Embed Size (px)
Citation preview

Pl. Syst. Evol. 211:13-29 (1998) --Plant
Systematics and
Evolution © Springer-Verlag 1998 Printed in Austria
Studies on the reproductive biology of Melastomataceae in "cerrado" vegetation
RENATO GOLDENBERG and G~ORGE J. SHEPHEm~
Received October 3, 1996; in revised form May 20, 1997
Key words: Melastomataceae, Leandra, Miconia, Tibouchina. - Breeding systems, apomixis, pollen fertility.
Abstract: Controlled pollinations and observations of pollen tube growth, pollen fertility and cytological data were studied in 11 species of Melastomataceae growing in "cerrado" in SE Brazil. Seven species produced apomictic fruits with viable seeds. Four species were not apomictic, with two self-incompatible (gametophytic SI) and two self-compatible, one showing spontaneous self-pollination. The apomictic species had lower pollen fertility than sexual ortes - although in some cases differences were not significant - showing that low pollen fertility may be a useful indicator of apomixis if analysed using careful sampling supplemented by emasculation experiments. The apomictic species also showed meiotic irregularities, probably related to hybridization, polyploidy and the cause of low pollen fertility. Apomixis in this group probably is more associated with taxonomy and phylogeny than habitat or habit, since 85% of the apomictic species reported in the literature belong to the tribe Miconieae and occur in a wide range of vegetation types and have diverse habits. This suggests the existence of very extensive apomictic complexes comparable to those reported in mainly temperate genera of Asteraceae and Rosaceae.
The family Melastomataceae is composed of around 166 genera and 4200-4500 species, occurring mainly in tropical and subtropical regions (R~N~~-~R 1993). Reproductive systems have been investigated in about 100 species and it has been shown that apomixis is relatively frequent in this group (RENN~R 1989). Apomixis is a phenomenon that has been extensively studied (GUSTA~SON 1946, 1947a, b; NO~LER 1984) and is known to occur at rather high frequencies in members of the families Asteraceae, Rosaceae and Poaceae (RICHARDS 1986), and rather more sporadically in other angiosperm families (NY6~N 1954). The great diversity of mechanisms that these plants employ to reproduce asexually (NO~LER 1984) seems to indicate that this process has evolved independently in all of these groups. In a large proportion of these cases, apomixis is associated with polyploidy and hybridization.
The vast majority of the cases of apomixis that have been studied and the models based on them refer to species from temperate regions. For plants from tropical areas, there are indications that besides Poaceae and Melastomataceae,

14 R. GOLDENBERG & G. J. SHEPHERD"
apomixis may be common in Dipterocarpaceae (KAUR & al. 1978) and occurs sporadically in at least 30 genera (BAKER & al. 1983) belonging to diverse families including Bombacaceae (OLIVEIRA & al. 1992), Clusiaceae (HA & al. 1988), Rutaceae (PIEDADE & RANGA 1993) and Erythroxylaceae (BERRY & al. 1991).
Most of the tropical examples are forest species, and little seems to be known about the distribution of apomixis, and reproductive biology in general, in other tropical formations such as savanna. In Brazil, savanna formations - here known as "cerrados" - cover extensive areas and are very poofly investigated from the point of view of plant reproduction. In one of the few studies of reproductive systems in a cerrado community, OLIVEIRA (1991) found a predominance of xenogamous species, mainly visited by small insects and with zoochory. This author reported apomixis only for Miconia albicans (Melastomataceae) and Eriotheca pubescens (Bombacaceae).
The main objective of the present study was to investigate breeding systems in cerrado species of Melastomataceae and to determine the degree of occurrence of apomixis in these species and to compare these data with results for species from other formations reported in the literature. We particularly wished to begin investigation of a number of questions raised about the occurrence of apomixis in this family: - Is apomixis associated with particular habitats, being commoner in forest or cerrado? Alternatively, is there a closer relationship with habit or with taxonomic groupings within the family? To what extent is apomixis a really common phenomenon in neotropical representatives of this family? Is it possible to detect apomictic species on the basis of their pollen sterility?
Materials and methods
Field work was carried out in the municipality of Itirapina, Säo Paulo State, SE Brazil (22°15 t S; 47°49/W), in two cerrado areas in the Experimental Station of the Instituto Florestal. Both areas were on red latosol (sandy phase) at an altitude of 760 m a. s. 1. The climate in this region is Cwa in KöPeEN'S classification, with a mean annual temperature of 19.7 °C (July mean 16.2°C and February mean 19.7 °C) and a mean annual rainfall of 1425 mm, mainly concentrated in the rainy season (l199mm) from October to March (GIANOrTI 1988).
The two areas had been subject to some degree of disturbance. The first had a more open physiognomy, with signs of fites in previous years. The second consisted of a gradient between more open cerrado s. str. and taller, more closed "cerradäo", with an area of transition to gallery forest. This area also contained several gravel pits.
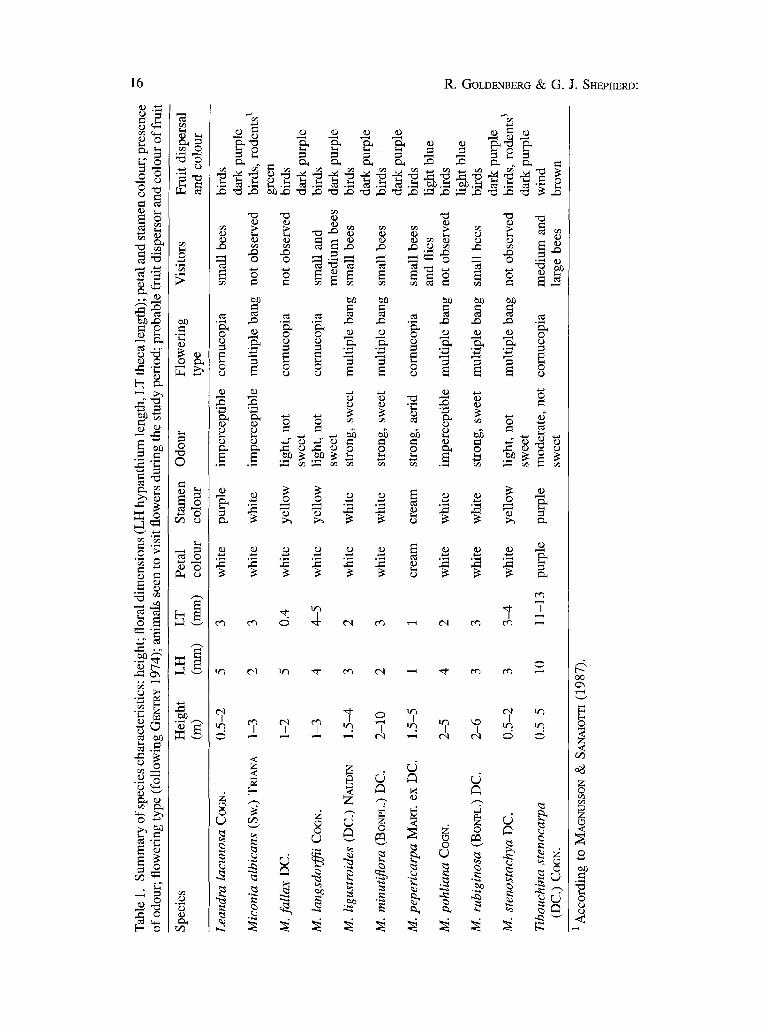
As far as possible, all of the species of Melastomataceae occurring in the area were studied. This gare a total of 11 species, nine of Miconia, one of Leandra and one of Tibouchina (Table 1), representing about 20% of the Melastomataceae reported to occur in cerrado vegetation (A. A. J. F. CASTRO, pers. comm.; A. A. J. E CASTRO • al., unpubl.)
Breeding systems were studied using controlled pollinations (self and crossed), no pollination (for apomixis) and spontaneous pollination, and these were compared with control flowers which were marked but otherwise left undisturbed. Inflorescences with various flowers were isolated in paper bags for these treatments. Since many of the flowers were very small, tests for apomixis were made by removing the stigmatic region before anthesis by cutting the bud and not by emasculation, a technique similar to that used by

Reproductive biology of Melastomataceae in Cerrado 15
RICHARDS (1986) when working with Asteraceae. In all cases, a few flowers were also individually emasculated and isolated to verify that the bud-cutting technique gave reliable results.
The number of individual plants and flowers used varied with the species and the number of plants available in the experimental area. As far as possible, at least five plants were used and all of the treatments were carried out on each individual, using a minimum of ten flowers per treatment. In M. pohliana and M. langsdorffii three and four individuals were used respectively. Fruit development was followed for 45-60 days after anthesis, and the resulting fruits collected just before full maturation to avoid losses to birds and other dispersal agents. Seeds from fruits produced in the control and apomixis treatments were germinated to ensure that the seeds from the apomixis treatment were, in fact, viable. Differences between treatments and individuals within treatments were tested using two- way analysis of variance without repetition ( S o l l & ROHLF 1981). Percentage fruit formation per individual per treatment was arcsine transformed.
Pollen fertility was observed using material fixed in 50% or 70% FAA and stained with acetic carmine. Five buds were analysed for each individual, with counts of 100 grains per bud. The buds were collected from a minimum of ten plants, except for M. pohliana (three individuals). Widely spaced individuals were used to avoid resampling clones established by vegetative reproduction, which may also occur in some species.
Meiosis was observed in two species using buds fixed in Carnoy no. 2 (6:3:1 - ethanol:chloroform: acetic acid - SOLT & WURDACK 1980) and stained in acetic carmine.
For the species with sexual reproduction, pollen tube growth was observed using fluorescence microscopy and staining with aniline blue, following MARTIN (1959).
Voucher specimens are deposited in the UEC herbarium (Botany Department, State University of Campinas). The collections are GOLDENBER6 28780 (Leandra lacunosa), 28775 (Miconia albicans), 28776 (M. fallax), 28492 (M. langsdorffii), 28488 (M. ligustroides), 28490 (M. minutiflora), 28485 (M. pepericarpa), 28483 (M. pohliana), 28486 (M. rubiginosa), 28493 (Tibouchina stenocarpa) and SHEPHERD & GOLDENBERG 28774 (M. stenostachya).
Results and discussion
Floral morphology and visitors. All of the species are shrubby with the exception of M. minutiflora which varies from a shrub to a small tree up to 10 m high, and all are characteristic of cerrado vegetation although M. minutiflora and M. langsdorffii also occur in secondary forest (Table 1).
The species of Miconia and Leandra, both belonging to the tribe Miconieae, have small flowers and stamens with thecae measuring from 0.1 to 0.5 cm. All have whitish petals (cream-coloured in M. pepericarpa) and cream-coloured (M. pepericarpa), yellow (M. fallax, M. stenostachya and M. langsdorffii), purple (L. lacunosa) or white anthers (remaining species). The flowers of M. pepericarpa give of t a strong acrid smell while those of M. ligustroides, M. minutiflora and M. rubiginosa have a sweetish odour and those of M. fallax, M. stenostachya and M. langsdorffii have a faint, not sweetened smell. No perceptible odour was detected in L. lacunosa, M. albicans and M. pohliana. Visits were entirely absent in M. albicans, M. pohliana, M. fallax and M. stenostachya, in spite of abundant flower production, and were very rare in L. lacunosa, M. ligustroides and M. rubiginosa. With the exception of M. pepericarpa, the flowers of the remaining species were visited by a number of bee species. All have berry-like fruits dispersed by animals,
16 R. Go~o~~ßzI~~ & G. J. SH~vrrzgo:
° ~~ o , . ~ i
~ , b ~ ~ ~
~~ ~~ g ~
~~ ~~
N ~
° ~
ù~
. ~ ~ ~ • .
ù~ ©
=
t"¢3 t ' ¢ 3
ù~
©
aù
r¢3
I
r..) ~ o
°~
o
Z o
o
<

Reproductive biology of MeIastomataceae in Cerrado 17
generally birds, except for M. albicans and M. fallax which are probably dispersed by rodents (MAGNUSSON & SANAIOTI'I 1987).
Miconia pepericarpa merits separate consideration as it is the only species to emit an acrid smell and its flowers produce a liquid which may be nectar. This liquid is produced by only some of the flowers and is viscous and very sweet. These characteristics would seem to indicate that M. pepericarpa may be pollinated by flies, the first indication of this pollination syndrome for Melastomataceae. It remains to be demonstrated if flies are effective pollinators for this species, since in addition to visits by several species of flies, wasps and small bees (which may buzz the flowers or not) were observed. For this species vibratile pollination does not appear to be necessary, as in other species of Miconia with rimose anthers or wide pores where a touch or any brusque movement is capable of liberating pollen. Miconia latecrenata, a species of the Atlantic Forest, is pollinated by bees which just walk over the dense inflorescences without vibratory movements (R. GOLDENBEaG, personal field observations).
Nectar production was reported by Mop, I & PIPoL¥ (1984) for M. minutiflora, but not observed in this study. It is possible that the material used here and that used by these authors may not belong to the same species because of difficulties with identification.
Tibouchina stenocarpa is the only species studied here belonging to the tribe Melastomeae. It has rauch larger flowers, with purple petals and stamens and a moderate, non-sweet, scent. It is visited by large bees which vibrate the stamens. The fruits of this species are capsules and the seeds dispersed by wind.
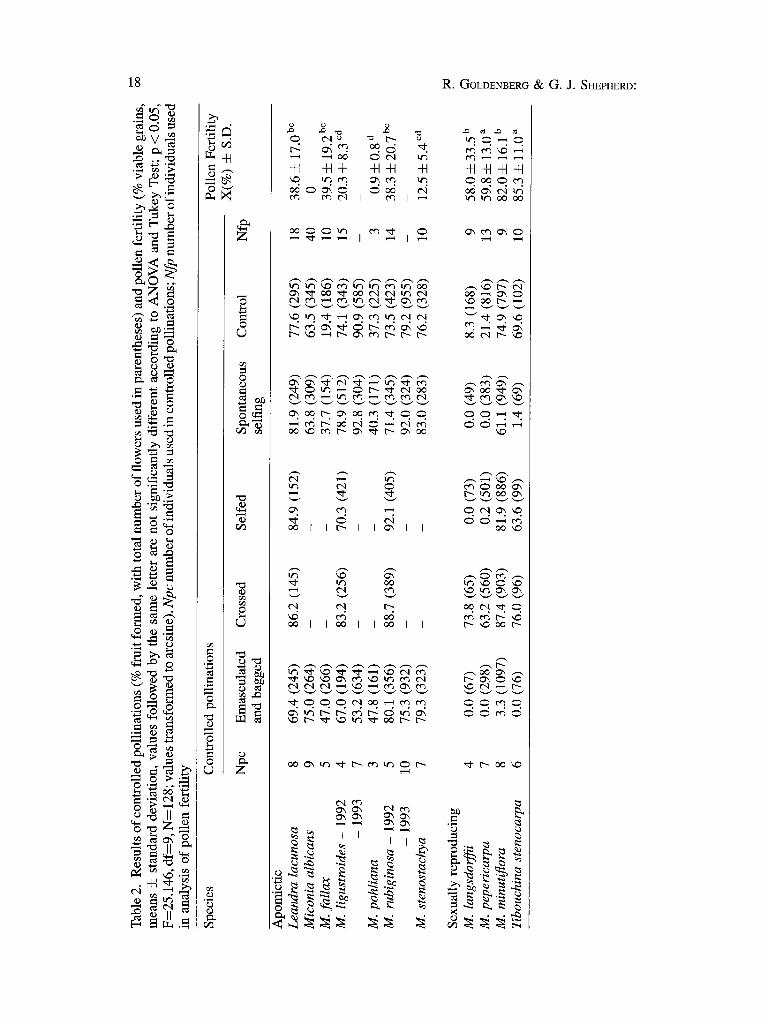
Reproduetive systems. The results of the experimental treatments are shown in Table 2. From these we conclude that four species have sexual reproduction, of which two are self-incompatible (SI) and two self-compatible (SC), and one of the latter shows spontaneous self-pollination. The remaining seven species all appear to be apomictic with high levels of fruit set when flowers are emasculated or have their stigmas removed. Miconia albicans and M. stenostachya have already been cited as apomicts in the literature by PENNER (1989) and BAUMGRATZ • SILVA (1988) respectively.
In the two self-incompatible species, M. pepericarpa and M. langsdorffii, practically no fruit-set was obtained from selfing and pollen tube growth ceased in the upper third of the style in selfed flowers. This suggests the presence of a gametophytic SI system (GIBBS 1990). SI systems appear to be relatively common in the family, with the largest number of cases reported in Miconieae and Rhexiae (Table 4). As far as we are aware, the genetic nature of SI in this family has never been studied and at present it is not possible to confirm gametophytic control.
In the two SC species, T. stenocarpa and M. minutiflora, fruit set from selfing was comparable to that obtained from cross-pollination and in control flowers. Miconia minutißora gave high fruit-set for spontaneous selfing, although this was somewhat lower than in the other treatments (ANOVA: d l -3 ; F=7.434; p < 0.001). For the genus Miconia there is one other documented case of spontaneous selfing in M. sintenisii in Costa Rica (DENT-ACOSTA & BP, ECKON 1991). This species has anthers with wide pores and, like M. pepericarpa, does not appear to need vibratile pollination. The very low fruit set obtained here in the apomixis treatment may indicate the presence of apomixis at very low frequency, but we feel that it is more
18
• ~ d -~ ~v~
0
©
,~ r~
'-~ 0
° I ~~.~
=c5
0
0
0 ¢)
!
B 2 ~ 2
-H +l+l -H-H -H
~ . ~ '-2.
I I
I I
~~.~~~ ~«
R. GOLDENBERG & G. J. SHEPHERD:
oO ',D
~ v v ~
O O ~ O

Reproductive biology of Melastomataceae in Cerrado 19
likely that this was the result of accidental contamination rather than apomixis, especially since the other apomictic species showed comparatively high levels of fruit-set. The population of M. minutißora cited by R E ~ a (1989) as apomictic actually belongs to M. regelli CoaN. (S. S. R E ~ a , pers. comm.).
Amongst the seven apomictic species, only M. ligustroides showed any significant difference between treatments, with a reduction, in the 1993 season, of fruit-set in the apomixis treatment. This may indicate some degree of sexual reproduction and thus, facultative apomixis, but it is impossible to confirm this without progeny tests using genetic markers. Miconia albicans appears to have obligate apomixis, since this species nearly always appears to fruit regularly and abundantly in natural conditions although no viable pollen was found in any of the 40 individuals which were sampled, at least during the two years in which the experiments were carried out. Observations of pollen samples from herbarium material also showed an apparent complete absence of well-formed pollen grains in practically all of the material, although a few well-formed grains (around 2%) were found in two samples from the state of Mato Grosso. We do not know if these do, in fact, represent viable pollen. It is possible, however, that some degree of sexual reproduction does exist in this species, particularly if pollen is donated by other species growing in the same area. The high degree of morphological variability generally found in natural populations of this species certainly suggests that several genotypes may be present, and that some degree of sexual reproduction might explain this. On the other hand, potential pollinator visits seem to be very rare, since none was observed during the whole experimental period which involved several weeks of field observations. It is possible that potential pollinators have learned to avoid Miconia albicans since its anthers are always practically empty and offer no reward. A similar avoidance of a pollen-sterile apomict was noted by BORGES (1991) in Leandra. The remaining apomictic species all show similar levels of fruit-set for all of the treatments, and it is difficult to reach any firm conclusions about the degree of sexual reproduction that still occurs in these species. The variation in pollen fertility noted in several species, however, suggests that there may be considerable variation between individuals, and it would be unwise to draw wide-ranging conclusions based on samples of only a few individuals.
The seeds obtained from apomictic fruits in six species germinated normally, with polyembryony observed in M. rubiginosa. In the seventh species (M. pohliana), it was not possible to obtain enough mature fruits for germination tests, but the fruits had perfect seeds.
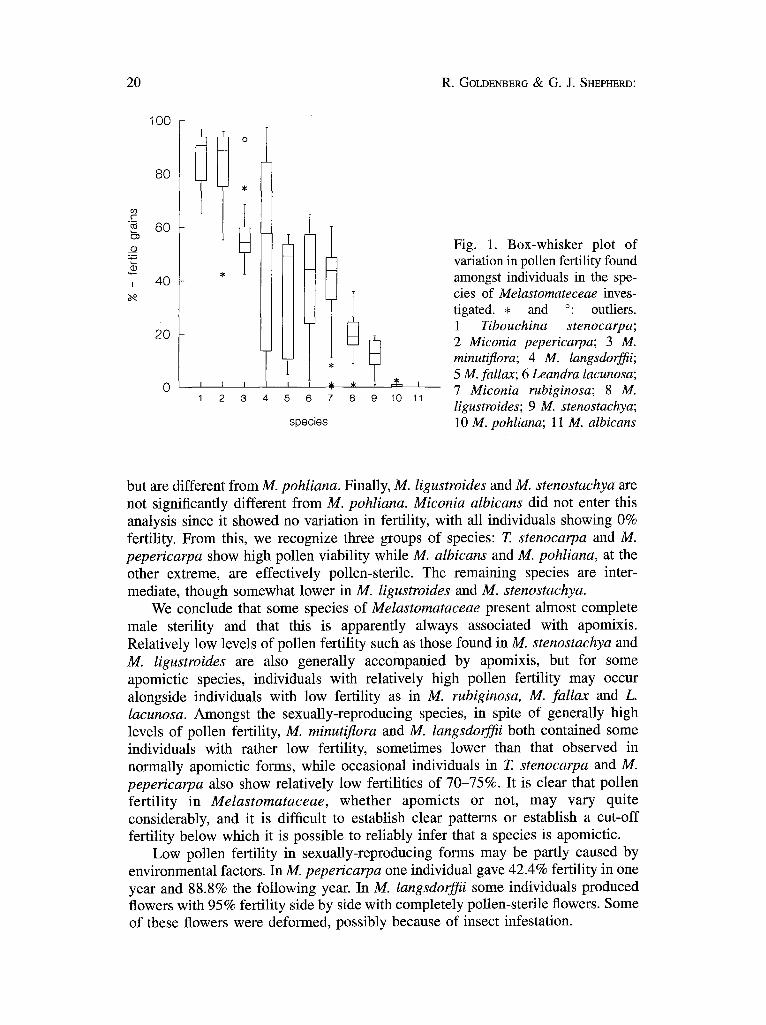
Pollen fertility. Pollen fertility was found to be extremely variable, both within and between species (Table 2). Figure 1 shows a comparison of pollen fertility for all 11 species.
The mean fertility values show significant differences (ANOVA: F = 25.146; p < 0.05). A comparison of means using the Tukey test (Table 2) shows that there is no significant difference between the means of T. stenocarpa and M. pepericarpa, both sexually reproducing, and that these differ from the rest. Miconia minutiflora and M. tangsdorffii are not significantly different from each other and also do not differ from M. rubiginosa, M. fallax and L. lacunosa. The first two have sexual reproduction while the rest are apomictic. Miconia rubiginosa, M. fallax and L. lacunosa do not differ from M. ligustroides and M. stenostachya (all apomictic),
20 R. GOLDENBERG • G. J. SHEPHERD:
C
100
80
60
40
20
I
0 i t i ~ 1 2 8 4 lO 11 5 6 7
spec ies
8 9
Fig. 1. Box-whisker plot of variation in pollen fertility found amongst individuals in the spe- cies of Melastomateceae inves- tigated. • and °: outliers. 1 Tibouchina stenocarpa; 2 Miconia pepericarpa; 3 M. minutiflora; 4 M. langsdorffii; 5 M. fallax; 6 Leandra lacunosa; 7 Miconia rubiginosa; 8 M. ligustroides; 9 M. stenostachya; 10 M. pohliana; 11 M. albicans
but are different from M. pohliana. Finally, M. ligustroides and M. stenostachya are not significantly different from M. pohliana. Miconia albicans did not enter this analysis since it showed no variation in fertility, with all individuals showing 0% fertility. From this, we recognize three groups of species: T. stenocarpa and M. pepericarpa show high pollen viability while M. albicans and M. pohliana, at the other extreme, are effectively pollen-sterile. The remaining species are inter- mediate, though somewhat lower in M. ligustroides and M. stenostachya.
We conclude that some species of Melastomataceae present almost complete male sterility and that this is apparently always associated with apomixis. Relatively low levels of pollen fertility such as those found in M. stenostachya and M. ligustroides are also generally accompanied by apomixis, but for some apomictic species, individuals with relatively high pollen fertility may occur alongside individuals with low fertility as in M. rubiginosa, M. fallax and L. lacunosa. Amongst the sexually-reproducing species, in spite of generally high levels of pollen fertility, M. minutiflora and M. langsdorffii both contained some individuals with rather low fertility, sometimes lower than that observed in norrnally apomictic forms, while occasional individuals in T. stenocarpa and M. pepericarpa also show relatively low fertilities of 70-75%. It is clear that pollen fertility in Melastomataceae, whether apomicts or not, may vary quite considerably, and it is difficult to establish clear patterns or establish a cut-off fertility below which it is possible to reliably infer that a species is apomictic.
Low pollen fertility in sexually-reproducing forms may be partly caused by environmental factors. In M. pepericarpa one individual gave 42.4% fertility in one year and 88.8% the following year. In M. langsdorffii some individuals produced flowers with 95% fertility side by side with completely pollen-sterile flowers. Some of these flowers were deformed, possibly because of insect infestation.

Reproductive biology of Melastomataceae in Cerrado 21
In the apomictic species, variation in pollen fertility may be caused by both environmental effects and meiotic irregularities. Examination of meiosis in M. fallax and M. stenostachya showed considerable variation in bivalent size, suggesting that at least some of the supposed bivalents may in fact have been univalents, and variable numbers of bridges and lagging chromosomes (possibly univalents) were common in anaphase I. In M. stenostachya, bridges were also observed in anaphase II, possibly resulting from paracentric inversions and double crossovers as suggested by BURNI-IAM (1962), but it was not possible to establish the origin of these. Tetrads with micronuclei were common in both species, probably resulting from the lagging chromosomes observed at anaphase I. Although meiosis was not observed in the remaining apomicts, it seems likely that similar irregularities occur.
In contrast to the pollen of the sexually-reproducing species, pollen in the apomicts was frequently irregular in size and shape, with many grains remaining only lightly coloured, leading to difficulty in distinguishing between fertile and sterile grains, and it is likely that pollen fertility as reported here was in fact considerably over-estimated.
The variability observed in this study makes it clear that it is dangerous to estimate pollen fertility without careful sampling of a large number of flowers and individuals, preferably at different seasons or in different years and including several populations. Fruit formation in spite of low pollen fertility may be a useful indicator of apomixis in Melastomataceae, especially when precocious fruit development is noted in unopened buds, but these observations must be supplemented by emasculation experiments to confirm that apomixis really does o c c u r .
Apomixis, polyploidy and hybridization. A strong association between apomixis and polyploidy and/or hybridization has been noted in many groups of plants (STEBBrNs 1971). In Melastomataceae, REYr¢ER (1989) has stated that polyploidy does not appear to be more frequent in apomictic species than in those where apomixis does not occur. However, if the analysis is restricted to tribe Miconieae, a much clearer relationship is evident. The most frequent chromosome number in this tribe is n=17 and, for this reason, it has been considered to be the basic number in the group (ALMEDA & Cn~AN~ 1992). These authors suggest that the number n=23 found in Clidemia ruddae - apparently an apomictic species - may be of polyploid origin. Similarly, other apomictic species for which counts are available almost all have a haploid chromosome number higher than n=17 while species shown to have sexual reproduction always have the basic number (Table 3). Exceptions are Clidemia capitellata and Maieta guianensis, apomictic species with n=17. It seems, therefore, that within tribe Miconieae a very close association exists between higher chromosome numbers, probably of polyploid origin, and apomixis.
Chromosome counts from M. albicans and other species of Melastomataceae (SOLT & Wtn~ACK 1980) show that samples from different regions may have different numbers. In rauch the same way, other species show apparent hetero- geneity in breeding system, as noted for M. dodecandra and M. laevigata (Table 3). It is possible that populations of widespread species may vary in ploidy level and in degree of apomixis, and this may explain some of the exceptions noted above,

22 R. GOI~DZN~ERG & G. J. SrmpI~EgD:
Table 3. Chromosome numbers for some species of Melastomataceae, tribe Miconieae. All counts cited are from SoLT & WURDACK (1980), except C. ruddae (ALME9A & CHUANG 1992), M. fallax and M. stenostachya (original). Miconia dodecandra is reported to have sexual reproduction by SOB~VmA & AR~OYO (1982), but Atlantic Rain Forest populations show very low pollen fertility (pers. obs.); M. laevigata is cited as sexually reproducing by RzN~~ (1989) and as apomictic by DENT-A¢osTA & B~CKO~ (1991)
Species Chromosome no.
Apomictic species Clidemia capitellata n=17 C. ruddae n=23 Maieta guianensis 2n=34 M. poeppigii 2n=46 Miconia albicans 2n=34 and 2n=48 M. fallax n=ca. 17 M. ibaguensis 2n=62 M. prasina 2n= 48-52 M. rubiginosa 2n=50 M. spinulosa 2n=46 M. stenostachya n=ca.26
Non-apomictic species Miconia araguensis n=17 M. calvescens 2n=34 M. ciliata n=17 M. macrodon n=17 M. minutiflora n=17 M. theaezans n=17 M. tuberculata n=17
Species with conflicting information Miconia dodecandra 2n=68 M. laevigata 2n=48
since reports of chromosome counts and breeding behaviour were not based on the same populations.
The counts made here for M. fallax and M. stenostachya gave numbers of n=c. 17 and n=c. 26 respectively. Although it was not possible to establish a precise number for M. stenostachya, the count obtained suggests that the population may be triploid (3x=52, close to 34 + 17). The meiotic irregularities observed in both are a strong indication of a hybrid origin for these species.
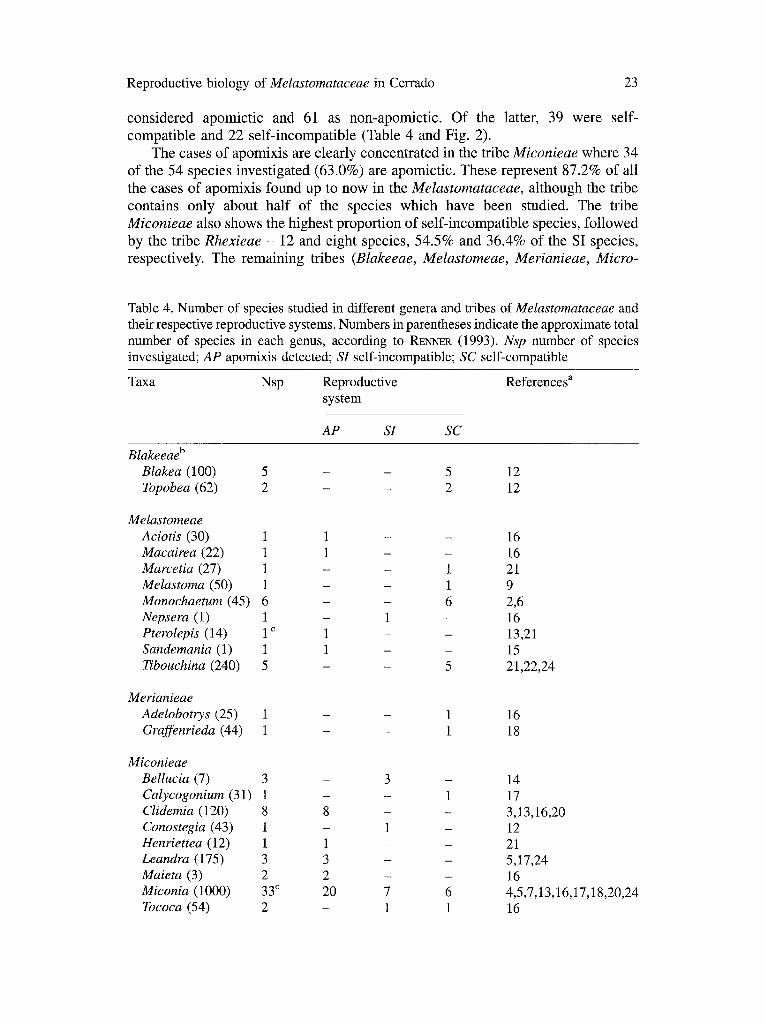
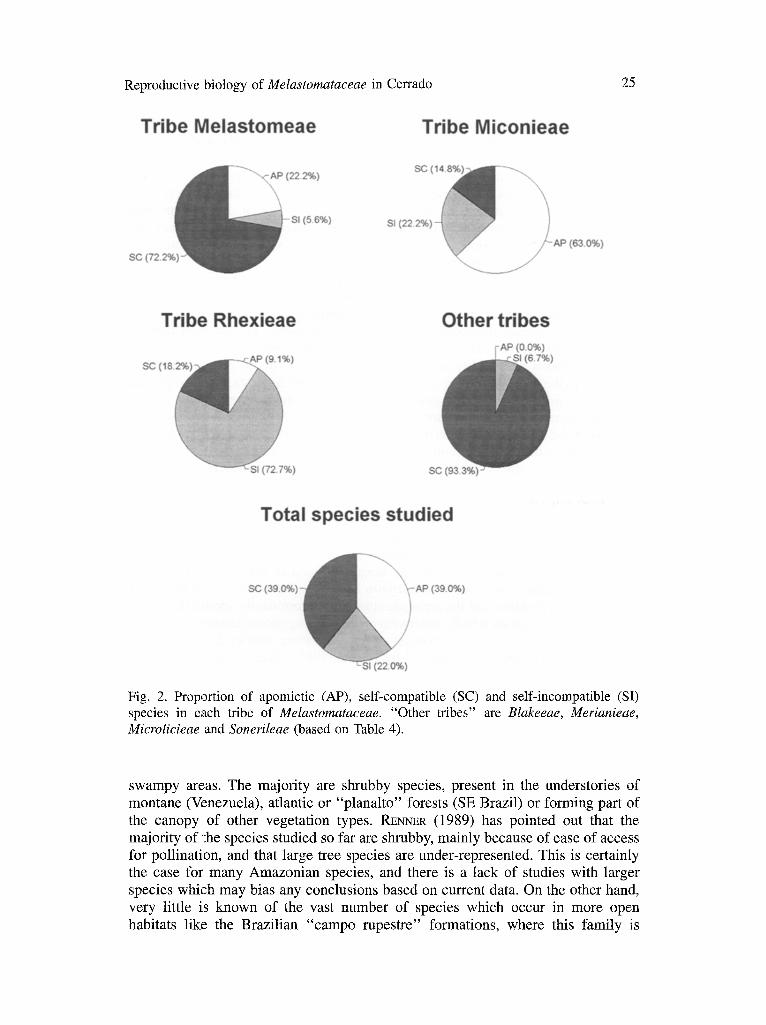
Apomixis in the Melastomataceae in general. The high proportion of apomictic species of Melastomataceae occurring in cerrado is orte of the more striking aspects of the data reported here. In fact, these data confirm a tendency which has been noted in other studies but not commented on in much detail. Amongst the 100 species of Melastomataceae whose breeding system has been investigated and reported in the literature or in the present study, 39 were
Reproductive biology of Melastomataceae in Cerrado 23
c o n s i d e r e d a p o m i c t i c a nd 61 as n o n - a p o m i c t i c . O f the latter, 39 w e r e self-
c o m p a t i b l e a nd 22 s e l f - i n c o m p a t i b l e (Table 4 and Fig. 2).
T h e cases o f a p o m i x i s are c l ea r ly c o n c e n t r a t e d in the t r ibe Miconieae w h e r e 34
o f the 54 spec ies i nves t iga t ed (63 .0%) are a p o m i c t i c . T h e s e r ep resen t 8 7 . 2 % o f all
the cases o f a p o m i x i s f o u n d up to n o w in the Melastomataceae, a l t h o u g h the t r ibe
c o n t a i n s o n l y a b o u t h a l f o f the spec ies w h i c h h a v e b e e n s tudied. T h e tr ibe
Miconieae also s h o w s the h i g h e s t p r o p o r t i o n o f s e l f - i n c o m p a t i b l e species , f o l l o w e d
b y the t r ibe Rhexieae - 12 and e igh t spec ies , 5 4 . 5 % and 3 6 . 4 % o f the SI spec ies ,
r e spec t ive ly . T h e r e m a i n i n g t r ibes (Blakeeae, Melastomeae, Merianieae, Micro-
Table 4. Number of species studied in different genera and tribes of Melastomataceae and their respective reproductive systems. Numbers in parentheses indicate the approximate total number of species in each genus, according to R e c k (1993). Nsp number o f species investigated; AP apomixis detected; SI self-incompatible; SC self-compatible
Taxa Nsp Reproduct ive References a system
AP SI SC
Blakeeae b Blakea (100) 5 - - 5 12 Topobea (62) 2 - - 2 12
Melastomeae Aciotis (30) 1 1 - - 16 Macairea (22) 1 1 - - 16 Marcetia (27) 1 - - 1 21 Melastoma (50) 1 - - 1 9 Monochaetum (45) 6 - - 6 2,6 Nepsera (1) 1 - 1 - 16 Pterolepis (14) 1 c 1 - - 13,21 Sandemania (1) 1 1 - - 15 Tibouchina (240) 5 - - 5 21,22,24
Merianieae Adelobotrys (25) 1 - - 1 Graffenrieda (44) 1 - - 1
16 18
Miconieae Bellucia (7) 3 - 3 - 14 Calycogonium (31) 1 - - 1 17 Clidemia (120) 8 8 - - 3,13,16,20 Conostegia (43) 1 - 1 - 12 Henriettea (12) 1 1 - - 21 Leandra (175) 3 3 - - 5,17,24 Maieta (3) 2 2 - - 16 Miconia (1000) 33 c 20 7 6 4,5,7,13,16,17,18,20,24 Tococa (54) 2 - 1 1 16
24
Table 4 (continued)
R. GOLDENBERG & G. J. SHEPHERD;
Taxa Nsp Reproductive References a system
AP SI SC
Microlicieae Rhynchanthera (15) 3 - - 3 10,16,21 TrembIeya (11) 1 - - 1 23
Rhexieae Rhexia (13) 11 c 1 8 2 8,11
Sonerileae Bertolonia (18) 1 - - 1 19 Centradenia (4) 3 - 1 2 1,6
a References. 1 ALMEDA (1977); 2 ALMEDA (1978); 3/~kLMEDA & CHUANG (1992); 4 BAUMGRAWZ & SILVA (1986/1988); 5 BORGES (1991); 6 DARWIN (1876, apud RENNER 1989); 7 DENT=AcoSTA & BRECKON (1991); 8 ETHRIDGE & HERR (1968); 9 GROSS (1993); 10 GU~MARÄES & al. (1993); I 1 KRAL & BOSTICK (1969, apud RENNER 1989); 12 LUMER (1982, apud RENNER 1989); 13 RAMIREZ Æ BRITO (1990); 14 RENNER (1987); 15 RENNER (1987); 16 RENNER (1989); 17 SARAWA (1994); 18 SOBREVmA & ARROYO (1982); 19 ZIEGLER (1925, apud RENNER 1989); 20 MELO (1995); 21 PINHEIRO (1995); 22 P. J. F. GUIMARÄES, pers. comm.; 23 J. SEMIR, pers. comm.; 24 original b Membership of tribes follows RENNER (1993) ° Some species were investigated in more than one study and gave different results. When a smdy encountered apomixy or not, for this Table the citation of apomixis had priority. This is the case of Pterolepis glomerata, Miconia laevigata, M. langsdorffii and Rhexia mariana
licieae and Sonerileae) have only four apomictic and two self-incompatible species in a total of 35. The remaining 29 species appear to be sexually-reproducing and self-compatible.
Generalizations based on a sample of only 100 species in a family containing 4200-4600 species must be treated with considerable caution, but the distribution observed in Table 4 appears to be consistent with pattems which have previously been observed in other apomictic groups. In the first place, apomictic groups often show a high frequency of self-incompatibility (RICHARDS 1986), and this appears to be confirmed in the case of tribe Miconieae, even with a relatively small number of
species. Moaiz (1992) has suggested that some taxonomic groups appear to present pre-
adaptations which facilitate the development of apomictic embryos, and this appears to be supported by the fact that 75% of 300 cases of apomixis described in the literature belong to the families Asteraceae, Rosaceae and Poaceae (RICHARDS 1986). This also seems to be the case for Melastomataceae, and is particularly striking for the tribe Miconieae. There is little evidence of any clear relationship with habitat or habit. The apomictic Melastomataceae occur in diverse habitats in tropical America. Of the species reported to be apomictic, 16 occur in savannas, ten in forest formations, seven in disturbed/ruderal formations and seven in marshes or
Reproductive biology of Melastomataceae in Cerrado
Tribe Melastomeae Tribe Miconieae
25
SC (14.8% (22.2%)
SI (5.6%) SI (22.2%)
BAP (63.0%)
SC (72.2%)
Tribe Rhexieae
sc (18.
Other tribes rAP (0.0%)
. )
sc (93.3%)
Total species studied
SC (39 0%)-~IAP/ (39 0%)
(22.0%)
Fig. 2. Proportion of apomictic (AP), self-compatible (SC) and self-incompatible (SI) species in each tribe of Melastomataceae. "Other tribes" are Blakeeae, Merianieae, Microlicieae and Sonerileae (based on Table 4).
swampy areas. The majority are shrubby species, present in the understories of montane (Venezuela), atlantic or "planalto" forests (SE Brazil) or forming part of the canopy of other vegetation types. REN~R (1989) has pointed out that the majority of 1Lhe species studied so far are shrubby, mainly because of ease of access for pollination, and that large tree species are under-represented. This is certainly the case for many Amazonian species, and there is a lack of studies with larger species which may bias any conclusions based on current data. On the other hand, very little is known of the vast number of species which occur in more open habitats like the Brazilian "campo rupestre" formations, where this family is

26 R. GOLDENBERG Æ G. J. SHEPHERD:
particularly weil represented and where most of the species are small to medium- sized shrubs. In spite of the possible bias in the current data set, the wide variation in habitats and habits of the species studied so far suggests that the major factor determining the distribution of apomixis in this family is phylogenetic. The fact that a species belongs to the tribe Miconieae appears to be a rauch more reliable predictor of the occurrence of apomixis than habit or habitat. In other tropical groups, this also appears to be the case in Dipterocarpaceae, a South-Asian rain forest family, where apomixis appears to be relatively widespread, occurring in emergents and understorey species (KAUR & al. 1978).
In the predominantly herbaceous apomictic groups that have received intensive study in temperate regions, it has been noted that many agamospecies have relatively wide distributions in comparison to sexually reproducing relatives, and may occur at higher altitudes/latitudes or more xerophytic or perturbed habitats (BIERZYCHUDEK 1987). This may represent greater colonizing ability of the apomictic forms or inability of sexually-reproducing forms to compete in these habitats. In the case ofAntennaria (Asteraceae) in North America, for example, the sexually reproducing species generally occur in more southerly parts of the distribution range while agamospecies are commoner in the northern regions, as if following the retreat of the Pleistocene glaciation (BAYER 1990). Similar patterns have been noted for polyploids and their diploid relatives (STEBBINS 1971). Amongst the apomictic species of tribe Miconieae, it seems clear that several supposed agamospecies such as M. albicans, M. fallax, M. stenostachya and M. rubiginosa have wider distributions than sexual taxa, occurring from Central America to southeast Brazil. This is also the case for M. laevigata and M. prasina (REN-~rER 1989). This pattern is less clear for L. lacunosa, M. ligustroides and M. pohliana that are restricted to the central-west and southeastern regions of Brazil. Of the sexually-reproducing species, M. langsdorffii and M. pepericarpa have more restricted distributions while M. minutißora has a distribution similar to that of M. albicans. It is possible that the areas occupied by predominantly apomictic species may represent regions which underwent extensive vegetation change during the Pleistocene while sexually-reproducing species are more restricted to areas in or near Pleistocene refuges (as defined by PRANCE 1987). There is certainly a very high level of diversity in the tribe Miconieae in some of the proposed refuge areas in the Atlantic Forest region, from the states of Espirito Santo to Säo Paulo in SE Brazil, but we know very little of breeding systems in these areas and further discussion must await more intensive studies of species of the Atlantic forest. The taxonomy of the group, especially in Miconia and Leandra, is also still in a state of flux and extensive revisions are necessary for a more complete discussion of breeding systems and geographical distribution.
It is also unclear, at present, to what extent the breeding system may vary within currently accepted species. The morphological variation observed in populations of M. albicans suggests that some degree of sexual reproduction may occur in some populations in some part of their ranges, but these must be very restricted since all the material studied up to now shows almost complete pollen sterility.
Little is known of the mechanisms of apomixis in tropical Melastomataceae. It has been suggested that tropical apomicts generally have adventitious embryony

Reproductive biology of Melastomataceae in Cerrado 27
while apospory and diplospory are commoner in temperate groups, but this is based on very limited investigations (BAKER & al. 1983 do not cite any Melastomata- ceae). Besides some polyembryony reports (RZNNZR 1989) which indicate adventitious embryony, the only data we have been able to locate on Melastomataceae are those of BORGES (1991; and BOR~ZS & al., unpubl.), who found evidence for nucellar embryos (adventitious embryony) and normal sexual embryos in Miconia discolor, while Leandra australis showed no formation of normal embryo sacs but produced nucellar embryos and embryos derived from unreduced embryo sacs which were probably diplosporous. In the latter case, the plants appeared to be totally apomictic, with no viable pollen produced. Both of these species occurred in semideciduous mesophytic forest or marshy areas on the borders of these forests. No data are available for cerrado species.
Conclusions
If the data reported here and in the literature available at present are representative of breeding systems in Melastomataceae as a whole, we may expect a total of around c. 1300 apomictic species within the tribe Miconieae, considering that this tribe possesses c. 2100 species (RENNER 1993) and that the proportion of apomictic species is constant in the tribe (63.0%). In this case, the family Melastomataceae and, in particular, the tribe Miconieae represent a major apomictic complex comparable to those in better-known families of temperate regions.
This is a predominantly woody group and nearly all of the apomicts detected in this family are small trees or shrubs, in strong contrast with families such as Asteraceae and Poaceae where most apomictic forms are herbaceous. Many of the habitat correlations observed for temperate apomictic groups are not applicable in this case, and discussions of the evolution of apomixis must take into account the different environmental conditions under which this complex has developed.
Extensive apomixis may also account for some of the taxonomic difficulty encountered in genera such as Miconia and Leandra where numerous rather similar species have been described and which may, in part, represent apomictic clones mixed with sexually reproducing forms. The family Melastomataceae in the neotropics therefore offers an interesting opportunity to test a number of ideas on the evolution and importance of apomixis in a rather different setting from that of the majority of apomictic complexes studied until now.
We thank ELIANA F. MARTnSS for her assistance with the cytological work, YA~A E BRESSAN for technical support, VOL~R BrrTP.ICH, AaY G. DA SmVA, ISABELA G. VARASSIN and two anonymous referees for comments on the manuscript. We thank Instituto Florestal (SP) and its staff in Itirapina for accomodation and permission to work in the cerrado. The first author received a grant from CAPES. Financial support was provided by FAEPAJNICAMR
References
ALMZDA, F., 1977: Systemafics of the neotropical genus Centradenia (Melastomataceae). - J. Arnold Arbor. 85: 73-108.
- 1978: Systemafics of the genus Monochaetum (Melastomataceae) in Mexico and Centra] Amefica. - Univ. Califonia Publ. Bot. 75: 1-134.

28 R. GOLDENBERG & G. J. SHEPHERD:
- CHVANC, T. I., 1992: Chromosome numbers and their systematic significance in some Mexican Melas tomataceae . - Syst. Bot. 17: 583-593.
BAKER, H. G., BAWA, K. S., FRANKIE, G. W., OPLER, P. A., 1983: Reproductive biology of plants in tropical forests. - In GOLLEY, F. B., (Ed.): Tropical rain forest ecosystems - structure and function. - Amsterdam: Elsevier.
BAUMGRATZ, J. E A., SILVA, N. M. E, 1988: Ecologia da polinizagäo e biologia da reprodu~äo de Miconia s tenostachya DC. (Melastomataceae) . - Rodriguesia 64/66: 1 1 - 2 3 .
BAYER, R. J., 1990: Patterns of clonal diversity in the Antennar ia rosea (Asteraceae) polyploid agamic complex. - Amer. J. Bot. 77: 1313-1319.
BERRY, P. E., TOBE, H., GOMEZ, J. A., 1991: Agamospermy and loss of distyly in Erythroxylum undulatum from Northern Venezuela. - Amer. J. Bot. 78: 595-600.
BIERZYCHUDEK, P., 1987: Patterns in plant parthenogenesis. - In STEARnS, S. C., (Ed.): The evolution of sex and its consequences. - Basel: Birkhäuser.
BORGES, H. B. N., 1991: Biologia reprodutiva de quatro espécies de Melas tomataceae . - M.Sc. Thesis, Universidade Estadual de Campinas, Brazil.
BURNHAM, C. R., 1962: Discussions in cytogenetics. - Minneapolis: Burgess. DENT-ACOSTA, S. R., BRECKON, G. J., 1991: Reproductive biology of six species of
Melas tomataceae in Western Porto Rico. - In: First International Melas tomataceae Symposium. - Washington: Smithsonian Institution.
ETHRIDGE, A. L., HERR, J. M., 1968: The development of the ovule and megagametophyte in Rhexia mariana. - Canad. J. Bot. 46: 133-139.
GEI~RY, A. H., 1974: Flowering phenology and diversity in tropical Bignoniaceae. -
Biotropica 6: 64-68. GIANNOTTI, E., 1988: Composi~äo flor/stica e estrutura fitossociológica da vegeta~äo de
cerrado e mata ciliar da Estaqäo Experimental de Itirapina (SP). - M.Sc. Thesis, Universidade Estadual de Campinas, Brazil.
GIBBS, P., 1990: Self-incompatibility in flowering plants: a neotropical perspective. - Revista Brasil. Bot. 13: 125-136.
GROSS, C. L., 1993: The breeding system and pollinators of M e l a s t o m a affine (Melas tomataceae) , a pioneer shrub in Tropical Australia. - Biotropica 25: 468-473.
GUtMARÄES, E J. E, STRANGHETTI, V., RANGA, N. T., 1993: Poliniza~äo e reprodu~äo de Rhynchanthera d ichotoma (LAM.) DC. (Melastomataceae) . - In: Anais do XX Colóquio de Incentivo ä Pesquisa. - Säo José R. Preto: UNESE
GUSTAFZSSON, A., 1946: Apomixis in higher plants. Part I. The mechanism of apomixis. - Acta Univ. Lund. 42: 1-66.
- 1947a: Apomixis in higher plants. Part II. Causal aspects. - Acta Univ. Lund. 43: 72- 178.
- 1947b: Apomixis in higher plants. Part III. Biotype and species formation. - Acta Univ. Lund. 43: 183-349.
HA, C. O., SANDS, V. E., SOEPA~MO, E., JOY6, K., 1988: Reproductive pattems of selected understorey trees in the Malaysian rain forest: the apomictic species. - Bot. J. Linn. Soc. 9 7 : 317-331.
KAUR, A., HA, C. O., JONC, K., SAYDS, V. E., CHAN, H. T., SOEPADMO, E., ASHTOn, P. S., 1978: Apomixis may be widespread among trees of the climax rain forest. - Nature 271: 404-442.
MAGNUSSON, W. E., SANAIOTTI, T. M., 1987: Dispersal of Miconia seeds by the rat Bolomys
lasiurus. - J. Trop. Ecol. 3: 277-278. MARTIN, E N., 1959: Staining and observing pollen tubes in the style by means of
fluorescence. - Stain Technol. 34: 125-128.

Reproductive biology of Melastomataceae in Cerrado 29
MELO, G. E A., 1995: Biologia floral e sistema reprodutivo de cinco espécies de Melastomataceae na mata de Dois Irmäos, Recife, PE. - M.Sc. Thesis, Universidade Federal de Pernambuco, Brazil.
MooiE, M., 1992: The evolution of asexual reproduction in plants. - London: Chapman & Hall.
MORI, S. A., PLPOLY, J. S., 1984: Observations on the Big Bang flowering of Miconia
minutiflora. - Brittonia 36: 337-341. NOaLER, G. A., 1984: Gametophytic apomixis. - In JOHRI, B. M., (Ed.): Embryology of
angiosperms. - Berlin, Heidelberg, New York: Springer. NYOP~N, A., 1954: Apomixis in angiosperms. Part II. - Bot. Rev. 20: 577-649. OLIV~rRA, R E. A. M., 1991: The pollination and reproductive biology of a cerrado woody
community in Brazil. - Ph.D. Thesis, University of St. Andrews, UK. - GmBS, E E., BARBOSA, A. A., TALAVERA, S., 1992: Contrasting breeding systems in two
Eriotheca (Bombacaceae) species of the Brazilian cerrados. - Pl. Syst. Evol. 179: 207- 219.
PIEDADE, L. H., RANGA, N. T., 1993: Ecologia da poliniza~äo de Galipea jasminiflora ENGLER (Rutaceae). - Revista Brasil. Bot. 16: 151-158.
PINHEmO, M. C. B., 1995: Biologia da reprodugäo de cinco espécies da Melastomataceae da Restinga de Maricä - RJ. - Doctoral Thesis, Universidade Estadual de Campinas, Brazil.
PRANCE, G. T., 1987: Biogeography of neotropical plants.- In WH1TMORE, T. C., PRANCE, G. T., (Eds): Biogeography and Quatemary history in Tropical Amefica. - Oxford: Clarendon Press.
RAMIREZ, N., BRITO, Y., 1990: Reproductive biology of a tropical palm swamp community in the Venezuelan llanos. - Amer. J. Bot. 77: 1260-1271.
RENNER, S. S., 1987: Reproductive biology of Bellucia (Melastomamceae) . - Acta Amazonica 16-17: 197-208.
- 1987: Sandemania hoehnei (Melastomataceae: Tibouchineae): taxonomy, distribution, and biology. - Brittonia 39: 441-446.
- 1989: A survey of reproductive biology in neotropical M e l a s t o m a m c e a e and M e m e c y l a c e a e . - Ann. Missouri Bot. Gar& 76: 496-518.
- 1993: Phylogeny and classification of the Melastomataceae and MemecyIaceae. - Nordic J. Bot. 13: 519-540.
RICHARDS, A. J., 1986: Plant breeding systems. - London: Allen & Unwin. SARArVA, L. C., 1994: Biologia da reproduqäo de arbustos e ärvores do cerrado em
Corumbataf, Estado de S. Paulo. - Doctoral Thesis, Universidade Estadual Paulista, Brazil.
SOBREVILA, C., ARROYO, M. T. K., 1982: Breeding systems in a montane tropical cloud forest in Venezuela. - Pl. Syst. Evol. 140: 19-37.
SOKAL, R. R., ROHLF, F. J., 1981: Biometry. - New York: Freeman. SOLT, M. L., WURDACK, J. J., 1980: Chromosome number in Melastomataceae. - Phytologia
47: 199-220. STEBmNS, G. L., 1971: Chromosomal evolution in higher plants. - London: Arnold.
Address ,of the authors: R. GOLDENBERG, G. J. SHEPHERD, Departamento de Botänica, Iß, UNICAMP, Caixa Postal 6109, CEP 13083-970, Cämpinas, SP, Brazil.
Accepted June 10, 1997 by V. H. HEYWOOD





























