Embed Size (px)
Citation preview

A
H1teAttIw©
K
1
f(2itra
0d
Available online at www.sciencedirect.com
Molecular Immunology 45 (2008) 2990–2997
Slower rescue of ER homeostasis by the unfolded protein responsepathway associated with common variable immunodeficiency
Juliana S. Kuribayashi a, Cıntia R. Bombardieri a, Gisele V. Baracho a,b, Julio Aliberti c,Fabiana S. Machado c, Jorge Kalil b,d, Luiza Guilherme b,d, Cristina M. Kokron b,d,
Luiz V. Rizzo a,b,d, Maristela M. de Camargo a,∗a Department of Immunology, Institute of Biomedical Sciences, University of Sao Paulo,
Avenida Professor Lineu Prestes 1730, Sao Paulo 05508-900, Brazilb Institute for Investigation in Immunology (iii), University of Sao Paulo, Sao Paulo 05403-001, Brazil
c Division of Molecular Immunology, Cincinnati Children’s Hospital Medical Center and University of Cincinnati College of Medicine,Cincinnati 45229-3039, OH, USA
d Division of Clinical Immunology and Allergy, School of Medicine,University of Sao Paulo, Sao Paulo 05403-001, Brazil
Received 23 November 2007; received in revised form 11 January 2008; accepted 15 January 2008Available online 5 March 2008
bstract
Common variable immunodeficiency (CVID) is a primary immunodeficiency characterized by hypogammaglobulinemia and recurrent infections.erein we addressed the role of unfolded protein response (UPR) in the pathogenesis of the disease. Augmented unspliced X-box binding protein(XBP-1) mRNA concurrent with co-localization of IgM and BiP/GRP78 were found in one CVID patient. At confocal microscopy analysis
his patient’s cells were enlarged and failed to present the typical surface distribution of IgM, which accumulated within an abnormally expandedndoplasmic reticulum. Sequencing did not reveal any mutation on XBP-1, neither on IRE-1� that could potentially prevent the splicing to occur.nalysis of spliced XBP-1, IRE-1� and BiP messages after LPS or Brefeldin A treatment showed that, unlike healthy controls that respond to
hese endoplasmic reticulum (ER) stressors by presenting waves of transcription of these three genes, this patient’s cells presented lower rates ofranscription, not reaching the same level of response of healthy subjects even after 48 h of ER stress. Treatment with DMSO rescued IgM andgG secretion as well as the expression of spliced XBP-1. Our findings associate diminished splicing of XBP-1 mRNA with accumulation of IgMithin the ER and lower rates of chaperone transcription, therefore providing a mechanism to explain the observed hypogammaglobulinemia.2008 Elsevier Ltd. All rights reserved.
ta(ddJS
eywords: B lymphocyte; Immunoglobulin; Immunodeficiency; ER
. Introduction
Common variable immunodeficiency (CVID) is the mostrequent primary immunodeficiency diagnosed in adultsCunningham-Rundles and Bodian, 1999; Kainulainen et al.,001; Sneller, 2001; Tiller and Liddle, 2000). The clin-cal spectrum of CVID is broad, arising from childhood
o late adult life. Most patients have a history of recur-ent respiratory tract infections, but it may also includeutoimmune phenomena, bowel infections and granuloma-∗ Corresponding author. Tel.: +55 11 3091 7434; fax: +55 11 3091 7224.E-mail address: [email protected] (M.M. de Camargo).
Ctelfwv
161-5890/$ – see front matter © 2008 Elsevier Ltd. All rights reserved.oi:10.1016/j.molimm.2008.01.013
ous disease. The diagnosis of CVID can only be madefter other immunodeficiencies such as hyper-IgM syndromeKaneko et al., 2005), X-linked agammaglobulinemia (vaner Hilst et al., 2002) and X-linked lymphoproliferative syn-rome (Aghamohammadi et al., 2003; Elenitoba-Johnson andaffe, 1997; Fontan-Casariego, 2001; Morra et al., 2001;oresina et al., 2002) have been excluded. Patients withVID may have normal or decreased levels of B cells in
he periphery (Caballero et al., 1984; Farrant, 1991; Farrantt al., 1989, 1985, 1994). In some patients with normal
evels of B cells, these have a mature phenotype bearing sur-ace immunoglobulins, while some other only possess cellsith an immature phenotype characterized by an enlargedolume, restricted usage of VH families and decreased mutat-
ar Imm
ig
m1taafXauooatL1
pbotacmEhC
2
2
ptbbiocNbfhs
2e
owPum
p1
2
wfl(
2
rtaa(atATC5CIt5GGCmms
2
LmFnr(t
3
soab
J.S. Kuribayashi et al. / Molecul
ng capacity at the hypervariable regions of the immunoglobulinenes (Braun et al., 1991, 1992a,b).
Terminally differentiated B cells have an active secretoryachinery due to intense immunoglobulin production. The IRE-
�/XBP-1 signalling loop ensures the effective folding andransport of these molecules (Gass et al., 2004; Rutkowskind Kaufman, 2004). X-box binding protein 1 (XBP-1) is
transcription factor involved in terminal plasma cell dif-erentiation as demonstrated by studies with chimera miceBP-1−/−/RAG-2−/− (Reimold et al., 2001). Plasma cells arebsent in these animals, which secrete very little immunoglob-lin of any isotype (Reimold et al., 2001). Augmented levelsf XBP-1 transcripts were detected in plasma cells infiltratesf inflammatory diseases, such as rheumatic arthritis (Kim etl., 1999). Additionally, it was also detected in primary cul-ure of murine B lymphocytes stimulated with anti-CD40 orPS and in several myeloma cell lines (Reimold et al., 2001,996).
Given its well-defined role on the antibody response it isossible that defective XBP-1 expression could result in anti-ody secretion disorders. In this report, we describe the findingsbtained through a novel molecular screening of 74 adult par-icipants. We found diminished splicing of XBP-1 mRNA to bessociated with accumulation of IgM in the ER and lower rates ofhaperone transcription in one subject that presented hypogam-aglobulinemia. We hypothesize that a slower recovery of theR homeostasis is one biochemical mechanism to explain theypogammaglobulinemia observed among some patients withVID.
. Materials and methods
.1. Subjects
Patients in this study were diagnosed with CVID since theyresent clinically relevant immunodeficiency signs and symp-oms plus serum IgG and IgA levels two standard deviationsellow the average for their age, if serum levels were normalefore the age of two. Additional criteria were: low or absentsohemagglutinins and poor antibody response to vaccines. Allther causes for hypogammaglobulinemia were ruled out. Ourohort included both male and female patients ages 18 to 54.one of the patients had a history of cancer. The internal reviewoard of the institutions involved approved all procedures per-ormed here. Blood collection was performed after the patientsave read and signed an informed consent form designed for thistudy.
.2. Isolation of peripheral-blood mononuclear cells andx vivo B cells, EBV-immortalization of B cells
Peripheral-blood mononuclear cells (PBMC) werebtained by centrifugation in Isolymph® gradient. PBMC
ere harvested at the interface, washed three times inBS and immediately processed. B cells were isolatedsing magnetic beads (MACS Miltenyi), according to theanufacturer’s protocol. Immortalization of B cells wasBXrt
unology 45 (2008) 2990–2997 2991
erformed according to standard protocol (Bird et al.,981).
.3. Flow cytometry analysis of PBMC
Cell-surface markers of peripheral-blood mononuclear cellsere determined by immunofluorescence antibody staining andow cytometry. Analysis was done with the software FlowJoTreeStar).
.4. RT- and Q-PCR analysis and sequencing
Total RNA was isolated from 107 cells using Trizol®
eagent (Invitrogen). cDNA was generated using 200 ng ofotal RNA. Total (unspliced and spliced) XBP-1 cDNAs weremplified in 35 cycles and mRNA content was normalizedgainst human glyceraldeyde 3-phosphate dehydrogenaseGAPDH). cDNA signals were captured by densitometrynalysis of agarose gels stained with ethidium bromide. Forotal (unspliced and spliced) XBP-1 the primers used were 5′-AACAGAGTAGCAGCTCAGACTGC-3′ and 5′-GTATCTC-AAGACTAGGGGCTTGGTA-3′. For unspliced XBP-1, 5′-AGAGGTGCACGTAGTCTGAGTGCTG-3′. For GAPDH,′-ACCACAGTCCATGCCATCAC-3′ and 5′-TCCACCAC-CTGTTGCTGTA-3′. The expression levels of spliced XBP-1,
RE-1� and BiP mRNA were measured by real-time PCR usinghe primers: XBP-1s 5′-GAGTCCGCAGCAGGTG-3′ and′-GGGCTTGGTATATATGTGG-3′; IRE-1� 5′-CGGCCTTT-CAGATAGTCTC-3′ and 5′-ACGTCCCCAGATTCACT-TC-3′; BiP 5′-CGAGGAGGAGGACAAGAAGG-3′ and 5′-ACCTTGAACGGCAAGAACT-3′. The quantity of specificRNA was normalized as a ratio to the amount of GAPDHRNA as described by Livak and Schmittgen (2001). For
equencing, amplicons were generated using Pfx polymerase.
.5. Confocal microscopy
PBMC, purified B cells or EBV B cells were exposed toPS for 48 h, cytospun onto slides, air dried and fixed with coldethanol. Cytospins were stained with anti-human-IgM-Alexaluor 488 and anti-BiP/GRP78-Alexa Fluor 546 followed byuclear counterstaining with DAPI. Image analysis and fluo-escence overlay was performed using Axiovision 4.2 softwareZeiss) and displayed as monochrome single color panels and/orhree-color overlays.
. Results
When cultured for 48 h in the presence of LPS, PBMChow a marked shift from unspliced to the spliced formf the protein XBP-1 (XBP-1p60) both in healthy individu-ls (n = 41) and CVID patients (n = 32). This shift in XBP-1and size occurs during differentiation of antibody-secreting
cells and results from the removal of a 26 bp intron fromBP-1 mRNA resulting in an open reading frame of 376 aa
ather than the shorter 261 aa. The splicing reflects the activa-ion of the unfolded protein response (UPR) pathway (Calfon
2992 J.S. Kuribayashi et al. / Molecular Immunology 45 (2008) 2990–2997
Table 1Analysis of leukocytes subsets from patient P
Surface markers (% of gated lymphocytes) Patient P year 2004 Patient P year 2007 Reference valuea
CD3 41.7 44.3 38–70CD3CD4 18.2 21.65 35–45CD3CD8 22.6 20.4 20–30CD3CD25 0.45 1.04 0.4–6.0CD3CD69 0.3 nd 0.5–6.0CD4annexinV 9.5 nd 7.5–17.5CD4CD45RA 44.1 26.15 24–50CD4CD45RO 1.1 27.7 9.5–35CD8CD57 14.3 5.63 5–11CD14 7 18.85 0.3–15CD16CD56 0 13.95 2.9–13.3CD19 15.8 8.05 6.4–15CD19CD25 2.6 2.6 0–1.5CD40L 0.5 nd 4–11CD70 2.5 nd 0–4CD95 31 nd 0–39
nd = not determined, flow-cytometric analysis of peripheral-blood mononuclear cells with antibodies against CD3, CD4, CD8, CD14, CD16, CD19, CD25, CD27,CD40L, CD45RA, CD45RO, CD56, CD57, CD70, CD95 and annexin-V revealed a typical profile for Brazilian CVID patients, normal levels of mature circulatingB D8 r
eias
boisabwoi
wcwXsannX(c(stfwta1
m2
ftafwBiorIfrawac
a(iuamIrg2
cells and lower counts of CD3+CD4+ with consequent inversion in the CD4/Ca Kokron et al. (2004) (25).
t al., 2002; Gass et al., 2002). One exception was foundn our cohort, patient P, who continued to lack detectablemounts of XBP-1p60 even after ER stress by LPS (data nothown).
Patient P, currently 20 years old, was diagnosed with CVIDy age of 16, after presenting recurrent pneumonias (averagef 4 per year), chronic sinusitis and frequent episodes of bowelnfection lasting longer than 10 days. At time of diagnosis hererum antibody levels were 328 mg/dl of IgG, 26 mg/dl of IgMnd 21 mg/dl of IgA. The analysis of T and B lymphocyte subsetsy flow cytometry revealed a profile typical for Brazilian patientsith CVID (Kokron et al., 2004), characterized by normal levelsf mature B cells and low CD3+CD4+ counts with a consequentnversion in the CD4/CD8 ratio (Table 1).
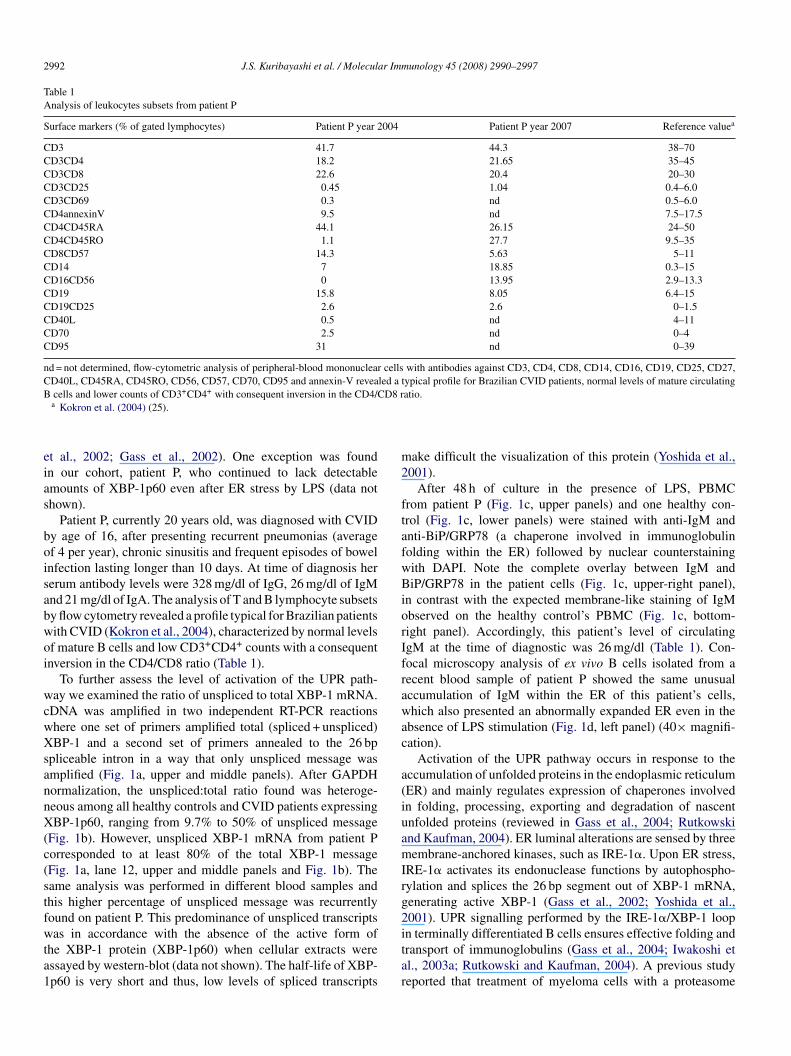
To further assess the level of activation of the UPR path-ay we examined the ratio of unspliced to total XBP-1 mRNA.
DNA was amplified in two independent RT-PCR reactionshere one set of primers amplified total (spliced + unspliced)BP-1 and a second set of primers annealed to the 26 bp
pliceable intron in a way that only unspliced message wasmplified (Fig. 1a, upper and middle panels). After GAPDHormalization, the unspliced:total ratio found was heteroge-eous among all healthy controls and CVID patients expressingBP-1p60, ranging from 9.7% to 50% of unspliced message
Fig. 1b). However, unspliced XBP-1 mRNA from patient Porresponded to at least 80% of the total XBP-1 messageFig. 1a, lane 12, upper and middle panels and Fig. 1b). Theame analysis was performed in different blood samples andhis higher percentage of unspliced message was recurrentlyound on patient P. This predominance of unspliced transcripts
as in accordance with the absence of the active form ofhe XBP-1 protein (XBP-1p60) when cellular extracts weressayed by western-blot (data not shown). The half-life of XBP-p60 is very short and thus, low levels of spliced transcripts
itar
atio.
ake difficult the visualization of this protein (Yoshida et al.,001).
After 48 h of culture in the presence of LPS, PBMCrom patient P (Fig. 1c, upper panels) and one healthy con-rol (Fig. 1c, lower panels) were stained with anti-IgM andnti-BiP/GRP78 (a chaperone involved in immunoglobulinolding within the ER) followed by nuclear counterstainingith DAPI. Note the complete overlay between IgM andiP/GRP78 in the patient cells (Fig. 1c, upper-right panel),
n contrast with the expected membrane-like staining of IgMbserved on the healthy control’s PBMC (Fig. 1c, bottom-ight panel). Accordingly, this patient’s level of circulatinggM at the time of diagnostic was 26 mg/dl (Table 1). Con-ocal microscopy analysis of ex vivo B cells isolated from aecent blood sample of patient P showed the same unusualccumulation of IgM within the ER of this patient’s cells,hich also presented an abnormally expanded ER even in the
bsence of LPS stimulation (Fig. 1d, left panel) (40× magnifi-ation).
Activation of the UPR pathway occurs in response to theccumulation of unfolded proteins in the endoplasmic reticulumER) and mainly regulates expression of chaperones involvedn folding, processing, exporting and degradation of nascentnfolded proteins (reviewed in Gass et al., 2004; Rutkowskind Kaufman, 2004). ER luminal alterations are sensed by threeembrane-anchored kinases, such as IRE-1�. Upon ER stress,
RE-1� activates its endonuclease functions by autophospho-ylation and splices the 26 bp segment out of XBP-1 mRNA,enerating active XBP-1 (Gass et al., 2002; Yoshida et al.,001). UPR signalling performed by the IRE-1�/XBP-1 loop
n terminally differentiated B cells ensures effective folding andransport of immunoglobulins (Gass et al., 2004; Iwakoshi etl., 2003a; Rutkowski and Kaufman, 2004). A previous studyeported that treatment of myeloma cells with a proteasome
J.S. Kuribayashi et al. / Molecular Immunology 45 (2008) 2990–2997 2993
Fig. 1. Role of the UPR pathway on CVID. Representative results of only some of the subjects are shown. (a) Two independent RT-PCR reactions were carriedout in order to amplify total XBP-1 message (spliced + unspliced) or only unspliced message, upper and middle panel, respectively. GAPDH was amplified fornormalization purposes, lower panel. (b) Densitometric analysis of results shown on upper panel. The mRNA levels were normalized to that of GAPDH and the ratiocalculated as [spliced = total − unspliced]. (c) PBMC from patient P (upper panels) or healthy control (lower panels) were stimulated with LPS for 48 h and stainedw lowedg rescee ulturew
is(watbsct0gai
t
wXtLpipbpctb
ith anti-IgM (green fluorescence) and anti-BiP/GRP78 (red fluorescence) folreyscale photographs obtained for each marker, the right panels depict the fluoxpanded ER before (left panel) and after 48 h of LPS stimulation (right panel). Cere taken at 40× magnification.
nhibitor blocked the IRE-1�/XBP-1 pathway and reduced theplicing of XBP-1 mRNA, leading these cells to apoptosisLee et al., 2003a). Therefore, disruption of the UPR path-ay results in poor plasma cell development and function,
nd ultimately, in immunodeficiency. We asked whether muta-ions of XBP-1 mRNA from patient P could prevent splicingy IRE-1� and explain the higher ratios of unspliced mes-age found. XBP-1 amplicons obtained from patient P wereloned and sequenced. From 18 clones sequenced, all containedhe unspliced message but none presented mutations within or.6 kb up- or downstream of the 26 bp intron (not shown), sug-esting this patient’s XBP-1 mRNA sequence does not present
ny mutation that could potentially prevent or inhibit splic-ng.B cells from patient P and one healthy control were immor-alized by EBV infection. These cells were stimulated in vitro
rvac
by nuclear counterstaining with DAPI (blue fluorescence). Each panel showsnce colored overlay. (d) B cells from patient P present enlarged phenotype andd purified B cells from patient P and stained as in panel C. All microphotographs
ith 10 mg/ml LPS for 12 and 48 h and the levels of splicedBP-1, IRE-1� and BiP were determined by quantitative real-
ime RT-PCR (Q-PCR) (Fig. 2a). Upon ER stress triggered byPS stimulation, spliced XBP-1, IRE-1� and BiP messageseaked at 12 h in the healthy control cells, rapidly decreas-ng afterwards, in reflection of the activation of the UPRathway and resolution of the ER stress (Fig. 2a, round sym-ols). In contrast, EBV cells from patient P did not present aeak at 12 h (Fig. 2a, triangle symbols). In fact, this patient’sells have not yet increased the transcriptional levels of thesehree genes at 48 h after LPS treatment (Fig. 2a, triangle sym-ols), suggesting a slower response of the UPR pathway. The
ole of unspliced XBP-1 message is still a theme of contro-ersy. There’s the possibility that unspliced XBP-1 transcriptsct like a dominant negative, regulating the response of aell to ER stressors (Lee et al., 2003a; Tirosh et al., 2006).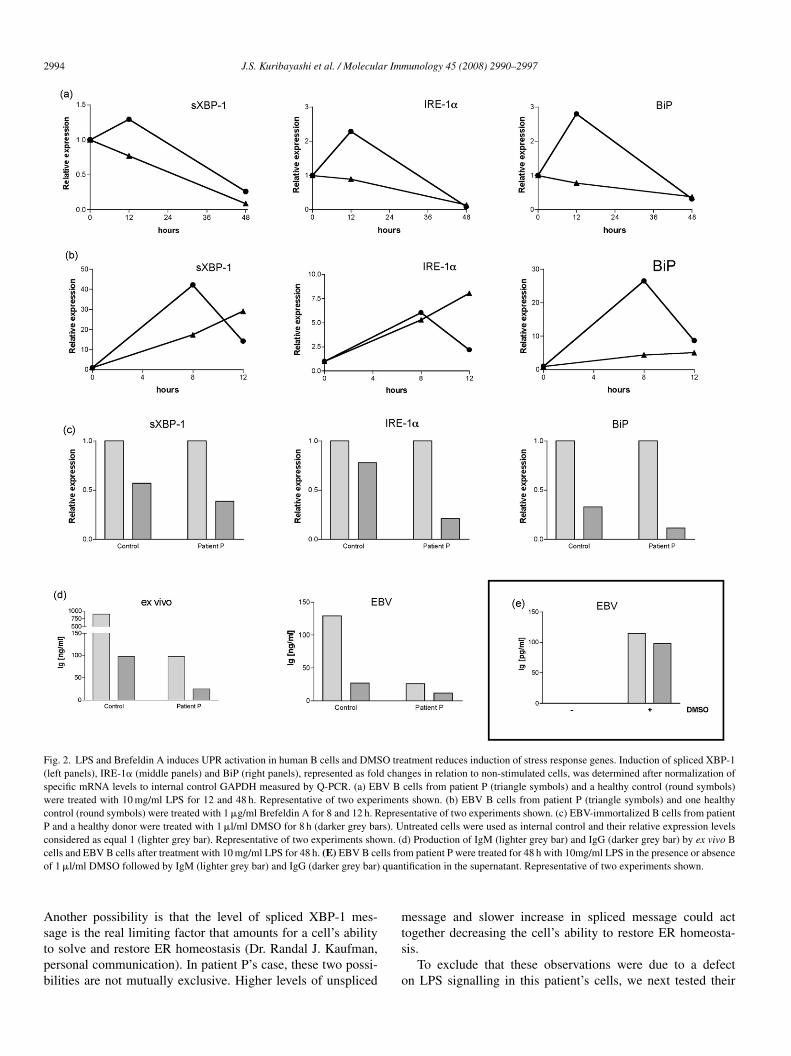
2994 J.S. Kuribayashi et al. / Molecular Immunology 45 (2008) 2990–2997
Fig. 2. LPS and Brefeldin A induces UPR activation in human B cells and DMSO treatment reduces induction of stress response genes. Induction of spliced XBP-1(left panels), IRE-1� (middle panels) and BiP (right panels), represented as fold changes in relation to non-stimulated cells, was determined after normalization ofspecific mRNA levels to internal control GAPDH measured by Q-PCR. (a) EBV B cells from patient P (triangle symbols) and a healthy control (round symbols)were treated with 10 mg/ml LPS for 12 and 48 h. Representative of two experiments shown. (b) EBV B cells from patient P (triangle symbols) and one healthycontrol (round symbols) were treated with 1 �g/ml Brefeldin A for 8 and 12 h. Representative of two experiments shown. (c) EBV-immortalized B cells from patientP and a healthy donor were treated with 1 �l/ml DMSO for 8 h (darker grey bars). Untreated cells were used as internal control and their relative expression levelsconsidered as equal 1 (lighter grey bar). Representative of two experiments shown. (d) Production of IgM (lighter grey bar) and IgG (darker grey bar) by ex vivo Bc lls froo quan
Astpb
m
ells and EBV B cells after treatment with 10 mg/ml LPS for 48 h. (E) EBV B cef 1 �l/ml DMSO followed by IgM (lighter grey bar) and IgG (darker grey bar)
nother possibility is that the level of spliced XBP-1 mes-
age is the real limiting factor that amounts for a cell’s abilityo solve and restore ER homeostasis (Dr. Randal J. Kaufman,ersonal communication). In patient P’s case, these two possi-ilities are not mutually exclusive. Higher levels of unsplicedts
o
m patient P were treated for 48 h with 10mg/ml LPS in the presence or absencetification in the supernatant. Representative of two experiments shown.
essage and slower increase in spliced message could act
ogether decreasing the cell’s ability to restore ER homeosta-is.To exclude that these observations were due to a defectn LPS signalling in this patient’s cells, we next tested their

ar Imm
rBclthauoBmrssattroRawaosc(g
ti(iif(p1adtoas
aaoaB(aIgsto
PXmaddpmti
4
pmscpodg2
e1iEmi(
AiCitpto
CtdiPeitrpl
J.S. Kuribayashi et al. / Molecul
esponse to LPS-independent stress caused by Brefeldin A.refeldin A disrupts the protein transport from ER to Golgiomplex, which then collapses, causing a retrograde accumu-ation of proteins within the ER lumen, leading this organelleo stress. EBV B cells (Fig. 2b) from patient P and oneealthy control were treated with 1 �g/ml Brefeldin A for 8nd 12 h. Similarly to our previous observations with LPS stim-lation, the stress caused by Brefeldin A increased splicingf XBP-1 message and induced higher levels of IRE-1� andiP transcription by healthy control cells at 8 h after treat-ent, with rapid decrease of these three parameters afterwards,
eflecting the restoration of ER homeostasis (Fig. 2b, roundymbols). In contrast, cells from patient P (Fig. 2b, triangleymbols) presented slower slopes of increase in spliced XBP-1nd BiP transcripts, suggesting a decreased ability to resolvehe ER stress, mirroring the observations with LPS. Impor-antly, the levels of IRE-1� message increases at a higherate in this patient’s cells and does not decrease after 8 h asbserved in healthy control cells (Fig. 2b, middle panel). TheNase activity of IRE-1� is required for activation of the UPRnd augmented levels of IRE-1� mRNA are often associatedith decreased RNase activity of the protein (Tirasophon et
l., 2000). We hypothesized that mutations on either kinaser endonuclease domains of IRE-1� could explain the tran-cription kinetics observed. We sequenced PCR ampliconsontaining both domains and found no mutations in patient PGenBank access numbers for IRE-1� gene gi:39775951 andi:51511734).
Chemical chaperones are low molecular weight moleculeshat stabilize misfolded proteins and promote their proper fold-ng and transport out of the ER (Welch and Brown, 1996). DMSOdimethilsulfoxide, (CH3)2SO) is currently being tested in clin-cal trials for its ability to act like a chemical chaperone andts potential as treatment of diseases caused by protein mis-olding, like cystic fibrosis and nephrogenic diabetes insipidusBernier et al., 2004; Morello et al., 2000). EBV cells fromatient P and a healthy control were treated in vitro with�l/ml DMSO for 8 h and the levels of spliced XBP-1, IRE-1�nd BiP before (Fig. 2c, lighter grey bars) and after (Fig. 2c,arker grey bars) treatment were determined. We found thatreatment of cells with DMSO rescued these cells’ ER home-stasis as indicated by lower levels of spliced XBP-1, IRE-1�nd BiP messages, reflecting resolution of basal level of ERtress.
EBV-immortalization did not alter these cells profile for IgMnd IgG production triggered by LPS stimulation (Fig. 2d). Thectivation of EBV cells from patient P with LPS in the presencef DMSO restored these cells’ ability to secrete IgM and IgG,s measured by ELISA in 48 h culture supernatant (Fig. 2e).iP transcription levels are considerably low in patient P cells
Fig. 2a and b, right panels, triangle symbols) and this might beclue to explain how DMSO overcomes this defect and restores
g secretion. The induction of spliced XBP-1 by DMSO sug-
ests that it is possible to induce this transcript and restore Igecretion, which goes along the absence of mutations found inhe splicing site of xbp1 and in the kinase/endonuclease domainsf ire1a.Phwl
unology 45 (2008) 2990–2997 2995
The next step would be to revert the XBP-1 defect in patient’s cells by transducing these cells with the spliced form ofBP-1 and checking whether increased amounts of splicedessage would rescue immunoglobulin secretion. So far, our
ttempts at accomplishing this failed because the patient’s cellsid not tolerate transfection using liposomes or retroviral trans-uction. We believe that given the poor physiological stateresented by these cells ex vivo, a rescue of function experi-ent will require a more sophisticated approach than a simple
ransfection/over-expression and thus, these are ongoing exper-ments.
. Discussion
We speculate herein a possible mechanism underlying theathology of one CVID patient. In our patient P, as in chimeraice lacking the spliced form of XBP-1, the absence of active
pliced XBP-1 is associated with diminished levels of cir-ulating immunoglobulins (Reimold et al., 2001). The dataresented here provide the first evidence that maintenancef ER homeostasis in human immunoglobulin-secreting cellsoes rely upon the UPR pathway, in accordance to those dataenerated in mice (Iwakoshi et al., 2003a; Reimold et al.,001).
Earlier descriptions of CVID patients’ cells describednlarged B cells with immature phenotype (Braun et al., 1991,992a,b). Our data suggest that the enlarged phenotype, at leastn some patients, might be the result of an abnormally expandedR caused by these cells inability to properly fold and exportassive amounts of nascent immunoglobulins, leading to an
ncrease in ER volume to accommodate this protein overloadFig. 1c and d).
Other players of the UPR pathway, such as PERK andTF6, remain to be studied. UPR dysfunction has been
mplicated in other diseases that are more common thanVID, such as cystic fibrosis and nephrogenic diabetes
nsipidus (Bernier et al., 2004; Morello et al., 2000), andhus may not be specific to all CVID patients. Interestingly,atient P has not presented, to this point, any symptom ofhe diseases more commonly associated with UPR pathol-gy.
Recently, Taubenheim and collaborators suggested thatVID could not be caused by a defect on XBP-1 gene since
his gene is crucial for B cell terminal maturation and anyefect would be lethal (Taubenheim et al., 2005). Our data isn agreement with this supposition in the sense that our patientdoes splice XBP-1, what is unquestionably proven by her oth-
rwise low amounts of spliced XBP-1 message. Our point heres that this patient’s rate of XBP-1 splicing is lower, leadingo a slower recovery of the ER homeostasis upon stress, lowerates of immunoglobulin folding/exporting and enlarged B cellshenotype. At this point we have no clear explanation for theower amounts of spliced XBP-1 transcripts found in-patient
. The mice data shows that the delay in re-establishing ERomeostasis is overcome by the cells by entering into apoptosis,hich ultimately contributes to the animals’ hypogammaglobu-inemia. We are currently checking apoptosis activation in this

2 ar Imm
pmooltfhtsmm
A
iRNo(SL
R
A
B
B
B
B
B
C
C
C
E
F
F
F
F
F
G
G
I
K
K
K
K
L
L
M
M
R
R
R
S
996 J.S. Kuribayashi et al. / Molecul
atient’s cells. Our data shows that IgM and unspliced XBP-1essage accumulation in our patient P is due to deregulation
f XBP-1/IRE1-� loop, providing clues about the mechanismsf hypogammaglobulinemia observed in this patient. Neverthe-ess, we believe it is important to bring our colleagues’ attentiono our findings. We propose CVID patients should be screenedor XBP-1 splicing defect through the in vitro assays describederein. Patients’ B cells should be exposed to DMSO and haveheir Ig production measured in order to expand these analy-es and test our hypothesis that a defect on the protein foldingachinery should be thought off when investigating hypogam-aglobulinemias.
cknowledgements
We thank the patients for giving us the privilege of conduct-ng these experiments and Drs. Anna Carla Goldberg, Gunnar. Mair, Marcia Lachtermacher-Triunfol and Rubens Belforteto for critical reading of this manuscript. Brazilian Ministryf Science and Technology; Sao Paulo State Council for ScienceFAPESP) (01/02584-2 for M.M.C.) and National Council forcientific and Technological Development (CNPq) (J.S.K. and.V.R.).
eferences
ghamohammadi, A., Kanegane, H., Moein, M., Farhoudi, A., Pourpak, Z.,Movahedi, M., Gharagozlou, M., Zargar, A.A., Miyawaki, T., 2003. Identi-fication of an SH2D1A mutation in a hypogammaglobulinemic male patientwith a diagnosis of common variable immunodeficiency. Int. J. Hematol. 78,45–47.
ernier, V., Lagace, M., Bichet, D.G., Bouvier, M., 2004. Pharmacological chap-erones: potential treatment for conformational diseases. Trends Endocrinol.Metab. 15, 222–228.
ird, A.G., McLchlan, S.M., Britton, S., 1981. Cyclosporin A promotes sponta-neous outgrowth “in vitro” of Epstein Barr virus-induced B-cell line. Nature289, 300–301.
raun, J., Berberian, L., King, L., Sanz, I., Govan 3rd., H.L., 1992a. Restricteduse of fetal VH3 immunoglobulin genes by unselected B cells in the adult pre-dominance of 56p1-like VH genes in common variable immunodeficiency.J. Clin. Invest. 89, 1395–1402.
raun, J., Galbraith, L., Valles-Ayoub, Y., Saxon, A., 1991. Human immunod-eficiency resulting from a maturational arrest of germinal center B cells.Immunol. Lett. 27, 205–208.
raun, J., Saxon, A., Wall, R., Morrison, S.L., 1992b. The second century ofthe antibody. Molecular perspectives in regulation, pathophysiology, andtherapeutic applications. West. J. Med. 157, 158–168.
aballero, F.M., Brown, W.R., Kohler, P.F., Hayward, A.R., 1984. B cell num-bers and responses in patients with common variable immunodeficiencyand nodular lymphoid hyperplasia of the bowel. J. Clin. Lab. Immunol. 13,59–63.
alfon, M., Zeng, H., Urano, F., Till, J.H., Hubbard, S.R., Harding, H.P.,Clark, S.G., Ron, D., 2002. IRE1 couples endoplasmic reticulum loadto secretory capacity by processing the XBP-1 mRNA. Nature 415, 92–96.
unningham-Rundles, C., Bodian, C., 1999. Common variable immunodefi-ciency: clinical and immunological features of 248 patients. Clin. Immunol.92, 34–48.
lenitoba-Johnson, K.S., Jaffe, E.S., 1997. Lymphoproliferative disorders asso-ciated with congenital immunodeficiencies. Semin. Diagn. Pathol. 14,35–47.
arrant, J., 1991. T and B cell defects in common variable immunodeficiency.Immunol. Invest. 20, 143–150.
S
unology 45 (2008) 2990–2997
arrant, J., bryant, A., Almandoz, F., Spickett, G., Evans, S.W., Webster, A.D.,1989. B cell function in acquired “common-variable” hypogammaglobuline-mia: proliferative responses to lymphokines. Clin. Immunol. Immunopathol.51, 196–204.
arrant, J., Bryant, A.E., Lever, A.M., Edwards, A.J., Knight, S.C., Webster,A.D., 1985. Defective low-density cells of dendritic morphology from theblood of patients with common variable hypogammaglobulinemia: lowimmunoglobulin production on stimulation of normal B cells. Clin. Exp.Immunol. 61, 189–194.
arrant, J., Spickett, G., Matamoros, N., Copas, D., Hernandez, M., North,M., Chapel, H., Webster, A.D., 1994. Study of B and T cell phenotypesin blood from patients with common variable immunodeficiency (CVID).Immunodeficiency 5, 159–169.
ontan-Casariego, G., 2001. Primary immunodeficiencies clinical features andvariant forms. Allergol. Immunopathol. (Madr) 29, 101–107.
ass, J.N., Gifford, N.M., Brewer, J.W., 2002. Activation of an unfolded proteinresponse during differentiation of antibody-secreting B cells. J. Biol. Chem.277, 49047–49054.
ass, J.N., Gunn, K.E., Sriburi, R., Brewer, J.W., 2004. Stressed-out Bcells? Plasma-cell differentiation and the unfolded protein response. TrendsImmunol. 25, 17–24.
wakoshi, N.N., Lee, A.H., Vallabhajosyula, P., Otipoby, K.L., Rajewsky, K.,Glimcher, L.H., 2003a. Plasma cell differentiation and the unfolded pro-tein response intersect at the transcription factor XBP-1. Nat. Immunol. 4,321–329.
ainulainen, L., Nikoskelainen, J., Ruuskanen, O., 2001. Diagnostic findingsin 95 Finnish patients with common variable immunodeficiency. J. Clin.Immunol. 21, 145–149.
aneko, H., Kawamoto, N., Asano, T., Mabuchi, Y., Horikoshi, H., Teramoto,T., Matsui, E., Kondo, M., Fukao, T., Kasahara, K., Kondo, N., 2005. Leakyphenotype of X-linked agammaglobulinaemia in a Japanese family. Clin.Exp. Immunol. 140, 520–523.
im, H.J., Krenn, V., Steinhauser, G., Berek, C., 1999. Plasma cell develop-ment in synovial germinal centers in patients with rheumatoid and reactivearthritis. J. Immunol. 162, 3053–3062.
okron, C.M., Errante, P.R., Barros, M.T., Baracho, G.V., Camargo, M.M., Kalil,J., Rizzo, L.V., 2004. Clinical and laboratory aspects of common variableimmunodeficiency. An. Acad. Bras. Cienc. 76, 707–726.
ee, A.H., Iwakoshi, N.N., Anderson, K.C., Glimcher, L.H., 2003a. Proteasomeinhibitors disrupt the unfolded protein response in myeloma cells. Proc. Natl.Acad. Sci. U.S.A. 100, 9946–9951.
ivak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expression datausing real-time quantitative PCR and the 2(-Delta Delta C(T)) method.Methods 25, 402–408.
orello, J.P., Petaja-Repo, U.E., Bichet, D.G., Bouvier, M., 2000. Pharmaco-logical chaperones: a new twist on receptor folding. Trends Pharmacol. Sci.21, 466–469.
orra, M., Silander, O., Calpe, S., Choi, M., Oettgen, H., Myers, L., Etzioni,A., Buckley, R., Terhorst, C., 2001. Alterations of the X-linked lympho-proliferative disease gene SH2D1A in common variable immunodeficiencysyndrome. Blood 98, 1321–1325.
eimold, A.M., Iwakoshi, N.N., Manis, J., Vallabhajosyula, P., Szomolanyi-Tsuda, E., Gravallese, E.M., Friend, D., Grusby, M.J., Alt, F., Glimcher, L.H.,2001. Plasma cell differentiation requires the transcription factor XBP-1.Nature 412, 300–307.
eimold, A.M., Ponath, P.D., Li, Y.S., Hardy, R.R., David, C.S., Strominger, J.L.,Glimcher, L.H., 1996. Transcription factor B cell lineage-specific activatorprotein regulates the gene for human X-box binding protein 1. J. Exp. Med.183, 393–401.
utkowski, D.T., Kaufman, R.J., 2004. A trip to the ER: coping with stress.Trends Cell Biol. 14, 20–28.
neller, M.C., 2001. Common variable immunodeficiency. Am. J. Med. Sci.321, 42–48.
oresina, A., Lougaris, V., Giliani, S., Cardinale, F., Armenio, L.,Cattalini, M., Notarangelo, L.D., Plebani, A., 2002. Mutations ofthe X-linked lymphoproliferative disease gene SH2D1A mimick-ing common variable immunodeficiency. Eur. J. Pediatr. 161, 656–659.

ar Imm
T
T
T
T
v
Wcal chaperones on protein folding. Cell Stress Chaperones 1, 109–115.
Yoshida, H., Matsui, T., Yamamoto, A., Okada, T., Mori, K., 2001. XBP1 mRNA
J.S. Kuribayashi et al. / Molecul
aubenheim, N., von Hornung, M., Durandy, A., Warnatz, K., Corcoran, L.,Peter, H.H., Eibel, H., 2005. Defined blocks in terminal plasma cell differ-entiation of common variable immunodeficiency patients. J. Immunol. 175,5498–5503.
iller, T.L.J., Liddle, K.J., 2000. Immunodeficiency problems in children. J. S.C. Med. Assoc. 96, 225–228.
irasophon, W., Lee, K., Callaghan, B., Welihinda, A., Kaufman, R.J., 2000. The
endoribonuclease activity of mammalian IRE1 autoregulates its mRNA andis required for the unfolded protein response. Genes Dev. 14, 2725–2736.irosh, B., Iwakoshi, N.N., Glimcher, L.H., Ploegh, H.L., 2005. Rapid turnoverof unspliced XBP-1 as a factor that modulates the unfolded protein response.J. Biol. Chem. 281, 5852–5860.
unology 45 (2008) 2990–2997 2997
an der Hilst, J.C., Smits, B.W., van der Meer, J.W., 2002. Hypogammaglobu-linaemia: cumulative experience in 49 patients in a tertiary care institution.Neth. J. Med. 60, 140–147.
elch, W.J., Brown, C.R., 1996. Influence of molecular and chemi-
is induced by ATF6 and spliced by IRE1 in response to ER stress to producea highly active transcription factor. Cell 107, 881–891.





























