Embed Size (px)
Citation preview

Hydrobiologia 491: 119–132, 2003.E. van Donk, M. Boersma & P. Spaak (eds), Recent Developments in Fundamental and Applied Plankton Research.© 2003 Kluwer Academic Publishers. Printed in the Netherlands.
119
Spatial and seasonal variation of the zooplankton in the coastal zone andmain khors of Lake Nasser (Egypt)
Gamal M. El-Shabrawy1 & Henri J. Dumont2,∗1National Institute of Oceanography and Fisheries, Inland Water Branch, 101 El-Kasr Alainy St., Cairo, EgyptE-mail: [email protected] of Animal Ecology, Ghent University, K. L. Ledeganckstraat, 35, B-9000 Ghent, BelgiumE-mail: [email protected] (∗Author for correspondence)
Received 6 July 2001; in revised form 25 March 2002; accepted 4 May 2002
Key words: Lake Nasser, zooplankton (Rotifera, Copepoda, Cladocera), khors, community structure,temporal variation
Abstract
A prominent feature of Lake Nasser is the presence of khors (dendritic side extensions). We studied the zooplanktonof the larger khors and coastal zone of the main lake in 1996 and 1997, and found an assemblage of rotifers, clado-cerans and copepods that was partly tropical, partly temperate, at relatively high biomasses. Spatial differenceswere weak, but the upstream khors (Toushka and Korosko) were consistently richer than the downstream khors(El-Ramla and Kalabsha), with a rather sudden transition around km 150 at El Madiq. Summer standing cropswere higher than those in winter by a factor 2–3. The zooplankton of the littoral of the main channel showedthe same spatial pattern as that in the khors, being more abundant in spring (average 82 300 ind m−3) than inautumn (average 72 700 ind m−3). Zooplankton dry weight increased from 4 g m−2 at khor El-Ramla to 7 g m−2
at khor Toushka. These rather high values had low variation. The number of species, diversity and evenness allshowed a high degree of similarity among the khors and in the littoral of the main lake. The lake fish fauna is poor,lacking a pelagic planktivore. The predominance of medium-sized Copepoda (one calanoid, two cyclopoids) in thezooplankton suggests that fish predation is moderate. This is confirmed by the persistence of two Daphnia speciesat low abundance, although rather strong variations in time suggest that Cladocera suffer from summer predationby invertebrates (copepods) as well as vertebrates (mostly larval fish). Because the zooplankton is underutilised byhigher trophic levels, we suggest to assess the benefits of introducing a pelagic zooplanktivorous fish.
Introduction
The construction of the Aswan High Dam in southernEgypt resulted in the creation of the longest man-madelake in the world. Filled in late 1970s, Lake Nasser(22◦ 31′ to 23◦ 45′ N and 31◦ 30′ to 33◦ 15′ E) extendsin the hot and dry climate of the eastern Sahara forabout 180 km in Sudan and 300 km in Egypt.
The reservoir is highly dendritic, with a num-ber of flooded side valleys, known as khors (Fig. 1).The mean length of the khors increases downstreamfrom the south to the north, owing to the northwardlydeclining ancient riverbed (Entz, 1973). All are U-
shaped in cross section. The dimensions of the largerones are shown in Table 1. Some cover a consider-able area and represent auxiliary semi-isolated lakes.Toushka, Allaqi, and Kalabsha are the largest khors(Fig. 1). They are wide, with a gentle slope and a sandybottom, while others like Korosko and El-Sadake [notshown in Fig. 1] are steep, relatively narrow and witha rocky bottom (Latif, 1984).
The fish fauna of the Nile below the cataracts thatseparate the Blue and White Niles from their sourcelakes amounts to ca 115 species. For unknown reasons,clupeids are totally absent (Greenwood, 1976). Ori-ginally, some 57 species were found in the reservoir,

120
Table 1. Dimensions of selected Khors of Lake Nasser
Name of khors Distance from Length Characteristics at 180 m level
High dam (km) Surface area Perimeter Volume (km3)
(km) (km2) (km)
Khor el Ramla 12 25.72 101.2 284 0.96
Khor Kalabsha 44 47.2 620 517 7.16
Khor Allaqi 100 54.83 490.8 510 11.57
Khor Korosko 180 22.56 83.6 353 1.76
Khor Toushka 242 15.02 66.9 117 1.44
Figure 1. Map of Lake Nasser-Nubia, identifying the khors studied,as well as the nine sampling locations along the main axis of thelake (1 = High Dam, 2 = el Ramla, 3 = Khalabsha, 4 = Allaqi, 5 =el Madiq, 6 = Korosko, 7 = Amada, 8 = Toushka, 9 = Abu Simbel).Distances from High Dam shown in km.
but this number later declined to ca 26 species (AbuGideiri & Ali, 1975; Latif, 1976; Ali, 1984). Sincethe early days of impoundment, landings from lakeNasser have contributed significantly to fish consump-tion in Egypt. The annual yield fluctuated between30 000 t in 1983 and 20 000 in 1997 (Agaypi, 1995;
Barrani, pers. com.), with Oreochromis niloticus andSarotherodon galilaeus constituting ca. 90% of thetotal catch, followed by the piscivores Lates niloticusand Hydrocynus forskali. The coastal area of the mainchannel of the lake and its khors plays a major role forthese fishes. In particular, tilapias use khors as favour-ite habitats, avoiding the limnetic zone, and feedingand reproducing in the littoral.
Rzoska (1976) and Dumont (1986a) reviewed thezooplankton of the Nile, and published species lists forthe river, Lake Nasser-Nubia, plus the lake sources ofthe Nile. These reveal that the species composition hasbeen remarkably stable over time, and does not deviatefrom that in the 19th century. However, few studieshave dealt with the ecology of the zooplankton, partic-ularly in the littoral and main khors of Lake Nasser.Available information includes that khors are twicericher in zooplankton than the main channel (Samaan,1971), the distribution and seasonal variation of zo-oplankton in khor El-Ramla (Abdel-Mageed, 1992),the species composition and distribution of zooplank-ton in khor Kalabsha in relation to environmentalconditions (Iskaros, 1993), and the standing crop anddistribution of zooplankton in khor El-Ramla (Habib,1996). Here, we deal with the spatial and temporaldynamics of the zooplankton, expressed as abundanceand as biomass, in the coastal zone and main khorsof Lake Nasser, with an aim at better understandingthe structure of the community. We also suggest waysof improving the utilisation of the zooplankton of thelake by fish.
Material and methods
Khors are important structural elements of the lake:their surface area covers about 4900 km2, i.e. 79% oftotal lake surface, while their volume is 86.4 km3, ie.

121
Table 2. Average diversity measures of the zooplankton of the khors of Lake Nasser
Khor el Ramla Khor Kalabsha Khor Allaqi Khor Korosko Khor Toushka
Number of species 20.000 20.000 23.000 23.000 19.000
Diversity Index 0.903 0.907 0.912 0.891 0.961
Evenness 0.694 0.697 0.670 0.655 0.751
Richness 1.702 1.715 1.990 1.961 1.601
55% of total lake volume (Entz, 1973, 1976). Thereare about 85 longer khors, 48 of which are on the eastside (576 km in shoreline length) and 37 on the westside (394 km in shore length) (Latif, 1976).
Abiotic variables
Water temperature was measured with a standardthermometer, pH using a Shibata pH meter model6H, transparency by Secchi disc, dissolved oxygenby Winkler Method, COD by oxidation of a knownvolume of unfiltered sample using KMnO4 (resultsexpressed as mg O2 l−1), and Chlorophyll-a by extrac-tion in 90% acetone and fluorometry using a Turner IIIfluorimeter.
Statistical analysis
We computed Shannon–Wiener diversity, species rich-ness, and evenness. We used canonical correspond-ence analysis (CCOA) to partition the importanceof environmental factors on zooplankton assemblagestructure in the khors (see Podani, 2000, for de-tails). We computed simple correlation coefficientsbetween selected taxa to explore the possibility ofecological interactions and explored the (dis)similarityof the zooplankton communities between khors usingclustering.
Zooplankton
Samples were collected at seasonal intervals in 1996,during four 10-day cruises, at one station in each ofthe five largest khors (Fig. 1). Additionally, nine sta-tions along the near-coastal area of the main channelwere sampled during autumn 1996 and spring 1997(numbered 1–9 in Fig. 1). Samples were collected byvertical tows, from a depth of 10 m to the surface(roughly corresponding to the euphotic zone), duringdaytime, using a plankton net (55 µm in mesh size,1-m long, 0.3 m in mouth opening diameter). Previouswork in the main channel (El-Shabrawy, 2000) had
shown that most plankton biomass resides in the eu-photic layer. Therefore, little could be gained by theadditional effort of sampling the entire water column,and the biomass in the top ten meters is here con-sidered to be representative of the biomass of thewhole water column. Samples were preserved imme-diately after collection in 4% neutral formalin. AllRotifera, Cladocera and Copepoda were examined,identified, and measured under a microscope. We useddry weight estimates from Dumont et al.’s (1975) bodylength–weight relationships.
Results
Physical and chemical variables, and chlorophyll
Water temperatureSurface water temperatures ranged between 17.5 ◦C atkhor Kalabsha in winter, to 32.2 ◦C at khor Allaqi insummer. This amplitude of variation is close to that ofthe Sahelian Lake Chad (18–30 ◦C) (Carmouze et al.,1983). In the open water of the lake, vertical mixingis complete in winter. A thermocline becomes estab-lished in summer, with a maximum vertical differenceof ca. 13 ◦C (Abd Allah, 1994).
TransparencyThe water of the downstream khors (El-Ramla andKalabsha) was more transparent than that upstream(Korosko and Toushka). Transparency was higher inwinter, with a maximum of 3.6 m at khor El-Ramla.The lowest transparency, 1.0 m, occurred at khorToushka and at Abu Simbel in summer. Assumingthe depth of the euphotic zone to equal 3–5 timesthe Secchi-depth, it follows that our vertical plank-ton hauls roughly represent a transect throughout thatzone.
Hydrogen ion concentrationpH was always on the alkaline side and varied from7.3 to 8.9.

122
Figure 2. Comparison of the percentage seasonal abundance and biomass of the rotifers, cladocerans and copepods of Lake Nasser (averagedover the whole lake).
Dissolved oxygen (DO)The khors were well oxygenated during winter andspring (average 11.1 and 13.5 mg l−1), with lowestaverage values in summer and autumn around 7 mgl−1. Dissolved oxygen showed little variation amongkhors (Anonymous, 1996). In the main lake, temper-ature stratification is accompanied by the establish-ment of an anoxic hypolimnion. The saturated surfacelayer declines in thickness from upstream (20 m) todownstream (8–10 m near the high dam) (Latif, 1984).
Chemical Oxygen Demand (COD)
COD fluctuated between 0.1 mg O2 l−1 at KhorKorosko in Winter and 4.8 mg O2 l−1 at Khor Kalab-sha in autumn (Anonymous, 1996).
Chlorophyll-a
Chl-a gradually increased from a minimum of2.8 mg m−3 in winter to a maximum of 9.1 mg m−3
in autumn. The most upstream khors had the highestvalues (Korosko 9.2 and Toushka 8.4 mg m−3).

123
Zooplankton species composition
We recorded 27 species of Copepoda, Cladocera, andRotifera. These are similar to the species assemblagedocumented from the Nile since the 19th century,suggesting qualitative stability in zooplankton com-position of the river system (see Dumont, 1986, fora full list of taxa, and historical references).
Copepoda
Main khors: Copepoda (all stages combined) werethe most abundant taxon in all seasons and khors,comprising 67% of total zooplankton, and 53% of bio-mass (Figs 2, 3a–c). They were represented by thecyclopoids Thermocyclops neglectus (G.O. Sars) andMesocyclops ogunnus Onabamiro, both carnivorous asadults, and by the herbivorous calanoid Thermodiap-tomus galebi (Barrois). We separated the copepodidstages into groups, but not the nauplii stages.
T. galebi was the dominant taxon, 6 and 45% oftotal and adult copepod densities, respectively, witha mean abundance of 2900 ind m−3 (range 500–8300ind m−3) (Fig. 3). In spite of these relatively low dens-ities, this large grazer formed a substantial biomass,42–64% of total and adult copepod biomass. T. galebiis typically limnetic and was found in relatively highdensities during spring. In contrast, its stock was lowin summer.
Thermocyclops neglectus contributed 5–35% todensities and 9–14% to biomass. Its highest densityoccurred in winter, with a peak of 5500 ind m−3 atKhor El-Ramla.
Mesocyclops ogunnus was aestival, with a max-imum of 3600 ind m−3 at Khor Toushka in sum-mer. Small nauplius larvae numerically dominated thecopepod fauna, usually contributing more than 40%to total copepod densities. They peaked in summer, ata mean value of 33 300 ind m−3 and their abundancewas responsible for the modest biomass of the cope-pods as a group. Nauplii indeed contributed little tobiomass (6%), while cyclopoid and calanoid copepo-dids contributed 17–35 and 8–11.0% to total densitiesand biomass, respectively. Khors Allaqi and Koroskohad the highest density (average 22 000 and 21 000ind m−3) while the lowest, 14 900 ind m−3, occurredat Khor Toushka. The summer effect was represen-ted by a peak standing crop of cyclopoid copepodids,44 500 ind m−3 at Khor Toushka, which did not occurin all copepod groups. For example, spring was the topseason for calanoid copepodids, with a peak of 12 000ind m−3 at Khors Kalabsha and Allaqi.
Coastal zone of the main channel: Copepoda domin-ated the zooplankton, especially in autumn (averageabundance 53 000 ind m−3), with 72% of total dens-ities and biomass. That percentage decreased to 47%(abundance) and 50% (biomass) in spring, a trend sim-ilar to that for the total zooplankton of this area. T.galebi was perennial and common, but most abundantin the southern station (Abu Simbel), with a maximumcrop of 10 000 and 11 300 ind m−3 during autumn andspring. Thermocyclops neglectus and Mesocyclopsogunnus were irregularly distributed. Nauplii contrib-uted 32 and 59% to total Copepods during autumnand spring. The density of copepodid larvae was highduring spring (average 23 000 ind m−3) compared toautumn (average 16 900 ind m−3) but without indic-ation of separate generations. The highest naupliusdensities occurred at Korosko. In contrast to naupliuslarvae, cyclopoid and calanoid copepodids were wellrepresented in autumn, contributing 17 and 34% oftotal Copepoda. Their percentage frequency decreasedto 10% in spring. It appears that copepods were un-equally represented in time, but with continuous re-production and high copepodid mortalities.
Cladocera
Khors: Ten species were recorded, contributing 17%to total zooplankton densities, with mean abundance12 000 ind m−3 (Fig. 4a). The contribution of theCladocera to biomass was higher, on average 45%of the total. There was a spatial tendency towards anincrease in abundance upstream. In time, spring wasthe season with the highest standing crop, with a peakof 33 700 ind m−3 at Khor Korosko, dropping to aminimum in summer.
Diaphanosoma mongolianum Ueno (formerly lis-ted from the Nile as D.excisum) and Bosminalongirostris O.F.M. were the most common species. D.mongolianum contributed 53 and 62% to total clado-ceran densities and biomass, with an annual averageof 6300 ind m−3 or 140 mg m−3. D. mongolianumwas thus typically seen in spring. Khor Korosko hadthe highest density (average 10 200 ind m−3) and KhorEl-Ramla the lowest (Figs 3a, 4a).
Bosmina longirostris represented 18 and 6% oftotal cladoceran densities and biomass. Its highestdensity, 18 000 ind m−3, was observed at KhorToushka in autumn, but generally, cladoceran abund-ance showed irregular variation, essentially at Koroskoand Toushka. This, undoubtedly, results from pres-sures other than seasonality or topography, actingsynergistically or antagonistically (see ‘Discussion’).
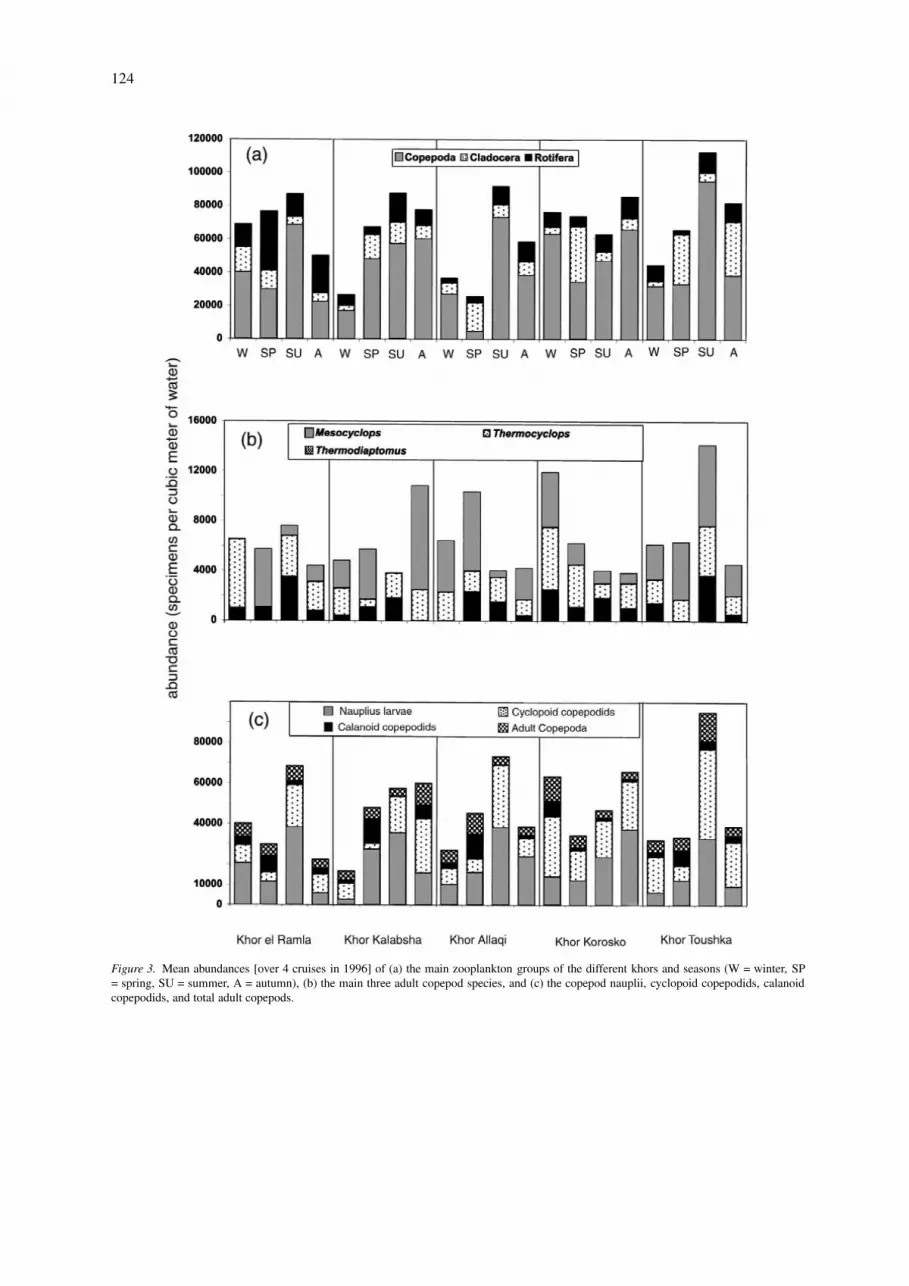
124
Figure 3. Mean abundances [over 4 cruises in 1996] of (a) the main zooplankton groups of the different khors and seasons (W = winter, SP= spring, SU = summer, A = autumn), (b) the main three adult copepod species, and (c) the copepod nauplii, cyclopoid copepodids, calanoidcopepodids, and total adult copepods.

125
Figure 4. Comparative abundance in different khors and seasons of (a) the main cladoceran species, and (b) the dominant rotifer species.Symbols as in Figure 3.
Coastal zone of the main channel: Contrary to cope-pods, Cladocera were weakly represented in autumn,contributing 13 and 27% to total zooplankton densityand biomass. Their density augmented in spring, to25 and 48% of total zooplankton density and biomass.Bosmina longirostris, Ceriodaphnia dubia, Daphnialongispina and Diaphanosoma mongolianum were themost common species. It is worth mentioning that,like in the khors, the comparatively large-sized Daph-nia longispina played a major role in the formationof Cladocera biomass (64 and 49%), in spite of itslow abundance (12 and 16%), and vice versa for B.longirostris.
Rotifera
Khors: Fourteen rotiferan species, contributing 16%
to total zooplankton, were recorded. Their biomassdid not exceed 4%. Khor el Ramla had the higheststanding crop, 21 200 ind m−3, while Khor Allaqihad the lowest, 7100 ind m−3 (Fig. 4b). Summer andautumn were rich with these small-sized organisms,which were poorly represented in winter. Keratellacochlearis dominated, constituting 76 and 12% of totalrotifer densities and biomass, with an annual averageof 8400 ind m−3, but a weight of only 0.9 mg m−3,respectively. K. cochlearis occurred in greatest densityat Khor El-Ramla and diminished upstream. Brachio-nus calyciflorus made up 9 and 11% of total dens-ities and biomass, respectively. It was totally absentduring winter and spring, had its highest density (av-erage 3600 ind m−3) in summer, and was weakly(faintly) represented in autumn. The large, raptorialAsplanchna girodi formed a substantial part of ro-

126
Figure 5. Clustering of the khors according to their zooplanktonfauna.
tifer biomass (67%), in spite of its low abundance(160 ind m−3). Some antagonism between rotifers andcladocerans resulted from the competitive grazing ad-vantage of cladocerans over rotifers (Fig. 4). Indeed,at times and places where cladoceran densities werehigh, rotifer densities tended to be low (correlationcoefficients negative and of the order of 0.2 to 0.3,explaining 5–10% of total variance).
Littoral of the main channel: The rotifer communitycontributed 14.0 and 0.7% to total zooplankton dens-ities and biomass during autumn, but its abundancesteeply increased to 27.3 and 2.1% in spring. The nu-merically dominant rotifer was clearly K. cochlearis,rarely accounting for less than 70% of the total, anda maximum density of 44 000 and 42 500 ind m−3 oc-curred at Korosko and Amada during spring. The otherrotifers were sporadic.
Species richness and other diversity indices
Khors: The highest species richness (23) was foundin Khors Allaqi and Korosko, while Khor Toushkahad the lowest (19 species). Khor Toushka, how-ever, showed the highest diversity and eveness, 0.96and 0.75, respectively. Khor Allaqi had the highestrichness value (1.99) while the lowest (1.6) was atKhor Toushka (Table 2). Cluster analysis of zooplank-ton diversity measures revealed three groups: KhorEl-Ramla, Khors Kalabsha and Allaqi, and KhorsKorosko and Toushka (Fig. 5). The highest similarity(about 92%) occurred within the second group, whileKhor El-Ramla showed the lowest similarity (ca. 70%)to the other khors. These figures are similar, illustrat-ing a high homogeneity among sub-environments.
Coastal zone of the main channel: Abou Simbel andKalabsha had diversity index values, 2.19 and 2.41
during autumn and spring, while Korosko and El-Ramla had lower values, 1.78 and 1.83 (Table 3).Eveness ranged between 0.66 and 0.79 during autumn,but increased to 0.69 and 0.87 in spring (Table 3).Regarding the similarity of species between sites,the cluster analysis revealed a high homogeneitybetween Korosko and Amada (similarity index 89.9)and Kalabsha and Allaqi (similarity index 83.9) dur-ing spring and autumn. The High Dam showed lowestsimilarity with the rest of the sites in autumn. Toushkaand Abou Simbel exhibited similar trends in spring.But as in the case of the khors, these values wererelatively average and unrevealing.
Canonical correspondence analysis identified pat-terns of variation in the zooplankton assemblage re-lative to environmental variables (Fig. 7). Axis 1explained 41.02% of the variance in species scores andcorrelated well with environmental factors that variedover the seasons (i.e. water temperature, COD, andchlorophyll-a). The species-environmental correlationwith axis 1 was 0.89. Total Copepoda and Brachio-nus calyciflorus had the highest scores on axis 1.Ceriodaphnia dubia and Keratella cochlearis showedlowest scores on axis 1. Axis 2 explained 8.85% of thevariation in species score. The species-environmentalcorrelation was 0.73. Axis 2 correlated well with dis-solved oxygen, pH and Secchi depth. Species withhighest scores on axis 2 were Diaphanosoma mongo-lianum and Mesocyclops ogunnus.
Zooplankton distribution, seasonal variation andbiomass
Khors: The upstream khors (Toushka, Korosko) werericher in zooplankton than the downstream khors (El-Ramla and Kalabsha), with an overall average abund-ance of 76 200 ind m−3 (range 44 500–112 500 indm−3) (Fig. 2a–b). Mean standing crop was higherin summer (88 300 ind m−3) than in winter (50 500ind m−3). Average zooplankton density in the khors(69 900 ind m−3) was almost indistinguishable fromthat in the main channel (59 500 ind m−3). Zooplank-ton biomass increased from a minimum 0.4 g m−3 (4g m−2) dry weight in Khor El-Ramla in winter to amaximum of 0.7 g m−3 (7 g m−2) dry weight at KhorToushka in spring.
Littoral of the main channel (Fig. 6): Zooplankton wasmore abundant in spring than in autumn, with a meanof 82 300 ind m−3 (range 42 000–126 500 ind m−3)and 72 700 ind m−3 (range 41 900–121 000 ind m−3),

127
Tabl
e3.
Div
ersi
tyM
easu
res
ofth
eZ
oopl
ankt
onin
the
(sub
)litt
oral
ofth
em
ain
chan
nelo
fL
ake
Nas
ser
Stat
ion
Hig
hD
amel
Ram
laK
alab
sha
Alla
qiel
Mad
iqK
oros
koA
mad
aTo
ushk
aA
buSi
mbe
l
Spec
ies
Aut
.96
Spr.9
7A
ut.9
6Sp
r.97
Aut
.96
Spr.9
7A
ut.9
6Sp
r.97
Aut
.96
Spr.9
7A
ut.9
6Sp
r.97
Aut
.96
Spr.9
7A
ut.9
6Sp
r.97
Aut
.96
Spr.9
7
Spec
ies
num
ber
1216
1414
1316
1411
1615
1717
1311
1316
12
Eve
nnes
s0.
750.
770.
770.
690.
720.
870.
750.
790.
800.
660.
690.
760.
720.
780.
730.
790.
84
Div
ersi
tyin
dex
1.87
2.13
2.04
1.83
1.85
2.41
1.97
1.89
2.23
1.77
1.96
2.16
2.00
1.87
1.85
2.19
2.08

128
Table 4. Abundance of the zooplankton species (m−3) in the khors of Lake Nasser
Khor El-Ramla Khor Kalabsha Khor Allaqi Khor Korosko Khor Toushka
No. W. Wt. No. W. Wt. No. W. Wt. No. W. Wt No. W. Wt.
RotiferaAsplanchna girodi 0 0 0 0 0 0 400 12.1 400 12.1
Brachionus angularis 0 0 0 0 0 0 125 0.1 0 0
Brachionus calyciflorus 125 0.1 450 0.4 625 0.5 1625 1.3 2125 1.7
Brachionus patulus 200 0.1 500 0.3 450 0.2 500 0.3 450 0.3
Filinia longiseta 0 0 0 0 50 0.1 125 0.1 100 0.1
Filinia opoliensis 0 0 0 0 150 0.1 150 0.1 0 0
Hexarthra mira 0 0 0 0 0 0 250 0.1 0 0
Keratella cochlearis 19 875 1.9 7800 0.7 4525 0.4 4700 0.5 4875 0.5
Keratella tropica 0 0 200 0 275 0.1 625 0.1 725 0.1
Lecane luna 125 0.1 0 0 0 0 150 0.1 0 0
Lecane arcula 0 0 0 0 100 0.1 0 0 0 0
Lepadella patella 0 0 0 0 100 0.1 0 0 0 0
Monostyla bulla 0 0 0 0 125 0.1 0 0 0 0
Proalides sp. 825 0.1 375 0.1 675 0.1 675 0.1 0 0
Subtotal 21 150 2.3 9325 1.5 7075 1.8 9325 14.8 8675 14.8
CopepodaNauplius larvae 18 925 15.5 20 325 16.7 21 975 18.1 21 275 17.4 14 900 12.2
Cyclopoid copepodids 10 975 29.6 13 950 37.7 13 725 37.1 21 850 59 22 925 61.9
Calanoid copepodids 4150 30.3 4950 36.1 3975 29.1 2875 20.9 4025 29.4
Thermodiaptomus galebi 1675 63.1 3625 136.7 3350 126.3 1975 74.5 4100 154.6
Thermocyclops neglectus 2775 28.3 1825 18.6 1825 18.6 2925 29.8 2275 23.2
Mesocyclops sp. 1600 46.7 825 24.1 1075 31.4 1600 46.7 1375 40.2
Subtotal 40 100 0 45 500 269.9 45 925 260.6 52 500 248.3 49 600 321.5
CladoceraAlona sp. 100 0.3 125 0.4 0 0 0 0 0 0
Bosmina longirostris 1725 10.2 1250 7.4 1200 7.1 250 1.5 6625 39.1
Chydorus sphaericus 775 5.6 775 5.6 575 4.1 0 0 0 0
Ceriodaphnia quadrangula 1625 29.4 700 12.7 750 31.6 150 2.7 125 2.3
Ceriodaphnia reticulata 925 16.7 700 12.7 100 1.8 425 7.7 1375 24.9
Daphnia barbata 125 4 200 6.4 375 12 0 0 250 8
Daphnia longispina 1675 53.6 0 0 0 0 850 27.2 1500 48
Diaphanosoma excisum 325 9.4 125 3.6 1050 33.6 875 25.41 550 16
Diaphanosoma mongolianum 2075 60.2 5850 169.7 6150 178.4 10150 294.4 7475 216.8
Simocephalus vetulus 0 0 200 8.6 0 0 0 0 0 0
Subtotal 9350 189.4 9925 227.1 10 200 268.6 12 700 358.91 17 900 355.1
Total 70 600 189.4 64 750 498.5 63 200 530.9 74 525 622.01 76 175 691.4
Diversity index 0.903 0.907 0.912 0.891 0.961
Eveness 0.694 0.697 0.670 0.655 0.751
Number of species 20.000 20.000 23.000 23.000 19.000
Richness 1.702 1.715 1.990 1.961 1.601

129
Figure 6. Relative numerical abundance of rotifers, copepods, and cladocerans in spring (SP) and autumn (A) at 9 stations in the (sub)littoralof the main axis of Lake Nubia.
respectively. Zooplankton was richer upstream (23species) than downstream (20 species), with a ratherabrupt transition at km 150, near El Madiq (Fig. 1).Biomass ranged between 0.3 (Allaqi) to 1 g m−3 dryweight (Abu-Simbel) with a mean value of 0.5 g m−3
in autumn. Biomass was particularly high in the up-stream stations in spring, with mean value 0.6 g m−3
(Fig. 6).
Discussion
The distinction of the zooplankton of the lake in abiomass-wise richer upstream and poorer half down-stream is peculiar. This phenomenon is paralleled
by transparency: half upstream (as well the khors asthe main channel) is less transparent (carrying moresediment) and has higher chlorophyll than half down-stream. More particular food is therefore availableupstream, and higher turbidity produces a lower vis-ibility of the zooplankton to predators. In terms ofspecies densities, the more shallow and lacustrinekhors are richer than the main lake, mainly by thepresence of littoral rotifer species. The open lake hasa higher diversity index and higher eveness, however,with little difference between autumn and spring.
A seasonal influence is represented by the yearlyamplitude of water temperature, ranging from 17 to32 ◦C at the surface. This amplitude is comparableto the 12◦ amplitude (18–30 ◦C) of Lake Chad (Car-

130
Figure 7. CCOA plot of the main zooplankton species againstenvironmental variables. Bc = Brachionus calyciflorus, Cd = Ceri-odaphnia dubia, Dl = Daphnia longispina, Dm. = Diaphanosomamongolianum, Kt = Keratella tropica, Ms = Mesocyclops ogunnus,nau = nauplii, tcl = total cladocerans, tco = total copepods, Tg =Thermodiaptomus galebi, Tn = Thermocyclops neglectus, tr = totalrotifers, tz = total zooplankton, wt = water temperature.
mouze et al., 1983). Also, like in Lake Chad, the toptemperatures in Lake Nasser are tropical, whereas theminima remain high enough to allow zooplankton pro-duction to proceed, albeit at a slower pace. Thus, theyearly temperature variation may produce highs andlows, but is insufficient for creating succession of spe-cies. High summer temperatures only correlated with adecrease of Daphnia longispina, Daphnia barbata andCeriodaphnia dubia. According to Kalk (1979) andSaint-Jean (1983), the Daphnia species encountered inAfrican waters are relatively cold stenothermal, suchthat cooler periods of the year favour them. Lévêque& St Jean (1983) found that the Daphnia of LakeChad (and Bosmina as well) had their temperature op-timum close to 22 ◦C. D. barbata and D. lumholtzioccur in the equatorial source lakes of the Nile, inlakes on the east African plateau, and in irrigationchannels and flooded khors along the Nile, either atlow temperatures (winter), or irrespective of temper-ature on condition that no fish are present (Dumont,1986a). Therefore, a low temperature optimum com-bines with high predation in summer to structure thecycle of these two large Daphnia (see Gliwicz, 1994).That the larger, well visible Daphnia suffer more fromfish predation than smaller species, is a paradigm ofplankton ecology (Hrbacek, 1962; Brooks & Dodson,1965; Zaret, 1980; Lazzaro, 1987; for Africa, see Du-mont, 1992), while juvenile crustacean zooplanktersand rotifers may actually increase under fish predation(Nielsen et al., 2000). Invertebrate predation is moreselective for smaller prey size. In Lake Nasser, theimportant invertebrate predator Chaoborus is rare, andtherefore seems not to be a compelling influence. The
cladocerans Diaphanosoma mongolianum, and Bos-mina longirostris are preyed upon by the carnivorousMesocyclops ogunnus and Thermocyclops neglectus,as was observed in live samples aboard the cruiseship. This effect was quantified by negative correla-tion coefficients between both (r = −0.28 and −0.26),which explained ca. 10% of variance, at the limit ofstatistical significance.
Copepoda and Brachionus calyciflorus abundancereacted positively to water temperature, contrary toCeriodaphnia dubia, Bosmina longirostris and Kera-tella cochlearis. However, this may mean little morethan that the former reproduced more quickly thanthat they were eaten away, compared to the latter. Webelieve that antagonistic effects occur between tem-perature preference and predation pressure. CCOAanalysis revealed as well a positive relation betweentotal Cladocera as Diaphanosoma mongolianum, anddissolved oxygen, while nauplius larvae and Bra-chionus calyciflorus showed an ability to live underdissolved oxygen deficiency (Moustafa et al, 1998;El-Shabrawy, 2000). In the khors, Ceriodaphnia du-bia associated with clear water phases, in contrast tonauplii, which are among the few zooplankters thatwere abundant in deep, relatively unsaturated layers.Specifically, Diaphanosoma mongolianum and Bra-chionus calyciflorus related negatively to secchi depthbut positively to chlorophyll-a concentration, mean-ing they were attracted to algal food rather than totransparent water.
On balance, one incisive, if rather subtle effect onLake Nasser zooplankton seems to be due to pred-atory fish (Lazzaro, 1987). Which species could beinvolved? Drenner et al. (1982) offered Lake Kinneretzooplankton to Sarotherodon galilaeus in laboratorytrials. Not only did this omnivore eat a share ofzooplankton, but it took the ‘classical’ pathway: se-lectively removing large Cladocera first, next smallCladocera, and only then small copepods and cope-podids of large species. Why, then, do tilapias suchas Oreochromis niloticus and Sarotherodon galilaeus,together forming over 90% of Lake Nasser’s ichthy-omass (Ahmed, 1998), not have a more profoundeffect on the plankton? The most probable answeris that tilapias, especially those in their early stages,use zooplankton, but most are riverine and pond spe-cies that only occasionally venture into the open waterof lakes. They may drive large vulnerable cladocer-ans extinct, but not the small species, nor excellentswimmers-escapers such as most copepods. Inver-tebrate predators such as adult Mesocyclops ogunnus

131
might occasionally control some cladocerans and largerotifers, with inter-group competition between rotifersand cladocerans at times also important.
The absence of a pelagic planktivore from the lakeis striking. Such planktivores, usually clupeids, existin many river systems and lakes elsewhere in Africa.Why they do not exist in the Nile is unknown. Care-ful studies on the beneficial as well as detrimentaleffects of the introduction of such a planktivorousfish into lake Nasser might therefore offer a viablestarting point for a better use of the lake as a fish-eries resource in the future. One possible species isLimnothrissa miodon from Lake Tanganyika. It hasbeen introduced to and successfully exploited in LakesKariba and Cahora Bassa (Zambezi basin), wherethere are no signs of a detrimental effect of the sardineon the zooplankton (Balon & Coche, 1974; Jackson,1986). In Lake Kivu, in contrast, it has decimated thezooplankton, (Dumont, 1986b) and subsequently re-verted to cannibalism (De Iongh et al., 1983). A phaseof careful testing should therefore precede its possibleintroduction to Lake Nasser.
Acknowledgements
Our special thanks to Dr X. Lazzaro for his incis-ive and constructive comments on a couple of earlierversions of this paper.
References
Abd Allah, R. G., 1994. Heat, salt and water budget of Lake Nasser.M.Sci. Thesis, Assiut Univ., 110 pp.
Abdel-Mageed, A. A., 1992. Ecological studies on zooplanktonfrom Khor El-Ramla (Lake Nasser). M.Sci. Thesis, Al-AzharUniv., 219 pp.
Abdel-Mageed, A. A., 1995. Studies on zooplankton from LakeNasser Egypt. Ph.D. Thesis, Fac. Sci. Al-Azhar Univ., 215 pp.
Abu-Gideiri, Y. B. & M. T. Ali, 1975. A preliminary biologicalsurvey of Lake Nubia. Hydrobiologia 46: 535–541.
Ahmed, O. K., 1998. The population dynamics of Oreochromisniloticus and Sarotherodon galilaeus from Lake Nasser. Ab-stracts of background papers on the ecological basis and man-agement policy sustainable fish production in Lake Nasser.ICLARM, p. 5.
Agaypi, M. Z., 1995. A preliminary report of catch correspondingto the water level in the High Dam Lake. Working report. FishMgmt Center Aswan 4: 25–32.
Ali, M. T., 1984. Fishes and fisheries of Lake Nubia, Sudan.Hydrobiologia 110: 305–314.
Anonymous, 1996. Studies of the environmental characteristics,sediment, and fisheries of Lake Nasser, first annual report, NIOFto the General Authorities for development of High Dam Lake,Aswan, 165 pp., in Arabic.
Balon, E. K. & A. Coche (eds), 1974. Lake Kariba, a man-madetropical ecosystem in Africa. Monographiae Biologicae 24. DrW. Junk Publishers, The Hague: 767 pp.
Brooks, J. L. & S. I. Dodson, 1965. Predation, body size and thecomposition of plankton. Science 150: 26–35.
Carmouze J-P, J-R Durand & C. Léveque (eds), 1983. LakeChad. Ecology and Productivity of a shallow tropical ecosystem.Monographiae Biologicae 53: 575 pp. Dr W. Junk Publishers,The Hague.
De Iongh, H. H., P. C. Spliethoff & V. G. Frank, 1983. Feedinghabits of the clupeid Limnothrissa miodon (Boulenger) in LakeKivu. Hydrobiologia 102: 113–122.
Drenner, R. W., G. L. Vinyard, M. Gophen, & S. R. McComass,1982. Feeding behavior of the cichlid Sarotherodon galilaeus:selective predation on Lake Kinneret zooplankton. Hydrobiolo-gia 87: 17–20.
Dumont, H. J., 1986a. Zooplankton of the Nile system. In Davies,B. R. & K. F. Walker (eds), The Ecology of River Systems.Monographiae Biologicae 60: 75–88. Dr W. Junk Publishers,The Hague.
Dumont, H. J., 1986b. The Lake Tanganyika sardine in Lake Kivu:another ecodisaster for Africa? Envir. Conserv. 13: 143–148.
Dumont, H. J., 1992. The regulation of plant and animal speciesand communities in African shallow lakes and wetlands. Rev.Hydrobiol. trop. 25: 303–344.
Dumont, H. J., I. van de Velde & S. Dumont, 1975. The dryweight estimate of biomass in a selection of Cladocera, Cope-poda and Rotifera from the plankton, periphyton and benthos ofcontinental waters. Oecologia 19: 75–97.
El-Shabrawy, G. M., 2000. Seasonal and spatial variation inzooplankton structure in Lake Nasser. I. Pelagic area of the mainchannel. Egypt. J. aquat. Biol. Fish. 4: 61–84.
Entz, B. A. G., 1973. Morphometery of Lake Nasser. Project R.P.H,UNDP SF. F.A.O.
Entz, B., 1976. Lake Nasser and Lake Nubia. In Rzoska, J. (ed.),The Nile. Monographiae Biologicae 29: 271–298. Dr W. JunkPublishers, The Hague.
Gliwicz, Z. M., 1994. Relative significance of direct and indir-ect effects of predation by planktivorous fish on zooplankton.Hydrobiologia 272: 201–210.
Greenwood, P. H., 1976. Fish Fauna of the Nile. In Rzoska, J. (ed.),The Nile. Monographiae Biologicae 29: 127–141. Dr W. JunkPublishers, The Hague.
Habib, O. A., 1996. Changes of the distribution of zooplankton atfive stations of Khor El-Ramla of Lake Nasser. Working reportof fishery management center Vol. 5: 41–54.
Hrbacék, J., 1962. Species composition and the amount of zo-oplankton in relation to the fish stock. Rozpr. CSAV, Ser. Math.Nat. Sci. 72: 1–117.
Iskaros I. A., 1993. Ecological studies on the distribution of zo-oplankton and benthic fauna in Khor Kalabsha in Lake Nasser.Ph.D. Thesis, Fac. Sci. El-Mansoura Univ., 310 pp.
Jackson, P. B. N., 1986. Fish of the Zambezi System. In Davies,B. R. & K. F. Walker (eds), The Ecology of River Systems.Monographiae Biologicae 60: 269–288. Dr W. Junk Publishers,Dordrecht.
Kalk, M., 1979. Zooplankton in Lake Chilwa: adaptations tochange. In Kalk, M., A. J. MaLachlan & C. Howard-Williams(eds), Lake Chilwa. Monographiae Biologicae 35: 123–141. DrW. Junk Publishers, The Hague.
Latif, A. F. A., 1976. Fish and fisheries of Lake Nasser. In Rzoska,J. (ed.), The Nile. Monographiae Biologicae 29: 299–307. Dr W.Junk Publishers, The Hague.

132
Latif, A. F. A., 1978. Effect of impoundment on Nile biota in LakeNasser. In Proceedings of an International Symposium on the En-vironmental effects of Hydraulic Engineering Works. TennesseeValley authority Knoxville, Tennessee: 435–445.
Latif, A. F. A., 1984. Lake Nasser. The new man-made lake in Egypt(with reference to Lake Nubia). In El-Serveir, F. B. T. (ed.),Ecosystems of the World 32, Lakes and Reservoirs: 385–416.
Lauzanne, L., 1983. Trophic relations of fishes in Lake Chad. InCarmouze, J-P., J-R. Durand & C. Léveque (eds), Lake Chad.Monographiae Biologicae 53: 385–424. Dr W. Junk Publishers,The Hague.
Lazzaro, X., 1987. A review of planktivorous fishes: their evolution,feeding behaviours, selectivities, and impacts. Hydrobiologia146: 97–167.
Lévêque, C. & L. Saint-Jean, 1983. Secondary Production (Zo-oplankton and Benthos). In Carmouze, J-P. et al. (eds), LakeChad. Monographiae Biologicae 53: 385–424. Dr W. JunkPublishers, The Hague.
Moustafa, A. S., M. A. Hussein, A. H. Obuid Allah & A. A. Mah-moud, 1998 Taxonomical and ecological studies of fresh waterCladocera in Qena Governorate Upper Egypt. J. Egypt. Ger. Soc.Zool. 26(D): 1–19.
Nielsen, D. L., T. J. Hillman, F. J. Smith & R. J. Shiel, 2000. Theinfluence of a planktivorous fish on zooplankton assemblages inexperimental billabongs. Hydrobiologia 434: 1–9.
Podani, J., 2000. Introduction to the Exploration of MultivariateBiological Data. Backhuys, Leiden, 407 pp.
Saint-Jean, L., 1983. The zooplankton. In Carmouze, J. P. et al.(eds.) Lake Chad. Monographiae Biologicae 53: 199–232. Dr W.Junk Publishers, The Hague.
Samaan, A. A., 1971. Report on the trip of Lake Nasser to investig-ate its primary productivity during March, 1971. Report for LakeNasser Development Center, 11 pp.
Zaret, T., 1980. Predation and Freshwater Communities. YaleUniversity Press, New Haven, 187 pp.





























