Embed Size (px)
Citation preview

doi:10.1016/j.jmb.2004.12.013 J. Mol. Biol. (2005) 346, 761–772
Single Particle Analysis of Relaxed and ActivatedMuscle Thin Filaments
Alnoor Pirani1†, Chen Xu2†, Victoria Hatch1, Roger Craig3
Larry S. Tobacman4 and William Lehman1*
1Department of Physiology andBiophysics, Boston UniversitySchool of Medicine, 715 AlbanyStreet, Boston, MA 02118USA
2Rosenstiel Basic MedicalSciences Research CenterBrandeis University, 415 SouthStreet, Waltham, MA 02454USA
3Department of Cell BiologyUniversity of MassachusettsMedical School, 55 Lake AvenueNorth, Worcester, MA 01655USA
4Department of Physiology andBiophysics, University ofIllinois at Chicago, College ofMedicine, 835 South WolcottAvenue, Chicago, IL 60612USA
0022-2836/$ - see front matter q 2004 E
† A.P. and C.X. contributed equalAbbreviations used: B-state, block
Ca2C-induced, closed-state; M-stateopen-state; CH, calponin homologymicroscopy; IHRSR, iterative helicareconstruction; S1, myosin subfragm
E-mail address of the [email protected]
The movement of tropomyosin from actin’s outer to its inner domain playsa key role in sterically regulating muscle contraction. This movement, froma low Ca2C to a Ca2C-induced position has been directly demonstrated byelectron microscopy and helical reconstruction. Solution studies, however,suggest that tropomyosin oscillates dynamically between these positions atall Ca2C levels, and that it is the position of this equilibrium that iscontrolled by Ca2C. Helical reconstruction reveals only the averageposition of tropomyosin on the filament, and not information on the localdynamics of tropomyosin in any one Ca2C state. We have therefore usedsingle particle analysis to analyze short filament segments to reveal localvariations in tropomyosin behavior. Segments of Ca2C-free and Ca2C-treated thin filaments were sorted by cross-correlation to low and highCa2C models of the thin filament. Most segments from each data setproduced reconstructions matching those previously obtained by helicalreconstruction, showing low and high Ca2C tropomyosin positions for lowand high Ca2C filaments. However, w20% of segments from Ca2C-freefilaments fitted best to the high Ca2C model, yielding a corresponding highCa2C reconstruction. Conversely, w20% of segments from Ca2C-treatedfilaments fitted best to the low Ca2C model and produced a low Ca2C
reconstruction. Hence, tropomyosin position on actin is not fixed in eitherCa2C state. These findings provide direct structural evidence for theequilibration of tropomyosin position in both high and low Ca2C states,and for the concept that Ca2C controls the position of this equilibrium. Thisflexibility in the localization of tropomyosin may provide a means ofsterically regulating contraction at low energy cost.
q 2004 Elsevier Ltd. All rights reserved.
Keywords: actin; electron microscopy; 3D reconstruction; tropomyosin;troponin
*Corresponding authorIntroduction
Tropomyosin forms a continuous strand on thesurface of actin-based thin filaments that plays acrucial role in the regulation of muscle contraction.1
The connection between movement of tropomyosinon actin and a switching mechanism for turningcontraction on and off was first suggested by X-ray
lsevier Ltd. All rights reserve
ly to this work.ed-state; C-state,
, myosin-induced,; EM, electronl real space
ent 1.ing author:
diffraction studies on intact muscle fibers.2–4 Tropo-myosin can be resolved in electron micrographs ofthin filaments, and helical reconstruction of theseimages directly confirmed that it changes positionduring muscle activation.5–11 In relaxed muscle,tropomyosin covers myosin-crossbridge bindingsites on actin, which is thought to block cyclicactin–myosin interaction and consequently contrac-tion.5,7 During activation, tropomyosin moves awayfrom these sites in a stepwise fashion that isinduced first by Ca2C binding to troponin andthen by myosin binding to actin.7 Tropomyosinmovement and myosin binding are synergistic,which, in part, explains the cooperativity ofmuscle’s regulatory switching mechanism.8,12–14
Plasticity in the location of tropomyosin on actinappears to be an inherent part of the muscle
d.

762 Single Particle Analysis of Muscle Thin Filaments
regulatory system. Strong, stereo-specific inter-actions between actin and tropomyosin, demandingsignificant free energy expenditure to switch tropo-myosin position, do not appear to be responsible forthe association of the two proteins. In contrast,weak electrostatic interactions between chargedamino acids on actin and tropomyosin appear tolink the proteins together, accounting for the smallenergy cost needed to relocate tropomyosin on actinduring activation and relaxation.15 Despite therelatively weak association of tropomyosin andactin monomers, the binding of the tropomyosinstrand on actin filaments is appreciable.16,17 Sevenpseudo-repeats in the elongated tropomyosinmolecule interact quasi-equivalently with sevenadjacent actin monomers in each long-pitch helicalstrand of the filament, and successive tropomyosinmolecules are linked end-to-end along each strand(w26 tropomyosin molecules/mm filament).18–21
Hence the binding of the tropomyosin ensembleon the thin filament depends on multipleweak interactions, allowing the local position oftropomyosin to be easily perturbed by thermalenergy.
Tropomyosin is thought to be poised on actin inone or another position that can be easily per-turbed.8,22–25 In thin filaments of relaxed striatedmuscle, troponin-I of Ca2C-free troponin biasestropomyosin away from the “closed”, calcium-induced (C-state) position on the inner domain ofactin toward the “blocking” (B-state) position on theouter domain.26 During activation, Ca2C releasesthis constraint, and tropomyosin assumes the Ca2C-induced position, exposing most but not all of themyosin-binding site. The initial binding of myosinheads then biases tropomyosin further from theblocking configuration toward the myosin-inducedopen state (M-state) position, and the myosin-binding site completely opens, resulting in fullactivation.7,8 Biochemical studies suggest that theposition of tropomyosin is not completely fixed inany of these configurational states, and that theroles of troponin and myosin are to alter smallenergy differences between states, hence favoringone or another configuration. Measurements of theinfluence of tropomyosin and troponin-tropo-myosin on the kinetics of the actin–myosininteractions are consistent with this scheme.22,23
Thus tropomyosin may not be statically arrangedon actin in any one regulatory state.
Devising a structural approach to address thequestion of local variation in tropomyosin positionin a given Ca2C-state is challenging. Electronmicroscopy (EM) of filaments captures static“snapshots” of transient events. Three-dimensional(3D) helical reconstruction defines the averageposition, over a span of about 50 to 100 actinmonomers, of tropomyosin so captured.5–9,27,28 Themethod cannot identify different local positions,and thus, if tropomyosin were in a dynamicequilibrium between positions, favoring but neverbeing truly fixed in one position, averaging wouldmask the variability. Iterative helical real space
reconstruction (IHRSR)) of short thin filamentsegments (wseven actin molecules and onetropomyosin molecule long) is better suited todetermining local differences in configuration.29 Inthe current study, we divided EM images of thinfilaments into segments, which we treated assingle particles. The segments were then sorted bycross-correlation to thin filament models havingdifferent tropomyosin positions. We found that themajority of segments of Ca2C-free thin filamentsfitted best to B-state models and produced areconstruction with tropomyosin in the B-stateposition on the outer domain of actin. However, asignificant subpopulation of segments fitted moreprecisely to C-state models and yielded a recon-struction with tropomyosin in the C-state position.Conversely, most segments from Ca2C-treated thinfilaments fitted best to C-state models, but a subsetcorresponded better to B-state models. Thus theregulatory mechanism controlling musclecontractility appears to influence the relativedistribution of tropomyosin positions on thinfilaments, which are not, as often depictedschematically, fixed rigidly in switched-on orswitched-off states.
Results and Discussion
Electron microscopy and preliminary steps inimage processing
Rabbit skeletal muscle F-actin was mixed withpurified tropomyosin and troponin under con-ditions that saturate filaments with these regulatoryproteins (see Materials and Methods). Negativestaining of these filaments showed typical actinsubstructure (Figure 1). Troponin densities werealso observed, repeating at 40 nm intervals, inregister on opposite sides of the filaments. Inaddition, elongated strands, known to representtropomyosin, were also occasionally observed inelectron micrographs (see Moody et al.10 and Craigand Lehman11).
The polarity of the filaments was evident in someof the EM images, but these cases were rare.Filament polarity is easily determined, however,by helical reconstruction, which was carried out onall thin filaments processed. Sometimes polaritywas also checked by cross-correlation of relativelylong filament stretches (R70 nm) to models of actinor actin-tropomyosin. Determinations made by thetwo methods were always consistent. Polarityassessment (of short filament segments (30–40 nm)) is also part of the IHRSR protocol. Inthis case, about 90% of the segments from eachfilament exhibited polarity correctly, and thosesegments that did not were excluded in thereconstructions described below (and seeEgelman29).

Figure 1. Electron micrographs of F-actin-tropomyosin-troponin complexes. Negative staining of (a) Ca2C-freeand (b) Ca2C-treated filaments; note the laterally orienteddensities repeating at 40 nm intervals that represent theglobular end of troponin and the obliquely orientedstrands that are characteristic of tropomyosin (several ofeach are indicated by arrowheads and arrows, respect-ively). All filaments shown have their pointed end at thetop. The scale bar represents 50 nm. Motility assaysperformed on the F-actin-tropomyosin-troponin com-plexes used showed that the filaments were completelydependent on Ca2C for their movement by myosinsubfragment 1 (data not shown).
Single Particle Analysis of Muscle Thin Filaments 763
IHRSR reconstruction of Ca2C-free andCa2C-treated thin filaments
Ca2C-free filaments
Approximately 20,000, 30–40 nm long, segmentsfrom 408 Ca2C-free F-actin-troponin-tropomyosinfilaments were fitted by cross-correlation todifferent rotational views of various thin filamentmodels (see Materials and Methods for details). Aninitial reconstruction was then generated by back-projection and was found to be virtually the same asthose obtained previously by helical reconstruc-tion.5–8 Tropomyosin localized in the B-stateposition, on the inner edge of the outer domain ofsuccessive actin monomers along the filament(Figures 2, 3(a) and 4(a)). The location of tropo-myosin was found to be independent of tropo-
myosin’s position in the reference models: i.e. if thesegments were aligned to either a helical recon-struction of Ca2C-free or one of Ca2C-treated thinfilaments, the new reconstructions obtained wereindistinguishable from each other. Clearly, the actinportion of the data, which carries the greatestamount of density information, dominated thealignment process. This conclusion is supportedby showing that when F-actin models (no troponin-tropomyosin) were used as initial references toalign segments, reconstructions matching thoseabove were produced, again with tropomyosin inthe B-state position. Obtaining the same results,regardless of the position or presence of tropo-myosin in the initial reference, indicates that thestarting models did not bias the analysis.
This initial reconstruction generated was thenused as a new reference model in a second round ofreconstruction and the process repeated up to 40times (IHRSR). Iteration had little or no impact ontropomyosin and actin in the reconstructions,demonstrating the quality of the initial helicalreconstructions used. In all cases, tropomyosinlocalized to the same site on the outer domain ofactin (a helical reconstruction of the data wasemployed as the initial reference for the IHRSRreconstructions of Ca2C-free segments shown inFigures 2, 3(a) and 4(a); the definition of tropo-myosin on actin detected by the IHRSR method canbe best appreciated by referring to Figure 2). Thispattern of results was also obtained when atomicmodels of actin or actin-tropomyosin (previouslygenerated by fitting crystal structures of G-actin andtropomyosin to X-ray fiber diagrams of filaments,15
i.e. not helical reconstructions) were used as initialreferences to align filament segments. Again, thealigned segments produced B-state reconstructionsthat were independent of the presence or position oftropomyosin in the reference.
Ca2C-treated filaments
Approximately 7000 segments from 138 Ca2C-treated filaments were analyzed in a similar way.IHRSR of these segments always yielded a recon-struction that matched the helical reconstruction ofthese filaments. Here, tropomyosin was localized inthe C-state position on the outer aspect of the innerdomain of successive actin monomers (Figures 3(b)and 4(b)). Again this configuration of tropomyosinon actin persisted through all rounds of IHRSRiteration. Comparison of IHRSR reconstructions ofCa2C-free and Ca2C-treated thin filaments indi-cated that the average azimuthal movement oftropomyosin on actin is about 20 to 258, as wasfound previously.5,7
Sorting filament segments into classes
In the above reconstructions, thin filament seg-ments were aligned to a single reference model inorder to produce a reconstruction from a particulardata set. We also used the IHRSR method to

Figure 2. Surface views of theIHRSR reconstruction of Ca2C-freethin filaments showing the pos-ition of tropomyosin on actin atdifferent density thresholds.IHRSR was carried out on 18,00942 nm long segments derived fromfour different preparations ofEGTA-treated actin-tropomyosin-troponin (virtually identical resultswere obtained when 19,428 28 nmlong segments, were studied). TheIHRSR reconstructions showcharacteristic bilobed actin thatcan be divided into four subdo-mains (numbered 1 to 4 on oneactin monomer in (a)). In addition,longitudinally continuous tropo-myosin strands can be seen on theinner edge of the outer domain ofactin, interacting with subdomain1 and bridging over subdomain 2of each successive actin monomeralong the filament. Reconstruc-tions were contoured at 1.0, 2.4and 4.0 s above the mean densityamplitude, as indicated. The con-tact point of tropomyosin with
subdomain 1 of actin is especially evident at high threshold in (a) (black arrowheads). The lack of contact oftropomyosin with subdomains 3 and 4 on the actin inner domain is also apparent (white arrows). All reconstructionsshown here are oriented with the pointed end of the filament facing up. The resolution of the reconstruction was 18 Aaccording to the 0.5 Fourier shell correlation criterion. Surface rendering here (and in Figure 3) was done using theprogram WEB.45
764 Single Particle Analysis of Muscle Thin Filaments
determine whether the data would sort intodifferent possible classes, by cross-correlating totwo or more reference models. Our aim was todecide whether filaments in one particular Ca2C-state consisted of segments with just one or morethan one structure. When segments of Ca2C-freefilaments were run against B and C-state recon-structions as references (using either helical orIHRSR models), the majority fitted best to theB-state model and produced a new reconstructionindistinguishable from those described above(Figures 3(c) and 4(c)). However, a significantsubpopulation of segments (w22% of the total)clearly fitted best to the C-state model and yielded areconstruction with tropomyosin in the C-stateposition (Figures 3(d) and 4(d), Table 1). Conversely,when segments of Ca2C-treated filaments weresorted in this way, most segments aligned best tothe C-state model (Figures 3(f) and 4(f)) and yieldedreconstructions indistinguishable from this model.However, a significant percentage (w20% of thedata) fitted best to the B-state model (Figures 3(e)and 4(e)), and produced a B-state reconstruction.Thus the position of tropomyosin in models of thethin filament makes a difference when segments areallowed to select between B or C-state references.
Tabulation of the sorted data revealed thatoccasionally a consecutive string of the Ca2C-freesegments, corresponding to about 120 nm to 160 nmin the filament, aligned best to the C-state model,
and conversely similar stretches of Ca2C-treatedfilaments sorted to B-state models. These resultssuggested that interactions between successiveactin and tropomyosin molecules could persistover relatively long filament distances even whenthe configurational state was not favored. Thesefilament stretches were long enough to carry outhelical reconstruction, providing the opportunity tocorroborate the tropomyosin position assigned byIHRSR. Helical reconstruction, performed on asmall sample of these stretches from Ca2C-freefilaments, showed that in most of the cases tested(w75%), tropomyosin was positioned in the C-state(Figure 5(a)). In other cases, the position oftropomyosin was ambiguous, possibly because thedata may still have represented a mixture oftropomyosin positions (Figure 5(b)).
IHRSR was also carried out on S1-decorated thinfilaments (Figure 4(i)). This localized tropomyosinin an M-state position differing azimuthally fromthe C-state position by 108 to 158, as previouslyfound by helical reconstruction7 (Figure 4(j)).A model of actin-tropomyosin was then built forthe M-state (with the S1 removed). This, togetherwith C and B-state models, was used to furtherclassify the positions of tropomyosin in Ca2C-activated filaments to test if a mixture of B, C andM-state locations could be detected. Segments ofCa2C-treated filaments clearly sorted into threecategories based on their cross-correlation to the
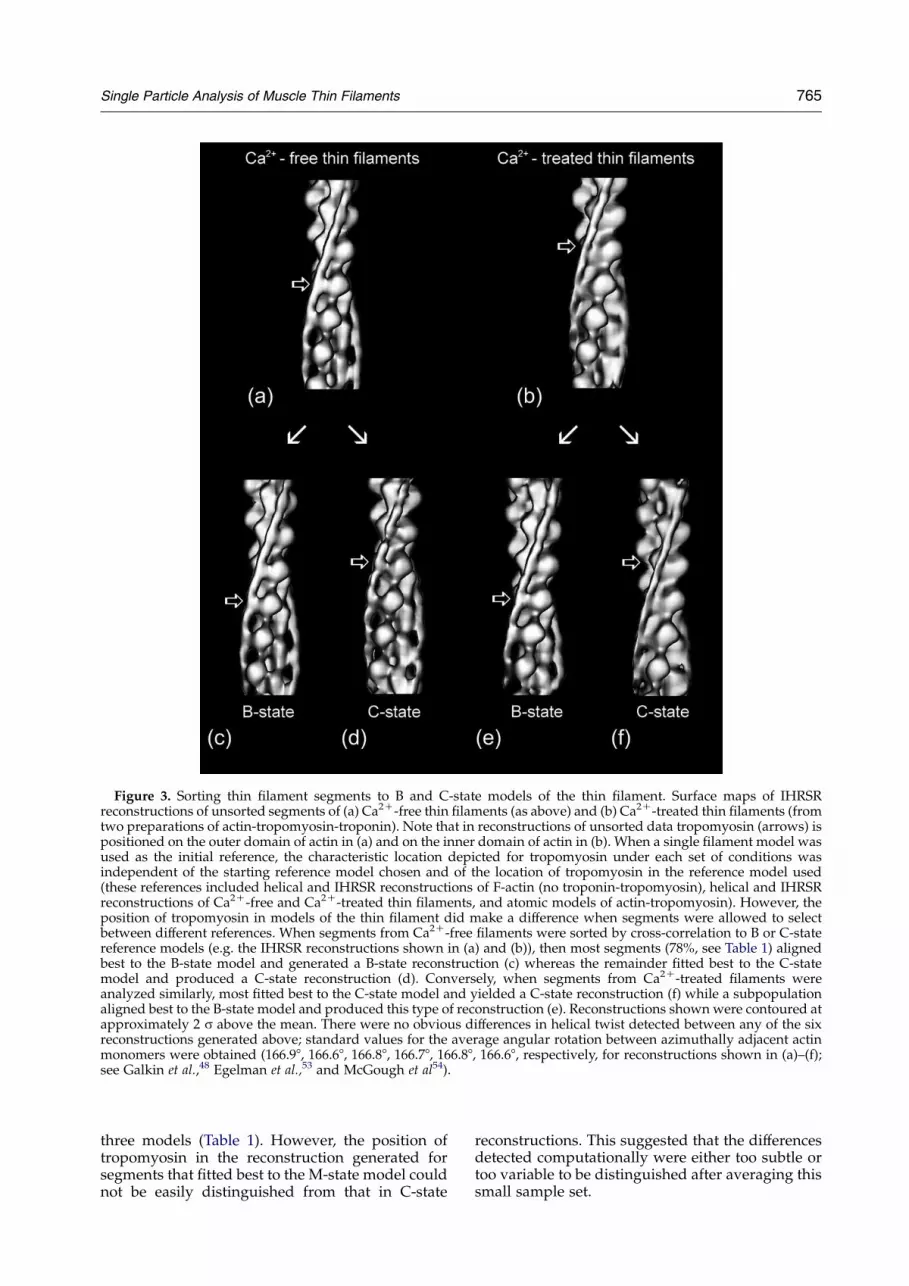
Figure 3. Sorting thin filament segments to B and C-state models of the thin filament. Surface maps of IHRSRreconstructions of unsorted segments of (a) Ca2C-free thin filaments (as above) and (b) Ca2C-treated thin filaments (fromtwo preparations of actin-tropomyosin-troponin). Note that in reconstructions of unsorted data tropomyosin (arrows) ispositioned on the outer domain of actin in (a) and on the inner domain of actin in (b). When a single filament model wasused as the initial reference, the characteristic location depicted for tropomyosin under each set of conditions wasindependent of the starting reference model chosen and of the location of tropomyosin in the reference model used(these references included helical and IHRSR reconstructions of F-actin (no troponin-tropomyosin), helical and IHRSRreconstructions of Ca2C-free and Ca2C-treated thin filaments, and atomic models of actin-tropomyosin). However, theposition of tropomyosin in models of the thin filament did make a difference when segments were allowed to selectbetween different references. When segments from Ca2C-free filaments were sorted by cross-correlation to B or C-statereference models (e.g. the IHRSR reconstructions shown in (a) and (b)), then most segments (78%, see Table 1) alignedbest to the B-state model and generated a B-state reconstruction (c) whereas the remainder fitted best to the C-statemodel and produced a C-state reconstruction (d). Conversely, when segments from Ca2C-treated filaments wereanalyzed similarly, most fitted best to the C-state model and yielded a C-state reconstruction (f) while a subpopulationaligned best to the B-state model and produced this type of reconstruction (e). Reconstructions shown were contoured atapproximately 2 s above the mean. There were no obvious differences in helical twist detected between any of the sixreconstructions generated above; standard values for the average angular rotation between azimuthally adjacent actinmonomers were obtained (166.98, 166.68, 166.88, 166.78, 166.88, 166.68, respectively, for reconstructions shown in (a)–(f);see Galkin et al.,48 Egelman et al.,53 and McGough et al54).
Single Particle Analysis of Muscle Thin Filaments 765
three models (Table 1). However, the position oftropomyosin in the reconstruction generated forsegments that fitted best to the M-state model couldnot be easily distinguished from that in C-state
reconstructions. This suggested that the differencesdetected computationally were either too subtle ortoo variable to be distinguished after averaging thissmall sample set.

Figure 4. Transverse sections (z-sections) through maps of IHRSR reconstructions: z-sections of unsorted data of (a)Ca2C-free and (b) Ca2C-treated actin-tropomyosin-troponin filaments; (c)–(f) z-sections of data sorted according to cross-correlation to B and C-state models, as in Figure 3. Actin subdomains numbered on the section in (a); (since azimuthallyopposite actin monomers along F-actin are staggered, transverse sections through subdomains 1 and 3 of one actinmonomer slice through subdomains 2 and 4 of the adjacent monomer). Black outlined arrows indicate tropomyosinpositions. Note the B-state association of tropomyosin with the outer domain of actin in (a), (c), and (e), and the C-statebinding to the inner domain in (b), (d) and (f). Note also that the C-state tropomyosin position for the Ca2C-data ((b) and(f)) is slightly further away from the outer actin domain than it is in (d) for the subpopulation of segments of Ca2C-freefilaments that fitted best to the C-state model. This is also apparent in the surface view (Figure 3(d)) and suggests thatparts of some segments in the latter group had mixed tropomyosin configurations. Transverse sections through IHRSRreconstructions of (g) actin-tropomyosin (no troponin) and (h) actin-tropomyosin-troponin-T153. Note that whiletropomyosin is in a B-state configuration in both (g) and (h), its position is better defined in the presence of the troponin-T fragment (h). (i) IHRSR reconstruction of S1-decorated thin filaments (original EM data from Vibert et al.7) Note thatafter S1 binding, M-state tropomyosin moves w108 over the actin inner domain away from the C-state; this shiftin tropomyosin position is especially evident by comparing maps of B and M-states by superimposition ( j). The S1-induced movement revealed by IHRSR is comparable to that originally observed by helical reconstruction,7 highlightingthe general consistency of the two techniques.
766 Single Particle Analysis of Muscle Thin Filaments
The troponin-binding site
In the above studies, no attempt was made toaccount for the contribution of troponin to our
Table 1. Percentage distribution of segments sorting todifferent regulatory states
Sample B-state C-state M-state
Ca2C-free filaments 78 22 –Ca2C-treated filaments 20 68 12
reconstructions or to align the troponin densitiesin segments to each other. Unlike tropomyosin,troponin is not continuously or uniformly distri-buted along thin filaments. The troponin complexcontains a globular head group (also called the core-domain, and comprising troponin-I, troponin-C,and the C-terminal domain of troponin-T) and anelongated tail (the N-terminal domains of troponinT).30 The elongated troponin-T tail is thought to lieparallel with tropomyosin and may contribute to itsmass observed in reconstructions (although wehave never with any certainty been able to

Figure 5. Helical reconstruction of C-state regions ofCa2C-free thin filaments. Helical reconstruction wasperformed on 120 to 160 nm-long filament stretches ofcontiguous segments found to be in the C-state by IHRSR;(a) z-section map representing an average of three helicalreconstructions showing tropomyosin (arrows) clearly inthe C-state position on the inner domain of actin,(b) another map showing a more ambiguous tropomyosinlocation.
Figure 6. Aligning filament segments to short models ofF-actin-tropomyosin-troponin. Ca2C-free filaments weredivided into 10.5 nm segments which were sorted bycross-correlation to two reference models of correspond-ing length; the B-state, actin-tropomyosin reconstructionwas used as one reference ((a)) and an identical referencewith an extra mass placed over subdomain-1 of adjacentactin monomers was used as the second ((b)); in the caseshown, the added density was derived from the CHdomain of spectrin,55 low pass filtered to 25 A resolutionto simulate an ellipsoidal mass. Reconstruction of thesegments that fitted best to the second reference revealedan extra density associated with the extreme outerdomain of actin (open arrow) adjacent to tropomyosin(narrow arrow), which was missing from the reconstruc-tion of those segments that sorted to the actin-tropomyo-sin model. When the reporter mass in the reference wasmoved to other sites on actin, little or no extra density wasfound in the reconstructions generated (not shown).Substituting mock constant density spherical masses upto about double the size of the CH domain gave similarresults and did not yield appreciable extra mass in thereconstructions. Surface rendering here (and in Figure 7)was done using the program Chimera.56
Single Particle Analysis of Muscle Thin Filaments 767
determine this). Troponin-I in Ca2C-free troponin islinked to every seventh actin monomer along eachof the two strands of the actin helix.1 Thisinteraction is thought to take place over actinsubdomains 1 and 2 in regions distinct from thepositions that we have determined for tropomyosinbinding, but this conclusion is not certain.31–33
As a first attempt to demarcate and align troponinin Ca2C-free filaments, very short filament seg-ments (10.5 nm long) were sorted between twomodels. Actin-tropomyosin (also 10.5 nm long) wasused as the first model and the same structure, witha relatively small ellipsoidal or spherical mass (withsizes ranging between w10 kDa and 20 kDa) addedon each of two adjacent actins in the model, as thesecond (Figure 6). Parts of filaments withouttroponin heads (six-sevenths of the data) weretherefore expected to sort to the first model andthose with troponin heads, to the latter, providedthe extra mass was appropriately placed. When theextra mass in the model was centered over N-terminal amino acids on subdomain 1 of actin, acontact point between troponin-I and actin,34 thesegments aligning to this model generated areconstruction with an extra density lying next tothe tropomyosin strands (Figure 6). The extra massdetected presumably represented protein close to orat the troponin binding site, and most likely part ofthe head group of the troponin complex. When thereporter was moved to other parts of the model (e.g.to subdomains 2 and 3), little or no extra densitywas detected in the reconstructions (data notshown). These results therefore agreed with othersthat troponin binds to the extreme outer domain ofactin, at a site separated from the tropomyosinbinding site.31 Since the troponin and tropomyosindensities were distinct from each other, the resultsalso suggest that globular troponin densities makelittle or no contribution to the tropomyosin strandsdetermined above (see Squire & Morris35). This
preliminary determination of the troponin-bindingsite proved useful in generating more complex thinfilament models.
The conclusion that troponin on thin filamentscontributes minimally to densities ascribed totropomyosin11 was tested further. IHRSR recon-structions of actin-tropomyosin, complexed withthe N-terminal fragment of troponin-T (TnT1-153,which lacks troponin’s globular core domain36),clearly demonstrated tropomyosin density compar-able to that seen in filaments containing wholetroponin, and also showed that this tropomyosinwas in the B-state position (Figure 4(h)), aspreviously shown by helical reconstruction.37

768 Single Particle Analysis of Muscle Thin Filaments
This confirmed that the globular core domain oftroponin is unlikely to contribute significantly to thetropomyosin density observed in the B-state. More-over, since the troponin-T fragment used was onlyone-quarter the mass of tropomyosin, the tropo-myosin density observed is unlikely to includesignificant contribution from the troponin taildomain. Troponin-T may be responsible for defin-ing tropomyosin strands clearly (presumably bystabilizing it on filaments), as tropomyosin was notas well localized in actin-tropomyosin controlslacking any troponin (Figure 4(g)).
Figure 7. Reconstructions of filament segments alignedto long models of F-actin-tropomyosin-troponin. Ca2C-free filaments were divided into 28 nm segments, whichwere sorted by cross-correlation to ten actin monomerlong actin-tropomyosin reference models with troponinplaced on two azimuthally adjacent actin monomers (andnot the others). In the model used, crystal structures ofactin, tropomyosin and the core-domain of troponin wereused to simulate the thin filament and low pass filtered to25 A resolution. The actin-tropomyosin-troponin modelwas rotated and translated to represent different views ofa troponin pair along the entire length of the filamentmodel; these were then used to align filament segments,accounting for troponin in addition to actin and tropo-myosin. (a) and (b) Two orthogonal views of thereconstruction generated; note the triskelion shapeddensity attributable to troponin (open arrows) and thecontinuous tropomyosin (narrow arrows) on the actinmonomers on either side of the troponin density. Thethree lobes of the troponin density may correspond to thetroponin-IT arm and the two globular (N and C) domainsof troponin-C that are observed in the crystal structure ofthe core-domain.52
The tropomyosin molecule binds to actinmonomers uniformly along its length
Narita et al.32 proposed that the tropomyosinmolecule is kinked when associated with actin, andthat this effect is most apparent in Ca2C-free thinfilaments. They suggested that the N andC-terminal halves of each tropomyosin occupieddifferent sites on actin monomers, the kink betweenthe two halves being formed by a joint formedbetween troponin and a particularly flexibledomain on tropomyosin. Such effects, if present,would have been averaged out in our previouslypublished helical reconstructions. Averaging can beavoided, however, using the single particleapproach, if the segments are aligned with respectto troponin molecules and if they are of sufficientlength to follow tropomyosin on either side oftroponin. A model of F-actin-tropomyosin wasagain generated to simulate thin filaments, herewith a more realistically sized extra density (basedon troponin crystal structures) placed on sub-domain 1 of each of two azimuthally adjacentactin monomers but not on the others over a tenactin monomer long stretch (i.e. a 28 nm long modelwith five actin monomers on each long pitch strandof F-actin, with troponin contacting just two of themonomers). The model was rotated and translatedto provide filament projections with the troponinstructures starting on the first and second actinmonomers and ending on the last two monomers.These were used to align 28 nm long segments ofthe Ca2C-free thin filaments to take into accounttroponin densities in addition to those of actin andtropomyosin (while the models and segments weresufficiently short to insure that they contained onlyone troponin for each tropomyosin strand). Recon-structions for each axial position of troponin weregenerated and brought to a common origin to alignthe troponin pairs. The process was repeated withthe troponin density thus detected now substitutedfor the corresponding mass used in the modelfilament of the first round of alignment. Newreconstructions were produced showing densitiesattributed to troponin on the two central actinmonomers of the ten actin long stretch togetherwith longitudinally continuous tropomyosinstrands on actin (Figure 7). Even though segmentswere aligned to models of filaments with discretelypositioned troponin (as by Narita et al.32) the
tropomyosin in the reconstructions was not dis-jointed but localized in a consistent position alongthe inner edge of subdomain 1 of successive actinmonomers, on either side of the troponin density.We therefore find no evidence for a non-uniform,kinked conformation of tropomyosin proposed byNarita et al.32 We note that the tropomyosin densityover each actin monomer in our reconstruction isobvious and that each of the actin monomers in ourmaps is well defined.
In the studies reported here, the alignment oftroponin was used as a tool to study the associationof tropomyosin on thin filaments. Examining thestructure of troponin in more detail is planned,including attempts to dock troponin crystal struc-tures into densities contained in the reconstructions.

Single Particle Analysis of Muscle Thin Filaments 769
Conclusion
We have demonstrated that single particleanalysis can be employed to determine the positionof tropomyosin on actin. Using IHRSR, we con-firmed previous observations made by helicalreconstruction that had defined the averageposition of tropomyosin in B, C and M-states overrelatively long stretches of thin filaments.5–9,27 Ournew results support the three-state steric schemedescribing muscle regulation, as previouslyenvisioned.7,8 Examining short filament lengths byIHRSR offered a better chance to determine the localdistribution of tropomyosin positions on actin andto examine the three-state model in further detail.Multi-reference cross-correlation of our datarevealed that tropomyosin is not confined to onefixed location in either of the regulatory statestested, even though a particular configuration wasfavored. The demonstration that parts of filamentsmaintained under relaxing conditions are in theC-state and, conversely, that parts treated withCa2C are in the B-state, is consistent with the viewthat the positions of tropomyosin on actin are indynamic equilibrium and that transitions betweenregulatory states presumably are rapid.22,23,38 Thissuggests that thin filament regulation is governedby the statistical–mechanical behavior of its com-ponents.23,39,40 Our view is that the tropomyosinmolecule is not kinked, but that the tropomyosinstrand, formed by end-to-end molecular linkage, issemi-flexible. Thus, thermal energy can cause localfluctuations in tropomyosin position on actin, whiletroponin, Ca2C and myosin perturb the distributionof positions to define an average regulatoryconfiguration governing muscle activity. We expectthat strong myosin head interaction will tend torestrict the fluctuation of tropomyosin on actin inactivated muscles.
Most standard presentations of steric regulationof muscle contraction depict tropomyosin strandsas rigid cables repositioning themselves over themyosin-binding sites on actin in an all-or-nonefashion. Our current work demonstrates that thisview is an over-simplification. Our objective now isto develop robust atomic models of the thinfilament that include troponin in addition totropomyosin, and to characterize the dynamics ofthe entire system.
Materials and Methods
Protein preparation and electron microscopy
Rabbit skeletal muscle F-actin, and bovine cardiactropomyosin and troponin were purified by standardmethods.41,42 Recombinant troponin-T153 consisting of theN-terminal 153 residues of cardiac troponin-T wasprepared as described.36 Thin filaments were reconsti-tuted by mixing F-actin (24 mM) first with tropomyosin(7.5 mM) and then with troponin (7.5 mM) in a solutionconsisting of 100 mM NaCl, 3 mM MgCl2, 0.2 mM EGTA,1 mM NaN3, 5 mM Pipes/5 mM sodium phosphate
buffer (pH 7.1) with or without CaCl2 added to 0.4 mM.Filaments were allowed to incubate for five to ten minutesat room temperature (w25 8C) before making a 20-folddilution with additional buffer and immediately applyingto carbon-coated electron microscope grids. Samples werenegatively stained with 1% (w/v) uranyl acetate andgrids dried at 80% relative humidity to aid in spreadingthe stain.10 Electron micrograph images of decoratedfilaments were recorded on a Philips CM120 electronmicroscope at 60,000! magnification under low doseconditions (w12 eK/A2) at a defocus of 0.5 mm.
Image reconstruction
Helical reconstruction
Micrographs were digitized using a Zeiss SCAI scannerto yield a pixel size corresponding to 0.7 nm in thefilaments. Well-preserved regions of the filaments wereselected, straightened and boxed as previously.43,44
Helical reconstruction was carried out by standardmethods28 as described.7,8
Single particle analysis
IHRSR was carried out on the digitized images asdescribed by Egelman.29 Filaments were boxed anddivided into 42 nm squares with a 6 nm interval betweensuccessive “cuts” along filaments so that there was anoverlap of 38 nm between adjacent segments. In general,helical reconstructions were used as initial referencemodels for the first round of IHRSR. Filament segmentswere fitted to different rotational views of the referencerepresented by 90 2D projections of the model turned 3608in 48 intervals. Cross-correlation and alignment wereperformed between the boxed filament segments and theprojections using the AP NQ routine in the SPIDER suiteof programs (System for Processing Image Data fromElectron Microscopy and Related Fields.45) Segmentsaligning with incorrect polarity or with in-plane rotationsof greater than G48 from their preset vertical orientationwere discarded; generally this amounted to about 10% ofthe total number of segments tested. The remainingsegments were back-projected using the BP 3F routine inSPIDER and used to generate an initial reconstruction.A search for helical symmetry was performed and thenapplied to the reconstruction to yield a refined helicallysymmetric volume.29 The symmetrized reconstructionwas used as a reference model for the next round ofalignment and image reconstruction.29 This process wasiterated until no changes were seen from one round to thenext. This usually took two or three rounds, but 20 to 40rounds were routinely used. Resolution was determinedusing the 0.5 criterion for Fourier shell correlation of datathat had been split into two equal groups and com-pared.46,47
The IHRSR method is also well suited for sortingfilament segments into subpopulations that displaydifferent modes of binding protein attachment and/orvariable actin–actin interaction or conformation.48 In thestudies presented here, two or more actin-tropomyosinreference models were used to categorize and separatedata by cross-correlation. The sorted data were used togenerate respective reconstructions by back-projection asabove.
Alignment of troponin
To localize the troponin-binding site on actin, filament

770 Single Particle Analysis of Muscle Thin Filaments
segments were aligned to references that contained extradensities to simulate troponin. In the models made(Figure 6), the B-state reconstruction of actin-tropomyosingenerated by IHRSR was truncated to a length of 10.5 nmand modified by addition of density on the twoazimuthally adjacent actin monomers. Either the crystalstructure of the CH-domain of spectrin (PDB databaseentry 1AA2) (which when reduced to 25 A resolution49,50
formed a uniform ellipsoidal mass) or constant densityspheres generated by SPIDER45 served as “reporters” fortroponin in the models. To separate short, 10.5 nmsegments with troponin from those without, an unmodi-fied IHRSR reconstruction of actin-tropomyosin served asa second reference (see Figure 6). Projections of these tworeferences representing 90 rotational views were used forcross-correlation and alignment of the 10.5 nm longfilament segments. The data were sorted and back-projected to generate reconstructions of actin-tropomyosin with and without densities attributableto troponin. The new reconstructions were then usedas references and the process was iterated up to tentimes.
Troponin–tropomyosin interaction on actin wasexamined further, making use of the contact site oftroponin on actin determined above to now locate a morerealistic model of troponin on a longer stretch of actin.The atomic structure of F-actin51 and crystal structures oftropomyosin19 and the troponin core domain (i.e. thetroponin globular head52) were used to generate a newmodel of troponin on the thin filament. Here, a tenmonomer long atomic model of the F-actin (i.e. with fourto five complete actin monomers on either long pitchhelix) was manually aligned to the B-state IHRSRreconstruction and to the preliminary reconstruction ofactin-tropomyosin-troponin generated above. The recon-structions were then used as templates to position thecrystal structure of tropomyosin, again manipulatedmanually to fit to the tropomyosin density, with troponinplaced on each of two azimuthally opposite actins (butnot on the others of the model). The axial position chosenfor the crystal structure of troponin along actin wascritical in making models that were effective in aligningthe troponin in filament segments, but the rotation oftroponin picked did not have as much impact. The entiremodel (derived from the crystal structures of actin,tropomyosin and troponin) was low-pass filtered to25 A resolution.49,50 Eight additional reference modelswere generated from this initial model where thetroponin pair occupied eight different equivalent bindingpositions on the other actin monomers of the tenmonomer long model. A total of 90 projections of eachyielded 810 reference images, which were then used toalign the data set. Projections of models of actin-tropomyosin (no troponin) were used to select outsegments that did not show troponin: 28 nm long thinfilament segments (i.e. representing about ten actinmonomers) were used for the cross-correlation. Thislength was short enough to ensure that no segment wouldcontain more than a single pair of globular troponindensities, while sufficiently long to delineate tropomyosinposition on actin at a distance from troponin. Reconstruc-tions corresponding to the nine models were generated byback-projection, brought to a common origin, anddisplayed so that the troponin pair was placed at thecenter of the filament. Helical symmetrization of IHRSRis not used here; otherwise, this refinement wouldsmear out the troponin. The reconstruction generatedwas then used as a new reference model and the processiterated.
Acknowledgements
This study was supported by NIH grantsHL36153 (to W.L.), AR34711 (to R.C.) andHL38834 (to L.S.T.); EM was carried out in theCore Electron Microscope Facility of the Universityof Massachusetts Medical School, supported byNIH Shared Instrumentation grant RR08426 (toR.C.) and Diabetes Endocrinology Research Center(University of Massachusetts) grant DK32520. Wethank Dr E. H. Egelman for providing programsneeded to perform IHRSR reconstruction.
References
1. Gordon, A. M., Homsher, E. & Regnier, M. (2000).Regulation of contraction in striated muscle. Physiol.Rev. 80, 853–924.
2. Huxley, H. E. (1972). Structural changes in actin- andmyosin-containing filaments during contraction. ColdSpring Harbor Symp. Quant. Biol. 37, 361–376.
3. Haselgrove, J. C. (1972). X-ray evidence for aconformational change in actin-containing filamentsof vertebrate striated muscle. Cold Spring HarborSymp. Quant. Biol. 37, 341–352.
4. Parry, D. A. D. & Squire, J. M. (1973). Structural role oftropomyosin in muscle regulation, analysis of X-raypatterns from relaxed and contracting muscles. J. Mol.Biol. 75, 33–55.
5. Lehman, W., Craig, R. & Vibert, P. (1994). Ca2C-induced tropomyosin movement in Limulus thinfilaments revealed by three-dimensional reconstruc-tion. Nature, 368, 65–67.
6. Lehman, W., Vibert, P., Uman, P. & Craig, R. (1995).Steric-blocking by tropomyosin visualized in relaxedvertebrate muscle thin filaments. J. Mol. Biol. 251,191–196.
7. Vibert, P., Craig, R. & Lehman, W. (1997). Steric-modelfor activation of muscle thin filaments. J. Mol. Biol.266, 8–14.
8. Lehman, W., Hatch, V., Korman, V., Rosol, M.,Thomas, L., Maytum, R. et al. (2000). Tropomyosinand actin isoforms modulate the localization oftropomyosin strands on actin filaments. J. Mol. Biol.302, 593–606.
9. Xu, C., Craig, R., Tobacman, L., Horowitz, R. &Lehman, W. (1999). Tropomyosin positions in regu-lated thin filaments revealed by cryoelectronmicroscopy. Biophys. J. 77, 985–992.
10. Moody, C., Lehman, W. & Craig, R. (1990). Caldesmonand the structure of vertebrate smooth muscle thinfilaments: electron microscopy of isolated thinfilaments. J. Muscle Res. Cell Motil. 11, 176–185.
11. Craig, R. & Lehman, W. (2002). The ultrastructuralbasis of actin filament regulation. In Results andProblems in Cell Differentiation: Molecular Interactionsof Actin (dos Remedios, C. G. & Thomas, D. D., eds),vol. 36, pp. 149–169, Springer Verlag, Heidelberg.
12. Landis, C. A., Bobkova, A., Homsher, E. & Tobacman,L. S. (1997). The active state of the thin filament isdestabilized by an internal deletion in tropomyosin.J. Biol. Chem. 272, 14051–14056.
13. Rosol, M., Lehman, W., Craig, R., Landis, C., Butters,C. & Tobacman, L. S. (2000). Three-dimensionalreconstruction of thin filaments containing mutanttropomyosin. Biophys. J. 78, 908–917.
14. Strand, J., Butters, C. A., Nili, M., Homsher, E. &

Single Particle Analysis of Muscle Thin Filaments 771
Tobacman, L. S. (2000). Effect of tropomyosinsequence on myosin S1 binding to actin. Evidencefor a myosin target zone. Biophys. J. 78, 364a.
15. Lorenz, M., Poole, K. J. V., Popp, D., Rosenbaum, G. &Holmes, K. C. (1995). An atomic model of theunregulated thin filament obtained by X-ray fiberdiffraction on oriented actin–tropomyosin gels. J. Mol.Biol. 246, 108–119.
16. Wegner, A. (1979). Equilibrium of the actin–tropo-myosin interaction. J. Mol. Biol. 131, 839–853.
17. Wegner, A. & Walsh, T. P. (1981). Interaction oftropomyosin-troponin with actin filaments.Biochemistry, 20, 5633–5642.
18. McLachlan, A. D. & Stewart, M. (1976). The 14-foldperiodicity in a-tropomyosin and the interaction withactin. J. Mol. Biol. 103, 271–298.
19. Phillips, J. G. N., Fillers, J. P. & Cohen, C. (1986).Tropomyosin crystal structure and muscle regulation.J. Mol. Biol. 192, 111–131.
20. Hitchcock-DeGregori, S. E. & Varnell, T. A. (1990).Tropomyosin has discrete actin binding sites withsevenfold and fourteenfold periodicities. J. Mol. Biol.214, 885–896.
21. Landis, C., Back, N., Homsher, E. & Tobacman, L. S.(1999). Effects of tropomyosin internal deletions onthin filament function. J. Biol. Chem. 274, 31279–31285.
22. McKillop, D. F. A. & Geeves, M. A. (1993). Regulationof the interaction between actin and myosin subfrag-ment-1: evidence for three states of the thin filament.Biophys. J. 65, 693–701.
23. Lehrer, S. S. & Geeves, M. A. (1998). The musclethin filament as a classical cooperative/allostericregulatory system. J. Mol. Biol. 277, 1081–1089.
24. Smith, D. A., Maytum, R. & Geeves, M. A. (2003).Cooperative regulation of myosin–actin interactionsby a continuous flexible chain I: actin–tropomyosinsystems. Biophys. J. 84, 3155–3167.
25. Smith, D. A. & Geeves, M. A. (2003). Cooperativeregulation of myosin–actin interactions by a con-tinuous flexible chain II: actin–tropomyosin–troponinand regulation by calcium. Biophys. J. 84, 3168–3180.
26. Robinson, J. M., Dong, W.-J., Xing, J. & Cheung, H. C.(2004). Switching of troponin I: Ca2C and myosin-induced activation of heart muscle. J. Mol. Biol. 340,295–305.
27. Milligan, R. A., Whittaker, M. & Safer, D. (1990).Molecular structure of F-actin and the location ofsurface binding sites. Nature, 348, 217–221.
28. Owen, C., Morgan, D. G. & DeRosier, D. J. (1996).Image analysis of helical objects. The Brandeis helicalpackage. J. Struct. Biol. 116, 167–175.
29. Egelman, E. H. (2000). A robust algorithm for thereconstruction of helical filaments using single-particle methods. Ultramicroscopy, 85, 225–234.
30. Flicker, P. F., Phillips, G. N. & Cohen, C. (1982).Troponin and its interactions with tropomyosin. Anelectron microscope study. J. Mol. Biol. 162, 495–501.
31. Lehman, W., Rosol, M., Tobacman, L. S. & Craig, R.(2001). Troponin organization on relaxed and acti-vated thin filaments revealed by electron microscopyand three-dimensional reconstruction. J. Mol. Biol.307, 739–744.
32. Narita, A., Yasunaga, T., Ishikawa, T., Mayanagi, K. &Wakabayashi, T. (2001). Ca2C-induced switching oftroponin and tropomyosin on actin filaments asrevealed by electron cryo-microscopy. J. Mol. Biol.308, 241–261.
33. Luo, Y., Leszyk, J., Li, B., Li, Z., Gergely, J. & Tao, T.(2002). Troponin-I interacts with the Met47 region of
skeletal muscle actin. Implications for the mechanismof thin filament regulation by calcium. J. Mol. Biol. 316,429–434.
34. Perry, S. V. (1999). Troponin I: inhibitor or facilitator.Mol. Cell Biochem. 190, 9–32.
35. Squire, J. M. & Morris, E. P. (1998). A new look at thinfilament regulation in vertebrate skeletal muscle.FASEB J. 12, 761–771.
36. Hinkle, A., Goranson, A., Butters, C. A. & Tobacman,L. S. (1999). Roles for the troponin tail domain in thinfilament assembly and regulation. A deletional studyof cardiac troponin T. J. Biol. Chem. 274, 7157–7164.
37. Tobacman, L. S., Nihli, M., Butters, C., Heller, M.,Hatch, V., Craig, R. et al. (2002). The troponin taildomain promotes a conformational state of the thinfilament that suppresses myosin activity. J. Biol. Chem.277, 27636–27642.
38. Geeves, M. A. & Lehrer, S. S. (1994). Dynamics of themuscle thin filament regulatory switch: the size of thecooperative unit. Biophys. J. 67, 273–282.
39. Huynh, Q., Butter, C. A., Leiden, J. M. & Tobacman,L. S. (1996). Statistical mechanical analysis of a TnCsite II mutant. Biophys. J. 70, 1447–1455.
40. Tobacman, L. S. & Butters, C. A. (2000). A new modelof cooperative myosin-thin filament binding. J. Biol.Chem. 275, 27587–27593.
41. Spudich, J. A. & Watt, S. (1971). The regulation ofrabbit skeletal muscle contraction. I. Biochemicalstudies of the interaction of the tropomyosin–tropo-nin complex with actin and the proteolytic fragmentsof myosin. J. Biol. Chem. 242, 4866–4871.
42. Tobacman, L. S. & Adelstein, R. S. (1986). Mechanismof regulation of cardiac actin–myosin subfragment 1by troponin-tropomyosin. Biochemistry, 25, 798–802.
43. Hodgkinson, J. L., Marston, S. B., Craig, R., Vibert, P.& Lehman, W. (1997). Three-dimensional imagereconstruction of reconstituted smooth muscle thinfilaments: effects of caldesmon. Biophys. J. 72,2398–2404.
44. Egelman, E. H. (1986). An algorithm for straighteningimages of curved filamentous structures.Ultramicroscopy, 19, 367–374.
45. Frank, J., Radermacher, M., Penczek, P., Zhu, J., Li, Y.,Ladjadj, M. & Leith, A. (1996). SPIDER and WEB:processing and visualization of images in 3D electronmicroscopy and related fields. J. Struct. Biol. 116,190–199.
46. Van Heel, M. (1987). Similarity between images.Ultramicroscopy, 21, 95–100.
47. Frank, J. (1996). Three-dimensional Electron Microscopyof Macromolecular Assemblies, Academic Press, SanDiego.
48. Galkin, V. E., Orlova, A., Lukoyanova, N., Wriggers,W. & Egelman, E. H. (2001). Actin depolymerizingfactor stabilizes an existing state of F-actin and canchange the tilt of F-actin subunits. J. Cell Biol. 153,75–86.
49. Wriggers, W., Milligan, R. A. & McCammon, J. A.(1999). Situs: A package for docking crystal structuresinto low-resolution maps from electron microscopy.J. Struct. Biol. 125, 185–195.
50. Chacon, P. & Wriggers, W. (2002). Multi-resolutioncontour-based fitting of macromolecular structures.J. Mol. Biol. 317, 375–384.
51. Holmes, K. C., Angert, I., Kull, F. J., Jahn, W. &Schroder, R. R. (2003). Electron cryo-microscopyshows how strong binding of myosin to actin releasesnucleotide. Nature, 425, 423–427.
52. Takeda, S., Yamashita, A., Maeda, K. & Maeda, Y.

772 Single Particle Analysis of Muscle Thin Filaments
(2003). Structure of the core domain of human cardiactroponin in the Ca2C-saturated form. Nature, 424,35–41.
53. Egelman, E. H., Francis, N. & DeRosier, D. J. (1982).F-actin is a helix with a random variable twist. Nature,298, 131–135.
54. McGough, A., Pope, B., Chiu, W. & Weeds, A. (1997).Cofilin changes the twist of F-actin: implications foractin filament dynamics and cellular function. J. CellBiol. 138, 771–781.
55. Carugo, K. D., Banuelos, S. & Saraste, M. (1997).Crystal structure of a calponin homology domain.Nature Struct. Biol. 4, 175–179.
56. Huang, C. C., Couch, G. S., Petterson, E. F. & Ferrin,T. E. (1996). Chimera: an extensible molecular model-ing program application constructed using standardcomponents. Pacific Symp. Biocomput. 1, 724.
Edited by W. Baumeister
(Received 1 October 2004; received in revised form 29 November 2004; accepted 7 December 2004)





























