Embed Size (px)
Citation preview

[CANCER RESEARCH 32, 868-876, April 1972]
Studies on the Mechanism of Inhibition by PolyenicAntibiotics of Nucleic Acid Biosynthesis in AscitesTumor Cells
Roberto Strom, Argante Bozzi, Antonio Scioscia Santoro, Carlo Grifo, Bruno Mondovi, and Alessandro RossiFanelli
Istituto di Chimica Biologica, University of Rome and Centro di Biologica Molecolare del C. N. R., Rome 00185, Italy
SUMMARY
The polyenic antibiotic lucensomycin caused a severeinhibition of uridine or thymidine incorporation into RNA orDNA of ascites tumor cells. The inhibition appeared to berelated to the ability of the polyene to interact with the cellmembrane, the permeability of which was increased; simpleleakage of precursors or of enzymes from the cells did notfully account for the kinetics of inhibition.
Lucensomycin did not affect DNA or RNA synthesis,starting from triphosphonucleotides, by isolated nuclei. In thepresence of the polyene, the cells became permeable toexternally added nucleotides, which they could utilize forRNA or DNA synthesis. Nucleosides, even at highconcentrations, could not substitute for nucleotides.
INTRODUCTION
In previous work from this laboratory (20-22), oxygenconsumption and DNA synthesis by Novikoff hepatoma cellsand Ehrlich ascites tumor cells were inhibited, although to adifferent extent, by 2 polyenic antibiotics, filipin andlucensomycin. These findings were tentatively correlated (21)to an increase in permeability of the cell membranes caused bythe 2 polyenes, which are known to act onphospholipid-cholesterol membranes. Regenerating liver cells,which were used as "normal" (i.e., nonmalignant) controls for
Novikoff hepatoma cells, were relatively insensitive to the 2antibiotics. Lucensomycin, more selective than filipin, had noeffect, even at high concentrations, on regenerating liver cellsor on oxygen uptake by tumor cells.
This paper reports the results of experiments aimed atelucidating the mechanism by which lucensomycin inhibitsDNA and RNA synthesis in ascites tumor cells. Ascites cellswere preferred to cells from solid tumors because ascites cellswere accessible, their population is relatively homogeneous,and the cell membrane is reliably intact. Besides Ehrlich ascitestumor cells from mice, which had been the subject of ourprevious work (21), Yoshida ascites tumor cells from rats werealso studied, since they could be obtained in larger amountsfrom a small number of animals (and therefore with a lowheterogeneity of the cell population) and could be bettercompared to normal lever cells (25).
MATERIALS AND METHODS
ChemicalsEagle's minimal essential medium, Medium 109, was
obtained from Grand Island Biological Co., Grand Island, N.Y.; Hanks' balanced salts solution and fetal calf serum werefrom Microbiological Associates, Bethesda, Md.; uridine-5-3H(specific activity, 5 Ci/mmole), thymidine-methyl-3 H (specificactivity, 5 Ci/mmole), UMP-4-14C (specific activity, 16.7mCi/mmole), and UTP-4-14C (specific activity, 51 to 53
mCi/mmole) were from the Radiochemical Centre, Amersham,England; TTP-methyl-3 H (specific activity, 10 to 13
Ci/mmole) was from New England Nuclear, Boston, Mass.;UTP, ATP, GTP, and CTP were from Boehringer und Soehne,Mannheim, Germany.; TTP, dATP, dCTP, dGTP, andfluorescein diacetate were from Koch-Light Laboratories,Colnbrook, England; Nonidet P-40 was from British DrugHouses, Poole, England; PPO and POPOP were fromNuclear-Chicago Corp., Des Plaines, 111.;and Soluene was fromPackard Instrument Co., Downers Grove, 111.Other chemicalswere obtained from E. Merck, Darmstadt, Germany, or fromBritish Drug Houses.
Lucensomycin (etruscomycin) and 7V-acetyllucensomycin,in nitrogen-filled, sealed vials each containing 10 mg of thepolyene, were kindly supplied by Professor F. Arcamone andProfessor M. Ghione of Farmitalia, Milan, Italy. The vials werekept at 4°,opened just before use, and dissolved in 0.6 to 1.0ml DMSO.1
Cell and Nuclei Preparation
Yoshida ascites tumor cells and Ehrlich ascites tumor cellswere harvested 6 to 8 days after i.p. transplantation of 4 to 7million cells in male Wistar rats or Swiss mice, respectively.Cell viability was checked by the dye exclusion method, withthe use of trypan blue, Img/ml, in 0.9% NaCl solution,according to the method of Tennant (29). Onlynonhemorrhagic cell samples containing more than 90% viablecells were used. The cells were washed in the cold bysuspension in KRB and were submitted to low-speedcentrifugation (1500 rpm), resuspended in KRB, passed
Received July 12, 1971 ¡acceptedDecember 1, 1971.
'The abbreviations used are: DMSO, dimethyl sulfoxide; KRB,Krebs-Ringer bicarbonate solution supplemented with benzylpenicillin,100 i.u./ml; LDH, láclate dehydrogenase (L-lactate:NADoxidoreductase, EC 1.1.1.27).
868 CANCER RESEARCH VOL. 32
Research. on November 9, 2015. © 1972 American Association for Cancercancerres.aacrjournals.org Downloaded from

Action ofPolyenic Antibiotics on Ascites Cells
through an 80 mesh nylon net and an 18-gauge needle,counted in a Burker chamber, diluted if necessary, and kept inice until use, i.e., for not more than 5 hr.
For preparation of isolated nuclei, 2 different procedureswere used, Procedure A and Procedure B. In Procedure A,Ehrlich ascites cells, sedimented by low-speed centrifugation inthe cold, were resuspended in 10 volumes of a buffercontaining 0.25 M sucrose, 50 mM Tris, 20 mM KC1,5 mMMgCl2, and 0.2 mM CaCl2, pH 7.4 (Solution A), equilibratedfor 20 min in a pressure bomb with nitrogen at 65atmospheres, and then rapidly expelled through a needle valve.The cavitation caused by the rapid release of nitrogendissolved in the suspension was effective in disrupting theplasma membrane of the cells (26), while preserving mostnuclei in an intact state (2).
The suspension was submitted to 10 strokes in a TeflonPotter-Elvehjem homogenizer at low speed (400 rpm), to freethe nuclei from cytoplasmic residues, and then layered over anequal volume of 1 M sucrose and centrifuged at 700 X g for 30min. The pellet, resuspended in 2.4 M sucrose containing 1mM MgCl2, was centrifuged twice at 50,000 X g for 60 min inthe SW25 rotor of the Spinco Model 40 ultracentrifuge (31)and was then suspended in a buffer containing 50 mM Tris,30% (v/v) glycerol, and 10 mM mercaptoethanol, pH 7.8(Buffer B) (11). Light microscopy on either fresh preparationsor those after Papanicolaou staining showed that the finalpellet thus obtained consisted of naked nuclei, sometimes overa sparse background of nuclear debris, with no whole cellspresent.
Nuclei from Yoshida ascites cells were prepared similarly,but in order to increase the yield, it was necessary to subjectthe cells to hypotonie shock prior to cell disruption. Thesedimented cells were therefore resuspended in 8 volumes ofH20; after 20 min in the cold, 4 times concentrated SolutionA was added to regain isosmolarity, and high-pressure nitrogenwas applied as already described.
In Procedure B, the cells, suspended in 8 volumes ofSolution A, were mixed with an equal volume of solution A towhich the surface-active agent Nonidet P40 had been added.The optimal final concentration of Nonidet P-40 for maximallysis of cell plasma membranes and minimal disruption ofnuclei was 0.5% for Ehrlich ascites cells and 1% for Yoshidaascites cells. For optimal yield, the latter cells were subjectedto hypotonie shock, as indicated above, prior to addition ofNonidet. After 5 min at room temperature, centrifugationthrough 1 M sucrose and then in 2.4 M sucrose was performedas described for Procedure A.
Procedure B, as compared to Procedure A, gave higheryields of pure nuclei with low amounts of nuclear debris andpractically no cytoplasmic contamination. Nuclei prepared bythe 2 methods behaved in an identical manner as far astriphosphoribonucleotide incorporation into polynucleotidewas concerned.
Evaluation of Cell Permeability
Fluorescein Efflux. The cells were loaded with fluoresceinby incubation with fluorescein diacciate, essentially asdescribed by Celada and Rotman (4).
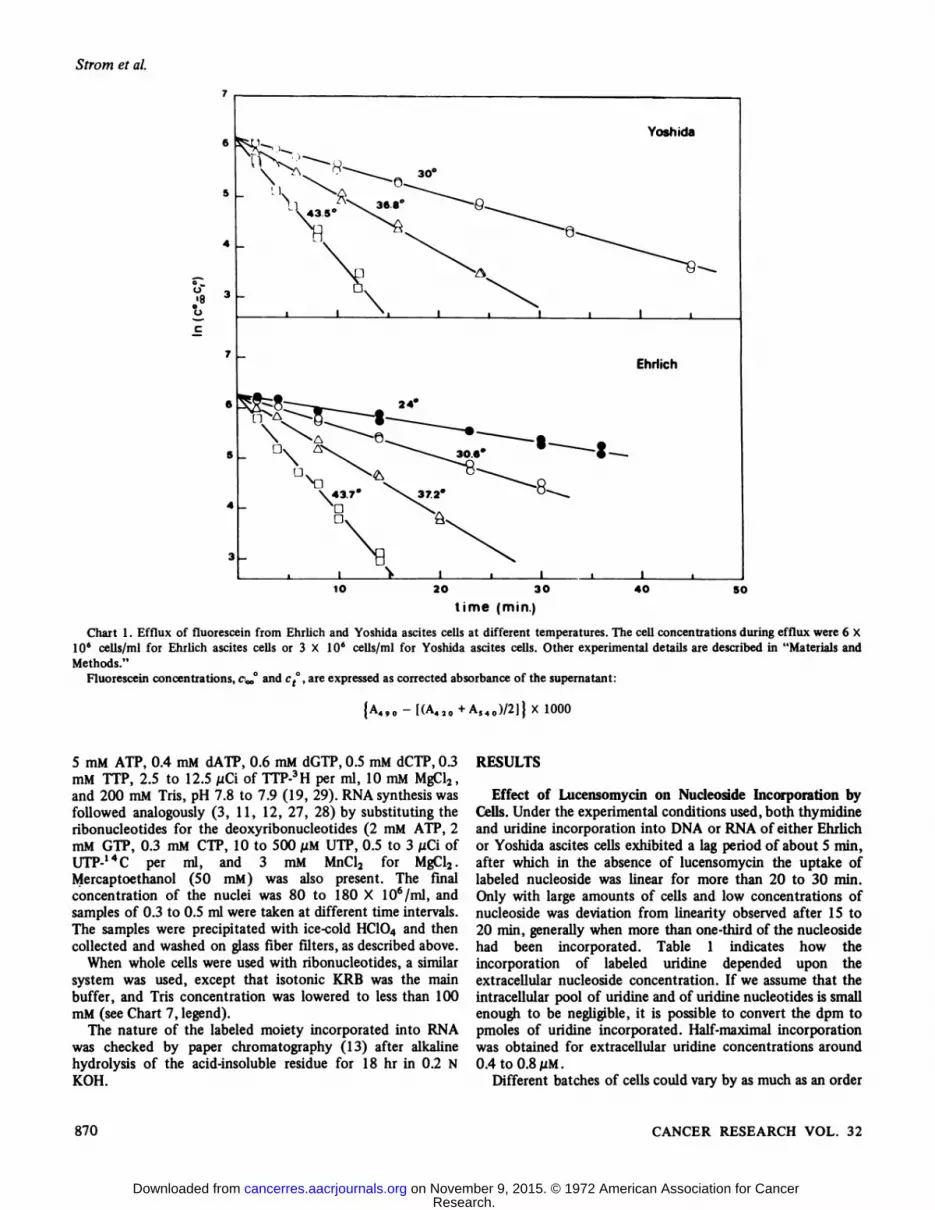
As shown by Rotman and Papermaster (24) for single cells,the concentration of fluorescein within the cells decreasesexponentially with time at rates proportional to theconcentration remaining in the cells. If ct°is the concentrationof fluorescein in the supernatant at the various times and Coo°is the concentration after at least 60 min at 45°,plotting In(e»0- ct°)versus time yields, therefore, a straight line (Chart
1). Its slope k, which is the rate constant of the exit process,allows an estimate of the diffusion coefficient of fluoresceinthroughout the cell membrane (A. Sciuscià Santoro, R. Strom,and C. Grifo. An Appraisal of Some Methods for EvaluatingCell Membrane Permeability, submitted for publication).
Leakage of Enzymes from the Cells. The amount of LDH inthe extracellular compartment was evaluated by the following,in 3-ml cuvets at 340 nm, pH 7.0, and 25°: the oxidation of 2X IO"4 M 0-NADH by 7.5 X IO"4 M pyruvate in the presence
of 50 ¡ilof the supernatant from a cell suspension in KRB.Incorporation of Labeled Nucleosides and Nucleotides into
Nucleic Acids. The cells, suspended in KRB, were added to 9volumes of KRB at 30°containing either thymidine-3H oruridine-3H, at concentrations varying between 0.2 and 6
¿iCi/mland the appropriate amount of lucensomycin inDMSO. Controls were always run with addition of DMSOalone, although the low amounts used (1 to 10 id/ml) werewithout effect. Use of of Hanks' balanced salts solution orEagle's minimal essential medium with or without fetal calf
serum did not, under our experimental conditions, modify therate of incorporation of either uridine or thymidine.
The incubation mixture was kept at 30°with moderate
shaking; repetitive samples were taken at 1- to 4-min intervalsand were mixed immediately with 7 volumes of ice-cold 0.4 MHC104. In a few experiments, sampling was begun beforeaddition of lucensomycin (for further details, see Chart 2,legend). The precipitate was then either washed bycentrifugation, defatted according to Tyner et al. (30), anddissolved in 1 ml of formamide at 140°for 5 to 8 hr, as
previously described (20, 21), or collected by filtration onWhatman GH 83 glass fiber filters, extensively washed withcold HC104, and then digested with 1 ml of Soluene at 55°for
20 min. Toluene (10.4 ml) containing 6 g PPO and 0.1 gPOPOP per liter were added (together with 8.6 ml of 95%ethanol for the formamide-dissolved samples), and the vialswere counted on a Packard Tri-Carb Model 3380 liquidscintillation counter. Correction of cpm to dpm was done byautomatic external standardization but was always controlledby a channel ratio method.
Autoradiography was performed by smearing microscopeslides with the cells from the incorporation experiments; afterfixation in ethanol:acetic acid (3:1), the slides were passedthrough cold 70% ethanol, dried, thoroughly washed with tapwater and then with distilled water, dried again, and finallycovered with L4 emulsion from Ilford Ltd., Ilford, England.After a week at room temperature in the dark, theautoradiographs were developed, with 0.45% diaminophenol(amidol) as the reducer, and fixed with sodium thiosulfate (7).Light staining with hemalum was finally performed.
DNA synthesis by isolated nuclei was studied by addition ofthe nuclear suspension, kept at 0°in Buffer B, to 4 to 6volumes of a solution, equilibrated at 30°,and containing 3 to
APRIL 1972 869
Research. on November 9, 2015. © 1972 American Association for Cancercancerres.aacrjournals.org Downloaded from
Strom et al.
30°
V;eo
Yoshida
Ehrlich
10 20 30
time (min.)40 SO
Chart l. Efflux of fluorescein from Ehrlich and Yoshida ascites cells at different temperatures. The cell concentrations during efflux were 6 X10s cells/ml for Ehrlich ascites cells or 3 X IO6 cells/ml for Yoshida ascites cells. Other experimental details are described in "Materials andMethods."
Fluorescein concentrations, c^ and ct°,are expressed as corrected absorbance of the supernatant:
{A490-[(A420+AS40)/2]}X 1000
5 mM ATP, 0.4 mM dATP, 0.6 mM dGTP, 0.5 mM dCTP, 0.3mM TTP, 2.5 to 12.5 pCi of TTP-3H per ml, 10 mM MgCl2,
and 200 mM Tris, pH 7.8 to 7.9 (19, 29). RNA synthesis wasfollowed analogously (3, 11, 12, 27, 28) by substituting theribonucleotides for the deoxyribonucleotides (2 mM ATP, 2mM GTP, 0.3 mM CTP, 10 to 500 juM UTP, 0.5 to 3 ¿/CiofUTP-14C per ml, and 3 mM MnCl2 for MgCl2.
Mercaptoethanol (50 mM) was also present. The finalconcentration of the nuclei was 80 to 180 X 106/ml, and
samples of 0.3 to 0.5 ml were taken at different time intervals.The samples were precipitated with ice-cold HC104 and thencollected and washed on glass fiber filters, as described above.
When whole cells were used with ribonucleotides, a similarsystem was used, except that isotonic KRB was the mainbuffer, and Tris concentration was lowered to less than 100mM (see Chart 7, legend).
The nature of the labeled moiety incorporated into RNAwas checked by paper chromatography (13) after alkalinehydrolysis of the acid-insoluble residue for 18 hr in 0.2 N
KOH.
RESULTS
Effect of Lucensomycin on Nucleoside Incorporation byCells. Under the experimental conditions used, both thymidineand uridine incorporation into DNA or RNA of either Ehrlichor Yoshida ascites cells exhibited a lag period of about 5 min,after which in the absence of lucensomycin the uptake oflabeled nucleoside was linear for more than 20 to 30 min.Only with large amounts of cells and low concentrations ofnucleoside was deviation from linearity observed after 15 to20 min, generally when more than one-third of the nucleosidehad been incorporated. Table 1 indicates how theincorporation of labeled uridine depended upon theextracellular nucleoside concentration. If we assume that theintracellular pool of uridine and of uridine nucleotides is smallenough to be negligible, it is possible to convert the dpm topmoles of uridine incorporated. Half-maximal incorporationwas obtained for extracellular uridine concentrations around0.4 to 0.8/xM.
Different batches of cells could vary by as much as an order
870 CANCER RESEARCH VOL. 32
Research. on November 9, 2015. © 1972 American Association for Cancercancerres.aacrjournals.org Downloaded from

of magnitude in the rate of uridine or thymidineincorporation. The values reported for the experiment given inTable 1 are, for instance, rather low, and might be higher evenby a factor of 6 with other batches of Ehrlich ascites cells. Thereasons for such a variability have not been investigated.
Table 1Dependence of uridine incorporation on extracellular nucleoside
concentration in intact Ehrlich ascites cellsEhrlich ascites cells (13 X IO6) suspended in 6 ml KRB, were
incubated at 30°with 5 /jCi of uiidine-3H and different concentrationsof "cold" uridine. The results are expressed as increase/min ofradioactivity incorporated into the RNA from 10' cells (Adpm/min/106 cells), or, assuming that the intracellular nucleoside poolcan be neglected, as pmoles uridine incorporated per min per IO6 cells.
UridineTotal uridine Specific activity incorporatedconcentration of uridine dpm/min/10' (pmoles/min/
(MM) (/jCi/Mitiole) cells IO6cells)
0.241.078.5783.5450001111138.914.3756291415.40.0690.1190.1350.172
Figs. 1 and 2. Autoradiographs of Ehrlich ascites cells after a 3-minpulse of thymidine-3 H (5 nCi/ml) in the absence (Fig. 1) or in thepresence (Fig. 2) of lucensomycin (2.5 Mg/106 cells). Incorporation was
started by addition of the cells to the medium containing labeledthymidine and, if indicated, lucensomycin. Other experimental detailsas described in "Materials and Methods."
Action ofPolyenic Antibiotics on Ascites Cells
Preliminary autoradiographical investigation indicated thatunder our experimental conditions most cells (> 80%) wereable to incorporate the labeled nucleoside. With thymidine-3 H,
after short incubation times (3 min), the label was foundmainly at the periphery of the nucleus (Fig. 1). Thisobservation can probably be compared to that of Comings andKakefuda (6), but its importance in the mechanism of DNAsynthesis requires more detailed investigation.
In all the systems investigated, addition of lucensomycinproduced inhibition of nucleoside incorporation (Chart 2). Itcan be shown that the percentage of inhibition did not dependon the concentration of the polyene in the medium but on theratio between the amount of lucensomycin and the number ofcells (Table 2); a similar behavior, as has been shown for otherpolyenes (17,18), is also valid for lysis of bovine erythrocytes.
Thymidine and uridine incorporation by Ehrlich ascites cellswere inhibited to a comparable extent by equal amounts oflucensomycin (Chart 2, A and B). Inhibition of uridineincorporation by Yoshida ascites cells instead requiredsomewhat higher amounts of lucensomycin per cell (Chart2C); this may be correlated to the microscopic observation ofthe larger size of these cells as compared to Ehrlich ascitescells.
Autoradiographic experiments confirmed that in thepresence of lucensomycin the cells became unable toincorporate 3H-labeled nucleosides into their nuclei. Moreover,
the cells appeared swollen and with enlarged nuclei (Fig. 2), ashas previously been described (21) in fresh, nonfixed
30
.20
16
12
L-s e
12
8 o
Ea
10 20 30lime (min.)
10 20 30
Chart 2. Kinetics of inhibition by lucensomycin or metabolicinhibitors, of uridine or thymidine incorporation by Ehrlich or Yoshidaascites cells. All assays at 30°were initiated by addition of the cells to
KRB containing the labeled nucleoside (2 nG/m\). After 15 min, duringwhich repetitive sampling was performed, more KRB, containing equalconcentrations of labeled nucleoside and either DMSO alone, 2 pl/nú(shown as L = 0), or lucensomycin in DMSO (the figure after Lindicating the lucensomycin/cell ratio, as ¿iglucensomycin/IO6 cells) orNaCN (5 X 10"" M, final concentration or 2,4-dinitrophenol (DNP) (5X 5~5 M final concentration). The incorporation values are given as
dpm in the acid-insoluble residue from 10' cells. A and £>,Ehrlich
ascites cells, thymidine incorporation; B, Ehrlich ascites cells, uridineincorporation; C, Yoshida ascites cells, uridine incorporation. Arrows,addition of lucensomycin; values on the ordinate should be divided bythe negative exponentials to obtain the correct dpm values.
APRIL 1972 871
Research. on November 9, 2015. © 1972 American Association for Cancercancerres.aacrjournals.org Downloaded from
Strom et al.
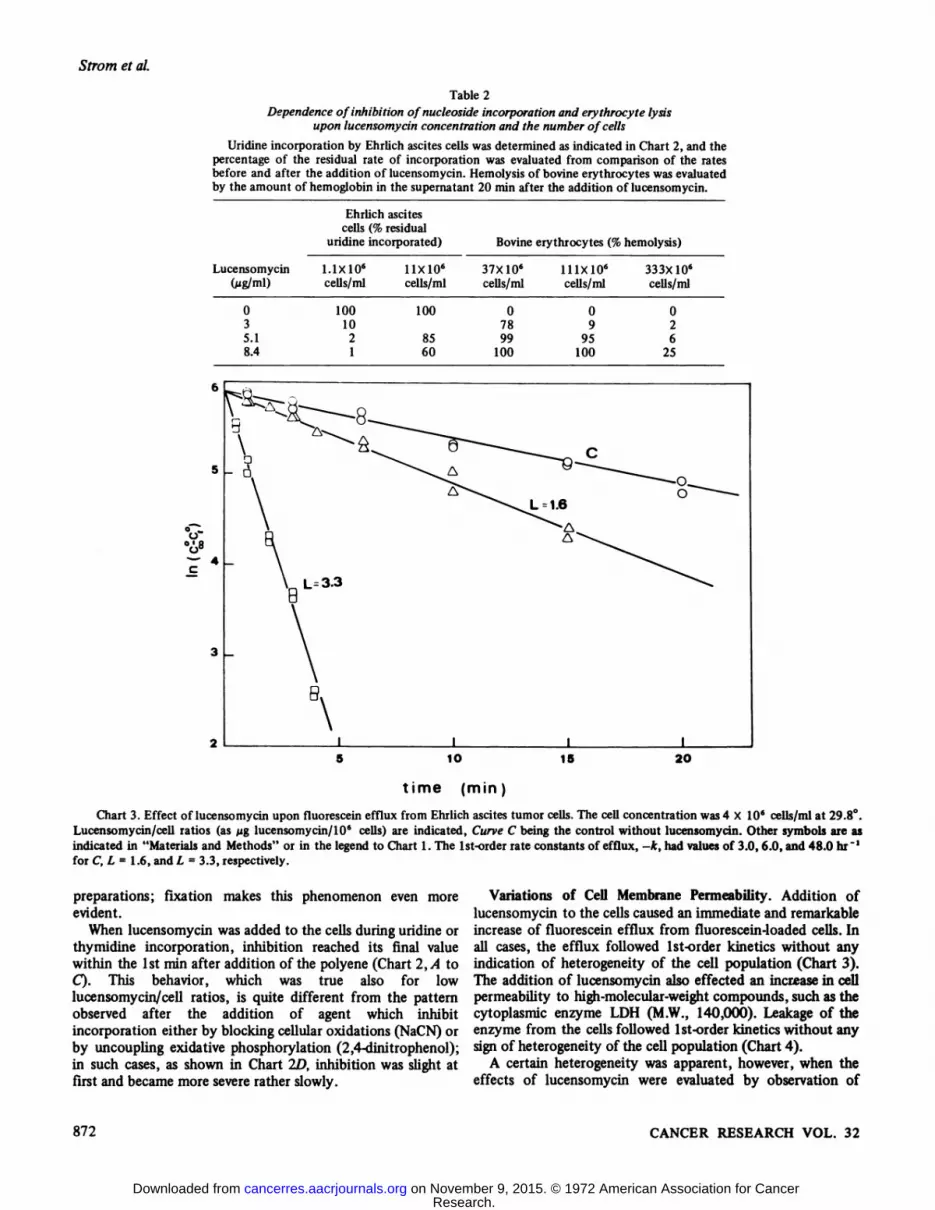
Table 2Dependence of inhibition ofnucleoside incorporation and erythrocyte lysis
upon lucensomycin concentration and the number of cells
Uridine incorporation by Ehrlich ascites cells was determined as indicated in Chart 2, and thepercentage of the residual rate of incorporation was evaluated from comparison of the ratesbefore and after the addition of lucensomycin. Hemolysis of bovine erythiocytes was evaluatedby the amount of hemoglobin in the supernatant 20 min after the addition of lucensomycin.
Ehrlich ascitescells (% residual
uridine incorporated) Bovine erythrocytes (% hemolysis)
Lucensomycin(Mg/ml)0
35.18.41.1X10«
cells/ml100
1021nxio6cells/ml10085
6037X106
cells/ml0
7899100lllxlO6cells/ml0
995
100333X10«
cells/ml02
625
0°~ 4
10
time (min)
15 2O
Chart 3. Effect of lucensomycin upon fluorescein efflux from Ehrlich ascites tumor cells. The cell concentration was 4 X 10' cells/ml at 29.8°.Lucensomycin/cell ratios (as fig lucensomycin/106 cells) are indicated, Curve C being the control without lucensomycin. Other symbols are asindicated in "Materials and Methods" or in the legend to Chart 1. The Ist-order rate constants of efflux, -k, had values of 3.0, 6.0, and 48.0 hr "'
for C, L = 1.6, and L = 3.3, respectively.
preparations; fixation makes this phenomenon even moreevident.
When lucensomycin was added to the cells during uridine orthymidine incorporation, inhibition reached its final valuewithin the 1st min after addition of the polyene (Chart 2, A toC). This behavior, which was true also for lowlucensomycin/cell ratios, is quite different from the patternobserved after the addition of agent which inhibitincorporation either by blocking cellular oxidations (NaCN) orby uncoupling exidative phosphorylation (2,4-dinitrophenol);in such cases, as shown in Chart 2D, inhibition was slight atfirst and became more severe rather slowly.
Variations of Cell Membrane Permeability. Addition oflucensomycin to the cells caused an immediate and remarkableincrease of fluorescein efflux from fluorescein-loaded cells. Inall cases, the efflux followed Ist-order kinetics without anyindication of heterogeneity of the cell population (Chart 3).The addition of lucensomycin also effected an increase in cellpermeability to high-molecular-weightcompounds, such as thecytoplasmic enzyme LDH (M.W., 140,000). Leakage of theenzyme from the cells followed Ist-order kinetics without anysign of heterogeneity of the cell population (Chart 4).
A certain heterogeneity was apparent, however, when theeffects of lucensomycin were evaluated by observation of
872 CANCER RESEARCH VOL. 32
Research. on November 9, 2015. © 1972 American Association for Cancercancerres.aacrjournals.org Downloaded from

Action ofPolyenic Antibiotics on Ascites Cells
0.O
40time (minutes)
30O
Chart 4. Effect of lucensomycin on LDH leakage from Ehrlichascites cells. Cells were incubated with or without lucensomycin(polyene/cell ratios, as jig lucensomycin/106 cells, are shown on thechart) at 30°.At different time intervals, samples were centrifuged, and
the supernatant was assayed for LDH activity. Values are expressed asdecrease of absorbance at 340 nm/min (A A/min), after addition of 50Ml of the supernatant to the pyruvate-NADH mixture under theconditions indicated in "Materials and Methods."
single cells by phase-contrast microscopy or in the presence oftrypan blue, 1 mg/ml. The cells appeared enlarged to a variableextent, and dye uptake by the nucleus and by the cytoplasm(21) was neither simultaneous nor uniform in all cells. Theexperimental conditions (time, temperature, oxygénation)could not, however, be carefully controlled during theobservation, and the heterogeneity observed, never verymarked, is therefore of doubtful significance.
The sensitivity of these membrane properties appeared, insome experiments, to be slightly lower compared to uridine orthymidine incorporation; a certain variability betweendifferent batches of cells makes this small difference doubtful,so that the increase in cell membrane permeability and theinhibition of nucleic acid biosynthesis appear to be strictlycorrelated.
A further parallelism between the action of polyene on cellmembrane permeability and its ability to inhibit nucleosideincorporation into nucleic acids of ascites tumor cells wasfound by investigations on the ./V-acetyl derivative oflucensomycin. This compound, which is slightly acidic andmore soluble in water than the amphoteric lucensomycin, isapproximately 40 times less effective on a molar basis thanlucensomycin in increasing cell membrane permeability, andhad, at concentrations up to 10 fig/106 cells (i.e., 14nmoles/106 cells), no effect on uridine incorporation by
Ehrlich ascites cells.
The structural formula of lucensomycin is (14):
OH
W-Acetyl derivatives of mycosamine-containing polyenes showless biological activity (as antifungal antibiotics) than theparent compounds (23).
Incorporation of Nucleotides into Nucleic Acids by IsolatedNuclei and by Lucensomycin-treated Cells. Incorporation ofthe uridylic moiety of labeled UTP by nuclei isolated fromeither Ehrlich or Yoshida cells in the presence of appropriateconcentrations of all 4 triphosphoribonucleo tides was not
400O
3000
2000
1OOO
Ea•¿�a
160O
1200
800
400 _
Ehrlich
- Yoshida
400
200
o
M»O
200 Eo.
too
5 10
time (min20
Chart 5. Incorporation of the uridylic moiety of UTP by nucleiisolated from Yoshida or Ehrlich ascites cells. Nuclei were prepared bynitrogen cavitation, and the assays were performed as described in"Materials and Methods." The UTP concentration was 0.13 mM, andthe final specific activity of UTP-14C was 3.85 ^Ci/Mmole. Nucleiconcentrations during assays were 18 X 10' (Yoshida) or 8 X 10'
(Ehrlich) nuclei/ml, o, no other addition; A, + DMSO, 2 Ml/ml; »,+lucensomycin, 40 Mg/ml, in DMSO. Incubation at 30°was started by
the addition of nuclei. Results are expressed either as dpm or as pmolesof labeled nucleotide incorporated into the acid-insoluble residue from0.5-ml samples.
APRIL 1972 873
Research. on November 9, 2015. © 1972 American Association for Cancercancerres.aacrjournals.org Downloaded from

Strom et al.
160O -
1200
Eo.
800
400
80
oC
m£oEo.
5 10time (min )
15
Chart 6. Incorporation of the thymidylic moiety of TTP by nucleiisolated from Ehrlich ascites cells. The TTP concentration was 0.3 mMand specific activity of TTP-3H was 8.6 MCi/Mmole. Nuclei were
prepared by nitrogen cavitation. Concentration of nuclei in the assaywas 7.5 X 10'/ml. The incubation mixture is given in "Materials andMethods." Samples of 0.5 ml each were taken. Symbols are as in Chart
5.
Table 3Binding capacity for lucensomycin of whole Ehrlich ascites
cells and isolated nucleiThe amount of lucensomycin required for 50% inhibition of uridine
incorporation was determined in the presence of different amounts ofEhrlich ascites cells or of isolated nuclei from Ehrlich ascites cells.
Cells/ml10
X10'6X10'IX
10«Lucensomycin(Mg/ml)for
50%Nuclei/mlinhibition1.187.128
X 10' 2.4
significantly modified by addition of lucensomycin (Chart 5).Similar results were also obtained for the incorporation of thethymidylic moiety of labeled TTP into DNA of nuclei fromEhrlich ascites cells (Chart 6). The procedure used for theisolation of the nuclei (nitrogen cavitation or Nonidet P-40)had no influence upon the results.
Since in a well-chosen range of lucensomycin concentrationsit is possible to evaluate the amount of free lucensomycin byits effect on nucleoside incorporation by intact cells, this assayisolated nuclei and of whole cells to bind the polyene. Asshown in Table 3, it is possible to calculate that 1 ng(i.e., 1.5nmoles) lucensomycin is bound by at least 23 million nucleiand by only 0.5 million cells.
Lucensomycin-treated cells became capable of incorporatingthe uridylic moiety of UTP into RNA (Chart 7). Under ourconditions, the rate of this incorporation was (with UTP
between 10 and 200 AIM) remarkably constant, i.e., around0.33 pmole UMP incorporated per min per 1 million Ehrlichascites cells. Paper chromatography confirmed that the labelwas contained in a nucleoside monophosphate moiety, whichcould be freed by mild alkaline hydrolysis.
The presence of all 4 triphosphoribonucleotides was notmandatory, labeled UMP being able in the presence of ATP,GTP, and CTP to substitute for labeled UTP without anyappreciable decrease in the rate of incorporation. As shown in
012
2 5 8 11 14 17 2 5 8 11 14 17
Chart 7. Incorporation of the uridine (A), UMP (B), uridylic moietyof UTP (C), or thymidylic moiety of TTP (£>)into lucensomycin-treated Ehrlich ascites cells. In A, B, and C, the incorporation mediumwas KRB to which 3.7 mM ATP, 2.2 mM GTP, 0.3 mM CTP, 50 mMmercaptoethanol, and 3 mM MnCl2, 60 mM Tris-HCl buffer (final pH7.6) and either 76 MMuridine-3H (specific activity, 182 ^Ci/Mmole) or140 MM UMP-'"C (specific activity, 16.6 nCi/nmo\e) or 158 MMUTP-^C (specific activity, 153 MCi/Mmole)were added. In D, the
incorporation medium was KRB to which 5 mM ATP, 0.5 mM dATP,0.5 mM dGTP, 0.5 mM dCTP, 10 mM MgCl2, and 90 mM Tris-HClbuffer (final pH 7.7), and 75 MM TTP-3H (specific activity, 42
MCi/Mmole)had been added.Incorporation was started by the addition of the cells to a final
concentration of 20 X 10' cells/ml to the medium equilibrated at 30°.
Lucensomycin was either absent (o) or present (•)in the medium at aconcentration of 276 Mg/ml (13.8 Mg/10' cells).
Results are expressed as dpm incorporated in the acid-insolubleresidue from 0.25-ml samples. The values on the ordinate should bedivided by the exponential to obtain the correct dpm values.
Table 4Dependence of uridine incorporation on extracellular nucleoside
concentration in lucensomycin-treated Ehrlich ascites cells
The procedure was the same as that described for Chart 1A, exceptfor uridine concentration. The final number of cells was 4.8 X 10' /ml.
Samples of 0.5 ml were used. The rate of incorporation in the presenceof lucensomycin is expressed as percentage of controls without thepolyene. In the uridine range considered, all controls had similarincorporation values.
Uridine(MM)76
156808
1446Incorporation
rate(% of control)5
12.549.063.4
874 CANCER RESEARCH VOL. 32
Research. on November 9, 2015. © 1972 American Association for Cancercancerres.aacrjournals.org Downloaded from

Action ofPolyenic Antibiotics on Ascites Cells
Table 4, under these conditions even uridine, at very high REFERENCESconcentrations, could be incorporated into RNA oflucensomycin Ehrlich ascites cells at a rate comparable to thatof intact cells. Lucensomycin-treated cells could also performthe incorporation of the thymidylic moiety of TTP into DNA.
DISCUSSION
The inhibitory effect of lucensomycin on the incorporationof nucleosides into nucleic acids of ascites tumor cells appearsto be strictly correlated to the action of the polyene on thecell membrane. The rapidity of onset of the effect oflucensomycin on nucleic acid biosynthesis by the cellsindicated that the inhibition observed is not due to leakage ofeither small-molecular-weight compounds or cytoplasmicenzymes, since leakage could be shown to be, under controlledconditions, relatively slow. A possible explanation could bethe inability of the cells, because of increased leakiness, toconcentrate the labeled nucleoside in intracellular values highenough to allow efficient phosphorylation and subsequentincorporation into DNA or RNA. Although specific effects ofpolyenes upon transport systems have been shown in intestinalmucosa (1) and yeast cells (17), in our case lucensomycinseems to act mainly, if not exclusively, by increasing passivediffusion.
Lucensomycin treatment makes the cells able to utilizenucleotides for RNA and DNA synthesis. UMP, but noturidine, in concentrations up to 0.3 mM, can substitute forUTP. The nucleotide kinase system is therefore intact, whileuridine kinase is able to work only at very high uridineconcentrations.
Lucensomycin-treated cells can be compared to toluenizedbacteria (14, 16); toluene has a more graded effect, so thattoluene-treated bacteria may become permeable only tomolecules with a molecular weight of < 2000 (9).
If polyene-treated cells are compared to isolated nuclei fromthe same cells, 2 main differences are apparent: a greaterability to utilize nucleoside monophosphates as substitutes fortriphosphates for incorporation into nucleic acids (due mostprobably to preservation of nucleotide kinase activity inlucensomycin-treated cells) and a longer linearity of thekinetics of incorporation of nucleoside triphosphates intonucleic acids.
It has been proposed (5, 6, 8, 14, 15, 23) thatnucleoprotein-lipid association at the nuclear membrane levelmay be of importance in initiating DNA synthesis inmammalian cells. Although under our conditionsautoradiography of Ehrlich ascites cells after a pulse oftritiated thymidine seems to confirm this possibility (Fig. 1),no evidence is available thus far to ascertain whetherlucensomycin has any effect on the nuclear membrane or onits association to DNA.
ACKNOWLEDGMENTS
The authors thank Mr. Vincenzo Peresempio for his skillful technicalassistance, and Professor Stelio Crifò,Institute of Otorhinolaiyngology,University of Rome, for the microscopic observations and photographs.
1. Adams, T. H., Wong, R. G., and Norman A. W. Studies on theMechanism of Action of Calciferol. II. Effects of the PolyeneAntibiotic, Filipin, on Vitamin D-mediated Calcium Transport. J.Biol. Chem., 245: 4432-4442,1970.
2. Avis, P. J. G. Pressure Homogenization of Mammalian Cells. In: G.D. Birnie and S. M. Fox (eds.), Subcellular Components, pp. 1-12.
London: Butterworth & Co., Ltd., 1969.3. Burgess, R. R. A New Method for the Large Scale Purification of
Escherichia coli Deoxyribonucleic Acid-dependent RibonucleicAcid Polymerase. J. Biol. Chem., 244: 6160-6167,1969.
4. Celada, F., and Rotman, B. A Fluorochiomatic Test forImmunocytotoxicity against Tumor Cells and Leucocytes inAgarose Plates. Proc. Nati. Acad. Sei. U. S., 57: 630-636,1967.
5. Comings, D. E., and Kakefuda, T. Initiation of DeoxyribonucleicAcid Replication at the Nuclear Membrane in Human Cells. J. Mol.Biol., 33: 225-229,1968.
6. DuPraw, E. J. The Organization of Nuclei and Chromosomes inHoneybee Embryonic Cells. Proc. Nati. Acad. Sei. U. S., 53:161-168,1965.
7. Ficq, A. Autoradiography. In: J. Brächetand A. E. Mirsky (eds.),The Cell, Vol. 1, pp. 67-90. New York: Academic Press, Inc.,
19598. Friedman, D. L., and Mueller, G. C. Studies on the Nature of
Replicating DNA of HeLa Cells. Biochim. Biophys. Acta, 174:253-263,1969.
9. Gachelin, G. A New Assay of the Phosphotransferase System inEscherichia coli. Biochem. Biophys. Res. Commun., 34: 382-387,1969.
10. Gaudiano, G., Bravo, P., Quilico, A., Golding, B. T., and Rickards,R. W. Struttura della Lucensomicina-Nota III. Gazz. Chim. Ital.,96: 1470-1491, 1966.
11. Goldberg, M. L., Moon, H. D., and Rosenau, W. Partial Purificationof RNA Polymerase from Rat Liver Nuclei. Biochim. Biophys.Acta, / 71: 192-194,1969.
12. Goldthwait, D. A., Antony, D. D., and Wu, C. W. Studies with theRNA Polymerase. In: L. Silvestri (ed.), RNA-Polymerase andTranscription, pp. 10-27; Amsterdam: North Holland PublishingCo., 1970.
13. Hall, R. H., and Khorana, H. G. Nucleoside Polyphosphates. II. ASynthesis of Uridine-5'-di- and -triphosphates. J. Am. Chem. Soc.,
76: 5056-5060, 1954.
14. Jackson, R. W., and De Moss, J. A. Effects of Toluene onEscherichia coli. L Bacteriol., 90: 1420-1425, 1965.
15. Jackson, V., Earnhardt, J., and Chalkley, R. A DNA-JJpid ProteinContaining Material Isolated from Calf Thymus NuclearChromatin. Biochem. Biophys. Res. Commun., 33: 253-259,1968.
16. Kohiyama, M., and Kolber, A. R. Temperature Sensitive Mutantof the DNA Replication System in Escherichia coli. Nature, 228:1157-1160, 1970.
17. Lampen, J. O. Interference by Polyenic Antifungal Antibiotics(Especially Nystatin and Filipin) with Specific MembraneFunctions. Symp. Soc. Gen. Microbiol., 16: 111-130, 1966.
18. Lechevalier, H., Borowski, E., Lampen, J. O., and Schaffner, C. P.Water Soluble W-Acetyl Derivatives of Heptaene MacrolideAntifungal Antibiotics: Microbiological Studies. Antibiot. Chemotherapy, 11: 640-647, 1961.
19. Loeb, L. A. Purification and Properties of Deoxyribonucleic AcidPolymerase from Nuclei of Sea Urchin Embryos. J. Biol. Chem.,244: 1672-1681,1969.
20. Mondovì,B., Finazzi Agro, A., Rotilio, G., Strom, R., Moricca,G., and Rossi Fanelli, A. The Biochemical Mechanism of Selective
APRIL 1972 875
Research. on November 9, 2015. © 1972 American Association for Cancercancerres.aacrjournals.org Downloaded from

Strom et al.
Heat Sensitivity of Cancer Cells. II. Studies on Nucleic Acids andProtein Synthesis. European J. Cancer, 5: 137-146,1969.
21. Mondovi, B., Strom, R., Finazzi Agro, A., Caiafa, P., De Sole, P.,Bozzi, A., Rotilio, G., and Rossi Fanelli, A. Effect of PolyenicAntibiotics on Ehrlich Ascites and Novikoff Hepatoma Cells.Cancer Res., 31: 505-509, 1971.
22. Mondovi, B., Strom, R., Rotilio, G., Finazzi Agro, A., CavaliereR., and Rossi Fanelli, A. The Biochemical Mechanism of SelectiveHeat Sensitivity of Cancer Cells. I. Studies on Cellular Respiration.European J. Cancer, 5; 129-136,1969.
23. Ormerod, M. G., and Lehmann, A. R. The Release of HighMolecular Weight DNA from a Mammalian Cell (L 5178 Y).Attachment of the DNA to the Nuclear Membrane. Biochim.Biophys. Acia, 228: 331-343,1971.
24. Rotman, B., and Papermaster, B. W. Membrane Properties of LivingMammalian Cells as Studied by Enzymatic Hydrolysis ofFluorogenic Esters. Proc. Nati. Acad. Sei. U. S., 55: 134-141,1966.
25. Sato, S., Matsushima, T., and Sugimura, T. Hexokinase IsozymePatterns of Experimental Hepatomas of Rats. Cancer Res., 29:1437-1446, 1969.
26. Steck, T. L., and Wallach, D. F. H. The Isolation of PlasmaMembranes, in: H. Busch (ed.), Methods in Cancer Research, Vol.5, pp. 93-153. New York: Academic Press, Inc., 1970.
27. Stein, H., and Hausen, P. A Factor from Calf Thymus StimulatingDNA-dependent RNA Polymerase Isolated from This Tissue.European J. Biochem., 14: 270-277,1970.
28. Teng, C., Bloch, D. P. and Roychoudhury, R. DNA Synthesis inNuclei Isolated from Ehrlich Ascites Tumor Cells. Biochim.Biophys. Acta, 224: 232-245,1970.
29. Tennant, J. R. Evaluation of the Trypan Blue Technique forDetermination of Cell Viability. Transplantation, 2: 685-694,1964.
30. Tyner, E. P. Heidelberger, C. and La Page, G. A. IntraceUularDistribution of Radioactivity in Nucleic Acids Nucleotides andProtein following Simultaneous Administration of P3ÃŽandGlycine-2-C1 *. Cancer Res., 13: 186-192,1953.
31. Widnell, C. C., and Tata, J. R. Studies on the Stimulation byAmmonium Sulphate of the DNA-dependent RNA Polymerase ofIsolated Rat Liver Nuclei. Biochim. Biophys. Acta, 123: 478-492,1966.
876 CANCER RESEARCH VOL. 32
Research. on November 9, 2015. © 1972 American Association for Cancercancerres.aacrjournals.org Downloaded from

1972;32:868-876. Cancer Res Roberto Strom, Argante Bozzi, Antonio Scioscia Santoro, et al. of Nucleic Acid Biosynthesis in Ascites Tumor CellsStudies on the Mechanism of Inhibition by Polyenic Antibiotics
Updated version
http://cancerres.aacrjournals.org/content/32/4/868
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on November 9, 2015. © 1972 American Association for Cancercancerres.aacrjournals.org Downloaded from





















![Antibiotics proof[1]](https://img.pdfslide.net/doc/110x75/634560b16cfb3d406409a867/antibiotics-proof1.jpg)







