Embed Size (px)
Citation preview

doi:10.1006/jmbi.2001.5114 available online at http://www.idealibrary.com on J. Mol. Biol. (2001) 314, 9±21
Simultaneous and Functional Binding of SmpB andEF-Tu �GTP to the Alanyl Acceptor Arm of tmRNA
Sharief Barends1, A. Wali Karzai2, Robert T. Sauer2, Jacek Wower3
and Barend Kraal1*
1Department of BiochemistryLeiden Institute of ChemistryLeiden University, PO Box9502, 2300 RA Leiden, TheNetherlands2Department of BiologyMassachusetts Institute ofTechnology, CambridgeMA 02139, USA3Department of Animal andDairy Sciences, Program inCell and Molecular BiosciencesAuburn University, 210Upchurch Hall, AuburnAL 36849-5415, USA
Present address: A. W. Karzai, 41Building, Department of BiochemistSUNY at Stony Brook, NY 11790, U
Abbreviations used: AlaRS, Ala-tPEP, phosphoenolpyruvate; PK, pyrtmRNA, transfer-messenger RNA.
E-mail address of the [email protected]
0022-2836/01/010009±13 $35.00/0
Transfer-messenger RNA (tmRNA) mimics functions of aminoacyl-tRNAand mRNA, subsequently, when rescuing stalled ribosomes on a 30 trun-cated mRNA without stop codon in bacteria. In addition, this mechanismmarks prematurely terminated proteins by a C-terminal peptide tag as asignal for degradation by speci®c cellular proteases. For Escherichia coli,previous studies on initial steps of this ``trans-translation'' mechanismrevealed that tmRNA alanylation by Ala-tRNA synthetase and bindingof Ala-tmRNA by EF-Tu �GTP for subsequent delivery to stalled ribo-somes are inef®cient when compared to analogous reactions with canoni-cal tRNAAla. In other studies, protein SmpB and ribosomal protein S1appeared to bind directly to tmRNA and to be indispensable for trans-translation. Here, we have searched for additional and synergistic effectsof the latter two on tmRNA alanylation and its subsequent binding toEF-Tu �GTP. Kinetic analysis of functioning combined with band-shiftexperiments and structural probing demonstrate, that tmRNA mayindeed form a multimeric complex with SmpB, S1 and EF-Tu �GTP,which leads to a considerably enhanced ef®ciency of the initial steps oftrans-translation. Whereas S1 binds to the mRNA region of tmRNA, wehave found that SmpB and EF-Tu both interact with its acceptor armregion. Interaction with SmpB and EF-Tu was also observed at the accep-tor arm of Ala-tRNAAla, but there the alanylation ef®ciency was inhibitedrather than stimulated by SmpB. Therefore, SmpB may function as anessential modulator of the tRNA-like acceptor arm of tmRNA during itssuccessive steps in trans-translation.
# 2001 Academic Press
Keywords: Ala-tRNA synthetase; mutant tmRNA; ribosomal protein S1;trans-translation; tRNAAla
*Corresponding authorIntroduction
Transfer-messenger RNA (tmRNA) was ®rstidenti®ed in Escherichia coli as the rescue moleculemediating a so-called trans-translation mechan-ism1 ± 4 and appears to occur in all eubacteria.5,6
The mechanism is activated when ribosomes arestalled either at the 30 end of a truncated messengerRNA without an in-frame stop codon or at clusters
0 Life Sciencery and Cell Biology,SA.
RNA synthetase;uvate kinase;
ing author:
of rare codons in mRNA.7 Alanyl-tmRNA(Figure 1) enters the ribosome at the non-pro-grammed and/or empty A-site and donates its ala-nine residue to the nascent peptide chain.Thereafter, the ribosome hops from the dead-endof the mRNA to a short reading frame in thetmRNA and is ®nally released by termination at aproper stop codon. The truncated protein productis thus fused to a C-terminal peptide tag encodedby tmRNA, which makes it a substrate for proteindegradation (for review and databases see Karzaiet al.,4 Williams8 and Zwieb & Wower9). The mech-anism functions for the removal of truncated andunwanted protein intermediates, and can regulatespeci®c gene expression. For instance, in the caseof coupled transcription-translation of the lacrepressor gene, tmRNA action plays a role in theregulation of active repressor levels.10
# 2001 Academic Press

Figure 1. Predicted secondary structure of E. colitmRNA adopted from Karzai et al.4 The alanylatedtRNA-like region and the reading frame of the tag pep-tide together with the four pseudoknot structures (desig-nated with pk1-pk4) are indicated. The broken lineencompassing nucleotides 1-89 and 300-363 con®nes theshortened version, tmRNA(�90-299).
10 SmpB as a Modulator of the tmRNA Acceptor Arm
During the ®rst steps of the mechanism of trans-translation, tmRNA displays functional interactionswith several proteins, including alanyl-tRNAsynthetase (AlaRS), elongation factor Tu in com-plex with GTP (EF-Tu �GTP), small protein B(SmpB), and ribosomal protein S1.11 ± 15 The ®rsttwo proteins, AlaRS and EF-Tu �GTP, are of gener-al importance for peptide chain elongation. S1,however, is implicated in translation initiation bykeeping the mRNA complexed with the 30 S ribo-
somal subunit,16 and has no af®nity for canonicaltRNAs. Intriguingly, S1 appeared to bind withhigh af®nity to the interior part of tmRNA(Kd � 10 nM) and to be essential for stable associ-ation of Ala-tmRNA with the ribosome.15 In E. coli,SmpB is a small and stable protein with a molecu-lar mass of 18 kDa, encoded by smpB. This gene islocated just upstream of ssrA, encoding tmRNA. Inall eubacteria tested, a gene related to smpB fromE. coli has been found. SmpB is essential fortmRNA-mediated protein tagging.14
Previously we reported that the kinetic inter-actions of AlaRS and EF-Tu �GTP with tmRNA andAla-tmRNA, respectively, are signi®cantly lessef®cient than those with tRNAAla and Ala-tRNAAla,and discussed how those results could be causedpartially by the large differences in molecular sizeand diffusion coef®cient of the two RNAs.13 Wespeculated that additional protein factors mightcompensate for the lower ef®ciency of tmRNA inaminoacylation and binding to EF-Tu �GTP.
Here, we have investigated the effects of proteinsSmpB and S1 on the ®rst two steps of the trans-translation mechanism, the alanylation of tmRNAby AlaRS and the subsequent binding of Ala-tmRNA to EF-Tu �GTP. In an attempt to delimitthe SmpB binding site on the tmRNA molecule, ashortened tmRNA lacking the messenger regionand three of the four pseudoknot structures wasstudied and RNase T1 footprinting was performed.E. coli tRNAAla was used throughout this study asa reference molecule. The results indicate thatSmpB activates the functioning of the acceptor-arm-like structure of the tmRNA molecule.
Results
Does SmpB stimulate complex formationbetween EF-Tu �GTP and Ala-tmRNA?
Our previous work13 on the recognition of Ala-tmRNA by EF-Tu �GTP showed that the af®nity ofthis interaction was about 30-fold lower than in theanalogous situation with a regular tRNA, such asAla-tRNAAla. To assess whether additional factors,like SmpB and S1, might enhance this bindingaf®nity, we used two different assays for complexformation, one based on protection by EF-Tu �GTPof the [3H]alanylated 30 end against RNase Acleavage, and the other on its protection byEF-Tu �GTP against non-enzymatic deacylation byhydroxyl ions. As a control and for comparison,both protection assays were performed also onAla-tmRNA and Ala-tRNAAla complexed withSmpB alone.
The RNase A protection assay was carried out at4 �C with a set of increasing concentrations ofSmpB and one ®xed concentration (5 mM) of EF-Tu �GTP. Results from such experiments aredepicted in Figure 2(a). EF-Tu �GTP, used at thisconcentration with Ala-tmRNA, provides abouthalf-maximal protection of the Ala-30ApC phos-phoester bond against RNase A cleavage. The
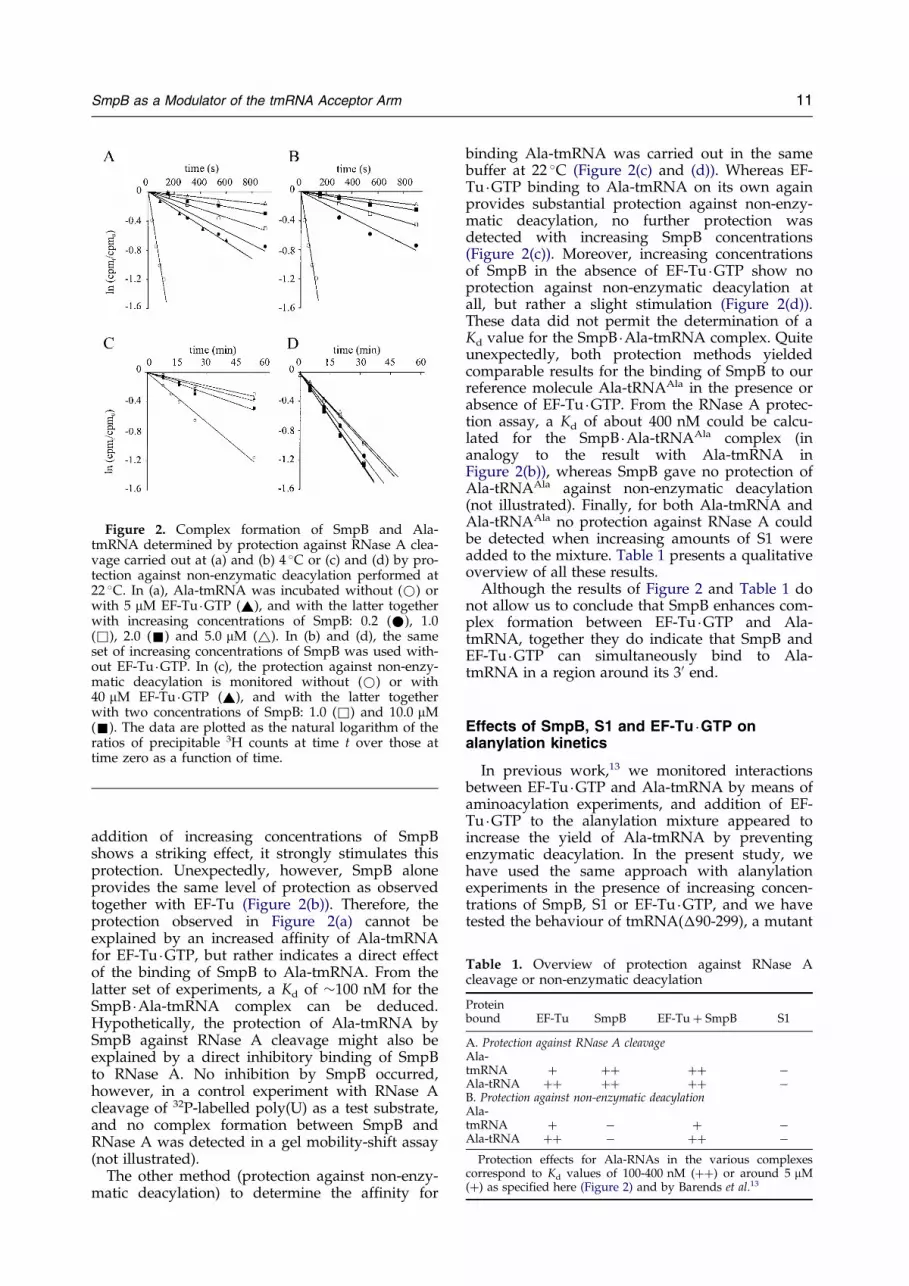
Figure 2. Complex formation of SmpB and Ala-tmRNA determined by protection against RNase A clea-vage carried out at (a) and (b) 4 �C or (c) and (d) by pro-tection against non-enzymatic deacylation performed at22 �C. In (a), Ala-tmRNA was incubated without (*) orwith 5 mM EF-Tu �GTP (~), and with the latter togetherwith increasing concentrations of SmpB: 0.2 (*), 1.0(&), 2.0 (&) and 5.0 mM (~). In (b) and (d), the sameset of increasing concentrations of SmpB was used with-out EF-Tu �GTP. In (c), the protection against non-enzy-matic deacylation is monitored without (*) or with40 mM EF-Tu �GTP (~), and with the latter togetherwith two concentrations of SmpB: 1.0 (&) and 10.0 mM(&). The data are plotted as the natural logarithm of theratios of precipitable 3H counts at time t over those attime zero as a function of time.
Table 1. Overview of protection against RNase Acleavage or non-enzymatic deacylation
Proteinbound EF-Tu SmpB EF-Tu � SmpB S1
A. Protection against RNase A cleavageAla-tmRNA � �� �� ÿAla-tRNA �� �� �� ÿB. Protection against non-enzymatic deacylationAla-tmRNA � ÿ � ÿAla-tRNA �� ÿ �� ÿ
Protection effects for Ala-RNAs in the various complexescorrespond to Kd values of 100-400 nM (��) or around 5 mM(�) as speci®ed here (Figure 2) and by Barends et al.13
SmpB as a Modulator of the tmRNA Acceptor Arm 11
addition of increasing concentrations of SmpBshows a striking effect, it strongly stimulates thisprotection. Unexpectedly, however, SmpB aloneprovides the same level of protection as observedtogether with EF-Tu (Figure 2(b)). Therefore, theprotection observed in Figure 2(a) cannot beexplained by an increased af®nity of Ala-tmRNAfor EF-Tu �GTP, but rather indicates a direct effectof the binding of SmpB to Ala-tmRNA. From thelatter set of experiments, a Kd of �100 nM for theSmpB �Ala-tmRNA complex can be deduced.Hypothetically, the protection of Ala-tmRNA bySmpB against RNase A cleavage might also beexplained by a direct inhibitory binding of SmpBto RNase A. No inhibition by SmpB occurred,however, in a control experiment with RNase Acleavage of 32P-labelled poly(U) as a test substrate,and no complex formation between SmpB andRNase A was detected in a gel mobility-shift assay(not illustrated).
The other method (protection against non-enzy-matic deacylation) to determine the af®nity for
binding Ala-tmRNA was carried out in the samebuffer at 22 �C (Figure 2(c) and (d)). Whereas EF-Tu �GTP binding to Ala-tmRNA on its own againprovides substantial protection against non-enzy-matic deacylation, no further protection wasdetected with increasing SmpB concentrations(Figure 2(c)). Moreover, increasing concentrationsof SmpB in the absence of EF-Tu �GTP show noprotection against non-enzymatic deacylation atall, but rather a slight stimulation (Figure 2(d)).These data did not permit the determination of aKd value for the SmpB �Ala-tmRNA complex. Quiteunexpectedly, both protection methods yieldedcomparable results for the binding of SmpB to ourreference molecule Ala-tRNAAla in the presence orabsence of EF-Tu �GTP. From the RNase A protec-tion assay, a Kd of about 400 nM could be calcu-lated for the SmpB �Ala-tRNAAla complex (inanalogy to the result with Ala-tmRNA inFigure 2(b)), whereas SmpB gave no protection ofAla-tRNAAla against non-enzymatic deacylation(not illustrated). Finally, for both Ala-tmRNA andAla-tRNAAla no protection against RNase A couldbe detected when increasing amounts of S1 wereadded to the mixture. Table 1 presents a qualitativeoverview of all these results.
Although the results of Figure 2 and Table 1 donot allow us to conclude that SmpB enhances com-plex formation between EF-Tu �GTP and Ala-tmRNA, together they do indicate that SmpB andEF-Tu �GTP can simultaneously bind to Ala-tmRNA in a region around its 30 end.
Effects of SmpB, S1 and EF-Tu �GTP onalanylation kinetics
In previous work,13 we monitored interactionsbetween EF-Tu �GTP and Ala-tmRNA by means ofaminoacylation experiments, and addition of EF-Tu �GTP to the alanylation mixture appeared toincrease the yield of Ala-tmRNA by preventingenzymatic deacylation. In the present study, wehave used the same approach with alanylationexperiments in the presence of increasing concen-trations of SmpB, S1 or EF-Tu �GTP, and we havetested the behaviour of tmRNA(�90-299), a mutant

Figure 3. Effects of SmpB and S1 on alanylation ef®-ciencies. Alanylation of 1 mM tmRNA (&, &) and 1 mMtmRNA(�90-299) (~, ~) was performed with 1 mMAlaRS, and alanylation of 1 mM tRNAAla (*, *) with0.25 mM AlaRS. Alanylation ef®ciencies were determinedfrom the initial rates at 37 �C as a function of increasingconcentrations of SmpB (®lled symbols) or S1 (opensymbols) in the presence of 6 % (v/v) glycerol. The ef®-ciency is de®ned here as the relative rate of Ala-RNAformation in comparison to that rate in the absence ofSmpB and S1, and an ef®ciency of 1.0 corresponds to0.11, 0.30 and 0.67 mM/minute for tmRNA,tmRNA(�90-299) and tRNAAla, respectively.
Figure 4. Alanylation kinetics of (a) 1 mM tmRNA and(b) 1 mM tmRNA(�90-299) in the presence of 1 mMAlaRS. The extent of alanylation (molar % of total RNA)was determined in the absence of any additional protein(*) or in the presence of 1 mM SmpB (*), 40 mM EF-Tu �GTP (&), or a combination of both (&). Broken linesshow the theoretical sum of the separate stimulation bySmpB and EF-Tu �GTP on the ®nal extent of the alanyla-tion.
Figure 5. Concomitant effects of EF-Tu �GTP andSmpB on the ef®ciency of tmRNA alanylation. Alanyla-tion of 1 mM tmRNA was measured at 37 �C as a func-tion of increasing concentrations of (a) EF-Tu �GTP or (b)SmpB in the presence of 1 mM AlaRS and 6 % (v/v) gly-cerol. In (a), the initial rates of alanylation were deter-mined in either the absence (*) or presence of 1 mMSmpB (*) and in (b) in the absence (*) or presence (*)of 20 mM EF-Tu �GTP; an ef®ciency of 1.0 corresponds toan apparent rate of 0.10 mM/minute Ala-tmRNAformed. Broken lines represent theoretical results in caseof purely additive effects by EF-Tu �GTP and SmpB.
12 SmpB as a Modulator of the tmRNA Acceptor Arm
tmRNA lacking the messenger region and pseudo-knot structures 2-4. Figure 3 shows that S1 has nopronounced effect on the alanylation ef®ciencies oftmRNA, tmRNA(�90-299) or tRNAAla. The SmpBprotein, however, appears to have a strongly sti-mulating effect on the alanylation of tmRNA,whereas for tRNAAla it does not stimulate andeven inhibits the alanylation when added at con-centrations higher than 1 mM. For tmRNA(�90-299), the stimulatory effect of SmpB is even higherthan that measured for tmRNA.
In a subsequent set of experiments, we analysedthe stimulatory effects of EF-Tu �GTP and SmpB,either separately or together, on the level oftmRNA alanylation as a function of time.Figure 4(a) shows that the steady state betweenalanylation and deacylation is shifted to a muchhigher level of alanylation by the combined actionof these two proteins (black squares). Interestingly,the sum of the separate stimulatory effects by EF-Tu �GTP and SmpB (see the broken line) is some-what less than the values of their combined action.In other words, the interactions of these proteinsare synergistic, or at least additive, and certainlynot mutually exclusive. In Figure 4(b), the samesort of additive/synergistic stimulation can be seenfor the alanylation of tmRNA(�90-299). The effectsare even stronger here, and a level of almost 100 %alanylation can be reached. Furthermore, thecurves for the separate effects of EF-Tu �GTP andSmpB on alanylation have a different shape, which
indicates that the two complementary stimulationmechanisms have a different background.
In order to better understand the concomitanteffects of EF-Tu �GTP and SmpB on the alanylationof tmRNA, we measured the alanylation ef®ciencyas a function of the concentration of either protein,without or with the other protein at a constant andpractically saturating concentration (Figure 5). Itcan be concluded from Figure 5(a) that the stimu-lating effect of increasing concentrations of EF-Tu �GTP on tmRNA alanylation is shifted to ahigher level for the whole concentration range bythe presence of saturating SmpB. In fact, the com-

SmpB as a Modulator of the tmRNA Acceptor Arm 13
bined presence of both proteins leads to an evengreater ef®ciency than just the additive effect. Thesame conclusion can be drawn from Figure 5(b),where the alanylation ef®ciency was determined asfunction of increasing SmpB concentrations in theabsence or presence of a saturating concentrationEF-Tu �GTP. As observed in Figure 3, the stimu-latory effect of SmpB disappears when present athigher concentrations. Again, the results demon-strate that EF-Tu �GTP and SmpB do not competefor binding to tmRNA, but can collaborate instimulation of the alanylation reaction. Similarexperiments with tRNAAla as a reference moleculeshowed just additive results for the effects on ala-nylation by EF-Tu �GTP (stimulating at all concen-trations) and by SmpB (neutral, but inhibitory athigher concentrations).
For a more quantitative study of the effects ofSmpB on the alanylation reaction by AlaRS, westudied the apparent reaction rates as a function ofincreasing substrate concentration of either the ala-nine acceptor RNA alone or in a 1:1 stoichiometricratio with SmpB. Figure 6 shows the Michaelis-Menten curves for tmRNA, tmRNA(�90-299) andtRNAAla together with the calculated kcat/KM
values. Whereas in the absence of SmpB, tmRNAand tmRNA(�90-299) have kcat/KM values about50-fold lower than that for tRNAAla, in the pre-sence of SmpB, this improves to 30-fold and 12-fold lower, respectively. The kcat/KM value fortRNAAla itself is not in¯uenced signi®cantly by thepresence of an equimolar amount of SmpB, thoughthe latter decreases the Vmax and thus the apparentkcat (as appears from the different plateaus of thetwo curves in Figure 6(c)) and, at the same time,lowers the KM (as appears from the difference oftheir initial slopes).
The positive effect of SmpB on the alanylation ofthe tmRNA acceptor arm is mainly due to anenhanced kcat (with only modest effect on KM) andis especially striking for the mutant tmRNA withthe deleted 90-299 region.
Figure 6. Michaelis-Menten plots of the apparent alanylatRNAAla at 37 �C as a function of the RNA concentration, eibation buffer contained 14 % (v/v) glycerol; in (a) and (b)kcat/KM values (103 Mÿ1 sÿ1) are printed below the correspon
Band-shift analysis of SmpB interactionswith tmRNA(���90-299)
In a ®rst attempt to delimit the SmpB bindingsite on the tmRNA structure, we compared bindingaf®nities of SmpB for tmRNA and tmRNA(�90-299) (see Figure 1). The latter, an internally shor-tened version of tmRNA, contains only the tRNA-like acceptor arm extended with pseudoknot pk1,a structure essential for alanylation,17 and it lacksthe open reading frame for the tag peptide and thepseudoknot structures pk2, pk3 and pk4, whichare recognized by ribosomal protein S1.15 Weincluded tRNAAla as a reference molecule with ala-nyl acceptor arm. From the results of a gel mobi-lity-shift assay,14 we already knew that SmpB has ahigh binding af®nity for tmRNA but not for bulktRNA from yeast. Here, we extended these obser-vations by showing that addition of increasingamounts of SmpB results in complex formationand retardation of all three species, tmRNA,tmRNA(�90-299) and tRNAAla, from E. coli(Figure 7). From the concentration-dependence ofthese shifts and the scanned band intensities,apparent Kd values could be calculated that areabout the same for all three (�50 nM). In addition,we con®rmed the speci®city of the SmpB bindingand found no retardation with a bulk mixture ofyeast tRNA (not illustrated). Apparently, theeukaryotic tRNAAla in this mixture has no highaf®nity for SmpB and, as known, eukaryotic organ-isms do not contain tmRNAs like eubacteria.
SmpB binding sites on tmRNA as determinedby footprinting
From the protection assays and alanylationexperiments carried out in the presence of SmpB,we deduced that SmpB binding to tmRNA wouldinvolve the tRNA-like moiety of tmRNA. More-over, tmRNA(�90-299), which lacks the internalpart of the full-length molecule, appeared to havean af®nity for SmpB similar to that of the full-
tion rates of (a) tmRNA, (b) tmRNA(�90-299) and (c)ther free (*) or in a 1:1 ratio with SmpB (*). The incu-1 mM AlaRS was present, in (c) 0.25 mM. The calculatedding curves.

Figure 7. Gel retardation patterns of SmpB binding totmRNA, tmRNA(�90-299), or tRNAAla. The 32P-labelledRNAs were complexed with different concentrations ofSmpB, separated on a 1 % (w/v) agarose gel and visual-ized by phosphor imaging.
Figure 8. Footprints of SmpB in complex with (a)tmRNA or (b) tRNAAla as detected by RNase T1 diges-tion. The cleavage patterns were, in each case, obtainedfor either free RNA or RNA with an equimolar amountof SmpB, under conditions of increasing concentrationsof RNase T1 as indicated over the lanes. Lanes L containthe alkaline degradation ladder and lanes C theuntreated RNA sample. The numbering corresponds tothe nucleotide positions indicated in Figure 9(a) and (d).
14 SmpB as a Modulator of the tmRNA Acceptor Arm
length product (Figure 7), and its alanylation isstimulated in a similar way by addition of SmpB(Figures 3 and 4).
To localisze further the binding of SmpB totmRNA, we used RNase T1 footprinting. Thedigestion pattern of 30-end labelled tmRNA withincreasing concentrations of RNase T1 shows aclear footprint when SmpB is present (Figure 8(a)).The few faint bands visible in the control lanes(samples incubated without RNase T1) do notinterfere and represent non-speci®c minor cuts. Wedo not observe bands of decreasing intensity withincreasing RNase T1 concentration, which excludesthe interference by any secondary cut in themolecule.18 The localisation of this footprint is indi-cated in the secondary structure model of tmRNAand in a predicted spatial model (Figure 9(a)-(c)).In the SmpB � tmRNA complex, nucleotides G355,G351, G336 and G333 are protected by SmpBagainst RNase T1 cleavage. These nucleotides arelocated around the acceptor stem structure of thetRNA-like structure of tmRNA (®lled triangles inFigure 9(a), red nucleotides in Figure 9(b) and (c)).Further upstream, the available G nucleotides areno longer protected in the SmpB � tmRNA complex.Footprint analysis in the 50-end region of tmRNAdid not show additional effects of SmpB complexformation either. For tmRNA(�90-299), similarfootprinting results were observed (not illustrated).
In parallel, we performed an RNase T1 footprintassay on SmpB � tRNAAla (Figure 8(b)). In the lattercase, upon addition of equimolar amounts ofSmpB, only relatively weak or no protection ofavailable G nucleotides against RNase T1 digestioncould be detected. In great contrast, SmpB binding
to tRNAAla appeared to expose several G nucleo-tides to RNase T1, again located mainly in theacceptor arm and variable loop (open diamonds inFigure 9(d), green nucleotides in Figure 9(e) and(f)). Examining our collection of footprint pictures,we found that the relatively weak protections, suchas those at U34, G43 and G53 in Figure 8(b) werenot suf®ciently signi®cant for presentation inFigure 9(d)-(f). These observations imply thatSmpB binds to tRNAAla in a different manner,albeit with a comparable af®nity, than to tmRNA.
Direct evidence for simultaneous binding ofSmpB and EF-Tu to Ala-tmRNA
So far, various lines of experimental evidencepresented here point clearly to a simultaneousbinding of SmpB and EF-Tu to the alanylatedacceptor arm of tmRNA. In a ®nal experiment wetried to obtain direct and physical evidence forsuch a simultaneous binding. We chose a gel mobi-lity-shift assay in order to detect a super-shift bythe combined presence of the two proteins. Weanticipated dif®culties posed by the relativelyweak af®nity of EF-Tu �GTP for Ala-tmRNA, itslack of af®nity for deacylated tmRNA and the easy

Figure 9. Mapping of SmpB footprints together withAlaRS and EF-Tu contact sites on the structures oftmRNA and tRNAAla. SmpB footprinting results ofFigure 8 are indicated in the secondary structures of (a)the predicted tRNA-like region of tmRNA and (d)tRNAAla. SmpB protection against RNase T1 cleavage isindicated by (~), unprotected RNase T1 cleavage by(~), and SmpB-induced enhanced RNase T1 cleavage by(}). The three-dimensional location of these sites,together with contact sites of AlaRS and EF-Tu, areshown in (b) and (c) for tmRNA and in (e) and (f) fortRNAAla, each pair representing two orthogonal top andside-views. The model of the tRNA-like region oftmRNA was taken from Zwieb & Wower9 and is basedon chemical and enzymatic probing,30 phylogeneticevidence8,9 and UV-induced intramolecular cross-linking.31 The model of tRNAAla is based on the crystalstructure of free tRNAPhe from yeast.32 The putativeinteraction sites for AlaRS (yellow) and EF-Tu (shadedcircles) are drawn in accordance with the data known
Figure 10. Mobility-shift analysis of complex for-mation between SmpB, EF-Tu �GTP and the alanylatedacceptor arm of tmRNA. Samples contained 2 mMtmRNA(�90-299) in the (a) presence or (b) absence ofalanylation activity, and with AlaRS, SmpB and EF-Tu �GTP at the indicated concentrations. RNA-contain-ing complexes were analyzed on a 1.2 % (w/v) agarosegel after ethidium bromide staining.
SmpB as a Modulator of the tmRNA Acceptor Arm 15
non-enzymatic deacylation of preparations of (par-tially) alanylated tmRNA.
In order to reveal higher-order complex for-mation between Ala-tmRNA, SmpB and EF-Tu, itwas important to keep the extent of alanylation ashigh as possible and to use at least tenfold higherconcentrations of the individual proteins than theirapparent Kd values in Ala-tmRNA complexes withSmpB or EF-Tu �GTP separately. Also, relativelyhigh tmRNA concentrations were used, whichenabled detection of RNA-containing bands byethidium bromide staining. Because the alanylationof tmRNA by AlaRS is stimulated considerably bythe presence of SmpB and/or EF-Tu, we decidedto have an active AlaRS system present duringcomplex formation of the (partially) alanylatedtmRNA with SmpB and/or EF-Tu. We decided touse the shortened tmRNA(�90-299) in these band-shift assays, because its alanylation is stimulatedeven better by SmpB and EF-Tu (up to 100 %,Figure 4(b)) and because eventual mobility-shiftson gel would be more pronounced than with themuch larger (363 nt) tmRNA molecule.
Representative results are shown in Figure 10.The ®rst two lanes of Figure 10(a) show no signi®-cant band-shift effects of the addition of EF-Tu �GTP to Ala-tmRNA(�90-299) (preparationwith about 50 % alanylation; see Figure 4(b)). Incomparison, however, to the corresponding twocontrol lanes of Figure 10(b) (no alanylation), theband at position 1 in (a) lane 2 has a smeared tailthat might originate from a rapid dissociation ofthe weak EF-Tu �GTP �Ala-tmRNA(�90-299) com-plex during electrophoresis. In Figure 10(a), lanes3-5, increasing concentrations (0.5-2.0 mM) of SmpB
for Phe-tRNAPhe and tRNAAla, respectively.26,29 SmpBbinding leads to protection against RNase T1 cleavage ofnucleotides marked in (b) and (c) red or leads toenhanced cleavage as marked in (e) and (f) green. (b),(c), (e) and (f) were prepared with the programme

16 SmpB as a Modulator of the tmRNA Acceptor Arm
lead to the expected full complex formation withAla-tmRNA(�90-299) and a corresponding band-shift of the RNA from position 1 to position 2.The same band-shift occurs with deacylatedtmRNA(�90-299) in the absence of AlaRS(Figure 10(b), lane 3), as observed before. Interest-ingly, in Figure 10(a), lanes 3-5 an additional shiftappears at position 3 that is absent from (b), lane 3.Apparently, this complex is due to the combinedpresence of AlaRS and SmpB, and indeed weknow that the alanylation of tmRNA(�90-299) byAlaRS is stimulated about twofold by an equimolaramount of SmpB (up to a level of about 75 % ala-nylation, Figure 4(b)). The Ala-tmRNA(�90-299)produced has probably lost its af®nity for AlaRSbut not that for SmpB (band at position 2).
In the last three lanes in Figure 10(a), one seesthe effect of the additional presence of EF-Tu �GTPin the situation already analyzed in the previouslanes 3-5. Smearing of the RNA-containing bandsis now much more pronounced and in the last lanewith the highest concentration of SmpB, a newsmeared band has shifted to position 4. As thelatter band is absent from the control experimentof Figure 10(b), last lane, with unchargedtmRNA(�90-299), we see this band as evidence fora complex of Ala-tmRNA(�90-299) with EF-Tu �GTP and SmpB together. An additional pre-sence of AlaRS in this complex is unlikely, becauseAlaRS would not bind to the alanylated acceptorarm and the conditions of that particular exper-iment result in almost 100 % alanylatedtmRNA(�90-299) as can be deduced fromFigure 4(b).
Discussion
For the tmRNA-mediated trans-translation mech-anism, a newly identi®ed protein factor, SmpB,was found to be crucial. Accordingly, an smpB del-etion mutant exhibits the same phenotype as atmRNA knock-out strain.14 Interestingly, ribosomalprotein S1 was shown to bind tmRNA with a simi-lar af®nity and the same sort of speci®city as doesSmpB.15 Both proteins bind to deacylated tmRNAand are important for Ala-tmRNA binding to 70 Sribosomes. These results, together with our pre-vious report13 on the relatively low af®nities ofboth tmRNA and Ala-tmRNA for binding toAlaRS and EF-Tu �GTP, respectively, led us tospeculate about the involvement of SmpB and/orany other protein factor in the subsequent pro-cesses of tmRNA alanylation and complex for-mation with EF-Tu �GTP. Here, we report theeffects of both auxiliary proteins, SmpB and S1, onthese processes.
In our search for effects of SmpB on Ala-tmRNAcomplex formation with EF-Tu �GTP, we were sur-prised to ®nd that SmpB itself can protect the 30end of Ala-tmRNA against RNase A-mediatedcleavage but not against non-enzymatic deacyla-tion. With the former assay, a Kd of 100 nM was
calculated for the SmpB �Ala-tmRNA complex,which is close to the 20-50 nM value calculatedfrom the band shift assays performed with deacy-lated tmRNA (Karzai et al.14 and this study).Apparently, the alanyl residue itself does not muchin¯uence the interaction between tmRNA andSmpB. The strong protection by SmpB of the Ala-30ApC end against RNase A cleavage was notfurther improved signi®cantly by the presence ofEF-Tu �GTP. Such an additional improvementwould in fact be dif®cult to measure in view of themuch weaker protection by EF-Tu �GTP with its100-fold lower af®nity for Ala-tmRNA.
Unexpectedly, binding and protection by SmpBwas found for Ala-tRNAAla. Whereas the band-shift assay (Figure 7) yielded similar values(� 50 nM) for the af®nity of SmpB to deacylatedtmRNA or tRNAAla, in the case of the alanylatedRNAs, the RNase A protection assay yielded Kd
values of 100 nM (Ala-tmRNA) and 400 nM (Ala-tRNAAla) for the corresponding af®nity of SmpB(Figure 2(b) and Results). The effect of alanylationon tRNAAla seems thus less favourable for SmpBbinding than in the case of tmRNA. Again, also forAla-tRNAAla no protection at all was found againstnon-enzymatic hydrolysis of the aminoacyl esterbond and, just as with Ala-tmRNA, even a ten-dency for enhanced deacylation could be observed.This indicates that in both types of SmpB �RNAcomplexes the 30 ends are well accessible to smallmolecules. On the other hand, the well-establishedprotection against non-enzymatic deacylation byternary complex formation with EF-Tu �GTP washardly affected by adding increasing concen-trations of SmpB. The two different protectionassays together thus suggest that both proteinsbind around the 30 end of the alanyl acceptor armin a mutually non-exclusive way (Table 1).
Because of the unexpectedly strong protection bySmpB of the 30 end of tmRNA against enzymaticdegradation, we investigated whether SmpB atincreasing concentrations would affect the enzy-matic alanylation at the 30 end. The results areremarkable, in the sense that SmpB appears tostimulate the latter enzymatic reaction when pre-sent at a 1:1 ratio with 1 mM tmRNA. At muchhigher SmpB to tmRNA ratios, the stimulationfades away (Figures 3 and 5). This explains whyno stimulation was observed in a previous studywith 2 mM SmpB and 0.36 mM tmRNA.14 At thesetmRNA concentrations, much above the Kd value(�50 nM) for tmRNA �SmpB complex formation, itis the molar ratio that determines to what extent a1:1 complex can be formed, by titration. We didnot test whether the loss of stimulation at the high-er SmpB concentrations was due to the binding ofan additional SmpB molecule to tmRNA; and otherexplanations might be envisaged. In parallel exper-iments with tRNAAla, no SmpB effect on the alany-lation reaction was observed under conditionswhere a 1:1 complex can be formed, but by furtherraising the SmpB concentration, total inhibitionsoon occurs (Figure 3). The biological relevance of

SmpB as a Modulator of the tmRNA Acceptor Arm 17
the latter inhibition has not been further investi-gated in this work with its focus on tmRNA. Oneshould note, that this inhibition occurs at an SmpBto tRNAAla ratio higher than 1 and at higher SmpBconcentrations (>1 mM) than needed for the initialstimulation of alanylation of tmRNA in Figure 3.Inhibition in vivo could therefore be predicted fortRNAAla only if SmpB to tmRNA ratios were con-siderably higher than 1.
Just as with the protection experiments dis-cussed above, we sought to analyse the effect ofthe simultaneous presence of SmpB and EF-Tu �GTP on the alanylation kinetics by AlaRS. Ourresults clearly point to an additive and even smallsynergistic effect of their combined presence rela-tive to their individual stimulation of the alanyla-tion ef®ciency of tmRNA (Figures 4 and 5). TheSmpB and EF-Tu �GTP contributions both posi-tively in¯uence the initial rate of the alanylationand the steady state level of alanylation and deacy-lation reactions (Figure 4), in a manner that doesnot involve the internal region (nucleotides 90-299)of tmRNA.
The synergistic effects between SmpB and EF-Tu �GTP as measured in Figure 5 (with one proteinat a constant and saturating concentration and theother at increasing concentrations) apparently donot in¯uence their individual binding af®nity fortmRNA in a substantial way. The EF-Tu �GTP con-centration at which half-maximal stimulation oftmRNA alanylation occurs, is about 5 mM, in theabsence or the presence of SmpB. This value of5 mM corresponds to the apparent Kd value foundearlier for the EF-Tu �GTP �Ala-tmRNA complexformation.13 The maximal stimulation by SmpB oftmRNA alanylation occurs at an about equimolarratio, either in the absence or the presence of EF-Tu �GTP. This corresponds to the expected titrationbehaviour of components with Kd 4 100 nM.
Our in vitro RNA transcripts do not contain theT and modi®cations in their T loop such as thenative tmRNA and tRNAAla, and one might won-der whether this lack would make them poorersubstrates for AlaRS and more dependent on acti-vator proteins. However, a direct involvement ofthe modi®ed nucleotides in alanylation is unlikely,since very similar kinetic parameters for the modi-®ed and unmodi®ed versions of tRNAAla havebeen reported.19 The post-transcriptional modi®-cations probably have only a minor, if any, contri-bution to the overall stability of the RNA, since acomparison of UV melting pro®les of nativetmRNA and the in vitro transcript shows nodifference.13,20 The activity of our in vitro tmRNAtranscript is better than that of the in vivo tmRNAproducts described in the literature thus far. Atleast 55 % of our preparation is active in alanyla-tion, whereas other groups report up to 40 % attheir alanylation plateau of native preparations.11,21
The alanylation level of our tmRNA(�90-299) tran-script even reaches 95 % (Figure 4), which inaddition points to very little heterogeneity at the 30end of this T7 transcript preparation. Because the
alanylation levels of our tmRNA transcripts aresigni®cantly higher than those reported for nativetmRNA, there is no obvious reason to hypothesizethat the T-loop modi®cations of the latter categorywould circumvent a need for activation by SmpBon the acceptor arm.
The speci®c activation of tmRNA alanylation bySmpB is demonstrated convincingly by absence ofactivation or even inhibition upon SmpB bindingto tRNAAla (Figure 6). Notably, the SmpB stimu-lation of the alanylation ef®ciency of tmRNA isabout fourfold for the tmRNA(�90-299) variantand is mainly due to an effect on the apparent kcat.This might, for example, be caused by betteraccessibility or conformational adaptation of theSmpB-bound RNA substrate in contact withthe AlaRS enzyme. Evidence for such anSmpB � tmRNA �AlaRS complex is indeed availablein Figure 10(a) by the band-shift at position 3. Wesee interesting analogies between this activatingeffect of SmpB and that of other recently discov-ered proteins that facilitate tRNA binding and sub-sequent aminoacylation reaction on a synthetase.For example, Arc1p from Saccharomyces cerevisiaeenhances methionyl-tRNA synthetase activity byincreasing its tRNA af®nity,22,23 the eukaryotic pro-EMAPII protein enhances the aminoacylationactivity of arginyl-tRNA synthetase 2.5-fold bydecreasing the apparent KM value,24 and theTrbp111 protein from the extreme thermophilicbacterium Aquifex aeolicus does not seem to haveany preferences for a special tRNA, but generallyrecognizes the L-shaped structure of tRNAs.25
Although these proteins do not display signi®cantsequence similarity to SmpB, their functioning isreminiscent of that of SmpB during tmRNA alany-lation.
In our experiments with S1, we saw no effects ofits binding to tmRNA in the protection assay ofthe 30 end (Figure 2) or the alanylation assay(Figure 3). This ®ts with the localization of its bind-ing site on tmRNA as reported by Wower et al.15
The latter study showed that the high af®nity of S1for tmRNA (Kd about 10 nM) is lost for the mutanttmRNA(�90-299), which locates the binding regionin the interior parts of tmRNA rather than in itstRNA-like part. Moreover, cross-linking exper-iments in that study revealed that the S1 proteinbinds to an interior region, encompassing tmRNAnucleotides 60-247.
From the crystal structures of Phe-tRNA andCys-tRNA in their respective complexes with EF-Tu and a GTP analogue,26,27 one can see that EF-Tucontacts the aminoacyl-tRNA on one side of theacceptor arm from its 30 end and along acceptorstem and T stem up to the variable loop. ThistRNA contact had been predicted by extensiveenzymatical probing analysis.28 Our RNase T1 foot-print of SmpB on tmRNA borders that side of theacceptor arm. However, we have clear evidencethat SmpB and EF-Tu �GTP do not compete in theirbinding to the acceptor arm, as measured in theassays on protection of the alanylated 30 end (no

18 SmpB as a Modulator of the tmRNA Acceptor Arm
interference, Figure 2 and Table 1) and in theactivity assays on their stimulation of alanylation(additive or even synergistic effects at saturatingconcentrations, Figures 4 and 5). We also havephysical evidence for a simultaneous binding ofSmpB and EF-Tu �GTP by the super-shift oftmRNA(�90-299) in Figure 10(a) at position 4.
The resolution of structure mapping with enzy-matic protection or footprinting methods is far toolow for model building, but we think of a pro-tein �Ala-tmRNA complex with room for SmpBand EF-Tu �GTP on one side of the acceptor armand an independent position for S1 around themessenger region coding for the tag peptide.15
Such a complex is ®t for entering a non-pro-grammed vacant ribosomal A-site. In view of theacceptor arm location of SmpB, it seems unlikelythat SmpB would act as a protein mimic of ananticodon arm structure to stabilize A-site bindingof the Ala-tmRNA �protein complex.
It is interesting to note (see Figure 9(b) and (c))that the proposed contact area for SmpB is at a siteof the alanyl-acceptor arm opposite to the arearecognised by AlaRS.29 As already discussed,SmpB may thus bind to uncharged tmRNAtogether with AlaRS (see super-shift in Figure 10(a)at position 3) and stimulate the reaction catalysedby the latter, perhaps by preventing the backwardreaction (deacylation) at the hydrolytic centre ofAlaRS. The model in Figure 9(b) and (c) alsoexplains the observation that only EF-Tu is able toprotect the Ala 30-ester bond of Ala-tmRNA againstnon-enzymatic deacylation by small H2O or OH ÿ
nucleophiles, whereas both EF-Tu and SmpB canprotect the penultimate 30C from surround andcleavage by the active centre of RNase A.
The alanyl acceptor arms of Ala-tRNA and Ala-tmRNA are equally well protected against RNaseA cleavage by SmpB, but their modes of SmpBbinding are different. SmpB does not stimulate butrather inhibits the alanylation by AlaRS, andRNase T1 footprinting reveals exposure of nucleo-tides around the T stem rather than shielding asfor tmRNA (see Figure 9(d)-(f)). Obviously, therespective alanyl acceptor arms of tmRNA andtRNAAla are not identical and an important differ-ence may occur in the tertiary folding around theso-called elbow region.
From our experiments, we conclude that SmpBdoes not primarily enhance the relatively modestaf®nity of tmRNA for AlaRS and EF-Tu �GTP, butat least independently increases the ef®ciency ofthe alanylation reaction (effect on kcat/KM) and pro-tects the Ala-tmRNA product against enzymaticdeacylation (effect on apparent kcat). This probablyoccurs by modulating the tRNA-like region oftmRNA. Possible effects of this modulation on thethree-dimensional structure are under investi-gation. Other effects of SmpB on Ala-tmRNAduring A-site positioning remain open for furtherresearch.
Materials and Methods
Isolation of proteins
C-terminally (His)6-tagged versions of EF-Tu andAlaRS from E. coli were obtained from the expressionplasmids pKECAHis34 and pQE-875.35 Expression andpuri®cation were carried out as described.13 SmpB andthe C-terminally (His)6-tagged S1 protein from E. coliwere isolated as described earlier.14,15 All proteins wereat least 98 % pure as judged by SDS-PAGE analysis. Pro-tein concentrations were determined spectrophotometri-cally (on the basis of contributions to the molarextinction coef®cient at 280 nm of 5600 Mÿ1 cmÿ1 perTrp and 1420 Mÿ1 cmÿ1 per Tyr residue in the proteinmolecule), and by the Coomassie Protein Assay (Pierce);these assays gave similar values. EF-Tu �GDP wasactivated to EF-Tu �GTP immediately before use byincubation at 37 �C for 15 minutes in buffer A (50 mMTris-HCl (pH 7.6), 7 mM MgCl2, 60 mM NH4Cl) comple-mented with 100 mM GTP, 6 mM phosphoenolpyruvate(PEP) and 10 mg/ml of pyruvate kinase (PK). After15 minutes of incubation, the mixture was kept on icebefore use.
Transcription
The full-length tmRNA molecule from E. coli and itsshortened derivative lacking nucleotides 90-299 wereexpressed from the plasmids pT710Sa#21 and ptm(�90-299),15 respectively (see Figure 1). E. coli tRNAAla wastranscribed from the pALA119 plasmid.36 Primers SB1(50-GATTACGAATTCTAATACGACTCACTATAGGGGCTGATTCTG GATTCGACGGG-30) and SB2 (50-GGTGGAGCTGGCGAGAGTTGAACCC-30) were used for PCRampli®cation of DNA templates for the transcriptionof tmRNA and tmRNA(�90-299) species lacking the30-terminal adenosine residue. Primers SB3 (50GAATTCCTGCAGTAATACGACTCACTATACGGG-30) andSB4 (50-GGTGGAGCTAAGCGGGATCGAACCG-30)were used for PCR ampli®cation of the DNA templatefor the transcription of tRNAAla lacking the 30-terminaladenosine residue. Primers were ordered from Gibco-BRL. All RNA species were obtained by run-off tran-scription in vitro (using MvaI-linearized plasmid DNAfor full-length species) by means of the T7 RiboMAXlarge-scale RNA-production system (Promega). Puri®-cation of transcripts was as described.37 Integrity andquantity of the transcripts were checked on ethidiumbromide-stained agarose gels and by spectrophotometricmeasurements of the denatured RNA (1 mg/ml of RNAin 98 % (v/v) formamide corresponding to 25 absorbanceunits at 260 nm). In vitro synthesis of internally (a-32P)-labelled RNAs was carried out by means of randomincorporation of [a-32P]UTP (3000 Ci/mmol) using theT7 RiboMAX kit.
32P labelling of 50 and 30 ends
For 50-end labelling, transcripts were ®rst depho-sphorylated with calf intestinal phosphatase in buffer Aat 37 �C for 30 minutes. The RNA was extracted withphenol (saturated with 0.2 M sodium acetate buffer,pH 5.0) and precipitated with ethanol. Subsequent label-ling was carried out in 50 ml of buffer A with 0.1 mM50-dephosphorylated RNA transcript, 0.1 mM [g-32P]ATP(4500 Ci/mmol) and 0.2 unit/ml of phage T4 polynucleo-tide kinase. After one hour incubation at 37 �C, reactionswere stopped by the addition of an equal volume of stop

SmpB as a Modulator of the tmRNA Acceptor Arm 19
solution (98 % (v/v) formamide, 10 mM EDTA, 0.025 %(w/v) xylene cyanol FF, 0.025 % (w/v) bromophenolblue) and the (50-32P)-phosphorylated RNAs were puri-®ed by 5 % (w/v) PAGE under denaturing conditions.
Transcripts lacking the 30-terminal adenosine residuewere labelled by (a-32P) adenylation. Reactions were car-ried out in 100 ml of buffer A supplemented with 0.1 mM[a-32P]ATP (3000 Ci/mmol), 0.1 mM RNA, 0.2 unit/ml ofRNAguard and 0.1 mg/ml of CTP/ATP:tRNA nucleotidyltransferase from yeast. After 16 hours incubation at 4 �C,samples were treated with one volume of phenol (pH 5.0)and one volume of chloroform. After an additionalchloroform extraction, the aqueous layer was appliedonto a Micro Bio-Spin (P-30) column (Bio-Rad), toremove unincorporated [a-32P]ATP. Enzymes and radio-isotopes were from Amersham-Pharmacia.
Band-shift assays
Conditions for measuring Kd values for SmpB �RNAcomplex formation were adopted from Karzai et al.14
Samples (20 ml) of 0.1 nM (a-32P)-labelled RNA andincreasing concentrations of SmpB in buffer A with 5 %(v/v) glycerol were incubated at room temperature for15 minutes and loaded onto a 1 % (w/v) agarose gel in20 mM Tris-borate (pH 7.5), 0.5 mM MgCl2. After elec-trophoresis at 100 V in the same buffer at room tempera-ture for 30-60 minutes, the 32P-labelled bands wereanalysed by a Personal Molecular Imager FX (Bio-Rad).
For the detection of higher-order complex formationbetween Ala-tmRNA, SmpB and EF-Tu, much higherconcentrations of these components were used, in thepresence of an active alanylation system with AlaRS.Alanylation and subsequent further complex formationtook place in a ®nal volume of 50 ml of buffer A comple-mented with 1 mM Ala, 2 mM ATP, 100 mM GTP, 6 mMPEP and 0.5 mg of PK, and with 2 mM tmRNA(�90-299)together with 1 mM AlaRS and/or SmpB (0.5, 1.0, or2.0 mM) and/or 50 mM EF-Tu, as indicated. Before theaddition of tmRNA(�90-299) and AlaRS to these mix-tures, the latter were pre-incubated at 37 �C for 15 min-utes for a full conversion of EF-Tu �GDP to EF-Tu �GTP.After completion of the mixtures, they were incubatedfor another 30 minutes at 37 �C and then chilled on ice.After addition of 5 ml of dimethyl sulphoxide, sampleswere loaded onto a 1.2 % (w/v) agarose gel in 20 mMTris-borate (pH 7.2), 0.5 mM MgCl2. Electrophoresis tookplace at 150 V and 4 �C for 20 minutes, in the same buf-fer complemented with a pre-incubated mixture of10 mM GTP, 20 mM PEP and 2 mg/ml of PK. RNA-con-taining bands were visualised afterwards by ethidiumbromide staining.
Footprint analysis
RNase T1 digestions of tmRNA and tRNAAla in theabsence or the presence of SmpB were carried out inorder to detect SmpB binding sites on these RNA mol-ecules. Experimental procedures were adopted fromWikman et al.28 Reaction volumes of 20 ml of buffer Awith 0.1 mM (a-32P) 30-end labelled RNA (see above)were incubated without or with an equimolar amount ofSmpB at room temperature for 15 minutes. The reactionwas started by the addition of RNase T1 at concen-trations in a range from 1 to 100 m units/ml. After tenminutes incubation at 22 �C, the digestions were stoppedby the addition of 20 ml of stop solution. After loadingonto a 15 % (w/v) polyacrylamide gel containing 8 M
urea, electrophoresis was carried out in 45 mM Tris-borate (pH 8.3), 1 mM EDTA. Reference lanes containeda sample of non-digested RNA as a control and an alka-line degradation ladder of the RNA sample (obtainedby incubation at 95 �C for three minutes with freshlyprepared 50 mM Na2CO3/NaHCO3, 1:9, v/v) for theassignment of cleavage sites. The results shown arerepresentative for experiments that were reproduced atleast three times.
Besides footprint analysis of SmpB binding in the 30end-labelled region of tmRNA, footprint analysis in the50 region was performed by means of a primer extensionmethod. After the RNase T1 digestion (see above),samples were extracted with one volume of phenol(pH 5.0) and one volume chloroform, and subsequentlyprecipitated with ethanol. Primer hybridization wasdone in 4 ml of 50 mM Tris-HCl (pH 7.0), 60 mMKCl, 10 mM DTT containing 0.2 pmol of digested RNAand 1 pmol of (50-32P) end-labelled primer SB5(50-GCGGCTTTTTACGAGGCC-30) complementary toG61-C79. Samples were incubated at 50 �C for15 minutes followed by further incubation at 37 �C for15 minutes. Reverse transcription was performed at37 �C in the presence of one unit of AMV reverse tran-scriptase (Promega). Reactions were stopped after 35minutes by the addition of one volume of stop solution.DNA sequencing of the 50 tmRNA region was carriedout by using the same (50-32P) end-labelled primer (SB5),as described by the supplier (Pharmacia). Samples wereanalysed by denaturing 10 % (w/v) PAGE and phospho-imager detection.
Aminoacylation
Aminoacylation kinetics of the RNA species weredetermined as described.13 When either EF-Tu �GTP orSmpB was present during the alanylation reaction,RNAs were pre-incubated with that protein at 22 �C for15 minutes. Reaction ef®ciencies were measured as initialrates at 37 �C and kcat/KM values were calculated bycomputerized data ®tting on the basis of the Michaelis-Menten equation. Preparations of alanylated RNAspecies were isolated as described.13 The percentage ofalanylation was calculated from the molar amount ofcovalently linked [3H]Ala (as measured by precipitation,®ltration and liquid scintillation counting13) and themolar amount of total RNA (as determined from theabsorbance at 260 nm).
Alanyl-RNA protection assays
The effects of SmpB, S1 and EF-Tu �GTP on complexformation with Ala-tmRNA or Ala-tRNAAla were testedby a special RNase A protection assay.13,38 Reactionswere carried out in a total volume of 200 ml. First, 5 nM[3H]Ala-RNA was equilibrated with protein at the indi-cated concentration in buffer A at 4 �C for 20 minutes.The reaction was started by the addition of 6 nM RNaseA, and the decay of precipitable [3H]Ala-RNA with timewas followed. The protection by EF-Tu �GTP and SmpBof the Ala-RNA ester bond against non-enzymaticdeacylation at 22 �C was assayed under the same bufferA conditions, and again the decay rate of precipitable[3H]Ala-RNA was measured.39

20 SmpB as a Modulator of the tmRNA Acceptor Arm
Acknowledgments
We thank Drs L. Ribas de Pouplana and P. Schimmelfor the expression plasmid of AlaRS, and Dr J. Horowitzfor that of tRNAAla. We thank G. Girard for help withfootprinting experiments, P.W.G. Verlaan for the gift ofCTP/ATP:tRNA nucleotidyl transferase, S.H.E. van denWorm for help with the 3D modelling programme andDrs C.W.A. Pleij and G.P. van Wezel for discussions(Leiden University). This work was supported by a grantto S.B. and B.K. from the Council for Chemical Sciencesof the Netherlands Organization for Scienti®c Research(CW-NWO, 328-035) and by a grant to J.W. from theNational Institutes of Health, USA (GM 58267).
References
1. Tu, G. F., Reid, G. E., Zhang, J. G., Moritz, R. L. &Simpson, R. J. (1995). C-terminal extension of trun-cated recombinant proteins in Escherichia coli with a10Sa RNA decapeptide. J. Biol. Chem. 270, 9322-9326.
2. Keiler, K. C., Waller, P. R. & Sauer, R. T. (1996).Role of a peptide tagging system in degradation ofproteins synthesized from damaged messengerRNA. Science, 271, 990-993.
3. Muto, A., Ushida, C. & Himeno, H. (1998). A bac-terial RNA that functions as both a tRNA and anmRNA. Trends Biochem. Sci. 23, 25-29.
4. Karzai, A. W., Roche, E. D. & Sauer, R. T. (2000).The SsrA-SmpB system for protein tagging, directeddegradation and ribosome rescue. Nature Struct. Biol.7, 449-455.
5. Zwieb, C., Wower, I. K. & Wower, J. (1999). Com-parative sequence analysis of tmRNA. Nucl. AcidsRes. 27, 2063-2071.
6. Keiler, K. C., Shapiro, L. & Williams, K. P. (2000).tmRNAs that encode proteolysis-inducing tags arefound in all known bacterial genomes: a two-piecetmRNA functions in Caulobacter. Proc. Natl Acad. Sci.USA, 97, 7778-7783.
7. Roche, E. D. & Sauer, R. T. (1999). SsrA-mediatedpeptide tagging caused by rare codons and tRNAscarcity. EMBO J. 18, 4579-4589.
8. Williams, K. P. (2000). The tmRNA website. Nucl.Acids Res. 28, 168.
9. Zwieb, C. & Wower, J. (2000). tmRDB (tmRNA data-base). Nucl. Acids Res. 28, 169-170.
10. Abo, T., Inada, T., Ogawa, K. & Aiba, H. (2000).SsrA-mediated tagging and proteolysis of LacI andits role in the regulation of lac operon. EMBO J. 19,3762-3769.
11. Komine, Y., Kitabatake, M., Yokogawa, T.,Nishikawa, K. & Inokuchi, H. (1994). A tRNA-likestructure is present in 10Sa RNA, a small stableRNA from E. coli. Proc. Natl Acad. Sci. USA, 91,9223-9227.
12. Rudinger-Thirion, J., GiegeÂ, R. & Felden, B. (1999).Aminoacylated tmRNA from Escherichia coli interactswith prokaryotic elongation factor Tu. RNA, 5, 989-992.
13. Barends, S., Wower, J. & Kraal, B. (2000). Kineticparameters for tmRNA binding to alanyl-tRNAsynthetase and elongation factor Tu from Escherichiacoli. Biochemistry, 39, 2652-2658.
14. Karzai, A. W., Susskind, M. M. & Sauer, R. T.(1999). SmpB, a unique RNA-binding protein essen-
tial for the peptide-tagging activity of SsrA(tmRNA). EMBO J. 18, 3793-3799.
15. Wower, I. K., Zwieb, C. W., Guven, S. A. & Wower,J. (2000). Binding and cross-linking of tmRNA toribosomal protein S1, on and off the Escherichia coliribosome. EMBO J. 19, 6612-6621.
16. Subramanian, A. R. (1983). Structure and functionsof ribosomal protein S1. Prog. Nucl. Acid Res. Mol.Biol. 28, 101-142.
17. Nameki, N., Takdaki, T., Himeno, H. & Muto, A.(2000). Three of four pseudoknots in tmRNA areinterchangeable and are substitutable with single-stranded RNAs. FEBS Letters, 470, 345-349.
18. Boutorin, A. S., Clark, B. F. C., Ebel, J. P., Kruse,T. A., Petersen, H. U., Remy, P. & Vassilenko, S.(1981). A study of the interaction of Escherichia colielongation factor-Tu with aminoacyl-tRNAs by par-tial digestion with cobra venom ribonuclease. J. Mol.Biol. 152, 593-608.
19. Tamura, K., Asahara, H., Himeno, H., Hasegawa, T.& Shimizu, M. (1991). Identity elements of Escheri-chia coli tRNAAla. J. Mol. Recogn. 4, 129-132.
20. Felden, B., Hanawa, K., Atkins, J. F., Himeno, H.,Muto, A., Gesteland, R. F. et al. (1998). Presence andlocation of modi®ed nucleotides in Escherichia colitmRNA: structural mimicry with tRNA acceptorbranches. EMBO J. 17, 3188-3196.
21. Nameki, N., Felden, B., Atkins, J. F., Gesteland, R. F.,Himeno, H. & Muto, A. (1999). Functional andstructural analysis of a pseudoknot upstream of thetag-encoded sequence in E. coli tmRNA. J. Mol. Biol.286, 733-744.
22. Simos, G., Segref, A., Fasiolo, F., Hellmuth, K.,Shevchenko, A., Mann, M. & Hurt, E. C. (1996). Theyeast protein Arc1p binds to tRNA and functions asa cofactor for the methionyl- and glutamyl-tRNAsynthetases. EMBO J. 15, 5437-5448.
23. Wang, C. C. & Schimmel, P. (1999). Species barrierto RNA recognition overcome with non-speci®cRNA binding domains. J. Biol. Chem. 274, 16508-16512.
24. Park, S. G., Jung, K. H., Lee, J. S., Jo, Y. J., Motegi,H., Kim, S. & Shiba, K. (1999). Precursor ofpro-apoptotic cytokine modulates aminoacylationactivity of tRNA synthetase. J. Biol. Chem. 274,16673-16676.
25. Morales, A. J., Swairjo, M. A. & Schimmel, P. (1999).Structure-speci®c tRNA-binding protein from theextreme thermophile Aquifex aeolicus. EMBO J. 18,3475-3483.
26. Nissen, P., Kjeldgaard, M., Thirup, S., Polekhina, G.,Reshetnikova, L., Clark, B. F. C. & Nyborg, J. (1995).Crystal structure of the ternary complex of Phe-tRNA. Phe EF-Tu, and a GTP analog. Science, 270,1464-1472.
27. Nissen, P., Thirup, S., Kjeldgaard, M. & Nyborg, J.(1999). The crystal structure of Cys-tRNACys �EF-Tu �GDPNP reveals general and speci®c features inthe ternary complex and in tRNA. Struct. Fold. Des.7, 143-156.
28. Wikman, F. P., Siboska, G. E., Petersen, H. U. &Clark, B. F. C. (1982). The site of interaction ofaminoacyl-tRNA with elongation factor Tu. EMBO J.1, 1095-1100.
29. Pleiss, J. A., Wolfson, A. D. & Uhlenbeck, O. C.(2000). Mapping contacts between Escherichia colialanyl tRNA synthetase and 20 hydroxyls using acomplete tRNA molecule. Biochemistry, 39, 8250-8258.

SmpB as a Modulator of the tmRNA Acceptor Arm 21
30. Felden, B., Himeno, H., Muto, A., McCutcheon, J. P.,Atkins, J. F. & Gesteland, R. F. (1997). Probing thestructure of the Escherichia coli 10Sa RNA (tmRNA).RNA, 3, 89-103.
31. Zwieb, C., Guven, S. A., Wower, I. K. & Wower, J.(2001). Three-dimensional folding of the tRNA-likedomain of Escherichia coli tmRNA. Biochemistry, 40,9587-9595.
32. Shi, H. J. & Moore, P. B. (2000). The crystal structureof yeast phenylalanine tRNA at 1.93 AÊ ngstromresolution: a classic structure revisited. RNA, 6,1091-1105.
33. Koradi, R., Billeter, M. & WuÈ thrich, K. (1996).MOLMOL: a program for display and analysisof macromolecular structures. J. Mol. Graph. 14,51-55.
34. Boon, K., Vijgenboom, E., Madsen, L. V., Talens, A.,Kraal, B. & Bosch, L. (1992). Isolation and functionalanalysis of histidine-tagged elongation factor Tu.Eur. J. Biochem. 210, 177-183.
35. Ribas de Pouplana, L. & Schimmel, P. (1997). Recon-struction of quaternary structures of class II tRNAsynthetases by rational mutagenesis of a conserveddomain. Biochemistry, 36, 15041-15048.
36. Liu, J. C., Liu, M. & Horowitz, J. (1998). Recognitionof the universally conserved 30-CCA end of tRNAby elongation factor EF-Tu. RNA, 4, 639-646.
37. Olsthoorn, R. C., Mertens, S., Brederode, F. T. & Bol,J. F. (1999). A conformational switch at the 30 end ofa plant virus RNA regulates viral replication. EMBOJ. 18, 4856-4864.
38. Vorstenbosch, E. L., Potapov, A. P., de Graaf, J. M.& Kraal, B. (2000). The effect of mutations in EF-Tuon its af®nity for tRNA as measured by two noveland independent methods of general applicability.J. Biochem. Biophys. Methods, 42, 1-14.
39. Pingoud, A. & Urbanke, C. (1979). The determi-nation of binding parameters from protection exper-iments. A quantitative assay for ternary complexformation of elongation factor Tu, GTP and amino-acyl-tRNA. Anal. Biochem. 92, 123-127.
Edited by J. Doudna
(Received 13 June 2001; received in revised form 21 September 2001; accepted 24 September 2001)





























