Embed Size (px)
Citation preview

Biochem. J. (2015) 468, 345–352 doi:10.1042/BJ20150030 345
The tumour suppressor CHD5 forms a NuRD-type chromatin remodellingcomplexVenkatadri Kolla*, Koumudi Naraparaju*, Tiangang Zhuang*, Mayumi Higashi*, Sriharsha Kolla*, Gerd A. Blobel†and Garrett M. Brodeur*1
*Division of Oncology, Department of Pediatrics, The Children’s Hospital of Philadelphia, University of Pennsylvania, Philadelphia 19104, PA, U.S.A.†Division of Hematology, Department of Pediatrics, The Children’s Hospital of Philadelphia, University of Pennsylvania, Philadelphia 19104, PA, U.S.A.
Eukaryotic gene expression is developmentally regulated, inpart by chromatin remodelling, and its dysregulation has beenlinked to cancer. CHD5 (chromodomain helicase DNA-bindingprotein 5) is a tumour suppressor gene (TSG) that maps toa region of consistent deletion on 1p36.31 in neuroblastomas(NBs) and other tumour types. CHD5 encodes a protein withchromatin remodelling, helicase and DNA-binding motifs that ispreferentially expressed in neural and testicular tissues. CHD5is highly homologous to CHD3 and CHD4, which are the coresubunits of nucleosome remodelling and deacetylation (NuRD)complexes. To determine if CHD5 forms a similar complex,we performed studies on nuclear extracts from NBLS, SY5Y(both with endogenous CHD5 expression), NLF (CHD5 null) andNLF cells stably transfected with CHD5 cDNA (wild-type andV5–histidine-tagged). Immunoprecipitation (IP) was performedwith either CHD5 antibody or antibody to V5/histidine-taggedprotein. We identified NuRD components both by GST–FOG1(Friend Of GATA1) pull-down and by IP. We also performed
MS/MS analysis to confirm the presence of CHD5 or other proteincomponents of the NuRD complex, as well as to identify othernovel proteins. CHD5 was clearly associated with all canonicalNuRD components, including metastasis-associated protein(MTA)1/2, GATA zinc finger domain containing 2A (GATAD2A),histone deacetylase (HDAC)1/2, retinoblastoma-binding protein(RBBP)4/7 and methyl DNA-binding domain protein (MBD)2/3,as determined by Western blotting and MS/MS. Our datasuggest CHD5 forms a NuRD complex similar to CHD4.However, CHD5–NuRD may also have unique proteinassociations that confer functional specificity and may contributeto normal development and to tumour suppression in NB andother cancers.
Key words: chromodomain helicase DNA-binding protein5 (CHD5), neuroblastoma, nuclear localization, nucleosomeremodelling histone deacetylase (NuRD) complex, tumoursuppressor.
INTRODUCTION
Neuroblastoma (NB) is the most common extracranial solidtumour of childhood. This tumour of the sympathetic nervoussystem accounts for 8%–10% of childhood cancers and 15% ofchildhood cancer deaths [1,2]. NBs may regress spontaneously,especially in infants, or grow relentlessly despite intensive,multimodality therapy. We and others have identified distinctpatterns of genomic changes that underlie these disparate clinicalbehaviours [1,3]. Deletion of the short arm of chromosome 1(1p) has been observed in 35% of primary NBs and 70 %of NB-derived cell lines [4–6]. We first identified 1p deletionas a characteristic change in advanced stage NBs [7], whichpresumably reflects loss of one or more tumour suppressor genes(TSGs) from this region. We used DNA-based polymorphisms[6,8] to refine the smallest region of consistent deletion (SRD)of 1p36 to a ∼2 Mb region on 1p36.31 and we identified CHD5(chromodomain helicase DNA-binding protein 5) as a bona fideTSG from this region in NBs [9–12]. CHD5 expression is low orabsent from NB cell lines and most high-risk tumours and lowexpression is associated with unfavourable features and outcome[9–11,13,14]. Bagchi et al. [15] also identified CHD5 as a TSG
on the orthologous region of mouse chromosome 4 using achromosome engineering approach. Furthermore, CHD5 has beenimplicated as a TSG in a variety of other cancers, such as gliomasand cancers of the colon, breast, lung, ovary, prostate, stomach,larynx and gall bladder [15–27].
CHD5 is a member of the chromodomain-helicase-DNA-binding (CHD) family [11,28,29]. Currently the CHD family hasnine members and they are divided into three subfamilies [27,30].CHD1 and CHD2 comprise the first subfamily, which containsa classic DNA-binding domain. The second subfamily consistsof CHD3 (Mi2α) and CHD4 (Mi2β), which have two PHD-zinc finger motifs and each forms a nucleosome remodelling anddeacetylation (NuRD) complex. The third subfamily consists ofCHD6–CHD9, which was originally identified based on structuralhomology to other known CHD members [29,31]. Of all theCHD family members, CHD5 is most homologous to CHD3 andCHD4. Indeed, CHD3–CHD5 are the only members that havetwo PHD domains near the N-terminus, in addition to pairedchromodomains and a split SWItch/Sucrose NonFermentable(SWI/SNF)-like helicase/ATPase domain [32]. Thus, CHD5belongs most appropriately in the second subfamily. However,CHD5 is expressed almost exclusively in the nervous system
Abbreviations: aa, amino acids; CHD, chromodomain-helicase-DNA binding; CHD4/5, chromodomain helicase DNA-binding protein4/ 5; GAPDH,glyceraldehyde-3-phosphate dehydrogenase; HDAC, histone deacetylase complex; IF, immunofluorescence; IP, immunoprecipitation; MBD, methyl DNA-binding protein; MTA, metastasis-associated protein; NB, neuroblastoma; NF-kB, nuclear factor kappa-light-chain-enhancer of activated B-cells; NuRD,nucleosome remodelling and deacetylation; RBBP, retinoblastoma-binding protein; RPMI, Roswell Park Memorial Institute medium; TSG, tumour suppressorgene.
1 To whom correspondence should be addressed (email [email protected]).
c© The Authors Journal compilation c© 2015 Biochemical Society
Bio
chem
ical
Jo
urn
al
ww
w.b
ioch
emj.o
rg

346 V. Kolla and others
and in testis, whereas CHD3 and CHD4 are expressed moreubiquitously [27,33].
Gene regulation is a highly co-ordinated process that involvesordered recruitment of transcriptional machinery to maintainproper chromatin structure and chromatin remodelling proteinsplay crucial roles in this process [34]. NuRD complexes werefirst identified more than 15 years ago [34,35] and they alterchromatin structure and gene expression in part by causing ATP-dependent remodelling of nucleosomes [36]. Most studies havefocused on the Mi2β/CHD4–NuRD complex, which has bothATPase and histone deacetylase activity [34]. Other canonicalNuRD components are: metastasis-associated proteins 1 and 2(MTA1/2), retinoblastoma-binding proteins (RBBP)4 and RBBP7(formerly RbAp48 and RbAp46 respectively), GATAD2A and -B (formerly p66α/β) and methyl DNA-binding domain proteins(MBD2/3) [28,37,38].
NuRD complexes have been implicated in regulating genetranscription, genome stability, DNA damage and DNA repair[28,37,39–41]. Altered NuRD function is associated with a varietyof cancers, as described above [15–27]. CHD5 has also beenshown to regulate the expression of neural-specific genes that areimplicated in aging, as well as in Alzheimer’s disease [42,43].However, it has been unclear if CHD5 forms a NuRD complexsimilar to CHD3 and CHD4 or whether CHD5 forms uniqueprotein associations. In the present study, we provide evidencethat CHD5 forms a bona fide NuRD complex with all thesame canonical proteins as CHD4. Furthermore, CHD5 formsnovel protein associations that may account for the functionaldifferences between CHD4– and CHD5–NuRD complexes.
EXPERIMENTAL
Reagents
Cell culture media [Roswell Park Memorial Institute medium(RPMI-1640)], antibiotics and FBS were obtained fromInvitrogen Inc. Parental NB cell lines were maintained in ourlaboratory but are also available from the A.T.C.C. Restrictionenzymes and other molecular biology reagents were purchasedfrom Roche Applied Sciences, Promega Inc. and New EnglandBiolabs Inc. ‘Complete’ protease inhibitor cocktail tablets wereobtained from Roche Applied Sciences. Halt protease andphosphatase inhibitor cocktail was obtained from Sigma–AldrichCompany. Nuclear extraction kits were obtained from ThermoScientific. NuPAGE gels (4%–12%), buffers and prestainedRainbow molecular mass markers were obtained from Invitrogen.
Antibodies
Rabbit polyclonal CHD5 antibody and goat primary polyclonalantibodies for MTA-1, MTA-2, RBBP4, RBBP7, histonedeacetylase (HDAC)1, HDAC2, MBD2 and MBD3 werefrom Santa Cruz Biotechnology. CHD4 antibody was fromBethyl Laboratories. Polyclonal antibodies for HDAC1,HDAC2, RBBP7, RBBP4/7 and MDB3 were from CellSignaling Technology. GATAD2A antibody was from UpstateBiotechnology. Tagged V5–His was from Invitrogen. Secondaryantibodies were from GE Healthcare Life Sciences and SantaCruz.
Plasmid cloning
CHD5–ORF2 was amplified directly from brain total RNA(by cDNA cloning) and subcloned after PCR amplification.
Expression plasmids for the CHD5 transcript were generated in aeukaryotic V5–His-tagged pcDNA3.1 expression plasmid at NotIand XhoI sites. DNA sequencing from both ends and restrictionenzyme analyses confirmed transcript fidelity. GST and GST–FOG1 [N-terminal 45 amino acids (aa)] plasmids were describedpreviously [44].
Cell culture and transfections
We transfected the NLF NB cell line [9–12] either with a CHD5expression vector (NLF–CHD5), with or without a V5–His-tag or with empty pcDNA 3.1 vector. NB cell lines (NBLS,SY5Y and NLF) were cultured in RPMI-1640 + 10% FBSand gentamicin, along with the appropriate selection antibiotics.Transfections of NLF cells were performed using Lipofectamine2000 (Invitrogen). Cells were cultured for 6–8 weeks in thepresence of G418 (400 μg/ml) to select single cell clones. Singlecell suspensions were cultured in the presence of neomycin toobtain clones derived from single cells.
Immunofluorescence
Parental NLF and NLF–CHD5 were cultured on cover slipsin six-well plates. Forty-eight hours after plating, cells werefixed with 4% paraformaldehyde (Electron Microscopy Sciences)in PBS for 15 min at room temperature. Cells were washedgently twice and permeabilized with 0.2 % Triton X-100 in 1%BSA, followed by immunostaining using either rabbit anti-CHD5(1:1000) or tagged monoclonal anti-mouse V5–His (1:1000;Invitrogen) followed by Alexa Fluor 488, goat anti-rabbit IgG(A11008) and Fluor 546, goat anti-mouse IgG (A11030) assecondary antibodies (1:400–1:2000). Coverslips were mountedand exposed to ProLong®Gold Antifade Reagent with DAPI(0.1 μg/ml, Molecular Probes) for 10 min to stain nuclei. Imageswere captured using a phase contrast microscope and analysedusing Olympus 1×70 with Slidebook4 software (UniversalImaging).
Cell extract preparation
Subcellular fractionated proteins (cytosol, membrane, nuclearand cytoskeleton) were extracted using commercially availablekits from ThermoFisher Scientific. Nuclear and cytosolicfractions were prepared with two buffer (low- and high-salt)extraction, as described previously [44]. Protein concentrationswere determined using Bradford Protein Assay Reagent withSmartSpec Plus spectrophotometer (BIO-RAD Laboratories).
Immunoprecipitation
Immunoprecipitations (IPs) of CHD5– and CHD4–NuRDcomplexes were performed using nuclear protein (100–2000 μg)from NBLS, SY5Y, parental NLF, as well as NLF–CHD5 cells.Typical IP buffer contained protease (Sigma P-8340; 1:500)and phosphatase inhibitors (50 mM Na fluoride, 10 mM Napyrophosphate, 5 mM Na vanadate). Extracts were pre-clearedwith agarose beads and incubated overnight using either anti-CHD5 antibody (1–25 μl) or V5–His tag antibody (1–10 μl) in thepresence of protease inhibitors. Bound complexes were washedand lysed in 20 μl lysis buffer and aliquots were applied to either4%–12% gradient or SDS/PAGE (10 % gel).
c© The Authors Journal compilation c© 2015 Biochemical Society

CHD5–NuRD complex in neuroblastoma cell lines 347
Western analysis and GST–FOG1 pull-downs
Western blot analyses for CHD5 and CHD5–NuRD componentswere performed following PAGE, as described previously[44,45], using NuPAGE Bis-Tris gels with MOPS-SDS runningbuffer (Invitrogen). Proteins were transferred to nitrocellulosemembranes (GE Healthcare Life Sciences) and probed withantibodies as described above using ECL detection (Invitrogen).We performed in vitro binding studies of GST, GST–FOG1(45 aa N-terminal fragment) a GATA1 cofactor, to pull downa CHD4 (Mi-2β), as described earlier [44]. We used a GST–FOG1 construct to pull down complete NuRD complexes fromNB cells. Twenty micrograms of either GST or GST–FOG1 wasincubated with 2 mg of NLF or NLF–CHD5 nuclear extractovernight in buffer containing 150 mM NaCl, 50 mM Tris/HCl(pH 7.5), 0.5 % Igepal (Sigma), protease inhibitor cocktail (P-8340, Sigma, 1:500) and 1 mM DTT. Bound proteins were washedtwice in the above buffer with 350 mM NaCl, then three timeswith 650 mM NaCl buffer, followed by an additional wash in350 mM NaCl, then separated by SDS/PAGE and stained withSimplyBlue (Invitrogen), a modified form of Coomassie Blue.Aliquots were also analysed by Western blot with appropriateNuRD component antibodies.
Protein sequence analysis by LC-MS/MS
LC-MS/MS analysis to identify proteins associated with CHD4and CHD5 was performed at the Taplin Biological MassSpectrometry Facility (Harvard Medical School). Nuclear extractswere immunoprecipitated with specific antibody and subjectedto SDS/PAGE (10 % gel) followed by Coomassie/Simply Bluestaining. Excised gel bands were cut into approximately 1 mm3
pieces. Gel pieces were then subjected to a modified in-gel trypsindigestion procedure [46]. Gel pieces were washed and dehydratedwith acetonitrile for 10 min followed by removal of acetonitrile.Pieces were then completely dried in a speed-vac. Rehydrationof the gel pieces was with 50 mM ammonium bicarbonatesolution containing 12.5 ng/μl modified sequencing-grade trypsin(Promega) at 4 ◦C. After 45 min, the excess trypsin solutionwas removed and replaced with 50 mM ammonium bicarbonatesolution to just cover the gel pieces. Samples were then placed in a37 ◦C room overnight. Peptides were later extracted by removingthe ammonium bicarbonate solution, followed by one wash witha solution containing 50% acetonitrile and 1 % formic acid. Theextracts were dried in a speed-vac (∼1 h). The samples were thenstored at 4 ◦C until analysis.
On the day of analysis, the samples were reconstituted in 5–10 μl of HPLC solvent A (2.5% acetonitrile, 0.1 % formic acid).A nanoscale reverse-phase HPLC capillary column was createdby packing 5 μm of C18 spherical silica beads into a fused silicacapillary (100 μm inner diameter × ∼25 cm length) with a flame-drawn tip [47]. After equilibrating the column, each sample wasloaded via a Famos auto sampler (LC Packings) on to the column.A gradient was formed and peptides were eluted with increasingconcentrations of solvent B (97.5 % acetonitrile, 0.1 % formicacid).
As peptides eluted, they were subjected to ESI and then enteredinto an LTQ Orbitrap Velos Pro ion-trap mass spectrometer(ThermoFisher Scientific). Peptides were detected, isolated andfragmented to produce an MS/MS of specific fragment ions foreach peptide. Peptide sequences (and hence protein identity)were determined by matching the acquired fragmentation patternwith protein databases using the software program, Sequest(ThermoFisher) [48]. All databases include a reversed versionof all the sequences and the data were filtered to between a 1 %
and 2% false discovery rates (FDRs). Results were tabulated incolumns, indicating gene symbol/name, total peptides identifiedand unique peptides for specific protein.
RESULTS
CHD5 expression is localized to the nucleus
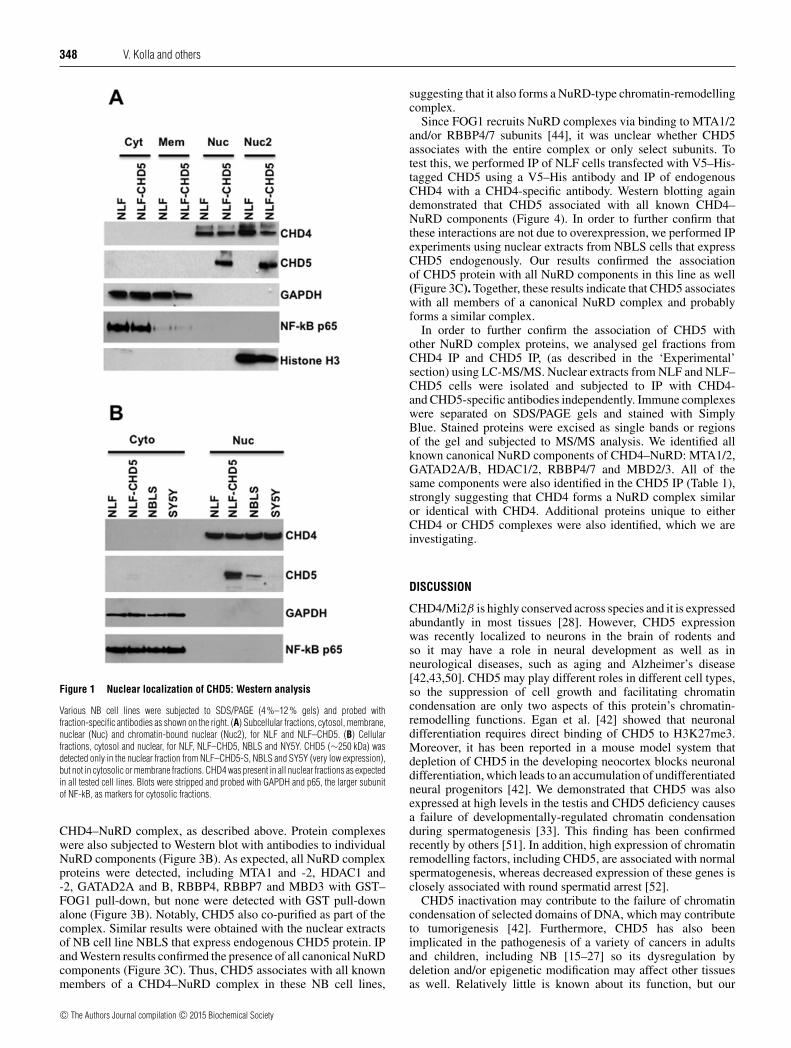
In order to determine the subcellular localization of CHD5, weperformed a series of Western analyses using various cell fractions(cytosol, membrane, nuclear and chromatin bound nuclear). Ourresults indicate that the CHD5 was detected only in the nuclearfractions from NLF–CHD5, but absent from parental NLF cells,whereas CHD4 was detected in both NLF–CHD5 and parentalNLF cells (Figures 1A and 1B). CHD5 protein was detected inboth nuclear and chromatin-bound nuclear fractions, but it wasnot detected in appreciable amounts in cytoplasmic or membranecompartments. To validate this subcellular fractionation, blotswere stripped and probed with cytosol-specific antibodies suchas glyceraldehyde-3-phosphate dehydrogenase (GAPDH) andthe larger subunit of NF-kB (nuclear factor kappa-light-chain-enhancer of activated B-cells) p65. As anticipated, we detectedGAPDH and NF-kB proteins only in the cytosolic fractions(Figure 1A), indicating that our cell fractions were specific toeach subcellular compartment. In order to confirm that theseobservations were not due to CHD5 overexpression in NLFtransfected cells, we performed Western blot analysis usingnuclear extracts from NBLS and SY5Y cells that expressendogenous CHD5. We observed that both NBLS and SY5Ycells expressed CHD4 and CHD5 only in the nucleus, althoughthe expression level of CHD5 was much lower in NBLS and SY5Ycells when compared with NLF–CHD5 ( Figure 1B). Expressionpatterns of cytosol-specific markers GAPDH and NF-kB p65 wereconsistent whether the cells expressed CHD5 endogenously or byoverexpression (Figure 1B).
To further confirm these observations of nuclear localizationof CHD5, we performed immunofluorescence (IF) studies witha CHD5-specific polyclonal antibody. Parental NLF cells did notexpress CHD5 protein and served as negative control. IF resultsfrom transfected NLF–CHD5 cells demonstrated that CHD5 wasexpressed only in the nucleus (Figure 2A). This was confirmedby analysis of independent NLF clones expressing CHD5 taggedwith the V5–His epitope using a V5–His antibody, which alsodemonstrated exclusive expression in the nucleus (Figure 2B). Wealso performed parallel experiments with the NBLS cell line thatexpresses CHD5 endogenously and we obtained similar results(Figure 2C). These results demonstrate that CHD5 is expressedexclusively in the nucleus, consistent with a role in chromatinremodelling, similar to CHD4.
CHD5 associates with all NuRD complex proteins
CHD4–NuRD binds to the transcription cofactor FOG1 via ahighly conserved short motif at the FOG1 N-terminus [44] and thisregion is required for the transcription functions of FOG1 [49].Fusion of this peptide to GST allows purification of the CHD4–NuRD complex to high purity by a single step. To determine ifCHD5 forms a NuRD complex similar to CHD4, we performedGST–FOG1 pull-downs with nuclear extracts from parental NLFand NLF–CHD5, followed by stringent washing with 150–650 mM NaCl. Bound proteins were subjected to SDS/PAGE andstained with SimplyBlue (Figure 3A). Well-separated, stainedproteins (Figure 3A) were also excised and identified byMS/MS. All bands were identified as known components of the
c© The Authors Journal compilation c© 2015 Biochemical Society
348 V. Kolla and others
Figure 1 Nuclear localization of CHD5: Western analysis
Various NB cell lines were subjected to SDS/PAGE (4 %–12 % gels) and probed withfraction-specific antibodies as shown on the right. (A) Subcellular fractions, cytosol, membrane,nuclear (Nuc) and chromatin-bound nuclear (Nuc2), for NLF and NLF–CHD5. (B) Cellularfractions, cytosol and nuclear, for NLF, NLF–CHD5, NBLS and NY5Y. CHD5 (∼250 kDa) wasdetected only in the nuclear fraction from NLF–CHD5-S, NBLS and SY5Y (very low expression),but not in cytosolic or membrane fractions. CHD4 was present in all nuclear fractions as expectedin all tested cell lines. Blots were stripped and probed with GAPDH and p65, the larger subunitof NF-kB, as markers for cytosolic fractions.
CHD4–NuRD complex, as described above. Protein complexeswere also subjected to Western blot with antibodies to individualNuRD components (Figure 3B). As expected, all NuRD complexproteins were detected, including MTA1 and -2, HDAC1 and-2, GATAD2A and B, RBBP4, RBBP7 and MBD3 with GST–FOG1 pull-down, but none were detected with GST pull-downalone (Figure 3B). Notably, CHD5 also co-purified as part of thecomplex. Similar results were obtained with the nuclear extractsof NB cell line NBLS that express endogenous CHD5 protein. IPand Western results confirmed the presence of all canonical NuRDcomponents (Figure 3C). Thus, CHD5 associates with all knownmembers of a CHD4–NuRD complex in these NB cell lines,
suggesting that it also forms a NuRD-type chromatin-remodellingcomplex.
Since FOG1 recruits NuRD complexes via binding to MTA1/2and/or RBBP4/7 subunits [44], it was unclear whether CHD5associates with the entire complex or only select subunits. Totest this, we performed IP of NLF cells transfected with V5–His-tagged CHD5 using a V5–His antibody and IP of endogenousCHD4 with a CHD4-specific antibody. Western blotting againdemonstrated that CHD5 associated with all known CHD4–NuRD components (Figure 4). In order to further confirm thatthese interactions are not due to overexpression, we performed IPexperiments using nuclear extracts from NBLS cells that expressCHD5 endogenously. Our results confirmed the associationof CHD5 protein with all NuRD components in this line as well(Figure 3C). Together, these results indicate that CHD5 associateswith all members of a canonical NuRD complex and probablyforms a similar complex.
In order to further confirm the association of CHD5 withother NuRD complex proteins, we analysed gel fractions fromCHD4 IP and CHD5 IP, (as described in the ‘Experimental’section) using LC-MS/MS. Nuclear extracts from NLF and NLF–CHD5 cells were isolated and subjected to IP with CHD4-and CHD5-specific antibodies independently. Immune complexeswere separated on SDS/PAGE gels and stained with SimplyBlue. Stained proteins were excised as single bands or regionsof the gel and subjected to MS/MS analysis. We identified allknown canonical NuRD components of CHD4–NuRD: MTA1/2,GATAD2A/B, HDAC1/2, RBBP4/7 and MBD2/3. All of thesame components were also identified in the CHD5 IP (Table 1),strongly suggesting that CHD4 forms a NuRD complex similaror identical with CHD4. Additional proteins unique to eitherCHD4 or CHD5 complexes were also identified, which we areinvestigating.
DISCUSSION
CHD4/Mi2β is highly conserved across species and it is expressedabundantly in most tissues [28]. However, CHD5 expressionwas recently localized to neurons in the brain of rodents andso it may have a role in neural development as well as inneurological diseases, such as aging and Alzheimer’s disease[42,43,50]. CHD5 may play different roles in different cell types,so the suppression of cell growth and facilitating chromatincondensation are only two aspects of this protein’s chromatin-remodelling functions. Egan et al. [42] showed that neuronaldifferentiation requires direct binding of CHD5 to H3K27me3.Moreover, it has been reported in a mouse model system thatdepletion of CHD5 in the developing neocortex blocks neuronaldifferentiation, which leads to an accumulation of undifferentiatedneural progenitors [42]. We demonstrated that CHD5 was alsoexpressed at high levels in the testis and CHD5 deficiency causesa failure of developmentally-regulated chromatin condensationduring spermatogenesis [33]. This finding has been confirmedrecently by others [51]. In addition, high expression of chromatinremodelling factors, including CHD5, are associated with normalspermatogenesis, whereas decreased expression of these genes isclosely associated with round spermatid arrest [52].
CHD5 inactivation may contribute to the failure of chromatincondensation of selected domains of DNA, which may contributeto tumorigenesis [42]. Furthermore, CHD5 has also beenimplicated in the pathogenesis of a variety of cancers in adultsand children, including NB [15–27] so its dysregulation bydeletion and/or epigenetic modification may affect other tissuesas well. Relatively little is known about its function, but our
c© The Authors Journal compilation c© 2015 Biochemical Society

CHD5–NuRD complex in neuroblastoma cell lines 349
Figure 2 Nuclear localization of CHD5-IF
(A) Parental NLF cells (top row) and NLF–CHD5 (second row) were cultured for 48 h. Cells were immunostained with CHD5 as indicated. Nuclei were stained with DAPI (left panels). Nuclearlocalization of CHD5 was observed when cells overexpressed CHD5 (middle panels), whereas no nuclear staining was observed in control NLF cells (upper middle panel). Merged images are shownon right. (B) IF images indicating nuclear expression with tagged V5–His antibody. (C) Nuclear expression of CHD5 in NBLS cells expressing endogenous CHD5.
results strongly suggest that CHD5 functions as part of a NuRD-type chromatin-remodelling complex. Nevertheless, the exactmechanism by which CHD5 functions as a TSG in NBs or othercancers is still unclear. Paul et al. [53] recently reported thatPHD-mediated histone 3 binding, involving chromatin mediatedtranscriptional regulation, is required for CHD5-mediated tumoursuppression. Whether CHD5 functions independently of a NuRD-type complex is unknown at present.
We demonstrate, in the present study, by both GST–FOG1 pull-down experiments and IP studies, that CHD5 is associated withall canonical members of a CHD4–NuRD complex (MTA1/2,HDAC1/2, GATAD2A/B, RBBP4/7, MBD2/3). Thus, basedon its homology with CHD4 (Supplementary Table S1), itsnuclear localization and its association with all typical CHD4–NuRD components, it is extremely likely that CHD5 forms aNuRD-type chromatin-remodelling complex. This is consistent
c© The Authors Journal compilation c© 2015 Biochemical Society

350 V. Kolla and others
Figure 3 Detection of CHD4– and CHD5–NuRD components: GST–FOG1 pull-down
(A) Nuclear extracts of NLF–CHD5 were subjected to either GST alone or GST–FOG1 pull-down followed by PAGE and stained with Simply Blue. All canonical NuRD components from CHD4/5–NuRDcomplexes were readily visible (marked with arrows). (B) GST–FOG1 bound protein complexes from NLF–CHD5 and NLF parental cells were subjected to 4 %–12 % PAGE. The identity of individualproteins from the NuRD complex was confirmed by Western blot analysis. CHD5 was detected only in NLF–CHD5-S extracts but absent from NLF extracts. Additional bands in the CHD5 lane areprobably due to proteolytic degradation products and additional bands with MTA1/2 probably represent alternate isoform recognition. (C) Identification of individual canonical NuRD componentsin NBLS cells. Nuclear extracts were immunoprecipitated with either IgG or CHD5 antibody as indicated and complexes were subjected to SDS/PAGE (4 %–12 % gels) followed by Western blotanalysis. MTA1/2, GATAD2A/B, HDAC1/2, RBBP4/7 and MBD2/3 were detected as individual canonical NuRD components along with CHD5.
Figure 4 Detection of CHD5–NuRD components: IP and Western analysis
Nuclear extracts from NLF–CHD5 and NLF were immunoprecipitated as indicated. Subsequentproteins from IP and GST–FOG1 pull-down (left lane) were subjected to 4 %–12 % PAGE.Extracts from NLF following GST–FOG1 pull-down served as negative control. Individual NuRDcomponents were detected by Western blotting with antibodies specific to each NuRD component.The boxes around the NLF–CHD5 and NLF lanes highlight the identification of all canonicalNuRD components in NLF–CHD5 and not in NLF as a negative control, as these cells do notexpress CHD5 endogenously. FT, flow through.
Table 1 IP/MS identified known canonical NuRD components
Nuclear extracts expressing CHD5 were immunoprecipitated with CHD5 (A) or CHD4 (B)antibodies independently followed by SDS/PAGE analysis. Gels were stained with Simply Blueand excised gel bands were sent for MS analysis. MS analysis revealed the list of knowncanonical NuRD components as shown below. Total, number of total peptides present in a givensample application; Unique, number of peptides that are unique and specific to the indicatedgene/protein description.
A. CHD5–NuRD
Gene symbol Description Unique Total
CHD5 Chromodomain helicase DNA-binding protein 5 56 97MTA1 Metastasis-associated 1 18 21MTA2 Metastasis-associated 2 27 34GATAD2A/p66α GATA zinc finger domain containing 2A 23 41GATAD2B/p66β GATA zinc finger domain containing 2B 15 26HDAC1 Histone deacetylase 1 11 16HDAC2 Histone deacetylase 2 7 11RBBP4/RbAp48 Retinoblastoma-binding protein 4 14 20RBBP7/RbAp46 Retinoblastoma-binding protein 7 4 5MBD2 Methyl-CpG-binding domain protein 2 4 4MBD3 Methyl-CpG-binding domain protein 3 7 8
B. CHD4–NuRD
Gene symbol Description Unique Total
CHD4 Chromodomain helicase DNA-binding protein 4 91 275MTA1 Metastasis-associated 1 20 30MTA2 Metastasis-associated 2 28 45GATAD2A/p66α GATA zinc finger domain containing 2A 27 77GATAD2B/p66β GATA zinc finger domain containing 2B 25 37HDAC1 Histone deacetylase 1 15 28HDAC2 Histone deacetylase 2 10 26RBBP4/RbAp48 Retinoblastoma-binding protein 4 18 42RBBP7/RbAp46 Retinoblastoma-binding protein 7 5 6MBD2 Methyl-CpG-binding domain protein 2 7 8MBD3 Methyl-CpG-binding domain protein 3 9 14
c© The Authors Journal compilation c© 2015 Biochemical Society

CHD5–NuRD complex in neuroblastoma cell lines 351
with the report by Potts et al. [43] who also found anassociation of CHD5 with several NuRD components. It isunknown whether CHD5 competes with CHD4 or CHD3 forNuRD components, but all NuRD components appear to beexpressed abundantly. Our data provide evidence that CHD5must have different functions than CHD4 in NBs as well as othertissues.
First, CHD4 is expressed ubiquitously in almost all tissues inthe body, whereas CHD5 expression is very restricted, with thehighest expression in the nervous system and testis [11,33,42,43].Second, NBs with 1p deletion and virtually no CHD5 expressionhave abundant expression of CHD4, yet they grow readily bothin vitro and in vivo [9,10,13]. When these same NBs weretransfected with CHD5, clonigenicity and tumorigenicitywere dramatically suppressed [9]. Indeed, CHD5 transfectionhas a similar effect on other cancers with 1p36 deletion andCHD5 transcriptional silencing [19–24,54–56]. Thus, CHD4–NuRD cannot simply substitute for CHD5–NuRD in thesecells.
One formal possibility was that CHD5 did not associate withall the same proteins as did CHD4 in a NuRD complex. However,our data demonstrate by both Western analysis and MS analysisthat all components known to be associated with CHD4–NuRD(MTA1/2, GATAD2A/B, HDAC1/2, RBBP4/7 and MBD2/3)were also associated with CHD5 (Table 1). Nevertheless, CHD4is expressed abundantly in NLF, yet cell growth, colony formationand tumour formation is inhibited only when CHD5 is introduced[9]. This suggests that CHD5 may have protein interactions withother proteins or functions that are entirely independent of itsrole as part of a NuRD complex and we are exploring bothpossibilities.
The functional characterization of CHD5, alone or as part ofa CHD5–NuRD complex, would provide insight into the roleof CHD5 in epigenetic modification of gene expression in normaldevelopment and in cancer, including NB. Targeting CHD5 forup-regulation would be theoretically possible, because even intumours with 1p deletion and loss of one copy, the remaining alleleis rarely if ever mutated. Instead, the promoter of the remainingallele is frequently methylated, resulting in epigenetic silencing[9,14] and transcription could be silenced by other mechanisms,such as histone modification. Therefore, reversing the epigeneticmodifications that result in transcriptional silencing of CHD5 [19–23] should restore expression of an intact allele and presumablyrestore growth control. Thus, it will be important to identify thegenes and proteins that regulate CHD5 expression, the genes thatCHD5 regulates and the proteins with which CHD5 interacts, tofully understand the role of CHD5 in normal development as wellas malignant transformation.
AUTHOR CONTRIBUTION
Venkatadri Kolla designed and performed all experiments. Koumudi Naraparaju conductedand assisted Western blot analysis. Tiangang Zhuang, Mayumi Higashi and SriharshaKolla assisted in conducting experiments and preparation of manuscript. Gerd Blobelgave valuable suggestions for the present study and provided the GST–FOG1 reagent forthese analyses. Garrett Brodeur conceived of the initial experimental design, supervisedthe experiments, analysed the data, interpreted results and helped write and edit themanuscript to its final form.
ACKNOWLEDGEMENTS
Authors thank Dr Linda Gonzales from the Division of Neonatology for her expert helpin fluorescence microscopy. We also thank Ross Tomaino at Taplin Biological MassSpectrometry Facility, at Harvard Medical School, Boston, MA for peptide analysis.
FUNDING
This work was supported by the National Institutes of Health [grant number R01-CA39771];the Alex’s Lemonade Stand Foundation; and the Audrey E. Evans Chair in MolecularOncology (to G.M.B).
REFERENCES
1 Brodeur, G.M. (2003) Neuroblastoma: biological insights into a clinical enigma. Nat. Rev.Cancer 3, 203–216 CrossRef PubMed
2 Brodeur, G.M., Hogarty, M.D., Mosse, Y.P. and Maris, J.M. (2011) Neuroblastoma. InPrinciples and Practice of Pediatric Oncology (Pizzo, P.A. and Poplack, D.G., eds),pp. 886–922, Lippincott, Williams and Wilkins, Philadelphia
3 Mosse, Y.P., Greshock, J., Margolin, A., Naylor, T., Cole, K., Khazi, D., Hii, G., Winter, C.,Shahzad, S., Asziz, M.U. et al. (2005) High-resolution detection and mapping of genomicDNA alterations in neuroblastoma. Genes Chromosomes Cancer 43, 390–403CrossRef PubMed
4 Maris, J.M., White, P.S., Beltinger, C.P., Sulman, E.P., Castleberry, R.P., Shuster, J.J.,Look, A.T. and Brodeur, G.M. (1995) Significance of chromosome 1p loss ofheterozygosity in neuroblastoma. Cancer Res. 55, 4664–4669 PubMed
5 Attiyeh, E.F., London, W.B., Mosse, Y.P., Wang, Q., Winter, C., Khazi, D., McGrady, P.W.,Seeger, R.C., Look, A.T., Shimada, H. et al. (2005) Chromosome 1p and 11q deletionsand outcome in neuroblastoma. N. Engl. J. Med. 353, 2243–2253 CrossRef PubMed
6 Fong, C.T., Dracopoli, N.C., White, P.S., Merrill, P.T., Griffith, R.C., Housman, D.E. andBrodeur, G.M. (1989) Loss of heterozygosity for the short arm of chromosome 1 in humanneuroblastomas: correlation with N-myc amplification. Proc. Natl. Acad. Sci. U.S.A. 86,3753–3757 CrossRef PubMed
7 Brodeur, G.M., Sekhon, G. and Goldstein, M.N. (1977) Chromosomal aberrations inhuman neuroblastomas. Cancer 40, 2256–2263 CrossRef PubMed
8 White, P.S., Maris, J.M., Beltinger, C., Sulman, E., Marshall, H.N., Fujimori, M., Kaufman,B.A., Biegel, J.A., Allen, C., Hilliard, C. et al. (1995) A region of consistent deletion inneuroblastoma maps within human chromosome 1p36.2–36.3. Proc. Natl. Acad. Sci.U.S.A. 92, 5520–5524 CrossRef PubMed
9 Fujita, T., Igarashi, J., Okawa, E.R., Gotoh, T., Manne, J., Kolla, V., Kim, J., Zhao, H.,Pawel, B.R., London, W.B. et al. (2008) CHD5, a tumor suppressor gene deleted from1p36.31 in neuroblastomas. J. Natl. Cancer Inst. 100, 940–949 CrossRef PubMed
10 Okawa, E.R., Gotoh, T., Manne, J., Igarashi, J., Fujita, T., Silverman, K.A., Xhao, H.,Mosse, Y.P., White, P.S. and Brodeur, G.M. (2008) Expression and sequence analysis ofcandidates for the 1p36.31 tumor suppressor gene deleted in neuroblastomas. Oncogene27, 803–810 CrossRef PubMed
11 Thompson, P.M., Gotoh, T., Kok, M., White, P.S. and Brodeur, G.M. (2003) CHD5, a newmember of the chromodomain gene family, is preferentially expressed in the nervoussystem. Oncogene 22, 1002–1011 CrossRef PubMed
12 White, P.S., Thompson, P.M., Gotoh, T., Okawa, E.R., Igarashi, J., Kok, M., Winter, C.,Gregory, S.G., Hogarty, M.D., Maris, J.M. and Brodeur, G.M. (2005) Definition andcharacterization of a region of 1p36.3 consistently deleted in neuroblastoma. Oncogene24, 2684–2694 CrossRef PubMed
13 Garcia, I., Mayol, G., Rodriguez, E., Sunol, M., Gershon, T.R., Rios, J., Cheung, N.K.,Kieran, M.W., George, R.E., Perez-Atayde, A.R. et al. (2010) Expression of theneuron-specific protein CHD5 is an independent marker of outcome in neuroblastoma.Mol. Cancer 9, 277 CrossRef PubMed
14 Koyama, H., Zhuang, T., Light, J.E., Kolla, V., Higashi, M., McGrady, P.W., London, W.B.and Brodeur, G.M. (2012) Mechanisms of CHD5 Inactivation in neuroblastomas. Clin.Cancer Res. 18, 1588–1597 CrossRef PubMed
15 Bagchi, A., Papazoglu, C., Wu, Y., Capurso, D., Brodt, M., Francis, D., Bredel, M., Vogel,H. and Mills, A.A. (2007) CHD5 is a tumor suppressor at human 1p36. Cell 128,459–475 CrossRef PubMed
16 Law, M.E., Templeton, K.L., Kitange, G., Smith, J., Misra, A., Feuerstein, B.G. andJenkins, R.B. (2005) Molecular cytogenetic analysis of chromosomes 1 and 19 in gliomacell lines. Cancer Genet. Cytogenet. 160, 1–14 CrossRef PubMed
17 Lang, J., Tobias, E.S. and Mackie, R. (2011) Preliminary evidence for involvement of thetumour suppressor gene CHD5 in a family with cutaneous melanoma. Br. J. Dermatol.164, 1010–1016 CrossRef PubMed
18 Ng, D., Yang, X.R., Tucker, M.A. and Goldstein, A.M. (2008) Mutation screening ofCHD5 in melanoma-prone families linked to 1p36 revealed no deleterious coding orsplice site changes. BMC Res. Notes 1, 86 CrossRef PubMed
19 Wu, X., Zhu, Z., Li, W., Fu, X., Su, D., Fu, L., Zhang, Z., Luo, A., Sun, X., Fu, L. and Dong,J.T. (2012) Chromodomain helicase DNA binding protein 5 plays a tumor suppressor rolein human breast cancer. Breast Cancer Res. 14, R73 CrossRef PubMed
20 Mokarram, P., Kumar, K., Brim, H., Naghibalhossaini, F., Saberi-firoozi, M., Nouraie, M.,Green, R., Lee, E., Smoot, D.T. and Ashktorab, H. (2009) Distinct high-profile methylatedgenes in colorectal cancer. PLoS One 4, e7012 CrossRef PubMed
c© The Authors Journal compilation c© 2015 Biochemical Society

352 V. Kolla and others
21 Cai, C., Ashktorab, H., Pang, X., Zhao, Y., Sha, W., Liu, Y. and Gu, X. (2012)MicroRNA-211 expression promotes colorectal cancer cell growth in vitro and in vivo bytargeting tumor suppressor CHD5. PLoS One 7, e29750 CrossRef PubMed
22 Wang, X., Lau, K.K., So, L.K. and Lam, Y.W. (2009) CHD5 is down-regulated throughpromoter hypermethylation in gastric cancer. J. Biomed. Sci. 16, 95 CrossRef PubMed
23 Zhao, R., Yan, Q., Lv, J., Huang, H., Zheng, W., Zhang, B. and Ma, W. (2012) CHD5, atumor suppressor that is epigenetically silenced in lung cancer. Lung Cancer 76,324–331 CrossRef PubMed
24 Gorringe, K.L., Choong, D.Y., Williams, L.H., Ramakrishna, M., Sridhar, A., Qiu, W.,Bearfoot, J.L. and Campbell, I.G. (2008) Mutation and methylation analysis of thechromodomain-helicase-DNA binding 5 gene in ovarian cancer. Neoplasia 10,1253–1258 PubMed
25 Wong, R.R., Chan, L.K., Tsang, T.P., Lee, C.W., Cheung, T.H., Yim, S.F., Siu, N.S., Lee,S.N., Yu, M.Y., Chim, S.S. et al. (2011) chd5 downregulation associated with poorprognosis in epithelial ovarian cancer. Gynecol. Obstet. Invest. 72, 203–207CrossRef PubMed
26 Robbins, C.M., Tembe, W.A., Baker, A., Sinari, S., Moses, T.Y., Beckstrom-Sternberg, S.,Beckstrom-Sternberg, J., Barrett, M., Long, J., Chinnaiyan, A. et al. (2011) Copy numberand targeted mutational analysis reveals novel somatic events in metastatic prostatetumors. Genome Res. 21, 47–55 CrossRef PubMed
27 Kolla, V., Zhuang, T., Higashi, M., Naraparaju, K. and Brodeur, G.M. (2014) Role ofCHD5 in human cancers: 10 years later. Cancer Res. 74, 652–658 CrossRef PubMed
28 Lai, A.Y. and Wade, P.A. (2011) Cancer biology and NuRD: a multifaceted chromatinremodelling complex. Nat. Rev. Cancer 11, 588–596 CrossRef PubMed
29 Marfella, C.G. and Imbalzano, A.N. (2007) The Chd family of chromatin remodelers.Mutat. Res. 618, 30–40 CrossRef PubMed
30 Hall, J.A. and Georgel, P.T. (2007) CHD proteins: a diverse family with strong ties.Biochem. Cell Biol. 85, 463–476 CrossRef PubMed
31 Ramirez, J., Dege, C., Kutateladze, T.G. and Hagman, J. (2012) MBD2 and multipledomains of CHD4 are required for transcriptional repression by Mi-2/NuRD complexes.Mol. Cell Biol. 32, 5078–5088 CrossRef PubMed
32 Woodage, T., Basrai, M.A., Baxevanis, A.D., Hieter, P. and Collins, F.S. (1997)Characterization of the CHD family of proteins. Proc. Natl. Acad. Sci. U.S.A. 94,11472–11477 CrossRef PubMed
33 Zhuang, T., Hess, R.A., Kolla, V., Higashi, M., Raabe, T.D. and Brodeur, G.M. (2014)CHD5 is required for spermiogenesis and chromatin condensation. Mech. Dev. 131,35–46 CrossRef PubMed
34 Tong, J.K., Hassig, C.A., Schnitzler, G.R., Kingston, R.E. and Schreiber, S.L. (1998)Chromatin deacetylation by an ATP-dependent nucleosome remodelling complex. Nature395, 917–921 CrossRef PubMed
35 Xue, Y., Wong, J., Moreno, G.T., Young, M.K., Cote, J. and Wang, W. (1998) NURD, anovel complex with both ATP-dependent chromatin-remodeling and histone deacetylaseactivities. Mol. Cell. 2, 851–861 CrossRef PubMed
36 Hargreaves, D.C. and Crabtree, G.R. (2011) ATP-dependent chromatin remodeling:genetics, genomics and mechanisms. Cell Res. 21, 396–420 CrossRef PubMed
37 Denslow, S.A. and Wade, P.A. (2007) The human Mi-2/NuRD complex and generegulation. Oncogene 26, 5433–5438 CrossRef PubMed
38 Allen, H.F., Wade, P.A. and Kutateladze, T.G. (2013) The NuRD architecture. Cell. Mol. LifeSci. 70, 3513–3524 CrossRef PubMed
39 Wang, L., He, S., Tu, Y., Ji, P., Zong, J., Zhang, J., Feng, F., Zhao, J., Gao, G. and Zhang, Y.(2013) Downregulation of chromatin remodeling factor CHD5 is associated with a poorprognosis in human glioma. J. Clin. Neurosci. 20, 958–963 CrossRef PubMed
40 Stanley, F.K., Moore, S. and Goodarzi, A.A. (2013) CHD chromatin remodelling enzymesand the DNA damage response. Mutat. Res. 750, 31–44CrossRef PubMed
41 Li, D.Q. and Kumar, R. (2010) Mi-2/NuRD complex making inroads into DNA-damageresponse pathway. Cell Cycle 9, 2071–2079CrossRef PubMed
42 Egan, C.M., Nyman, U., Skotte, J., Streubel, G., Turner, S., O’Connell, D.J., Rraklli, V.,Dolan, M.J., Chadderton, N., Hansen, K. et al. (2013) CHD5 is required for neurogenesisand has a dual role in facilitating gene expression and polycomb gene repression. Dev.Cell 26, 223–236 CrossRef PubMed
43 Potts, R.C., Zhang, P., Wurster, A.L., Precht, P., Mughal, M.R., Wood, III, W.H., Zhang, Y.,Becker, K.G., Mattson, M.P. and Pazin, M.J. (2011) CHD5, a brain-specific paralog of Mi2chromatin remodeling enzymes, regulates expression of neuronal genes. PLoS One 6,e24515 CrossRef PubMed
44 Hong, W., Nakazawa, M., Chen, Y.Y., Kori, R., Vakoc, C.R., Rakowski, C. and Blobel, G.A.(2005) FOG-1 recruits the NuRD repressor complex to mediate transcriptional repressionby GATA-1. EMBO J. 24, 2367–2378 CrossRef PubMed
45 Laemmli, U.K. (1970) Cleavage of structural proteins during the assembly of the head ofbacteriophage T4. Nature 227, 680–685 CrossRef PubMed
46 Shevchenko, A., Wilm, M., Vorm, O. and Mann, M. (1996) Mass spectrometricsequencing of proteins silver-stained polyacrylamide gels. Anal. Chem. 68, 850–858CrossRef PubMed
47 Peng, J. and Gygi, S.P. (2001) Proteomics: the move to mixtures. J. Mass Spectrom. 36,1083–1091 CrossRef PubMed
48 Eng, J.K., McCormack, A.L. and Yates, J.R. (1994) An approach to correlate tandem massspectral data of peptides with amino acid sequences in a protein database. J. Am. Soc.Mass Spectrom. 5, 976–989 CrossRef PubMed
49 Miccio, A., Wang, Y., Hong, W., Gregory, G.D., Wang, H., Yu, X., Choi, J.K., Shelat, S.,Tong, W., Poncz, M. and Blobel, G.A. (2010) NuRD mediates activating and repressivefunctions of GATA-1 and FOG-1 during blood development. EMBO J. 29, 442–456CrossRef PubMed
50 Vestin, A. and Mills, A.A. (2013) The tumor suppressor Chd5 is induced during neuronaldifferentiation in the developing mouse brain. Gene Expr. Patterns 13, 482–489CrossRef PubMed
51 Li, W., Wu, J., Kim, S.Y., Zhao, M., Hearn, S.A., Zhang, M.Q., Meistrich, M.L. and Mills,A.A. (2014) Chd5 orchestrates chromatin remodelling during sperm development. Nat.Commun. 5, 3812 PubMed
52 Steilmann, C., Cavalcanti, M.C., Bergmann, M., Kliesch, S., Weidner, W. and Steger, K.(2010) Aberrant mRNA expression of chromatin remodelling factors in round spermatidmaturation arrest compared with normal human spermatogenesis. Mol. Hum. Reprod. 16,726–733 CrossRef PubMed
53 Paul, S., Kuo, A., Schalch, T., Vogel, H., Joshua-Tor, L., McCombie, W.R., Gozani, O.,Hammell, M. and Mills, A.A. (2013) Chd5 requires PHD-mediated histone 3 binding fortumor suppression. Cell Rep. 3, 92–102 CrossRef PubMed
54 Mulero-Navarro, S. and Esteller, M. (2008) Chromatin remodeling factor CHD5 issilenced by promoter CpG island hypermethylation in human cancer. Epigenetics 3,210–215 CrossRef PubMed
55 Qu, Y., Dang, S. and Hou, P. (2013) Gene methylation in gastric cancer. Clin. Chim. Acta424, 53–65 CrossRef PubMed
56 Wang, J., Chen, H., Fu, S., Xu, Z.M., Sun, K.L. and Fu, W.N. (2011) The involvement ofCHD5 hypermethylation in laryngeal squamous cell carcinoma. Oral. Oncol. 47, 601–608CrossRef PubMed
Received 8 January 2015/30 March 2015; accepted 31 March 2015Published as BJ Immediate Publication 31 March 2015, doi:10.1042/BJ20150030
c© The Authors Journal compilation c© 2015 Biochemical Society

Supplementary Table S1: Shown here is the schematic diagram of amino acid alignment map for CHD3, CHD4 and CHD5 proteins indicating the sequence homology. Consensus sequence is shown in the bottom lane and the color description is as follows based on the nature of the R-group of amino acid. Yellow Sulphor M, C (Met, Cys) B/W Aliphatic G, A, V, L, I (Gly, Ala, Val, Leu, Ile) Pink Hyroxyl S, T (Ser, Thr) Green Circular P (Pro) Dark Blue Basic K, R, H (Lys, Arg, His) Red Acidic E, G (Asp, Glu) Orange Benzene F, W, Y (Phe, Trp, Tyr) Light Blue Amide N, Q (Asn, Gln)

hs CHD3,CHD4 and CHD5 aa alignmentTue, Feb 3, 2015 4:00 PMPage 1
Page 1
ABCDEFGHIJKLMNOPQRSTUVWXYZ abcdefghijklmnopqrstuvwxyz.0123456789hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
202040
4060 6080
80100
100120 120140
140160
160180 180200
200220
220240 240
M A S P L R D E E E E E E E M V V S E E E E E E E E E G D E
E E E E E V E A A D E D D E E D D D E G V L G R G P G H D R
G R D R H S P - P G C H L F P P P P P P P P P L P P P P P P
P P P D K D D I R - L L P S A L G V K K R K R G P - K K Q K
E N K P G K P R K R K K R D S E E E F G S E R D E Y R E K S
E S G G S E Y G T G P G R K R R R K H R E K K E K K T K R R
K K G E G D G G Q K Q V E Q K S S A T L L L T W G L E D V E
H V F S E E D Y H T L T N Y K A F S Q F M R P L I A K K N P
M A S G L G - - - S P S P C S A G S E E E D M D A L L N N S
L P P P H P E N E E D P E E D L S E T E T P K L K K K K K P
K K P R D P K I P K S K R Q K K E R M L L C R Q L G D S S G
E G P E F V E E E E E V A L R S D S E G S D Y T P G K K K K
K K L G P K K E K K S K S K R K E E E E E E D D D D D S K E
P - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - K S S A Q L L E D W G M E D I D
H V F S E E D Y R T L T N Y K A F S Q F V R P L I A A K N P
M R G P V G - - - - T E E E L P R L F A E E M E N E D E M S
E E E D G G L E A F D D F F P V E P V S L P K K K K P K K L
K E N K C K G K R K K K E G S N D E L S - - - - - - - - - -
- - - - - - E N E E D L E E K S E S E G S D Y S P N K K K K
K K L K D K K E K K A K R K K K D E D E D D N D D G C L K E
P - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - K S S G Q L M A E W G L D D V D
Y L F S E E D Y H T L T N Y K A F S Q F L R P L I A K K N P
M A S P L G D E E E E E . . S E E E E M E E . S
E E E . . E A . D D . E . . . . . P K . K K K K
K . R P K K .
P . E E E . L . S . S E G S D Y . P K K K K
K K L K K E K K K R K . K E E . E . E D D K E
P S G G S E Y G T G P G R K R R R K H R E K K E K K T K R R
K K G E G D G G Q K Q V E Q K S S A Q L L . . W G L E D V D
H V F S E E D Y H T L T N Y K A F S Q F . R P L I A K K N P
K

hs CHD3,CHD4 and CHD5 aa alignmentTue, Feb 3, 2015 4:00 PMPage 2
Page 2
ABCDEFGHIJKLMNOPQRSTUVWXYZ abcdefghijklmnopqrstuvwxyz.0123456789hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
180
180200
200220
220240 240260
260280
280300 300320
320340
340360 360380
380400
400420 420
K K G E G D G G Q K Q V E Q K S S A T L L L T W G L E D V E
H V F S E E D Y H T L T N Y K A F S Q F M R P L I A K K N P
K I P M S K M M T I L G A K W R E F S A N N P F K G S A A A
V A A A A A A A A A A V A E Q V S A A V S S A T P I A P S G
P P A L P P P P A A D I Q P P P I R R A K T K E G K G P G H
K R R S K S P R V P D G R K K L R G K K M A P L K I K L G L
L G G K R K K G G S Y V F Q S D E G P E P E A E E S D L D S
G S V H S A S G R P D G - - P V R T K K L K R G R P G R K K
- - - - - - - - - - - - - - K S S A Q L L E D W G M E D I D
H V F S E E D Y R T L T N Y K A F S Q F V R P L I A A K N P
K I A V S K M M M V L G A K W R E F S T N N P F K G S S G A
S V A A A A A A A V A V V E S M V T A T E V A P P - - - - -
- - - - P P P V - - - - - E V P I R K A K T K E G K G P N A
R R K P K G S P R V P D A K K P K P K K V A P L K I K L G G
F G S K R K R S S S E D D D L D V - - - - - - - E S D F D D
A S I N S Y S - V S D G S T S R S S R S R K K L R T T K K K
- - - - - - - - - - - - - - K S S G Q L M A E W G L D D V D
Y L F S E E D Y H T L T N Y K A F S Q F L R P L I A K K N P
K I P M S K M M T V L G A K W R E F S A N N P F K G S S A A
A A A A A V A A A V E T V T I S P P L - A V S P P - - - - -
- - - - - - - - - Q V P Q P V P I R K A K T K E G K G P G V
R K K I K G S K - - D G K K K G K G K K T A G L K F R F G G
I S N K R K K G S S S E E D E R - - - - - - - E E S D F D S
A S I H S A S V R S E C S - - - - A A L G K K S K R R R K K
K K G E G D G G Q K Q V E Q K S S A Q L L . . W G L E D V D
H V F S E E D Y H T L T N Y K A F S Q F . R P L I A K K N P
K I P M S K M M T V L G A K W R E F S A N N P F K G S S A A
. A A A A A A A A V A V V E A V A P P I A P S G
P P A L P P P A Q P V P I R K A K T K E G K G P G .
R R K K G S . D G . K K . K G K K A P L K I K L G G
. G K R K K G S S . . D D G P E P E A E E S D F D S
A S I H S A S . R S D G S T . . . K K . R R K K
K

hs CHD3,CHD4 and CHD5 aa alignmentTue, Feb 3, 2015 4:00 PMPage 3
Page 3
ABCDEFGHIJKLMNOPQRSTUVWXYZ abcdefghijklmnopqrstuvwxyz.0123456789hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
360
360380
380400
400420 420440
440460
460480 480500
500520
520540 540560
560580
580600 600
L G G K R K K G G S Y V F Q S D E G P E P E A E E S D L D S
G S V H S A S G R P D G - - P V R T K K L K R G R P G R K K
K K V L G C P A V A G E E E V D G Y E T D H Q D Y C E V C Q
Q G G E I I L C D T C P R A Y H L V C L D P E L D R A P E G
K W S C P H C E K E G V Q W E A K E E E E E Y E E E G E E E
G E K - E E E D D - H M E Y C R V C K D G G E L L C C D A C
I S S Y H I H C L N P P L P D I P N G E W L C P R C T C P V
L K G R V Q K I L H W R W G E P P V A V P A P - Q Q A D G N
F G S K R K R S S S E D D D L D V - - - - - - - E S D F D D
A S I N S Y S - V S D G S T S R S S R S R K K L R T T K K K
K K G - - - - - E E E V T A V D G Y E T D H Q D Y C E V C Q
Q G G E I I L C D T C P R A Y H M V C L D P D M E K A P E G
K W S C P H C E K E G I Q W E A K E D N S E G E E I L E E V
G G D L E E E D D H H M E F C R V C K D G G E L L C C D T C
P S S Y H I H C L N P P L P E I P N G E W L C P R C T C P A
L K G K V Q K I L I W K W G Q P P S P T P V P - R P P D A D
I S N K R K K G S S S E E D E R - - - - - - - E E S D F D S
A S I H S A S V R S E C S - - - - A A L G K K S K R R R K K
K R I - - - - - - - - - D D G D G Y E T D H Q D Y C E V C Q
Q G G E I I L C D T C P R A Y H L V C L D P E L E K A P E G
K W S C P H C E K E G I Q W E P K D D D D E E E E - - - - -
G G C E E E E D D - H M E F C R V C K D G G E L L C C D A C
P S S Y H L H C L N P P L P E I P N G E W L C P R C T C P P
L K G K V Q R I L H W R W T E P P A P F M V G L P G P D V E
. G K R K K G S S . . D D G P E P E A E E S D F D S
A S I H S A S . R S D G S T . . . K K . R R K K
K K . L G C P A . . V D G Y E T D H Q D Y C E V C Q
Q G G E I I L C D T C P R A Y H L V C L D P E L E K A P E G
K W S C P H C E K E G I Q W E A K E D . . E E E . E E
G G E E E D D H H M E F C R V C K D G G E L L C C D A C
P S S Y H I H C L N P P L P E I P N G E W L C P R C T C P .
L K G K V Q K I L H W R W G E P P . P P V P L P D . .
P

hs CHD3,CHD4 and CHD5 aa alignmentTue, Feb 3, 2015 4:00 PMPage 4
Page 4
ABCDEFGHIJKLMNOPQRSTUVWXYZ abcdefghijklmnopqrstuvwxyz.0123456789hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
540
540560
560580
580600 600620
620640
640660 660680
680700
700720 720740
740760
760780 780
I S S Y H I H C L N P P L P D I P N G E W L C P R C T C P V
L K G R V Q K I L H W R W G E P P V A V P A P - Q Q A D G N
P D V P P P R P L Q G R S E R E F F V K W V G L S Y W H C S
W A K E L Q L E I F H L V M Y R N Y Q R K N D M D E P P P L
D Y G S G E D D G K S D K R K V K D P H Y A E M E E K Y Y R
F G I K P E W M T V H R I I N H S V D K K G N Y H Y L V K W
R D L P Y D Q S T W E E D E M N I P E Y E E - H K Q S Y W R
H R E L I M G E D P A Q P R K Y K K K K K E L Q - - - G D G
P S S Y H I H C L N P P L P E I P N G E W L C P R C T C P A
L K G K V Q K I L I W K W G Q P P S P T P V P - R P P D A D
P N T P S P K P L E G R P E R Q F F V K W Q G M S Y W H C S
W V S E L Q L E L H C Q V M F R N Y Q R K N D M D E P P S G
D F G - G - D E E K S R K R K N K D P K F A E M E E R F Y R
Y G I K P E W M M I H R I L N H S V D K K G H V H Y L I K W
R D L P Y D Q A S W E S E D V E I Q D Y D L F - K Q S Y W N
H R E L - M R G E E G R P - - - G K K L K K V K L R K L E R
P S S Y H L H C L N P P L P E I P N G E W L C P R C T C P P
L K G K V Q R I L H W R W T E P P A P F M V G L P G P D V E
P S L P P P K P L E G I P E R E F F V K W A G L S Y W H C S
W V K E L Q L E L Y H T V M Y R N Y Q R K N D M D E P P P F
D Y G S G D E D G K S E K R K N K D P L Y A K M E E R F Y R
Y G I K P E W M M I H R I L N H S F D K K G D V H Y L I K W
K D L P Y D Q C T W E I D D I D I P Y Y D N L K Q A Y W - G
H R E L M L G E D T R L P K R L L K K G K K L R D D K Q E K
P S S Y H I H C L N P P L P E I P N G E W L C P R C T C P .
L K G K V Q K I L H W R W G E P P . P P V P L P D . .
P . P P P K P L E G R P E R E F F V K W . G L S Y W H C S
W V K E L Q L E L . H V M Y R N Y Q R K N D M D E P P P .
D Y G S G . D D G K S . K R K N K D P . Y A E M E E R F Y R
Y G I K P E W M M I H R I L N H S V D K K G V H Y L I K W
R D L P Y D Q T W E D D . . I P . Y D . K Q S Y W
H R E L M G E D . P . . . K K . K K L . K . E .
P

hs CHD3,CHD4 and CHD5 aa alignmentTue, Feb 3, 2015 4:00 PMPage 5
Page 5
ABCDEFGHIJKLMNOPQRSTUVWXYZ abcdefghijklmnopqrstuvwxyz.0123456789hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
720
720740
740760
760780 780800
800820
820840 840860
860880
880900 900920
920940
940960 960
R D L P Y D Q S T W E E D E M N I P E Y E E - H K Q S Y W R
H R E L I M G E D P A Q P R K Y K K K K K E L Q - - - G D G
P P S S P T N D P T V K Y E T Q P R F I T A T G G T L H M Y
Q L E G L N W L R F S W A Q G T D T I L A D E M G L G K T I
Q T I V F L Y S L Y K E G H T K G P F L V S A P L S T I I N
W E R E F Q M W A P K F Y V V T Y T G D K D S R A I I R E N
E F S F E D N A I K G G K K A F K M K R E A Q V K F H V L L
T S Y E L I T I D Q A A L G S I R W A C L V V D E A H R L K
R D L P Y D Q A S W E S E D V E I Q D Y D L F - K Q S Y W N
H R E L - M R G E E G R P - - - G K K L K K V K L R K L E R
P P E T P T V D P T V K Y E R Q P E Y L D A T G G T L H P Y
Q M E G L N W L R F S W A Q G T D T I L A D E M G L G K T V
Q T A V F L Y S L Y K E G H S K G P F L V S A P L S T I I N
W E R E F E M W A P D M Y V V T Y V G D K D S R A I I R E N
E F S F E D N A I R G G K K A S R M K K E A S V K F H V L L
T S Y E L I T I D M A I L G S I D W A C L I V D E A H R L K
K D L P Y D Q C T W E I D D I D I P Y Y D N L K Q A Y W - G
H R E L M L G E D T R L P K R L L K K G K K L R D D K Q E K
P P D T P I V D P T V K F D K Q P W Y I D S T G G T L H P Y
Q L E G L N W L R F S W A Q G T D T I L A D E M G L G K T V
Q T I V F L Y S L Y K E G H S K G P Y L V S A P L S T I I N
W E R E F E M W A P D F Y V V T Y T G D K E S R S V I R E N
E F S F E D N A I R S G K K V F R M K K E V Q I K F H V L L
T S Y E L I T I D Q A I L G S I E W A C L V V D E A H R L K
R D L P Y D Q T W E D D . . I P . Y D . K Q S Y W
H R E L M G E D . P . . . K K . K K L . K . E .
P P . T P T V D P T V K Y E . Q P Y I D A T G G T L H P Y
Q L E G L N W L R F S W A Q G T D T I L A D E M G L G K T V
Q T I V F L Y S L Y K E G H S K G P F L V S A P L S T I I N
W E R E F E M W A P D F Y V V T Y T G D K D S R A I I R E N
E F S F E D N A I R G G K K A F R M K K E A Q V K F H V L L
T S Y E L I T I D Q A I L G S I . W A C L V V D E A H R L K
N
hs CHD3,CHD4 and CHD5 aa alignmentTue, Feb 3, 2015 4:00 PMPage 6
Page 6
ABCDEFGHIJKLMNOPQRSTUVWXYZ abcdefghijklmnopqrstuvwxyz.0123456789hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
900
900920
920940
940960 960980
9801000
10001020 10201040
10401060
10601080 10801100
11001120
11201140 1140
E F S F E D N A I K G G K K A F K M K R E A Q V K F H V L L
T S Y E L I T I D Q A A L G S I R W A C L V V D E A H R L K
N N Q S K F F R V L N G Y K I D H K L L L T G T P L Q N N L
E E L F H L L N F L T P E R F N N L E G F L E E F A D I S K
E D Q I K K L H D L L G P H M L R R L K A D V F K N M P A K
T E L I V R V E L S P M Q K K Y Y K Y I L T R N F E A L N S
R G G G N Q V S L L N I M M D L K K C C N H P Y L F P V A A
M E S P K L P S G A Y E G G A L I K S S G K L M L L Q K M L
E F S F E D N A I R G G K K A S R M K K E A S V K F H V L L
T S Y E L I T I D M A I L G S I D W A C L I V D E A H R L K
N N Q S K F F R V L N G Y S L Q H K L L L T G T P L Q N N L
E E L F H L L N F L T P E R F H N L E G F L E E F A D I A K
E D Q I K K L H D M L G P H M L R R L K A D V F K N M P S K
T E L I V R V E L S P M Q K K Y Y K Y I L T R N F E A L N A
R G G G N Q V S L L N V V M D L K K C C N H P Y L F P V A A
M E A P K M P N G M Y D G S A L I R A S G K L L L L Q K M L
E F S F E D N A I R S G K K V F R M K K E V Q I K F H V L L
T S Y E L I T I D Q A I L G S I E W A C L V V D E A H R L K
N N Q S K F F R V L N S Y K I D Y K L L L T G T P L Q N N L
E E L F H L L N F L T P E R F N N L E G F L E E F A D I S K
E D Q I K K L H D L L G P H M L R R L K A D V F K N M P A K
T E L I V R V E L S Q M Q K K Y Y K F I L T R N F E A L N S
K G G G N Q V S L L N I M M D L K K C C N H P Y L F P V A A
V E A P V L P N G S Y D G S S L V K S S G K L M L L Q K M L
E F S F E D N A I R G G K K A F R M K K E A Q V K F H V L L
T S Y E L I T I D Q A I L G S I . W A C L V V D E A H R L K
N N Q S K F F R V L N G Y K I D H K L L L T G T P L Q N N L
E E L F H L L N F L T P E R F N N L E G F L E E F A D I S K
E D Q I K K L H D L L G P H M L R R L K A D V F K N M P A K
T E L I V R V E L S P M Q K K Y Y K Y I L T R N F E A L N S
R G G G N Q V S L L N I M M D L K K C C N H P Y L F P V A A
M E A P K L P N G Y D G S A L I K S S G K L M L L Q K M L
K

hs CHD3,CHD4 and CHD5 aa alignmentTue, Feb 3, 2015 4:00 PMPage 7
Page 7
ABCDEFGHIJKLMNOPQRSTUVWXYZ abcdefghijklmnopqrstuvwxyz.0123456789hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
1080
10801100
11001120
11201140 11401160
11601180
11801200 12001220
12201240
12401260 12601280
12801300
13001320 1320
R G G G N Q V S L L N I M M D L K K C C N H P Y L F P V A A
M E S P K L P S G A Y E G G A L I K S S G K L M L L Q K M L
R K L K E Q G H R V L I F S Q M T K M L D L L E D F L D Y E
G Y K Y E R I D G G I T G A L R Q E A I D R F N A P G A Q Q
F C F L L S T R A G G L G I N L A T A D T V I I F D S D W N
P H N D I Q A F S R A H R I G Q A N K V M I Y R F V T R A S
V E E R I T Q V A K R K M M L T H L V V R P G L G S K A G S
M S K Q E L D D I L K F G T E E L F K D E N E - - - - - - -
R G G G N Q V S L L N V V M D L K K C C N H P Y L F P V A A
M E A P K M P N G M Y D G S A L I R A S G K L L L L Q K M L
K N L K E G G H R V L I F S Q M T K M L D L L E D F L E H E
G Y K Y E R I D G G I T G N M R Q E A I D R F N A P G A Q Q
F C F L L S T R A G G L G I N L A T A D T V I I Y D S D W N
P H N D I Q A F S R A H R I G Q N K K V M I Y R F V T R A S
V E E R I T Q V A K K K M M L T H L V V R P G L G S K T G S
M S K Q E L D D I L K F G T E E L F K D E A T D - - - - - -
K G G G N Q V S L L N I M M D L K K C C N H P Y L F P V A A
V E A P V L P N G S Y D G S S L V K S S G K L M L L Q K M L
K K L R D E G H R V L I F S Q M T K M L D L L E D F L E Y E
G Y K Y E R I D G G I T G G L R Q E A I D R F N A P G A Q Q
F C F L L S T R A G G L G I N L A T A D T V I I Y D S D W N
P H N D I Q A F S R A H R I G Q N K K V M I Y R F V T R A S
V E E R I T Q V A K R K M M L T H L V V R P G L G S K S G S
M T K Q E L D D I L K F G T E E L F K D D V E G M M S Q G Q
R G G G N Q V S L L N I M M D L K K C C N H P Y L F P V A A
M E A P K L P N G Y D G S A L I K S S G K L M L L Q K M L
K K L K E G H R V L I F S Q M T K M L D L L E D F L E Y E
G Y K Y E R I D G G I T G . L R Q E A I D R F N A P G A Q Q
F C F L L S T R A G G L G I N L A T A D T V I I Y D S D W N
P H N D I Q A F S R A H R I G Q N K K V M I Y R F V T R A S
V E E R I T Q V A K R K M M L T H L V V R P G L G S K . G S
M S K Q E L D D I L K F G T E E L F K D E . E M M S Q G Q
R

hs CHD3,CHD4 and CHD5 aa alignmentTue, Feb 3, 2015 4:00 PMPage 8
Page 8
ABCDEFGHIJKLMNOPQRSTUVWXYZ abcdefghijklmnopqrstuvwxyz.0123456789hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
1260
12601280
12801300
13001320 13201340
13401360
13601380 13801400
14001420
14201440 14401460
14601480
14801500 1500
V E E R I T Q V A K R K M M L T H L V V R P G L G S K A G S
M S K Q E L D D I L K F G T E E L F K D E N E - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
G E N K E - E D S S V I H Y D N E A I A R L L D R N Q D A T
E D T D V Q N M N E Y L S S F K V A Q Y V V R E E D K I E E
I E R E - - I I K Q E E N V D P D Y W E K L L R H H Y E Q Q
Q E D L A R N L G K G K R V R K Q V N Y N D A A Q E D Q D N
Q S E Y S - - - - - - - V G S E E E D E D F D E R P E - G R
V E E R I T Q V A K K K M M L T H L V V R P G L G S K T G S
M S K Q E L D D I L K F G T E E L F K D E A T D - - - - - -
- - - - - - - - - - - - - G G - - - - - - - - - - - - - - -
G D N K E G E D S S V I H Y D D K A I E R L L D R N Q D E T
E D T E L Q G M N E Y L S S F K V A Q Y V V R E E E M G E E
E E V E R E I I K Q E E S V D P D Y W E K L L R H H Y E Q Q
Q E D L A R N L G K G K R I R K Q V N Y N D G S Q E D R D W
Q D D Q S D N Q S D Y S V A S E E G D E D F D E R S E A P R
V E E R I T Q V A K R K M M L T H L V V R P G L G S K S G S
M T K Q E L D D I L K F G T E E L F K D D V E G M M S Q G Q
R P V T P I P D V Q S S K G G N L A A S A K K K H G S T P P
G D N K D V E D S S V I H Y D D A A I S K L L D R N Q D A T
D D T E L Q N M N E Y L S S F K V A Q Y V V R E E D - V E E
V - R E - - I I K Q E E N V D P D Y W E K L L R H H Y E Q Q
Q E D L A R N L G K G K R I R K Q V N Y N D A S Q E D Q E W
Q D E L S D N Q S E Y S I G S E D E D E D F E E R P E G Q S
V E E R I T Q V A K R K M M L T H L V V R P G L G S K . G S
M S K Q E L D D I L K F G T E E L F K D E . E M M S Q G Q
R P V T P I P D V Q S S K G G N L A A S A K K K H G S T P P
G D N K E . E D S S V I H Y D D A I R L L D R N Q D A T
E D T E L Q N M N E Y L S S F K V A Q Y V V R E E D . E E
. E R E R E I I K Q E E N V D P D Y W E K L L R H H Y E Q Q
Q E D L A R N L G K G K R I R K Q V N Y N D A S Q E D Q D W
Q D E S D N Q S . Y S V G S E E E D E D F D E R P E . R
R

hs CHD3,CHD4 and CHD5 aa alignmentTue, Feb 3, 2015 4:00 PMPage 9
Page 9
ABCDEFGHIJKLMNOPQRSTUVWXYZ abcdefghijklmnopqrstuvwxyz.0123456789hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
1440
14401460
14601480
14801500 15001520
15201540
15401560 15601580
15801600
16001620 16201640
16401660
16601680 1680
Q E D L A R N L G K G K R V R K Q V N Y N D A A Q E D Q D N
Q S E Y S - - - - - - - V G S E E E D E D F D E R P E - G R
R Q - - S K R Q L R N E K D K P L P P L L A R V G G N I E V
L G F N T R Q R K A F L N A V M R W G M P P Q D A F T T Q W
L V R D L R G K T E K E F K A Y V S L F M R H L C E P G A D
G S E T F A D G V P R E G L S R Q Q V L T R I G V M S L V K
K K V Q E F E H I N G R W S M P E L M P D P S A D S K R S S
R A S S P T K T S P T T P E A S A T N S P C T S K P A T P A
Q E D L A R N L G K G K R I R K Q V N Y N D G S Q E D R D W
Q D D Q S D N Q S D Y S V A S E E G D E D F D E R S E A P R
R P - - S R K G L R N D K D K P L P P L L A R V G G N I E V
L G F N A R Q R K A F L N A I M R Y G M P P Q D A F T T Q W
L V R D L R G K S E K E F K A Y V S L F M R H L C E P G A D
G A E T F A D G V P R E G L S R Q H V L T R I G V M S L I R
K K V Q E F E H V N G R W S M P E A E V E - - - E N K K M S
Q P G S P S P K T P T P S T P G D T Q P N T P A - P V P P A
Q E D L A R N L G K G K R I R K Q V N Y N D A S Q E D Q E W
Q D E L S D N Q S E Y S I G S E D E D E D F E E R P E G Q S
G R R Q S R R Q L K S D R D K P L P P L L A R V G G N I E V
L G F N A R Q R K A F L N A I M R W G M P P Q D A F N S H W
L V R D L R G K S E K E F R A Y V S L F M R H L C E P G A D
G A E T F A D G V P R E G L S R Q H V L T R I G V M S L V R
K K V Q E F E H V N G K Y S T P D L I P E G P - E G K K S G
E V I S S D P N T P V P A S P A H L L P A P L G L P D K M E
Q E D L A R N L G K G K R I R K Q V N Y N D A S Q E D Q D W
Q D E S D N Q S . Y S V G S E E E D E D F D E R P E . R
R R Q S R R Q L R N D K D K P L P P L L A R V G G N I E V
L G F N A R Q R K A F L N A I M R W G M P P Q D A F T T Q W
L V R D L R G K S E K E F K A Y V S L F M R H L C E P G A D
G A E T F A D G V P R E G L S R Q H V L T R I G V M S L V R
K K V Q E F E H V N G R W S M P E L P E A E K K S S
. . S P . P T P T P . P . T . P . P . P A

hs CHD3,CHD4 and CHD5 aa alignmentTue, Feb 3, 2015 4:00 PMPage 10
Page 10
ABCDEFGHIJKLMNOPQRSTUVWXYZ abcdefghijklmnopqrstuvwxyz.0123456789hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
1620
16201640
16401660
16601680 16801700
17001720
17201740 17401760
17601780
17801800 18001820
18201840
18401860 1860
K K V Q E F E H I N G R W S M P E L M P D P S A D S K R S S
R A S S P T K T S P T T P E A S A T N S P C T S K P A T P A
P S E K G E G I R T P L E K E E A E N Q E E K P E K N S R I
G E K M E T E A D A P S P A P S L G E R L E P R K I P L E D
E V P G V P G E M E P E P G Y R G D R E K S A T E S T P G E
R G E E K P L D G Q E H R E R P E G E T G D L G K R E D V K
G D R E L R P G P R D E P R S N G R R E E K T E K P R F M F
N I A D G G F T E L H T L W Q N E E R A A I S S G K L N E I
K K V Q E F E H V N G R W S M P E A E V E - - - E N K K M S
Q P G S P S P K T P T P S T P G D T Q P N T P A - P V P P A
E D G I K I E E N S L - - K E E E S I E G E K E V K - S T A
P E T A I E C T Q A P A P A S E D E K V V V E P P E G E E K
V E K A E V K E R T E E - P M E T E P K G A A D V - - - - E
K V E E K S A I D L T P I V - V E D K E E K K E E E E K K E
V M L Q - N G E T P K D L N D E - - K Q K K N I K Q R F M F
N I A D G G F T E L H S L W Q N E E R A A T V T K K T Y E I
K K V Q E F E H V N G K Y S T P D L I P E G P - E G K K S G
E V I S S D P N T P V P A S P A H L L P A P L G L P D K M E
A Q L G Y M D E K D P G A Q K P R Q P L E V - - Q A L P A A
L D R V E - - - - - - - - - S E D K H - - - E S P A S K E R
A R E E R P E E T E K A P P S P E Q L P R E E V L P E K E K
I L D K L E L S L I H S R G D S S E L R P D D T K A E E K E
P I E T Q Q N G D K E E D D E G K K E D K K - G K F K F M F
N I A D G G F T E L H T L W Q N E E R A A V S S G K I Y D I
K K V Q E F E H V N G R W S M P E L P E A E K K S S
. . S P . P T P T P . P . T . P . P . P A
. . . E . . P . K E E E E K K S A
. E . . E A P P A S E D . . E P . E .
. . P E E E P P . . A . E
. . E E K L . . R . E . D K E . K E
. . G . . E . . . . K K . K R F M F
N I A D G G F T E L H T L W Q N E E R A A . S S G K . Y E I
W

hs CHD3,CHD4 and CHD5 aa alignmentTue, Feb 3, 2015 4:00 PMPage 11
Page 11
ABCDEFGHIJKLMNOPQRSTUVWXYZ abcdefghijklmnopqrstuvwxyz.0123456789hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
1800
18001820
18201840
18401860 18601880
18801900
19001920 19201940
19401960
19601980 19802000
20002020
20202040 2040
G D R E L R P G P R D E P R S N G R R E E K T E K P R F M F
N I A D G G F T E L H T L W Q N E E R A A I S S G K L N E I
W H R R H D Y W L L A G I V L H G Y A R W Q D I Q N D A Q F
A I I N E P F K T E A N K G N F L E M K N K F L A R R F K L
L E Q A L V I E E Q L R R A A Y L N L S Q E P A H P A M A L
H A R F A E A E C L A E S H Q H L S K E S L A G N K P A N A
V L H K V L N Q L E E L L S D M K A D V T R L P A T L S R I
P P I A A R L Q M S E R S I L S R L A S K G T E P H P T P A
V M L Q - N G E T P K D L N D E - - K Q K K N I K Q R F M F
N I A D G G F T E L H S L W Q N E E R A A T V T K K T Y E I
W H R R H D Y W L L A G I I N H G Y A R W Q D I Q N D P R Y
A I L N E P F K G E M N R G N F L E I K N K F L A R R F K L
L E Q A L V I E E Q L R R A A Y L N M S E - P S H P S M A L
N T R F A E V E C L A E S H Q H L S K E S - A G N K P A N A
V L H K V L K Q L E E L L S D M K A D V T R L P A T I A R I
P P V A V R L Q M S E R N I L S R L A N R A P E P T P Q Q V
P I E T Q Q N G D K E E D D E G K K E D K K - G K F K F M F
N I A D G G F T E L H T L W Q N E E R A A V S S G K I Y D I
W H R R H D Y W L L A G I V T H G Y A R W Q D I Q N D P R Y
M I L N E P F K S E V H K G N Y L E M K N K F L A R R F K L
L E Q A L V I E E Q L R R A A Y L N M T Q D P N H P A M A L
N A R L A E V E C L A E S H Q H L S K E S L A G N K P A N A
V L H K V L N Q L E E L L S D M K A D V T R L P S M L S R I
P P V A A R L Q M S E R S I L S R L T N R A G D P T I Q Q G
. . G . . E . . . . K K . K R F M F
N I A D G G F T E L H T L W Q N E E R A A . S S G K . Y E I
W H R R H D Y W L L A G I V H G Y A R W Q D I Q N D P R Y
A I L N E P F K . E . N K G N F L E M K N K F L A R R F K L
L E Q A L V I E E Q L R R A A Y L N M S Q . P H P A M A L
N A R F A E V E C L A E S H Q H L S K E S L A G N K P A N A
V L H K V L N Q L E E L L S D M K A D V T R L P A T L S R I
P P V A A R L Q M S E R S I L S R L A N R A E P T P Q Q .
A

hs CHD3,CHD4 and CHD5 aa alignmentTue, Feb 3, 2015 4:00 PMPage 12
Page 12
ABCDEFGHIJKLMNOPQRSTUVWXYZ abcdefghijklmnopqrstuvwxyz.0123456789hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD3
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD4
hsCHD5
hsCHD5
hsCHD5
hsCHD5
hsCHD5
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
consensus
1980
19802000
20002020
20202040 20402060
20602080
20802100 21002120
2120
V L H K V L N Q L E E L L S D M K A D V T R L P A T L S R I
P P I A A R L Q M S E R S I L S R L A S K G T E P H P T P A
Y P P G P Y A T P P G Y G A A F S A A P V G A L A A A G A N
Y S Q M P A G S F I T A A T N G P P V L V K K E K E M V G A
L V S D G L D R K E P R A G E V I C I D D
V L H K V L K Q L E E L L S D M K A D V T R L P A T I A R I
P P V A V R L Q M S E R N I L S R L A N R A P E P T P Q Q V
A Q Q Q - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - -
V L H K V L N Q L E E L L S D M K A D V T R L P S M L S R I
P P V A A R L Q M S E R S I L S R L T N R A G D P T I Q Q G
A F G S S Q M Y S N N F G P N F R G P G P G G I V N Y N Q M
P L G P Y V T D I - - - - - - - - - - - - - - - - - - - - -
- - - - - - - - - - - - - - - - - - - - -
V L H K V L N Q L E E L L S D M K A D V T R L P A T L S R I
P P V A A R L Q M S E R S I L S R L A N R A E P T P Q Q .
A . G F . G . . .
. I T A A T N G P P V L V K K E K E M V G A
L V S D G L D R K E P R A G E V I C I D D





























