells with 1�Laemmli buffer. Modified histones were detected with anti–acety-lated histone H3K14, anti–acetylated histone H4, anti–phosphohistoneH3S10, anti–phosphoS10-acetylK14 histone H3 (Upstate Biotechnology),anti– c-fos (Santa Cruz), anti–phospho-RSK-2(Ser380), and anti–phospho-MSK1(Ser376) (Cell Signaling) antibodies, according to themanufacturer’s instructions. Normalization of protein loading wasobtained using an anti–GRB-2 antibody (Santa Cruz), anti-histoneH1 antibody (Upstate Biotechnology), and anti–RSK-2 and anti-MSK-1 antibodies (Cell Signaling). For detailed protocol, see onlinedata supplement (available at http://www.circresaha.org).
Pull-Down AssayCells were exposed to SS for 1 hour or kept in ST. Nuclear extractswere obtained as previously described21 and incubated with 5�-biotinylated oligonucleotides containing a CREB site (see onlinesupplemental data for detailed protocol).
Oligonucleotide sequences were as follows: AGAGATTGCCT-GACGTCAGAGAGCTAG and CTAGCTCTCTGACGTCAG-
GCAATCTCT. Bound complexes were boiled in 1� Laemmlibuffer and detected with anti-CREB and anti-CBP antibodies (SantaCruz), according to manufacturer’s instructions.
In Vitro Acetylation AssayWhole-cell extracts were obtained as described.2 HAT activity wasdetected using the HAT-check activity assay kit (Pierce) according tothe manufacturer’s instructions. The amount of radiolabeled H4peptide was measured using a liquid scintillation counter (TRI-CARB, Packard). See online supplemental data for detailed protocol.
ChIp AssayHUVECs (3�107 cells) were starved overnight in EBM-2 supple-mented with 0.5% FBS. The day after, medium was replaced withEBM-2 supplemented with 2% FBS, and HUVECs were exposed toSS for 60 minutes or kept in ST. Chromatin immunoprecipitation(ChIp) was performed as described.22 A primer pair, GGGTTGACT-GGTAGCAGATAAGTGTTGAG and TCTGGGCAGTTA-GAGAGAAGGTGAAAAG, was used to amplify a 200-bp promoterfragment of c-jun. Another primer pair, GCAGCCGGGCGGCCG-CAGAA and GTCTCGACCCATCCTCGTGCC, was used to am-plify a 216-bp promoter fragment of c-fos.
Statistical AnalysisResults were analyzed by 1-way ANOVA. Post hoc tests accordingto the Student-Newman-Keuls method were used to assess statisti-cally significant differences among different groups. A value ofP�0.05 was considered statistically significant.
ResultsSS Induces Phosphorylation of Histone H3 andAcetylation of Histones H3 and H4The effect of SS on histone modifications was examined byWestern blot analyses using protein extracts obtained fromHUVECs exposed to a laminar SS of 10 dyne/cm2 per secondfor 30 to 240 minutes or kept in ST, either in the presence orabsence of TSA. The pattern of histone H3 phosphorylationwas first evaluated because histone H3 phosphorylation andphosphoacetylation account for chromatin modifications inresponse to extracellular stimuli.15 In the presence of SS,histone H3 became phosphorylated within 30 minutes (Figure1A). Surprisingly, in cells treated with TSA, the level ofphosphorylation was weaker (Figure 1A, right). It has beenreported that in case of multiple modifications, the epitoperecognized by the anti–phospho H3 antibody at position 10(S10) may be masked by the acetyl group at position 14(K14), making antibody binding to the phosphorylatedepitope less efficient.19 To verify whether in our experimentalconditions histone H3 was simultaneously phosphorylated onS10 and acetylated on K14, we performed Western blotanalyses using an anti–phospho-acetyl histone H3 antibody.We found that this histone became both phosphorylated andacetylated in HUVECs treated with TSA and exposed to SS(Figure 1B). Phosphorylation and acetylation of histones areregulated by coordinated but independent mechanisms23;therefore, the level of histone H3 acetylation alone was alsoanalyzed. SS stimulated histone H3 acetylation on K14 witha peak between 60 and 120 minutes, and this phenomenonwas further increased in the presence of TSA (Figure 1C). Inaddition, the acetylation level of the histone H4, which,together with H3, constitutes the inner scaffold of nucleo-somes, was evaluated.16 Notably, in ECs treated with TSA,
156 Circulation Research July 25, 2003
by guest on November 16, 2015http://circres.ahajournals.org/Downloaded from
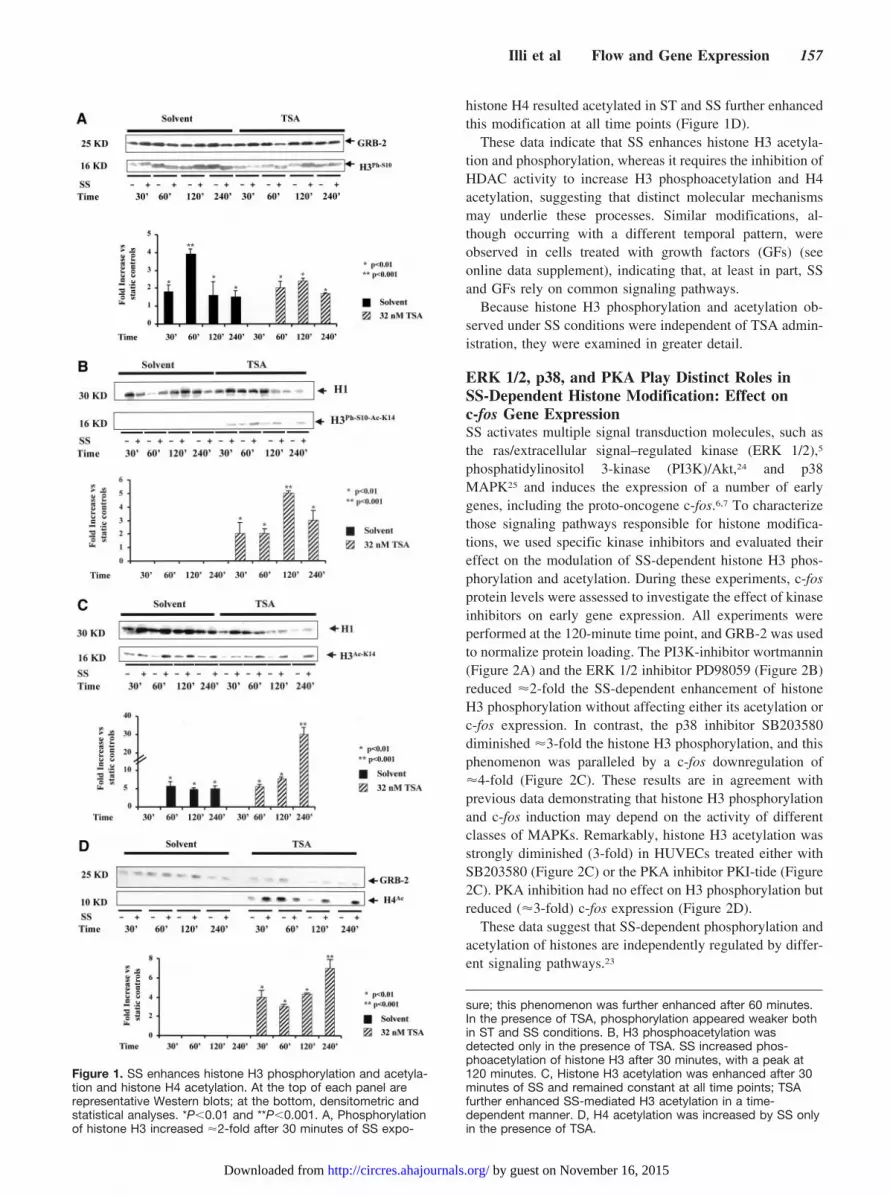
histone H4 resulted acetylated in ST and SS further enhancedthis modification at all time points (Figure 1D).
These data indicate that SS enhances histone H3 acetyla-tion and phosphorylation, whereas it requires the inhibition ofHDAC activity to increase H3 phosphoacetylation and H4acetylation, suggesting that distinct molecular mechanismsmay underlie these processes. Similar modifications, al-though occurring with a different temporal pattern, wereobserved in cells treated with growth factors (GFs) (seeonline data supplement), indicating that, at least in part, SSand GFs rely on common signaling pathways.
Because histone H3 phosphorylation and acetylation ob-served under SS conditions were independent of TSA admin-istration, they were examined in greater detail.
ERK 1/2, p38, and PKA Play Distinct Roles inSS-Dependent Histone Modification: Effect onc-fos Gene ExpressionSS activates multiple signal transduction molecules, such asthe ras/extracellular signal–regulated kinase (ERK 1/2),5
phosphatidylinositol 3-kinase (PI3K)/Akt,24 and p38MAPK25 and induces the expression of a number of earlygenes, including the proto-oncogene c-fos.6,7 To characterizethose signaling pathways responsible for histone modifica-tions, we used specific kinase inhibitors and evaluated theireffect on the modulation of SS-dependent histone H3 phos-phorylation and acetylation. During these experiments, c-fosprotein levels were assessed to investigate the effect of kinaseinhibitors on early gene expression. All experiments wereperformed at the 120-minute time point, and GRB-2 was usedto normalize protein loading. The PI3K-inhibitor wortmannin(Figure 2A) and the ERK 1/2 inhibitor PD98059 (Figure 2B)reduced �2-fold the SS-dependent enhancement of histoneH3 phosphorylation without affecting either its acetylation orc-fos expression. In contrast, the p38 inhibitor SB203580diminished �3-fold the histone H3 phosphorylation, and thisphenomenon was paralleled by a c-fos downregulation of�4-fold (Figure 2C). These results are in agreement withprevious data demonstrating that histone H3 phosphorylationand c-fos induction may depend on the activity of differentclasses of MAPKs. Remarkably, histone H3 acetylation wasstrongly diminished (3-fold) in HUVECs treated either withSB203580 (Figure 2C) or the PKA inhibitor PKI-tide (Figure2C). PKA inhibition had no effect on H3 phosphorylation butreduced (�3-fold) c-fos expression (Figure 2D).
These data suggest that SS-dependent phosphorylation andacetylation of histones are independently regulated by differ-ent signaling pathways.23
Figure 1. SS enhances histone H3 phosphorylation and acetyla-tion and histone H4 acetylation. At the top of each panel arerepresentative Western blots; at the bottom, densitometric andstatistical analyses. *P�0.01 and **P�0.001. A, Phosphorylationof histone H3 increased �2-fold after 30 minutes of SS expo-
sure; this phenomenon was further enhanced after 60 minutes.In the presence of TSA, phosphorylation appeared weaker bothin ST and SS conditions. B, H3 phosphoacetylation wasdetected only in the presence of TSA. SS increased phos-phoacetylation of histone H3 after 30 minutes, with a peak at120 minutes. C, Histone H3 acetylation was enhanced after 30minutes of SS and remained constant at all time points; TSAfurther enhanced SS-mediated H3 acetylation in a time-dependent manner. D, H4 acetylation was increased by SS onlyin the presence of TSA.
Illi et al Flow and Gene Expression 157
by guest on November 16, 2015http://circres.ahajournals.org/Downloaded from

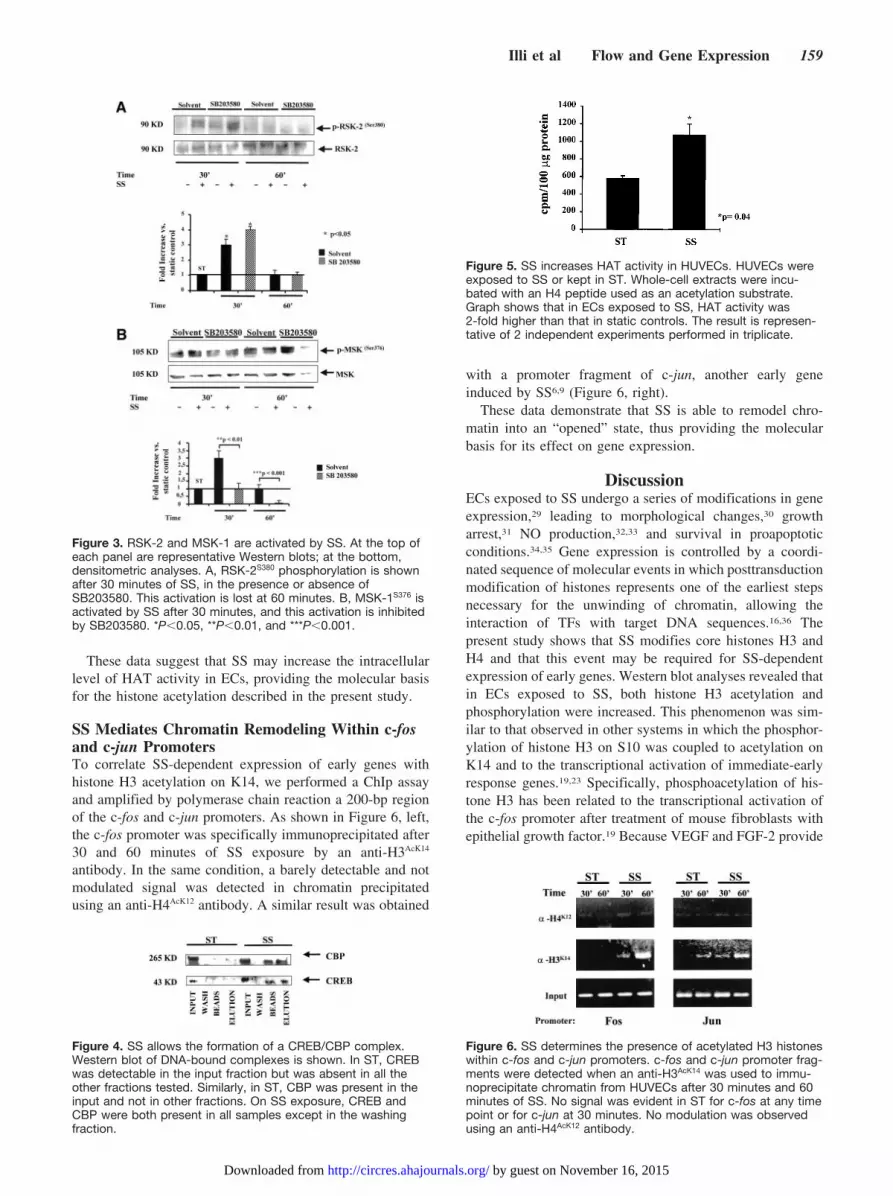
RSK-2 and MSK-1 Are Activated by SSPhosphorylation of histone H3 on S10 depends on RSK-2and/or MSK-1 kinase activity.17,26 Moreover, MSK-1 maymediate CREB activation,27 which, recruiting CBP, couldenhance histone acetylation. Phosphorylation of RSK-2 de-pends on ERK 1/2 signaling, whereas the phosphorylation ofMSK-1 relies either on ERK 1/2 or p38 kinase activity.26
Because in our experiments the p38 inhibitor SB203580interfered either with H3 phosphorylation or acetylation,experiments were performed in the presence of SB203580 toevaluate RSK-2 and MSK-1 activation in HUVECs exposedto SS or kept in ST. Western blot analysis indicates that after30 minutes of SS, RSK-2 and MSK-1 became phosphorylatedat serine residues 380 and 376, respectively. This event wastransient, and the signal rapidly decreased between 30 and 60minutes. As expected, inhibition of p38 activity by SB203580treatment did not alter RSK-2 activation in the presence of SS(Figure 3A), whereas MSK-1 phosphorylation at serine resi-due 376 was completely abolished. Phosphorylation ofMSK-1 and RSK-2 in the presence of SS was apparentlylimited to serines 376 and 380, respectively, inasmuch as nodetectable signals were obtained from other serine andthreonine residues known as phosphorylation targets in thesemolecules (not shown). These results suggest that althoughthe ERK 1/2 pathway may regulate H3 phosphorylation viaRSK-2 activation, p38 and MSK-1 could be mainly involvedin H3 phosphorylation and acetylation.
SS Stimulates Formation of CREB/CBPTranscriptional Complex and Enhances HATActivity in ECsThe expression of c-fos depends on CREB transcriptionalactivity,28 which is activated by PKA. Because we observedthat inhibition of PKA activity strongly interfered with c-fosexpression (Figure 2D), we reasoned that CREB transcrip-tional complexes could be activated by SS. To demonstratethis hypothesis, we performed a pull-down assay on nuclearextracts derived from HUVECs exposed to SS or kept in ST.Figure 4 shows that in the presence of SS, CREB recruits theCREB-binding protein CBP and this complex binds to asynthetic cAMP response element (CRE). This result indi-cates that SS induces the formation of transcriptional com-plexes potentially carrying HAT activity on specific DNAtarget sequences. To verify the hypothesis that an increasedHAT activity could account for histone H3 acetylation inresponse to SS, an acetylation assay was performed in vitrousing whole-cell extracts obtained from ECs exposed to SS.Figure 5 shows that the level of acetylation of a biotinylatedhistone H4 peptide used as a substrate was �2-fold higher inthe presence of protein extracts obtained from ECs exposed toSS than those kept in ST.
both phosphorylation and acetylation of histone H3 and c-fosexpression in response to SS. D, PKA inhibitor PKI-tide (5�mol/L) inhibits SS-mediated H3 acetylation and c-fos expres-sion, without affecting H3 phosphorylation. E, Graph showsresult of densitometric and statistical analyses of 3 independentexperiments. *P�0.01 and **P�0.001.
Figure 2. SS-dependent histone H3 phosphorylation and acety-lation and c-fos expression are differentially regulated. A and B,PI3K inhibitor wortmannin (50 nmol/L, A) and ERK inhibitorPD98059 (50 �mol/L, B) abrogate SS effect on H3 phosphoryla-tion; in contrast, no effects are observed on H3 acetylation andc-fos induction. C, p38 inhibitor SB203580 (5 �mol/L) reduces
158 Circulation Research July 25, 2003
by guest on November 16, 2015http://circres.ahajournals.org/Downloaded from
These data suggest that SS may increase the intracellularlevel of HAT activity in ECs, providing the molecular basisfor the histone acetylation described in the present study.
SS Mediates Chromatin Remodeling Within c-fosand c-jun PromotersTo correlate SS-dependent expression of early genes withhistone H3 acetylation on K14, we performed a ChIp assayand amplified by polymerase chain reaction a 200-bp regionof the c-fos and c-jun promoters. As shown in Figure 6, left,the c-fos promoter was specifically immunoprecipitated after30 and 60 minutes of SS exposure by an anti-H3AcK14
antibody. In the same condition, a barely detectable and notmodulated signal was detected in chromatin precipitatedusing an anti-H4AcK12 antibody. A similar result was obtained
with a promoter fragment of c-jun, another early geneinduced by SS6,9 (Figure 6, right).
These data demonstrate that SS is able to remodel chro-matin into an “opened” state, thus providing the molecularbasis for its effect on gene expression.
DiscussionECs exposed to SS undergo a series of modifications in geneexpression,29 leading to morphological changes,30 growtharrest,31 NO production,32,33 and survival in proapoptoticconditions.34,35 Gene expression is controlled by a coordi-nated sequence of molecular events in which posttransductionmodification of histones represents one of the earliest stepsnecessary for the unwinding of chromatin, allowing theinteraction of TFs with target DNA sequences.16,36 Thepresent study shows that SS modifies core histones H3 andH4 and that this event may be required for SS-dependentexpression of early genes. Western blot analyses revealed thatin ECs exposed to SS, both histone H3 acetylation andphosphorylation were increased. This phenomenon was sim-ilar to that observed in other systems in which the phosphor-ylation of histone H3 on S10 was coupled to acetylation onK14 and to the transcriptional activation of immediate-earlyresponse genes.19,23 Specifically, phosphoacetylation of his-tone H3 has been related to the transcriptional activation ofthe c-fos promoter after treatment of mouse fibroblasts withepithelial growth factor.19 Because VEGF and FGF-2 provide
Figure 3. RSK-2 and MSK-1 are activated by SS. At the top ofeach panel are representative Western blots; at the bottom,densitometric analyses. A, RSK-2S380 phosphorylation is shownafter 30 minutes of SS, in the presence or absence ofSB203580. This activation is lost at 60 minutes. B, MSK-1S376 isactivated by SS after 30 minutes, and this activation is inhibitedby SB203580. *P�0.05, **P�0.01, and ***P�0.001.
Figure 4. SS allows the formation of a CREB/CBP complex.Western blot of DNA-bound complexes is shown. In ST, CREBwas detectable in the input fraction but was absent in all theother fractions tested. Similarly, in ST, CBP was present in theinput and not in other fractions. On SS exposure, CREB andCBP were both present in all samples except in the washingfraction.
Figure 5. SS increases HAT activity in HUVECs. HUVECs wereexposed to SS or kept in ST. Whole-cell extracts were incu-bated with an H4 peptide used as an acetylation substrate.Graph shows that in ECs exposed to SS, HAT activity was2-fold higher than that in static controls. The result is represen-tative of 2 independent experiments performed in triplicate.
Figure 6. SS determines the presence of acetylated H3 histoneswithin c-fos and c-jun promoters. c-fos and c-jun promoter frag-ments were detected when an anti-H3AcK14 was used to immu-noprecipitate chromatin from HUVECs after 30 minutes and 60minutes of SS. No signal was evident in ST for c-fos at any timepoint or for c-jun at 30 minutes. No modulation was observedusing an anti-H4AcK12 antibody.
Illi et al Flow and Gene Expression 159
by guest on November 16, 2015http://circres.ahajournals.org/Downloaded from

potent proliferation and differentiation stimuli to ECs, weinvestigated whether these GFs could have effects similar tothose of SS on histone H3. We found that VEGF enhancedboth phosphorylation and acetylation of histone H3 but witha kinetic different from that observed under SS conditions.FGF-2 showed a transient induction of H3 phosphorylationwithout affecting its acetylation (see online data supplement).Conversely, SS activation of ECs was able to generate a moreenhanced and prolonged effect on histone H3 acetylation andphosphorylation than that induced by GFs alone. Therefore,SS and GFs, although probably sharing common signalingpathways, may have distinct effects on histones. However,further experiments are required to elucidate this point.
SS-dependent histone H3 phosphorylation and acetylationappeared linked to the activation of distinct signaling path-ways, controlled respectively by PI3K and by ERK and p38or p38 and PKA. Blocking the MAPK/ERK pathway and H3phosphorylation did not alter the acetylation of K14 onhistone H3, indicating that these events may be independentlyregulated.23 In fact, ERKs and p38 may differentially regulatedistinct downstream kinases involved in DNA structuremodification. The MAPK/ERK pathway activates RSK-2,and this kinase, when released from its interaction with CBP,can phosphorylate histone H3 on S10. This dissociative eventmay lead, in turn, to the acetylation of K14 on the same corehistone.18 However, phosphorylation of histone H3 can alsobe regulated by MSK-1, via p38 MAPK.26,27 In this regard,inhibiting p38 function resulted in a marked reduction of bothhistone H3 phosphorylation and acetylation during SS, indi-cating that the nuclear kinase MSK-1 is important for the SSeffect on the posttransductional modification of core histones.
Recent data suggest that PKA may be activated by SS.37
PKA is a well-known activator of CREB and CBP/p300,38,39
and in our experiments, SS apparently stimulates the forma-tion of CREB/CBP complexes, which may ultimately tetherHAT activity on target DNA regions. This observation issupported by the evidence that histone H3 acetylation on K14decreased on PKA inhibition. The formation of transcrip-tional complexes containing HAT molecules, which coordi-nate and integrate multiple signaling pathways with thetranscriptional machinery,16 and the increase in intracellular
acetylation activity occurring in the presence of SS mayprovide the molecular basis for the effect of flow on geneexpression. A growing body of evidence points out thatchanges in the folding of chromatin are required as apreliminary step toward the expression of target genes inpresence of specific stimuli.15 ChIp experiments clearly showthat in the presence of SS, acetylated histone H3 is presentwithin c-fos and c-jun promoter sequences, indicating that SSmay contribute to the remodeling of chromatin, which ac-quires an “opened” structure within the promoter region ofspecific genes. However, the status of chromatin acetylationresults from the balance of HAT and HDAC activity,15 and toelucidate this aspect in the context of SS requires further andthorough investigation. In the present report, it is, in fact,noteworthy that other SS-dependent modifications on histoneH3 and H4 were detected in presence of TSA. This evidencesuggests possible additional mechanisms of regulation thatmay not be directly controlled by SS. TSA, in fact, is known,per se, to alter biological responses in different biologicalsystems,40,41 eg, anticipating the timing of gene expressionduring embryo development.42 It is then possible that certainmodifications of chromatin occurring in the presence ofHDAC inhibitors, as described for the smooth muscle–specific SM22 gene locus in 10T1/2 cells,43 may not berelated to the effect of physiological stimuli.
In conclusion, the present study describes the effect oflaminar SS on histone modification and on chromatin struc-ture, providing the molecular basis for SS-mediated generegulation in ECs (see Figure 7 for summary).
AcknowledgmentsThis study has been partially supported by Ministero della Salute“Progetti di Ricerca Finalizzata” ICS 120.1, 1/RF00208, ICS 120.4/RA00-90, and “Ricerca Corrente” RC 2002-4.5. Simona Nanni issupported by the “Fondazione Italiana per la Ricerca sul Cancro”(FIRC).
References1. Chen KD, Li YS, Kim M, Li S, Yan S, Chien S, Shyy YJJ. Mechano-
transduction in response to shear stress. J Biol Chem. 1999;274:18393–18400.
2. Shay-Salit A, Shushy M, Wolfovitz E, Yahav H, Breviario F, Dejana E,Resnick N. VEGF receptor 2 and the adherens junction as a mechanical
Figure 7. Schematic representation of SS signalingpathways leading to histone modifications. Thisschematic representation summarizes the resultsof the present study. According to this model,PI3K, ERK 1/2, and RSK-2, as well as p38 andMSK-1, regulate phosphorylation (P) of H3 on S10.However, c-fos gene expression did not dependon phosphorylation of histone H3, inasmuch asinhibition of PI3K and ERK 1/2 pathways did notabrogate c-fos protein expression. Conversely, his-tone H3 acetylation (Ac) on K14 was regulated byPKA, p38, and MSK-1. PKA regulates the forma-tion of CREB/CBP complexes. Blocking p38 andPKA activity downregulates c-fos protein expres-sion. In conclusion, SS-dependent induction ofc-fos gene may depend on histone H3 acetylationand/or phosphoacetylation via p38/MSK-1 or PKA/CREB/CBP pathways.
160 Circulation Research July 25, 2003
by guest on November 16, 2015http://circres.ahajournals.org/Downloaded from

transducer in vascular endothelial cells. Proc Natl Acad Sci U S A. 2002;99:9462–9467.
3. Li S, Kim M, Hu YL, Jalali S, Schlaepfer DD, Hunter T, Chien S, ShyyYJJ. Fluid shear stress activation of focal adhesion kinase. J Biol Chem.1997;272:30455–30462.
4. Kano Y, Katoh K, Masuda M, Fujiwara K. Macromolecular compositionof stress fiber–plasma membrane attachment sites in endothelial cells insitu. Circ Res. 1996;79:1000–1006.
5. Jalali S, Li YS, Sotoudeh M, Yuan S, Li S, Chien S, Shyy YJJ. Shearstress activates p60src-Ras-MAPK signaling pathway in vascular endo-thelial cells. Arterioscler Thromb Vasc Biol. 1998;18:227–234.
6. Nagel T, Resnick N, Dewey CF Jr, Gimbrone MA Jr. Vascular endothe-lial cells respond to spatial gradients in fluid shear stress by enhancedactivation of transcription factors. Arterioscler Thromb Vasc Biol. 1999;19:1825–1834.
7. Hsieh HJ, Li NQ, Frangos JA. Pulsatile and steady flow induces c-fosexpression in human endothelial cells. J Cell Physiol. 1993;154:143–151.
8. Resnick N, Gimbrone MA Jr. Hemodynamic forces are complex regu-lators of endothelial gene expression. FASEB J. 1995;9:874–882.
9. Lan Q, Mercurius KO, Davies PF. Stimulation of transcription factorsNF-�B and AP-1 in endothelial cells subjected to shear stress. BiochemBiophys Res Commun. 1994;20:950–956.
10. Khachigian LM, Resnick N, Gimbrone MA Jr, Collins T. Nuclearfactor-�B interacts functionally with the platelet-derived growth factorB-chain shear stress response element in vascular endothelial cellsexposed to fluid shear stress. J Clin Invest. 1995;96:1169–1175.
11. De Nigris F, Lerman LO, Ignarro SW, Sica G, Lerman A, Palinski W,Ignarro LJ, Napoli C. Beneficial effects of antioxidants and L-arginine onoxidation-sensitive gene expression and endothelial NO synthase activityat sites of disturbed shear stress. Proc Natl Acad Sci U S A. 2003;100:1420–1425.
12. Korenaga R, Ando J, Kosaki K, Isshiki M, Takada Y, Kamiya A.Negative transcriptional regulation of the VCAM-1 gene by fluid shearstress in murine endothelial cells. Am J Physiol. 1997;273:C1506–C1515.
13. Topper JN, Cai J, Qiu Y, Anderson KR, Xu YY, Deeds JD, Feeley R,Gimeno CJ, Woolf EA, Tayber O, Mays GG, Sampson BA, Schoen FJ,Gimbrone MA Jr, Falb D. Vascular MADs: two novel MAD-relatedgenes selectively inducible by flow in human vascular endothelium. ProcNatl Acad Sci U S A. 1997;94:9314–9319.
14. Brown JD, DiChiara MR, Anderson KR, Gimbrone MA Jr, Topper JN.MEKK-1, a component of the stress (stress-activated protein kinase/c-junN-terminal kinase) pathway, can selectively activate Smad-2-mediatedtranscriptional activation in endothelial cells. J Biol Chem. 1999;274:8797–8805.
15. Jenuwein T, Allis CD. Translating the histone code. Science. 2001;293:1074–1079.
16. Chen H, Tini M, Evans RM. HATs on and beyond chromatin. Curr OpinCell Biol. 2001;13:218–224.
17. Sassone Corsi P, Mizzen CA, Cheung P, Crosio C, Monaco L, Jacquot S,Hanauer A, Allis CD. Requirement of Rsk-2 for epidermal growth factor-activated phosphorylation of histone H3. Science. 1999;285:886–891.
18. Merienne K, Pannetier S, Harel-Bellan A, Sassone-Corsi P. Mitogen-regulated Rsk2-CBP interaction controls their kinase and acetylaseactivity. Mol Cell Biol. 2001;21:7089–7096.
19. Clayton AL, Rose S, Barrat MJ, Mahadevan LC. Phosphoacetylation ofhistone H3 on c-fos and c-jun associated nucleosomes upon gene acti-vation. EMBO J. 2000;19:3714–3726.
20. Cheung P, Tanner KG, Cheung WL, Sassone-Corsi P, Denu JM, AllisCD. Synergistic coupling of histone H3 phosphorylation and acetylationin response to epidermal growth factor stimulation. Mol Cell. 2000;5:905–915.
21. Schreiber E, Matthias P, Muller MM, Shaffner W. Rapid detection ofoctamer binding proteins with “mini-extracts,” prepared from a smallnumber of cells. Nucleic Acids Res. 1998;17:6419.
22. Nanni S, Narducci M, Della Pietra L, Moretti F, Grasselli A, De Carli P,Sacchi A, Pontecorvi A, Farsetti A. Signaling through estrogen receptorsmodulates telomerase activity in human prostate cancer. J Clin Invest.2002;110:219–227.
23. Thomson S, Clayton AL, Mahadevan LC. Independent dynamic regu-lation of histone phosphorylation and acetylation during immediate-earlygene induction. Mol Cell. 2001;8:1231–1241.
24. Fisslthaler B, Dimmeler S, Hermann C, Busse R, Fleming I. Phosphor-ylation and activation of the endothelial nitric oxide synthase by fluidshear stress. Acta Physiol Scand. 2000;168:81–88.
25. Surapisitchat J, Hoefen RJ, Pi X, Yoshizumi M, Yan C, Berk BC. Fluidshear stress inhibits TNF-� activation of JNK but not ERK 1/2 or p38 inhuman umbilical vein endothelial cells: inhibitory crosstalk amongMAPK family members. Proc Natl Acad Sci U S A. 2002;98:6476–6481.
26. Thomson S, Clayton AL, Hazzalin CA, Rose S, Barrat M, MahadevanLC. The nucleosomal response associated with immediate-early geneinduction is mediated via alternative MAP kinase cascades: MSK1 as apotential histone H3/HMG-14 kinase. EMBO J. 1999;17:4779–4793.
27. Deak M, Clifton AD, Lucocq JM, Alessi DR. Mitogen- and stress-acti-vated protein kinase-1 (MSK1) is directly activated by MAPK andSAPK2/p38, and may mediate activation of CREB. EMBO J. 1998;15:4426–4441.
28. Dwarki VJ, Montminy M, Verma IM. Both the basic region and the“leucine zipper” domain of the cyclic AMP response element binding(CREB) protein are essential for transcriptional activation. EMBO J.1990;9:225–232.
29. Garcia-Cardena G, Comander J, Anderson KR, Blackman BR, GimbroneMA Jr. Biomechanical activation of vascular endothelium as a deter-minant of its functional phenotype. Proc Natl Acad Sci U S A. 2001;98:4478–4485.
30. Li S, Hen BPC, Azuma N, Hu YL, Wu SZ, Sumpio BE, Shyy YJJ, ChienS. Distinct roles for the small GTPases Cdc42 and Rho in endothelialresponse to shear stress. J Clin Invest. 1999;103:1141–1150.
31. Lin K, Hsu PP, Chen BP, Yuan S, Usami S, Shyy YJJ, Li YS, Chien S.Molecular mechanism of endothelial growth arrest by laminar shearstress. Proc Natl Acad Sci U S A. 2000;97:9385–9389.
32. Davis ME, Cai H, Drummond GR, Harrison DG. Shear stress regulatesendothelial nitric oxide synthase expression through c-src by divergentsignaling pathways. Circ Res. 2001;89:1073–1080.
33. Go YM, Boo YC, Park H, Maland MC, Patel R, Pritchard KA, Fujio Y,Walsh K, Darley-Usmar V, Jo H. Protein kinase B/Akt activates c-junNH2-terminal kinase by increasing NO production in response to shearstress. J Appl Physiol. 2001;91:1574–1581.
34. Bartling B, Tostlebe H, Darmer D, Holtz J, Silber RE, Morawietz H.Shear stress-dependent expression of apoptosis-regulating genes in endo-thelial cells. Biochem Biophys Res Commun. 2000;278:740–746.
35. Urbich C, Walter DH, Zeiher A, Dimmeler S. Laminar shear stressupregulates integrin expression: role in endothelial cell adhesion andapoptosis. Circ Res. 2000;87:683–689.
36. Narlikar GJ, Fan HI, Kingston RE. Cooperation between complexes thatregulate chromatin structure and transcription. Cell. 2002;108:475–487.
37. Boo YC, Sorescu G, Boyd N, Shiojima I, Walsh K, Du J, Jo H. Shearstress stimulates phosphorylation of endothelial nitric oxide synthase atser1179 by Akt-independent mechanisms: role of protein kinase A. J BiolChem. 2002;27:3388–3396.
38. Liu JS, Park EA, Gurney AL, Roesler WJ, Hanson RW. Cyclic AMPinduction of phosphoenolpyruvate carboxykinase (GTP) gene tran-scription is mediated by multiple promoter elements. J Biol Chem. 1991;266:19095–190102.
39. Parker D, Jhala US, Radhakrishnan I, Yakke MB, Reyes C, Shulman AI,Cantley LC, Wright PE, Montminy M. Analysis of an activator:coacti-vator complex reveals an essential role for secondary structure in tran-scriptional activation. Mol Cell. 1998;2:353–359.
40. Rossig L, Li H, Fisslthaler B, Urbich C, Fleming I, Forstermann U, ZehierAM, Dimmeler S. Inhibitors of histone deacetylation downregulate theexpression of endothelial nitric oxide synthase and compromise endothe-lial cell function in vasorelaxation and angiogenesis. Circ Res. 2002;91:837–844.
41. Kim MS, Kwon HJ, Lee YM, Baek JH, Jang JE, Lee SW, Moon EJ, KimHS, Lee SK, Chung HY, Kim CW, Kim KW. Histone deacetylases induceangiogenesis by negative regulation of tumor suppressor genes. Nat Med.2001;7:437–443.
42. Iezzi S, Cossu G, Nervi C, Sartorelli V, Puri PL. Stage-specific modu-lation of skeletal myogenesis by inhibitors of nuclear deacetylases. ProcNatl Acad Sci U S A. 2002;99:7757–7762.
43. Ping Q, Li L. Histone acetylation and recruitment of serum responsivefactor and CREB-binding protein onto SM22 promoter during SM22 geneexpression. Circ Res. 2002;90:858–865.
Illi et al Flow and Gene Expression 161
by guest on November 16, 2015http://circres.ahajournals.org/Downloaded from

MATERIAL AND METHODS
VEGF and FGF-2 treatments.
HUVEC were cultured in EGM-2 medium, kept in ST and treated with 10 ng/ml VEGF and FGF-2
for 30 to 240 min.
Western Blots
Total protein extracts were obtained by lysing cells with 1X Laemmli buffer. Samples were boiled
10 minutes and centrifuged 5 minutes before gel loading. Histones were resolved on a 18%
denaturing polyacrilamide gel, while c-fos, RSK-2, and MSK-1 were resolved on a 10% denaturing
polyacrilamide gel. Gels were blotted onto a nitrocellulose membrane (Hybond-C-extra,
Amersham) for 3 hours at 400 mA, at +4°C. Membranes were saturated in 3% non fatty dry milk
and probed with anti-acetylated histone H3K14, anti-acetylated histone H4, anti-phospho histone
H3S10, anti-phosphoS10-acetylK14 histone H3 (Upstate Biotechnology) anti-c-fos (Santa Cruz), anti
phspho-RSK-2(Ser380), anti-phospho-MSK1(Ser376) (Cell Signalling) antibodies, according to
manufacturer’s instructions. Normalization of protein loading was obtained using an anti-GRB-2
antibody (Santa Cruz), anti-histone H1 antibody (Upstate Biotechnology), anti-RSK-2 and anti-
MSK-1 antibodies (Cell Signalling).
“Pull down” assay
Cells were exposed to SS for one hour or kept in static conditions. Nuclear extracts were obtained
as previously described (21) and dialyzed over-night at + 4°C against 20 mmol/L Hepes pH 7.9, 50
mmol/L KCl, 40% glycerol, 0.2 mmol/L EDTA pH 8.
100 µl of dynabeads (DYNAL-M280, Oxoid) were equilibrated in 10 mmol/L Tris.HCl pH 7.5, 1
mmol/L EDTA, 2 mol/L NaCl and conjugated with 100 pmoles of double strand oligonucleotide

containing a cAMP Response Element (CRE) two hours at room temperature. Conjugated oligos
were washed three times with 10 mM/L Tris HCl pH 7.5, 1 mM/L EDTA, 2 mol/L NaCl and
blocked with 5% BSA in 20 mmol/L Hepes pH 7.9, 50 mM/L KCl, 40% Glycerol, 0.2 mmol/L
EDTA pH 8 one hour at + 4°C. After blocking, oligos were recovered with a magnet (Oxoid),
washed once as above and incubated over-night at + 4°C with 200 µg of recovered nuclear proteins
(50 pmoles for each experimental condition).
The day after, bound complexes were washed once with 100mmol/L KCL, 20 mmol/L Hepes pH
7.9, 40% Glycerol, 0.2 mmol/L EDTA pH 8, 0.01% NP-40. Beads were resuspended in 20 µl of
water and and treated with DNAse I (Roche Biochemicals) 30’ at 37°C in 40 mmol/L MgCl2 2
mmol/L DTT to release bound complexes.Reaction was stopped with 1.5 mol/L CH3COONa pH
5.2, 50 mmol/L EDTA pH 8, 1% SDS. Recovered nuclear complexes were boiled in Laemmli
buffer 1X and resolved on a 10% denaturing polyacrilamide mini-gel. Gel was blotted onto a
nitrocellulose membrane (Hybond C-Extra, Amersham Biosciences), for 2 hours at 200 mA at +
4°C. Membrane was blocked in PBS/0,05% Tween-20 plus 5% non fatty dry milk for 1 hour at
room temperature and probed with anti-CREB and anti-CBP antibodies (Santa Cruz),according to
manufacturer’s instructions.
Oligos sequences were: AGAGATTGCCTGACGTCAGAGAGCTAG and
CTAGCTCTCTGACGTCAGGCAATCTCT. CRE sequence is underlined.
“In vitro” acetylation assay
Whole cell extracts were obtained as described (2). Histone acetyl transferase activity was detected
using the HAT-check activity assay kit (PIERCE) according to manufacturer’s instruction. Briefly,
cell extracts (100 µg) were incubated with a biotynilated-histone H4 peptide, as substrate, in
presence of 250 nCi of 3H-AcetylCoA, for 1 hour, at 30°C. The mixture was then rocked for 30
minutes at room temperature with 100 µl of streptoavidin-conjugated agarose beads and extensively

washed with RIPA buffer. The amount of radiolabelled H4 peptide was measured using a liquid
scintillation counter TRI-CARB (PACKARD).

RESULTS
VEGF, FGF-2 and SS have different effects on histone H3 modifications.
Epithelial Growth Factor (EGF) has been reported to be a potent inducer of histone H3
phosphorylation.1, 2 Since VEGF and FGF-2 are potent proliferation and differentiation stimuli for
ECs, we investigated their effect on histone H3 modifications in HUVEC. As shown in Fig. 1
VEGF induced about 2 fold acetylation and phosphorylation of histone H3 after 30 min. of
treatment with a peak at 60 min (~ 3 fold). FGF-2 had no effect on H3 acetylation, enhancing only
H3 phosphorylation from 1,5 to 3 fold at any time point. Therefore, these GFs, although sharing
share common features with SS in their capacity of modifying histone H3 result different in terms
of kinetic and intensity. Specifically, SS effects in H3 modifications are more prolonged and
stronger than those of GFs alone.

REFERENCES
1. Sassone Corsi P, Mizzen CA, Cheung P, Crosio C, Monaco L, Jacquot S, Hanauer A, Allis CD.
Requirement of Rsk-2 for epidermal growth factor-activated phosphorylation of histone H3.
Science. 1999; 285: 886-891.
2. Merienne K, Pannetier S, Harel-Bellan A, Sassone-Corsi P. Mitogen-regulated Rsk2-CBP
interaction controls their kinase and acetylase activity. Mol Cell Biol. 2001; 21: 7089-7096.

FIGURE LEGENDS
Fig 1E: VEGF and FGF-2 induce acetylation and phosphorylation of histone H3
Figure shows that upon VEGF treatment histone H3 acetylation increases after 30 min. of treatment
with a peak at 60 min.; this penomenon decreases at 120 min. H3 phosphorylation significantly
increases about 1,5 fold and 2,5 fold after 30 and 60 min. respectively and is lost at 120 min. FGF-2
has no effect on H3 acetylation; conversely, enhances phosphorylation at any time point. P value
was *(p<0.05).

Capogrossi and Carlo GaetanoBarbara Illi, Simona Nanni, Alessandro Scopece, Antonella Farsetti, Paolo Biglioli, Maurizio C.
Regulation of Gene ExpressionMediated Chromatin Remodeling Provides Molecular Basis for Flow-Dependent−Shear Stress
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2003 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.0000080933.82105.292003;93:155-161; originally published online June 12, 2003;Circ Res.
http://circres.ahajournals.org/content/93/2/155World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2003/07/24/93.2.155.DC1.htmlData Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer process is available in the
click Request Permissions in the middle column of the Web page under Services. Further information about this located,Editorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Research Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on November 16, 2015http://circres.ahajournals.org/Downloaded from





























