Embed Size (px)
Citation preview

THE JOURNAL OF COMPARATIVE NEUROLOGY 215:228-243 (1983)
Somatosensory Properties of the Inferior Olive of the Cat
REUBEN GELLMAN, JAMES C. HOUK, AND ALAN R. GIBSON Neuroscience Program and Department of Physiology, Northwestern University
Medical School, Chicago, Illinois 60611
ABSTRACT We examined the somatosensory properties of 391 neurons in the in-
ferior olive in 20 cats that were anesthetized with barbiturate or decere- brated. A response consisted of a single spike with a variable number of wavelets followed by a long refractory period. Neurons responsive to natural somatosensory stimuIi were recorded in all olivary subdivisions. The dorsal accessory olive (DAO) contained the highest proportion of responsive units (96%), compared with 66% for the medial accessory olive (MAO) and 43% for the principal olivary (PO) nucleus. Within the rostral DAO we found a refined cutaneous map of the entire contralateral body surface. In the caudal DAO responsiveness to manipulation of deep tissues became prominent, and both individual limbs and bilateral pairs were represented. In the me- dial region of the PO responsiveness to taps predominated and bilaterally symmetrical fields were frequent. The lateral PO was unresponsive under the conditions of these experiments. The MA0 was distinguished by a greater complexity of receptive field and by a preponderance of deep over cutaneous modality. The lateral part of caudal MA0 contained cells with interesting spatial patterns of excitation and inhibition, whereas most cells in the rostral MA0 had purely excitatory fields. A teleceptive area receiving visual and auditory input was recognized in the medial MA0 and nearby structures such as the dorsal cap. Contact and proprioceptive signals arriving via climbing fibers may provide the cerebellum with information necessary to relate the body to external objects.
Key words: climbing fibers, cerebellum, neurophysiology, somatotopy
Sensory input to the cerebellum arrives via two systems, the climbing fibers and the mossy fibers. The mossy fibers provide a wide range of inputs that arise from many areas of the brain; the climbing fibers, in contrast, arise solely from the inferior olive (Courville and Faraco-Cantin, '78). Climbing fibers exert a strong synaptic influence on Pur- kinje cells in the cerebellar cortex (Eccles et al., '66) and also provide an input to the cerebellar nuclei (see Brodal and Kawamura, '80, for review). Normal cerebellar activ- ity seems to depend on the integrity of the olive (Wilson and Magoun, '45; Carrea et al., '47; Brooks et al., '80). Despite its obvious importance in cerebellar function, few direct studies of the physiological properties of the inferior olive have been reported.
Most physiological studies of the olivocerebellar system have been based on identifying and recording the "complex spike" produced in Purkinje cells when they are synapti- cally activated by climbing fibers. Sensory stimuli which produce complex spikes include pinching skin and deep tissues (Thach, '67; Oscarsson, '68, '69), taps against the footpads (Larson et al., '69a), and light puffs of air on hairy skin (Eccles et al., '72). Climbing fibers and olivary cells
responding to the stimulation of muscle afferents have been reported (Armstrong et al., '68; Ishikawa et al., '72a,b), and passive movement of a limb also activates climbing fibers (Rushmer et al., '76; Kolb and Rubia, '80). Both large (Thach, '67) and small (Eccles et al., '72) receptive fields have been reported. Rushmer et al. ('80) noted cells with receptive fields on more than one limb, and such fields were occasionally discontinuous.
The studies of complex spikes provide only indirect in- formation about the organization of sensory input to the inferior olive. In addition, many earlier studies utilized electrical shocks to peripheral nerves and thus did not provide detailed information about receptive field struc- ture and modality. Furthermore, Armstrong and Rawson ('79) have reported that complex spikes are sometimes in- distinguishable from simple spikes on extracellular re- cording. Although much has been learned about the climb- ing fiber system, a clear picture of its sensory organization has yet to emerge. We have studied the responses of single
Accepted December 20, 1982.
0 1983 ALAN R. LISS, INC.
INFERIOR OLIVE ORGANIZATION 229
olivary neurons to natural stimuli with the intent of de- termining the organization of somatosensory information within the inferior olive.
MATERIALS AND METHODS Preparation
The experiments were performed on 20 adult cats (weights 1.8-4 kg). All but two of these were anesthetized with sodium pentobarbital (35 mgikg, IV) which was supple- mented as necessary throughout the experiment. The anesthetized preparations were maintained at a level where pinching the paw produced withdrawal. The animals were not paralyzed and showed no sign of discomfort. The re- maining two animals were decerebrated under nitrous ox- ide-halothane anesthesia, and recordings were carried out without anesthesia. Heart rate and rectal temperature were monitored, and temperature was maintained within the physiological range with a heating pad. The typical du- ration of an experiment was 36 hours.
The cats were mounted in a Kopf stereotaxic apparatus. The olive was approached through the cerebellum with the electrode mounted so that its tip pointed forward a t an angle of 15" to the vertical. This angle permitted rostra1 parts of the inferior olive to be studied without having to remove portions of the skull near the transverse sinuses and the bony tentorium. The dorsal approach through the cerebellum was chosen in preference to the ventral ap- proach since preparation time is short, trauma to the an- imal is reduced, and the position of the electrode can be referenced in standard stereotaxic planes.
Recording procedures We used Epoxylite-insulated tungsten microelectrodes
(impedance 1-3 M a ) for extracellular recording. In most experiments tracks were spaced 0.5-1 mm apart, and these were continued through all olivary laminae encountered. In a number of later experiments we were concerned with detailed organization within the dorsal accessory olive (DAO), and in these cases penetrations were about 0.2 mm apart and were restricted to the DAO.
As we became more adept at recording from this region, a number of features were found to be helpful in locating the olive during the electrode penetration. Stereotaxic co- ordinates were used as a first approximation; the anter- oposterior coordinates did not vary by more than 2 mm. Once the dorsal surface of the brainstem had been located, it served as an initial landmark for depth, as the olive is typically 3-4 mm below this. Upon approaching the ex- pected location of the inferior olive the microelectrode was advanced slowly. Entry into the olive was signalled by a characteristic background activity, a low-frequency sput- tering sound, much like popcorn popping. The cessation or beginning of this background activity enabled us to note when the electrode was passing out of one olivary lamella or into another. We were able to confirm that these silent regions corresponded to spaces between lamellae with his- tological reconstructions aided by small lesions (see be- low).
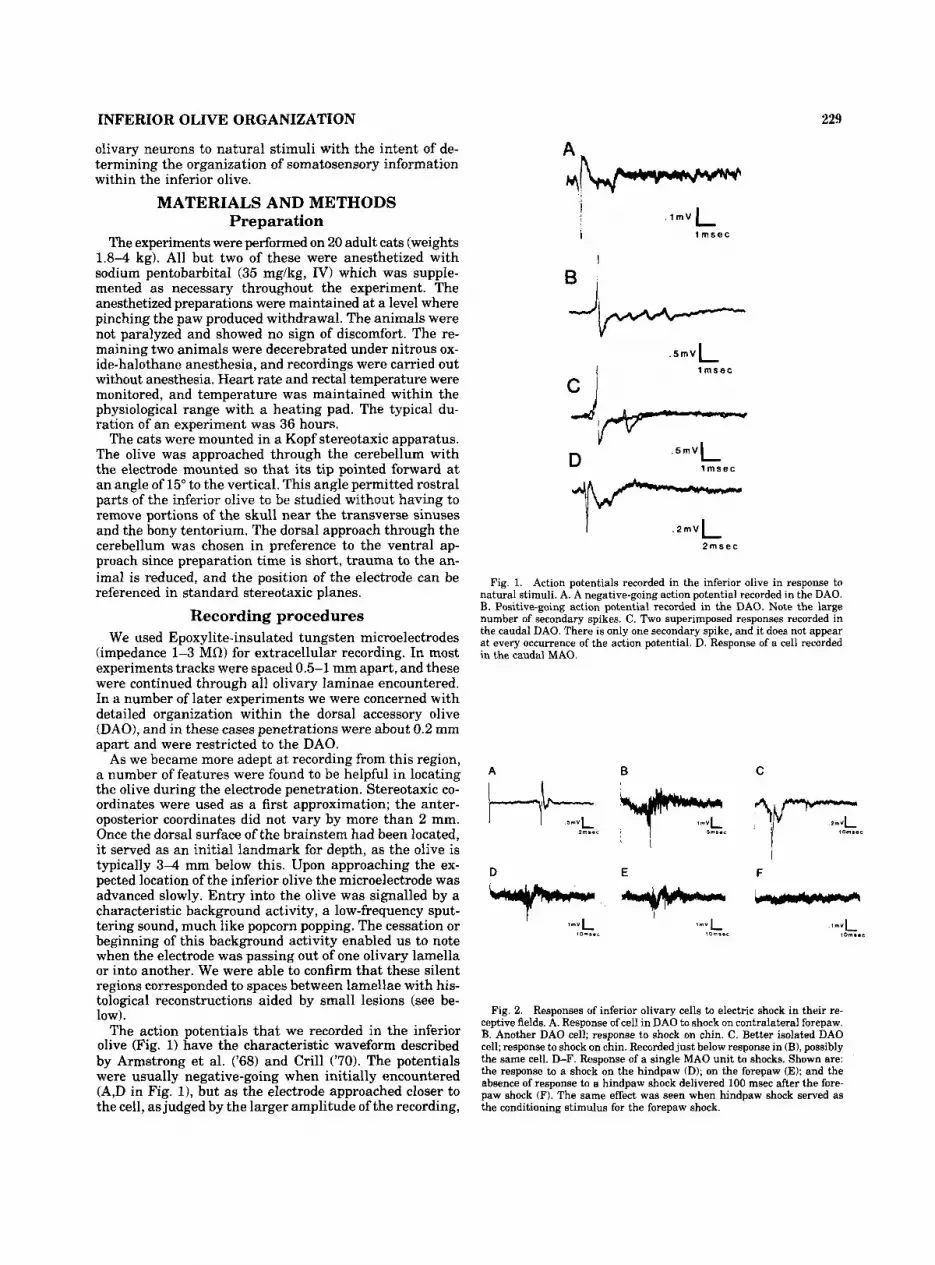
The action potentials that we recorded in the inferior olive (Fig. 1) have the characteristic waveform described by Armstrong et al. ('68) and Crill ('70). The potentials were usually negative-going when initially encountered (A,D in Fig. I), but as the electrode approached closer to the cell, as judged by the larger amplitude of the recording,
. I ~ V L j i 1 msec
I
B I
Y D
. 5 m ~ (
l m s e c
. 2 m ~ L
2msec I
Fig. 1. Action potentials recorded in the inferior olive in response to natural stimuli. A. A negative-going action potential recorded in the DAO. B. Positive-going action potential recorded in the DAO. Note the large number of secondary spikes. C. Two superimposed responses recorded in the caudal DAO. There is only one secondary spike, and it does not appear a t every occurrence of the action potential. D. Response of a cell recorded in the caudal MA0
A B
E - C
I F -
Fig. 2. Responses of inferior olivary cells to electric shock in their re- ceptive fields. A. Response of cell in DAO to shock on contralateral forepaw. B. Another DAO cell; response to shock on chin. C. Better isolated DAO cell; response to shock on chin. Recorded just below response in (B), possibly the same cell. D-F. Response of a single MA0 unit to shocks. Shown are: the response to a shock on the hindpaw (D); on the forepaw (E); and the absence of response to a hindpaw shock delivered 100 msec after the fore- paw shock (F). The same effect was seen when hindpaw shock served as the conditioning stimulus for the forepaw shock.

1; ii !i ~
0.8 I 1 1 1 I 1 7 J7
6 8 10.8 12 .6 14 .7
l l l l 1 1 1 14.7P 0.6L d C
1.7L "10
2.1L
2.7L
DAO
MAO
Fig. 3. A series of frontal and parasagittal sections presented for com- the levels of the frontal sections. Abbreviations: cMAO, caudal part of- parison. The figures alongside the frontal sections indicate the frontal plane; MAO; DAO, dorsal accessory olive; dc, dorsal cap; dmcc, dorsomedial cell those along the parasagittal sections indicate the distance from the midline. column; np, nucleus p; PO, principal olivary nucleus; rMAO, rostra1 part Each line drawn through the frontal sections corresponds to the level of of medial accessory olive; vlo, ventrolateral outgrowth. one parasagittal section. The scale above the parasagittal sections indicates
Fig. 4. Examples of histological sections used for determining location of a recorded response. Marking lesions are located to the right of the numbers (1-9). In A: (1) Lesion identifying recording track; (2) DAO cell responding to touch on dorsal surface of forelimb; (3) cell in ventral lamella
of PO responding upon release of squeeze of forelimb. In B: (4), (5), (7) Lesions identifying recording tracks; (6) unresponsive cells; (8) cell inhib- ited by squeeze of both hindlimbs; (9) cell with field shown in Figure 14 for cat 1014.

INFERIOR OLIVE ORGANIZATION 231
A
SOMATOSENSORY CELLS
DAO 0.7L "L"a - l 1.4L
DAO
0-
rMAO
2.0L
D A O D A O
P
2.7L
B
TELECEPTIVE CELLS
D A O 0.7L - -a 1.4L
DAO DAO - 2.OL
D A O
2.7L
C
UNRESPONSIVE CELLS
DAO 0.7L
r M A O c M A O
1.4L D A O D A O
2.0L
DAO D A O
2.7L
Fig. 5 . Overall summary showing histologically verified tracks and re- cording locations projected onto a series of four composite parasagittal sections. A. Somatosensory cells: 0, contiguous receptive fields; A, non-
contiguous receptive fields or cells with mixed excitatory and inhibitory input. B. Teleceptive cells: 0, auditory; A, visual; t, combined auditory and visual input. C. Unresponsive cells with spontaneous activity.
the action potential often reversed to positive-going (B,C in Fig. 1). In Figure 1B four wavelets ("secondary spikes") can be seen following the initial spike. The number of secondary spikes produced by a given cell was variable as shown by the two superimposed traces in Figure 1C. In one trace a secondary spike can be seen, while in the second only the primary spike occurred. Only cells with action potentials similar to these, and that were clearly located within the olive based on histological verification, are de- scribed here. We refer to each action potential together with its secondary spikes or wavelets as one spike or one action potential for the purpose of describing responses.
Sensory stimuli such as taps and squeezes were contin- ually applied during penetrations. In some experiments a flashing light and/or a device tapping the earbars (as an auditory stimulus) were used to search for teleceptive in- put. Once an olivary cell was located, its receptive field was determined (manually or by using a fine probe), and we attempted to define an optimal stimulus. Cells respond- ing to superficial stimuli such as puffs of air or light touch were defined as cutaneous; those responding to manipu- lation of muscles or joints we called deep. Light touch, which seemed to be equivalent to bending hairs, was sub-
jectively defined, but in later experiments with an aes- thesiometer (Stoelting 18011) it was determined to be ap- proximately equivalent to a force of 25 mg. Cells responding to sharp taps but not touch were not placed in either cat- egory. Their responses may have been due to stimulation of Pacinian corpuscles or synchronous activation of pri- mary endings on muscle spindles. For cells with somato- sensory properties ("somatosensory cells") a standarized data sheet was completed, which noted the stereotaxic lo- cation of the cell, described the types of stimuli to which it responded, noted its latency to electric shock, and in- dicated its receptive field on a schematic cat.
Response latencies were determined for somatosensory stimuli by shocking percutaneously in the receptive field through two hypodermic needles. Single shocks (-1 mamp, 100-pec pulses) were provided by a Grass S88 stimulator driving a PS-IU6 constant-current unit. The response typ- ically consisted of a field potential, with one or more su- perimposed spikes. Figure 2A shows a large spike elicited by a shock; large fields with spikes in response to a shock are shown in Figure 2B,C. The fields and responses are similar to those obtained in the caudal part of the DAO by Armstrong et al. ('68).

232 R. GELLMAN, J.C. HOUK, AND A.R. GIBSON
Histological verification and construction of maps
During the experiment small lesions were placed at points of interest or elsewhere along the track by passing current ( - 5pamp for 5 seconds, typically) through the electrode to aid in histological reconstruction. Lesions were placed immediately after recording a response before advancing the electrode or during withdrawal of the electrode after completion of recording in the track. In the latter case the lesion was placed only if the cells were physiologically identical to those recorded while advancing the electrode, since about 1 mm of hysteresis exists between identical points on the withdrawal of the electrode compared with its advance. On some tracks additional lesions were placed above the olive to aid in identification. Following all ex- periments the animals were perfused through the aorta with saline followed by formalin, and the tissue was pro- cessed conventionally. Sections were stained with luxol fast blue and counterstained with cresyl violet. Only units that could be localized histologically with a great degree of confidence were included in our data base.
We used the parasagittal plane of section in all cases but one. One series of parasagittal sections was used as a “model” inferior olive, and the locations of all tracks, le- sions, and recording sites were plotted onto this map. This plane was chosen as it was the only standard stereotaxic plane in which our tracks (placed at 15” to the frontal plane) could readily be seen. Furthermore, since the olive is longitudinally oriented, we found that we could most readily comprehend its organization in the parasagittal plane. Since the olive is traditionally shown in the frontal plane while some of our data are presented in the para- sagittal, a series of sections in the latter plane is shown in Figure 3. This figure also shows an abbreviated frontal series for orientation. The horizontal projections of the DAO and MA0 used in several figures were constructed from the complete set of parasagittal series from which the samples in Figure 3 were taken. Anterior and posterior edges of the subnuclei were plotted as a function of dis- tance from the midline. The points were then connected to form the horizontal projections yielding an outline sim- ilar to that introduced by Brodal (’40). AP (anterior-pos- terior) coordinates are taken from Berman (’68). Some data is presented on an abbreviated series of parasagittal con- structions (Fig. 5). These are composites derived from av- eraging a number of adjacent sections and reflect the ap- proximate structure of the olive at the indicated laterality.
Figure 4 shows two parasagittal sections through the inferior olive typical of those we used to identify recording sites and to construct the maps on which we report. The upper section (Fig. 4A) shows three lesions on one track: The uppermost was used to distinguish this track from another nearby. The middle lesion shows the location of a cell in the DAO, while the ventralmost lesion is in the ventral lamella of the PO. Note that the latter two lesions are approximately 300 pm apart. In the lower section, in addition to three lesions in the caudal MAO, three track identifying lesions can be seen.
RESULTS We recorded cells responsive to somatosensory stimuli
in all the major subnuclei of the inferior olive-namely, the principal olive (PO), the medial accessory olive (MAO),
and the dorsal accessory olive (DAO). The main purpose of this study was to map the nature and distribution of this somatosensory input, but we also examined respon- siveness to visual and auditory stimulation and noted the location of cells that were spontaneously active but un- responsive to the stimuli applied by us. The subnuclei were quite different from each other in respect to the distri- bution of these three categories of cellular activity as sum- marized by the parasagittal composites shown in Figure 5. Our total sample of 391 neurons recorded during 136 electrode tracks through the inferior olive is projected onto these sections, with each point representing one or more neurons recorded a t that location. The majority of active neurons were responsive to some form of somatosensory stimulation (287, or 73%; referred to as somatosensory cells in this paper, and shown in Fig. 5A), 4% were re- sponsive to flashes or clicks (teleceptive cells; Fig. 5B),and 23% failed to respond to any of the stimuli which we ap- plied (Fig. 5 0 . The proportion of responsive neurons var- ied considerably with location within the olive. Nearly all (96%) cells were responsive in the DAO, contrasting with 66% in the MAO, and only 43% in the PO. Not shown in Figure 5 are 14 tracks which we showed histologically to have passed through certain subdivisions of the olive but on which no units were recorded. Two of these passed through the DAO, six through the MAO, and six through the PO.
Dorsal accessory nucleus Different combinations of cutaneous and deep stimuli
provided adequate stimulation for 150 of 156 cells recorded in this region, and those responsive to natural stimuli could also be activated by electrical stimuli applied through a pair of hypodermic needles inserted within the receptive field. With both natural and electrical stimulation we con- firmed previous reports (Armstrong and Harvey, ’66; Arm- strong et al., ’68) that olivary neurons have long refractory periods and will not respond to stimuli repeated a t rates above 7-8lsecond. At lower rates (up to 2-3Jsecond) the response to both types of stimulation is very reliable. We found that neurons responding to light cutaneous stimu- lation could be activated in a one to one manner with each presentation of the stimulus, while those responding to stimulation of deep structures sometimes failed to respond to repeated stimuli, even at these low frequencies. This may have resulted from a failure to locate and stimulate the most sensitive portion of the receptive field.
We used electrical stimuli to determine the response latency of cells to stimulation in their receptive fields. The latency for a given neuron was relatively constant. For cells with short latencies (<15 msec) the variability was no more than 1-2 msec from the mean, and even for cells with rather long latencies (40-50 msec) the variability did not exceed 5 msec. A histogram of the mean latencies for DAO cells with receptive fields on the face, forelimb, and hindlimb is shown in the upper part of Figure 6. The modal values were 12 msec for the face and forelimb and 25 msec for the hindlimb. The mean values of the latencies for these regions are given in Figure 6.
We found that 29% of DAO cells responded exclusively to deep input (squeezing or moving the limb) while 35% responded well to light cutaneous stimuli such as puffs of air or stroking the skin. For the remaining cells, which responded to taps in the receptive field, it was not clear

INFERIOR OLIVE ORGANIZATION
FACE
233
FORELIMB HINDLIMB 6 0
55
25
20
15
1 0
5
n 4 7 UNITS
MEAN 23 1 m s e c
UNITS
MEAN 12.9 msec
7 3 UNITS
MEAN 15 8 m s e c
1
I 0
0 10 20 30 40 50 60
25 1 5 UNITS
0 10 20 30 40 5 0 60
24 UNITS
MEAN 28 7 m s e c (74.5 msec)
10 UNITS
MEAN 38.7 m s e c
25
20
15
n 10
2 5
2 0
15
1 0
MEAN 21.6 mseC
: L 4 0 10 2 0 30 4 0 50 6 0 100 300 0 1 0 2 0 3 0 4 0 50 6 0
m i e c
0 1 0 20 30 4 0 5 0 6 0
msec msec
Fig. 6. Histograms showing latencies of inferior olivary cells to electric shocks delivered in receptive field. Top row: cells in D A O bottom row: cells in MAO.
3.0- LATERAL
2.5-
2.0-
1.5-
1 0 -
0 0 0 CUTANEOUS(& DEEP?) 0 0 DEEP ONLY
O I O 0.5 -
I 0.0
I 10.0
I 11.0
I 12.0
I I 13.0 POSTERIOR 14.0
Fig. 7. Dorsal view of the DAO showing location of cells with exclusively deep input versus those with a difinite cutaneous input. Each symbol indicates the property of one cell recorded at that point. Note the preponderance of cells with deep input caudally, and those with light cutaneous input rostrally.

234 R. GELLMAN, J.C. HOUK, AND A.R. GIBSON
whether the response was to higher-threshold cutaneous receptors or to deep input. In Figure 7 the distribution of cells that had clear cutaneous input and those with exclu- sively deep input are shown on a dorsal projection of the DAO. It is apparent that in the rostral part of the DAO cells which have cutaneous input predominate, while in the caudal part most cells seem to have exclusively deep input. At the caudal tip there is a small zone of cells with cutaneous input.
Cells responsive to very light cutaneous stimuli also responded to hard squeezes and manipulation of the joint. In order to discover whether this was due to a deep input or another form of stimulus to the cutaneous receptors, we skinned the forelimb of one animal while recording from a cell that responded strongly to light cutaneous stimuli. After the skin had been peeled back, preserving much of its innervation and blood supply, responses could still be obtained from the olivary unit when the skin flap was stroked. Tapping or pinching the muscles also produced a reliable response in the DAO unit as did rotation of the joint. Cutting the tendon and pulling on these muscles, in contrast, produced no response unless very substantial tugs were applied. The receptors in question may have been high-threshold muscle receptors, although the high threshold could also have resulted from a depression of low-threshold muscle spindle activity transmission through a central pathway. We noted that the extent of the area that was driven by deep stimuli corresponded well with that of the superficial receptive field.
All but four of the responsive DAO cells showed exclu- sively excitatory responses to stimulation. The responses generally consisted of a modulation of the firing rate above the low spontaneous level of activity which was in the range 0-llsecond. The form of this modulation varied, with cells responsive to squeezing of muscles showing a tonic response at rates of 1-3/second as long as the stimulus was maintained. Cells responsive to light cutaneous input responded tonically to a prolonged stroke, but a main- tained touch or light pressure at one site in the receptive field produced only one (or occasionally two) spikes when contact occurred. This light pressure often evoked a tonic background low-amplitude activity, seemingly implying that the response of the afferents to the DAO is not mim- icked by the DAO cells themselves. Sharp taps evoked large responses consisting of spikes from more than one unit and a large field potential. Responses of DAO neurons in the two decerebrated cats were similar to those in the anesthetized cats. Spontaneous activity, however, seemed slightly depressed in the decerebrated cats.
Receptive field organization. Receptive fields of DAO cells generally had well-defined boundaries. Examples of fields from the rostral part of the DAO, all of which were purely contralateral, are shown in Figure 8A (hindlimb) and B (forelimb). Note that the field may vary substan- tially in size, occupying only part of one paw (bottom panel of Fig. 8B, top panel in Fig. 8A), an entire limb (Fig. 8A, bottom right; Fig. 8B, upper panel) or even larger areas (Fig. 11). The smallest receptive field we encountered cov- ered only one claw; at the other extreme we found receptive fields covering the entire trunk on the contralateral side. Fields generally contained only part of a limb and were usually restricted to either the dorsal or the ventral sur- face. The panels of Figure 8 are typical in this respect.
Figure 8 further shows that receptive fields located prox- imally on the limb were larger than those located distally. Such large fields usually included the paw, although fields covering the forelimb above the wrist were occasionally seen.
Figure 9 shows the receptive fields of a number of cells recorded in the caudal part of the DAO. Although small, restricted receptive fields are occasionally seen in the cau- dal DAO (Fig. 9A), the fields are generally larger than those in the rostral DAO, and 20% of cells in this caudal region have noncontiguous receptive areas (Fig. 9B,D) compared with 10% in the rostral part. Of 65 cells in this region, 15% had bilateral input with both continuous (Fig. 9C) and noncontiguous (Fig. 9D) fields. No cells in the rostral DAO had bilateral input.
The study of recording sites within the DAO revealed a clear somatotopic organization. Figure 10 shows a dorsal projection of the locations of tracks on which somatosen- sory cells were recorded in the DAO. Each symbol indicates the receptive area(s) of the cell(s) recorded on the track in question. A small area representing the head and face is most medial. A forelimb area overlaps with the face area and extends about 1 mm more laterally. The most lateral part of the DAO is devoted to the hindlimb. This soma- totopic organization is present in the caudal pole of the DAO as well, although overlap between the zones repre- senting different body areas is more common in this region. Axial areas are represented a t the rostral edge of the DAO, the trunk being represented medial to the tail. Thus the rostral half of the DAO contains a complete and detailed representation of the surface of the entire contralateral half of the body.
Closely spaced penetrations in individual animals re- vealed an organization even more detailed than that shown in Figure 10. Two examples of this detail are illustrated in Figure 8A and B, which summarize the data from two experiments in which penetrations were spaced about 200 pm apart. In the forelimb zone (Fig. 8B) successive pen- etrations from medial to lateral (from bottom to top in Fig. 8) revealed cells with receptive fields that extended to include successively more proximal regions of the limb. Thus the paw was represented medially in small receptive fields, while the entire limb, including the paw, was rep- resented more laterally in large fields. In the hindlimb region of one animal (Fig. 8A) a mirror image of the or- ganization in the forelimb region was found, with the prox- imal parts of the limb represented rnedially (bottom of figure) and the paw more laterally (top).
The detailed organization along the anteroposterior di- mension within the rostral part of the DAO for both the forelimb and the hindlimb is also shown in Figure 8. As one moves from anterior to posterior (left to right in the figures) in the DAO, the receptive field moves from the dorsal surface of the limb around the lateral aspect to the ventral surface.
As mentioned above, cells with noncontiguous receptive fields were often seen in the caudal part of the DAO. Such fields were also occasionally (10% of 91 cells) seen in ros- tral DAO, although all of these cases occurred at the bor- ders between zones representing the different body regions contained in their receptive fields. Figure 11 gives rep- resentative examples of cells located at border regions be- tween body areas.

INFERIOR OLIVE ORGANIZATION
A
B
235
I M
n
Fig. 8. Detailed organization of (A) hindlimb and (B) forelimb areas in rostra1 part of the DAO. The darkened area on each pair of figurines in- dicates the receptive field of one cell. All these cells responded to light cutaneous stimulation. In both regions there is a gradient from the dorsal
to the ventral surface of the limb with anterior to posterior progression in the DAO, while the proximal-distal location on the limb varies with me- diolateral position. The insert at the bottom left schematically locates the recording locations in DAO.

236 R. GELLMAN, J.C. HOUK, AND A.R. GIBSON
Fig. 9. Examples of receptive fields from the caudal pole of DAO. Note noncontiguous and bilateral fields (B-D). A and C reponded to light cutaneous stimulation, B and D to hard squeeze.
30- L A T E R A L
2 5-
2 0-
1 5 -
A A A A
1 0 -
0 5-
I I I I I I 8 0 10.0 11.0 12 0 13.0 POSTERIOR 14.0
Fig. 10. Somatotopic organization in DAO. Each point summarizes the receptive fields of all cells recorded on a penetration at that point. Pooled data from 20 cats. Insert (not to scale) summarizes somatotopic organization. Dashed border between face and forelimb zones indicates that there were many forelimb cells in the face area.

INFERIOR OLIVE ORGANIZATION 237
Medial accessory nucleus Of the 150 cells that we recorded in the MA0 (118 in
the caudal division, 32 in the rostral division) 66% were responsive to one or more of the stimuli which we tested. The bulk of these (88% of responsive units) responded to somatosensory stimulation, and the remainder responded to visual or auditory stimulation (Fig. 5). We include in the latter group visual neurons located in the dorsal cap and the adjacent ventrolateral outgrowth, even though these regions may not strictly be part of the MAO. Un- responsive cells were found in both the rostral and caudal divisions of the MAO. In the caudal M A 0 39 of 118 cells were unresponsive, and most of these were concentrated in the rostral half of this region, which included the te- leceptive area defined later (see Fig. 14). Responses of MA0 cells to somatosensory stimulation were frequently not as reliable as those in the DAO; electric shocks repeated at low rates (l/second or less) and repeated natural stimu- lation often failed to evoke a response on every trial. This was due in part to the longer refractory periods of MA0 as opposed to DAO cells.
Somatosensory neurons most commonly (44% of 87 cells) responded to natural stimulation with an increase in firing rate. A large group (26%), however, showed a reduction in firing rate. The remaining cells responded on release of an applied stimulus (7%) or responded in different ways to stimuli applied to different parts of their receptive fields (23%). For cells in the latter category both spontaneous activity and firing elicited by stimulation in an excitatory receptive area of a neuron were reduced by stimulation in “inhibitory” regions of the field. The reduction in firing rate seen in response to a maintained squeeze was very brief in some cells (duration about 1 second) and long- lasting in others (up to 20 seconds). Most of the cells which showed a reduced firing rate in response to a stimulus resumed their spontaneous rate upon termination of the stimulus (77% of 43 cells); the remainder showed a tran- sient excitation.
The somatosensory neurons in the MAO, whether ac- tivated or depressed by a stimulus, usually responded only to squeeze of muscles or other deep structures (57% of somatosensory units). A further 37% responded to taps or pressure, and only 6% responded to light cutaneous stimuli such as touch or puffs of air. A small number of cells responded to different stimuli in different parts of their receptive field; examples can be seen in Figure 12.
Response latencies of neurons in the MA0 were long and variable, as indicated in Figure 6 (lower part). This variability was evident in the population and also for a given cell from stimulus to stimulus even when stimuli were spaced several seconds apart to eliminate compli- cations due to refractoriness or habituation. We exclude inhibitory cells and those with unreliable responses from this illustration; as a result only 45% of the somatosensory population is represented. An analysis of the locations of these cells indicated no apparent distribution on the basis of latency: Cells of almost all latency ranges (20-30 msec; 30-40 msec; 40-60 msec; 100-300 msec) were evenly dis- tributed throughout the subnucleus. The only exceptions were the four cells with latencies between 10 and 20 msec, all of which were found in the caudal half of the MAO.
Receptive field organization. The receptive fields of the majority of neurons in the MA0 (67% of somatosensory cells) consisted of several noncontiguous receptive areas from the limbs, head and/or trunk as illustrated in Figure
Fig. 11. Location and receptive fields of five cells in DAO with input from more than one body area. These cells are found near the borders of the body representation zones.
J INHIBITORY EXCITATORY
Fig. 12. Examples of receptive fields in MAO. A-D from caudal division; E, F from rostral division. Note the complexity of receptive fields including the presence of inhibitory areas. Activation was by hard squeeze or sharp taps, except A and B, the forepaw in C, and the face in D, which were responsive to light taps or touch.

238 R. GELLMAN, J.C. HOUK, AND A.R. GIBSON
Y.,,
N-0 N-20 N.20 N - I 3
N-Ll N.12 N - I 1 100
m
W 0
: 8 0
; 0 0
4 0 2 ,
I: W
0 2 0 a a W
0
Fie. 13. Me teri
0 0 .5 1.0 1.6 2 .0 2.6 3.0 DISTANCE F R O M MIDLINE (mm)
OFORELIMB
.HINDLIMB
MIXED
somatotopy compare Y
in the three subnuclei. In all regions forelimb units are more common medially and hindlimb units are more common laterally. Note that the M A 0 does not extend beyond 2.5 mm, and the PO begins about 1 mm from the midline. The dashed line in the bottom histogram indicates that unresponsive units in the PO are found as far as 2.5 mm from the midline while no responsive neurons were found in this region. With the exception of this group, unresponsive neurons are not plotted on this figure.
2 0-
1 0 -
0 - I I I I I I I 8 0 1 0 0 1 1 0 1 2 0 1 3 0 1 4 0 1 5 0
Fig. 14. The upper row shows a particular combination of inhibitory and excitatory input that was found in three cats. The inhibitory response was obtained by hard squeezes, the excitatory responses by taps. In the lower part all cells with excitatory, inhibitory, and mixed responses are shown. The locations of the three cells shown in the upper part of the figure are indicated by the lines drawn from these figures. Note that these three cells are concentrated in a small region. Also note that there are few cells with inhibitory input in the rostra1 half of the nucleus, and that there appears to be a gradient of increasing inhibition moving caudally.
A
I lOmsec
II lOmsec
Fig. 15. Cells in M A 0 responding to a flash of light. The trace was triggered simultaneously with the flash. The two responses were recorded in the same cat. Note the difference in latency. A. Recorded in the cap lying dorsal to the MAO. B. Recorded in MAO, about 0.5 mm medial and 1.5 mm caudal to the site of A.

INFERIOR OLIVE ORGANIZATION 239
12C-F and Figure 14. Contiguous receptive fields were found for 33% of somatosensory cells (Fig. 12A,B). While these fields were occasionally small (Fig. 12A), in most cases responses could be obtained by stimulating almost any part of the limb. The lack of response of these neurons to light cutaneous stimulation and our inability to provide more localized deep stimulation prevented us from map- ping receptive fields as precisely as in the DAO.
We wished to verify that the noncontiguous receptive fields were indeed recorded from one cell, as appeared to be the case, and not from two separate neurons. Utilizing the 100-msec refractoriness of inferior olivary cells follow- ing activation, we examined the effect of a conditioning stimulus applied to one part of the receptive field on the neuronal response to stimulation in a different part of the field. The example in Figure 2D,E demonstrates the re- sponse of a cell with a noncontiguous receptive field to separate shocks to the forepaw (D) and hindpaw (El. Figure 2F demonstrates that the field potential is decreased in amplitude following a conditioning shock compared to Fig- ure 2D, and the unit seen in Figure 2D is not present in 2F. Clearly, the two body areas show a strong interaction, and the same unit is probably driven from both. While this example is from the MAO, the same phenomenon was observed in the DAO and PO.
The complexity of receptive fields in the MA0 precludes the description of a simple somatotopic organization in this region. Figure 13 shows that there is nonetheless a broad mediolateral somatotopy, wherein the proportion of cells receiving input from the hindlimb is greater laterally in the MAO, while those with input from the forelimb pre- dominate medially.
Cells with common receptive field properties tend to cluster in well-defined regions in the MAO, and the or- ganization was the same from animal to animal. The lo- cations of all MA0 cells which responded to somatosensory stimuli are plotted on Figure 14. Also shown are the lo- cations of three cells (from different cats) that combined excitation and inhibition in a particular manner (upper part of the figure). Note that these cells were found within a restricted zone extending about 0.4 mm rostrocaudally and 0.2 mm mediolaterally. Located near these cells are other units that combined excitation and inhibition (tri- angles in Fig. 14). Note that in the intermediate and ros- tral MA0 the responses were mostly excitatory (filled cir- cles), while purely inhibitory fields (open circles) tended to cluster in the caudal MAO.
Teleceptive cells in MAO. Visual and auditory cells also cluster in small, well-defined regions within the MA0 as shown in Figure 5B and indicated in summary fashion in Figure 14. Cells responding to a flash of light were found in the dorsal cap, the nearby ventrolateral outgrowth, and the adjacent MAO. Latencies were slightly shorter for the caudally located cells (40-45 msec; lower trace in Fig. 15) than for cells in the more rostral and lateral region (45-50 msec; upper trace). Throughout this teleceptive area the cells followed flashes up to about Blsecond.
In the caudal MAO, near nucleus p, we found six cells responding to auditory stimuli such as keys being jangled or hand claps. Timing their latency from a tap delivered on the frame of the stereotaxic instrument by a mechanical device showed that they responded a t latencies of 45-50 msec. A neighboring region (indicated in Fig. 5B) con- tained a cell which responded to both auditory and visual stimuli. The cells in these teleceptive areas had little spon- taneous activity.
Principal nucleus In the principal nucleus of the inferior olive a relatively
smaller proportion of cells were responsive to somatosen- sory stimuli (43% of 46 cells), and no responses to tele- ceptive stimulation were encountered. In more lateral parts of the PO the cells were largely unresponsive to our stimuli (Fig. 13). There appeared to be a slight difference between the ventral and dorsal lamella with respect to modalities (see below), but the types of receptive fields and organi- zation were similar. In all cases but one the response to a stimulus was an increase in firing rate; the exceptional cell responded upon termination of the stimulus.
Response latencies to shock were intermediate between those measured in DAO and those in MAO. For nine cells responsive to forelimb stimulation the mean was 25 msec. For three hindlimb cells, the mean was 23 msec.
Receptive fields were similar to the less-complicated fields observed in the MA0 and the more complicated ones found in the DAO. Of the responsive cells, nine were activated by taps, seven by squeeze, and two by touch. All but one of the cells responding to taps were located in the dorsal lamella; the cells responding to touch and six of those responding to squeeze were located in the ventral lamella. Of responsive cells, 30% had bilateral receptive fields. No cells were found with fields on the trunk. Although there was no clear somatotopic organization, Figure 13 shows that there was an increased probability of finding cells with fields on the forelimb medially and hindlimb lat- erally, as in the DAO and MAO.
DISCUSSION Our results support the presence of several distinct so-
matosensory representations within the three divisions of the inferior olivary nucleus. The most refined of these is the cutaneous map of the entire contralateral body surface present in the rostral dorsal accessory olive (rDAO). Not only are there individual face, forelimb, hindlimb, trunk, and tail regions, but within each limb zone there are sep- arate orderly progressions in the anterior-posterior and medial-lateral directions (Fig. 8). Moving caudally in the DAO (cDAO) there is a transition from cutaneous to deep responsiveness suggesting the existence of a second map of the contralateral body in which individual limbs, or occasionally bilateral pairs of limbs, are represented.
Our methods discriminate between a medial and lateral region of the principal olive. The medial region of the PO has a preponderance of responsiveness to taps, many bi- laterally symmetric fields, and conforms with the general medial forelimb and lateral hindlimb trend seen in the overlying DAO (Fig. 13). The lateral region appears to be unresponsive to natural stimulation under the conditions of our experiment.
The somatosensory representations in the MA0 are dis- tinguished by a greater complexity of the receptive fields (Fig. 12) and by a preponderance of deep over cutaneous modality. The most intriguing representation is found in the lateral region of the caudal MA0 (cMAO) which con- tains patterned fields made up of excitatory and inhibitory patches (Fig. 14). Purely inhibitory fields are also concen- trated within this region. The rostral and intermediolat- era1 regions (rMAO) appear to contain a separate soma- tosensory representation in which receptive fields consisting of purely excitatory patches are found. In addition to these somatosensory areas, the caudomedial nucleus p region of

240 R. GELLMAN, J.C. HOUK, AND A.R. GIBSON
analysis of pathways was used by Oscarsson and his col- leagues in most cases, while we used natural stimuli for the analysis of receptive fields. Further, the anatomically defined zones of the cerebellar cortex may not correspond precisely to the physiologically defined zones of Oscarsson and his colleagues. In a different study of climbing fiber responses in the vermis Robertson et al. ('82) found that in the medial part of lobule V only 17% of cells were re- sponsive to cutaneous stimulation. The fields were located on the back and the head and were often bilateral. Inhib- itory responses were not described. Their study was lim- ited to cells responsive to cutaneous stimulation, and this fact probably accounts for the small proportion of respon- sive cells.
In the B zone (input from cDAO) Oscarsson ('80) reported that the face, forelimb, trunk, hindlimb, and tail were represented by a series of parasagittal microzones. In the cDAO, in contrast, we found input only from the limbs and face, but not from axial regions. Similarly, in lateral vermal regions Robertson et al. ('82) recorded climbing fiber responses from the limbs only. In their study 25% of cells in this region were unresponsive, while in our study nearly all cells were responsive. This too may be due to the different types of sensory stimulation used.
The C2 zone has been described as being less responsive than the A zone (see Oscarsson and Sjolund, '77a; Ekerot and Larson, '79a). Purkinje cells in the C2 zone were equally responsive to stimuli on all four limbs (Larson et al., '69b), but these responses were obtained only in a small propor- tion of experiments. Long latency responses to bilateral input were reported in this zone by Ekerot and Larson ('79a), but these responses were not seen under barbiturate anesthesia. In our studies, in contrast, there was no clear difference between the rostral and caudal MA0 in res- ponsivity or latency. In both regions about 65% of active cells were responsive to some form of sensory stimulation, and latencies were also comparable (overall mean of 32 msec for rMAO, 34 msec for cMAO). Further, about 50% of responsive cells in the rMAO responded only to contra- lateral inputs.
Recordings in intermediate cortex by Rushmer et al. ('80) revealed receptive fields very similar to those that we have reported in the rDAO. In the studies of Oscarsson and his colleagues the C1 and C3 zones were generally the most responsive zones, corresponding to our findings that cells in the rDAO had low thresholds. The C3 zone also showed a fine somatotopic organization (Ekerot and Lar- son, '79b). It appears from a comparison of these studies with our own that the map formed by the climbing fiber system in the cerebellum represents a reorganization of the map seen in the DAO. The mediolateral somatotopy we report for the DAO appears to be transformed into an anteroposterior organization in the cerebellum (e.g., Os- carsson, '69). Small body regions are represented in elon- gated patches in the intermediate cerebellar cortex ac- cording to Rushmer et al. ('801, or in multiple parasagittal strips in zone C3 (Ekerot and Larson, '79b). Ekerot and Larson ('79b) described a further map in zone C1. The last- mentioned study implies the presence of multiple maps in the projection zone of the rDAO, in contrast to the single map in the rDAO itself. There is an additional projection from the rDAO to the paramedian lobule (cf. Brodal and Kawamura, '80). This diversity of maps may be accounted for by the divergence of climbing fibers which has been observed by Armstrong et al. ('74) using electrophysiolog-
the MA0 and the adjacent dorsal cap and ventrolateral outgrowth appear to constitute a segregated teleceptive region.
The finding of discrete sensory representations does not agree with the only previous mapping study of the olive based on natural stimulation (Sedgwick and Williams, '67). The reported absence of somatotopic organization seemed unlikely given the high degree of anatomical organization seen in both the afferent and efferent connections of the inferior olive (as reviewed below). The recognition of sev- eral somatosensory maps in the present study can be at- tributed to a large sample of neurons (391) and to precision in the histological reconstruction of recording sites, which we estimate to be on the order of 100 pm in individual animals.
The somatosensory map of the DAO that emerged was found to be a reliable guide in later mapping experiments, where we wished to make closely spaced electrode pene- trations to study the fine organization within the rDAO. In addition, we used the map and the location of a known area to determine the appropriate electrode placement for recording in other olivary regions.
Olivocerebellar relations Previous studies of the physiological properties of the
olivocerebellar system have largely focused on recording climbing fiber responses in the cerebellar cortex and have thus provided indirect information on the organization of the inferior olive with which our findings can be compared. Studies of the projection from subnuclei of the inferior olive to the feline cerebellum have shown that the cerebellar cortex can be divided into parasagittal strips named, from medial to lateral, A, B (in the vermis), C1-3 (intermediate cortex), and the D zones (lateral hemispheres). The vermal strips receive input from the caudal accessory nuclei-the A zone from the cMAO, the B zone from the cDAO. Strips in the intermediate cortex receive input from the rostral accessory nuclei-the C2 zone from rMAO, the C1 and C3 zones from rDAO. The D zones receive input from the PO- the D1 zone from the dorsal lamella, the D2 zone from the ventral lamella. (The olivocerebellar projection is re- viewed by Brodal and Kawamura, '80.) Most of these par- asagittal strips have extensions in the posterior lobe.
Recordings in all the above cerebellar zones have re- vealed climbing fiber responses to electrical shocks of limb nerves, though not all zones were equally responsive as judged by latency measurements. In experiments by Os- carsson and his colleagues in cats with only individual funiculi in the spinal cord left intact, the most responsive regions appear to be C1 and C3, followed by B, A, and D2; least responsive were C2 and D1 (Oscarsson and Sjolund, '77a; Ekerot and Larson, '79a). Our description of sensory responses in all olivary regions broadly parallels these findings. There are, however, a number of discrepancies that warrant discussion.
In the A zone (input from cMAO) reliable responses were found to shocks of nerves in the ipsilateral hindlimb only (Oscarsson, '69). In our study responses to stimuli on all four limbs, as well as the face and back, were found in the caudal MAO. We further found that many of the fields in this region contained inhibitory components or were en- tirely inhibitory, whereas inhibition was rarely seen in recordings from Purkinje cells located in the A zone (Os- carsson, '69). These differences may be attributed to dif- ferent methods of stimulation: Electrical stimuli for the

INFERIOR OLIVE ORGANIZATION
ical methods, and by Brodal et al. ('80) using a double- label retrograde tracing technique.
Ekerot and Larson ('79a) reported finding short-latency (10-20 msec) sensory responses in the D2 zone of the an- terior lobe, which receives input primarily from the prin- cipal nucleus (cf. Brodal and Kawamura, '80). They spec- ulated that the responses may stem from the DAO, since the principal nucleus was not known to receive somato- sensory input. These responses may in fact be transmitted through the medial part of the principal nucleus, which we have found to contain many responsive cells.
Sources of somatosensory responsiveness The somatosensory responses we recorded from olivary
neurons could be mediated by direct projections from the spinal cord via several known pathways, or they could reflect transmission through central neurons, where again several pathways may be considered. Considering anatom- ical studies in the cat, two-neuron routes have been de- scribed which relay in medial regions of spinal laminae IV-VII (Armstrong and Schild, '79) and in the dorsal col- umn nuclei (DCN) (Oscarsson, '69; Groenewegen et al., '75; Boesten and Voogd, '75; Berkley and Hand, '78). Less direct pathways in the cat traverse the reticular formation (Walberg, '82b), the cerebellar nuclei (Tolbert et al., '76; Berkley and Worden, '78), several mesencephalic sites (Saint-Cyr and Courville, '821, and the cerebral cortex (Sousa-Pinto and Brodal, '69), though the last mentioned is now thought to provide a rather minor input (Saint-Cyr and Courville, '80). Our consideration of possible corre- spondences between input pathways and responses to nat- ural stimulation will deal first with the direct pathways, since it is likely that the circuitous routes were relatively depressed by the barbiturate anesthesia used in our study.
The most likely source of the refined cutaneous map found in the rDAO is the somatotopically organized input from the DCN and the spinal trigeminal nucleus. The gra- cile nucleus projects laterally in the DAO (Groenewegen et al., '75; Boesten and Voogd, '75; Berkley and Hand, '78). while the cuneate nucleus projects more medially (Boesten and Voogd, '75; Berkley and Hand, '78). The trigeminal projection partially overlaps with that from the cuneate nucleus and extends more medially (Berkley and Hand, '78; see also Walberg, '82a). Our results support this so- matotopy and show a detailed topographic organization beyond that indicated by the anatomical studies. The pa - cile and cuneate nuclei are somatotopically organized (see Willis and Coggeshall, '78, for review) and receptive fields are purely ipsilateral. Distal receptive fields are small; proximal fields, particularly those on the trunk, tend to be larger. In these aspects neurons in the rDAO resemble those in the DCN. In addition, most cells in the cuneate and gracile nuclei respond to cutaneous stimulation, em- phasizing that these are a likely source of the responses that we observed in the DAO.
Dorsal column input is also distributed to the cDAO and cMAO, yet these regions do not respond as well as the DAO to light cutaneous stimulation under our experi- mental conditions. This may be an effect of the anesthesia, or it may indicate that the regions of the DCN projecting to these caudal portions of the inferior olive differ from those projecting to the rDAO. The deep responses observed in the cDAO and cMAO may nonetheless be transmitted via the DCN, which has been shown to contain cells re- sponsive to deep input. The somatotopic organization in
241
the cDAO is predicted by the anatomical studies cited above. In the cMAO, however, the cuneate and gracile projections overlap (Berkley and Hand, '781, although we have noted that there is a broad mediolateral somatotopy in this re- gion (Fig. 13).
The pathways from spinal neurons may convey the deep input that is found to some degree in the rDAO and more extensively in the cDAO and cMAO. Boesten and Voogd ('75) showed that the DAO and caudal two-thirds of the MA0 receive direct inputs from the spinal cord. The pro- jection from cervical spinal areas terminates in the same region of the DAO as those from the cuneate; that from the lumbar cord is coextensive with the gracile projection, thus supporting the mediolateral somatotopy that we have described. In the cMAO the lumbar projection is also lo- cated lateral to that from the cervical cord, a finding which might account for the trend toward a mediolateral organ- ization that we noted in the MAO.
Oscarsson and his colleagues have demonstrated several spinal pathways terminating in the DAO and cMAO using electrophysiological techniques. Many of the pathways were activated by flexor reflex afferents (Oscarsson, '69; 0 s - carsson and Sjolund, '77b). In our study we could not be certain whether the deep input was from high-threshold mechanoreceptors or from low-threshold muscle propri- oceptors that required a massive activation, produced by a squeeze, in order to drive spinal relay neurons above threshold. This question needs to be addressed in the alert animal.
Two regions that receive neither direct spinal nor dorsal column input, namely, the rMAO and the PO, nevertheless contained many neurons that were responsive to soma- tosensory stimuli. The responsive cells in these two regions were similar with respect to the type of receptive fields and modality, but differed in that some inhibitory fields were found in the rMAO but not the PO. The overall mean latency was also somewhat shorter in the PO than rMAO (22 vs. 33 msec). Possible sources for the responses in both PO and rMAO include the cerebellar nuclei, the reticular formation, sensory cortex, and mesencephalon. Transmis- sion via such indirect pathways may account for the long latency of many responses in the cMAO, rMAO, and PO. Another possible explanation for the very long latencies (>150 msec) found occasionally in the MA0 is transmis- sion via slowly conducting afferents, such as C-fibers (cf. O'Connor and Vangilder, '81).
Teleceptive representation We recorded responses to teleceptive input in the dorsal
cap, the ventrolateral outgrowth, and medial regions of the MA0 (nucleus p), (the region labeled T in Fig. 14). Responses to auditory stimuli were found in medial MAO, adjacent to nucleus p. This region projects to posterior vernal regions (cf. Brodal and Kawamura, '80) apparently corresponding with the cerebellar region in which Snider and Stowell ('44) recorded surface potentials in response to clicks. There is also a brief note reporting auditory climbing fiber responses in the paraflocculus (Azizi et al., '81). The source of the auditory input to the inferior olive might be deep collicular areas which are known to project to caudomedial regions of the MA0 (Saint-Cyr and Cour- ville, '82).
We recorded both field potentials and cellular responses to a flash of light in the dorsal cap, which agrees with previous findings in the rabbit (Barmack and Hess, '80).

242 R. GELLMAN, J.C. HOUK, AND A.R. GIBSON
dition seem to be less extensively represented, and the representation that is present probably has highly spe- cialized properties (see Barmack and Hess, '80). In other parts of the nervous system visual space, auditory space, and body surface are all represented as finely detailed maps, and much of this information is available to the cerebellum via its mossy fiber input (cf. Llinas and Simp- son, '81). Why is the climbing fiber input so heavily de- voted to body surface? To make accurate movements rel- ative to objects in the world one must represent the position of these objects relative to the position of the body. Perhaps contact and proprioceptive information arriving via the climbing fiber system provides the cerebellum with infor- mation necessary to relate the body to external objects.
These findings conflict with a report by Haddad et al. ('80) in which flash responses were not observed in the dorsal cap of the anesthetized cat. The source of visual input to the dorsal cap is probably the pretectal nuclei, particularly the nucleus of the optic tract. This nucleus has been shown to project to the dorsal cap in the rabbit (Mizuno et al., '73) and is known to respond to visual stimuli (Hoffmann and Schoppmann, '75). In the cat the pretectal nuclei pro- ject to the inferior olive (Hoffmann et al., '76; Saint-Cyr and Courville, %1), but it is not known whether the fibers terminate specifically in the dorsal cap. The deep layers of the superior colliculus, which project to a region of the MA0 near nucleus p (Saint-Cyr and Courville, '821, may also be considered a possible source of visual input to the olive.
An additional source of input to these areas is provided by the vestibular nuclei (Saint-Cyr and Courville, '79). Although we were not able to present vestibular stimu- lation in our experiments, it is likely that there are neu- rons in this region which are responsive to such a stimulus. This would lend credence to the idea that this region of the inferior olive is concerned with information regarding the orientation of the body in space, which is provided by the classical teleceptive senses, namely, vision and audi- tion, as well as the vestibular sense.
Functional considerations The massive anatomical linkage from the inferior olive
to the cerebellar cortex provided by climbing fibers insures an intimate relationship between olivary activity and the presumed motor functions of the cerebellum (cf. Arm- strong, '74; Szentagothai and Arbib, '74). Just what these functions might be has remained elusive. Recent specu- lation has centered around two basic ideas. One concept postulates that olivary activity acts as a detection andlor trigger mechanism for corrective movements (Llinas, '70; Rushmer et al., '76; Houk, '79; Barmack and Hess, '80; Oscarsson, '80). The other idea postulates that climbing fiber activity exerts a long-term influence on transmission between the mossy fiber system and Purkinje cells, for example, as a mechanism for motor learning (Marr, '69; Gilbert and Thach, '77; Ito, '81). The information reported here provides important constraints on these and other functional theories.
Single units in the inferior olive do not seem to provide information about stimulus parameters such as modality or intensity, as they fire only one or two spikes for any adequate stimulus, be it a light puff or a sharp tap. This would seem to mean that these neurons are signalling the occurrence of events. In the case of the DAO, for example, the event in question would be contact with an object. Alternatively, the population could signal intensity, as indicated by Boylls ('801, who found that in multiunit re- cordings in the MA0 neural activity was correlated with the step-cycle, while no such correlation was seen in study- ing single units. This may also be the implication of the findings of Rushmer et al. ('76) and Kolb and Rubia ('801, who demonstrated that the activity of a single climbing fiber, averaged over many trials, appeared to code for var- ious parameters of movement.
The most striking characteristic of the inferior olive emerging from our study is the highly detailed map of the body surface, which is particularly evident in the DAO. Even the MA0 and PO have many cells which respond to somatosensory stimuli. On the other hand, vision and au-
ACKNOWLEDGMENTS This work was supported by USPHS grant Pol-NS17489
to JCH. The authors thank Dannie Hansma for prepara- tion of the figures.
LITERATURE CITED Armstrong, D.M. (19741 Functional significance of the connections of the
inferior olive. Physiol. Rev. 54:358-417. Armstrong, D.M., and R.J. Harvey (1966) Responses in the inferior olive
to stimulation of cerebellar and cerebral cortices in the cat. J. Physiol. 187:553-574.
Armstrong, D.M., J.C. Eccles, R.J. Harvey, and P.B.C. Matthews (1968) Responses in the dorsal accessory olive of the cat to stimulation of hind limb afferents. J. Physiol. 194:125-145.
Armstrong, D.M., R.J. Harvey, and R.F. Schild (1974) Topographical lo- calization in the olivo-cerebellar projection: An electrophysiological study in the cat. J. Comp. Neurol. 154:287-302.
Armstrong, D.M., and J.A. Rawson (1979) Activity patterns of cerebellar cortical neurones and climbing fibre afferents in the awake cat. J. Physiol. 289:425-448.
Armstrong, D.M., and R.F. Schild (1979) Spino-olivary neurones in the lumbo-sacral cord of the cat demonstrated by retrograde transport of horseradish peroxidase. Brain Res. 168:176-179.
Azizi, A,, R.A. Burne, and D.J. Woodward (1981) Visual and auditory input and output relations of the lateral cerebellum: The paraflocculus. SOC. Neurosci. Abstr. 7t77.
Barmack, N.H., and D.T. Hess (1980) Multiple unit activity evoked in dorsal cap of inferior olive of the rabbit by visual stimulation. J. Neu- rophysiol. 43:151-164.
Berkley, K.J., and P.J. Hand (1978) Projections to the inferior olive of the cat. 11. Comparisons of input from the gracile, cuneate and spinal tri- geminal nuclei. J. Comp. Neurol. 180:253-264.
Berkley, K.J., and I.G. Worden (1978) Projections to the inferior olive of the cat. 1. Comparisons of input from the dorsal column nuclei, the lateral cervical nucleus, the spino-olivary pathways, the cerebral cortex and the cerebellum. J. Comp. Neurol. 180:237-252.
Berman, A.L. (1968) The Brain Stem of the Cat. A Cytoarchitectonic Atlas with Stereotaxic Coordinates. Madison: Univ. of Wisconsin Press.
Boesten, A.J.P., and J. Voogd (1975) Projections of the dorsal column nuclei and the spinal cord on the inferior olive in the cat. J. Comp. Neurol. 161:215-238.
Boylls, C.C. (1980) Contributions to locomotor coordination of an olivocer- ebellar projection to the vermis in the cat: Experimental results and theoretical proposals. In J. Courville, C. de Montigry, and Y. Lamarre (eds): The Inferior Olivary Nucleus: Anatomy and Physiology. New York: Raven Press. pp. 321-348.
Brodal, A. (1940) Experimentelle Untersuchungen ueber die Olivo-cere- bellare Lokalisation. Z. Gesamte Neurol. Psychiatr. 169:l-153.
Brodal, A,, and K. Kawamura (1980) Olivocerebellar Projection: A Review. New York Springer-Verlag.
Brodal, A,, F. Walberg, K.J. Berkley, and A. Pelt (1980) Anatomical dem- onstration of branching olivocerebellar fibres by means of a double retrograde labelling technique. Neuroscience 52193-2202.

INFERIOR OLIVE ORGANIZATION 243
Man-, D. (1969) A theory of cerebellar cortex. J. Physiol. 202:437470. Mizuno, N., K. Mochizuki, C. Akimoto, and R. Matsushima (1973) Pretectal
projections to the inferior olive in the rabbit. Exp. Neurol. 39:498-506. O’Connor, G.A., and J.C. Vangilder (1981) Spinal cord conduction pathway
for cerebellar evoked potentials from C-fibers. Brain Res. 21 7t192-195. Oscarsson, 0. (1968) Termination and functional organization of the ven-
tral spino-olivocerebellar path. J. Physiol. 196t45-78. Oscarsson, 0. (1969) Termination and functional organization of the dorsal
spino-olivocerebellar path. J. Physiol. 200~129-149. Oscarsson, 0. (1980) Functional organization of olivary projection to the
cerebellar anterior lobe. In J. Courville, C. de Montigry, and Y. Lamarre (eds): The Inferior Olivary Nucleus: Anatomy and Physiology. New York: Raven Press. pp. 279-289. system in the cat. I. Identification of five paths and their termination in the cerebellar anterior lobe. Exp. Brain Res. 28t469-486.
Oscarsson, O., and B. Sjolund (1977b) The ventral spino-olivocerebellar system in the cat. 111. Functional characteristics of the five paths. Exp. Brain Res. 28:505-520.
Robertson, L.T., K.D. Laxer, and D.S. Rushmer (1982) Organization of climbing fiber input from mechanoreceptors to lobule V vermal cortex of the cat. Exp. Brain. Res. 46:281-291.
Rushmer, D.S., W.J. Roberts, and G.K. Augter (1976) Climbing fiber re- ponses of cerebellar Purkinje cells to passive movement of the cat fore- paw. Brain Res. 106:l-20.
Rushmer, D.S., M.H. Woollacott, L.T. Robertson, and K.D. Laxer (1980) Somatotopic organization of climbing fiber projections from low thresh- old cutaneous afferents to pars intermedia of cerebellar cortex in the cat. Brain Res. 181:17-30.
Saint-Cyr, J.A., and J. Courville (1979) Projection from the vestibular nuclei to the inferior olive in the cat: An autoradiographic and horse- radish peroxidase study. Brain Res. 165t189-200.
Saint-Cyr, J.A., and J. Courville (1980) Projections from the motor cortex, midbrain, and vestibular nuclei to the inferior olive in the cat: Ana- tomical organization and functional correiates. In J. Courville e t al. (eds): The Inferior Olivary Nucleus: Anatomy and Physiology. New York: Raven Press. pp. 97-124.
Saint-Cyr, J.A., and J. Courville (1981) Sources of descending afferents to the inferior olive from the upper brain stem in the cat as revealed by the retrograde transport of horseradish peroxidase. J. Comp. Neurol. 198:567-581.
Saint-Cyr, J.A., and J. Courville (1982) Descending projections to the in- ferior olive from the mesencephalon and superior colliculus in the cat. Exp. Brain Res. 45~333348.
Sedgwick, E.M., and T.D. Williams (1967) Responses of single units in the inferior olive to stimulation of the limb nerves, peripheral skin recep- tors, cerebellum, caudate nucleus and motor cortex. J. Physiol. 189~261-279.
Snider, R.S., and A. Stowell (1944) Receiving areas of the tactile, auditory and visual systems in the cerebellum. J. Neurophysiol. 7:331357.
Sousa-Pinto, A,, and A. Brodal (1969) Demonstration of a somatotopical pattern in the cortico-olivary projection in the cat: An experimental- anatomical study. Exp. Brain Res. 8:364-386.
Szentagothai, J., and M.A. Arbib (1974) Conceptual Models of Neural Or- ganization. Neurosci. Res. Prog. Bull. 12.
Thach, W.T. (1967) Somatosensory receptive fields of single units in cat cerebellar cortex. J. Neurophysiol. 30~675-696.
Tolbert, D.L., L.C. Massopust, M.G. Murphy, and P.A. Young (1976) The anatomical organization of the cerebello-olivary projection in the cat. J. Comp. Neurol. 170:525-544.
Walberg, F. (1982a) The trigemino-olivary projection in the cat as studied with retrograde transport of horseradish peroxidase. Exp. Brain Res. 45:lOl-107.
Walberg, F. (198213) Olivary afferents from the brain stem reticular for- mation. Exp. Brain Res. 47t130-136.
Willis, W.D., and R.E. Coggeshall(1978) Sensory Mechanisms ofthe Spinal Cord. New York: Plenum Press.
Wilson, W.C., and H.W. Magoun (1945) The functional significance of the inferior olive in the cat. J. Comp. Neurol. 83:69-77.
Brooks, V.B., P.R. Kennedy, and H.-G. Ross (1980) Does cooling monkey’s inferior olive increase biceps stretch reflex? In J. Courville e t al. (eds): The Inferior Olivary Nucleus: Anatomy and Physiology. New York: Raven Press. pp. 361-365.
Carrea, R.M.E., M. Reissig, and F.A. Mettler (1947) The climbing fibers of the simian and feline cerebellum: Experimental inquiry into their or- igin by lesions of the inferior olives and deep cerebellar nuclei. J. Comp. Neurol. 87:321-365.
Courville, J., and F. Faraco-Cantin (1978) On the origin of the climbing fibers of the cerebellum. An experimental study in the cat with a n autoradiographic tracing method. Neuroscience 3r797-809.
Crill, W.E. (1970) Unitary multiple-spiked responses in cat inferior olive nucleus. J. Neurophysiol. 33~199-209.
Eccles, J.C., R. Llinas, and K. Sasaki (1966) The excitatory synaptic action of climbing fibers on the Purkinje cells of the cerebellum. J. Physiol. 182.968-296.
Eccles, J.C., N.H. Sabah, R.F. Schmidt, and H. Taborikova (1972) Cuta- neous mechanoreceptors influencing impulse discharges in cerebellar cortex. 111. In Purkinje cells by climbing fiber input. Exp. Brain Res. 15r484497.
Ekerot, C.-F., and B. Larson (1979a) The dorsal spino-olivocerebellar sys- tem in the cat. I . Functional organization and termination in the an- terior lobe. Exp. Brain Res. 36t201-217.
Ekerot, C.-F., and B. Larson (1979b) The dorsal spino-olivocerebellar sys- tem in the cat. 11. Somatotopical organization. Exp. Brain Res. 36:219-232.
Gilbert, P.F.C., and W.T. Thach (1977) Purkinje cell activity during motor learning. Brain Res. 128t309-328.
Groenewegen, H.J., A.J.P. Boesten, and J. Voogd (1975) The dorsal column nuclear projections to the nucleus ventralis posterior lateralis thalami and the inferior olive in the cat: An autoradiographic study. J. Comp. Neurol. 162:505-518.
Haddad, G.M., J.L. Demer, and D.A. Robinson (1980) The effect of lesions of the dorsal cap of the inferior olive on the vestibulo-ocular and op- tokinetic systems of the cat. Brain Res. 185:265-275.
Hoffmann, K.-P., and A. Schoppmann (1975) Retinal input to direction selective cells in the nucleus tractus opticus of the cat. Brain Res. 99~359-366.
Hoffmann, K.-P., K. Behrend, and A. Schoppmann (1976) A direct afferent visual pathway from the nucleus of the optic tract to the inferior olive in the cat. Brain Res. 115t150-153.
Houk, J.C. (1979) Motor control processes: New data concerning motoservo mechanism and a tentative model for SR processing. In R.E. Talbot and D.R. Humphrey (eds): Posture and Movement. New York: Raven Press. pp. 231-241.
Ishikawa, K., S. Kawaguchi, and M.J. Rowe (1972a) Actions of afferent impulses from muscle receptors on cerebellar Purkinje cells. I. Re- sponses to muscle vibration. Exp. Brain Res. 15:177-193.
Ishikawa, K., S. Kawaguchi, and M.J. Rowe (1972b) Actions of afferent impulses from muscle receptors on cerebellar Purkinje cells. 11. Re- sponses to muscle contraction: Effects mediated via the climbing fiber pathway. Exp. Brain Res. 16~104-114.
Ito, M. (1981) Experimental evidence for cerebellar motor learning. In 0. Pompeiano and C.A. Marsan (eds): Brain Mechanisms and Perceptual Awareness. New York: Raven Press. pp. 261-267.
Kolb, F.P., and F.J. Rubia (1980) Information about peripheral events conveyed to the cerebellum via the climbing fiber system in the decer- ebrate cat. Exp. Brain Res. 38:363-373.
Larson, B., S. Miller, and 0. Oscarsson (1969a) Termination and functional organization of the dorsolateral spino-oiivocerebellar path. J. Physiol. 203t611-640.
Larson, B., S. Miller, and 0. Oscarsson (1969b) A spinocerebellar climbing fiber path activated by the flexor reflex afferents from all four limbs. J. Physiol. 203:641-649.
Llinas, R. (1970) Neuronal operations in cerebellar transactions. In F.O. Schmitt (ed): The Neurosciences: Second Study Program. New York: Rockefeller University Press. pp. 409-426.
Llinas, R.R., and J.I. Simpson (1981) Cerebellar control of movement. In A.L. Towe and E.S. Luschei (eds): Handbook of Behavioral Neurobiol- ogy, Vol. 5. New York Plenum Press. pp. 231-302.





























