Embed Size (px)
Citation preview

ISSN: 1524-4571 Copyright © 2005 American Heart Association. All rights reserved. Print ISSN: 0009-7330. Online
TX 72514Circulation Research is published by the American Heart Association. 7272 Greenville Avenue, Dallas,
DOI: 10.1161/01.RES.0000194324.29363.82 published online Nov 3, 2005; Circ. Res.
Roland Vetter, Joachim Jankowski, Martin Paul and Gilbert Schönfelder Stefan Verlohren, Norbert Fuhr, Malgorzata Lanowska, Andreas Nonnenmacher, Oliver Krysiak, Anja Bretschneider, Enhong Zhong, Jessie Webb, Hartmut Hopp,
With Preeclampsia From Additional Migration by VEGFDownregulation of FLT-1 and Prevents Activated Neutrophils From Women Soluble Vascular Endothelial Growth Factor Receptor-1 (sFLT-1) Mediates
http://circres.ahajournals.org/cgi/content/full/01.RES.0000194324.29363.82/DC1Data Supplement (unedited) at:
http://circres.ahajournals.org
located on the World Wide Web at: The online version of this article, along with updated information and services, is
http://www.lww.com/reprintsReprints: Information about reprints can be found online at
[email protected]. E-mail:
Fax:Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters
http://circres.ahajournals.org/subscriptions/Subscriptions: Information about subscribing to Circulation Research is online at
by on May 16, 2011 circres.ahajournals.orgDownloaded from

Soluble Vascular Endothelial Growth Factor Receptor-1(sFLT-1) Mediates Downregulation of FLT-1 and Prevents
Activated Neutrophils From Women With PreeclampsiaFrom Additional Migration by VEGF
Oliver Krysiak,* Anja Bretschneider,* Enhong Zhong, Jessie Webb, Hartmut Hopp, Stefan Verlohren,Norbert Fuhr, Malgorzata Lanowska, Andreas Nonnenmacher, Roland Vetter, Joachim Jankowski,
Martin Paul, Gilbert Schonfelder
Abstract—Neutrophil activation and increased migration is associated with preeclampsia and is resolved after delivery.Preeclampsia is an inflammatory disorder where altered levels of vascular endothelial growth factor (VEGF) and thecirculating soluble fms-like tyrosine kinase 1 (sFlt-1) have a pathogenic role. VEGF, by binding to FLT-1, inducesleukocytic chemotaxis. We studied expression and function of FLT-1 in maternal neutrophils during preeclampsia andnormal pregnancies. Analysis of maternal neutrophils showed the relationship between FLT-1 expression and week ofgestation. Preeclamptic women express lower FLT-1 and sFLT-1 in neutrophils. In contrast, serum levels of sFLT-1 inpatients with preeclampsia are increased and, therefore, inhibit upregulation of FLT-1 in neutrophils by neutralizingVEGF. VEGF-dependent FLT-1 expression is regulated by changing FLT-1-promoter activity. Promoter activity isdecreased by sFLT-1. In vitro experiments demonstrated that migration of neutrophils is regulated by VEGF via FLT-1and excess of sFLT-1. Thus, VEGF-dependent migration of neutrophils is decreased during preeclampsia as aconsequence of excess circulating sFlt1. But, they still increase migration by fMLP and, therefore, migration ofneutrophils from preeclamptic women is highly activated when compared with the normotensive group. In conclusion,besides being involved in inducing an antiangiogenic state in the serum, excess of sFLT-1 seems to prevent activatedneutrophils from women with preeclampsia from additional migration by VEGF. We provide evidence that neutrophilsmay be involved in the pathophysiology of pregnancy-related hypertensive disorders. (Circ Res. 2005;97:0-0.)
Key Words: migration � neutrophils � preeclampsia � pregnancy � VEGF receptor 1
Preeclampsia (PE) is a hypertensive disorder of unknownetiology affecting 5% to 10% of all pregnancies. Patho-
physological changes include elevated systemic vascularresistance, generalized vasoconstriction, activation of thecoagulation cascade, maternal endothelial dysfunction, and apoorly perfused fetoplacental unit.1 Preeclampsia is charac-terized by altered cytokine production2 and marked neutro-phil activation.3–6 They release proteases and reactive oxygenspecies, which can cause vascular endothelial injury, inap-propriate endothelial activation or transformation.7–9 Thelatter processes are important for physiological adaptationsduring normal pregnancy creating a low-resistance arteriolarsystem, which allows adequate blood supply to the growingfetus.10,11
Recently, case-control studies have shown associationsbetween polymorphisms of the vascular endothelial growth
factor (VEGF) gene and preeclampsia.12 VEGF is a multi-functional cytokine that plays a pivotal role in angiogenesis invivo.13 VEGF is expressed by different cells, eg, smoothmuscle, endothelial cells, monocytes/macrophages, and poly-morphonuclear neutrophils (PMNs). VEGF exerts its biolog-ical effects through Fms-like tyrosine kinase 1 (FLT-1)/VEGF receptor-1 and VEGF receptor-2 (FLK-1/KDR).13
KDR is considered as the principal receptor of the VEGF-dependent angiogenic signals resulting in normal vesselgrowth and maintenance.13 In contrast, FLT-1 has recentlygained attention because of its overall implication in patho-logical angiogenesis and inflammation.13 Thus, VEGF bybinding to FLT-1, induces monocyte chemotaxis and modu-lates transendothelial PMN migration and activation.14–16
Furthermore, VEGF-induced macrophage migration isstrongly suppressed in FLT-1(TK�/�) mice.17
Original received December 13, 2004; resubmission received August 23, 2005; revised resubmission received October 6, 2005; accepted October 24,2005.
From the Institute of Clinical Pharmacology and Toxicology (O.K., A.B., E.Z., J.W., R.V., M.P., G.S.) and the Department of Gynecology andObstetrics (H.H., S.V., N.F.), Medizinische Klinik fur Endokrinologie und Nephrologie (J.J.), Charite-Universitaetsmedizin Berlin, Germany.
*Both authors contributed equally to this work.Correspondence to Dr Gilbert Schonfelder, Institute of Clinical Pharmacology and Toxicology, Campus Benjamin Franklin, Charite-
Universitatsmedizin Berlin, Garystrasse 5, 14195 Berlin, Germany. E-mail [email protected]© 2005 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/01.RES.0000194324.29363.82
1 by on May 16, 2011 circres.ahajournals.orgDownloaded from

Previous studies emphasized the physiological significanceof VEGF during pregnancy. FLT-1 is found in the blood ofpregnant but not in nonpregnant women and men. In theserum it is known as soluble FLT-1 (sFlt1).18 sFLT-1 func-tions as a “decoy” during development and prevents VEGFfrom binding to its signaling receptor KDR.19,20 This scaven-ger role might be involved in the pathogenesis of preeclamp-sia.13,21,22 Increased sFlt1 production by the preeclampticplacenta scavenges VEGF and placental growth factor(PlGF), thereby lowering circulating levels of unboundVEGF and PlGF. This altered balance causes generalizedendothelial dysfunction resulting in multi-organ disease.13 Itwas demonstrated in pregnant rats, that sFLT-1 treatmentinduces hypertension, proteinuria, and glomerularendotheliosis.21
Because of the reports about the physiological impact ofthe VEGF-system during pregnancy and its role in activatingmonocytes, we hypothesize the existence of differences in theexpression, regulation, and function of FLT-1 in maternalneutrophils between normotensive and preeclamptic pregnan-cies. We studied, therefore, the relation between expressionof FLT-1 in PMNs and sFLT-1 of healthy and preeclampticwomen. We asked how VEGF and sFLT-1 are able toregulate chemotaxis of maternal PMNs in healthy and pre-eclamptic women.
Materials and MethodsDetailed Materials and Methods are available in the online datasupplement available at http://circres.ahajournals.org.
The investigation conforms with the principles outlined in theDeclaration of Helsinki. Maternal blood was obtained during andimmediately after delivery from healthy women (n�179) and pa-tients with severe PE (n�42) at the Department of Obstetrics andwas used for further analysis. Clinical data were collected at routineobstetric visits and are summarized in the Table. All pregnantwomen were normotensive and nonproteinuric before the week 24 ofpregnancy. Preeclampsia was defined according to criteria of theAmerican College of Obstetricians and Gynecologists. Patients withother medical complications were not included.
For RT-PCR analysis and cell culture experiments, PMNs werecarefully isolated from whole blood samples immediately afterobtaining them from the women.
Flow CytometryBlood samples were tested by direct immunofluorescence staining ofwhole blood using fluorochrome-conjugated antibodies. All analyseswere performed on a FACScan flow cytometer (Becton Dickinson).The results are expressed as fluorescence histograms plotted on a logscale.
Briefly, maternal PMNs were identified based on the typicalmorphology in the forward scatter/side scatter cytogram and withmonoclonal antibodies against the neutrophil surface markersCD11B (�M- Integrin, MAC-1; Becton-Dickinson) and CD16 (FCreceptor IIIb; Immunotech) according to the manufactures instruc-tions. Antibodies against the lymphocyte surface marker CD4(Becton-Dickinson) were used as negative control for PMNs. Humanserum was used to minimize nonspecific antibody binding to Fcreceptors. Nonspecific fluorescence was assessed by substituting anonbinding isotype-matched control FITC conjugated to sheepF(ab)2 anti-mouse IgG (Sigma) for the primary mAb. A polyclonal(goat) primary antibody raised against the internal domain of FLT-1(SC-316G; Santa Cruz) was conjugated with FITC and used for flowcytometry.
ImmunofluorescenceIsolated maternal PMNs were fixed with 4% paraformaldehyde. Thecells were subsequently incubated with the primary antibody raisedagainst FLT-1 (SC-316G; Santa Cruz) and examined with a laserscanning confocal microscope (LSM-META, Carl Zeiss).
RT-PCRTotal cellular RNA was isolated from isolated maternal PMNs.Human microvascular endothelial cells were used as a positivecontrol for FLT-1 and KDR expression.
For RT-PCR analysis, first-strand cDNA was synthesized asdescribed previously.23 RT-PCR and quantitative RT-PCR (Taqman)was used to compare mRNA levels of FLT-1 and sFLT-1 in PMNsof normotensive and preeclamptic women. TaqMan analysis wasperformed according to the manufacturer’s instructions, with the useof an Applied Biosystems 7700system (PerkinElmer).
Enzyme-Linked Immunosorbent AssaySandwich enzyme-linked immunosorbent assay (ELISA) for thedetection of total sFLT-1 was performed according to manufacturer’s(RELIATech) specifications. Aliquots of serum from normotensiveand preeclamptic women were assayed in duplicate.
Cell Culture and Assays
Comparison of VEGF-Mediated FLT-1 Inductionon PMNsAccording to previous experiments with human vascular endothelialcells, where VEGF upregulates FLT-1 expression,24 we testedwhether VEGF can affect FLT-1 expression in PMNs. Isolatedmaternal PMNs were cultured. Expression of FLT-1 on PMNs wasassessed by incubating cultures with 0.6 nM recombinant VEGF165
(Biochrome AG) or preincubation with sFLT-1 (1,000pg/mL) for 2hours before incubating with VEGF or aqua bidest (negative con-trol). After 24 hours FLT-1 expression was measured by quantitativeflow cytometry.
Measurement of Neutrophil Chemotaxis inBoyden ChamberTo compare FLT-1-dependent migration of PMNs from preeclampticand healthy women, chemotaxis was assessed using multidish 6-wellcell culture plates with a 3-�m pore diameter polycarbonate mem-brane transwell apparatus (cell culture insert, NUNC) as directed bythe manufacturer. Recombinant VEGF165 at different concentrations(0.3 and 0.6 nM; Biochrome AG) or aqua bidest (negative control)was applied to the lower wells of the chamber. To study the effectsof sFLT-1 in maternal serum on PMN chemotaxis, neutrophils fromnormotensive women were preincubated with sFLT-1 (1000 pg/mL)for 2 hours before incubating with 0.6 nM VEGF, or pretreated with
Characteristics of the Normotensive Pregnant andPreeclamptic Group
CaseControl
(n�179)Preeclampsia
(n�42)
Maternal age, y 30�5 30�4
Maternal systolic blood pressure, mm Hg 113�13 150�10
Maternal diastolic blood pressure, mm Hg 68�10 93�12
Proteinuria in 24 hours, g/L — 0.30�0.02
Edema 0% 73%
Gestational age when blood sampleswere obtained, wk
35.7�3.3 34.8�3.1
Minimum 6 23
Maximum 43 41
Grava 2.0�1.3 2.4�1.7
Data denote means�SD.
2 Circulation Research December 9/23, 2005
by on May 16, 2011 circres.ahajournals.orgDownloaded from

serum from normotensive and preeclamptic women for 2 hours.After incubation for 24 hours, the cells that had migrated through thefilter were collected and counted. PMN response to other cytokineswas investigated by using N-formylmethionyl-leucyl-phenylalanine(10 ng/mL, fMLP; Sigma), a positive control for a neutrophilchemoattractant.
Transcriptional Regulation of FLT-1To examine the putative functional significance of enhanced FLT-1expression after VEGF exposure on the transcriptional level, westudied its promoter activity in HL-60 cells. Based on the sequenceof the human FLT-1 promoter [gi 1088437, D64016.1], we per-formed a genomic PCR. The resulting PCR products (1196 bp of the5�-regulatory region directly upstream of the putative translationinitiation start codon) were subcloned into the luciferase reportervector pGL3basic (Promega) and the identity of the inserts wasconfirmed by sequencing.25
HL-60 cells (DSMZ) were cultured in RPMI1640 containing 10%FBS for 72 hours. Then, HL-60 cells were suspended in serum-freeRPMI1640 medium, mixed with FLT-1- reporter-plasmid and elec-trophoresed in the Gene Pulser Xcell System (Bio-Rad). For stan-dardizing the transfection efficiency, pSV-Beta-Galactosidase vector(Promega) was cotransfected. The cells were immediately culturedeither with 0.6 nM recombinant VEGF165 or pretreated with sFLT-1(1000 pg/mL) for 2 hours before incubating with VEGF or with aquabidest (negative control) for 24 hours. We analyzed luciferase andbeta-galactosidase activity as previously described.25
Screening of the Promoter of the FLT-1 Gene forGenetic VariationsInformation about genetic variations were obtained either by data-base search in the NCBI’s dbSNP, HGMD (Human Gene MutationDatabase), and HGBASE (Human Genic Bi-Allelic Sequences) or bySingle-Strand Conformation Polymorphism (SSCP) analysis.
SSCP AnalysisGenomic DNA from all study subjects was extracted by standardtechniques.25 From the published sequences of the FLT-1 promotergene (D64016) overlapping fragments, �320 bp in length wereenzymatically amplified to cover the entire promoter region in 166healthy (332 alleles) women and 33 patients (66 alleles) withpreeclampsia.
All PCR amplifications were performed by standard techniques25
using specific primers (supplementary Table I), followed by single-strand conformation polymorphism (SSCP) analysis as previouslydescribed. DNA from patients presenting different migration patternson the polyacrylamide gels were then sequenced twice (both DNAstrands with sense and antisense primers) with the use of anautomated sequencing device (ABI PRISM 377, Perkin-Elmer).
Statistical AnalysisData analysis was performed using the Statistical Package for SocialSciences for Windows (SPSS; versions 11.0) and Sigma Plot forWindows Version 8.0 (Systat Software GmbH). Values are given asmeans�SEM if not otherwise indicated. Statistical analyses betweenthe hypertensive and control group were performed using analysis ofvariance, the Mann–Whitney test or Student t test. In addition,statistical analysis of the migration assay was preformed using theKruskal-Wallis test. P�0.05 was regarded as the level ofsignificance.
ResultsIdentification of FLT-1 in Maternal NeutrophilsAs shown in Figure 1A, forward scatter and side scatter wereused to identify maternal PMNs. Figure 1B and 1C illustratethe gates used to characterize PMNs by CD16 and CD11bexpression, whereas CD4 expression was not detected (Figure1D). Our flow cytometry revealed specific FLT-1 expression
in the maternal CD16�/CD11b� PMNs (Figure 1E and 1F).Substituting control FITC conjugated to sheep F(ab)2 anti-mouse IgG instead of primary antibody revealed no immu-nofluorescence in the flow cytometry (Figure 1G).
Immunohistochemistry demonstrated specific staining forFLT-1 in the cytoplasm and cell membrane of PMNs (Figure1H) by using laser scanning confocal microscopy.
Specificity of the immunoreactions used for both methodswas assessed by using different amounts of primary antibody(data not shown) and peptide pre-absorbed antiserum againstFLT-1-peptide and antiserum against CD11b pre-absorbedwith FLT-1-peptide (supplemental Figure I).
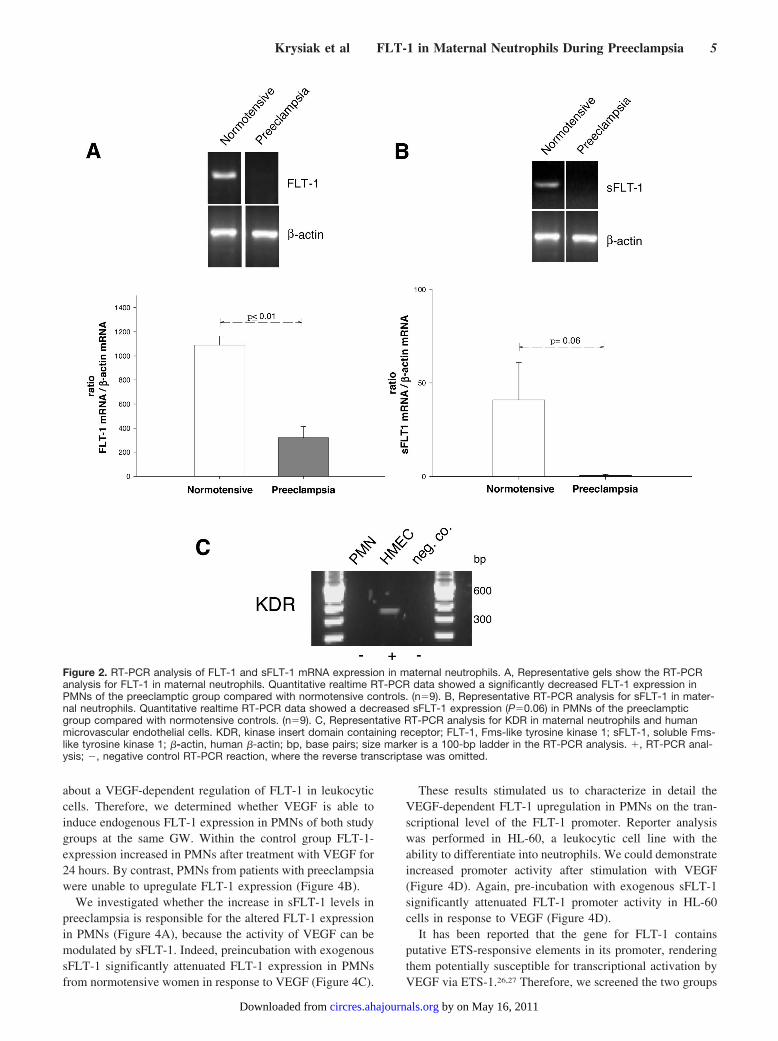
Characterization of FLT-1 Expression in MaternalNeutrophils of Hypertensive PregnanciesFigure 2A and 2B present FLT-1, sFLT-1 and KDR expres-sion on mRNA level in maternal PMNs of healthy womenwith the preeclamptic pregnancies. RT-PCR analysis revealedexpression of FLT-1 and sFLT-1 only (Figure 2). Omission ofthe reverse transcriptase from the reaction mixture resulted inno amplification (data not presented). Sequence analysis ofthe obtained KDR, FLT-1, and sFLT-1 RT-PCR productsconfirmed total homology with the published human mRNAsequences.
Quantitative results of FLT-1 and sFLT-1 mRNA levels byrealtime RT-PCR is given in Figure 2. Statistical analysis ofthe data showed a significantly decreased FLT-1 and sFLT-1expression in PMNs of the preeclamptic group comparedwith normotensive controls, when related to �-actin mRNAexpression.
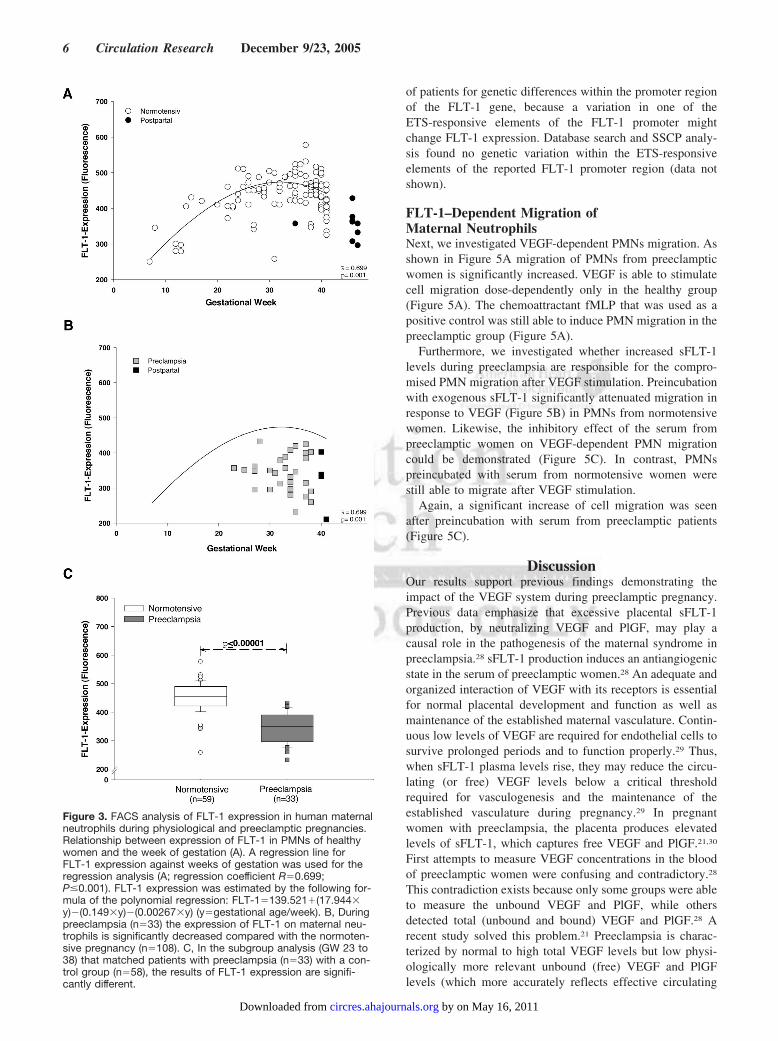
Furthermore, FLT-1 protein expression in maternal PMNsfrom 179 healthy women were studied for between gesta-tional week (GW) 6 and 42 and compared with the pre-eclamptic group between GW 23 and 41 using FACS analy-sis. Additionally, PMNs from 9 healthy women were studied24 hours after giving birth. The plot in Figure 3A demon-strates the relationship between FLT-1 expression and weekof gestation in healthy women. FLT-1 expression increasedduring the first two trimesters, peaked at 30 to 33 weeks, anddecreased thereafter with the lowest expression after givingbirth (Figure 3A).
In contrast, PMNs from patients with preeclampsia ex-pressed significantly lower FLT-1 levels (Figure 3B) whencompared with the healthy group (Figure 3B). In subgroupanalysis in patients with and without preeclampsia ofmatched gestational age, FLT-1 expression in the PMNs wassignificantly lower in preeclamptic women (Figure 3C) ascompared with healthy women.
Another subgroup analysis (gestational week 30 and 38)compared FLT-1 expression with sFLT-1 serum levels duringnormotensive and preeclamptic pregnancies (Figure 4A).Soluble FLT-1 serum concentrations are elevated in the PEgroup when compared with the normotensive women. Incontrast, FLT-1 expression in PMNs is decreased in the PEgroup.
FLT-1 Expression in Maternal PMNs AfterVEGF StimulationIt was demonstrated that VEGF exposure leads to higherlevels of FLT-1 in cultured ECs,24 but nothing was known
Krysiak et al FLT-1 in Maternal Neutrophils During Preeclampsia 3
by on May 16, 2011 circres.ahajournals.orgDownloaded from
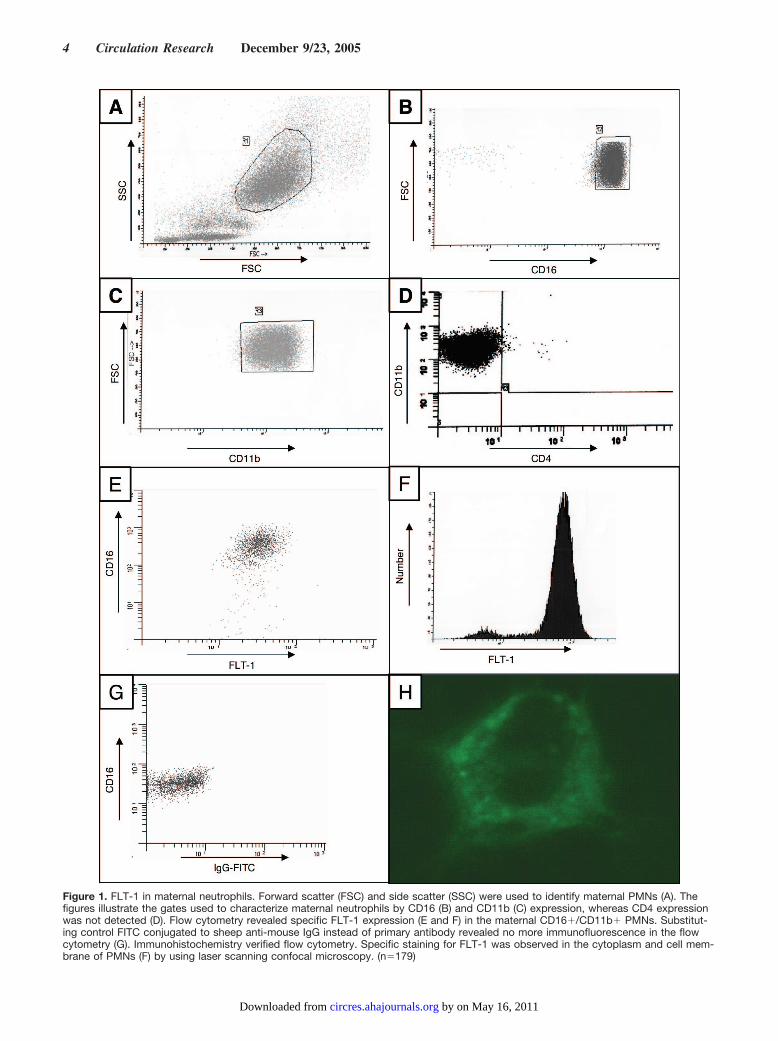
Figure 1. FLT-1 in maternal neutrophils. Forward scatter (FSC) and side scatter (SSC) were used to identify maternal PMNs (A). Thefigures illustrate the gates used to characterize maternal neutrophils by CD16 (B) and CD11b (C) expression, whereas CD4 expressionwas not detected (D). Flow cytometry revealed specific FLT-1 expression (E and F) in the maternal CD16�/CD11b� PMNs. Substitut-ing control FITC conjugated to sheep anti-mouse IgG instead of primary antibody revealed no more immunofluorescence in the flowcytometry (G). Immunohistochemistry verified flow cytometry. Specific staining for FLT-1 was observed in the cytoplasm and cell mem-brane of PMNs (F) by using laser scanning confocal microscopy. (n�179)
4 Circulation Research December 9/23, 2005
by on May 16, 2011 circres.ahajournals.orgDownloaded from
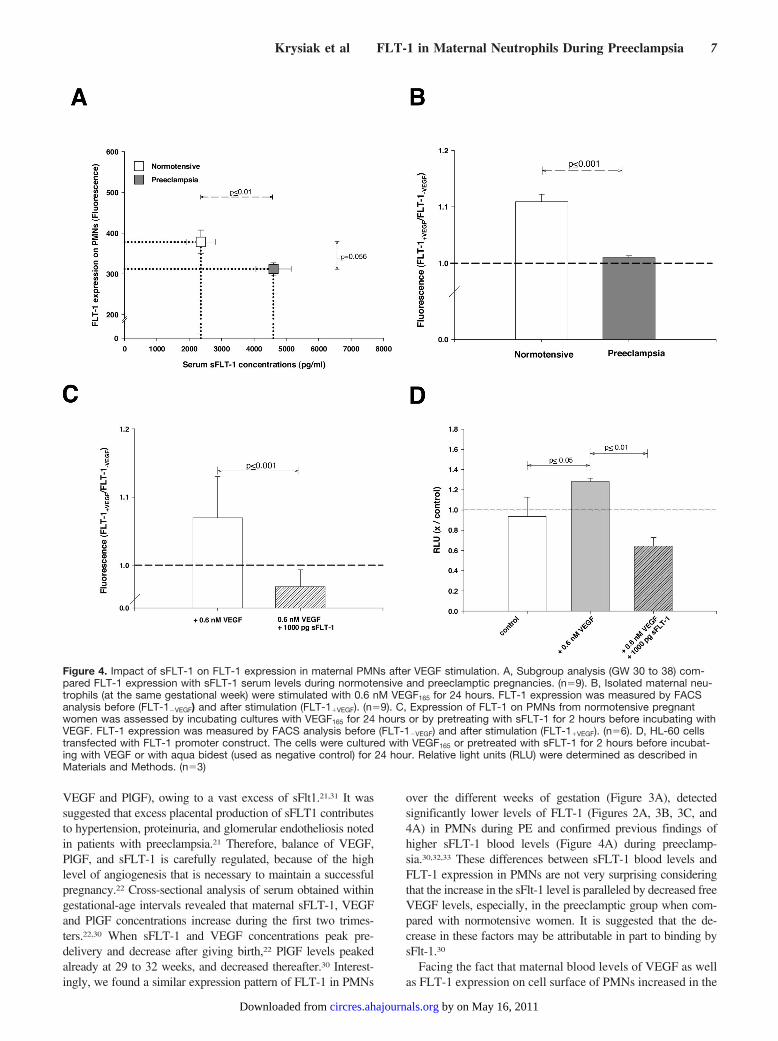
about a VEGF-dependent regulation of FLT-1 in leukocyticcells. Therefore, we determined whether VEGF is able toinduce endogenous FLT-1 expression in PMNs of both studygroups at the same GW. Within the control group FLT-1-expression increased in PMNs after treatment with VEGF for24 hours. By contrast, PMNs from patients with preeclampsiawere unable to upregulate FLT-1 expression (Figure 4B).
We investigated whether the increase in sFLT-1 levels inpreeclampsia is responsible for the altered FLT-1 expressionin PMNs (Figure 4A), because the activity of VEGF can bemodulated by sFLT-1. Indeed, preincubation with exogenoussFLT-1 significantly attenuated FLT-1 expression in PMNsfrom normotensive women in response to VEGF (Figure 4C).
These results stimulated us to characterize in detail theVEGF-dependent FLT-1 upregulation in PMNs on the tran-scriptional level of the FLT-1 promoter. Reporter analysiswas performed in HL-60, a leukocytic cell line with theability to differentiate into neutrophils. We could demonstrateincreased promoter activity after stimulation with VEGF(Figure 4D). Again, pre-incubation with exogenous sFLT-1significantly attenuated FLT-1 promoter activity in HL-60cells in response to VEGF (Figure 4D).
It has been reported that the gene for FLT-1 containsputative ETS-responsive elements in its promoter, renderingthem potentially susceptible for transcriptional activation byVEGF via ETS-1.26,27 Therefore, we screened the two groups
Figure 2. RT-PCR analysis of FLT-1 and sFLT-1 mRNA expression in maternal neutrophils. A, Representative gels show the RT-PCRanalysis for FLT-1 in maternal neutrophils. Quantitative realtime RT-PCR data showed a significantly decreased FLT-1 expression inPMNs of the preeclamptic group compared with normotensive controls. (n�9). B, Representative RT-PCR analysis for sFLT-1 in mater-nal neutrophils. Quantitative realtime RT-PCR data showed a decreased sFLT-1 expression (P�0.06) in PMNs of the preeclampticgroup compared with normotensive controls. (n�9). C, Representative RT-PCR analysis for KDR in maternal neutrophils and humanmicrovascular endothelial cells. KDR, kinase insert domain containing receptor; FLT-1, Fms-like tyrosine kinase 1; sFLT-1, soluble Fms-like tyrosine kinase 1; �-actin, human �-actin; bp, base pairs; size marker is a 100-bp ladder in the RT-PCR analysis. �, RT-PCR anal-ysis; �, negative control RT-PCR reaction, where the reverse transcriptase was omitted.
Krysiak et al FLT-1 in Maternal Neutrophils During Preeclampsia 5
by on May 16, 2011 circres.ahajournals.orgDownloaded from
of patients for genetic differences within the promoter regionof the FLT-1 gene, because a variation in one of theETS-responsive elements of the FLT-1 promoter mightchange FLT-1 expression. Database search and SSCP analy-sis found no genetic variation within the ETS-responsiveelements of the reported FLT-1 promoter region (data notshown).
FLT-1–Dependent Migration ofMaternal NeutrophilsNext, we investigated VEGF-dependent PMNs migration. Asshown in Figure 5A migration of PMNs from preeclampticwomen is significantly increased. VEGF is able to stimulatecell migration dose-dependently only in the healthy group(Figure 5A). The chemoattractant fMLP that was used as apositive control was still able to induce PMN migration in thepreeclamptic group (Figure 5A).
Furthermore, we investigated whether increased sFLT-1levels during preeclampsia are responsible for the compro-mised PMN migration after VEGF stimulation. Preincubationwith exogenous sFLT-1 significantly attenuated migration inresponse to VEGF (Figure 5B) in PMNs from normotensivewomen. Likewise, the inhibitory effect of the serum frompreeclamptic women on VEGF-dependent PMN migrationcould be demonstrated (Figure 5C). In contrast, PMNspreincubated with serum from normotensive women werestill able to migrate after VEGF stimulation.
Again, a significant increase of cell migration was seenafter preincubation with serum from preeclamptic patients(Figure 5C).
DiscussionOur results support previous findings demonstrating theimpact of the VEGF system during preeclamptic pregnancy.Previous data emphasize that excessive placental sFLT-1production, by neutralizing VEGF and PlGF, may play acausal role in the pathogenesis of the maternal syndrome inpreeclampsia.28 sFLT-1 production induces an antiangiogenicstate in the serum of preeclamptic women.28 An adequate andorganized interaction of VEGF with its receptors is essentialfor normal placental development and function as well asmaintenance of the established maternal vasculature. Contin-uous low levels of VEGF are required for endothelial cells tosurvive prolonged periods and to function properly.29 Thus,when sFLT-1 plasma levels rise, they may reduce the circu-lating (or free) VEGF levels below a critical thresholdrequired for vasculogenesis and the maintenance of theestablished vasculature during pregnancy.29 In pregnantwomen with preeclampsia, the placenta produces elevatedlevels of sFLT-1, which captures free VEGF and PlGF.21,30
First attempts to measure VEGF concentrations in the bloodof preeclamptic women were confusing and contradictory.28
This contradiction exists because only some groups were ableto measure the unbound VEGF and PlGF, while othersdetected total (unbound and bound) VEGF and PlGF.28 Arecent study solved this problem.21 Preeclampsia is charac-terized by normal to high total VEGF levels but low physi-ologically more relevant unbound (free) VEGF and PlGFlevels (which more accurately reflects effective circulating
Figure 3. FACS analysis of FLT-1 expression in human maternalneutrophils during physiological and preeclamptic pregnancies.Relationship between expression of FLT-1 in PMNs of healthywomen and the week of gestation (A). A regression line forFLT-1 expression against weeks of gestation was used for theregression analysis (A; regression coefficient R�0.699;P�0.001). FLT-1 expression was estimated by the following for-mula of the polynomial regression: FLT-1�139.521�(17.944�y)�(0.149�y)�(0.00267�y) (y�gestational age/week). B, Duringpreeclampsia (n�33) the expression of FLT-1 on maternal neu-trophils is significantly decreased compared with the normoten-sive pregnancy (n�108). C, In the subgroup analysis (GW 23 to38) that matched patients with preeclampsia (n�33) with a con-trol group (n�58), the results of FLT-1 expression are signifi-cantly different.
6 Circulation Research December 9/23, 2005
by on May 16, 2011 circres.ahajournals.orgDownloaded from
VEGF and PlGF), owing to a vast excess of sFlt1.21,31 It wassuggested that excess placental production of sFLT1 contributesto hypertension, proteinuria, and glomerular endotheliosis notedin patients with preeclampsia.21 Therefore, balance of VEGF,PlGF, and sFLT-1 is carefully regulated, because of the highlevel of angiogenesis that is necessary to maintain a successfulpregnancy.22 Cross-sectional analysis of serum obtained withingestational-age intervals revealed that maternal sFLT-1, VEGFand PlGF concentrations increase during the first two trimes-ters.22,30 When sFLT-1 and VEGF concentrations peak pre-delivery and decrease after giving birth,22 PlGF levels peakedalready at 29 to 32 weeks, and decreased thereafter.30 Interest-ingly, we found a similar expression pattern of FLT-1 in PMNs
over the different weeks of gestation (Figure 3A), detectedsignificantly lower levels of FLT-1 (Figures 2A, 3B, 3C, and4A) in PMNs during PE and confirmed previous findings ofhigher sFLT-1 blood levels (Figure 4A) during preeclamp-sia.30,32,33 These differences between sFLT-1 blood levels andFLT-1 expression in PMNs are not very surprising consideringthat the increase in the sFlt-1 level is paralleled by decreased freeVEGF levels, especially, in the preeclamptic group when com-pared with normotensive women. It is suggested that the de-crease in these factors may be attributable in part to binding bysFlt-1.30
Facing the fact that maternal blood levels of VEGF as wellas FLT-1 expression on cell surface of PMNs increased in the
Figure 4. Impact of sFLT-1 on FLT-1 expression in maternal PMNs after VEGF stimulation. A, Subgroup analysis (GW 30 to 38) com-pared FLT-1 expression with sFLT-1 serum levels during normotensive and preeclamptic pregnancies. (n�9). B, Isolated maternal neu-trophils (at the same gestational week) were stimulated with 0.6 nM VEGF165 for 24 hours. FLT-1 expression was measured by FACSanalysis before (FLT-1�VEGF) and after stimulation (FLT-1�VEGF). (n�9). C, Expression of FLT-1 on PMNs from normotensive pregnantwomen was assessed by incubating cultures with VEGF165 for 24 hours or by pretreating with sFLT-1 for 2 hours before incubating withVEGF. FLT-1 expression was measured by FACS analysis before (FLT-1�VEGF) and after stimulation (FLT-1�VEGF). (n�6). D, HL-60 cellstransfected with FLT-1 promoter construct. The cells were cultured with VEGF165 or pretreated with sFLT-1 for 2 hours before incubat-ing with VEGF or with aqua bidest (used as negative control) for 24 hour. Relative light units (RLU) were determined as described inMaterials and Methods. (n�3)
Krysiak et al FLT-1 in Maternal Neutrophils During Preeclampsia 7
by on May 16, 2011 circres.ahajournals.orgDownloaded from
same manner during the first two trimesters30 (Figure 3A), wehave to remember a recent study showing increased FLT-1expression in endothelial cells after VEGF exposure.24 There-fore, we became interested to understand whether VEGF isalso able to induce FLT-1 expression in maternal PMNsduring normal and preeclamptic pregnancies. Increased FLT-1-expression in normal PMNs was found after incubationwith VEGF conditioned medium (Figure 4B). In contrast,PMNs from preeclamptic patients were unable to upregulateFLT-1 expression (Figure 4B). Thus, we addressed whetherthe elevated sFLT-1 in patients with preeclampsia inhibitsupregulation of FLT-1 in neutrophils by neutralizing VEGF.Indeed, our experiments demonstrated that sFLT-1 inhibitsVEGF-dependent upregulation of FLT-1 (Figure 4C) bychanging its promoter activity (Figure 4D), because the genefor FLT-1 contains putative ETS-responsive elements in itspromoter, rendering them potentially susceptible for tran-scriptional activation by VEGF via ETS-1.27 These resultsstimulated us to screen the two groups of patients for geneticdifferences within the promoter region of the FLT-1 gene. Infact, a variation in one of the ETS-responsive elements of theFLT-1 promoter could be responsible for changes of FLT-1expression. Because the general catalog of genome variationwith the complete contents of dbSNP, HGMD, and HGBASEis available, we can easily screen those databases for pub-lished genetic variations. However, our bioinformatics anal-ysis failed to find any kind of genetic variation within thewhole FLT-1 promoter region (data not shown). Genotypingof the patients’ DNA by SSCP analysis and additionalsequencing confirmed that there are no variations in theETS-responsive elements (data not shown). Nevertheless,future studies performing large-scale associations with pro-moter analysis of genetic variations of the ETS-1 gene shouldbe initiated for a better understanding of the impact of ETS-1on the transcriptional regulation of FLT-1 during hyperten-sive pregnancies.
So far, the functional consequence of the attenuated FLT-1expression in PMNs during preeclampsia remained unsolved.We knew that resting monocytes normally express low levelsof FLT-1;15 and VEGF, by binding to FLT-1, inducesmonocyte chemotaxis and modulates transendothelial migra-tion.14–16 Our results confirmed that FLT-1 is involved inVEGF-induced chemotaxis of peripheral blood cells,14 be-cause these cells express only FLT-1 but not KDR (Figure 2Aand 2C) and migrate in response to exogenous VEGF (Figure5A and 5B). Thus, when FLT-1 is downregulated in pre-eclamptic PMNs no VEGF-dependent chemotaxis could beobserved (Figure 5A), although fMLP (a standard chemokineused as a positive control) was still able to induce normalPMNs migration. Furthermore, sFLT-1 (Figure 5B) or me-dium conditioned with preeclamptic serum rich in sFLT-1(Figure 5C) decreased neutrophil migration by neutralizingVEGF. These findings are in accordance with recent reportsshowing decreased endothelial cell migration after incubationwith sFLT-1 or conditioned medium from preeclampticplacenta.34
How does the decreased FLT-1 expression in neutrophilsof preeclamptic women fit into the understanding of thepathophysiology of the maternal syndrome of preeclampsia?
Figure 5. Neutrophil chemotaxis in response to VEGF, sFLT-1and fMLP. A, PMNs from control (n�7) and preeclamptic (n�6)women at same gestational age were exposed to medium withdifferent doses of VEGF (0.3 and 0.6 nM) and aqua bidest (neg-ative control). Neutrophil chemotactic response was expressedas percentage of migrated PMNs. Each result represents anaverage of two independent experiments per patient. B, VEGF-induced migration of PMNs from normotensive women (n�3)with and without preincubation of sFLT-1. (Relative cell migra-tion; migration�VEGF/migration�VEGF). C, VEGF-induced migrationof PMNs from healthy pregnant women (n�3) after preincuba-tion with serum from normotensive and preeclamptic women.
8 Circulation Research December 9/23, 2005
by on May 16, 2011 circres.ahajournals.orgDownloaded from

In general, pregnancy is associated with a generalized inflam-matory response.35 One reason is the maintenance of theperipheral tolerance to fetal alloantigen. Thus, during gesta-tion physiological adaptation of innate and adaptive immunesystem is a necessary key element for maternal homeostasis.35
PMNs and monocytes as parts of the innate immune systemappear to be the most profoundly affected cells as shown bydistinct changes in their phenotype and function. In particu-lar, proper receptor-driven cell functions35 such as apoptosisand VEGF/FLT1-dependent chemotaxis are required.
Compared with normal pregnancy, the systemic inflamma-tory response is much more pronounced during preeclamp-sia.36 The reason for this extensive systemic inflammationremains unknown. It is believed that a hypoxic and dysfunc-tional preeclamptic placenta releases increased amounts ofproinflammatory components: plasma factors, like IL-6,TNF-�, and sFLT-1,8 cell debris, such as syncytiotrophoblastmembrane microparticles, and even cytotrophoblasts into thematernal circulation. Increased plasma levels of sFLT-1,besides other released proinflammatory factors, then alterneutrophil function (Figures 4 and 5).
We therefore speculate that the consequence of this pla-cental disposal plus the impaired VEGF response of neutro-phils (Figures 4B and 5A) increases the maternal inflamma-tory burden.36 This leads to a higher PMNs activation and, inturn, a potentiated interaction with the vascular endotheliumresulting in increased infiltration of the maternal systemicvasculature,37 and contributes to vascular endothelial dys-function, which is central for the pathophysiology of pre-eclampsia. Indeed, our results based on absolute cell migra-tion measurements revealed a significantly increasedmigration of PMNs from preeclamptic women when com-pared with controls (Figure 5A). Furthermore, medium con-ditioned with preeclamptic serum increased even migration ofPMNs from normotensive pregnant women (Figure 5C). Thiscould be attributable to increased plasma concentrations ofcytokines, such as TNF-�.8
In summary, increased sFLT-1 appears to alter the neutro-phil function resulting in loss of VEGF-dependent migrationduring preeclampsia. Moreover, sFLT-1 appears to down-regulate FLT-1 expression in PMNs because of its VEGFneutralizing action.
AcknowledgmentsResearch was supported by the Charite-Universitaetsmedizin Berlin.The authors thank Doris Webb for critically reading ofthis manuscript.
References1. Dekker GA, Sibai BM. Etiology and pathogenesis of preeclampsia:
current concepts. Am J Obstet Gynecol. 1998;179:1359–1375.2. Conrad KP, Miles TM, Benyo DF. Circulating levels of immunoreactive
cytokines in women with preeclampsia. Am J Reprod Immunol. 1998;40:102–111.
3. Greer IA, Haddad NG, Dawes J, Johnstone FD, Calder AA. Neutrophilactivation in pregnancy-induced hypertension. Br J Obstet Gynaecol.1989;96:978–982.
4. Clark P, Boswell F, Greer IA. The neutrophil and preeclampsia. SeminReprod Endocrinol. 1998;16:57–64.
5. Greer IA, Dawes J, Johnston TA, Calder AA. Neutrophil activation isconfined to the maternal circulation in pregnancy-induced hypertension.Obstet Gynecol. 1991;78:28–32.
6. Mellembakken JR, Hogasen K, Mollnes TE, Hack CE, Abyholm T,Videm V. Increased systemic activation of neutrophils but not com-plement in preeclampsia. Obstet Gynecol. 2001;97:371–374.
7. Wang Y, Gu Y, Zhang Y, Lewis DF. Evidence of endothelial dysfunctionin preeclampsia: decreased endothelial nitric oxide synthase expression isassociated with increased cell permeability in endothelial cells frompreeclampsia. Am J Obstet Gynecol. 2004;190:817–824.
8. Lee VM, Quinn PA, Jennings SC, Ng LL. Neutrophil activation andproduction of reactive oxygen species in pre-eclampsia. J Hypertens.2003;21:395–402.
9. Wang Y, Adair CD, Coe L, Weeks JW, Lewis DF, Alexander JS.Activation of endothelial cells in preeclampsia: increased neutrophil-en-dothelial adhesion correlates with up-regulation of adhesion moleculeP-selectin in human umbilical vein endothelial cells isolated from pre-eclampsia. J Soc Gynecol Investig. 1998;5:237–243.
10. Sibai B, Dekker G, Kupferminc M Pre-eclampsia. Lancet. 2005;26:365:785–799.
11. Wang Y, Gu Y, Philibert L, Lucas MJ. Neutrophil activation induced byplacental factors in normal and pre-eclamptic pregnancies in vitro.Placenta. 2001;22:560–565.
12. Brenchley PE, Ralph SA, Summers A. VEGF polymorphisms and renalpathologies. Advances in nephrology from the Necker Hospital. 2002;32:17–23.
13. Luttun A, Autiero M, Tjwa M, Carmeliet P Genetic dissection of tumorangiogenesis: are PlGF and VEGFR-1 novel anti-cancer targets? BiochimBiophys Acta. 2004;4:1654:79–94.
14. Ancelin M, Chollet-Martin S, Herve MA, Legrand C, El Benna J, Perrot-Applanat M. Vascular endothelial growth factor VEGF189 induceshuman neutrophil chemotaxis in extravascular tissue via an autocrineamplification mechanism. Lab Invest. 2004;84:502–512.
15. Barleon B, Sozzani S, Zhou D, Weich HA, Mantovani A, Marme D.Migration of human monocytes in response to vascular endothelialgrowth factor (VEGF) is mediated via the VEGF receptor flt-1. Blood.1996;87:3336–3343.
16. Clauss M, Weich H, Breier G, Knies U, Rockl W, Waltenberger J, RisauW. The vascular endothelial growth factor receptor Flt-1 mediates bio-logical activities. Implications for a functional role of placenta growthfactor in monocyte activation and chemotaxis. J Biol Chem. 1996;271:17629–17634.
17. Hiratsuka S, Minowa O, Kuno J, Noda T, Shibuya M Flt-1 lacking thetyrosine kinase domain is sufficient for normal development and angio-genesis in mice. Proc Natl Acad Sci U S A. 1998;4:95:9349–9354.
18. Banks RE, Forbes MA, Searles J, Pappin D, Canas B, Rahman D,Kaufmann S, Walters CE, Jackson A, Eves P, Linton G, Keen J, WalkerJJ, Selby PJ. Evidence for the existence of a novel pregnancy-associatedsoluble variant of the vascular endothelial growth factor receptor, Flt-1.Mol Hum Reprod. 1998;4:377–386.
19. Carmeliet P, Moons L, Luttun A, Vincenti V, Compernolle V, De Mol M,Wu Y, Bono F, Devy L, Beck H, Scholz D, Acker T, DiPalma T,Dewerchin M, Noel A, Stalmans I, Barra A, Blacher S, VandendriesscheT, Ponten A, Eriksson U, Plate KH, Foidart JM, Schaper W,Charnock-Jones DS, Hicklin DJ, Herbert JM, Collen D, Persico MG.Synergism between vascular endothelial growth factor and placentalgrowth factor contributes to angiogenesis and plasma extravasation inpathological conditions. Nat Med. 2001;7:575–583.
20. Clauss M. Functions of the VEGF Receptor-1 (FLT-1) in the Vasculature.Trends Cardiovasc Med. 1998;8:241–245.
21. Maynard SE, Min JY, Merchan J, Lim KH, Li J, Mondal S, LibermannTA, Morgan JP, Sellke FW, Stillman IE, Epstein FH, Sukhatme VP,Karumanchi SA. Excess placental soluble fms-like tyrosine kinase 1(sFlt1) may contribute to endothelial dysfunction, hypertension, and pro-teinuria in preeclampsia. J Clin Invest. 2003;111:649–658.
22. McKeeman GC, Ardill JE, Caldwell CM, Hunter AJ, McClure N. Solublevascular endothelial growth factor receptor-1 (sFlt-1) is increasedthroughout gestation in patients who have preeclampsia develop. Am JObstet Gynecol. 2004;191:1240–1246.
23. Schonfelder G, John M, Hopp H, Fuhr N, van-Der-Giet M, Paul M.Expression of inducible nitric oxide synthase in placenta of women withgestational diabetes. Faseb J. 1996;10:777–784.
24. Barleon B, Siemeister G, Martiny-Baron G, Weindel K, Herzog C,Marme D Vascular endothelial growth factor up-regulates its receptorfms-like tyrosine kinase 1 (FLT-1) and a soluble variant of FLT-1 inhuman vascular endothelial cells. Cancer Res. 1997;1:57:5421–5425.
25. Funke-Kaiser H, Reichenberger F, Kopke K, Herrmann SM, Pfeifer J,Orzechowski HD, Zidek W, Paul M, Brand E Differential binding of
Krysiak et al FLT-1 in Maternal Neutrophils During Preeclampsia 9
by on May 16, 2011 circres.ahajournals.orgDownloaded from

transcription factor E2F-2 to the endothelin-converting enzyme-1bpromoter affects blood pressure regulation. Hum Mol Genet. 2003;15:423–433.
26. Valter MM, Hugel A, Huang HJ, Cavenee WK, Wiestler OD, Pietsch T,Wernert N Expression of the Ets-1 transcription factor in human astro-cytomas is associated with Fms-like tyrosine kinase-1 (Flt-1)/vascularendothelial growth factor receptor-1 synthesis and neoangiogenesis.Cancer Res. 1999;1:5608–5614.
27. Wakiya K, Begue A, Stehelin D, Shibuya M A cAMP response elementand an Ets motif are involved in the transcriptional regulation of flt-1tyrosine kinase (vascular endothelial growth factor receptor 1) gene.J Biol Chem. 1996;29:30823–30828.
28. Levine RJ, Karumanchi SA. Circulating angiogenic factors in pre-eclampsia. Clin Obstet Gynecol. 2005;48:372–386.
29. Luttun A, Carmeliet P. Soluble VEGF receptor Flt1: the elusive pre-eclampsia factor discovered? J Clin Invest. 2003;111:600–602.
30. Levine RJ, Maynard SE, Qian C, Lim KH, England LJ, Yu KF,Schisterman EF, Thadhani R, Sachs BP, Epstein FH, Sibai BM, SukhatmeVP, Karumanchi SA Circulating angiogenic factors and the risk of pre-eclampsia. N Engl J Med. 2004;12:672–683.
31. Jelkmann W. Pitfalls in the measurement of circulating vascular endo-thelial growth factor. Clin Chem. 2001;47:617–623.
32. Chung JY, Song Y, Wang Y, Magness RR, Zheng J. Differentialexpression of vascular endothelial growth factor (VEGF), endocrinegland derived-VEGF, and VEGF receptors in human placentas fromnormal and preeclamptic pregnancies. J Clin Endocrinol Metab. 2004;89:2484–2490.
33. Helske S, Vuorela P, Carpen O, Hornig C, Weich H, Halmesmaki E.Expression of vascular endothelial growth factor receptors 1, 2 and 3 inplacentas from normal and complicated pregnancies. Mol Hum Reprod.2001;7:205–210.
34. Ahmad S, Ahmed A. Elevated placental soluble vascular endothelialgrowth factor receptor-1 inhibits angiogenesis in preeclampsia. Circ Res.2004;95:884–891.
35. Luppi P How immune mechanisms are affected by pregnancy. Vaccine.2003;28:3352–3357.
36. Redman CW, Sargent IL Latest advances in understanding preeclampsia.Science. 2005;10:1592–1594.
37. Leik CE, Walsh SW. Neutrophils infiltrate resistance-sized vessels ofsubcutaneous fat in women with preeclampsia. Hypertension. 2004;44:72–77.
10 Circulation Research December 9/23, 2005
by on May 16, 2011 circres.ahajournals.orgDownloaded from

Online Data Supplement Krysiak et al.
1
Expanded Materials and Methods
For RT-PCR analysis and cell culture experiments PMNs were carefully isolated from whole
blood samples immediately after obtaining them from the women. Neutrophils (>95% purity,
>95% viability ± 2 as per trypan blue and propidiumiodid exclusion) were purified over a
gradient using Polymorphprep® (AXIS-SHIELD PoC AS, Norway) and characterized by flow
cytometry for CD4, CD11b, CD16 and FLT-1 expression.
Flow cytometry
Blood samples were tested by direct immunofluorescence staining of whole blood using
fluorochrome-conjugated antibodies. All analyses were performed on a FACScan flow
cytometer (Becton Dickinson, Mountain View, USA). The results were expressed as
fluorescence histograms plotted on a log scale. Compensation and PMT voltages were
standardized before each run using CalibriteTM beads (Becton Dickinson, Germany).
Maternal PMNs were identified based on the typical morphology in the FSC/SSC cytogram
and with monoclonal antibodies against neutrophil surface markers conjugated with
fluorochromes phycoerithrin for CD11B (αM- Integrin, MAC-1; Becton-Dickinson, Germany)
and PC5 for CD16 (FC receptor IIIb; Immunotech, Germany) according to the manufactures
instructions. Antibodies against the lymphocyte surface marker CD4, conjugated with FITC
(Becton-Dickinson, Germany), were used as negative control for PMNs. Anti-mouse IgG1,
conjugated with PE or FITC was used as isotype control. For the procedure, 10 µl of
fluorochrome-conjugated antibody was added to 50 µl of whole blood according to the
manufactures protocol.
Different monoclonal antibodies against the external domain of FLT-1 from commercial
source (V4262, Sigma Aldrich, Germany) or donated by A. Menrad (Schering AG, Germany)
were tested for characterization of FLT-1 expression in maternal PMNs. Binding was
assessed either by the colour indirect method performing a secondary detection with FITC
conjugated to sheep F(ab)2 anti-mouse IgG (Sigma, Germany) or direct FITC conjugated
by on May 16, 2011 circres.ahajournals.orgDownloaded from

Online Data Supplement Krysiak et al.
2
primary antibodies against FLT-1. Human serum was used to minimize nonspecific antibody
binding to Fc receptors. Nonspecific fluorescence was assessed by substituting a nonbinding
isotype-matched control FITC conjugated to sheep F(ab)2 anti-mouse IgG (Sigma, Germany)
for the primary mAb. Nevertheless, nonspecific binding to the Fc receptors on PMNs could
not be prevented.
A polyclonal (goat) primary antibody raised against the internal domain of FLT-1 (SC-316G;
Santa Cruz, Germany) was conjugated with FITC by using The Zenon Goat IgG Labeling Kit
(Molecular Probes Europe BV, The Netherlands) and used for flow cytometry. Briefly, for the
intracellular staining, the CD11b + / CD16 + cells were permeabilized, washed with Cellwash
(BD Biosciences Pharmingen, Germany), and stained sequentially with FITC-conjugated
polyclonal antibody against FLT-1 containing Lysing solution G (BD Biosciences
Pharmingen, Germany) at 4°C for 30 min.
Specificity of the obtained immunoreactions was assessed by using different amounts of first
antibody against FLT-1, peptide pre-absorbed antiserum against FLT-1-peptide (SC-316P;
Santa Cruz, Germany; 2:1 and 1:1 competition for 30 min at room temperature), peptide pre-
absorbtion of antiserum against CD11b with FLT-1-peptide (negative control) or substituting
control FITC conjugated to sheep F(ab)2 anti-mouse IgG (Sigma Aldrich, Germany) instead
of primary antibody.
All samples were tested by quantitative flow cytometry for FLT-1- and CD16b-expression.
The intensity levels for all fluorescence markers were standardized for each run using
Calibrite beads (Becton-Dickinson Biosciences, Germany). The mean channel fluorescence
(MCF) for each conjugated antibody binding to neutrophils was determined by comparing
sample MCF of four levels of calibrated beads containing a known number of fluorochrome
molecules per bead (Quantibrite Beads, Becton-Dickinson Biosciences, Germany). An MCF
ratio of 101 or higher was voted as a "positive" reaction. This value invariably exceeded that
obtained with normal control serum by at least 3.0 SD. Analysis was performed using the
quantitative calibration option of “Win List” (Verity Software House, United States of
America).
by on May 16, 2011 circres.ahajournals.orgDownloaded from

Online Data Supplement Krysiak et al.
3
Immunofluorescence
Isolated maternal PMNs were fixed with fresh 4% paraformaldehyde, rinsed in phosphate-
buffered saline and then incubated for 1 h in blocking buffer (PBS containing 1% BSA and
10% goat serum; DAKO, Germany). The cells were subsequently incubated with the
polyclonal primary antibody raised against the internal domain of FLT-1, diluted in PBS
containing 0.1 % Triton X-100 at a concentration of 1:100. Cells were washed with PBS, and
incubated with a secondary FITC-conjugated anti-goat IgG Ab (Jackson ImmunoResearch
Europe Ltd, UK) at a dilution of 1:200 in PBS containing 0.1 % Triton X-100. Subsequentley,
cells were washed, and mounted to be examined with a laser scanning confocal microscope
(LSM-META, Carl Zeiss, Germany).
Negative control reactions for FLT-1-immunostaining were conducted on isolated PMNs with
substitution of either PBS or normal goat serum or preabsorbtion of the primary antibodiy
with an excess of FLT-1 peptide (SC-316P; Santa Cruz, Germany).
Reverse transcription-polymerase chain reaction (RT-PCR)
Total cellular RNA was isolated from isolated maternal CD11+/ CD16+/ CD4-/ FLT-1+ PMNs
using TRIZOL reagent (Invitrogen, Germany). Human microvascular endothelial cells were
used as a positive control for FLT-1 and KDR expression.
For RT-PCR analysis, first-strand cDNA was synthesized as described previously 1.
Control PCR assays were performed to estimate cDNA amount, as well as RT efficiency and
cDNA quality, using specific primers to human β-actin 2. Reactions were carried out as
previously described 1.
RT-PCR and quantitative RT-PCR (Taqman) was used to compare mRNA levels of FLT-1 in
PMNs of normotensive and preeclamptic women. Primer and probes (FLT-1 forward: 5´-
TTCCTTTGGATGAGCAGTGTGA-3´ and reverse: 3´-GTGGCCAATGTGGGTCAAGA-5´,
probe:5-6-FAMTGATGCCAGCAAGTGGGAGTTTG-TAMRA-3; sFLT-1 forward: 5-
GGCTGACTCTAGAATTTCTGGAATCT-3 and reverse: 5-
by on May 16, 2011 circres.ahajournals.orgDownloaded from

Online Data Supplement Krysiak et al.
4
GTGGTACAATCATTCCTTGTGCTT-3, probe: 5-6-FAM-
AACTTGGAAAAAATGCCGACGGAAGG-TAMRA-3; KDR forward: 5-GAA AGC ATC GAA
GTC TCA TGC-3´ and reverse: 5-GTGGTACAATCATTCCTTGTGCTT-3; β-actin forward: 5-
ATCGTCCACCGCAAATGCTT-3; reverse: 5-CAACCGACTGCTGTCACCTTCA-3; probe: 5-
6-FAM-CACCCTTTCTTGACAAAACCTAACTTGCGC-TAMRA-3) were designed with the
use of the program Primer Express 1.0 (PE Applied Biosystems). RT-PCR was carried out as
previously described 3. TaqMan analysis was carried out according to the manufacturer’s
instructions, with the use of an Applied Biosystems 7700 system (PerkinElmer). The
amplification was performed with the following time course: 94°C for 3 minutes and 40 cycles
of 94°C for 30 s, 60°C for 1 minute and 72°C for 1 minute. Each sample was tested in
triplicate. Quantitation of the amount of target in unknown samples is accomplished by using
the Standard Curve Method. Plasmid DNA (pCR II vector; Invitrogen, Netherlands)
containing either the sFLT-1, FLT-1- or human β-actin-RT-PCR product was used to prepare
absolute standards. Expression levels of sFLT-1, FLT-1 were normalized to β-actin
expression. For realtime PCR analysis of β-actin cDNA was diluted 1:10 due to the higher
amount of β-actin expression.
Specificity of the FLT-1, sFLT-1 and KDR RT-PCR products was confirmed by sequencing
using an automated sequencing device (ABI PRISM 377, Applied Biosystems, Germany).
Enzyme-Linked Immunosorbent Assay
Sandwich ELISA for the detection of total (free and complexed) sVEGFR-1 was performed
according to manufacturer’s (RELIATech) specifications. Aliquots of serum from
normotensive and preeclamptic women were assayed in duplicate.
Cell Culture and Assays
Comparison of VEGF-mediated FLT-1 induction on PMNs
According to previous experiments with human vascular endothelial cells, where VEGF up-
regulates FLT-1 expression 4, we tested whether or not VEGF can affect FLT-1 expression in
by on May 16, 2011 circres.ahajournals.orgDownloaded from

Online Data Supplement Krysiak et al.
5
PMNs. Isolated maternal PMNs were cultured in RPMI 1640 (Seromed, Germany) containing
5 % FCS, 1% L-Glutamin (2mM) and 1% Penicillin/Streptomycin at 37 °C. Expression of
FLT-1 on PMNs was assessed by incubating cultures with 0.6 nM recombinant VEGF165
(Biochrome AG, Germany) or by pretreating with sFLT-1 (1,000 pg/mL) for 2 hours before
incubating with VEGF or aqua bidest (negative control). After 24 hours, FLT-1 expression
was measured by quantitative flow cytometry.
Measurement of neutrophil chemotaxis in Boyden chamber
To compare FLT-1-dependent migration of PMNs from preeclamptic and healthy women,
chemotaxis was assessed using multidish-6-well-cell culture plates with a 3-µm pore
diameter polycarbonate membrane transwell apparatus (cell culture insert, NUNC, Germany)
as directed by the manufacturer. 1.5 x 106 cells/well were placed over the filter in the upper
wells of the chamber and chemotactic were agents added to the lower chamber.
Recombinant VEGF165 at different concentrations (0.3 / 0.6 nM; Biochrome AG, Germany)
or aqua bidest (negative control) was applied to the lower wells of the chamber. To study the
effects of sFLT-1 and maternal serum on PMNs´ chemotaxis, neutrophils from normotensive
women were pretreated with sFLT-1 (1,000 pg/mL) for 2 hours before incubating with VEGF
(0.6 nM) or pretreated with serum from normotensive and preeclamptic women for 2 hours.
After incubation for 24 h the cells that had migrated through the filter were collected by
centrifugation and counted. The data were expressed as the average number of cells that
had migrated into three wells. Viability at the end of the assay in both chambers remained
>95 ± 2%. The cells that had migrated nonspecifically were excluded.
PMN responses to other cytokines was investigated by using N-formylmethionyl-leucyl-
phenylalanine (10 ng/ml, fMLP; Sigma, Germany) instead of VEGF165 as a positive control
for a neutrophil chemoattractant.
Transcriptional Regulation of FLT-1
by on May 16, 2011 circres.ahajournals.orgDownloaded from

Online Data Supplement Krysiak et al.
6
To examine the putative functional significance of enhanced FLT-1 expression after VEGF
exposure on the transcriptional level, we studied its promoter activity in HL-60 cells.
Based on the sequence of the human FLT-1 promoter [gi 1088437,|D64016.1], we performed
a genomic PCR (5’-GCGCACGCGTGTGGCAACTTTGGGTTACCCAAC-3’ and 5’-
GCGCAAGCTTTGTGAGAAGCAGACAGCTGAGCA-3’ containing restriction sites for HindIII
respectively MluI at their 5’-end) using genomic templates. The resulting PCR products (1196
bp of the 5'-regulatory region directly upstream of the putative translation initiation start
codon) were subcloned into the luciferase reporter vector pGL3basic (Promega, Madison,
USA) and the identity of the inserts was confirmed by sequencing 5.
HL-60 cells (DSMZ- Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH,
Germany) were cultured at 0.5 x 105 cells/ ml RPMI 1640 (1% L-Glutamin, 1%
Penicillin/Streptomycin) containing 10 % FBS for 72 hours. Next, HL-60 cells (2 x 107 cells)
were suspended in serum-free RPMI 1640 medium, mixed with pGL3-FLT-1 reporter plasmid
and electroporated in the Gene Pulser Xcell System (Bio-Rad, Germany). For standardizing
the transfection efficiency, pSV-Beta-Galactosidase vector (Promega, Germany) was
cotransfected. The cells were immediately transferred to a 12-well plate and cultured either
with 0.6 nM recombinant VEGF165 or pretreated with sFLT-1 (1000 pg/mL) for 2 hours
before incubating with VEGF or with aqua bidest (used as negative control) for 24h,
respectively.
We analyzed luciferase and beta-galactosidase activity as previously described 5. The
transfection data represent the mean relative luciferase activity (RLA) values of 3
independent experiments (each n=3). With respect to statistical analysis of transient
transfection experiments, a two-tailed t-test was performed and statistical significance was
assumed at P<0.05.
Screening of the promoter of the FLT-1 gene for genetic variations
by on May 16, 2011 circres.ahajournals.orgDownloaded from

Online Data Supplement Krysiak et al.
7
Information about genetic variations were obtained either by database search in the NCBI's
dbSNP 6, HGMD (Human Gene Mutation Database) 7, and HGBASE (Human Genic Bi-Allelic
Sequences) 8 or by Single-Strand Conformation Polymorphism (SSCP) -analysis.
SSCP-analysis
Genomic DNA from all study subjects was extracted by standard techniques 5. From the
published sequences of the FLT-1 promoter gene (D64016) overlapping fragments, ~320 bp
in length were enzymatically amplified to cover the entire promoter region in all 166 healthy
(332 alleles) women and 33 patients (66 alleles) with preeclampsia.
All PCR amplifications were performed by standard techniques 5 using of specific primer
(table S1), followed by single-strand conformation polymorphism (SSCP) analysis as
previously described. DNA from patients presenting different migration patterns on the
polyacrylamide gels were then sequenced twice (both DNA strands with sense and antisense
primers) with the use of an automated sequencing device (ABI PRISM 377, Perkin-Elmer).
Statistical Analysis
Data analysis was performed using the Statistical Package for Social Sciences (SPSS,
version 11.0 for Windows, United States of America) and Sigma Plot for Windows Version
8.0 (Systat Software GmbH, Germany). Values are given as means + SEM if not otherwise
indicated. Statistical analyses between the hypertensive and control group were performed
using analysis of variance, the Mann-Whitney test or Student´s t test. In addition, statistical
analysis of the migration assay was perfomed using the Kruskal-Wallis-test. P < 0.05 was
regarded as the level of significance.
by on May 16, 2011 circres.ahajournals.orgDownloaded from

Online Data Supplement Krysiak et al.
8
References
(1) Schonfelder G, John M, Hopp H, Fuhr N, van-Der-Giet M, Paul M. Expression of inducible
nitric oxide synthase in placenta of women with gestational diabetes. Faseb J. 1996;10:777-
84.
(2) Raff T, van der Giet M, Endemann D, Wiederholt T, Paul M. Design and testing of beta-actin
primers for RT-PCR that do not co-amplify processed pseudogenes. Biotechniques.
1997;23:456-60.
(3) Schonfelder G, Fuhr N, Hadzidiakos D, John M, Hopp H, Paul M. Preeclampsia is associated
with loss of neuronal nitric oxide synthase expression in vascular smooth muscle cells of the
human umbilical cord. Histopathology. 2004;44:116-28.
(4) Barleon B, Siemeister G, Martiny-Baron G, Weindel K, Herzog C, Marme D. Vascular
endothelial growth factor up-regulates its receptor fms-like tyrosine kinase 1 (FLT-1) and a
soluble variant of FLT-1 in human vascular endothelial cells. Cancer Res. 1997;57:5421-5.
(5) Funke-Kaiser H, Reichenberger F, Kopke K, Herrmann SM, Pfeifer J, Orzechowski HD, Zidek
W, Paul M, Brand E. Differential binding of transcription factor E2F-2 to the endothelin-
converting enzyme-1b promoter affects blood pressure regulation. Hum Mol Genet.
2003;12:423-33.
(6) Sherry ST, Ward MH, Kholodov M, Baker J, Phan L, Smigielski EM, Sirotkin K. dbSNP: the
NCBI database of genetic variation. Nucleic Acids Res. 2001;29:308-11.
(7) Human Gene Mutation Database. 2004.
(8) Human Genic Bi-Allelic Sequences. 2004.
by on May 16, 2011 circres.ahajournals.orgDownloaded from
Online Data Supplement Krysiak et al.
9
Additional Figures and supporting information:
Figure S1
Figure S1: FACS analysis of FLT-1 expression in maternal neutrophils using peptide
pre-absorbed antibodies against FLT-1.
Specificity of the obtained immunoreactions within the flow cytometry was assessed by using
different amounts of peptide pre-absorbed antiserum against FLT-1 (2:1 and 1:1 competition
for 30 min at room temperature; A, C and E). In contrast, pre-absorbtion of the CD11b
antibody with the FLT-1 peptide (2:1 and 1:1 competition for 30 min at room temperature; B,
D and F) did not change its immunoreaction. We conclude, that immunreactions against FLT-
1 are highly specific. Data are representative of three independent experiments.
by on May 16, 2011 circres.ahajournals.orgDownloaded from
Online Data Supplement Krysiak et al.
11
Additional Tables: Table S1: FLT-1 Primer sequences for SSCP-analysis
No.
Primer pair
Primer sequence
5´- 3´
Position
Gene bank: D64016
1 gtggcaactttgggttaccca (sense)
tgaccacaaccggctgaaat (antisense)
1 - 21
295 - 275
2 ccggttgtggtcagagtcact(sense)
cgcccaagtcatttcctca (antisense)
283 - 304
485 - 466
3 actgaggaaatgacttgggcg (sense)
cctgaccccttcagactgtcc (antisense)
465 - 486
665 - 644
4 ttcagggtctttgctcct (sense)
cttactcagctctgggggc (antisense)
648 - 666
857 - 838
5 gcaaggggagacagccggactgc (sense)
ctcctgcccatcgcgctcagga (antisense)
693 - 716
884 - 862
6 tttcgggcggcccccagagct (sense)
cacttcctaccccggcacctccttct (antisense)
830 - 851
1125 - 1098
7 agcctgggaggaagaag (sense)
tccagccaggagacaac (antisense)
1059 - 1076
1357 - 1340
by on May 16, 2011
circres.ahajournals.orgD
ownloaded from
























![Mianserin in breast milk [letter]](https://img.pdfslide.net/doc/110x75/634774a86e55ee279302c2dd/mianserin-in-breast-milk-letter.jpg)




