Embed Size (px)
Citation preview

R
EPRODUCTIONRESEARCHSonic hedgehog supplementation of oocyte and embryo culturemedia enhances development of IVF porcine embryos
Ngoc Tan Nguyen1,2, Neng-Wen Lo3, Sing-Ping Chuang1, Ya-Lan Jian1 and Jyh-Cherng Ju1
1Department of Animal Science, National Chung Hsing University, 250 Kuokuang Road, Taichung 402, Taiwan, ROC,2Agricultural Biotechnology Center, Institute of Agricultural Sciences for Southern Vietnam, 121 Nguyen Binh KhiemStreet, Ho Chi Minh City, Vietnam and 3Department of Animal Science and Biotechnology, Tunghai University,181 Section 3, Taichung Harbor Road, Taichung 407, Taiwan, ROC
Correspondence should be addressed to J-C Ju; Email: [email protected]
Abstract
We investigated the expression of sonic hedgehog (SHH) receptor PTCH1 and its co-receptor smoothened (SMO) in fertilized porcine
embryos. Effects of exogenous SHH on embryonic development and expressions of survival- and pluripotency-related genes were also
determined. We found that PTCH1 and SMO are expressed from two-cell to blastocyst embryos. When oocytes or fertilized embryos
were respectively cultured in the maturation or embryo culture medium supplemented with SHH (0.5 mg/ml), their blastocyst rates and
total cell numbers increased (P!0.05) compared with the untreated control. When cultured simultaneously in the in vitro maturation
(IVM) and in vitro culture (IVC) media supplemented with SHH, the oocytes gained increased blastocyst rates and total cell numbers in
an additive manner, with reduced apoptotic indices (P!0.05). Interestingly, SHH treatment did not affect the expression of the BCL2L1
(BCL-XL) gene, yet reduced BAX expression. Blastocysts cultured with various SHH regimes had similar pluripotency-related gene
(POU5F1 (OCT-4) and CDX2) expression levels, but blastocysts derived from SHH treatment during IVM had higher ZPF42 (REX01)
expression (P!0.05). The highest ZPF42 expression was observed in the blastocysts derived from SHH-supplemented IVC and from
dual IVM and IVC treatments. The levels of acetylated histone 3 (AcH3K9/K14) increased in the two-cell and the four-cell embryos
when IVM and/or IVC media were supplemented with SHH (P!0.05). Our findings indicate that SHH conferred a beneficial effect
on preimplantation development of porcine embryos, particularly when both IVM and IVC media were supplemented with SHH, and
the effects may be further carried over from IVM to the subsequent embryonic development.
Reproduction (2011) 142 87–97
Introduction
The porcine animal model has been broadly applied inmany aspects of biochemical research (Prather et al.2003), and porcine embryos hold great promise forembryonic stem cell technologies (Beebe et al. 2009).Despite efforts that have been made to optimize thein vitro production (IVP) system of embryos, the yieldand the quality of IVP embryos are still low whencompared with their in vivo-produced counterparts(Abeydeera et al. 1998, Kim et al. 2006, Ock et al.2007). It is well known that oocyte and embryo qualityhas been closely associated with the expression of aseries of genes during early embryogenesis (Lonerganet al. 2006, Zheng & Dean 2007), exerting a significantinfluence on the pre- and the post-implantationembryonic development to term (Krisher 2004, Sirardet al. 2006, Mtango et al. 2008).
A great majority of improvements in oocyte exploita-tion rely heavily on the accumulated knowledge
q 2011 Society for Reproduction and Fertility
ISSN 1470–1626 (paper) 1741–7899 (online)
concerning the in vitro culture (IVC) system of theoocytes and embryos up to the blastocyst stage (VanLangendonckt et al. 1997, Lonergan et al. 2003, Rizoset al. 2003). By altering the culture conditions of oocytematuration and embryo development, patterns of geneexpression could be manipulated and reprogrammedto closely mimic in vivo conditions, and, as a result,enhance embryo quality. In spite of the common use ofdefined media for in vitro maturation (IVM) of oocytesand IVC of embryos in many species, porcine oocyte-and/or blastocyst-promoting agents to improve theyield and quality of IVP embryos are still under extensiveinvestigation (Thompson 2000). However, little con-sensus has been reached regarding the mechanisms ofaction of growth factors, cytokines, vitamins, or aminoacids added to culture media (Craig et al. 2004, Kimet al. 2006, Gupta et al. 2007, Uhm et al. 2007).
Hedgehog (Hh) signaling molecules exist in the rodent(Wijgerde et al. 2005, Russell et al. 2007, Ren et al.2009) and bovine (Spicer et al. 2009) reproductive
DOI: 10.1530/REP-11-0049
Online version via www.reproduction-online.org
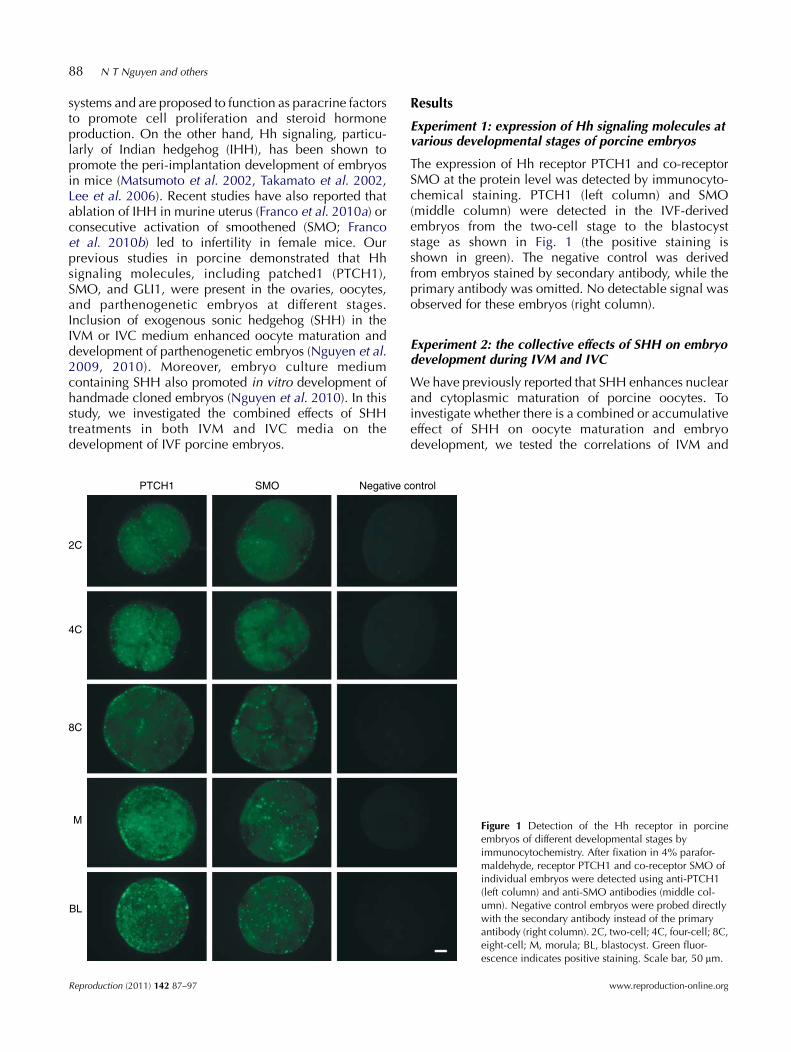
88 N T Nguyen and others
systems and are proposed to function as paracrine factorsto promote cell proliferation and steroid hormoneproduction. On the other hand, Hh signaling, particu-larly of Indian hedgehog (IHH), has been shown topromote the peri-implantation development of embryosin mice (Matsumoto et al. 2002, Takamato et al. 2002,Lee et al. 2006). Recent studies have also reported thatablation of IHH in murine uterus (Franco et al. 2010a) orconsecutive activation of smoothened (SMO; Francoet al. 2010b) led to infertility in female mice. Ourprevious studies in porcine demonstrated that Hhsignaling molecules, including patched1 (PTCH1),SMO, and GLI1, were present in the ovaries, oocytes,and parthenogenetic embryos at different stages.Inclusion of exogenous sonic hedgehog (SHH) in theIVM or IVC medium enhanced oocyte maturation anddevelopment of parthenogenetic embryos (Nguyen et al.2009, 2010). Moreover, embryo culture mediumcontaining SHH also promoted in vitro development ofhandmade cloned embryos (Nguyen et al. 2010). In thisstudy, we investigated the combined effects of SHHtreatments in both IVM and IVC media on thedevelopment of IVF porcine embryos.
PTCH1
2C
4C
8C
M
BL
SMO Negative c
Reproduction (2011) 142 87–97
Results
Experiment 1: expression of Hh signaling molecules atvarious developmental stages of porcine embryos
The expression of Hh receptor PTCH1 and co-receptorSMO at the protein level was detected by immunocyto-chemical staining. PTCH1 (left column) and SMO(middle column) were detected in the IVF-derivedembryos from the two-cell stage to the blastocyststage as shown in Fig. 1 (the positive staining isshown in green). The negative control was derivedfrom embryos stained by secondary antibody, while theprimary antibody was omitted. No detectable signal wasobserved for these embryos (right column).
Experiment 2: the collective effects of SHH on embryodevelopment during IVM and IVC
We have previously reported that SHH enhances nuclearand cytoplasmic maturation of porcine oocytes. Toinvestigate whether there is a combined or accumulativeeffect of SHH on oocyte maturation and embryodevelopment, we tested the correlations of IVM and
ontrol
Figure 1 Detection of the Hh receptor in porcineembryos of different developmental stages byimmunocytochemistry. After fixation in 4% parafor-maldehyde, receptor PTCH1 and co-receptor SMO ofindividual embryos were detected using anti-PTCH1(left column) and anti-SMO antibodies (middle col-umn). Negative control embryos were probed directlywith the secondary antibody instead of the primaryantibody (right column). 2C, two-cell; 4C, four-cell; 8C,eight-cell; M, morula; BL, blastocyst. Green fluor-escence indicates positive staining. Scale bar, 50 mm.
www.reproduction-online.org
Table 1 Effects of sonic hedgehog supplementation in maturation and culture media on the development of porcine embryos in vitro.
SHH (0.5 mg/ml)
IVM IVC Total (N)Cleavage rate,
% (n)Blastocyst rate,
% (n)Cell number per
blastocystApoptotic index,
% (n)
K K 226 79.3G3.4a (179) 25.0G1.4a (56) 46.4G3.3a 4.5G0.4a (19)C K 214 84.3G3.1a (181) 37.3G1.2b (79) 57.4G3.7b 3.6G0.3bc (22)K C 220 81.2G2.1a (179) 33.8G2.1b (73) 59.1G2.2ab 3.2G0.3ab (25)C C 214 85.5G1.8a (182) 42.8G1.7c (90) 69.7G3.2c 2.4G0.2c (27)
Numbers in parentheses represent the number of embryos in each category. Data are meanGS.E.M. from six replicates. a–cWithin a column, meanswithout the same superscript letters differed (P!0.05). IVM, in vitro maturation; IVC, in vitro culture.
SHH enhances development of IVF porcine embryos 89
IVC media in the presence or absence of SHH with theend point development of porcine embryos in vitro. Asshown in Table 1, consistent with what we had foundpreviously, presence of SHH in IVM or IVC mediumalone enhanced blastocyst formation (37.3 and 33.8%respectively) compared with the control group (25%,P!0.05). The improvement of blastocyst formationreached the highest level when SHH was included inboth IVM and IVC media (IVMC/IVCC) compared withthe control (42.8 vs 25%, P!0.05) and the other groups,IVMC/IVCK and IVMK/IVCC. In addition, the total cellnumber per blastocyst increased with the presence ofSHH in IVM (54.1G3.7), in IVC (59.1G2.2) and in bothmedia (69.7G3.2) yet remained unaltered in the controlgroup (46.4G3.3, P!0.05). However, no differenceswere observed among the three treatment groups. Theapoptotic indices were significantly reduced when SHHwas present in IVC (3.2G0.3%) or in the IVMC/IVCC
group (2.4G0.2%) compared with the control group(4.6G0.4%). The apoptotic indices were similarbetween the IVMC/IVCK (3.6G0.3%) and the IVMK/IVCC groups, and between the IVMC/IVCK and thecontrol groups (PO0.05).
An additional experiment was designed to evaluatethe fertilization rate after IVM, and the results arepresented in Table 2 and Fig. 2. Sperm penetrationrates were similar among treatment groups (85.5–87.5%). With the presence of SHH in IVM or in bothIVM and IVC media, the monospermic rate (85.0 and
Table 2 Effects of sonic hedgehog supplementation to in vitro maturation a
SHH (0.5 mg/ml)Number of
oocytes examinedPenetration
% (n)aIVM IVC
K K 96 86.4G0.5 (C K 92 85.5G2.2 (K C 88 87.5G1.7 (C C 95 86.5G1.7 (
Numbers in parentheses represent the actual number of oocytes examined.without the same superscript letters differed (P!0.05). IVM, in vitro maturaMonospermic rate (%): number of monospermic oocytes/total penetratedoocytes/total inseminated oocytes!100.
www.reproduction-online.org
85.7% respectively) increased significantly comparedwith the control group (67.4%) or the IVMK/IVCC group(68.7%). Similarly, a significant improvement of normalfertilization rate was also found when SHH was added inIVM medium alone (72.6%) or in both media (74.2%;control group, 58.2%).
Experiment 3: effects of SHH treatment in IVM andIVC media on histone acetylation profile
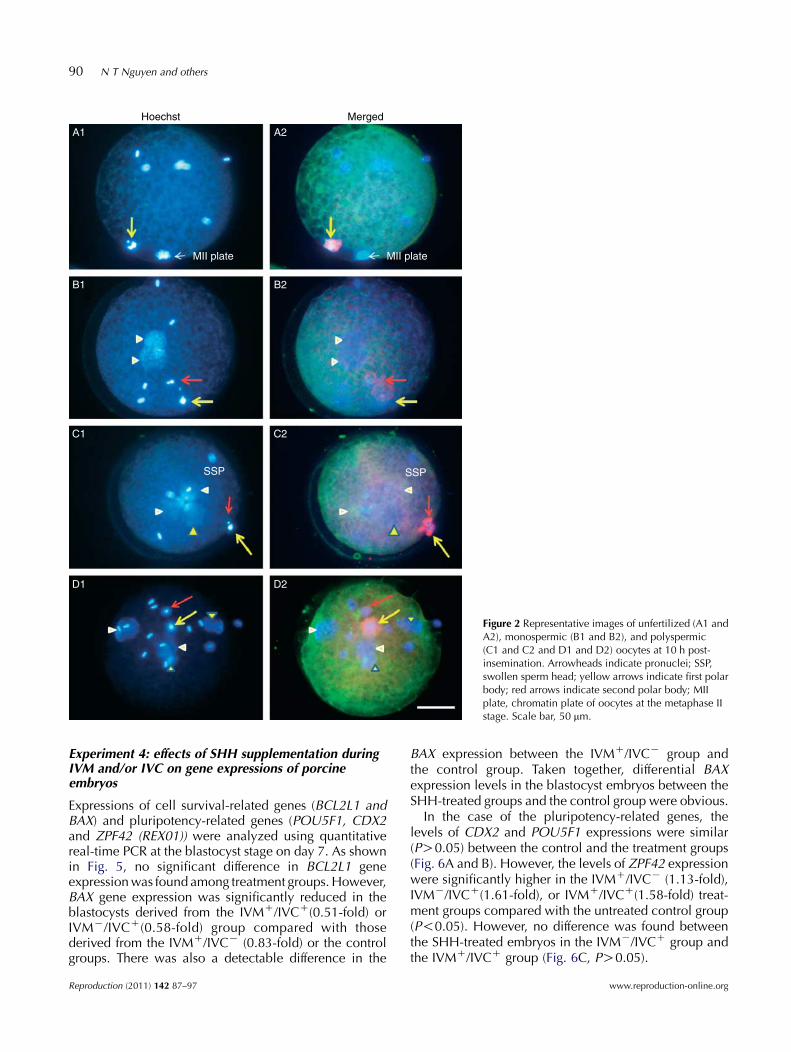
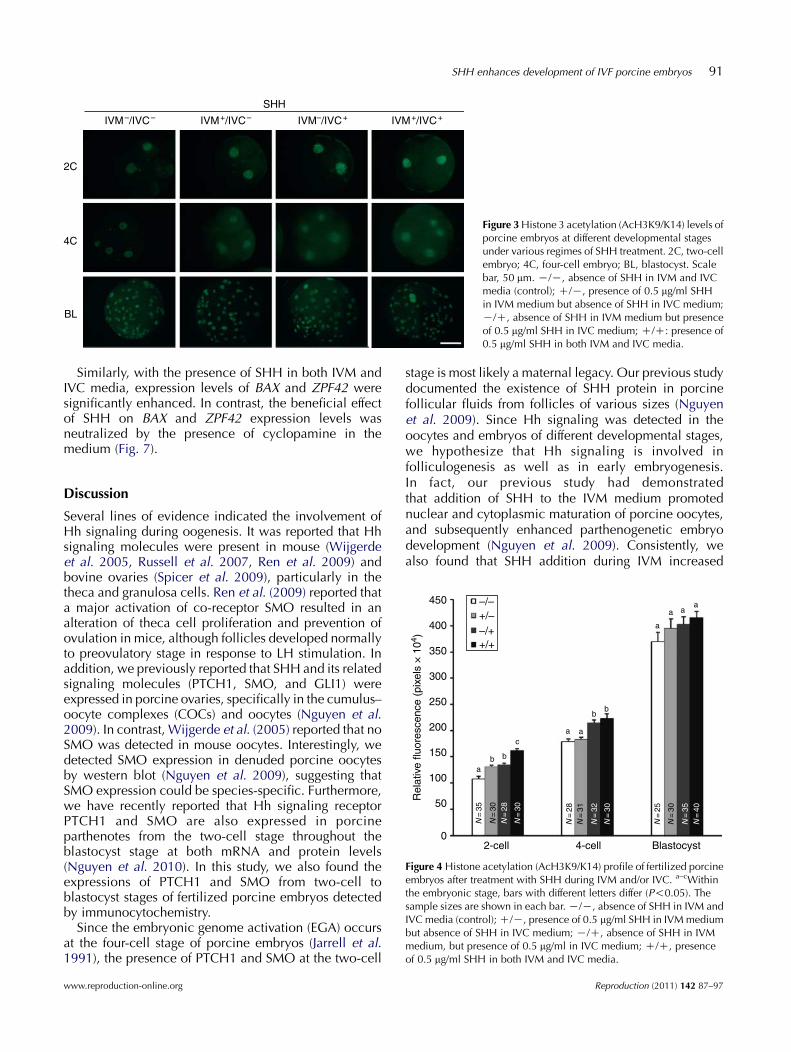
Levels of the histone 3 acetylation or AcH3K9/K14 ofporcine embryos at different stages were determined.As shown in Figs 3 and 4, the level of AcH3K9/K14 wassignificantly increased (P!0.05) at the two-cell stage inthe presence of SHH, either in IVM (130.5G3.7) orin IVC (134.6G3.2) medium alone, and in both media(161.1G4.7) compared with the control group (108.2G4.2). Among the SHH-treated groups, the AcH3K9/K14 levels in the dual SHH-treated embryos (IVMC/IVCC) were significantly higher than those in theembryos with single SHH treatment (IVMC/IVCK orIVMK/IVCC). At the four-cell stage, embryos derivedfrom IVMK/IVCC (214.1G5.8) and IVMC/IVCC treat-ments (223.3G8.0) had a significantly higher level ofacetylation pattern than those of the control (178.9G5.3)or from IVMC/IVCK treatment (182.9G5.0). However,embryos developing up to the blastocyst stage had onlya similar level of acetylation between the control andthe treatment groups.
nd culture media on IVF rates.
rate, Monospermic rate,% (n)
Normal fertilizationrate, %b
73) 67.4G2.7a (57) 58.2G2.5a
80) 85.0G2.1b (68) 72.6G2.3b
77) 68.7G5.5a (54) 60.0G4.2a
83) 83.7G1.4b (71) 74.2G1.2b
Data are meanGS.E.M. from four replicates. a,bWithin a column, meansation; IVC, in vitro culture.oocytes!100. bNormal fertilization rate (%): number of monospermic
Reproduction (2011) 142 87–97
A1 A2
B1 B2
C1 C2
D1 D2
SSP SSP
MII plateMII plate
Hoechst Merged
Figure 2 Representative images of unfertilized (A1 andA2), monospermic (B1 and B2), and polyspermic(C1 and C2 and D1 and D2) oocytes at 10 h post-insemination. Arrowheads indicate pronuclei; SSP,swollen sperm head; yellow arrows indicate first polarbody; red arrows indicate second polar body; MIIplate, chromatin plate of oocytes at the metaphase IIstage. Scale bar, 50 mm.
90 N T Nguyen and others
Experiment 4: effects of SHH supplementation duringIVM and/or IVC on gene expressions of porcineembryos
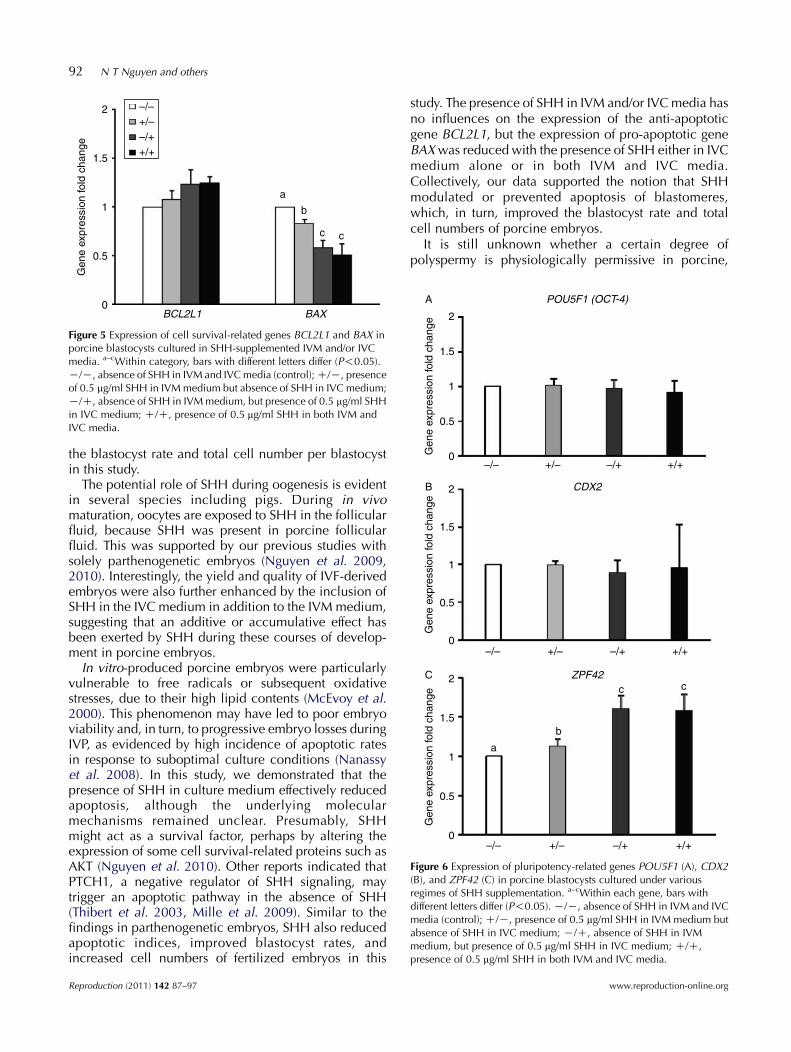
Expressions of cell survival-related genes (BCL2L1 andBAX) and pluripotency-related genes (POU5F1, CDX2and ZPF42 (REX01)) were analyzed using quantitativereal-time PCR at the blastocyst stage on day 7. As shownin Fig. 5, no significant difference in BCL2L1 geneexpression was found among treatment groups. However,BAX gene expression was significantly reduced in theblastocysts derived from the IVMC/IVCC(0.51-fold) orIVMK/IVCC(0.58-fold) group compared with thosederived from the IVMC/IVCK (0.83-fold) or the controlgroups. There was also a detectable difference in the
Reproduction (2011) 142 87–97
BAX expression between the IVMC/IVCK group andthe control group. Taken together, differential BAXexpression levels in the blastocyst embryos between theSHH-treated groups and the control group were obvious.
In the case of the pluripotency-related genes, thelevels of CDX2 and POU5F1 expressions were similar(PO0.05) between the control and the treatment groups(Fig. 6A and B). However, the levels of ZPF42 expressionwere significantly higher in the IVMC/IVCK (1.13-fold),IVMK/IVCC(1.61-fold), or IVMC/IVCC(1.58-fold) treat-ment groups compared with the untreated control group(P!0.05). However, no difference was found betweenthe SHH-treated embryos in the IVMK/IVCC group andthe IVMC/IVCC group (Fig. 6C, PO0.05).
www.reproduction-online.org
IVM–/IVC–
2C
4C
BL
IVM+/IVC– IVM+/IVC+IVM–/IVC+
SHH
Figure 3 Histone 3 acetylation (AcH3K9/K14) levels ofporcine embryos at different developmental stagesunder various regimes of SHH treatment. 2C, two-cellembryo; 4C, four-cell embryo; BL, blastocyst. Scalebar, 50 mm. K/K, absence of SHH in IVM and IVCmedia (control); C/K, presence of 0.5 mg/ml SHHin IVM medium but absence of SHH in IVC medium;K/C, absence of SHH in IVM medium but presenceof 0.5 mg/ml SHH in IVC medium; C/C: presence of0.5 mg/ml SHH in both IVM and IVC media.
SHH enhances development of IVF porcine embryos 91
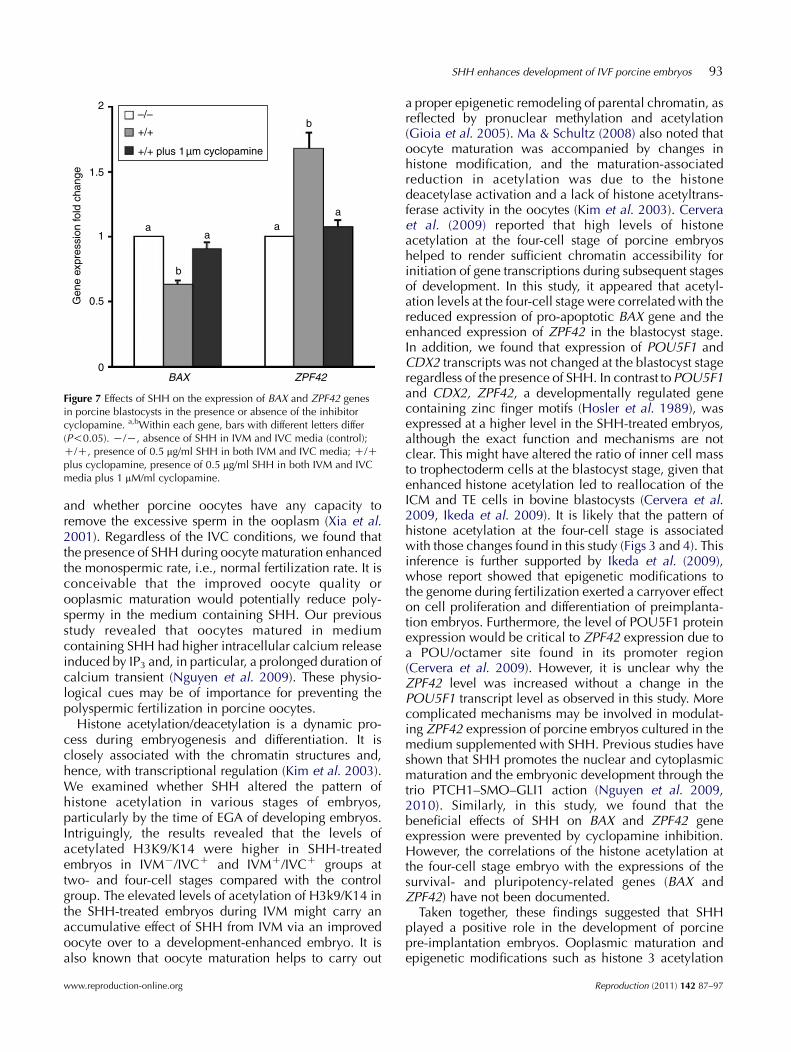
Similarly, with the presence of SHH in both IVM andIVC media, expression levels of BAX and ZPF42 weresignificantly enhanced. In contrast, the beneficial effectof SHH on BAX and ZPF42 expression levels wasneutralized by the presence of cyclopamine in themedium (Fig. 7).
450 –/–+/––/++/+
400
350
300
250
Rel
ativ
e flu
ores
cenc
e (p
ixel
s ×
104 )
200
150
100
2-cell
N=
35
N=
30N
=28
N=
30
N=
28N
=31
N=
32
N=
30
N=
25
N=
30
N=
35N
=40
4-cell Blastocyst
50
0
a
a
a
a aa
bb
bb
ca
Figure 4 Histone acetylation (AcH3K9/K14) profile of fertilized porcineembryos after treatment with SHH during IVM and/or IVC. a–cWithinthe embryonic stage, bars with different letters differ (P!0.05). Thesample sizes are shown in each bar. K/K, absence of SHH in IVM andIVC media (control); C/K, presence of 0.5 mg/ml SHH in IVM mediumbut absence of SHH in IVC medium; K/C, absence of SHH in IVMmedium, but presence of 0.5 mg/ml in IVC medium; C/C, presenceof 0.5 mg/ml SHH in both IVM and IVC media.
Discussion
Several lines of evidence indicated the involvement ofHh signaling during oogenesis. It was reported that Hhsignaling molecules were present in mouse (Wijgerdeet al. 2005, Russell et al. 2007, Ren et al. 2009) andbovine ovaries (Spicer et al. 2009), particularly in thetheca and granulosa cells. Ren et al. (2009) reported thata major activation of co-receptor SMO resulted in analteration of theca cell proliferation and prevention ofovulation in mice, although follicles developed normallyto preovulatory stage in response to LH stimulation. Inaddition, we previously reported that SHH and its relatedsignaling molecules (PTCH1, SMO, and GLI1) wereexpressed in porcine ovaries, specifically in the cumulus–oocyte complexes (COCs) and oocytes (Nguyen et al.2009). In contrast, Wijgerde et al. (2005) reported that noSMO was detected in mouse oocytes. Interestingly, wedetected SMO expression in denuded porcine oocytesby western blot (Nguyen et al. 2009), suggesting thatSMO expression could be species-specific. Furthermore,we have recently reported that Hh signaling receptorPTCH1 and SMO are also expressed in porcineparthenotes from the two-cell stage throughout theblastocyst stage at both mRNA and protein levels(Nguyen et al. 2010). In this study, we also found theexpressions of PTCH1 and SMO from two-cell toblastocyst stages of fertilized porcine embryos detectedby immunocytochemistry.
Since the embryonic genome activation (EGA) occursat the four-cell stage of porcine embryos (Jarrell et al.1991), the presence of PTCH1 and SMO at the two-cell
www.reproduction-online.org
stage is most likely a maternal legacy. Our previous studydocumented the existence of SHH protein in porcinefollicular fluids from follicles of various sizes (Nguyenet al. 2009). Since Hh signaling was detected in theoocytes and embryos of different developmental stages,we hypothesize that Hh signaling is involved infolliculogenesis as well as in early embryogenesis.In fact, our previous study had demonstratedthat addition of SHH to the IVM medium promotednuclear and cytoplasmic maturation of porcine oocytes,and subsequently enhanced parthenogenetic embryodevelopment (Nguyen et al. 2009). Consistently, wealso found that SHH addition during IVM increased
Reproduction (2011) 142 87–97
2 –/–+/––/++/+
a
b
c c
1.5
Gen
e ex
pres
sion
fold
cha
nge
1
0.5
0BCL2L1 BAX
Figure 5 Expression of cell survival-related genes BCL2L1 and BAX inporcine blastocysts cultured in SHH-supplemented IVM and/or IVCmedia. a–cWithin category, bars with different letters differ (P!0.05).K/K, absence of SHH in IVM and IVC media (control); C/K, presenceof 0.5 mg/ml SHH in IVM medium but absence of SHH in IVC medium;K/C, absence of SHH in IVM medium, but presence of 0.5 mg/ml SHHin IVC medium; C/C, presence of 0.5 mg/ml SHH in both IVM andIVC media.
–/– +/– –/+ +/+
–/– +/– –/+ +/+
POU5F1 (OCT-4)
CDX2
2
A
B
1.5
Gen
e ex
pres
sion
fold
cha
nge
1
0.5
0
2
1.5
Gen
e ex
pres
sion
fold
cha
nge
1
0.5
0
ZPF42
–/– +/– –/+ +/+
a
b
c c2
1.5
Gen
e ex
pres
sion
fold
cha
nge
1
0.5
0
C
Figure 6 Expression of pluripotency-related genes POU5F1 (A), CDX2(B), and ZPF42 (C) in porcine blastocysts cultured under variousregimes of SHH supplementation. a–cWithin each gene, bars withdifferent letters differ (P!0.05). K/K, absence of SHH in IVM and IVCmedia (control); C/K, presence of 0.5 mg/ml SHH in IVM medium butabsence of SHH in IVC medium; K/C, absence of SHH in IVMmedium, but presence of 0.5 mg/ml SHH in IVC medium; C/C,presence of 0.5 mg/ml SHH in both IVM and IVC media.
92 N T Nguyen and others
the blastocyst rate and total cell number per blastocystin this study.
The potential role of SHH during oogenesis is evidentin several species including pigs. During in vivomaturation, oocytes are exposed to SHH in the follicularfluid, because SHH was present in porcine follicularfluid. This was supported by our previous studies withsolely parthenogenetic embryos (Nguyen et al. 2009,2010). Interestingly, the yield and quality of IVF-derivedembryos were also further enhanced by the inclusion ofSHH in the IVC medium in addition to the IVM medium,suggesting that an additive or accumulative effect hasbeen exerted by SHH during these courses of develop-ment in porcine embryos.In vitro-produced porcine embryos were particularly
vulnerable to free radicals or subsequent oxidativestresses, due to their high lipid contents (McEvoy et al.2000). This phenomenon may have led to poor embryoviability and, in turn, to progressive embryo losses duringIVP, as evidenced by high incidence of apoptotic ratesin response to suboptimal culture conditions (Nanassyet al. 2008). In this study, we demonstrated that thepresence of SHH in culture medium effectively reducedapoptosis, although the underlying molecularmechanisms remained unclear. Presumably, SHHmight act as a survival factor, perhaps by altering theexpression of some cell survival-related proteins such asAKT (Nguyen et al. 2010). Other reports indicated thatPTCH1, a negative regulator of SHH signaling, maytrigger an apoptotic pathway in the absence of SHH(Thibert et al. 2003, Mille et al. 2009). Similar to thefindings in parthenogenetic embryos, SHH also reducedapoptotic indices, improved blastocyst rates, andincreased cell numbers of fertilized embryos in this
Reproduction (2011) 142 87–97
study. The presence of SHH in IVM and/or IVC media hasno influences on the expression of the anti-apoptoticgene BCL2L1, but the expression of pro-apoptotic geneBAX was reduced with the presence of SHH either in IVCmedium alone or in both IVM and IVC media.Collectively, our data supported the notion that SHHmodulated or prevented apoptosis of blastomeres,which, in turn, improved the blastocyst rate and totalcell numbers of porcine embryos.
It is still unknown whether a certain degree ofpolyspermy is physiologically permissive in porcine,
www.reproduction-online.org
–/–
+/+ plus 1µm cyclopamine
+/+
2
1.5
Gen
e ex
pres
sion
fold
cha
nge
1a
aa
a
b
b
BAX ZPF42
0.5
0
Figure 7 Effects of SHH on the expression of BAX and ZPF42 genesin porcine blastocysts in the presence or absence of the inhibitorcyclopamine. a,bWithin each gene, bars with different letters differ(P!0.05). K/K, absence of SHH in IVM and IVC media (control);C/C, presence of 0.5 mg/ml SHH in both IVM and IVC media; C/Cplus cyclopamine, presence of 0.5 mg/ml SHH in both IVM and IVCmedia plus 1 mM/ml cyclopamine.
SHH enhances development of IVF porcine embryos 93
and whether porcine oocytes have any capacity toremove the excessive sperm in the ooplasm (Xia et al.2001). Regardless of the IVC conditions, we found thatthe presence of SHH during oocyte maturation enhancedthe monospermic rate, i.e., normal fertilization rate. It isconceivable that the improved oocyte quality orooplasmic maturation would potentially reduce poly-spermy in the medium containing SHH. Our previousstudy revealed that oocytes matured in mediumcontaining SHH had higher intracellular calcium releaseinduced by IP3 and, in particular, a prolonged duration ofcalcium transient (Nguyen et al. 2009). These physio-logical cues may be of importance for preventing thepolyspermic fertilization in porcine oocytes.
Histone acetylation/deacetylation is a dynamic pro-cess during embryogenesis and differentiation. It isclosely associated with the chromatin structures and,hence, with transcriptional regulation (Kim et al. 2003).We examined whether SHH altered the pattern ofhistone acetylation in various stages of embryos,particularly by the time of EGA of developing embryos.Intriguingly, the results revealed that the levels ofacetylated H3K9/K14 were higher in SHH-treatedembryos in IVMK/IVCC and IVMC/IVCC groups attwo- and four-cell stages compared with the controlgroup. The elevated levels of acetylation of H3k9/K14 inthe SHH-treated embryos during IVM might carry anaccumulative effect of SHH from IVM via an improvedoocyte over to a development-enhanced embryo. It isalso known that oocyte maturation helps to carry out
www.reproduction-online.org
a proper epigenetic remodeling of parental chromatin, asreflected by pronuclear methylation and acetylation(Gioia et al. 2005). Ma & Schultz (2008) also noted thatoocyte maturation was accompanied by changes inhistone modification, and the maturation-associatedreduction in acetylation was due to the histonedeacetylase activation and a lack of histone acetyltrans-ferase activity in the oocytes (Kim et al. 2003). Cerveraet al. (2009) reported that high levels of histoneacetylation at the four-cell stage of porcine embryoshelped to render sufficient chromatin accessibility forinitiation of gene transcriptions during subsequent stagesof development. In this study, it appeared that acetyl-ation levels at the four-cell stage were correlated with thereduced expression of pro-apoptotic BAX gene and theenhanced expression of ZPF42 in the blastocyst stage.In addition, we found that expression of POU5F1 andCDX2 transcripts was not changed at the blastocyst stageregardless of the presence of SHH. In contrast toPOU5F1and CDX2, ZPF42, a developmentally regulated genecontaining zinc finger motifs (Hosler et al. 1989), wasexpressed at a higher level in the SHH-treated embryos,although the exact function and mechanisms are notclear. This might have altered the ratio of inner cell massto trophectoderm cells at the blastocyst stage, given thatenhanced histone acetylation led to reallocation of theICM and TE cells in bovine blastocysts (Cervera et al.2009, Ikeda et al. 2009). It is likely that the pattern ofhistone acetylation at the four-cell stage is associatedwith those changes found in this study (Figs 3 and 4). Thisinference is further supported by Ikeda et al. (2009),whose report showed that epigenetic modifications tothe genome during fertilization exerted a carryover effecton cell proliferation and differentiation of preimplanta-tion embryos. Furthermore, the level of POU5F1 proteinexpression would be critical to ZPF42 expression due toa POU/octamer site found in its promoter region(Cervera et al. 2009). However, it is unclear why theZPF42 level was increased without a change in thePOU5F1 transcript level as observed in this study. Morecomplicated mechanisms may be involved in modulat-ing ZPF42 expression of porcine embryos cultured in themedium supplemented with SHH. Previous studies haveshown that SHH promotes the nuclear and cytoplasmicmaturation and the embryonic development through thetrio PTCH1–SMO–GLI1 action (Nguyen et al. 2009,2010). Similarly, in this study, we found that thebeneficial effects of SHH on BAX and ZPF42 geneexpression were prevented by cyclopamine inhibition.However, the correlations of the histone acetylation atthe four-cell stage embryo with the expressions of thesurvival- and pluripotency-related genes (BAX andZPF42) have not been documented.
Taken together, these findings suggested that SHHplayed a positive role in the development of porcinepre-implantation embryos. Ooplasmic maturation andepigenetic modifications such as histone 3 acetylation
Reproduction (2011) 142 87–97

94 N T Nguyen and others
of the genome pre- and post-fertilization could bepersistent and carried over to the subsequent embryonicdevelopment, in particular, when both IVM and IVCmedia were supplemented with SHH to direct cellsurvival, proliferation, and differentiation during thepreimplantation development of porcine embryos.
Materials and Methods
Chemicals
All chemicals used in this study were purchased from SigmaChemicals, unless otherwise specified.
Oocyte maturation and embryo culture
Procedures for collection of ovaries and oocytes weredescribed previously (Nguyen et al. 2009, 2010). In brief,freshly harvested COCs were rinsed several times and culturedfor 22 h in the NCSU-23 medium supplemented with 10%porcine follicular fluid, cysteine (0.1 mg/ml), eCG (10 IU/ml),and hCG (10 IU/ml). The COCs were further cultured for anadditional 22 h in the same medium without hormones.
Sperm preparation, IVF, and embryo culture
Diluted semen from two boars of proven fertility was suppliedby a porcine artificial insemination center (Taichung, Taiwan),stored at 15 8C, and used for 2 days. Motile sperm wereobtained by centrifugation at 700 g with Percoll (P1644)discontinuous gradient (2 ml of 45% over 2 ml of 90%) for15 min. Spermatozoa were collected from the bottom of the90% fraction and washed in Dulbecco’s PBS (DPBS) supple-mented with 10% fetal bovine serum, followed by centri-fugation at 100 g for 5 min. After centrifugation, the spermpellet was resuspended in modified tris-buffered medium(mTBM) containing 113.1 mM NaCl, 3 mM KCl, 7.5 mMCaCl2.2H2O, 20 mM Tris, 11 mM glucose, 5 mM sodiumpyruvate, and 0.2% (w/v) BSA. After 44 h of maturation,oocytes were freed from cumulus cells, and matured oocyteswith first polar body were washed three times in mTBM thathad been pre-equilibrated for at least 12 h at 39 8C under 5%CO2. After washing, groups of 30 oocytes were randomlyplaced into 45 ml droplets of mTBM covered with pre-warmedmineral oil. After dilution (10!), 5 ml of sperm suspensionwere added to a 45 ml droplet of fertilization medium (mTBM)to give a final sperm concentration of 5!105 sperm/ml. Thegametes were co-cultured for 6 h at 39 8C in an incubatorcontaining 5% CO2 in humidified air. After 6 h, oocytes wereremoved from the fertilization medium and washed three timesin porcine zygote medium-3 (PZM-3) to remove excessivesperm. The presumptive zygotes were then cultured in 100 mldroplets (20–30 oocytes per droplet) of PZM-3 covered withmineral oil for 7 days. Cleavage and blastocyst rates wereevaluated under a stereomicroscope at days 2 and 7 afterinsemination (insemination dayZday 1).
An additional experiment was designed to evaluate thefertilization rate. At 10 h post-insemination, some oocytes wereselected and fixed in the fixative. The fixed oocytes were
Reproduction (2011) 142 87–97
subjected to triple staining and observed under a fluorescencemicroscope for nuclear evaluation, and were classified intothree categories, i.e. unfertilized oocytes, monospermicoocytes (two pronuclei, 2PN), and polyspermic oocytes(more than 2PN or 2PN plus swollen sperm heads). Therepresentative images are presented in Fig. 2.
Detection of histone acetylation and quantification
The effect of SHH on the acetylation of histone 3 at lysine 9 and14 (AcH3K9/K14) during pre-implantation development wasstudied. Embryos at different developmental stages were fixedwith 4% paraformaldehyde (PFA; EMS, Hatfield, Pennsylvania)in DPBS containing 0.1% polyvinyl alcohol (PVA) for 1 h at RT.They were then washed in washing solution (DPBS containing0.1% (w/v) PVA, 1% (w/v) BSA, and 0.1% (v/v) Tween-20) for1 h, and were permeated with 0.5% (v/v) Triton X-100 in DPBSfor 30 min at room temperature and blocked in workingsolution (DPBS–PVA supplemented with 2% BSA) at 4 8Covernight. The embryos were then incubated with anti-acetylhistone 3 (1:300, Cat #9677, Cell Signaling Technology, Inc.Beverly, MA, USA) for 1 h at 37 8C, washed in washing solutionfor 30 min, and then incubated with Alexa Flour 488-conjugated secondary antibody (1:200, Invitrogen) for 1 h atroom temperature (darkness), followed by two washes in thewashing solution. The samples were mounted on slides withmounting medium containing Hoechst. Immunofluorescentlabeling of AcH3K9/K14 was evaluated under an epifluores-cent microscope (Nikon, Tokyo, Japan), and micrographimages were captured. Fluorescence intensity was measuredby analysing the micrographs with ImageJ Software (ImageProcessing and Analysis in Java, Version 1.42 for Windows,NIH, Bethesda, MD, USA) after background subtraction. Dataare presented as a mean of relative fluorescence intensity (pixels).
Immunocytochemical staining
Patched (PTCH1) and SMO proteins
To detect PTCH1 and SMO, embryos at two-cell, four- to eight-cell, morula, and blastocyst stages were fixed in 4% (w/v) PFAin DPBS supplemented with 0.1% (w/v) PVA for 1 h at roomtemperature. After washing twice in washing solution, embryoswere permeated with 0.5% (v/v) Triton X-100 in DPBS for30 min. The embryos were then blocked in working solutionsupplemented with 5% (v/v) chicken serum at 4 8C overnight.After being washed twice in washing solution, the embryoswere incubated with primary antibody for PTCH1 (1:330;sc-13943; Santa Cruz Biotechnology, Inc., Santa Cruz, CA,USA) or SMO (1:330; sc-9016; Santa Cruz) at 4 8C overnight.They were then washed for 30 min and incubated with AlexaFlour 488-conjugated secondary antibody (1:200, Invitrogen)for 1 h (darkness) and then washed twice with washingsolution. Samples were mounted on slides with mountingmedium containing Hoechst, and images were captured underan epifluorescence microscope (Nikon).
Nuclear and cytoskeleton: triple staining
Triple staining procedure was performed as described by Juet al. (2003). In brief, microtubule stabilization buffer (MTSB,
www.reproduction-online.org

SHH enhances development of IVF porcine embryos 95
5!) and working solution (MTSB-XF) containing 0.5 M PIPES,25 mM magnesium chloride, 125 mM EGTA, 0.01% (w/v)aprotinin, 1 mM dithiothreitol, 50% deuterium oxide, 1 mMpaclitaxel, 0.1% (v/v) Triton X-100, and 2% (v/v) formaldehyde.Mounting medium contained 10 mg/ml Hoechst 33342, 50%(v/v) glycerol, and 25 mg/ml sodium azide in DPBS. Oocyteswere transferred into the fixative in an Eppendorf tube and wereincubated in water bath (38 8C) for 1 h, and stored at 4 8C for atleast 1 week before staining. For staining, the fixed oocyteswere incubated with anti-a- and anti-b-tubulin primaryantibodies at 4 8C overnight. The oocytes were then washedthree times with washing solution containing 2% (w/v) BSA,2% goat serum, 0.2% (w/v) milk powder, 0.2% (w/v) sodiumazide, and 0.1% (v/v) Triton X-100 in DPBS. The oocytes weresubsequently incubated with the FITC-conjugated secondaryantibody at 4 8C overnight. After being washed three times,microfilaments of the oocytes were stained with rhodamine–phalloidin (R147; Molecular Probes, Invitrogen) for 1 h forlabeling the filamentous actin and then washed three timesagain. Finally, the oocytes were mounted on the slide withmounting medium containing Hoechst 33342 and then sealedwith fingernail polish, and examined under an epifluorescencemicroscope (Nikon) with magnification at 40!.
RNA purification, reverse transcription and quantitativereal-time PCR
Total RNA was extracted by TRIzol Reagent (Invitrogen) andpurified by RNeasy Mini Kit (Qiagen). Purified RNA wasquantified using a ND-1000 spectrophotometer (NanodropTechnology, Wilmington, DE, USA) and qualitatively analyzedby Bioanalyzer 2100 (Agilent Technology, USA). To prepare acDNA pool from each RNA sample, total RNA (19 ng) wasreverse transcribed using MMLV reverse transcriptase (Pro-mega), and the resulting samples were diluted 2!103 timeswith DNase-free water. Each cDNA pool was stored at K20 8Cuntil further real-time PCR analysis.
Real-time PCR were performed on the Roche LightCyclerInstrument 1.5 using LightCycler FastStart DNA MasterPLUSSYBR Green I kit (Roche Cat. 03 515 885 001). Briefly, the 10 mlreaction mixture contained 2 ml master mix, 2 ml of 0.75 mM or2 ml of 0.50 mM forward primer and reverse primer with 5%
Table 3 Primer sequences, GenBank accession numbers, and expected pro
Target genes Primers Sequences (5 0 to 3 0)
BCL2L1 Forward GCAGGTATTGAACGReverse GCATCTCCTTGTCTA
BAX Forward CTACCAAGAAGTTGReverse ACGGCTGCGATCAT
CDX2 Forward CCTCTCGCCCACAAReverse TCCAACCGCACCTG
ZPF42 Forward GGTCTCAGCCTCAGReverse TGTAGCCAGCGTCC
POU5F1 Forward CGCAACGAGAGGAReverse CGCCAGAGGAAAG
ACTB Forward CCACGCCATCCTGCReverse CCATCTCCTGCTCGA
www.reproduction-online.org
DMSO respectively, and 6 ml cDNA sample solutions. Eachsample was performed in triplicate. The RT-PCR wereprogrammed at 95 8C for 10 min, 50 cycles of 95 8C for 10 s,60 8C for 15 s, and 72 8C for 10 s. At the end of the program,a melt curve analysis was performed. At the end of eachRT-PCR run, data were automatically analyzed by the system,and an amplification plot was generated for each cDNAsample. From each of these plots, the LightCycler3 DataAnalysis Software (Roche Diagnostics) automatically calcu-lated CP value (crossing point, the turning point correspondsto the first maximum of the second derivative curve), whichimplied the beginning of an exponential amplification. Thecomparative CT method was applied for quantification of geneexpression levels as described by Wee et al. (2006). Geneexpression fold changes were calculated using the formula2KðDDCT ExpKControlÞ, where
DCT Exp ZCT targetKCT ref of Exp sample
DCT Control ZCT targetKCT ref of control sample
Primer sequences, GenBank accession numbers, andexpected product sizes for the real-time PCR analysis arepresented in Table 3.
Specific experiments
Experiment 1: expression of Hh signaling molecules at differentdevelopmental stages of IVF-derived porcine embryos
The expressions of PTCH1 and SMO in IVF-derived two-cell,four- to eight-cell embryos, morulae and blastocysts wereinvestigated by immunocytochemical staining. At least thirtyembryos in each stage were used for detection.
Experiment 2: the collective effects of SHH on embryodevelopment accumulated during IVM and IVC
The optimized dose usage of SHH at 0.5 mg/ml was determinedfrom our previous studies (Nguyen et al. 2009, 2010) and usedthroughout all treatments in this study. Matured oocytes derivedfrom IVM with or without SHH treatment were subjected toIVF. After 6 h co-culture with sperm, the presumptive zygoteswere cultured in PZM-3 and randomly allocated into thefollowing culture groups: no SHH in both IVM and IVC
duct sizes for the qRT-PCR analysis.
GenBankaccession number
Productsize (bp)
AACTCTTCCG AJ_001203 102CGCTCTCCAGCGAGTGTC AJ_606301 85CCTCTGATGTTCAC TC_206866 82TCTTTACCTGTCAGC TC_317123 105TCTTCCTTTTGAG TC_168415 68GATACTGGTCT AK_237086 168AGTCCAG
Reproduction (2011) 142 87–97

96 N T Nguyen and others
(control, IVMK/IVCK), IVM with SHH and IVC withoutSHH (IVMC/IVCK), IVM without SHH and IVC with SHH(IVMK/IVCC), and SHH in both IVM and IVC (IVMC/IVCC).The cleavage and blastocyst rates were recorded at days 2and 7 respectively. The blastocysts were subjected toHoechst staining and TUNEL labeling for the determinationof the total cell number and apoptotic cells respectively(Nguyen et al. 2010). Apoptotic indices were calculated asfollows:
Apoptotic index (%)Z(number of positive nuclei/total cellnumber)!100.
At 10 h post-fertilization, a proportion of presumptivezygotes from each treatment were randomly selected forevaluation of fertilization rate.
Experiment 3: effects of SHH treatment in IVM and IVC mediaon acetylation profile of IVF embryos
Embryos from each treatmentas inExperiment 1werecollectedattwo-cell, four-cell, and blastocyst stages for histone acetylationassay. At least 25 embryos at each stage were used, and therepresentative numbers were given as shown in the Fig. 4.
Experiment 4: effects of SHH treatment in IVM and IVCon gene expressions of IVF embryos
Embryos collected and pooled at day 7 from each treatment(ten embryos per treatment) were incubated with cell lysisbuffer at 42 8C for 1 h, and then stored at K80 8C. Cell survival-related genes (BCL2L1 and BAX), pluripotency marker genePOU5F1, trophectoderm lineage marker CDX2, and inner cellmass lineage marker ZPF42 were analyzed by quantitative real-time PCR.
An additional experiment was designed to confirm thebeneficial effect of SHH on BAX and ZPF42 expression. Afterinsemination, matured oocytes under the presence of 0.5 mg/mlSHH were divided into two groups, in which one group wascultured in the IVC medium supplemented with 0.5 mg/mlSHH, and the other cultured in the medium with 0.5 mg/mlSHH plus 1 mM cyclopamine, an SHH inhibitor. The maturedoocytes cultured without SHH in both IVM and IVC mediawere the control group. The blastocysts harvested at day 7 asmentioned earlier were used to determine the expression levelsof BAX and ZPF42 genes.
Statistical analysis
All data were subjected to ANOVA using the general linearmodel procedure in SAS version 9 (SAS Institute, Cary, NC,USA), followed by Tukey’s test. Percentile data were arcsinetransformed before statistical analysis, and the P!0.05 wasconsidered significantly different between treatment groups.
Declaration of interest
The authors declare that there is no conflict of interest thatcould be perceived as prejudicing the impartiality of theresearch reported.
Reproduction (2011) 142 87–97
Funding
This study was partially supported by grants fromNational Science Council, Executive Yuan, Taiwan, ROC(NSC 98-2628-B005-019-MY3), and was performed in theTissue Engineering and Stem Cells Center, National ChungHsing University, Taichung, Taiwan.
Acknowledgements
The authors would like to thank Dr David Pei-Cheng Lin atChung-Shan Medical University (Taichung-Taiwan) for thecritical reading of this manuscript. We are also grateful toTaichung Meat Market Co. Ltd, Taichung, Taiwan, for kindlyproviding porcine ovaries for years.
References
Abeydeera LR, WangWH, Cantley TC, Rieke A, Prather RS & Day BN 1998Presence of epidermal growth factor during in vitro maturation of pigoocytes and embryo culture can modulate blastocyst development afterin vitro fertilization. Molecular Reproduction and Development 51395–401. (doi:10.1002/(SICI)1098-2795(199812)51:4!395::AID-MRD6O3.0.CO;2-Y)
Beebe LFS, Vassilev I, Mcilfatrick S & Nottle B 2009 Adding essential aminoacids at a low concentration improves the development of in vitrofertilized porcine embryos. Journal of Reproduction and Development55 373–377. (doi:10.1262/jrd.20176)
Cervera RP, Marti-Gutierrez N, Escorihuela E, Moreno R & Stojkovic M2009 Trichostatin A affects Histone acetylation and gene expression inporcine somatic cell nucleus transfer embryos. Theriogenology 721097–1110. (doi:10.1016/j.theriogenology.2009.06.030)
Craig J, Zhu H, Dyce PW, Petrik J & Li J 2004 Leptin enhances oocytesnuclear and cytoplasmic maturation via mitogen-activated proteinkinase. Endocrinology 145 5355–5363. (doi:10.1210/en.2004-0783)
Franco HL, Lee KY, Russell RB, White LD, Lanske B, Lydon JP, Jeong JW &DeMayo FJ 2010a Ablation of Indian hedgehog in the murineuterus results in decreased cell cycle progression, aberrant epidermalgrowth factor signaling, and increased estrogen signaling. Biology ofReproduction 82 783–790. (doi:10.1095/biolreprod.109.080259)
Franco HL, Lee KY, Rubel CA, Creighton CJ, White LD, Russell RB,Lewis MT, Lydon JP, Jeong JW & DeMayo FJ 2010b Constitutiveactivation of smoothened leads to female infertility and altered uterinedifferentiation in the mouse. Biology of Reproduction 82 991–999.(doi:10.1095/biolreprod.109.081513)
Gioia L, Barboni B, Turriani M, Capacchietti G, Pistilli MG, Berardinelli P& Mattioli M 2005 The capability of reprogramming the male chromatinafter fertilization is dependent on the quality of oocyte maturation.Reproduction 130 29–39. (doi:10.1530/rep.1.00550)
Gupta MK, Uhm SJ, Han DW & Lee HT 2007 Embryo quality andproduction efficiency of porcine parthenotes is improved by phytohe-magglutinin. Molecular Reproduction and Development 74 435–444.(doi:10.1002/mrd.20547)
Hosler BA, LaRosa GJ, Grippo JF & Gudas LJ 1989 Expression of REX-1, agene containing zinc finger motifs, is rapidly reduced by retinoic acid inF9 teratocarcinoma cells. Molecular and Cellular Biology 9 5623–5629.
Ikeda S, Tatemizo A, Iwamoto D, Taniguchi S, Hoshino Y, Amano T,Matsumoto K, Hosoi Y, Iritani A & Saeki K 2009 Enhancement of histoneacetylation by trichostatin A during in vitro fertilization of bovine oocytesaffects cell number of the inner cell mass of the resulting blastocysts.Zygote 17 209–215. (doi:10.1017/S0967199409005279)
Jarrell VL, Day BN & Prather RS 1991 The transition from maternal tozygotic control of development occurs during the 4-cell stage in thedomestic pig Sus scrofa: quantitative and qualitative aspects of proteinsynthesis. Biology of Reproduction 44 62–68. (doi:10.1095/biolre-prod44.1.62)
www.reproduction-online.org

SHH enhances development of IVF porcine embryos 97
Ju JC, Tsay C & Ruan CW 2003 Alterations and reversibility in thechromatin, cytoskeleton and development of pig oocytes treated withroscovitine. Molecular Reproduction and Development 64 482–491.(doi:10.1002/mrd.10234)
Kim JM, Liu H, Tazaki M, Nagata M & Aoki F 2003 Changes in histoneacetylation during mouse oocyte meiosis. Journal of Cell Biology 16237–46. (doi:10.1083/jcb.200303047)
Kim S, Lee SH, Kim JH, Jeong YW, Hashem MA, Koo OJ, Park SM, Lee EG,Hossein MS, Kang SK et al. 2006 Anti-apoptotic effect of insulin-likegrowth factor (IGF)-I and its receptor in porcine preimplantation embryosderived from in vitro fertilization and somatic cell nuclear transfer.Molecular Reproduction and Development 73 1523–1530. (doi:10.1002/mrd.20531)
Krisher R 2004 The effect of oocyte quality on development. Journal ofAnimal Science 82 E14–E23.
Lee K, Jeong J, Kwak I, Yu CT, Lanske B, Soegiarto DW, Toftgard R, Tsai MJ,Tsai S, Lydon JP et al. 2006 Indian hedgehog is a major mediatorof progesterone signaling in the mouse uterus. Nature Genetics 381204–1209. (doi:10.1038/ng1874)
Lonergan P, Rizos D, Gutierrez-Adan A, Moreira PM, Pintado B, de laFuente J & Boland MP 2003 Temporal divergence in the pattern ofmessenger RNA expression in bovine embryos cultured from the zygoteto blastocyst stage in vitro or in vivo. Biology of Reproduction 691424–1431. (doi:10.1095/biolreprod.103.018168)
Lonergan P, Fair T, Corcoran D & Evans ACO 2006 Effect of cultureenvironment on gene expression and developmental characteristicsin IVF-derived embryos. Theriogenology 65 137–152. (doi:10.1016/j.theriogenology.2005.09.028)
Ma PP & Schultz RM 2008 Histone deactylase 1 (HDAC1) regulates histoneacetylation, development, and gene expression in preimplantationmouse embryos. Developmental Biology 319 110–120. (doi:10.1016/j.ydbio.2008.04.011)
Matsumoto H, Zhao X, Das SK, Hogan BL & Dey SK 2002 Indian hedgehogas a progesterone-responsive factor mediating epithelial–mesenchymalinteractions in the mouse uterus. Developmental Biology 245 280–290.(doi:10.1006/dbio.2002.0645)
McEvoy TG, Coull GD, Broadbent PJ, Hatchbinson JS & Speake BK 2000Fatty acid composition of lipids in immature cattle, pig and sheep oocyteswith intact zona pellucida. Journal of Reproduction and Fertility 118163–170. (doi:10.1530/jrf.0.1180163)
Mille F, Thibert C, Fombonne J, Rama N, Guix C, Hayashi H, Corset V,Reed JC & Mehlen P 2009 The Patched dependence receptor triggersapoptosis through a DRAL–caspase-9 complex. Nature Cell Biology 11739–746. (doi:10.1038/ncb1880)
Mtango NR, Pottireddy S & Latham KE 2008 Oocyte quality and maternalcontrol of development. International Review of Cell and MolecularBiology 268 223–290. (doi:10.1016/S1937-6448(08)00807-1)
Nanassy L, Lee K, Javor A & Machaty Z 2008 Effects of activation methodsand culture conditions on development of parthenogenetic porcineembryos. Animal Reproduction Science 104 264–274. (doi:10.1016/j.anireprosci.2007.01.019)
Nguyen NT, Lin DPC, Yen SY, Tseng JK, Chuang JF, Hung HT, Lin TA,Chang HH & Ju JC 2009 Sonic hedgehog promotes porcine oocytematuration and early embryo development. Reproduction, Fertilityand Development 21 805–815. (doi:10.1071/RD08277)
Nguyen NT, Lin DPC, Siriboon C, Lo NW & Ju JC 2010 Sonic hedgehogimproves in vitro development of porcine parthenotes and handmadecloned embryos. Theriogenology 74 1149–1160. (doi:10.1016/j.ther-iogenology.2010.05.016)
Ock SA, Lee SL, Kim JG, Kumar BM, Balasubramanian S, Choe SY & Rho GJ2007 Development and quality of porcine embryos in different culturesystems and embryo-producing methods. Zygote 15 1–8. (doi:10.1017/S0967199406003911)
www.reproduction-online.org
Prather RS, Hawley RJ, Carter DB, Lai L & Greenstein JL 2003 Transgenicswine for medicine and agriculture. Theriogenology 19 115–123.(doi:10.1016/S0093-691X(02)01263-3)
Ren Y, Cowan RG, Harman RM & Quirk SM 2009 Dominant activation ofhedgehog signaling pathway in the ovary alters theca development andprevents ovulation. Molecular Endocrinology 23 711–723. (doi:10.1210/me.2008-0391)
Rizos D, Gutierrez-Adan A, Perez-Garnalo S, de la Fuente J, Boland MP &Lonergan P 2003 Bovine embryo culture in the presence or absence ofserum: implications for blastocyst development, cryotolerance, andmessenger RNA expression. Biology of Reproduction 68 236–243.(doi:10.1095/biolreprod.102.007799)
Russell MC, Cowan RG, Harman RM, Walker AL & Quirk SM 2007The hedgehog signaling pathway in the mouse ovary. Biology ofReproduction 77 226–236. (doi:10.1095/biolreprod.106.053629)
Sirard MA, Richard F, Blondin B & Robert C 2006 Contribution of theoocyte to embryo quality. Theriogenology 65 126–136. (doi:10.1016/j.theriogenology.2005.09.020)
Spicer LJ, Sudo S, Aad PY, Wang LS, Chun SY, Ben-Shlomo I, Klein C &Hsueh AJW 2009 The hedgehog-patched signaling pathway and functionin the mammalian ovary: a novel role for hedgehog proteins instimulating proliferation and steroidogenesis of theca cells. Reproduction138 329–339. (doi:10.1530/REP-08-0317)
Takamato N, Zhao BH, Tsai SY & Demayo FJ 2002 Identificationof Indian hedgehog as a progesterone-responsive gene in the murineuterus. Molecular Endocrinology 16 2338–2348. (doi:10.1210/me.2001-0154)
Thibert C, Teillet MA, Lapointe F, Mazelin L, Le Dounarin MN &Mehlen P 2003 Inhibition of neuroepithelial patched-induced apoptosisby sonic Hedgehog. Science 301 843–846. (doi:10.1126/science.1085405)
Thompson JG 2000 In vitro culture and embryo metabolism of cattleand sheep embryos-A decades of achievement. Animal ReproductionScience 60–61 263–275. (doi:10.1016/S0378-4320(00)00096-8)
Uhm SJ, Gupta MK, Yang JH, Lee SH & Lee HT 2007 Selenium improves thedevelopmental ability and reduces the apoptosis in porcine parthenotes.Molecular Reproduction and Development 74 1386–1394. (doi:10.1002/mrd.20701)
Van Langendonckt A, Donnay I, Schuurbiers N, Auquier P, Carolan C,Massip A & Dessy F 1997 Effects of supplementation with fetal calf serumon development of bovine embryos in synthetic oviduct fluid medium.Journal of Reproduction and Fertility 109 87–93. (doi:10.1530/jrf.0.1090087)
Wee G, Koo DB, Song BS, Kim JS, Kang MJ, Moon SJ, Kang YK, Lee KK &Han YM 2006 Inheritable histone H4 acetylation of somatic chromatinsin cloned embryos. Journal of Biological Chemistry 281 6048–6057.(doi:10.1074/jbc.M511340200)
Wijgerde M, Ooms M, Hoogerbrugge JW & Grootegoed JA 2005Hedgehog signaling in mouse ovary: Indian hedgehog and deserthedgehog from granulosa cells induce target gene expression indeveloping theca cells. Endocrinology 146 3558–3566. (doi:10.1210/en.2005-0311)
Xia P, Wang Z, Yang Z, Tan J & Qin P 2001 Untrastructural study ofpolyspermy during early embryo development in pigs, obverved byscanning electron microscope and transmission electron microscope.Cell Tissue Research 303 271–275. (doi:10.1007/s004410000315)
Zheng P & Dean J 2007 Oocyte-specific genes affect folliculogenesis,fertilization, and early development. Seminars in ReproductiveMedicine25 243–251. (doi:10.1055/s-2007-980218)
Received 21 February 2011
First decision 12 April 2011
Accepted 12 April 2011
Reproduction (2011) 142 87–97





























