Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Speleothems from Mawsmai and Krem Phyllut caves, Meghalaya,India: some evidences on biogenic activities
Sushmitha Baskar Æ R. Baskar Æ Natuschka Lee ÆP. K. Theophilus
Received: 22 February 2008 / Accepted: 27 May 2008 / Published online: 19 June 2008
� Springer-Verlag 2008
Abstract The Mawsmai cave and Krem Phyllut caves,
East Khasi hills, Meghalaya, India has so far not yet
attracted the attention of geomicrobiologists. Observations
and hypotheses on the possible influence of identified
microorganisms for speleothem formations in Meghalaya
are reported for the first time. XRD studies identified cal-
cite in speleothems and gypsum in cave wall deposits as the
dominant minerals. SEM-EDAX showed interesting mi-
crofabric features showing strong resemblance with
fossilised bacteria, calcified filaments, needle calcite and
numerous nano scale calcite crystals, highly weathered and
disintegrated crystals of calcite, that point towards a sig-
nificant microbial influence in its genesis. Thin section
petrography showed laminated stromatolitic features. The
microorganisms identified by conventional isolation and
further evaluation of isolates by molecular techniques
include Bacillus cereus, Bacillus mycoides, Bacillus
licheniformis, Micrococcus luteus, and Actinomycetes.
Microscopic observations also showed unidentifiable cocci
and four unidentifiable strains of CaSO4 (gypsum) precip-
itating bacteria. Experimental studies confirmed that these
bacteria are able to precipitate calcium minerals (calcite,
gypsum, minor amounts of dolomite) in the laboratory.
These results allow us to postulate that species like these
may contribute to active biogenic influence in the cave
formations at Meghalaya.
Keywords Geomicrobiology � Caves �Carbonate speleothems � Calcium minerals �Biogeochemistry � Bacteria
Introduction
Caves have always haunted the imagination of mankind—
already from the start of human history, these natural for-
mations provided shelter to early man and were sought
after earnestly, to provide a ‘safe home’. Caves are also
natural habitats for a wide spectrum of fascinating life
forms. Caves offer natural, experimental study systems of
the subsurface for fundamental geomicrobiological studies,
because they are relatively easy to access (Culver 1982;
Frey 1963). Calcium carbonate speleothems dominate in
most known caves of the world and a number of geomi-
crobiological studies have been reported in literature on
stalactites, stalagmites, helictites, moonmilk, pool fingers
and cave pearls. Microorganisms impact significantly their
environments and in turn microbial communities are
affected and shaped by the geochemistry of their environ-
ment. Recent studies have clearly identified some of the
factors that control the contribution of microbes to CaCO3
precipitation (Castanier et al. 2000; Engel et al. 2003,
2004; Baskar et al. 2005, 2006, 2007). Laboratory experi-
ments support that the microbial species isolated from
these minerals can produce similar crystals from organic
calcium salts in the lab (Rivadeneyra et al. 1993; Warth-
mann et al. 2000; Baskar et al. 2006).
Caves are generally considered as extreme environments
for life. They are mostly resource limited due to the
S. Baskar � R. Baskar (&) � P. K. Theophilus
Department of Environmental Science and Engineering,
Guru Jambheshwar University of Science and Technology,
Hisar 125001, Haryana, India
e-mail: [email protected]
N. Lee
Division Microbial Systems Ecology (MSE),
Department of Microbiology,
Technical University of Munich (TUM),
Am Hochanger 4, 85354 Freising, Germany
123
Environ Geol (2009) 57:1169–1186
DOI 10.1007/s00254-008-1413-y

prevailing aphotic conditions and subsequently there is no
photosynthesis; thus, this drives most cave ecosystems to
depend on allochthonous organic materials for energy
(Poulson and Lavoie 2000; Simon et al. 2003). The entry of
dripping, seeping and flowing water brings energy into the
caves. Groundwater discharges as springs into the passages
of some caves (Sarbu et al. 1996; Egemeier 1981; Angert
et al. 1998; Hose et al. 2000) also serve as high energy-
yielding substrates for some microorganisms. Microorgan-
isms control biogeochemical cycles and their diverse
metabolic pathways convert minerals into chemical forms
that are readily usable by other organisms (Elia 2002).
In the past decade, cave microbiology has emerged as
one of the frontier areas of geobiology research involving
the co-ordinated efforts of microbiologists, chemists and
geologists to address challenging questions regarding
microbial metabolism, biogeochemistry and their role in
mineral precipitation/dissolution. Bacterial communities in
caves are known to acquire energy by, e.g. transforming
aromatic compounds, fixing gases, and oxidising reduced
metals within rocks. By their interaction with minerals,
microbial species play an important role in reshaping the
mineral environment of caves and helps form features such
as stalactites, stalagmites and various cave wall deposits.
India has a large number of unexplored caves of which
the Meghalaya caves are the largest, which have so far not
yet attracted scientific attention. This study reports the first
geomicrobiological investigations, general geochemistry,
in vitro precipitation experiments, and provides further
indications on the significant role of bacteria as geochem-
ical agents in the genesis of the speleothems from two
caves in East Khasi hills, Meghalaya (India).
Study area and geology
There are more than 1,000 caves in Meghalaya (a few of
them forming one of the longest caves on the Indian sub-
continent) and the caves in the East Khasi Hills are Krem
Phyllut, Mawsmai, Krem Mawmluh, Krem Soh Shympi
and Krem Dam.
The Khasi Hills (Meghalaya Plateau) is an uplifted
Precambrian crystalline complex and forms the northeast-
ern extension of the Indian Peninsular Shield. It is an E–W
trending oblong horst block elevated about 600–1,800 m
above the Bangladesh plains in the south and separated
from Peninsular India by the Rajmahal-Garo gap (Ghosh
et al. 2005). The caves investigated in this study are
located on the East Khasi Hills (25�0700; 25�4100 North
Latitudes and 91�2100; 92�0900 East Longitudes), bounded
by Ri-Bhoi District on the north, Karbi Anglong District on
the north east, Jaintia Hills district on the east, Bangladesh
on the south and West Khasi Hills district on the west
(Fig. 1a–c). The Khasi group consists of sandstone and
conglomerate of Jadukata formation overlying the feld-
spathic sandstone of Mahadek formation. Isolated patches
of older Alluvium overlie the Tertiary rocks along the
southern fringes of Khasi Hills and recent alluvium is
found in the river valleys in the northern foothills region.
The Proterozoic meta sedimentary Shillong Group and
the basement Gneissic Complex make up most of the
Meghalaya plateau (Ghosh et al. 2005). The southern part
of the plateau is covered by Cretaceous Sylhet basalt and
Tertiary shelf sediments. A number of prominent linea-
ments trending NE–SW and E–W are present in the
plateau; the most prominent structural feature is the E–W
trending Dauki fault, which marks the southern border of
the plateau. The granite plutons occur in greater number in
the eastern part of the plateau compared to the western part.
Meghalaya has huge deposits of limestone and abundant
rainfall, which is the main reason for the Karst cave for-
mations. All the three hills, namely, Khasi, Jaintia and
Garo contain limestone of variable quantity and quality.
The important limestone deposits in the East Khasi Hills
are in Cherrapunji. This deposit is located in the
Mawmluh-Mawsmai Hills (study area) south of lower
Cherrapunji. The deposit is about 1.40 km in area. The
deposit is of a composite nature made up of limestone in
the upper part and dolomite in the lower part. These caves
are not so easily accessible for the public since they are
situated in a hilly, uninhabited area and are thus scarcely
visited. Here, we report our initial geomicrobiological
studies of two of the Meghalaya caves, the Mawsmai and
the Krem Phyllut.
Sites of study
Mawsmai cave
These caves are located amidst a thickly forested zone and
are quite small (160 m long, 15 m high, width 4–10 m),
but the inner parts are large enough to facilitate easy
movement within them. It is an important site for tourism,
but the cave speleothems are not allowed to be touched by
the public due to strict rules set by the state government
cave authorities. The main entry to these caves is located
close to the Mawsmai village and the entry is a fairly
narrow (1.8 m) vertical opening. The cave was totally
aphotic and has myriads of stalagmites and stalactites. The
only cave fauna observed were spiders and rodents in
rather sporadic amounts. The cave was moist and only
dripping water could be observed. The average annual
temperature of the inner cave is around 15–19�C (Table 1).
The length and diameter of the stalactites ranged from
7–10 cm length, 8–15 cm diameter (small) to 50–1.5 m
length, 50–100 cm diameter (large).
1170 Environ Geol (2009) 57:1169–1186
123

Krem Phyllut cave
This cave is also situated in Mawsmai village, south of
Cherrapunjee. This cave has a large section of fossil pas-
sage, two stream ways that join inside the cave and then
leave out of the cave. This cave was relatively long (total
length of 1,003 m, width 4.5 m, height 15 m) and had three
entrances (approximately of 2 m height, 2.5 m width).
Inside the cave the observed zones include a photic zone,
twilight zone with limited light penetration and a deeper
zone, which was totally aphotic. The average annual tem-
perature of the deep aphotic inner cave wall was
approximately 15–17�C and discharged two springs that
run approximately 500 m through the length of the cave.
The approximate length and diameter of the stalactites
ranged from 6–7 cm in length, 25–30 cm in diameter
(small) to 30–40 cm in length, 50 cm in diameter (large)
and the columns were 40–45 cm long and diameter of the
upper end was 40 cm and that of the lower end was 60 cm
(Table 1). The speleothems at the entrance were large and
towards the interior they were smaller in size. Interestingly,
some of the stalactites observed showed nodular and pop-
corn-type morphologies (Fig. 2a–c). The main cavern
extends to about 50–60 m and narrows to a much smaller
tunnel. Gypsum deposits on the cave walls were found
here. The characteristic fauna here consisted of sporadic
amounts of prawns and rats.
Methodology
Sampling
Samples for the studies were taken from minimal con-
taminated (not disturbed by human/anthropogenic
activities) areas of the cave and were collected with sterile
disposable gloves, sterile forceps, sterile ziplock sachets
and sterile autoclaved bottles. Ten aliquots of spring
waters (2 l each) and ten speleothem samples (0.5 kg
each) (including stalactites, stalagmites, cave wall
Fig. 1 a Location map of the study area. Inset shows its position
in India. b Geological map of Meghalaya (Ghosh et al. 2005).
c Geological map of the study cave area (Ghosh 1940; Garg and Jain
1995). 1 Alluvium, 2 Kopili formation (shale/sandstone), 3 Sylhet
limestone, 4 Therria formation (calcareous bands, sandstone, shale),
5 Langpar formation and Um Sohryngkew formation (calcareous
shale, marl and mudstone), 6 Sylhet trap, 7 Archaen, 8 K/T Sect., 9Um Sohryngkew river
Environ Geol (2009) 57:1169–1186 1171
123
deposits and pool sediments), three each from different
areas of the cave (photic, aphotic and twilight areas) were
collected in sterilised bottles, sterile ziplock sachets and
stored at 4�C for up to 30 days before different geo-
chemical and microscopic analysis. The microbial
community structure was characterised and identified
using a combination of conventional isolation efforts and
molecular techniques of isolated specimens, examination
of morphology by SEM, and subsequent quantitative
analyses by EDX.
Geochemical analysis
The major and trace element rock geochemistry of the
speleothems (Na2O, MgO, SiO2, K2O, CaO, P2O5, Al2O3,
TiO2, MnO, Fe2O3, Sr, Ni, Cu, Zn, Ga, Pb, Th, Rb, U, Y,
Zr, Nb) were analysed using an automated sequential X-ray
fluorescence spectrometer WDXRF (Siemens SRS 3000,
Wadia institute of Himalayan Geology, Dehradun). The
geochemistry of the spring, pool and dripping water sam-
ples (Mn, Cu, Zn, Cd, Ba, Sr, Cr, Ag, Mg, Si, V) were
analysed using ICP-MS (Wadia Institute of Himalayan
Geology, Dehradun). The acidity, alkalinity, hardness,
conductivity and pH of spring waters and total organic
carbon content of speleothems was determined by using
standard procedures at the Environmental Geology Labo-
ratory, Guru Jambheshwar University of Science and
Technology, Hisar, India.
Electron microscopy and mineralogy
Scanning electron microscopic (SEM ZEISS EVO 40
EP, resolution, 3.0 nm SE and HV; magnification,
7–1,000,0009; accelerating voltage, 0.2–30 kV; WIHG
Dehradun) studies were performed on the speleothems and
bacterially precipitated (in vitro) mineral samples. EDX
(EDX microanalyser Bruker LN2 Free X Flash 4010SDD
detector, resolution, 129 eV at Mn ka9 (5.98 keV) and XRD
(X0Pert Pan X-ray diffractometer, WIHG, Dehradun) was
used for quantitative estimation of the chemical composi-
tion of the minerals. The speleothem samples and samples
postulated to contain biomass were fixed onto aluminum
stubs with two-way adherent tabs with conductive paint,
and allowed to dry (Leveille et al. 2000). They were then
gold-coated by sputtering for approximately 2–3 min.
Some of the speleothem samples were acid-etched (3 and
10% HCl) before SEM examination (Melim et al. 2001).
For EDX the samples were coated with carbon (CC7650
carbon evaporation coater) and then transferred to the
sample chamber of the instrument prior to imaging. The
SEM was operated with a working distance of 15 mm for
optimum imaging and to minimize charging and sample
damage. For X-ray analysis, an accelerating voltage of
Table 1 General characteristics of speleothems from East Khasi Hill caves, Meghalaya
S.no. Sample no. Type Mineral Length (cm) Diameter
(cm)
Temperature
(�C)
Zone Distance from
cave entrance (m)
Mawsmai cave (MC1R)
1 MC1R-12 Stalagmite Calcite 7–50 8–100 17 Aphotic 90
2 MC1R-18 Stalactite Calcite 15–35 10–90 16 Aphotic 100
Krem Phyllut cave (MC2R)
3 MC2R-02 Stalactite Calcite 30–40 35–50 15–17 Aphotic 90–100
4 MC2R-08 Cave wall deposit Gypsum 12–15 15 16 Aphotic 260–270
5 MC2R-11 Column Calcite, aragonite 40–45 40 Upper,
50 lower end
16 Aphotic 330
6 MC2R-13 Pool water sediment SiO 20–25 10–20 17 Aphotic 360
7 MC2R-15 Stalagmite Gypsum 15–20 25–30 17 Aphotic 400
8 MC2R-23 Cave wall deposit Calcite 7–50 30 16 Aphotic 70–80
9 MC2R-25 Stalagmite 7–40 6–30 17 Twilight 20
Fig. 2 Speleothems, pool waters from Meghalaya caves. b Nodular
and popcorn crusts (encircled)
1172 Environ Geol (2009) 57:1169–1186
123

15 kV was used to obtain sufficient X-ray counts. Samples
for XRD were dried, powdered and scanned between 4�and 64� 2h at 1� 2h min-1. The minerals were identified
using the JCPDS–ICDD, XRPD database (2000).
Microbiological analysis
Attempts to isolate bacteria from the speleothems and cave
wall deposits from both of the caves were made for identi-
fying the culturable, aerobic, heterotrophic fraction of the
total microbial community. Stalactite, stalagmite and cave
wall deposit samples (1 g) were powdered in a sterilised set
of mortar and pestle, suspended in sterile 0.9% saline solu-
tion and briefly vortexed. Thiosulphate and sulphite agar
(tryptone 10 g l-1, sodium sulphite 1 g l-1, agar 20 g l-1)
based media were used to isolate bacterial isolates from the
cave wall deposits at pH values of 4–6. Standard nutrient-
rich media such as nutrient agar and B-4 agar (Boquet et al.
1973) were used to isolate bacteria from the speleothems.
The agar plates were inoculated with sample dilutions
ranging from 101 to 106 and incubated under aerobic con-
ditions between 15 and 25�C (photic, aphotic conditions
separately designed to mimic cave temperatures) for
2 weeks. The initial plate counts were approximately
12 9 102 g l-1 (sulphite agar), 16 9 101 g l-1 (thiosul-
phate agar), 40 9 102 g l-1 (nutrient agar), 32 9 102 g l-1
and contained a large variation of colony-morphotypes, of
five different colours (yellowish, pale cream, creamish,
whitish brown, red). Individual colonies were selected
(based on differences in colour, colony morphotypes) and
purified by repeated streaking. For short-term preservation,
the isolates were streaked on respective agar slants and
stored at 4�C for 30 days before renewed inoculation.
Microbe–mineral precipitation experiments were designed
using temperatures and salt concentrations to mimic the cave
environment. Bacterial isolates capable of precipitating
minerals were streaked for purity and further identified by
conventional and molecular techniques and the mineral type
and chemistry of the precipitated bio-mineral was deter-
mined using XRD and EDX. Altogether 30 purified isolates
were obtained and five of these isolates were deposited at the
strain collection MTCC (Microbial Type Culture Collection
and Gene Bank), Institute of microbial technology, IM-
TECH, Chandigarh, India. Strains 2, 4, and 8 were screened
by conventional and DNA sequencing identification meth-
ods, whereas strains 5 and 13 were characterised by
conventional phenotypical methods. The origin of the strains
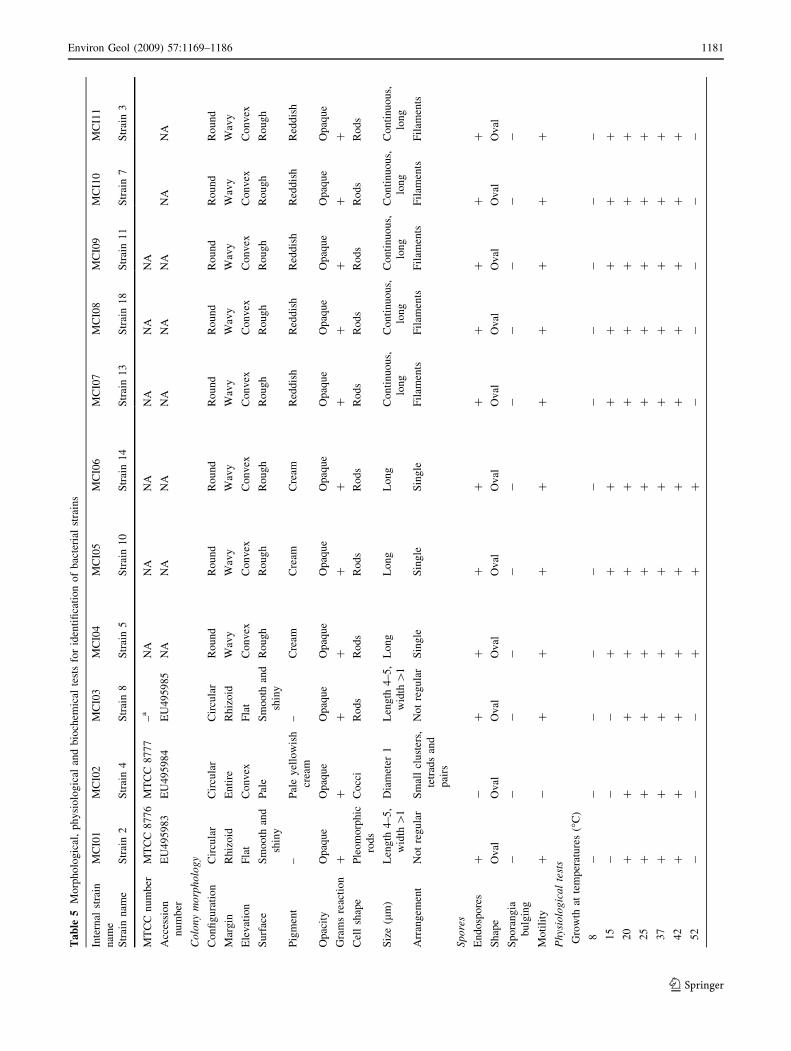
and their designations are listed in Table 2.
The isolates were identified using conventional (mor-
phological, biochemical and physiological tests) and DNA
sequencing identification methods, based on PCR amplifi-
cation of 16S rRNA genes (16S rDNA) and partial
sequencing (*800–1,000 bp). The partial 16S rRNA gene
sequences were compared with the large nucleotide data-
base of the National Center for Biotechnology Information
NCBI by submitting the sequences to BLAST (Altschul
et al. 1990, http://www.ncbi.nlm.nih.gov/). All (three in
all) sequences, as well as relevant reference 16S rRNA
gene sequences identified by BLAST search in NCBI were
imported to the phylogenetic software ARB (Ludwig et al.
2004; http://www.arb-home.de) for alignment and sub-
sequent calculation of a phylogenetic tree based on the
neighboring joining hood algorithm, to confirm the iden-
tities suggested by BLAST. The sequences retrieved in this
study have been submitted to NCBI, and their accession
numbers are: EU495983, EU495984 and EU495985.
Results and discussions
The purpose of this study was to investigate the biogenic
influence on the cave formation in two of the Meghalaya
caves in the Indian subcontinent. For this, we combined
geochemical, scanning electron microscopical, microbio-
logical and in vitro precipitation studies. Here, we present
our results from these studies on ‘‘Speleothem carbonates’’
as well as on ‘‘Gypsum and cave wall deposits’’. In ‘‘Bio-
genic influences on speleothem carbonates’’ and ‘‘Biogenic
influence on gypsum and cave wall deposits’’, we present
our hypotheses on biogenic influences on these systems.
Speleothem carbonates
Petrographic observations
It is crucial to determine the conditions under which spe-
leothems form by studying the fabrics preserved in the cave
precipitates/deposits. The criteria for recognising microbes
and microbial activity in speleothems and cave deposits
include documentation and recognition of mineralised
microbes, recognition of stromatolitic structures that are
wavy or laminated structures formed by microbes that trap
and bind detrital grains to a substrate or act as nucleation
sites for mineral precipitation and the identification of
fabrics/textures that are known to be indicative of micro-
bial activity (Jones 2001).
Petrography of our speleothem samples showed alter-
nating white and dark-laminated bands and voids (Fig. 3a,
b), and the presence of lithified structures and internal
fabrics similar to microbialites along with some organic
inclusions. Dark coloured clotted peloidal fabrics (Fig. 3c)
were observed, which may be interpreted as calcified
bacterial aggregates since such formations have quite often
been reported in stromatolites, thrombolites, travertines,
and in reefs (Chafetz and Buczynski 1992; Pedley 1992;
Riding 2000; Baskar et al. 2007).
Environ Geol (2009) 57:1169–1186 1173
123
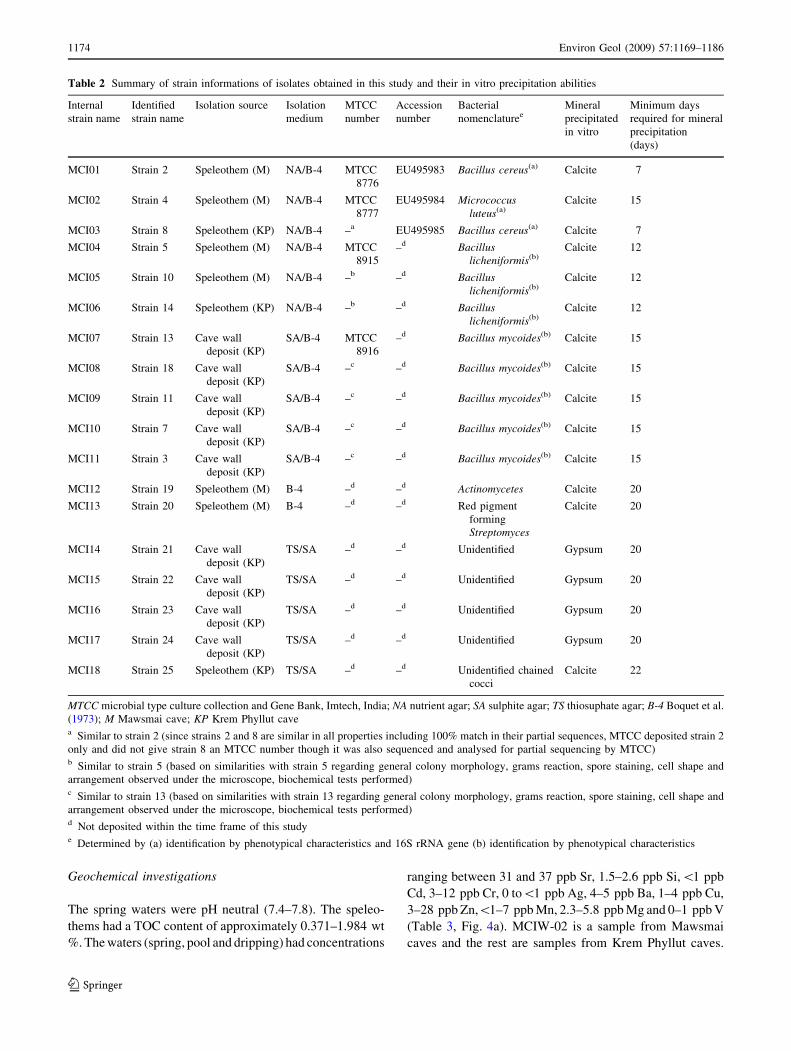
Geochemical investigations
The spring waters were pH neutral (7.4–7.8). The speleo-
thems had a TOC content of approximately 0.371–1.984 wt
%. The waters (spring, pool and dripping) had concentrations
ranging between 31 and 37 ppb Sr, 1.5–2.6 ppb Si,\1 ppb
Cd, 3–12 ppb Cr, 0 to\1 ppb Ag, 4–5 ppb Ba, 1–4 ppb Cu,
3–28 ppb Zn,\1–7 ppb Mn, 2.3–5.8 ppb Mg and 0–1 ppb V
(Table 3, Fig. 4a). MCIW-02 is a sample from Mawsmai
caves and the rest are samples from Krem Phyllut caves.
Table 2 Summary of strain informations of isolates obtained in this study and their in vitro precipitation abilities
Internal
strain name
Identified
strain name
Isolation source Isolation
medium
MTCC
number
Accession
number
Bacterial
nomenclatureeMineral
precipitated
in vitro
Minimum days
required for mineral
precipitation
(days)
MCI01 Strain 2 Speleothem (M) NA/B-4 MTCC
8776
EU495983 Bacillus cereus(a) Calcite 7
MCI02 Strain 4 Speleothem (M) NA/B-4 MTCC
8777
EU495984 Micrococcusluteus(a)
Calcite 15
MCI03 Strain 8 Speleothem (KP) NA/B-4 –a EU495985 Bacillus cereus(a) Calcite 7
MCI04 Strain 5 Speleothem (M) NA/B-4 MTCC
8915
–d Bacilluslicheniformis(b)
Calcite 12
MCI05 Strain 10 Speleothem (M) NA/B-4 –b –d Bacilluslicheniformis(b)
Calcite 12
MCI06 Strain 14 Speleothem (KP) NA/B-4 –b –d Bacilluslicheniformis(b)
Calcite 12
MCI07 Strain 13 Cave wall
deposit (KP)
SA/B-4 MTCC
8916
–d Bacillus mycoides(b) Calcite 15
MCI08 Strain 18 Cave wall
deposit (KP)
SA/B-4 –c –d Bacillus mycoides(b) Calcite 15
MCI09 Strain 11 Cave wall
deposit (KP)
SA/B-4 –c –d Bacillus mycoides(b) Calcite 15
MCI10 Strain 7 Cave wall
deposit (KP)
SA/B-4 –c –d Bacillus mycoides(b) Calcite 15
MCI11 Strain 3 Cave wall
deposit (KP)
SA/B-4 –c –d Bacillus mycoides(b) Calcite 15
MCI12 Strain 19 Speleothem (M) B-4 –d –d Actinomycetes Calcite 20
MCI13 Strain 20 Speleothem (M) B-4 –d –d Red pigment
forming
Streptomyces
Calcite 20
MCI14 Strain 21 Cave wall
deposit (KP)
TS/SA –d –d Unidentified Gypsum 20
MCI15 Strain 22 Cave wall
deposit (KP)
TS/SA –d –d Unidentified Gypsum 20
MCI16 Strain 23 Cave wall
deposit (KP)
TS/SA –d –d Unidentified Gypsum 20
MCI17 Strain 24 Cave wall
deposit (KP)
TS/SA –d –d Unidentified Gypsum 20
MCI18 Strain 25 Speleothem (KP) TS/SA –d –d Unidentified chained
cocci
Calcite 22
MTCC microbial type culture collection and Gene Bank, Imtech, India; NA nutrient agar; SA sulphite agar; TS thiosuphate agar; B-4 Boquet et al.
(1973); M Mawsmai cave; KP Krem Phyllut cavea Similar to strain 2 (since strains 2 and 8 are similar in all properties including 100% match in their partial sequences, MTCC deposited strain 2
only and did not give strain 8 an MTCC number though it was also sequenced and analysed for partial sequencing by MTCC)b Similar to strain 5 (based on similarities with strain 5 regarding general colony morphology, grams reaction, spore staining, cell shape and
arrangement observed under the microscope, biochemical tests performed)c Similar to strain 13 (based on similarities with strain 13 regarding general colony morphology, grams reaction, spore staining, cell shape and
arrangement observed under the microscope, biochemical tests performed)d Not deposited within the time frame of this studye Determined by (a) identification by phenotypical characteristics and 16S rRNA gene (b) identification by phenotypical characteristics
1174 Environ Geol (2009) 57:1169–1186
123

MC2W-01, a drip water sample is enriched in Zn compared
to other samples. Geochemically, the speleothems contained
45–54.67% Ca, 0.12–1.69% Fe2O3, 0.31–0.91% MgO,
0.002–0.131% Na2O, 0–3.24% Al2O3 and Sr 46–501 ppm
(Table 4, Fig. 4b). The pool sediments of the sampled caves
have 19.8% iron and are basically pyrite (Table 4). XRD
revealed calcite as the dominant mineral and traces of
dolomite, aragonite, iron titanium hydrate, copper minerals,
tin telluride, palladium hydride, quartz, and iron minerals.
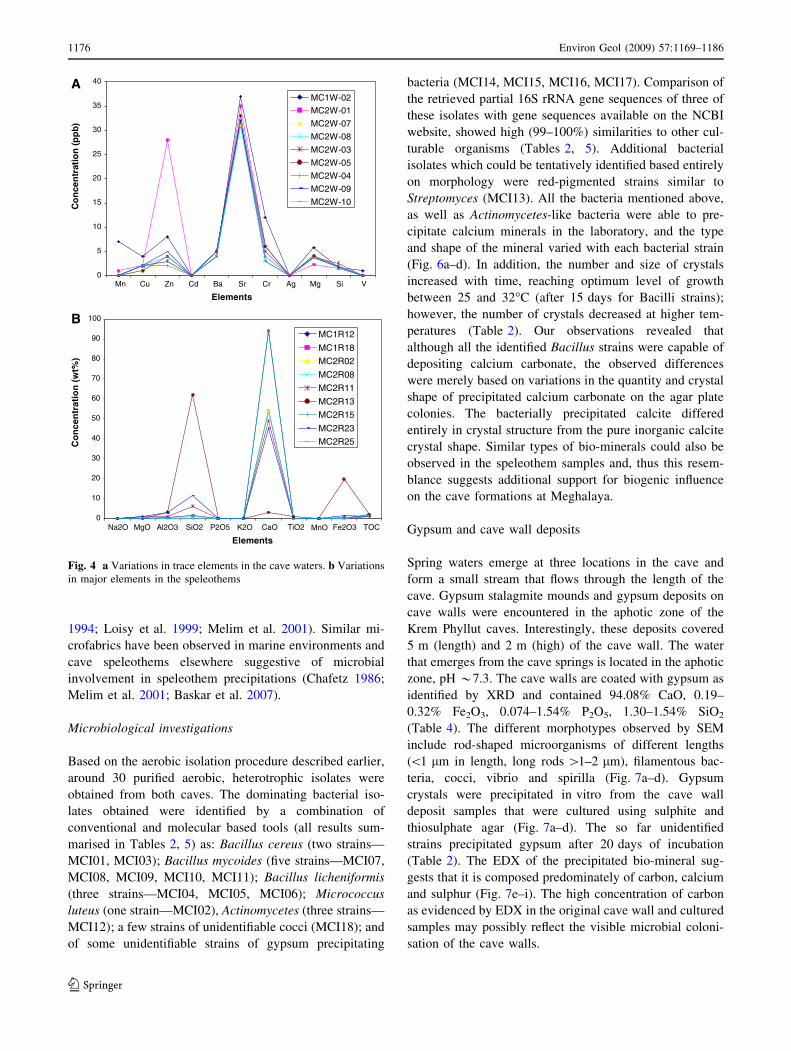
Electron microscopic observations
SEM revealed most strikingly a significant number of
microcrystalline needle fiber crystals, large micro-rod cal-
cites, dendritic calcite crystals, nano scale to microscopic
sized calcite crystals (Fig. 5a, b). All these structures may
possibly have been induced by the metabolic activities of
the associated bacterial communities. The presence of
needle, fibrous, micro-rod calcites, highly weathered and
disintegrated crystals of calcite, organic inclusions and
lithified structures that resemble stromatolitic microbial
mats indicate that they may have been formed by microbial
activities (Melim et al. 2001; Northup et al. 2000; Chafetz
1986; Northup and Lavoie 2001). The classification of
micro-rod calcite has been earlier reported, and its origin
has been attributed to rapid precipitation at high super-
saturation states during evaporation in soils or to calcifi-
cation of bacilliform bacteria (Loisy et al. 1999). The
present study also revealed the presence of micro-rods and
can be classified as follows: microcrystalline needle fiber
crystals, 0.1 m wide, 1–2 lm long; large needle fibers,
2–20 lm wide (Fig. 5a–c), 50–100 lm long; calcified fil-
aments 5–50 lm long (Fig. 5c), spiky calcite, rounded
balls of calcite, a number of bacterial ligaments, filaments
in the speleothems (Fig. 5d, e) (Verrecchia and Verrecchia
Fig. 3 a, b Stromatolitic
laminations. c Clotted chocolate
brown blebs of peloidal material
Table 3 Water analysis (ICP-MS), East Khasi Hill caves, Meghalaya
S.no. Mn Cu Zn Cd Ba Sr Cr Ag Mg Si V
MC1W-02 7 4 8 \1 4 37 12 0 5.8 1.7 1
MC2W-01 1 2 28 \1 4 35 6 0 2.3 1.5 \1
MC2W-07 \1 1 3 \1 4 31 6 \1 3.7 2.1 0
MC2W-08 \1 2 3 \1 4 31 6 \1 3.9 1.7 0
MC2W-03 \1 2 3 \1 5 32 5 0 3.7 2.6 0
MC2W-05 \1 1 4 \1 5 33 6 0 4.1 1.9 0
MC2W-04 \1 2 2 \1 4 32 4 0 3.8 1.7 0
MC2W-09 \1 2 5 \1 5 32 3 0 3.7 1.9 0
MC2W-10 \1 2 4 \1 5 31 3 0 3.7 1.9 0
Values are in ppb. Dripping waters MC1W-02, MC2W-01. Pool
waters MC2W-03, MC2W-04, MC2W-05, MC2W-07, MC2W-08.
Spring waters MC2W-09, MC2W-10
Environ Geol (2009) 57:1169–1186 1175
123
1994; Loisy et al. 1999; Melim et al. 2001). Similar mi-
crofabrics have been observed in marine environments and
cave speleothems elsewhere suggestive of microbial
involvement in speleothem precipitations (Chafetz 1986;
Melim et al. 2001; Baskar et al. 2007).
Microbiological investigations
Based on the aerobic isolation procedure described earlier,
around 30 purified aerobic, heterotrophic isolates were
obtained from both caves. The dominating bacterial iso-
lates obtained were identified by a combination of
conventional and molecular based tools (all results sum-
marised in Tables 2, 5) as: Bacillus cereus (two strains—
MCI01, MCI03); Bacillus mycoides (five strains—MCI07,
MCI08, MCI09, MCI10, MCI11); Bacillus licheniformis
(three strains—MCI04, MCI05, MCI06); Micrococcus
luteus (one strain—MCI02), Actinomycetes (three strains—
MCI12); a few strains of unidentifiable cocci (MCI18); and
of some unidentifiable strains of gypsum precipitating
bacteria (MCI14, MCI15, MCI16, MCI17). Comparison of
the retrieved partial 16S rRNA gene sequences of three of
these isolates with gene sequences available on the NCBI
website, showed high (99–100%) similarities to other cul-
turable organisms (Tables 2, 5). Additional bacterial
isolates which could be tentatively identified based entirely
on morphology were red-pigmented strains similar to
Streptomyces (MCI13). All the bacteria mentioned above,
as well as Actinomycetes-like bacteria were able to pre-
cipitate calcium minerals in the laboratory, and the type
and shape of the mineral varied with each bacterial strain
(Fig. 6a–d). In addition, the number and size of crystals
increased with time, reaching optimum level of growth
between 25 and 32�C (after 15 days for Bacilli strains);
however, the number of crystals decreased at higher tem-
peratures (Table 2). Our observations revealed that
although all the identified Bacillus strains were capable of
depositing calcium carbonate, the observed differences
were merely based on variations in the quantity and crystal
shape of precipitated calcium carbonate on the agar plate
colonies. The bacterially precipitated calcite differed
entirely in crystal structure from the pure inorganic calcite
crystal shape. Similar types of bio-minerals could also be
observed in the speleothem samples and, thus this resem-
blance suggests additional support for biogenic influence
on the cave formations at Meghalaya.
Gypsum and cave wall deposits
Spring waters emerge at three locations in the cave and
form a small stream that flows through the length of the
cave. Gypsum stalagmite mounds and gypsum deposits on
cave walls were encountered in the aphotic zone of the
Krem Phyllut caves. Interestingly, these deposits covered
5 m (length) and 2 m (high) of the cave wall. The water
that emerges from the cave springs is located in the aphotic
zone, pH *7.3. The cave walls are coated with gypsum as
identified by XRD and contained 94.08% CaO, 0.19–
0.32% Fe2O3, 0.074–1.54% P2O5, 1.30–1.54% SiO2
(Table 4). The different morphotypes observed by SEM
include rod-shaped microorganisms of different lengths
(\1 lm in length, long rods [1–2 lm), filamentous bac-
teria, cocci, vibrio and spirilla (Fig. 7a–d). Gypsum
crystals were precipitated in vitro from the cave wall
deposit samples that were cultured using sulphite and
thiosulphate agar (Fig. 7a–d). The so far unidentified
strains precipitated gypsum after 20 days of incubation
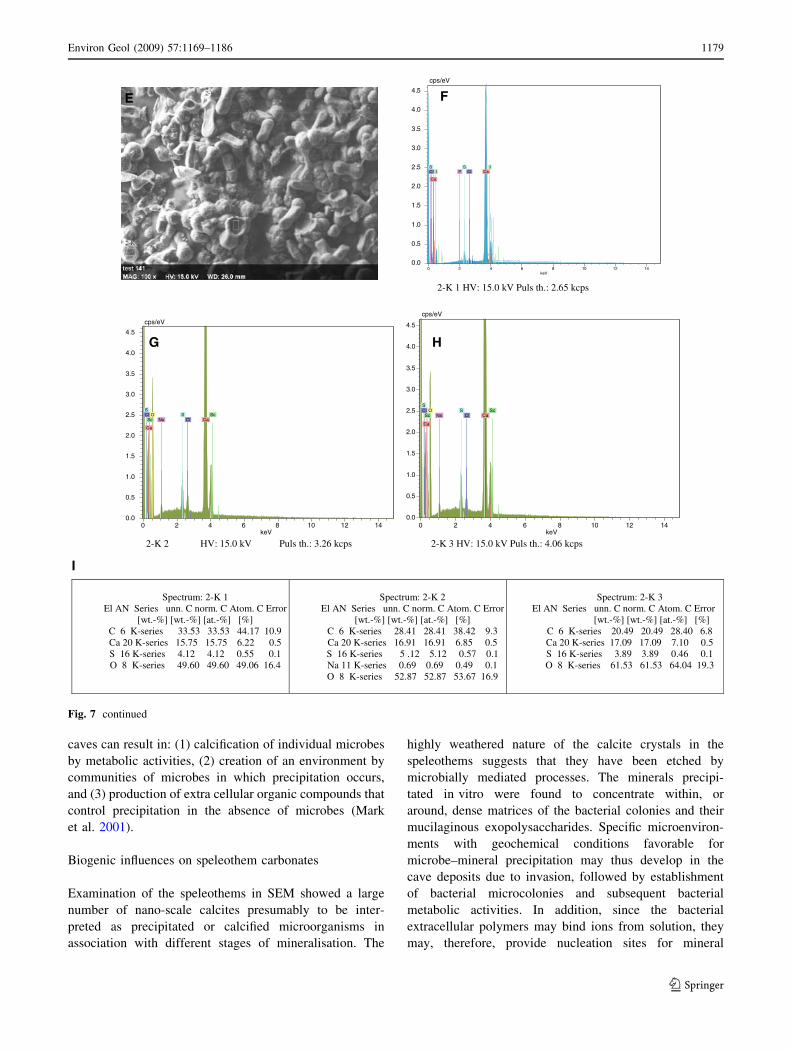
(Table 2). The EDX of the precipitated bio-mineral sug-
gests that it is composed predominately of carbon, calcium
and sulphur (Fig. 7e–i). The high concentration of carbon
as evidenced by EDX in the original cave wall and cultured
samples may possibly reflect the visible microbial coloni-
sation of the cave walls.
0
5
10
15
20
25
30
35
40
Mn
Elements
Co
nce
ntr
atio
n (p
pb)
MC1W-02
MC2W-01
MC2W-07
MC2W-08
MC2W-03
MC2W-05
MC2W-04
MC2W-09
MC2W-10
0
10
20
30
40
50
60
70
80
90
100
Na2O
Elements
Co
nce
ntr
atio
n (
wt%
)
MC1R12
MC1R18
MC2R02
MC2R08
MC2R11
MC2R13
MC2R15
MC2R23
MC2R25
VSiMgAgCrSrBaCdZnCu
TOCFe2O3MnOTiO2CaOK2OP2O5SiO2Al2O3MgO
A
B
Fig. 4 a Variations in trace elements in the cave waters. b Variations
in major elements in the speleothems
1176 Environ Geol (2009) 57:1169–1186
123
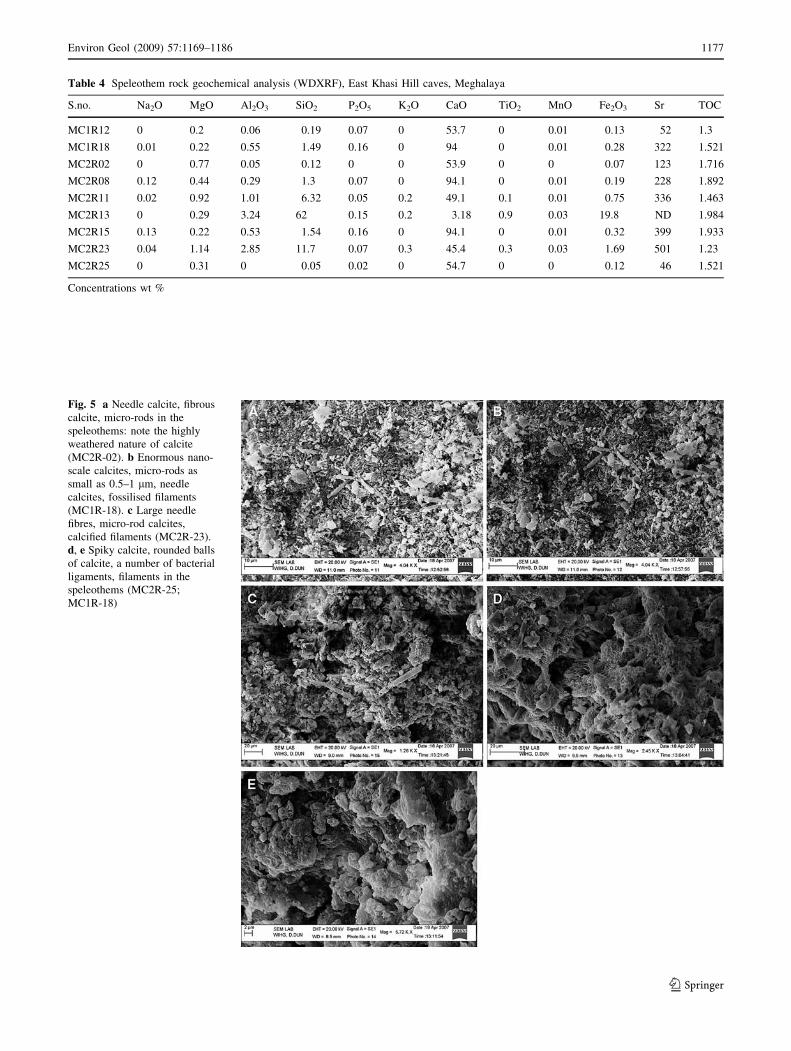
Table 4 Speleothem rock geochemical analysis (WDXRF), East Khasi Hill caves, Meghalaya
S.no. Na2O MgO Al2O3 SiO2 P2O5 K2O CaO TiO2 MnO Fe2O3 Sr TOC
MC1R12 0 0.2 0.06 0.19 0.07 0 53.7 0 0.01 0.13 52 1.3
MC1R18 0.01 0.22 0.55 1.49 0.16 0 94 0 0.01 0.28 322 1.521
MC2R02 0 0.77 0.05 0.12 0 0 53.9 0 0 0.07 123 1.716
MC2R08 0.12 0.44 0.29 1.3 0.07 0 94.1 0 0.01 0.19 228 1.892
MC2R11 0.02 0.92 1.01 6.32 0.05 0.2 49.1 0.1 0.01 0.75 336 1.463
MC2R13 0 0.29 3.24 62 0.15 0.2 3.18 0.9 0.03 19.8 ND 1.984
MC2R15 0.13 0.22 0.53 1.54 0.16 0 94.1 0 0.01 0.32 399 1.933
MC2R23 0.04 1.14 2.85 11.7 0.07 0.3 45.4 0.3 0.03 1.69 501 1.23
MC2R25 0 0.31 0 0.05 0.02 0 54.7 0 0 0.12 46 1.521
Concentrations wt %
Fig. 5 a Needle calcite, fibrous
calcite, micro-rods in the
speleothems: note the highly
weathered nature of calcite
(MC2R-02). b Enormous nano-
scale calcites, micro-rods as
small as 0.5–1 lm, needle
calcites, fossilised filaments
(MC1R-18). c Large needle
fibres, micro-rod calcites,
calcified filaments (MC2R-23).
d, e Spiky calcite, rounded balls
of calcite, a number of bacterial
ligaments, filaments in the
speleothems (MC2R-25;
MC1R-18)
Environ Geol (2009) 57:1169–1186 1177
123
Postulation of the biogenic role of microbes on the cave
formations in Meghalaya
The geological processes that are mediated by microbial
metabolism appear to be fundamentally related to the dis-
tribution of microbes throughout the cave (Jones 2001). It
is generally assumed that metabolic diversity is a reflection
of the different electron flow paths which are employed by
various microorganisms. Mineral surfaces provide periodic
arrays of reactive sites that absorb protons, water, metal
ions and organic molecules. Recent studies on stromatolite
forming microbial mats indicate that microbial activity in
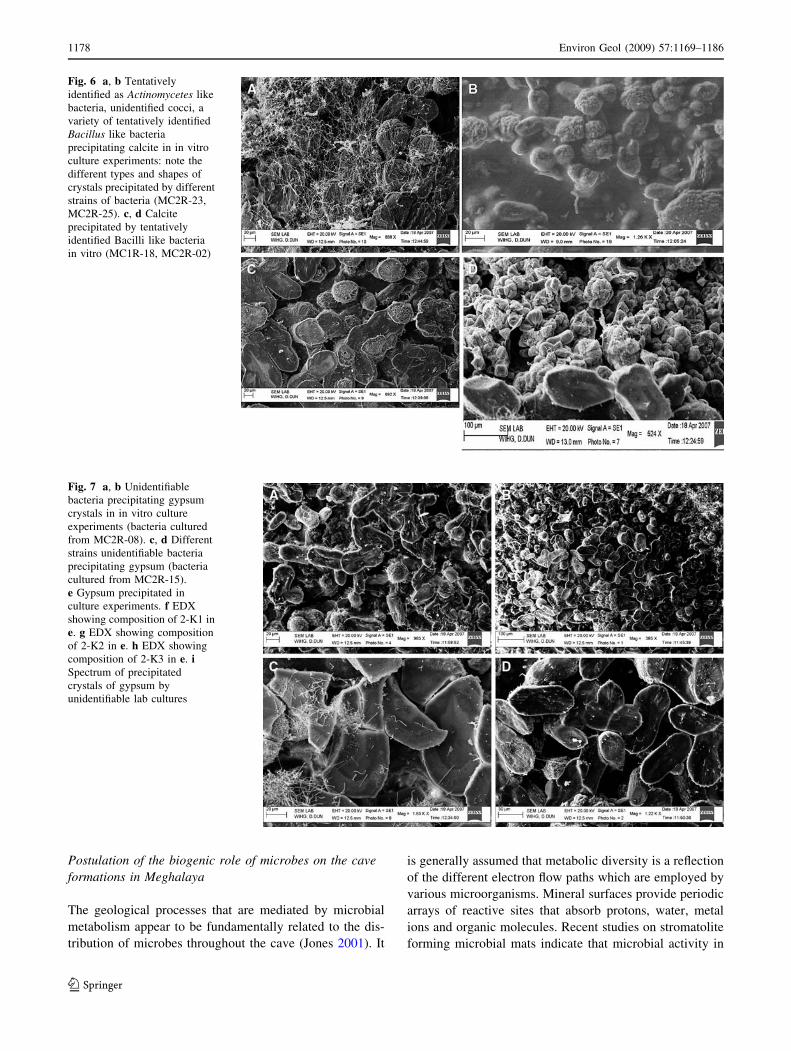
Fig. 6 a, b Tentatively
identified as Actinomycetes like
bacteria, unidentified cocci, a
variety of tentatively identified
Bacillus like bacteria
precipitating calcite in in vitro
culture experiments: note the
different types and shapes of
crystals precipitated by different
strains of bacteria (MC2R-23,
MC2R-25). c, d Calcite
precipitated by tentatively
identified Bacilli like bacteria
in vitro (MC1R-18, MC2R-02)
Fig. 7 a, b Unidentifiable
bacteria precipitating gypsum
crystals in in vitro culture
experiments (bacteria cultured
from MC2R-08). c, d Different
strains unidentifiable bacteria
precipitating gypsum (bacteria
cultured from MC2R-15).
e Gypsum precipitated in
culture experiments. f EDX
showing composition of 2-K1 in
e. g EDX showing composition
of 2-K2 in e. h EDX showing
composition of 2-K3 in e. iSpectrum of precipitated
crystals of gypsum by
unidentifiable lab cultures
1178 Environ Geol (2009) 57:1169–1186
123
caves can result in: (1) calcification of individual microbes
by metabolic activities, (2) creation of an environment by
communities of microbes in which precipitation occurs,
and (3) production of extra cellular organic compounds that
control precipitation in the absence of microbes (Mark
et al. 2001).
Biogenic influences on speleothem carbonates
Examination of the speleothems in SEM showed a large
number of nano-scale calcites presumably to be inter-
preted as precipitated or calcified microorganisms in
association with different stages of mineralisation. The
highly weathered nature of the calcite crystals in the
speleothems suggests that they have been etched by
microbially mediated processes. The minerals precipi-
tated in vitro were found to concentrate within, or
around, dense matrices of the bacterial colonies and their
mucilaginous exopolysaccharides. Specific microenviron-
ments with geochemical conditions favorable for
microbe–mineral precipitation may thus develop in the
cave deposits due to invasion, followed by establishment
of bacterial microcolonies and subsequent bacterial
metabolic activities. In addition, since the bacterial
extracellular polymers may bind ions from solution, they
may, therefore, provide nucleation sites for mineral
2-K 1 HV: 15.0 kV Puls th.: 2.65 kcps
2-K 2 HV: 15.0 kV Puls th.: 3.26 kcps 2-K 3 HV: 15.0 kV Puls th.: 4.06 kcps
Spectrum: 2-K 1 El AN Series unn. C norm. C Atom. C Error
[wt.-%] [wt.-%] [at.-%] [%] C 6 K-series 33.53 33.53 44.17 10.9 Ca 20 K-series 15.75 15.75 6.22 0.5 S 16 K-series 4.12 4.12 0.55 0.1 O 8 K-series 49.60 49.60 49.06 16.4
Spectrum: 2-K 2 El AN Series unn. C norm. C Atom. C Error
[wt.-%] [wt.-%] [at.-%] [%] C 6 K-series 28.41 28.41 38.42 9.3 Ca 20 K-series 16.91 16.91 6.85 0.5 S 16 K-series 5 .12 5.12 0.57 0.1 Na 11 K-series 0.69 0.69 0.49 0.1 O 8 K-series 52.87 52.87 53.67 16.9
Spectrum: 2-K 3 El AN Series unn. C norm. C Atom. C Error [wt.-%] [wt.-%] [at.-%] [%]
C 6 K-series 20.49 20.49 28.40 6.8 Ca 20 K-series 17.09 17.09 7.10 0.5 S 16 K-series 3.89 3.89 0.46 0.1 O 8 K-series 61.53 61.53 64.04 19.3
0 2 4 6 8 10 12 14keV
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
cps/eV
Ca
Ca
Sc Sc Cl
Cl S S
Na O
0 2 4 6 8 10 12 14keV
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
cps/eV
Ca
Ca
Sc Sc Cl
Cl S S
Na O
0 2 4 6 8 10 12 14keV
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
cps/eV
Ca
Ca
I I Cl Cl
S S P
E F
G H
I
Fig. 7 continued
Environ Geol (2009) 57:1169–1186 1179
123

crystallisation and growth. Bacteria may also act as
highly reactive geochemical interfaces (Beveridge1989;
Fein et al. 1997) and their extracellular polymers are
especially effective at binding ions from solution and
serving as nucleation surfaces for mineral formation
(Ferris et al. 1987; Beveridge 1989; Fortin et al. 1997).
Their metabolic activities can, therefore, induce localised
conditions that are favorable for mineral precipitation
(Thompson and Ferris 1990; Merz 1992; Fortin et al.
1997; Barker and Banfield 1998). Furthermore, it has
now become established that also biologically produced
exopolysaccharides (EPS) influence calcite precipitation
rates considerably.
Bacterially induced precipitation of calcium carbonate,
the so-called carbonatogenesis (Kowalchuk et al. 1997),
has drawn much attention in recent decades because of its
numerous implications. For example, certain strains of
Bacillus and Actinomycetes have been reported to precip-
itate calcium minerals (Cacchio et al. 2003; Rivadeneyra
et al. 1993; Baskar et al. 2006). The strain Bacillus cereus
is used as a conservation treatment for ornamental stone
(Kowalchuk et al. 1997), and the strain Bacillus mycoides
has been reported to be closely associated with carbonate
speleothems (Ehrlich 1996).
Interestingly, our isolation efforts yielded indeed some
strains (MCI01, MCI03—Bacillus cereus) that showed
nearly 100% identities to some different Bacillus strains
with at least some of the above-mentioned capacities.
Further physiological characterisation of some of the
metabolic capabilities of these strains, showed that strains
MCI01, MCI03, MCI04, MCI05, MCI06, MCI07, MCI08,
MCI09, MCI10, MCI11 are facultatively anaerobic, pro-
duce acid, follow the ammonification and nitrate reduction
metabolic pathway (Table 5) and are able to precipitate
calcite in vitro. This is a heterotrophic pathway that
involves the ammonification of amino acids and nitrate
reduction, and is often observed in geological sediments
(Fujita et al. 2000). These involve the production of met-
abolic CO2 and ammonia (NH3) which in the presence of
calcium ions lead to precipitation (Hammes and Verstraete
2002). Interestingly, it has indeed been reported that
Bacillus cereus precipitates calcite through the ammonifi-
cation and nitrate reduction metabolic pathway (Castanier
et al. 1999).
Two of our other isolates were identified as Bacillus
licheniformis and Micrococcus luteus. Since it has been
reported that Bacillus licheniformis is facultatively anaer-
obic and also reduces nitrate with/without N2 gas
production, these strains could, therefore, possibly also
precipitate calcite through the ammonification and nitrate
reduction metabolic pathway (Castanier et al. 1999).
Micrococcus luteus is a strict aerobe, and oxidizes carbo-
hydrates into CO2 and water and produces the enzyme
catalase. This CO2 production helps in the mediation of
carbonate precipitation.
Ten of the characterised isolates (MCI01, MCI03,
MCI04, MCI05, MCI06, MCI07, MCI08, MCI09, MCI10,
MCI11) may through the ammonification and nitrate
reduction metabolic pathways (Hammes and Verstraete
2002), induce an overall pH increase, which can in turn
shift the bicarbonate equilibrium. This, results in the for-
mation of carbonate ions which, in the presence of soluble
calcium ions, precipitate CaCO3, as summarised by the
reactions listed below:
CO(NH2Þ2 þ H2O! CO2 þ NH3
2NH3 þ CO2 þ H2O! 2NHþ4 þ CO2�3
HCO�3 þ Hþ þ 2NHþ4 þ OH� $ CO2�3 þ 2NHþ4 þ 2H2O
CO2�3 þ Ca2þ $ CaCO3:
As the above mentioned reactions indicate, microbial
carbonate precipitation always depend on metabolic CO2
production/CO2 consumption and the relative insolubility
of calcium carbonate, and their metabolic activities are
proven to participate at carbonate mineral precipitation
(Ehrlich 2002).
Although our aerobic isolation efforts on nutrient-rich
media have probably only retrieved a minor and possibly
even biased fraction of the total existing prokaryotic
diversity in the caves investigated in this study, we have
nevertheless obtained some isolates that seem to be at least
theoretically able to participate at precipitation reactions
that may take place in the cave. Thus, together with our
petrographic observations and SEM studies, these obser-
vations seem to add additional support to the generally
formed hypothesis about the biogenic influence of the
speleothem genesis.
Biogenic influence on gypsum and cave wall deposits
Microbes may play an active part in changing the nature of
limestone rocks, both by dissolving it as well as by con-
tributing to the creation of a range of deposits on the cave
walls. The amount of light entering the cave and the type
of microbe inhabiting each part of the cave system
may, however, differ. For example, the microbial biota
inhabiting totally dark zones are most likely mainly
chemolithotrophs (Northup et al. 1997). Since we did not
attempt to isolate chemolithotrophs in the present study, we
were not able to identify presumably chemolithotrophic
CaSO4 precipitating bacteria in the complete aphotic zone
where gypsum is precipitated on the cave walls. However,
the heterotrophic isolates that we obtained (MCI07,
MCI08, MCI09, MCI10, MCI11—one of the dominant
species in the microbial community obtained by the
1180 Environ Geol (2009) 57:1169–1186
123
Ta
ble
5M
orp
ho
log
ical
,p
hy
sio
log
ical
and
bio
chem
ical
test
sfo
rid
enti
fica
tio
no
fb
acte
rial
stra
ins
Inte
rnal
stra
in
nam
e
MC
I01
MC
I02
MC
I03
MC
I04
MC
I05
MC
I06
MC
I07
MC
I08
MC
I09
MC
I10
MC
I11
Str
ain
nam
eS
trai
n2
Str
ain
4S
trai
n8
Str
ain
5S
trai
n1
0S
trai
n1
4S
trai
n1
3S
trai
n1
8S
trai
n1
1S
trai
n7
Str
ain
3
MT
CC
nu
mb
erM
TC
C8
77
6M
TC
C8
77
7–
aN
AN
AN
AN
AN
AN
A
Acc
essi
on
nu
mb
er
EU
49
59
83
EU
49
59
84
EU
49
59
85
NA
NA
NA
NA
NA
NA
NA
NA
Co
lon
ym
orp
ho
log
y
Co
nfi
gu
rati
on
Cir
cula
rC
ircu
lar
Cir
cula
rR
ou
nd
Ro
un
dR
ou
nd
Ro
un
dR
ou
nd
Ro
un
dR
ou
nd
Ro
un
d
Mar
gin
Rh
izo
idE
nti
reR
hiz
oid
Wav
yW
avy
Wav
yW
avy
Wav
yW
avy
Wav
yW
avy
Ele
vat
ion
Fla
tC
on
vex
Fla
tC
on
vex
Co
nv
exC
on
vex
Co
nv
exC
on
vex
Co
nv
exC
on
vex
Co
nv
ex
Su
rfac
eS
mo
oth
and
shin
y
Pal
eS
mo
oth
and
shin
y
Ro
ug
hR
ou
gh
Ro
ug
hR
ou
gh
Ro
ug
hR
ou
gh
Ro
ug
hR
ou
gh
Pig
men
t–
Pal
ey
ello
wis
h
crea
m
–C
ream
Cre
amC
ream
Red
dis
hR
edd
ish
Red
dis
hR
edd
ish
Red
dis
h
Op
acit
yO
paq
ue
Op
aqu
eO
paq
ue
Op
aqu
eO
paq
ue
Op
aqu
eO
paq
ue
Op
aqu
eO
paq
ue
Op
aqu
eO
paq
ue
Gra
ms
reac
tio
n+
++
++
++
++
++
Cel
lsh
ape
Ple
om
orp
hic
rod
s
Co
cci
Ro
ds
Ro
ds
Ro
ds
Ro
ds
Ro
ds
Ro
ds
Ro
ds
Ro
ds
Ro
ds
Siz
e(l
m)
Len
gth
4–
5,
wid
th[
1
Dia
met
er1
Len
gth
4–
5,
wid
th[
1
Lo
ng
Lo
ng
Lo
ng
Co
nti
nu
ou
s,
lon
g
Co
nti
nu
ou
s,
lon
g
Co
nti
nu
ou
s,
lon
g
Co
nti
nu
ou
s,
lon
g
Co
nti
nu
ou
s,
lon
g
Arr
ang
emen
tN
ot
reg
ula
rS
mal
lcl
ust
ers,
tetr
ads
and
pai
rs
No
tre
gu
lar
Sin
gle
Sin
gle
Sin
gle
Fil
amen
tsF
ilam
ents
Fil
amen
tsF
ilam
ents
Fil
amen
ts
Sp
ore
s
En
do
spo
res
+-
++
++
++
++
+
Sh
ape
Ov
alO
val
Ov
alO
val
Ov
alO
val
Ov
alO
val
Ov
alO
val
Ov
al
Sp
ora
ng
ia
bu
lgin
g
--
--
--
--
--
-
Mo
tili
ty+
-+
++
++
++
++
Ph
ysio
log
ica
lte
sts
Gro
wth
atte
mp
erat
ure
s(�
C)
8-
--
--
--
--
--
15
--
-+
++
++
++
+
20
++
++
++
++
++
+
25
++
++
++
++
++
+
37
++
++
++
++
++
+
42
++
++
++
++
++
+
52
--
-+
++
--
--
-
Environ Geol (2009) 57:1169–1186 1181
123

Ta
ble
5co
nti
nu
ed
Inte
rnal
stra
in
nam
e
MC
I01
MC
I02
MC
I03
MC
I04
MC
I05
MC
I06
MC
I07
MC
I08
MC
I09
MC
I10
MC
I11
Str
ain
nam
eS
trai
n2
Str
ain
4S
trai
n8
Str
ain
5S
trai
n1
0S
trai
n1
4S
trai
n1
3S
trai
n1
8S
trai
n1
1S
trai
n7
Str
ain
3
Gro
wth
atN
aCl
(%)
2+
++
++
++
++
++
5+
++
++
++
++
++
7+
++
++
++
++
+
10
++
+-
--
--
Gro
wth
atp
H
4-
--
++
++
++
++
5+
-+
++
++
++
++
6+
-+
++
++
++
++
7+
++
++
++
++
++
8+
++
++
++
++
++
9+
++
++
++
++
++
Fac
ult
ativ
ely
An
aero
bic
+W
eak
++
++
++
++
+
Bio
chem
ica
lte
sts
Gro
wth
on
McK
on
key
agar
--
--
--
--
--
-
Ind
ole
test
--
--
--
--
--
-
Met
hy
lte
st+
-+
++
++
++
++
Vo
ges
Pro
skau
er
test
--
-+
++
++
++
+
Cit
rate
uti
lisa
tio
n
--
-+
++
--
--
-
H2S
pro
du
ctio
n-
--
--
-+
++
++
Cas
ein
hy
dro
lysi
s
+W
eak
+-
--
++
++
+
Esc
uli
n
hy
dro
lysi
s
+-
+
Gel
atin
hy
dro
lysi
s
++
++
++
++
++
+
Sta
rch
hy
dro
lysi
s
+-
++
++
++
++
+
Ure
ah
yd
roly
sis
--
--
--
--
--
-
Cat
alas
ete
st-
--
++
++
++
++
1182 Environ Geol (2009) 57:1169–1186
123

Ta
ble
5co
nti
nu
ed
Inte
rnal
stra
in
nam
e
MC
I01
MC
I02
MC
I03
MC
I04
MC
I05
MC
I06
MC
I07
MC
I08
MC
I09
MC
I10
MC
I11
Str
ain
nam
eS
trai
n2
Str
ain
4S
trai
n8
Str
ain
5S
trai
n1
0S
trai
n1
4S
trai
n1
3S
trai
n1
8S
trai
n1
1S
trai
n7
Str
ain
3
Ox
idas
ete
st+
-+
++
++
++
++
Nit
rate
red
uct
ion
+-
++
++
++
++
+
Arg
inin
e
dih
yd
rola
se
+-
++
++
++
++
+
Orn
ith
ine
dec
arb
ox
yla
se
+-
++
++
--
--
-
Ly
sin
e
dec
arb
ox
yla
se
+-
+-
--
--
--
-
Tw
een
20
hy
dro
lysi
s
No
gro
wth
No
gro
wth
No
gro
wth
No
gro
wth
No
gro
wth
No
gro
wth
No
gro
wth
No
gro
wth
No
gro
wth
No
gro
wth
No
gro
wth
Tw
een
40
hy
dro
lysi
s
++
+
Tw
een
60
hy
dro
lysi
s
++
+
Tw
een
80
hy
dro
lysi
s
-+
++
++
Aci
dfr
om
glu
cose
+-
+
Aci
dfr
om
lact
ose
+-
+
Gas
fro
m
glu
cose
--
--
--
--
--
-
Ph
osp
hat
ase
+-
+
Ty
rosi
ne
--
-
O/F
test
--
--
--
--
--
-
ON
PG
--
-
16
SrR
NA
gen
e
sim
ilar
ity
10
0%
to
Ba
cill
us
cere
us
99
%to
Mic
roco
ccu
slu
teu
s
99
%to
Ba
cill
us
cere
us
NA
NA
NA
NA
NA
NA
NA
NA
Bac
teri
al
no
men
clat
ure
Ba
cill
us
cere
usb
Mic
roco
ccu
slu
teu
sbB
aci
llu
sce
reu
sbB
aci
llu
sli
chen
ifo
rmis
cB
aci
llu
sli
chen
ifo
rmis
cB
aci
llu
sli
chen
ifo
rmis
cB
aci
llu
sm
yco
ides
cB
aci
llu
sm
yco
ides
cB
aci
llu
sm
yco
ides
cB
aci
llu
sm
yco
ides
cB
aci
llu
sm
yco
ides
c
NA
no
tav
aila
ble
aS
imil
arto
stra
in2
sin
cest
rain
2,
8ar
esi
mil
arin
all
pro
per
ties
,in
clu
din
g1
00
%m
atch
inth
eir
par
tial
seq
uen
ces,
MT
CC
dep
osi
ted
stra
in2
on
lyan
dd
idn
ot
giv
est
rain
8an
MT
CC
nu
mb
erb
Iden
tifi
cati
on
by
ph
eno
typ
ical
char
acte
rist
ics
and
16
SrR
NA
gen
ec
Iden
tifi
cati
on
by
ph
eno
typ
ical
char
acte
rist
ics
Environ Geol (2009) 57:1169–1186 1183
123

traditional aerobic isolation effort), showed high similarity
to Bacillus mycoides, which has been shown to be able to
produce H2S (Tables 2, 5). Thus microbial H2S oxidation
to sulphuric acid followed by subsequent reaction of the
sulphuric acid with limestone to precipitate gypsum could
indeed take place in the cave, as suggested by the following
chemical reactions:
H2Sþ O2 $ Hþ þ HSO�4 $ 2Hþ þ SO2�4
2Hþ þ SO2�4 þ CaCO3 $ Ca2þ þ SO2�
4 þ H2Oþ CO2
Ca2þ þ SO2�4 þ H2O! CaSO4 � 2H2O:
Furthermore, as previously mentioned, we observed by
SEM also unidentifiable strains of presumably CaSO4-
precipitating bacteria; this group could thus contribute as
an additional possible factor for gypsum precipitates. Thus,
biological activity can significantly accelerate the oxidation
of H2S, causing the production of H2SO4 as a by-product of
their metabolism. Therefore, cave gypsum precipitates may
also in some cases be considered as a consequence of
bacterial activity (Galdenzi et al. 1999). Pyrite FeS2, which
was identified in our pool sediments and was interspersed
with the limestone deposits, could also serve as another
oxidation source to sulphuric acid by the local bacterial
community, which in turn then may react with limestone to
form gypsum.
Generally, sulphur oxidizers retrieve their energy by
oxidising sulphur and acquiring their carbon from the air,
thus converting carbon dioxide into organic material. In the
dark aphotic cave ecosystems, bacteria, tentatively postu-
lated to have a ‘‘chemosynthetic’’ metabolic lifestyle may
thus possibly form the basis of the food chain in this habitat.
We anticipate that a wide range of different bacterial groups
may be found on the cave walls and springs in the dark,
pitch black aphotic zones, such as sulphate-reducing bac-
teria and sulphur-oxidising bacteria, with species such as
Thiothrix, Thiovulum, Desulfovibrio. These organisms may
utilize sulphate, thiosulphate (S2O3-), sulphite (SO3
-), or
other reducible sulphur-containing ions as terminal electron
acceptors in their respiratory metabolism. Based on these
metabolic activities, sulphur-containing ions can, therefore,
be reduced to hydrogen sulphide. The hydrogen sulphide
thereafter mixes with oxygen to form sulphuric acid, which
dissolves the limestone rock, leaving behind a deposit of
gypsum. A number of bacteria are known to function in the
transition zone between aerobic and anaerobic environ-
ments, particularly in the cave wall biofilms associated with
elemental sulphur and formation of gypsum crystals. The
sources of gypsum in caves differ depending on the specific
caves.
The proposed postulations in this study are based
on geochemical observations in the field, coupled with
laboratory experiments and drawing on the general
knowledge about cave environments and microbial activi-
ties. Based on our geochemical and initial microbiological
observations, at least some of the isolated bacteria seem to
be able to contribute to the proposed mineral precipitation
reactions, thus, confirming the role of microorganisms in
the mineral formations. However, since culture indepen-
dent molecular techniques have shown that only a minor
fraction of the total microbial diversity is culturable (see
e.g. Amann et al. 1996), future studies will be employed to
implement other tools, such as culture-independent tech-
niques to unravel further the hidden biodiversity of the
caves investigated in this study. This will expand our
knowledge considerably about the overall microbial pro-
cesses and influences in the cave, and confirm as well as
expand the basic hypotheses that we have laid in this paper.
Conclusions
In this paper, we present further evidences that support
current general hypotheses on the bacterial involvement in
speleothem cave formations, exemplified with novel
investigations of a so far non-explored cave, the Meghalaya
caves, in India. Our laboratory experiments confirm that (1)
carbonate precipitation occurs during microbial CaCO3
precipitation in a closely related group of bacteria fol-
lowing the ammonification/nitrate reduction pathway; and
(2) gypsum precipitates are formed by microbial mediated
geochemical reactions.
Several processes appear to be acting in tandem induc-
ing the precipitation of carbonates and gypsum observed in
the cave systems studied. This combination of microbio-
logical and inorganic transformation processes may explain
the biogenic involvement in the formation of the speleo-
thems and cave wall deposits in these and in other caves
systems. The observations suggest that the microbes and
their metabolic activities have definitely influenced the
formation of these cave deposits. Further studies, based on,
e.g. stable isotope chemistry and culture independent
molecular techniques will be employed to cast further light
on the mechanism and extent of the microbial role in the
speleothem genesis of the cave areas investigated in this
study, to confirm and expand the basic hypotheses laid in
this study.
Acknowledgments S. Baskar and R. Baskar thank the Wadia Insti-
tute of Himalayan Geology, Dehradun (WIHG), for laboratory and
library facilities (ICP-MS, XRD, SEM-EDX) and Institute of Micro-
bial Technology (IMTECH), Chandigarh for bacterial identification.
R. Baskar thanks the World Bank for financial assistance to attend the
FISH training programme in connection with this project at Technical
University, Munich, Germany. R. Baskar thanks UGC, New Delhi for
financial assistance in the form of Major research project.
1184 Environ Geol (2009) 57:1169–1186
123

References
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic
local alignment search tool. J Mol Biol 215:403–410
Amann R, Snaidr J, Wagner M, Ludwig W, Schleifer KH (1996)
In situ visualization of high genetic diversity in a natural
microbial community. J Bacteriol 178:3496–3500
Angert ER, Northup DE, Reysenbach AL, Peek AS, Goebel BM, Pace
NR (1998) Molecular phylogenetic analysis of a bacterial
community in Sulphur River, Parker Cave, Kentucky. Am
Mineral 83:1583–1592
Barker WW, Banfield JF (1998) Zones of chemical and physical
interaction at interfaces between microbial communities and
minerals: a model. Geomicrobiol J 15:223–244
Baskar S, Baskar R, Mauclaire L, McKenzie JA (2005) Role of
microbial community in stalctite formation, Sahastradharacaves,
Dehradun, India. Curr Sci 88:1305–1308
Baskar S, Baskar R, Mauclaire L, McKenzie JA (2006) Microbially
induced calcite precipitation by culture experiments—possible
origin for stalactites in Sahastradhara, Dehradun, India. Curr Sci
90:58–64
Baskar S, Baskar R, Kaushik A (2007) Evidences for microbial
involvement in the genesis of speleothem carbonates, Borra
Caves, Vishakapatanam, India. Curr Sci 92(3):350–355
Beveridge TJ (1989) Role of cellular design in bacterial metal
accumulation and mineralization. Annu Rev Microbiol 43:147–
171
Boquet E, Boronat A, Ramos-Cormenza A (1973) Production of
calcite (calcium carbonate) crystals by soil bacteria is a general
phenomenon. Nature 246:527–529
Cacchio P, Ercole P, Cappucio G, Lepidi A (2003) Calcium carbonate
precipitation by bacterial strains isolated from a limestone cave
and from a loamy soil. Geomicrobiol J 20(2):85–98
Castanier S, Le M’etayer-Levrel G, Perthuisot JP (1999) Ca
carbonates precipitation and limestone genesis—the microbiol-
ogist point of view. Sediment Geol 126:9–23
Castanier S, Le M’etayer-Levrel G, Perthuisot JP (2000) Bacterial
roles in the precipitation of carbonate minerals. In: Riding RE,
Awramik SM (eds) Microbial sediments. Springer, Heidelberg,
pp 32–39
Chafetz HS (1986) Marine peloids: a product of bacterially induced
precipitation of calcite. J Sediment Petrol 56:812–817
Chafetz HS, Buczynski C (1992) Bacterially induced lithification of
microbial mats. Palaios 7:277–293
Culver DC (1982) Cave life: evolution and ecology. Harvard
University press, Cambridge, p 189
Egemeier S (1981) Cave development by thermal waters. Natl Speleol
Soc Bull 43:31–51
Ehrlich HL (1996) In: Geomicrobiology, 3rd edn. Marcel Dekker,
New York, p 719
Ehrlich HL (2002) In: Geomicrobiology, 4th edn. Marcel Dekker,
New York, p 768
Elia TB (2002) Microbiology and geology: solid marriage made on
earth. ASM News 68:1 (American Society for microbiology)
Engel SE, Lee N, Porter ML, Stern AL, Bennett PC, Wagner M
(2003) Filamentous Epsilonproteobacteria dominate microbial
mats from sulfidic cave springs. Appl Environ Microbiol
69:5503
Engel AS, Porter ML, Stern LA, Quinlan S, Bennett PC (2004) Bacterial
diversity and ecosystem function of filamentous microbial mats
from aphotic (cave) sulfidic springs dominated by chemolithoauto-
trophic ‘‘Epsilonproteobacteria’’. FEMS Microbiol Ecol 51:31–53
Fein JB, Daughney CJ, Yee N, Davis TA (1997) A chemical
equilibrium model for metal adsorption onto bacterial surfaces.
Geochim Cosmochim Acta 61:3319–3328
Ferris FG, Fyfe WS, Beveridge TJ (1987) Bacteria as nucleation sites
for authigenic minerals in a metal-contaminated lake sediment.
Chem Geol 63:225–232
Fortin D, Ferris FG, Beveridge TJ (1997) Surface-mediated mineral
development by bacteria. In: Banfield JF, Nealson KH (eds)
Geomicrobiology: interactions between microbes and minerals.
Reviews in Mineralogy, vol 35. Min. Soceity of America,
Washington, DC, pp 161–180
Frey DG (1963) Limnology in North America. The University of
Wisconsin Press, Madison, p 734
Fujita Y, Ferris EG, Lawson RD, Colwell FS, Smith RW (2000)
Calcium carbonate precipitation by ureolytic subsurface bacteria.
Geomicrobiol J 17(4):305–318
Galdenzi S, Menichetti M, Sarbu S, Rossi A (1999) Frasassi caves: a
biogenic hypogean karst system? In: Audra P (ed) Proceedings
Eurpean conference Karst 99, Grands causes, Vercors, France:
Cagep, Universite de province, Etudes de Geographie physique,
travaux 1999, suppl 28, pp 101–106
Garg R, Jain KP (1995) Significance of the terminal cretaceous
calcareous nannofossil marker Micula prinsii at the Cretaceous-
Tertiary boundary in the Um Sohryngkew River section,
Meghalaya, India. Curr Sci 66(12):1012–1017
Ghosh AMN (1940) The stratigraphical position of the Cherra
sandstone, Assam. Rec GSI 75:1–19
Ghosh S, Fallick AE, Paul DK, Potts PJ (2005) Geochemistry and
origin of Neoproterozoic Granitoids of Meghalaya, Northeast
India: implications for linkage with amalgamation of Gondwana
Supercontinent. Gondwana Res 8(3):421–432
Hammes F, Verstraete W (2002) Key roles of pH and calcium
metabolism in microbial carbonate precipitation. Rev Environ
Sci Biotechnol 1:3–7
Hose LD, Palmer AN, Palmer MV, Northup DE, Boston PJ, DuChene
HR (2000) Microbiology and geochemistry in a hydrogen-
sulphide-rich karst environment. Chem Geol 169:399–423
JCPDS–ICDD. XRPD database (2000) International Centre for
Diffraction Data. 12 Campus Boulevard, Newtown Square, PA
19073–3273. Powder Diffraction File cards num, 5–586
Jones B (2001) Microbial activity in caves—a geological perspective.
Geomicrobiol J 18:345–357
Kowalchuk GJ, Stephen W, De Boer Prosser J, Embley T, Wolden-
dorp J (1997) Analysis of ammonia-oxidizing bacteria of the
beta subdivision of the class Proteobacteria in coastal sand
dunes by denaturing gradient gel electrophoresis and sequencing
of PCR-amplified 16S ribosomal DNA fragments. Appl Environ
Microbiol 63:1489–1497
Leveille RJ, Fyfe WS, Longstaffe FJ (2000) Geomicrobiology of
carbonate–silicate microbialites from Hawaiian basaltic sea
caves. Chem Geol 169:339–355
Loisy C, Verrechia EP, Dufour P (1999) Microbial origin for
pedogenic micrite associated with a carbonate paleosol (Cham-
pagne, France). Sediment Geol 126:193–204
Ludwig W, Strunk O, Westram R, Richter L, Meier H, Yadhukumar
A, Buchner A, Lai T, Steppi S, Jobb G, Forster W, Brettske I,
Gerber S, Ginhart AW, Gross O, Grumann S, Hermann S, Jost R,
Konig A, Liss T, Lußmann R, May M, Nonhoff B, Reichel B,
Strehlow R, Stamatakis A, Stuckmann N, Vilbig A, Lenke M,
Ludwig T, Bode A, Schleifer KH (2004) ARB: a software
environment for sequence data. Nucleic Acids Res 32:1363–
1371
Mark F, Martin N, Iwan S (2001) How microbial are microbial
sediments? Microbes in sedimentary geochemical processes.
Quantifying the significance of microbes in sedimentary geo-
chemical processes. In: Symposium, EUG XI–BG04
Melim LA, Shinglman KM, Boston PJ, Northup DE, Spilde MN,
Queen JM (2001) Evidence for microbial involvement in pool
Environ Geol (2009) 57:1169–1186 1185
123

finger precipitations, Hidden cave, New Mexico. Geomicrobiol J
18:311–329
Merz MUE (1992) The biology of carbonate precipitation by
cyanobacteria. Facies 26:81–102
Northup DE, Lavoie KH (2001) Geomicrobilogy of caves: a review.
Geomicrobiol J 18:199–222
Northup DE, Reysenbach AL, Pace NR (1997) Microorganisms and
speleothems. In: Hill CA, Forti P (eds) Cave minerals of the
world. National Speleological Society, Huntsville, pp 261–266
Northup DE, Dahm CN, Melim LA, Spilde MN, Crossey LJ, Lavoie
KH, Mallory LM, Boston PJ, Cunningham KI, Barns SM (2000)
Evidence for geomicrobiological interactions in Guadalupe
caves. J Cave Karst Stud 62(2):30–40
Pedley M (1992) Freshwater (phytoherm) reefs: the role of biofilms
and their bearing on marine reef cementation. Sediment Geol
79:255–274
Poulson TL, Lavoie KH (2000) The trophic basis of subsurface
ecosystems. In: Wilkens DC, Culver DC, Humphreys WF (eds)
Ecosystems of the world 30. Elsevier, Amsterdam, pp 231–249
Riding R (2000) Microbial carbonates: the geological record of
calcified bacterial–algal mats and biofilms. Sedimentology
47:179–214
Rivadeneyra MA, Delgado R, Delgado G, Del Moral A, Ferrer MR,
Ramos-Cormenza A (1993) Precipitation of carbonate by Bacillussp. isolated from saline soils. Geomicrobiol J 11:175–184
Sarbu SM, Kane TC, Kinkle BK (1996) A chemoautotrophically
based cave ecosystem. Science 272:1953–1955
Simon KS, Benfield EF, Macko SA (2003) Food web structure and the
role of epilthic biofilms in cave streams. Ecology 84:2395–2406
Thompson JB, Ferris FG (1990) Cyanobacterial precipitation of
gypsum, calcite, and magnesite from natural alkaline lake water.
Geology 18:995–998
Verrecchia EP, Verrecchia KE (1994) Needle fibre calcite: a critical
review and a proposed classification. J Sediment Res A 64:650–
664
Warthmann R, Lith YV, Vasconcelos C, McKenzie JA, Karpoff AM
(2000) Bacterially induced dolomite precipitation in anoxic
culture experiments. Geology 28:1091–1094
1186 Environ Geol (2009) 57:1169–1186
123





























