Embed Size (px)
Citation preview

Suckling Stimulus Suppresses MessengerRNA for Tyrosine Hydroxylase in
Arcuate Neurons During Lactation
KATHIE A. BERGHORN,1 WEI-WEI LE,2 THOMAS G. SHERMAN,3
AND GLORIA E. HOFFMAN2*1Laboratory for Pregnancy and Newborn Research, Department of Physiology, College of
Veterinary Medicine, Cornell University, Ithaca, New York 148532Department of Anatomy & Neurobiology, University of Maryland School
of Medicine, Baltimore MD 212013Departments of Physiology & Biophysics and Medicine, Division of Endocrinology &
Metabolism, Georgetown University Medical Center, Washington, DC 20007
ABSTRACTTyrosine hydroxylase (TH) mRNA in tuberoinfundibular dopamine (TIDA) neurons is
suppressed during lactation but rebounds upon pup removal. A time course of TH mRNAchanges after pup removal revealed three phases: (1) a nuclear phase (evident 1.5 hours afterpup removal, maximal at 3 hours) with TH mRNA appearing in 1 or 2 nuclear loci with littleor no change in cytoplasmic mRNA; (2) a cytoplasmic phase (noted 6 hours after pup removal,peaked 12–24 hours) with a significant increase in total TH mRNA levels mainly in thecytoplasm; and (3) a stabilization phase (24–48 hours after pup removal) when nuclearsignals were low and cytoplasmic RNA showed a slight decline with extension of RNAclusters into the cell dendrites. In rats whose pups could suckle only on one side, TH wasup-regulated only on the side contralateral to nipple blockade. These data indicate that aftersuckling terminates, TH up-regulation is evident at 1.5 hours, but 6 hours is needed beforethe cells transport sufficient mRNA into the cytoplasm. The rapid signaling of TH up-regulation stems from the fact that the TIDA neurons respond to neural signals fromtermination of suckling. J. Comp. Neurol. 438:423–432, 2001. © 2001 Wiley-Liss, Inc.
Indexing terms: in situ hybridization; nuclear transcription sites; hypothalamus; dopamine;
neuroendocrine; prolactin; gene expression
Tuberoinfundibular dopamine neurons (TIDA) inhibit pro-lactin secretion from the anterior pituitary. Their activitychanges under physiological conditions with large increaseson the day of ovulation (deMaria et al., 1998), during lacta-tion (Meites et al., 1972), and after stress (Lookingland et al.,1990; Kehoe et al., 1992). Of these conditions, lactation isunique. After parturition, TIDA neuronal function shiftsfrom tonic activity (as is seen during cycling) to maintainedsuppression as revealed by reductions of dopamine releasefrom the median eminence in response to suckling (de Greefet al., 1981; de Greef and Visser, 1981), decreased DA turn-over in the median eminence (Selmanoff and Wise, 1981;Demarest et al., 1983) and decreased expression of the rate-limiting enzyme for DA synthesis, tyrosine hydroxylase (TH)(Wang et al., 1993). After pup removal, TH expression re-turns (Wang et al., 1993), triggering inhibition of prolactinsecretion (Selmanoff and Wise, 1981) that contributes to thereturn of cycling.
A time course of TH mRNA changes initially indicatedthat mRNA levels first rose significantly at 6 hours afterpups were removed (Wang et al., 1993), a finding whichcould suggest that hormonal changes upon termination ofsuckling trigger the up-regulation of TH. Yet, evidencefrom other dopamine systems indicates that afferent neu-ral activity is key for stimulating TH expression.
Specifically, in the olfactory bulb, afferent activity de-termines the level of TH expression (Erlich et al., 1990;Baker and Farbman, 1993; Cho et al., 1996; Jin et al.,
Grant sponsor: NIH grants from NINDS; Grant number: NS 28477;Grant number: NS 28730.
*Correspondence to: Gloria E. Hoffman, Department of Anatomy andNeurobiology, University of Maryland, School of Medicine, 685 W. Balti-more Street, Baltimore MD 21201. E-mail: [email protected]
Received 22 January 2001; Revised 13 June 2001; Accepted 25 June 2001
THE JOURNAL OF COMPARATIVE NEUROLOGY 438:423–432 (2001)
© 2001 WILEY-LISS, INC.

1996). If TH expression in TIDA neurons is similarly reg-ulated, the TIDA neurons would respond rapidly to re-moval of the suckling stimulus, and up-regulation uponpup removal would proceed even when hormone levels aremaintained.
In several other systems, early changes in gene expres-sion are best reflected in changes in unprocessed hetero-nuclear RNA (Lee et al., 1991, 1992; Ma et al., 1997a,b). Soas to exploit the ability to detect heteronuclear TH RNA inaddition to retaining the capacity for measurement ofmature mRNA in the cytoplasm, we devised a quantita-tive approach that used a nonradioactive probe directedtoward a single exon of the TH gene. After determiningexactly when changes in heteronuclear RNA within thecell nucleus and mature mRNA in the cell cytoplasm ofTIDA neurons occurred after the suckling stimulus wasterminated, we devised a paradigm in which pups weredenied access to the teats on one side of the body to studythe neurally driven changes in TH expression neurons onthe TIDA cells ipsilateral and contralateral to the nippleblock.
MATERIALS AND METHODS
Animals
Adult female timed-pregnant Sprague-Dawley ratswere maintained on a 12-hour light:12-hour dark schedule(lights on 6:00 AM to 6:00 PM) and given free access to foodand water. All rats delivered on gestational day 21 or 22and litter size was adjusted to 8 pups per rat on postpar-tum day 2. The collection of some of the tissue in this studywas made at the University of Pittsburgh before Dr. Hoff-man’s move to the University of Maryland. Thus, both theUniversity of Pittsburgh’s and the University of Mary-land’s Committees on Animal Care and Use approved allexperimental paradigms according to NIH guidelines.
Experiment 1: Determining the timing of initial
mRNA up-regulation of TH gene expression. For de-termining the time of onset of changes in TH expressionafter pup removal, lactating dams (n 5 3/group) 9 dayspostpartum were either kept with their pups or had theirpups removed to a separate room for periods of 1.5, 3, 6,12, 24, or 48 hours before euthanasia. At the time ofeuthanasia, each rat was anesthetized with an overdose ofpentobarbital (100 mg/kg, i.p.) and perfused with 2.5%acrolein (EM Grade, Polysciences, Warrington, PA) inphosphate-buffered 4% paraformaldehyde (pH 6.7) as pre-viously described (Hoffman et al., 1992). After fixation, thebrain was removed, sunk in 30% aqueous sucrose and cutinto serial 25-mm sections on a freezing microtome. Serialsections were collected into 1-in-12 series, placed in tissueculture wells containing cryoprotectant solution (Watsonet al., 1986), and maintained at 220°C until in situ hy-bridization and/or immunocytochemistry procedures wereinitiated.
Experiment 2. Limiting the effect of unilateral suck-
ling to one side of the body. For determining the effectof the location of the suckling stimulus on TH expressionin TIDA neurons, a group of lactating rats (n 5 8) had theteats on one side of the body covered with Nexaband(Veterinary Products Laboratories, Phoenix, AZ) so thatelevations of the nipples were masked, preventing suck-ling on that side. To prevent competition for the intactside, pup number was reduced from 8 to 4 upon return.
Twenty-four hours later, the animals were anesthetizedand perfused for mRNA localization as described above.
In situ hybridization
Probe preparation. The pGEM3-TH39 construct con-tains a 475 base pair EcoRI/HindIII fragment correspond-ing to amino acids 219-377 of the rat TH enzyme. This THfragment was derived from the RR1.2 plasmid obtainedfrom Dr. D.M. Chikaraishi (Duke University). For anti-sense TH riboprobes (cRNA), the plasmid was linearizedwith HindIII and transcribed with T7 RNA polymerase,yielding a 509 nucleotide complementary RNA (cRNA).The in vitro transcription reaction mixture contained 2.0mM Biotin-14-CTP (Life Technologies, Gaithersburg,MD), 1 mg HindIII-linearized-pGEM3z-TH39, 5 mM DTT,50 units RNAsin ribonuclease inhibitor (Promega, Madi-son, WI), 4 units T7 RNA polymerase (Life Technologies),0.5 mM CTP, and 1.0 mM each of ATP, GTP, and UTP.The transcription reaction was stopped by the addition of1 ml of EDTA. For an mRNA sense probe, the pGEM3-TH39 plasmid was linearized with EcoRI and transcribedwith SP6 RNA polymerase.
Hybridization and detection. The general strategyinvolved a hybridization with biotinylated riboprobes fol-lowed by immunohistochemical detection of the biotin. Itis summarized as follows:
Day 1. Freely floating 25-mm tissue sections encom-passing the entirety of the hypothalamic arcuate nucleuswere removed from cryoprotectant and placed into mi-crobeakers containing potassium phosphate buffered sa-line (KPBS, 0.05 M, pH 7.4) in diethylpyrocarbonate(DEPC) -treated water. Cryoprotectant was rinsed fromthe sections, and the tissue was incubated in 1% sodiumborohydride (NaBH4) in KPBS for 20 minutes at roomtemperature (RT). The NaBH4 was rinsed out of the tissuesections and ISH procedures were initiated. Sections wererinsed twice in 0.1 M triethanolamine buffer (TEA, pH8.0) and then incubated in 0.25% acetic anhydride in 0.1 Mtriethanolamine buffer (TEA, pH 8.0) for 10 minutes atRT. The tissue was then rinsed in 23 SSC (0.3 M NaCl,0.33 M Na citrate, pH 7.0) for 10 minutes at RT. Prehy-bridization was conducted for 2 hours at 37°C by usinghybridization buffer (50% deionized formamide, 10% dex-tran sulfate, 13 Denhardt’s solution, 0.3 M NaCl, 8 mMTris pH 8.0, 0.8 mM EDTA, 15% DPEC H2O) containingheat denatured torula yeast RNA (0.1 mg/ml; 5 Prime-3Prime Inc, Boulder, CO). Tissue sections were then rinsedfor 10 minutes with 23 SSC. The biotinylated TH probe(final concentration 600 ng/kbase per ml) and torula yeastRNA were combined and heat denatured at 100°C for 5minutes, added to fresh hybridization buffer, and placedin beakers for incubation with the tissue sections. Thetissue was added, the microbeakers were covered withParafilm, and the hybridization was continued overnightat 37°C.
Day 2. Tissue was rinsed with 43 SSC for 40 minutes,followed by incubation in 10 mM Tris-HCl, pH 8.0, con-taining 0.5 M NaCl and 20 mg/ml DNase-free RNase A(Boehringer Mannheim, Indianapolis, IN) at 37°C for 30minutes. Sections were rinsed and then incubated inRNase buffer (0.5 M NaCl, 0.05 M EDTA, 10 mM Tris, pH8.0) for 30 minutes. After 40 minutes of rinsing with fourchanges of 23 SSC, the sections were incubated in 0.13SSC for 60 minutes at 42°C. Tissue was rinsed once with0.13 SSC at RT and then rinsed with KPBS for 60 min-
424 K.A. BERGHORN ET AL.

utes. Sections were then incubated in goat anti-biotin(Vector Laboratories, Burlingame, CA) at a concentrationof 1:60,000 in KPBS with 0.4% Triton X-100 at 4°C for40–48 hours.
Day 4. The tissue was rinsed with 10 changes of KPBSover 60 minutes and then incubated for 1 hour at RT inbiotinylated donkey anti-goat IgG (heavy and light chains;Vector Laboratories) at a concentration of 1:600 in KPBSwith 0.4% Triton X-100. After rinsing, tissue was incu-bated in avidin-biotin complex solution (4.5 ml/ml KPBSwith 0.4% Triton X-100; Elite ABC kit, Vector Laborato-ries) for 1 hour at RT. Tissue was first rinsed 3 or 4 timeswith KPBS, then 3 to 4 times with 0.175 M sodium ace-tate. The biotin antibody peroxidase complex was stainedwith a solution of 0.175 M sodium acetate containing 12.5mg/ml nickel sulfate, 0.1 mg/ml 3,3 diaminobenzidine-HCl(DAB), and 0.42 ml/ml of 3% H2O2. This staining solutionwas more dilute than is normally used for ABC immuno-cytochemistry to keep background staining to a minimum.Intermittent viewing under a microscope monitored thechromogen reaction. Staining proceeded generally for30–45 minutes. Placing the tissue into 0.175 M sodiumacetate stopped the chromogen reaction. After rinsingwith KPBS, the tissue was either transferred into salineand mounted onto subbed gelatin/chrome glass slides orimmunocytochemically stained for TH protein as de-scribed below. After drying overnight, the tissue slideswere dehydrated in graded alcohols, cleared in Histoclear(National Diagnostics, Atlanta, GA) and cover-slippedwith Histomount (National Diagnostics).
Immunocytochemistry
When conducting double labeling of slides for mRNAand TH protein, the following protocol was performed:After the nonradioactive ISH chromogen reaction, the tis-sue was rinsed in saline followed by KPBS for 60 minutesand then incubated in a monoclonal tyrosine hydroxylaseantibody (MAB 318; Chemicon, Temecula, CA) at a con-centration of 1:200,000 in KPBS with 0.4% Triton X-100 at4°C for 48 hours. After rinsing, the tissue was incubated inbiotinylated goat anti-mouse IgG (heavy and light chains;Vector Laboratories) at a concentration of 1:600 in KPBSwith 0.4% Triton X-100 for 1 hour at RT, rinsed, andincubated in avidin-biotin complex solution (4.5 ml/mlKPBS with 0.4% Triton X-100; Elite ABC kit, Vector Lab-oratories) for 1 hour at room temperature. After rinsing inKPBS, the tissue was placed into Tris-HCl (pH 7.2) andthe TH antibody complex was stained with 0.2 mg/ml DABin Tris-HCl and 0.83 ml/ml of 3% H2O2 for approximately5–7 minutes. Sections were monitored under a microscopeto make sure that the TH protein stain (DAB) was darkenough to distinguish, yet not so dark as evoke nonspecificbackground staining that obscured the in situ hybridiza-tion signal. The tissue was then transferred to Tris-HCl tostop the reaction, rinsed with KPBS, and mounted out ofsaline onto gelatin/chrome subbed slides. The sectionswere dried and cover-slipped as described above. To avoidthe necessity for color photography we used a slightlydifferent approach for data presented in the manuscript.After visualization of the mRNA with NiDAB, we rinsedthe sections and incubated them in anti-TH (1:150,000) for48 hours. The TH is then visualized with immunofluores-cence by using biotin amplification (Berghorn et al., 1994)with streptavidin CY3 or Texas Red as the fluorophore.
The biotinyl tyramine was purchased from New EnglandNuclear.
Image analysis
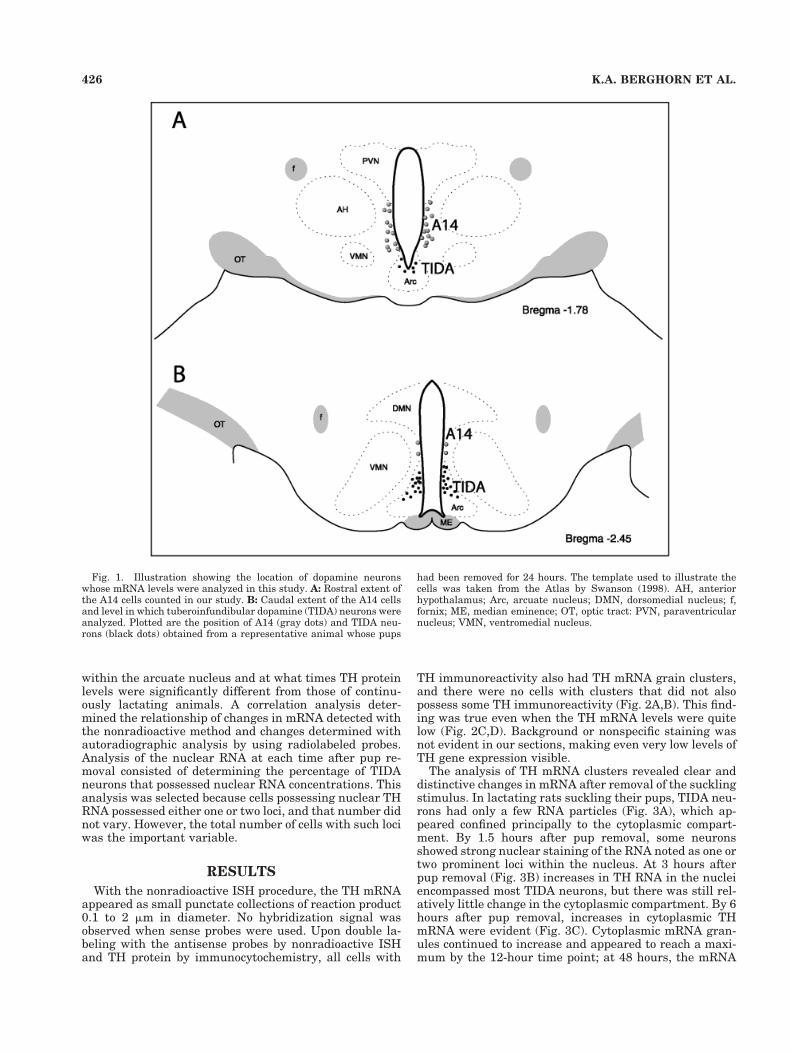
Analysis of the nonradioactive ISH data capitalized onthe fact that the cell’s RNA clusters could be resolved withNiDAB staining and counted with computerized imageanalysis. All analyses were performed on matched sec-tions taken from the mid-level arcuate nucleus where THcell numbers were maximal (Fig. 1B); all TIDA neuronswithin the arcuate nucleus were analyzed. The individualconducting the analysis was blind to the condition of theanimal. To encompass all the cells within each arcuatenucleus, multiple images were captured (by using a 403objective) from a Nikon Optiphot-equipped Edge Micro-scope (Edge Corp., Santa Monica, CA) or a Nikon Eclipse800 Microscope with a Photometrics Sensys Digital cam-era (Biovision Technologies, Exton, PA) and transferred toa Power Tower 200e computer (Power Computing, RoundRock, TX) for analysis. Signal Analytic’s IP Spectrum Soft-ware (Vienna, VA) was used for the analysis. An addi-tional analysis of the DA cells located within the periven-tricular zone of the hypothalamus (A14) was conducted. Inthat instance, cells were analyzed from the level of therostral arcuate nucleus to the mid TIDA group (as illus-trated in Fig. 1A,B) by using the same strategy describedabove.
Unlike autoradiographic silver grains, the nonradioac-tive ISH RNA product was distributed within rather thanabove the cells. As a result, analysis was performed onthose RNA clusters that were in focus at a single plane.Because the clusters’ darkness varied, we captured clus-ters in 10 gray level-unit bins starting from the darkestclusters and working progressively to the lightest grainclusters until all visible clusters were counted. A definedrange of accepted cluster sizes was set in the analysisprogram to eliminate counting of areas shadowed byunder- or overlying clusters or recounting grain clusterswhose borders were not as dark as the remainder of thecluster. The range of accepted cluster sizes was derivedfrom determinations in pilot experiments of individual THclusters that were well separated. For all the cells countedin a field, cell area measurements were obtained. Pilotstudies determined that the density of clusters did notvary considerably within each plane of the perikaryoncytoplasm, although RNA clusters were generally absentfrom the nucleus. So as not to dilute the counts with“negative” compartments, when a nucleus was in the focalplane its area was not included in the cell area measure-ments. Once the initial analysis was performed, the totalnumber of grains was summed for all the cells and clusterdensity (number of grains/cell area) was then calculated.To validate that nonradioactive ISH accurately revealedoverall changes in TH mRNA levels within TIDA neuronsafter pup removal, we performed a correlation analysis onthe nonradioactive ISH data (by using the summed clusternumber for each arcuate nucleus) from the present studyand grain counts/arcuate nucleus from a previous study(Wang et al., 1993) that used 35S-labeled probes in anidentical experimental paradigm.
Statistical analysis
A two-way analysis of variance followed by aBonferroni/Dunn post hoc analysis determined whetherthe time after pup removal affected the level of TH mRNA
425CHANGES IN TH mRNA DURING PUP REMOVAL
within the arcuate nucleus and at what times TH proteinlevels were significantly different from those of continu-ously lactating animals. A correlation analysis deter-mined the relationship of changes in mRNA detected withthe nonradioactive method and changes determined withautoradiographic analysis by using radiolabeled probes.Analysis of the nuclear RNA at each time after pup re-moval consisted of determining the percentage of TIDAneurons that possessed nuclear RNA concentrations. Thisanalysis was selected because cells possessing nuclear THRNA possessed either one or two loci, and that number didnot vary. However, the total number of cells with such lociwas the important variable.
RESULTS
With the nonradioactive ISH procedure, the TH mRNAappeared as small punctate collections of reaction product0.1 to 2 mm in diameter. No hybridization signal wasobserved when sense probes were used. Upon double la-beling with the antisense probes by nonradioactive ISHand TH protein by immunocytochemistry, all cells with
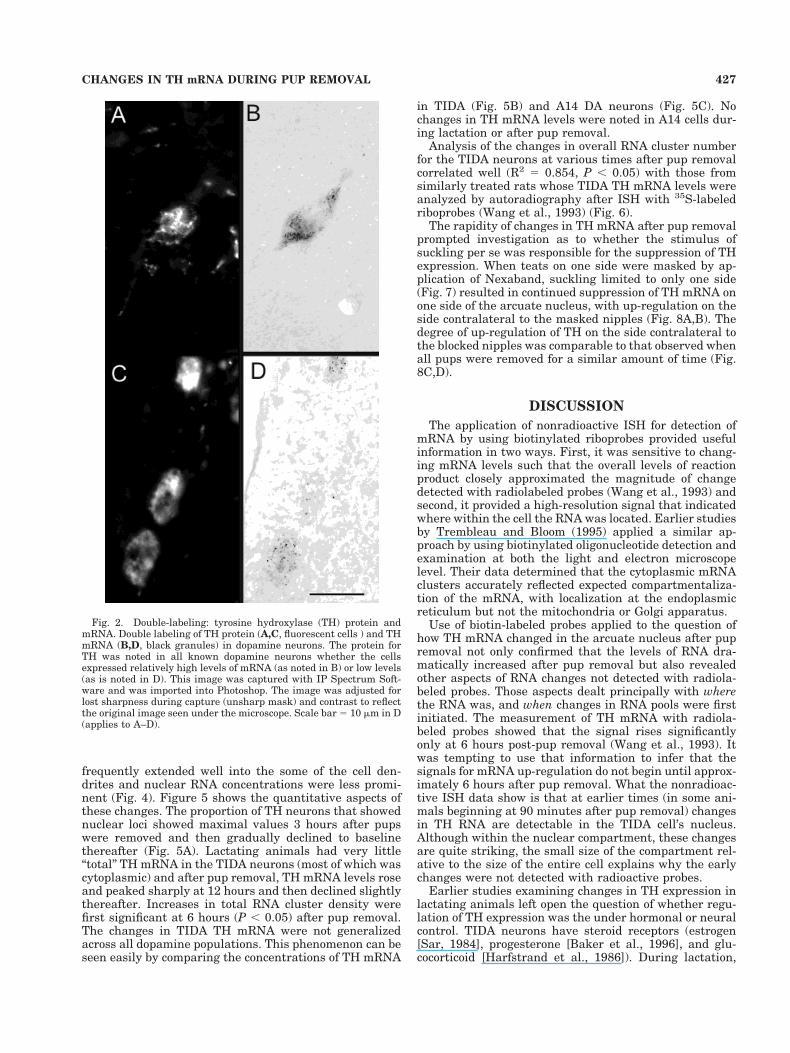
TH immunoreactivity also had TH mRNA grain clusters,and there were no cells with clusters that did not alsopossess some TH immunoreactivity (Fig. 2A,B). This find-ing was true even when the TH mRNA levels were quitelow (Fig. 2C,D). Background or nonspecific staining wasnot evident in our sections, making even very low levels ofTH gene expression visible.
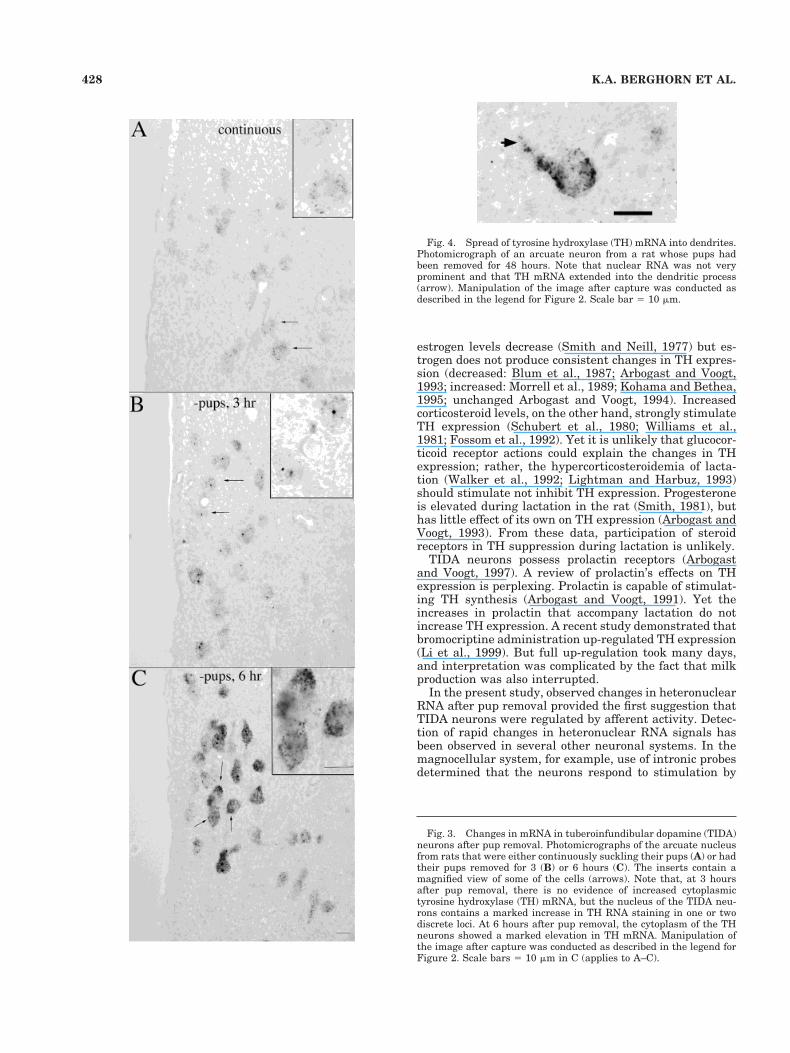
The analysis of TH mRNA clusters revealed clear anddistinctive changes in mRNA after removal of the sucklingstimulus. In lactating rats suckling their pups, TIDA neu-rons had only a few RNA particles (Fig. 3A), which ap-peared confined principally to the cytoplasmic compart-ment. By 1.5 hours after pup removal, some neuronsshowed strong nuclear staining of the RNA noted as one ortwo prominent loci within the nucleus. At 3 hours afterpup removal (Fig. 3B) increases in TH RNA in the nucleiencompassed most TIDA neurons, but there was still rel-atively little change in the cytoplasmic compartment. By 6hours after pup removal, increases in cytoplasmic THmRNA were evident (Fig. 3C). Cytoplasmic mRNA gran-ules continued to increase and appeared to reach a maxi-mum by the 12-hour time point; at 48 hours, the mRNA
Fig. 1. Illustration showing the location of dopamine neuronswhose mRNA levels were analyzed in this study. A: Rostral extent ofthe A14 cells counted in our study. B: Caudal extent of the A14 cellsand level in which tuberoinfundibular dopamine (TIDA) neurons wereanalyzed. Plotted are the position of A14 (gray dots) and TIDA neu-rons (black dots) obtained from a representative animal whose pups
had been removed for 24 hours. The template used to illustrate thecells was taken from the Atlas by Swanson (1998). AH, anteriorhypothalamus; Arc, arcuate nucleus; DMN, dorsomedial nucleus; f,fornix; ME, median eminence; OT, optic tract: PVN, paraventricularnucleus; VMN, ventromedial nucleus.
426 K.A. BERGHORN ET AL.
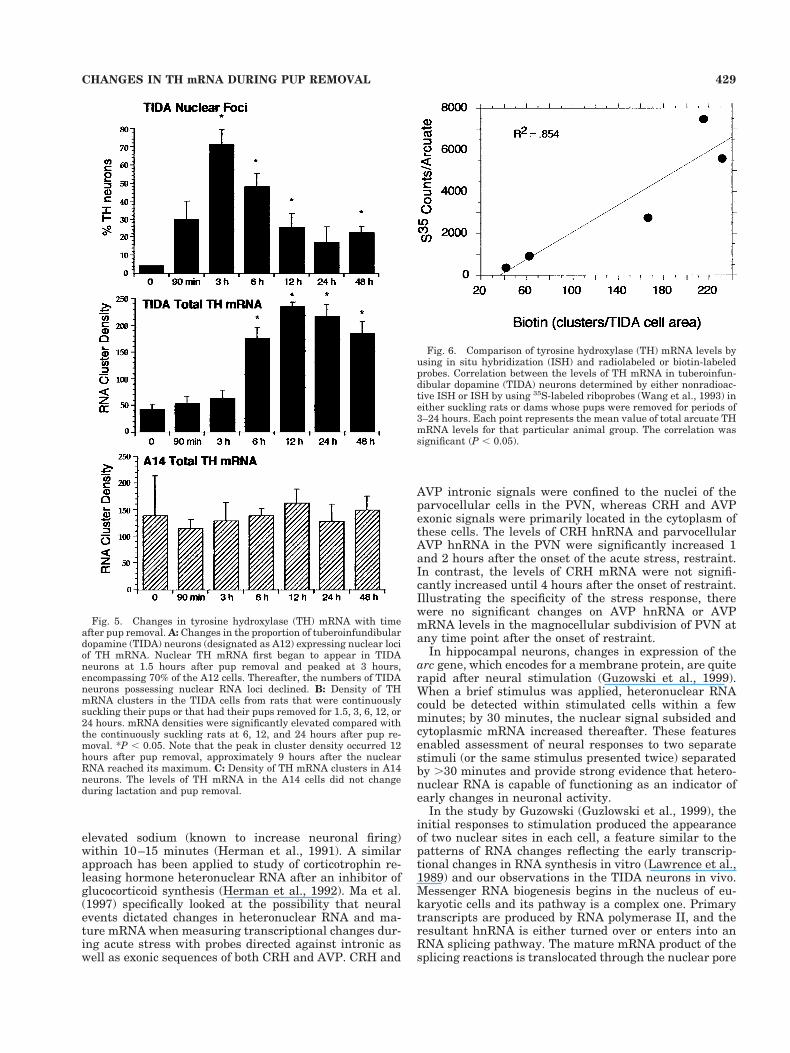
frequently extended well into the some of the cell den-drites and nuclear RNA concentrations were less promi-nent (Fig. 4). Figure 5 shows the quantitative aspects ofthese changes. The proportion of TH neurons that showednuclear loci showed maximal values 3 hours after pupswere removed and then gradually declined to baselinethereafter (Fig. 5A). Lactating animals had very little“total” TH mRNA in the TIDA neurons (most of which wascytoplasmic) and after pup removal, TH mRNA levels roseand peaked sharply at 12 hours and then declined slightlythereafter. Increases in total RNA cluster density werefirst significant at 6 hours (P , 0.05) after pup removal.The changes in TIDA TH mRNA were not generalizedacross all dopamine populations. This phenomenon can beseen easily by comparing the concentrations of TH mRNA
in TIDA (Fig. 5B) and A14 DA neurons (Fig. 5C). Nochanges in TH mRNA levels were noted in A14 cells dur-ing lactation or after pup removal.
Analysis of the changes in overall RNA cluster numberfor the TIDA neurons at various times after pup removalcorrelated well (R2 5 0.854, P , 0.05) with those fromsimilarly treated rats whose TIDA TH mRNA levels wereanalyzed by autoradiography after ISH with 35S-labeledriboprobes (Wang et al., 1993) (Fig. 6).
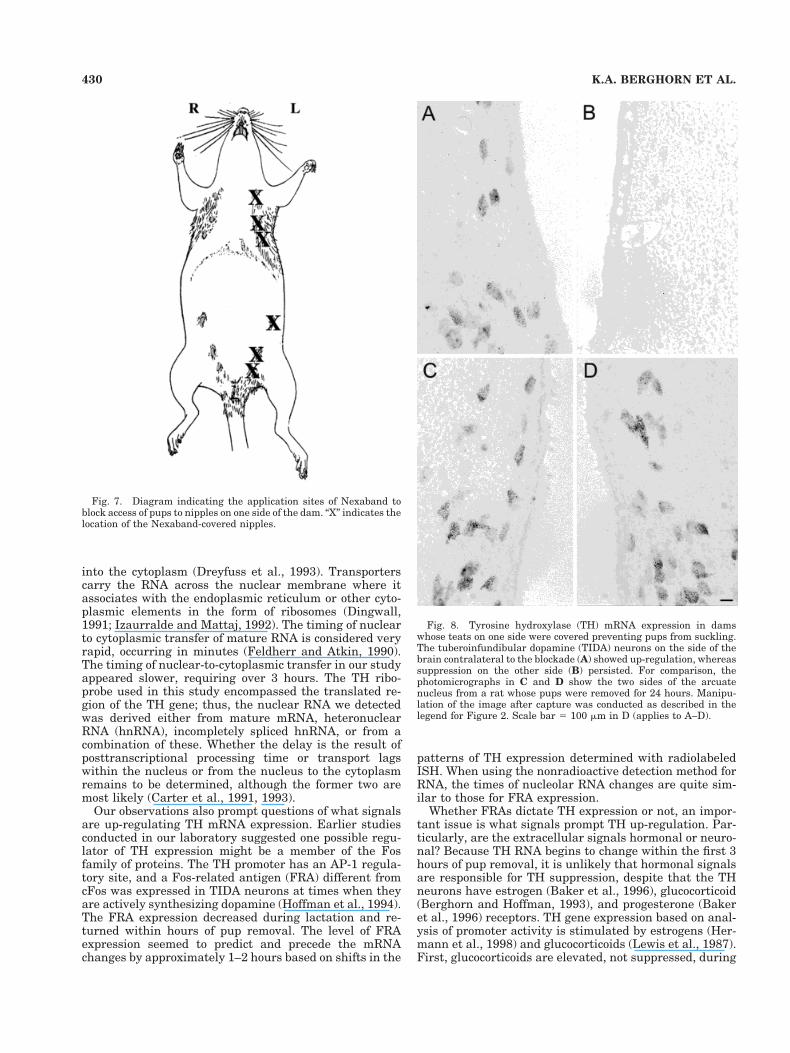
The rapidity of changes in TH mRNA after pup removalprompted investigation as to whether the stimulus ofsuckling per se was responsible for the suppression of THexpression. When teats on one side were masked by ap-plication of Nexaband, suckling limited to only one side(Fig. 7) resulted in continued suppression of TH mRNA onone side of the arcuate nucleus, with up-regulation on theside contralateral to the masked nipples (Fig. 8A,B). Thedegree of up-regulation of TH on the side contralateral tothe blocked nipples was comparable to that observed whenall pups were removed for a similar amount of time (Fig.8C,D).
DISCUSSION
The application of nonradioactive ISH for detection ofmRNA by using biotinylated riboprobes provided usefulinformation in two ways. First, it was sensitive to chang-ing mRNA levels such that the overall levels of reactionproduct closely approximated the magnitude of changedetected with radiolabeled probes (Wang et al., 1993) andsecond, it provided a high-resolution signal that indicatedwhere within the cell the RNA was located. Earlier studiesby Trembleau and Bloom (1995) applied a similar ap-proach by using biotinylated oligonucleotide detection andexamination at both the light and electron microscopelevel. Their data determined that the cytoplasmic mRNAclusters accurately reflected expected compartmentaliza-tion of the mRNA, with localization at the endoplasmicreticulum but not the mitochondria or Golgi apparatus.
Use of biotin-labeled probes applied to the question ofhow TH mRNA changed in the arcuate nucleus after pupremoval not only confirmed that the levels of RNA dra-matically increased after pup removal but also revealedother aspects of RNA changes not detected with radiola-beled probes. Those aspects dealt principally with wherethe RNA was, and when changes in RNA pools were firstinitiated. The measurement of TH mRNA with radiola-beled probes showed that the signal rises significantlyonly at 6 hours post-pup removal (Wang et al., 1993). Itwas tempting to use that information to infer that thesignals for mRNA up-regulation do not begin until approx-imately 6 hours after pup removal. What the nonradioac-tive ISH data show is that at earlier times (in some ani-mals beginning at 90 minutes after pup removal) changesin TH RNA are detectable in the TIDA cell’s nucleus.Although within the nuclear compartment, these changesare quite striking, the small size of the compartment rel-ative to the size of the entire cell explains why the earlychanges were not detected with radioactive probes.
Earlier studies examining changes in TH expression inlactating animals left open the question of whether regu-lation of TH expression was the under hormonal or neuralcontrol. TIDA neurons have steroid receptors (estrogen[Sar, 1984], progesterone [Baker et al., 1996], and glu-cocorticoid [Harfstrand et al., 1986]). During lactation,
Fig. 2. Double-labeling: tyrosine hydroxylase (TH) protein andmRNA. Double labeling of TH protein (A,C, fluorescent cells ) and THmRNA (B,D, black granules) in dopamine neurons. The protein forTH was noted in all known dopamine neurons whether the cellsexpressed relatively high levels of mRNA (as noted in B) or low levels(as is noted in D). This image was captured with IP Spectrum Soft-ware and was imported into Photoshop. The image was adjusted forlost sharpness during capture (unsharp mask) and contrast to reflectthe original image seen under the microscope. Scale bar 5 10 mm in D(applies to A–D).
427CHANGES IN TH mRNA DURING PUP REMOVAL
estrogen levels decrease (Smith and Neill, 1977) but es-trogen does not produce consistent changes in TH expres-sion (decreased: Blum et al., 1987; Arbogast and Voogt,1993; increased: Morrell et al., 1989; Kohama and Bethea,1995; unchanged Arbogast and Voogt, 1994). Increasedcorticosteroid levels, on the other hand, strongly stimulateTH expression (Schubert et al., 1980; Williams et al.,1981; Fossom et al., 1992). Yet it is unlikely that glucocor-ticoid receptor actions could explain the changes in THexpression; rather, the hypercorticosteroidemia of lacta-tion (Walker et al., 1992; Lightman and Harbuz, 1993)should stimulate not inhibit TH expression. Progesteroneis elevated during lactation in the rat (Smith, 1981), buthas little effect of its own on TH expression (Arbogast andVoogt, 1993). From these data, participation of steroidreceptors in TH suppression during lactation is unlikely.
TIDA neurons possess prolactin receptors (Arbogastand Voogt, 1997). A review of prolactin’s effects on THexpression is perplexing. Prolactin is capable of stimulat-ing TH synthesis (Arbogast and Voogt, 1991). Yet theincreases in prolactin that accompany lactation do notincrease TH expression. A recent study demonstrated thatbromocriptine administration up-regulated TH expression(Li et al., 1999). But full up-regulation took many days,and interpretation was complicated by the fact that milkproduction was also interrupted.
In the present study, observed changes in heteronuclearRNA after pup removal provided the first suggestion thatTIDA neurons were regulated by afferent activity. Detec-tion of rapid changes in heteronuclear RNA signals hasbeen observed in several other neuronal systems. In themagnocellular system, for example, use of intronic probesdetermined that the neurons respond to stimulation by
Fig. 4. Spread of tyrosine hydroxylase (TH) mRNA into dendrites.Photomicrograph of an arcuate neuron from a rat whose pups hadbeen removed for 48 hours. Note that nuclear RNA was not veryprominent and that TH mRNA extended into the dendritic process(arrow). Manipulation of the image after capture was conducted asdescribed in the legend for Figure 2. Scale bar 5 10 mm.
Fig. 3. Changes in mRNA in tuberoinfundibular dopamine (TIDA)neurons after pup removal. Photomicrographs of the arcuate nucleusfrom rats that were either continuously suckling their pups (A) or hadtheir pups removed for 3 (B) or 6 hours (C). The inserts contain amagnified view of some of the cells (arrows). Note that, at 3 hoursafter pup removal, there is no evidence of increased cytoplasmictyrosine hydroxylase (TH) mRNA, but the nucleus of the TIDA neu-rons contains a marked increase in TH RNA staining in one or twodiscrete loci. At 6 hours after pup removal, the cytoplasm of the THneurons showed a marked elevation in TH mRNA. Manipulation ofthe image after capture was conducted as described in the legend forFigure 2. Scale bars 5 10 mm in C (applies to A–C).
428 K.A. BERGHORN ET AL.
elevated sodium (known to increase neuronal firing)within 10–15 minutes (Herman et al., 1991). A similarapproach has been applied to study of corticotrophin re-leasing hormone heteronuclear RNA after an inhibitor ofglucocorticoid synthesis (Herman et al., 1992). Ma et al.(1997) specifically looked at the possibility that neuralevents dictated changes in heteronuclear RNA and ma-ture mRNA when measuring transcriptional changes dur-ing acute stress with probes directed against intronic aswell as exonic sequences of both CRH and AVP. CRH and
AVP intronic signals were confined to the nuclei of theparvocellular cells in the PVN, whereas CRH and AVPexonic signals were primarily located in the cytoplasm ofthese cells. The levels of CRH hnRNA and parvocellularAVP hnRNA in the PVN were significantly increased 1and 2 hours after the onset of the acute stress, restraint.In contrast, the levels of CRH mRNA were not signifi-cantly increased until 4 hours after the onset of restraint.Illustrating the specificity of the stress response, therewere no significant changes on AVP hnRNA or AVPmRNA levels in the magnocellular subdivision of PVN atany time point after the onset of restraint.
In hippocampal neurons, changes in expression of thearc gene, which encodes for a membrane protein, are quiterapid after neural stimulation (Guzowski et al., 1999).When a brief stimulus was applied, heteronuclear RNAcould be detected within stimulated cells within a fewminutes; by 30 minutes, the nuclear signal subsided andcytoplasmic mRNA increased thereafter. These featuresenabled assessment of neural responses to two separatestimuli (or the same stimulus presented twice) separatedby .30 minutes and provide strong evidence that hetero-nuclear RNA is capable of functioning as an indicator ofearly changes in neuronal activity.
In the study by Guzowski (Guzlowski et al., 1999), theinitial responses to stimulation produced the appearanceof two nuclear sites in each cell, a feature similar to thepatterns of RNA changes reflecting the early transcrip-tional changes in RNA synthesis in vitro (Lawrence et al.,1989) and our observations in the TIDA neurons in vivo.Messenger RNA biogenesis begins in the nucleus of eu-karyotic cells and its pathway is a complex one. Primarytranscripts are produced by RNA polymerase II, and theresultant hnRNA is either turned over or enters into anRNA splicing pathway. The mature mRNA product of thesplicing reactions is translocated through the nuclear pore
Fig. 5. Changes in tyrosine hydroxylase (TH) mRNA with timeafter pup removal. A: Changes in the proportion of tuberoinfundibulardopamine (TIDA) neurons (designated as A12) expressing nuclear lociof TH mRNA. Nuclear TH mRNA first began to appear in TIDAneurons at 1.5 hours after pup removal and peaked at 3 hours,encompassing 70% of the A12 cells. Thereafter, the numbers of TIDAneurons possessing nuclear RNA loci declined. B: Density of THmRNA clusters in the TIDA cells from rats that were continuouslysuckling their pups or that had their pups removed for 1.5, 3, 6, 12, or24 hours. mRNA densities were significantly elevated compared withthe continuously suckling rats at 6, 12, and 24 hours after pup re-moval. *P , 0.05. Note that the peak in cluster density occurred 12hours after pup removal, approximately 9 hours after the nuclearRNA reached its maximum. C: Density of TH mRNA clusters in A14neurons. The levels of TH mRNA in the A14 cells did not changeduring lactation and pup removal.
Fig. 6. Comparison of tyrosine hydroxylase (TH) mRNA levels byusing in situ hybridization (ISH) and radiolabeled or biotin-labeledprobes. Correlation between the levels of TH mRNA in tuberoinfun-dibular dopamine (TIDA) neurons determined by either nonradioac-tive ISH or ISH by using 35S-labeled riboprobes (Wang et al., 1993) ineither suckling rats or dams whose pups were removed for periods of3–24 hours. Each point represents the mean value of total arcuate THmRNA levels for that particular animal group. The correlation wassignificant (P , 0.05).
429CHANGES IN TH mRNA DURING PUP REMOVAL
into the cytoplasm (Dreyfuss et al., 1993). Transporterscarry the RNA across the nuclear membrane where itassociates with the endoplasmic reticulum or other cyto-plasmic elements in the form of ribosomes (Dingwall,1991; Izaurralde and Mattaj, 1992). The timing of nuclearto cytoplasmic transfer of mature RNA is considered veryrapid, occurring in minutes (Feldherr and Atkin, 1990).The timing of nuclear-to-cytoplasmic transfer in our studyappeared slower, requiring over 3 hours. The TH ribo-probe used in this study encompassed the translated re-gion of the TH gene; thus, the nuclear RNA we detectedwas derived either from mature mRNA, heteronuclearRNA (hnRNA), incompletely spliced hnRNA, or from acombination of these. Whether the delay is the result ofposttranscriptional processing time or transport lagswithin the nucleus or from the nucleus to the cytoplasmremains to be determined, although the former two aremost likely (Carter et al., 1991, 1993).
Our observations also prompt questions of what signalsare up-regulating TH mRNA expression. Earlier studiesconducted in our laboratory suggested one possible regu-lator of TH expression might be a member of the Fosfamily of proteins. The TH promoter has an AP-1 regula-tory site, and a Fos-related antigen (FRA) different fromcFos was expressed in TIDA neurons at times when theyare actively synthesizing dopamine (Hoffman et al., 1994).The FRA expression decreased during lactation and re-turned within hours of pup removal. The level of FRAexpression seemed to predict and precede the mRNAchanges by approximately 1–2 hours based on shifts in the
patterns of TH expression determined with radiolabeledISH. When using the nonradioactive detection method forRNA, the times of nucleolar RNA changes are quite sim-ilar to those for FRA expression.
Whether FRAs dictate TH expression or not, an impor-tant issue is what signals prompt TH up-regulation. Par-ticularly, are the extracellular signals hormonal or neuro-nal? Because TH RNA begins to change within the first 3hours of pup removal, it is unlikely that hormonal signalsare responsible for TH suppression, despite that the THneurons have estrogen (Baker et al., 1996), glucocorticoid(Berghorn and Hoffman, 1993), and progesterone (Bakeret al., 1996) receptors. TH gene expression based on anal-ysis of promoter activity is stimulated by estrogens (Her-mann et al., 1998) and glucocorticoids (Lewis et al., 1987).First, glucocorticoids are elevated, not suppressed, during
Fig. 7. Diagram indicating the application sites of Nexaband toblock access of pups to nipples on one side of the dam. “X” indicates thelocation of the Nexaband-covered nipples.
Fig. 8. Tyrosine hydroxylase (TH) mRNA expression in damswhose teats on one side were covered preventing pups from suckling.The tuberoinfundibular dopamine (TIDA) neurons on the side of thebrain contralateral to the blockade (A) showed up-regulation, whereassuppression on the other side (B) persisted. For comparison, thephotomicrographs in C and D show the two sides of the arcuatenucleus from a rat whose pups were removed for 24 hours. Manipu-lation of the image after capture was conducted as described in thelegend for Figure 2. Scale bar 5 100 mm in D (applies to A–D).
430 K.A. BERGHORN ET AL.

lactation. Thus, one would predict a decrease in glucocor-ticoid receptor-regulated TH expression after pup removalnot an increase. Progesterone, which has few direct ac-tions on TH regulation (Hermann et al., 1998) could beacting to block glucocorticoid effects on TH activitythrough prevention of glucocorticoid receptor binding toDNA (Hoffman et al., 1997). However, progesterone levelswould not be expected to fall in such a short time. Seventy-two hours is needed for progesterone levels to return tobaseline after pup removal (Lee et al., 1989). Prolactinlevels might be altered within that time frame, but phar-macologic blockade of prolactin secretion does not preventTH suppression during suckling (Li et al., 1997). In fact,bromocriptine-treated rats showed TH mRNA levels thatwere only slightly elevated 24 hours after suckling com-pared with suckled animals receiving vehicle.
More likely, the suppression of TH gene expression isdue to inhibitory afferent activity. Scientists have demon-strated marked changes in TH expression in the olfactorybulb as afferent stimulation varies (Erlich et al., 1990;Baker and Farbman, 1993; Cho et al., 1996; Jin et al.,1996). Preventing odorants from reaching the olfactoryepithelium results in a marked fall in TH expression.Reopening of the nares reestablishes olfactory functionand produces re-expression of TH mRNA and protein. Ourdata demonstrate that activity is also regulating TH ex-pression in the TIDA neurons. The prevention of sucklingon one set of teats markedly and rapidly up-regulated THexpression in TIDA neurons only on the side contralateralto the blocked nipples. These data suggest that the sen-sory stimulus to the nipples is inhibiting the TIDA neu-rons and rule out the possibility that only hormonal fac-tors drive the changes in TH expression. Analysis of theTH promoter reveals regulatory elements that respond tolevels of cyclic AMP as well as AP1 complexes (Ghee et al.,1998; Liu et al., 1998). It is through such a mechanismthat TH expression is likely determined in vivo.
Although we postulate that increased transcription ofTH is signaled at the time we observed increased levels ofRNA staining, it is also possible that some of the increasedRNA signals we observed resulted from a change in turn-over of mRNA owing to hormonal- or activity-dependentchanges in RNA degradation. Analysis of TH gene tran-scriptional activity by using intron-directed in situ hybrid-ization would address this possibility. Nonetheless, by 6hours after removal of the suckling stimulus, increasedTH RNA was located in the nucleus and cytoplasm. In apilot study, comparison of the data from Western blots tothe nonradioactive ISH data indicated that at 6 hoursafter pup removal, the increased TH mRNA in the TIDAcell cytoplasm was associated with an increase in theamount of TH protein in the median eminence (Hoffmanet al., 1999). What those changes suggest is that thecapacity to regulate prolactin is reestablished at thattime.
The TH mRNA extended into the dendritic compart-ment between 24 and 48 hours after pup removal. Thisobservation suggests that there are broad sites of THsynthesis extending beyond the perinuclear region. A re-duction in the RNA in nuclear sites occurred at this sametime and suggests some stabilization of RNA synthesis.Importantly, these relatively small changes in compart-mental TH RNA can be used to document dynamicchanges in TH regulation with varying endocrine status.
In summary, this study demonstrates that TH mRNA ischanging within a few hours of pup removal in a mannerconsistent with timing of the resurgence of dopamine’ssuppression of prolactin release. The study of TH expres-sion in dams with one set of teats masked preventingsuckling on that side determined that suckling stimula-tion and not hormonal factors produced the change in THexpression in the TIDA neurons.
ACKNOWLEDGMENTS
The authors thank Dr. Susan Wray, Dr. Frank Margo-lis, and Dr. Stanley Watson for their critical reading of themanuscript and constructive comments.
LITERATURE CITED
Arbogast LA, Voogt JL. 1991. Hyperprolactinemia increases and hypopro-lactinemia decreases tyrosine hydroxylase messenger ribonucleic acidlevels in the arcuate nuclei, but not the substantia nigra or zonaincerta. Endocrinology 128:997–1005.
Arbogast LA, Voogt JL. 1993. Progesterone reverses the estradiol-induceddecrease in tyrosine hydroxylase mRNA levels in the arcuate nucleus.Neuroendocrinology 58:501–510.
Arbogast LA, Voogt JL. 1994. Progesterone suppresses tyrosine hydroxy-lase messenger ribonucleic acid levels in the arcuate nucleus onproestrus. Endocrinology 135:343–350.
Arbogast LA, Voogt JL. 1997. Prolactin (PRL) receptors are colocalized indopaminergic neurons in fetal hypothalamic cell cultures: effect of PRLon tyrosine hydroxylase activity. Endocrinology 138:3016–3023.
Baker H, Farbman A. 1993. Olfactory afferent regulation of the dopaminephenotype in the fetal rat olfactory system. Neuroscience 52:115–134.
Baker RA, Le W-W, Berghorn KA, Hoffman GE. 1996. Selective co-localization of progesterone receptors on dopamine-containing neuronsof the hypothalamic arcuate nucleus in the rat. Endocr Soc Abstr1:P1–328.
Berghorn KA, Hoffman GE. 1993. Glucocorticoid receptor type II is presentin dopamine neurons but not in LHRH neurons. Endocr Soc Abstr17:1186.
Berghorn KA, Bonnet JH, Hoffman GE. 1994. cFos immunoreactivity isenhanced with biotin amplification. J Histochem Cytochem 42:1635–1642.
Blum M, McEwen B, Roberts J. 1987. Transcriptional analysis of tyrosinehydroxylase gene expression in the tuberoinfundibular dopaminergicneurons of the rat arcuate nucleus after estrogen treatment. J BiolChem 262:817–821.
Carter KC, Taneja KL, Lawrence JB. 1991. Discrete nuclear domains ofpoly(A) RNA and their relationship to the functional organization ofthe nucleus. J Cell Biol 115:1191–1202.
Carter KC, Bowman D, Carrington W, Fogarty K, McNeil JA, Fay FS,Lawrence JB. 1993. A three-dimensional view of precursor messengerRNA metabolism within the mammalian nucleus. Science 259:1330–1335.
Cho J, Min N, Franzen L, Baker H. 1996. Rapid down-regulation of ty-rosine hydroxylase expression in the olfactory bulb of naris-occludedadult rats. J Comp Neurol 369:264–276.
de Greef WJ, Visser TJ. 1981. Evidence for the involvement of the hypo-thalamic dopamine and thyrotropin-releasing hormone in suckling-induced release of prolactin. J. Endocrinol 91:213–223.
de Greef WJ, Plotsky PM, Neill JD. 1981. Dopamine levels in hypophysialstalk plasma and prolactin levels in peripheral plasma of the lactatingrat: effects of a simulated suckling stimulus. Neuroendocrinology 32:229–233.
Demarest KT, McKay DW, Riegle GD, Moore KE. 1983. Biochemical indi-ces of tuberoinfundibular dopaminergic neuronal activity during lacta-tion: a decreased responsiveness to prolactin. Neuroendocrinology 36:130–137.
deMaria JE, Livingstone JD, Freeman ME. 1998. Characterization of thedopaminergic input to the pituitary gland throughout the estrous cycleof the rat. Neuroendocrinology 67:377–383.
Dingwall C. 1991. If the cap fits. . . Curr Biol 1:65–66.
431CHANGES IN TH mRNA DURING PUP REMOVAL

Dreyfuss G, Matunis M, Pinol-Roma S, Burd C. 1993. hnRNP proteins andthe biogenesis of mRNA. Annu Rev Biochem 62:289–321.
Erlich M, Grillo M, Joh T, Margolis F, Baker H. 1990. Transneuronalregulation of neuronal specific gene expression in the mouse olfactorybulb. Mol Brain Res 7:115–122.
Feldherr C, Atkin D. 1990. EM visualization of nucleocytoplasmic trans-port processes. Elect Microsc 3:73–86.
Fossom L, Sterling C, Tank A. 1992. Regulation of tyrosine hydroxylasegene transcriptional rate and tyrosine hydroxylase mRNA stability bycyclic AMP and glucocorticoid. Mol Pharmacol 42:898–908.
Ghee M, Baker H, Miller JC, Ziff EB. 1998. AP-1, CREB and CBP tran-scription factors differentially regulate the tyrosine hydroxylase gene.Brain Res Mol Brain Res 55:101–114.
Guzowski JF, McNaughton BL, Barnes CA, Worley PF. 1999. Environment-specific expression of the immediate-early gene Arc in hippocampalneuronal ensembles. Nat Neurosci 2:1120–1124.
Harfstrand A, Fuxe K, Cintra A, Agnati LF, Zini I, Wikstrom AC, Okret S,Yu ZY, Goldstein M, Steinbusch H, et al. 1986. Glucocorticoid receptorimmunoreactivity in monoaminergic neurons of rat brain. Proc NatlAcad Sci USA 83:9779–9783.
Herman JP, Schafer MK, Watson SJ, Sherman TG. 1991. In situ hybrid-ization analysis of arginine vasopressin gene transcription usingintron-specific probes. Mol Endocrinol 5:1447–1456.
Herman JP, Schafer MK, Thompson RC, Watson SJ. 1992. Rapid regula-tion of corticotropin-releasing hormone gene transcription in vivo. MolEndocrinol 6:1061–1069.
Hermann B, Beyer C, Winkler A, Holsboer F, Rupprecht R. 1998. Regula-tion of the tyrosine hydroxylase by steroid hormones. Soc NeurosciAbstr 24:1598.
Hoffman GE, Smith MS, Fitzsimmons MD. 1992. Detecting steroidal ef-fects on immediate early gene expression in the hypothalamus. Neu-roprotocols 1:52–66.
Hoffman GE, Le WW, Abbud R, Lee WS, Smith MS. 1994. Use of Fos-related antigens (FRAs) as markers of basal activity: FRA changes indopamine neurons during proestrus, pregnancy and lactation. BrainRes 654:207–215.
Hoffman GE, Liu J, DeFranco D. 1997. Glucocorticoid receptors are trans-located by progesterone but show DNA binding deficits. Endocr SocAbstr P1–556.
Hoffman G, Le W, Anderson K. 1999. The suckling stimulus and notaltered hormone levels suppresses tyrosine hydroxylase expressionduring lactation in the rat. Endocr Soc Abstr 81:382.
Izaurralde E, Mattaj IW. 1992. Transport of RNA between nucleus andcytoplasm. Semin Cell Biol 3:279–288.
Jin BK, Franzen L, Baker H. 1996. Regulation of c-Fos mRNA and fosprotein expression in olfactory bulbs from unilaterally odor-deprivedadult mice. Int J Dev Neurosci 14:971–982.
Kehoe L, Janik J, Callahan P. 1992. Effects of immobilization stress ontuberoinfundibular dopaminergic (TIDA) neuronal activity and prolac-tin levels in lactating and non-lactating female rats. Life Sci 50:55–63.
Kohama SG, Bethea CL. 1995. Steroid regulation of tyrosine hydroxylasemessenger ribonucleic acid in dopaminergic subpopulations of monkeyhypothalamus. Endocrinology 136:1790–1800.
Lawrence JB, Singer RH, Marselle LM. 1989. Highly localized tracks ofspecific transcripts within interphase nuclei visualized by in situ hy-bridization. Cell 57:493–502.
Lee LR, Haisenleder DJ, Marshall JC, Smith MS. 1989. Effects of proges-terone on pulsatile luteinizing hormone (LH) secretion and LH subunitmessenger ribonucleic acid during lactation in the rat. Endocrinology124:2128–2134.
Lewis E, Harrington C, Chikaraishi D. 1987. Transcriptional regulation ofthe tyrosine hydroxylase gene by cyclic AMP. Proc Natl Acad Sci USA84:3550–3554.
Li C, Chen P, Smith M. 1997. Role of prolactin in the suckling-inducedincrease in NPY mRNA in the hypothalamus during lactation. EndocrSoc Abstr 79:P2–361.
Li C, Chen P, Smith M. 1999. Neuropeptide Y and tuberoinfundibulardopamine activities are altered during lactation: role of prolactin. En-docrinology 140:118–123.
Lightman SL, Harbuz MS. 1993. Expression of corticotropin-releasingfactor mRNA in response to stress. Ciba Found Symp 172:173–187.
Liu N, Tinti C, Joh T, Baker H. 1998. AP-1 and CRE elements mediateregulation of both basal and inducible tyrosine hydroxylase promoteractivities. Soc Neurosci Abstr 24:1598.
Lookingland KJ, Gunnet JW, Toney TW, Moore KE. 1990. Comparison ofthe effects of ether and restraint stress on the activity of tuberoinfun-dibular dopaminergic neurons in female and male rats. Neuroendocri-nology 52:99–105.
Ma XM, Levy A, Lightman SL. 1997a. Rapid changes in heteronuclearRNA for corticotrophin-releasing hormone and arginine vasopressin inresponse to acute stress. J Endocrinol 152:81–89.
Ma XM, Levy A, Lightman SL. 1997b. Rapid changes of heteronuclear RNAfor arginine vasopressin but not for corticotropin releasing hormone inresponse to acute corticosterone administration. J Neuroendocrinol9:723–728.
Meites J, Lu KH, Wuttke W, Welsch CW, Nagasawa H, Quadri SK. 1972.Recent studies of functions and control of prolactin secretion in rats.Recent Prog Horm Res 38:471–526.
Morrell JI, Rosenthal MF, McCabe JT, Harrington CA, Chikaraishi DM,Pfaff DW. 1989. Tyrosine hydroxylase mRNA in the neurons of thetuberoinfundibular region and zona incerta examined after gonadalsteroid hormone treatment. Mol Endocrinol 3:1426–1433.
Sar M. 1984. Estradiol is concentrated in tyrosine hydroxylase-containingneurons of the hypothalamus. Science 223:938–940.
Schubert D, LaCorbiere M, Klier FG, Steinbach JH. 1980. The modulationof neurotransmitter synthesis by steroid hormones and insulin. BrainRes 190:67–79.
Selmanoff M, Wise PM. 1981. Decreased dopamine turnover in the medianeminence in response to suckling in the lactating rat. Brain Res 212:101–115.
Smith MS. 1981. The effects of high levels of progesterone secretion duringlactation on the control of gonadotropin secretion in the rat. Endocri-nology 109:1509–1517.
Smith MS, Neill JD. 1977. Inhibition of gonadotropin secretion duringlactation in the rat: relative contribution of suckling and ovarian ste-roids. Biol Reprod 17:255–261.
Swanson L. 1998. Brain maps: structure of the rat brain. Amsterdam:Elsevier.
Trembleau A, Bloom FE. 1995. Enhanced sensitivity for light and electronmicroscopic in situ hybridization with multiple simultaneous non-radioactive oligodeoxynucleotide probes. J Histochem Cytochem 43:829–841.
Walker CD, Lightman SL, Steele MK, Dallman MF. 1992. Suckling is apersistent stimulus to the adrenocortical system of the rat. Endocri-nology 130:115–125.
Wang HJ, Hoffman GE, Smith MS. 1993. Suppressed tyrosine hydroxylase(TH) and increased NPY gene expression in the arcuate nucleus oflactating rats. Endocrinology 133:1657–1663.
Watson RE, Wiegand SJ, Clough RW, Hoffman GE. 1986. Use of cryopro-tectant to maintain long-term peptide immunoreactivity and tissuemorphology. Peptides 7:155–159.
Williams LR, Sandquist D, Black AC Jr, Williams TH. 1981. Glucocorti-coids increase tyrosine hydroxylase activity in cultured murine neuro-blastoma. J Neurochem 36:2057–2062.
432 K.A. BERGHORN ET AL.





























