Embed Size (px)
Citation preview

TheProstate 71:1559^1567 (2011)
SWAP70, Actin-Binding Protein,Function as anOncogeneTargetingTumor-SuppressivemiR-145
in ProstateCancer
Takeshi Chiyomaru,1 Shuichi Tatarano,1 Kazumori Kawakami,1 Hideki Enokida,1*Hirofumi Yoshino,1 Nijiro Nohata,2 Miki Fuse,2 Naohiko Seki,2 and
Masayuki Nakagawa1
1DepartmentofUrology,Graduate SchoolofMedicalandDental Sciences,KagoshimaUniversity,Kagoshima, Japan2Departmentof Functional Genomics,Graduate SchoolofMedicine,ChibaUniversity,Chiba, Japan
BACKGROUND. MiR-145 is down-regulated in various human cancers. We previouslydemonstrated that some actin-binding proteins were targeted by several microRNAs (miR-NAs), including miR-145, in bladder and prostate cancer (CaP). The aim of this study is todetermine a novel oncogenic gene targeted by miR-145 by focusing on actin-bindingproteins in CaP.METHODS. We focused on the SWAP switching B-cell complex 70 kDa subunit (SWAP70),which is an F-actin binding protein involved in activating B-cell transformation. A luciferasereporter assay was used to identify the actual binding sites between miR-145 and SWAP70mRNA. Cell viability was evaluated by cell proliferation, wound healing, and matrigel inva-sion assays in si-SWAP70 transfectants. A total of 75 clinical prostate specimens were sub-jected to immunohistochemistry of SWAP70.RESULTS. Molecular target searches of this miRNA and the luciferase reporter assayshowed that SWAP70 was directly regulated by miR-145. Silencing of SWAP70 studies dem-onstrated significant inhibitions of cell migration and invasion in CaP cell lines. TheSWAP70 positive-staining was significantly higher in percentage in the CaP than in benignprostate hyperplasia tissue.CONCLUSIONS. Down-regulation of miR-145 was a frequent event in CaP, and it mayhave a tumor suppressive function. SWAP70 may be a target of miR-145, and it mighthave a potential oncogenic function. The novel molecular networks though which miR-145 acts, may provide new insights into the underlying molecular mechanisms of CaP.Prostate 71: 1559–1567, 2011. # 2011 Wiley-Liss, Inc.
KEY WORDS: SWAP70; actin-binding protein; microRNA; miR-145; prostate cancer
INTRODUCTION
Prostate cancer (CaP) is one of the most commonmalignancies among men. The decrease in deathrates over the years reflects improvements in earlydetection and treatment [1]. However, global cancerstatistics indicated that in 2010, 11% of cancer deathsin men were attributable to CaP, the second highestrate of cancer mortality after lung cancer.
MicroRNAs (miRNAs) are a class of smallnon-coding RNA that regulates gene expressionthrough translational repression and mRNA cleav-age. MiRNAs play a critical role in a variety ofbiological processes, including metabolism, tissue
Additional Supporting Information may be found in the onlineversion of this article.
Grant sponsor: Ministry of Education, Science, Sports and Culture;Grant numbers: 20390427. 20591861.
Conflict of interest: The authors have no financial relationships todisclose.
*Correspondence to: Hideki Enokida, M.D., Ph.D., Associate Pro-fessor of Urology, Department of Urology, Graduate Schoolof Medical and Dental Sciences, Kagoshima University, 8-35-1Sakuragaoka, Kagoshima 890-8520, Japan.E-mail: [email protected] 20 January 2011; Accepted 7 February 2011DOI 10.1002/pros.21372Published online 25 February 2011 in Wiley Online Library(wileyonlinelibrary.com).
� 2011Wiley-Liss,Inc.

development, and differentiation, and contribute tothe development of various types of cancer [2]. MiR-145 is down-regulated and acts as a tumor suppres-sor in various human cancers including CaP [3],bladder cancer [4,5], esophageal squamous cell carci-noma [6], colon cancer [7,8], and breast cancer [9]. Inour previous study, we performed genome-widegene expression analysis to identify candidate genestargeted by miR-145 in CaP and found that 24 geneswere commonly down-regulated upon miR-145transfected PC3 and DU145 cell lines [10]. We havereported that among the candidates, FSCN1, anactin-binding protein, had an oncogenic functionand was directly regulated by miR-145 in CaP, aswell as in bladder cancer [5] and esophageal cancer[6]. On the other hand, miR-1, miR-133a, and miR-218 might function as tumor suppressors in bladdercancer through suppression of LASP, which is anactin-binding protein like FSCN1 [11]. Therefore, it isplausible that actin-binding protein is one of keycategories in which to search for novel moleculartargets related to miRNA. Thus, we focused on theSWAP switching B-cell complex 70 kDa subunit(SWAP70) gene, an actin-binding protein, as a prom-ising candidate from our gene expression profile inmiR-145 transfectants.
SWAP70 is a 70-kDa protein, originally isolatedfrom activated B lymphocytes [12], and it has beenmapped to a region of human chromosome 11q15.SWAP70 as well as FSCN1 have been shown to beassociated with a subset of actin filaments generatedin lamelipodia and the membrane ruffles of motilecells [13]. Over-expression of SWAP70 altered theactin organization and lamellipodial morphology[13]. The protein has been reported to localize withinmodulate cell adhesion to fibronectin, homotopic cellaggregation, and migration [14]. However, little isknown about the functional role of SWAP70 in CaP.We hypothesized that oncogenic SWAP70 is directlyregulated by tumor-suppressive miR-145 in CaP. Inthis study, we used a luciferase reporter assay todetermine whether SWAP70 actually has sitestargeted by miR-145. To investigate the functionalroles of SWAP70 in CaP, we performed a loss-of-function study using si-SWAP70-transfected CaPcell lines. Furthermore, we evaluated SWAP70protein expression in clinical CaP specimens byimmunohistochemistry.
MATERIALSANDMETHODS
CellCulture
We used two human CaP cell lines, PC3 andDU145, which were obtained from the AmericanType Culture Collection and were maintained in
RPMI-1640 medium supplemented with 10% fetalbovine serum in a humidified atmosphere of 5% CO2
and 95% air at 378C.
RNAExtraction
Total RNA including miRNA was extracted usingthe mirVanaTM miRNA isolation kit (Ambion, Aus-tin, TX) by following the manufacturer’s protocol.The integrity of the RNA was checked with RNA6000 Nano Assay Kit and a 2100 BioanalyzerTM (Agi-lent Technologies, Santa Clara, CA).
QuantitativeReal-TimeRT-PCR
TaqMan probes and primers for SWAP70 (P/N:Hs00929832_m1; Applied Biosystems, Foster City,CA) was assay-on-demand gene expression prod-ucts. All reactions were performed in triplicate, anda negative-control lacking cDNA was included. Wefollowed the manufacturer’s protocol for the PCRconditions. To normalize the data for quantificationof SWAP70 mRNA, we used human GUSB (P/N:Hs99999908_m1; Applied Biosystems) and the delta–delta Ct method to calculate the fold change.
MaturemiRNAandsiRNATransfection
As described elsewhere [4], the CaP cell lines weretransfected with LipofectamineTM RNAiMAX trans-fection reagent (Invitrogen, Carlsbad, CA) and Opti-MEMTM (Invitrogen) with 10 nM of mature miRNAmolecules. Pre-miRTM and negative-control miRNA(Applied Biosystems) were used in the gain-of-func-tion experiments, whereas SWAP70 siRNA (Cat;HSS118138 and HSS177150, Invitrogen) and negativecontrol siRNA (D-001810-10; Thermo Fisher Scien-tific, Waltham, MA) were used in the loss-of-functionexperiments. Cells were seeded in a 10-cm dish forprotein extraction (8 � 105 per dish), in a 6-well platefor the wound healing assay (20 � 104 per well), in a24-well plate for the mRNA extraction and luciferasereporter assay (5 � 104 per well), and in a 96-wellplate for the XTT assay (3,000 per well).
Cell Proliferation,Migration, and InvasionAssays
Cell proliferation was determined by using anXTT assay (Roche Applied Sciences, Tokyo, Japan)performed according to the manufacturer’s instruc-tions. Cell migration activity was evaluated bywound-healing assay. Cells were plated in six-welldishes, and the cell monolayer was scraped using aP-20 micropipette tip. The initial gap length (0 hr)and the residual gap length 24 hr after woundingwere calculated from photomicrographs. A cell inva-sion assay was carried out using modified Boyden
1560 Chiyomaruet al.
The Prostate

Chambers consisting of transwell-precoated matrigelmembrane filter inserts with 8-mm pores in 24-welltissue culture plates (BD Biosciences, Bedfold, MA).MEM containing 10% foetal bovine serum in thelower chamber served as the chemoattractant, as des-cribed previously [5]. All experiments were per-formed in triplicate.
WesternBlotAnalysis
After three days of transfection, protein lysate(30 mg) was separated by NuPAGE on 4–12% bis-trisgel (Invitrogen) and transferred to a polyvinylidenefluoride membrane. Immunoblotting was done withdiluted (1:200) polyclonal SWAP70 antibody(HPA006810; Sigma–Aldrich, St. Louis, MO) andGAPDH antibody (MAB374; Chemicon, Temecula,CA). The membrane was washed and then incubatedwith goat anti-rabbit IgG (H þ L)-HRP conjugate(BIO-RAD, Hercules, CA). Specific complexes werevisualized with an echochemiluminescence (ECL)detection system (GE Healthcare, Little Chalfont,UK), and the expression level of these genes wasevaluated by using ImageJ software (ver. 1.43;http://rsbweb.nih.gov/ij/index.html).
PredictionofmiRNATargets
The predicted target genes and their miRNA bind-ing site seed regions were investigated using Tar-getScan (release 5.1, http://www.targetscan.org/).The sequences of the predicted mature miRNAswere confirmed by referring to miRBase (release16.0, Sept 2010; http://microrna.sanger.ac.uk/).
PlasmidConstructionandDual-LuciferaseReporterAssay
MiR-145 target sequences were inserted betweenthe XhoI–PmeI restriction sites in the 3’UTR of thehRluc gene in the psiCHECKTM-2 vector (C8021;Promega, Madison, WI). Primer sequences for full-length 3’UTR of SWAP70 mRNA (GATCGCTCGAG-GAAGAACTGGAAAGAGAAAAAGACC and TA-GGTTTAAACTGTACAATGTTTTCATACGAATGTTTT)were designed. Specific miRNA target sequences(Supplemental Table I) for miR-145 were artificiallysynthesized and inserted in the vector. In addition,we constructed three mutated vectors in which thespecific sites targeted by the miRNAs were deleted.Following that, PC3 cells were transfected with 5 ngof vector, 10 nM of miRNA, and 1 ml of Lipofectami-neTM 2000 (Invitrogen) in 100 ml of Opti-MEM
TM
(Invi-trogen). The activities of firefly and Renillaluciferases in cell lysates were determined with adual-luciferase assay system (E1910; Promega).
Normalized data were calculated as the quotient ofRenilla/firefly luciferase activities.
Immunohistochemistry
A tissue microarray of 64 CaPs and 11 hyperpla-sias was obtained from US Biomax, Inc. (PR752;Rockville, MD). Detailed information on all tumorspecimens can be found at http://www.biomax.us/index.php. Immunostaining was done on the tissuemicroarray following the manufacturer’s protocol.The primary rabbit polyclonal antibodies againstSWAP70 (Sigma–Aldrich) were diluted by 1:25. Theslides were treated with biotinylated anti-rabbit IgG(H þ L)TM made in goat (Vector Laboratories, Burlin-game, CA). Diaminobenzidine–hydrogen peroxide(Sigma–Aldrich) was the chromogen, and the coun-terstaining was done with 0.5% hematoxylin. Immu-nostaining was evaluated according to the scoringmethod as described previously [15]. Each case wasscored on the basis of the intensity and area of stain-ing. The intensity of staining was graded on the fol-lowing scale: 0, no staining; 1þ, mild staining; 2þ,moderate staining; and 3þ, intense staining. The areaof staining was evaluated as follows: 0, no stainingof cells in any microscopic field; 1þ, <30% of cellsstained positive; 2þ, 30–60% stained positive; 3þ,>60% stained positive. The immunostaining scorewas also classified into two categories: that is, whena sum of intensity and area of staining score was twoor more, the sample was defined as positive, else itwas negative.
StatisticalAnalysis
The relationship between two variables and thenumerical values obtained by real-time RT-PCR wasanalyzed using the Mann–Whitney U test. Therelationship among three variables and the numericalvalues was analyzed using the Bonferroni-adjustedMann–Whitney U test. A x2-test was used to evaluatethe staining results of SWAP70 expression amongdifferent groups. Expert StatView1 analysis software(version 4; SAS Institute Inc., Cary, NC) was used inboth cases. In the comparison among three variables,a non-adjusted statistical level of significance ofP < 0.05 corresponds to a Bonferroni-adjusted levelof P < 0.0167.
RESULTS
SWAP70 asaTargetof Post-TranscriptionalRepressionbymiR-145
We focused on SWAP70 because it had been listedas a candidate actin-binding gene for miR-145 in thegenome-wide gene expression analysis that was done
miR-145Regulates SWAP70 inProstateCancer 1561
The Prostate

Fig. 1. MiR-145 might have directly targeted SWAP70. (A) Upper, SWAP70 mRNA expression after 24 hr transfection with 10 nM ofmiR-145. (A) Lower, SWAP70 protein expression after 72 hr transfection ofmiR-145.GAPDHwas used as a loading control.ThemRNA andprotein expression levels of SWAP70 were also suppressed in the transfectants. (B,C) MiR-145 binding sites in 3’UTR of SWAP70 mRNA.(B) A luciferase reporter assay using the vector encoding full-length 3’UTR of SWAP70 mRNA.The Renilla luciferase values were normal-izedby firefly luciferasevalues. (C) Luciferasereporter assays using thevectors encodingputative target sites of SWAP703’UTR: two targetsites formiR-145.
The Prostate
by using microarrays (the Gene Expression Omnibus(GEO) accession number, GSE24782). The mRNAand protein expression levels of SWAP70 were mark-edly down-regulated in the transfectants in compari-son with the controls (Fig. 1A). We performed aluciferase reporter assay to determine whetherSWAP70 mRNA has a target site for miR-145. Weused a vector encoding full-length 3’UTR of SWAP70mRNA and found that the luminescence intensitywas significantly reduced in the miR-145 transfectant(Fig. 1B). Furthermore, the luminescence intensitysignificantly decreased at the two sites targeted bymiR-145 (position 1,726–1,732 and 1,763–1,769)(Fig. 1C). In addition, the luminescence intensity wasnot decreased in double deleted vector by miR-145(Fig. 1C). These data suggest that miR-145 directlybinds to specific sites on 3’UTR of SWAP70 mRNA.
Effectof SWAP70KnockdownonCell Proliferation,Migration, and InvasionActivityinCaPCell Lines
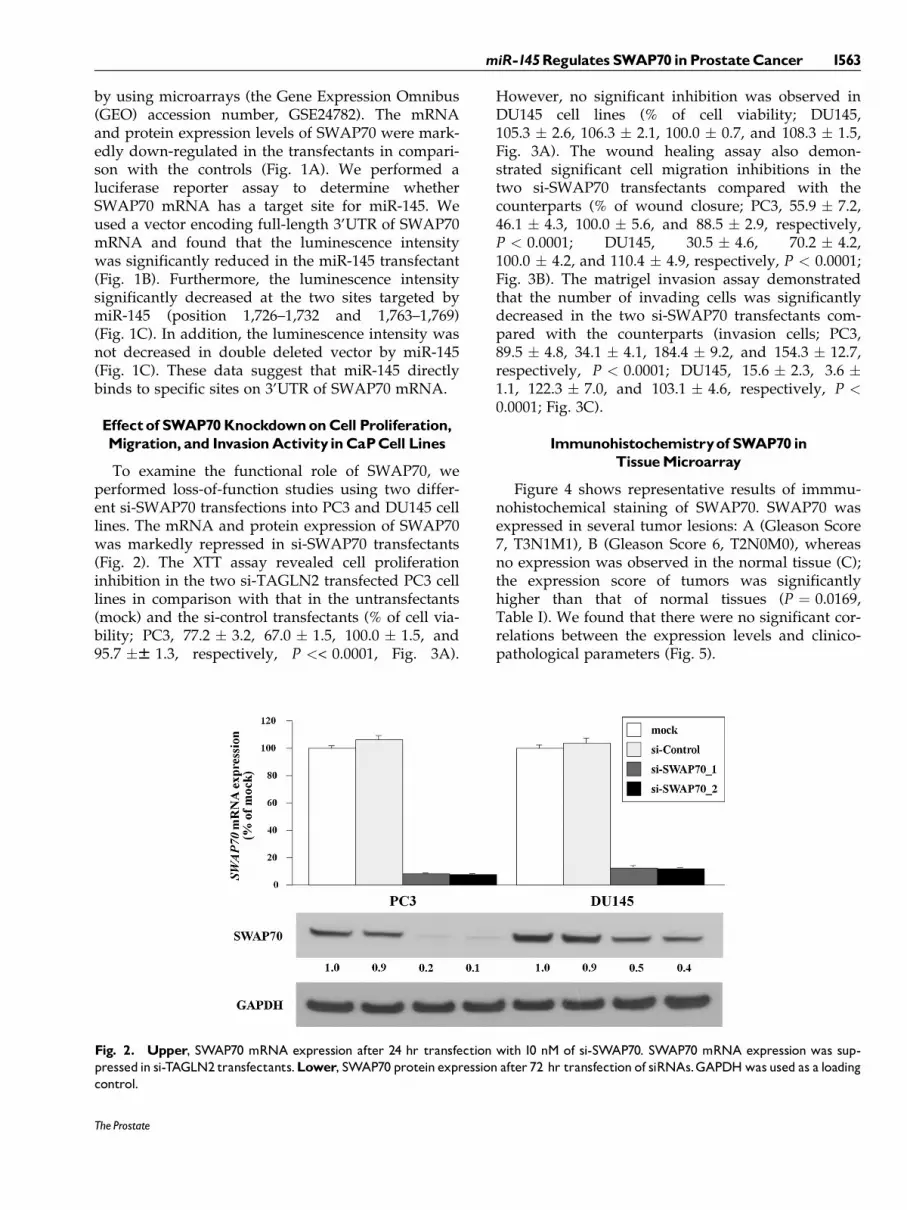
To examine the functional role of SWAP70, weperformed loss-of-function studies using two differ-ent si-SWAP70 transfections into PC3 and DU145 celllines. The mRNA and protein expression of SWAP70was markedly repressed in si-SWAP70 transfectants(Fig. 2). The XTT assay revealed cell proliferationinhibition in the two si-TAGLN2 transfected PC3 celllines in comparison with that in the untransfectants(mock) and the si-control transfectants (% of cell via-bility; PC3, 77.2 � 3.2, 67.0 � 1.5, 100.0 � 1.5, and95.7 �W 1.3, respectively, P << 0.0001, Fig. 3A).
However, no significant inhibition was observed inDU145 cell lines (% of cell viability; DU145,105.3 � 2.6, 106.3 � 2.1, 100.0 � 0.7, and 108.3 � 1.5,Fig. 3A). The wound healing assay also demon-strated significant cell migration inhibitions in thetwo si-SWAP70 transfectants compared with thecounterparts (% of wound closure; PC3, 55.9 � 7.2,46.1 � 4.3, 100.0 � 5.6, and 88.5 � 2.9, respectively,P < 0.0001; DU145, 30.5 � 4.6, 70.2 � 4.2,100.0 � 4.2, and 110.4 � 4.9, respectively, P < 0.0001;Fig. 3B). The matrigel invasion assay demonstratedthat the number of invading cells was significantlydecreased in the two si-SWAP70 transfectants com-pared with the counterparts (invasion cells; PC3,89.5 � 4.8, 34.1 � 4.1, 184.4 � 9.2, and 154.3 � 12.7,respectively, P < 0.0001; DU145, 15.6 � 2.3, 3.6 �1.1, 122.3 � 7.0, and 103.1 � 4.6, respectively, P <0.0001; Fig. 3C).
Immunohistochemistryof SWAP70 inTissueMicroarray
Figure 4 shows representative results of immmu-nohistochemical staining of SWAP70. SWAP70 wasexpressed in several tumor lesions: A (Gleason Score7, T3N1M1), B (Gleason Score 6, T2N0M0), whereasno expression was observed in the normal tissue (C);the expression score of tumors was significantlyhigher than that of normal tissues (P ¼ 0.0169,Table I). We found that there were no significant cor-relations between the expression levels and clinico-pathological parameters (Fig. 5).
Fig. 2. Upper, SWAP70 mRNA expression after 24 hr transfection with 10 nM of si-SWAP70. SWAP70 mRNA expression was sup-pressed in si-TAGLN2 transfectants.Lower, SWAP70 protein expression after 72 hr transfection of siRNAs.GAPDHwas used as a loadingcontrol.
miR-145Regulates SWAP70 inProstateCancer 1563
The Prostate

Fig. 3. (A^C) SWAP70-knockdown effects on CaP cell viability by si-RNA. (A) Cell proliferation determined by the XTTassay; (B) cellmigration activity determined by the wound healing assay; and (C) cell invasion activity determined by the matrigel invasion assay in PC3andDU145 cell lines transfectedwith si-SWAP70. �P < 0.0001.
1564 Chiyomaruet al.
The Prostate

DISCUSSION
Increasing evidence indicates that miR-145 isunderexpressed in various tumor tissues and func-tions as a tumor suppressor [3–9]. It has shown thatmiR-145 is capable of inhibiting tumor cell growthand invasion and mediating metastasis [16]. Interest-ingly, p53 directly binds to the promoter of miR-145[8]. Previous studies demonstrated that c-Myc [8],mucin 1 [17], RTKN [18], FLI1 [19], and IRS-1 [20],which have an oncogenic function, were the targetgenes of miR-145. Recent studies suggested that sev-eral miRNAs regulated genes classified as actin-bind-ing proteins with a conserved actin-binding domain
in their construct and that these genes were associ-ated with tumor growth acceleration or inhibition[5,11,21]. For example, the activities of actin-bindingproteins are controlled through various signalingpathways including the Rho-family small GTPases(RhoA, Rac, and Cdc42) that were targeted by sev-eral miRNAs [22,23]. The expression of WAVE3, anactin cytoskeleton remodeling and metastasis pro-moter protein, is regulated by the miR200 family,which has been found to regulate the epithelial tomesenchymal transition (EMT) during tumor pro-gression and metastasis [24]. MiR-143 and miR-145were involved in regulation of MYO6 expression andpossibly in the development of CaP [25]. On the ot-her hand, TPM1, a tumor suppressor actin-bindingprotein can be regulated by miR-21 that is over-expressed in several tumor tissues [26]. In this study,we focused on SWAP70 because it is an actin-bind-ing protein. Furthermore, SWAP70 is an actin-bind-ing protein like FSCN1 and seemed to play animportant role in CaP development. The gene profiletargeted by miR-145 in the CaP cell lines is verysimilar to our previous profiles from BC and ESCC[5,6]. Therefore, we looked back and found thatSWAP70 was markedly down-regulated in miR-145-transfected BC and ESCC cell lines (each GEO acces-sion number, GES19717 and GES20028) [5,6]. Whenwe combined the three microarray analyses, thereremained four genes (FSCN1, DTD1, SWAP70,NDUFA4) that were commonly down-regulated(Supplemental Table II). Among them, FSCN1 waslisted at the top of all arrays analyses, and it has beenreported to be a target oncogene of miR-145 [5,6,10].Our results strongly suggest that these actin-bindingproteins including SWAP70 play important roles intumor development in CaP as well as other humanmalignancies and that miR-145 functions as a criticalregulator of these oncogenic genes.
We demonstrated that the SWAP70 was highlyexpressed in human CaP and cell viabilities werereduced in si-SWAP70-transfected PC3 and DU145cell lines. These results suggest that SWAP70 has anoncogenic function through cell migration and in-vasion activity in CaP. However, there was no sig-nificant difference in cell proliferation activity in thesi-SWAP70-transfected DU145 cell line, suggesting
Fig. 4. Immunohistochemical staining of SWAP70 in tissuespecimens (left panel, original magnification �100; right panel,original magnification �400). (A) Positively stained tumor lesion(Gleason Score 7, T3N1M1); (B) positively stained tumor lesion(Gleason Score 6, T2N0M0); and (C) negative staining in hyper-plasia tissue. (A, B) Positive staining in tumor cells: weak (A),moderate (B).
TABLE I. Expression of SWAP70 inHumanProstateTissues
Histological evaluation
Expression of SWAP70
P-valuePositive, No. (%) Negative, No. (%)
Hyperplasia, n ¼ 11 0 (0.0) 11 (100.0) 0.0169Adenocarcinoma, n ¼ 64 23 (35.9) 41 (64.1)
miR-145RegulatesSWAP70 inProstateCancer 1565
The Prostate

that cell proliferation does not depend on SWAP70expression in some phenotypes. In our cohort, therewas no significant relationship between SWAP70expression and clinico-pathological parameters. Theexpression levels of SWAP70 were relatively higherin low Gleason’s grade and non-metastasis CaPs,suggesting that increased expression of SWAP70might be an early event in prostate carcinogenesis.Our sample number was too small to test for arelationship. The use of a stable SWAP70 knockdownsystem in vivo and the development of pharmaco-logic inhibitors of SWAP70 will be instrumental infurthering our understanding of the role of SWAP70dynamics in human CaP.
CONCLUSIONS
In summary, miR-145 may function as a tumorsuppressor through repression of oncogenic SWAP70in CaP. SWAP70 knockdown resulted in decreasedCaP cell migration and invasion activity. Novel mol-ecular networks of miRNAs may provide newinsights into the underlying molecular mechanismsof CaP.
ACKNOWLEDGMENTS
We thank Ms. Mutsumi Miyazaki for her excellentlaboratory assistance. This research was partiallysupported by the Ministry of Education, Science,
Sports and Culture Grants-in-Aid for ScientificResearch (B and C), 20390427 and 20591861, 2008.
REFERENCES
1. Jemal A, Siegel R, Xu J, Ward E: Cancer statistics, 2010.CA Cancer J Clin 2010;60(5):277–300.
2. Ryan BM, Robles AI, Harris CC: Genetic variation in micro-RNA networks: The implications for cancer research. Nat RevCancer 2010;10(6):389–402.
3. Zaman MS, Chen Y, Deng G, Shahryari V, Suh SO, Saini S,Majid S, Liu J, Khatri G, Tanaka Y, Dahiya R: The functionalsignificance of microRNA-145 in prostate cancer. Br J Cancer2010;103(2):256–264.
4. Ichimi T, Enokida H, Okuno Y, Kunimoto R, Chiyomaru T,Kawamoto K, Kawahara K, Toki K, Kawakami K, NishiyamaK, Tsujimoto G, Nakagawa M, Seki N: Identification of novelmicroRNA targets based on microRNA signatures in bladdercancer. Int J Cancer 2009;125(2):345–352.
5. Chiyomaru T, Enokida H, Tatarano S, Kawahara K, Uchida Y,Nishiyama K, Fujimura L, Kikkawa N, Seki N, Nakagawa M:miR-145 and miR-133a function as tumour suppressors anddirectly regulate FSCN1 expression in bladder cancer. Br JCancer 2010;102(5):883–891.
6. Kano M, Seki N, Kikkawa N, Fujimura L, Hoshino I, AkutsuY, Chiyomaru T, Enokida H, Nakagawa M, Matsubara H:miR-145, miR-133a and miR-133b: Tumor suppressive miR-NAs target FSCN1 in esophageal squamous cell carcinoma.Int J Cancer 2010;127(12):2804–2814.
7. Schepeler T, Reinert JT, Ostenfeld MS, Christensen LL, Silah-taroglu AN, Dyrskjot L, Wiuf C, Sorensen FJ, Kruhoffer M,
Fig. 5. Correlationbetween SWAP70 expression and clinicopathologic parameters inCaP specimens.
1566 Chiyomaruet al.
The Prostate

Laurberg S, Kauppinen S, Orntoft TF, Andersen CL: Diagnos-tic and prognostic microRNAs in stage II colon cancer. CancerRes 2008;68(15):6416–6424.
8. Sachdeva M, Zhu S, Wu F, Wu H, Walia V, Kumar S, Elble R,Watabe K, Mo YY: p53 represses c-Myc through induction ofthe tumor suppressor miR-145. Proc Natl Acad Sci USA 2009;106(9):3207–3212.
9. Spizzo R, Nicoloso MS, Lupini L, Lu Y, Fogarty J, Rossi S,Zagatti B, Fabbri M, Veronese A, Liu X, Davuluri R, CroceCM, Mills G, Negrini M, Calin GA: miR-145 participates withTP53 in a death-promoting regulatory loop and targets estro-gen receptor-alpha in human breast cancer cells. Cell DeathDiffer 2010;17(2):246–254.
10. Fuse M, Nohata N, Kojima S, Sakamoto S, Chiyomaru T,Kawakami K, Enokida H, Nakagawa M, Naya Y, Ichikawa T,Seki N: Restoration of miR-145 expression suppresses cell pro-liferation, migration, and invasion in prostate cancer by tar-geting FSCN1. Int J Oncol 2011;38(4):1093–1101.
11. Chiyomaru T, Enokida H, Kawakami K, Tatarano S, UchidaY, Kawahara K, Nishiyama K, Seki N, Nakagawa M: Func-tional role of LASP1 in cell viability and its regulation bymicroRNAs in bladder cancer. Urol Oncol 2010 [In press].
12. Borggrefe T, Wabl M, Akhmedov AT, Jessberger R: A B cell-specific DNA recombination complex. J Biol Chem 1998;273(27):17025–17035.
13. Hilpela P, Oberbanscheidt P, Hahne P, Hund M, KalhammerG, Small JV, Bahler M: SWAP-70 identifies a transitional sub-set of actin filaments in motile cells. Mol Biol Cell 2003;14(8):3242–3253.
14. Sivalenka RR, Jessberger R: SWAP-70 regulates c-kit-inducedmast cell activation, cell–cell adhesion, and migration. MolCell Biol 2004;24(23):10277–10288.
15. Zhang Y, Ye Y, Shen D, Jiang K, Zhang H, Sun W, Zhang J,Xu F, Cui Z, Wang S: Identification of transgelin-2 as a bio-marker of colorectal cancer by laser capture microdissectionand quantitative proteome analysis. Cancer Sci 2010;101(2):523–529.
16. Sachdeva M, Mo YY: miR-145-mediated suppression of cellgrowth, invasion and metastasis. Am J Transl Res 2010;2(2):170–180.
17. Sachdeva M, Mo YY: MicroRNA-145 suppresses cell invasionand metastasis by directly targeting mucin 1. Cancer Res2010;70(1):378–387.
18. Wang S, Bian C, Yang Z, Bo Y, Li J, Zeng L, Zhou H, ZhaoRC: miR-145 inhibits breast cancer cell growth through RTKN.Int J Oncol 2009;34(5):1461–1466.
19. Zhang J, Guo H, Zhang H, Wang H, Qian G, Fan X, HoffmanAR, Hu JF, Ge S: Putative tumor suppressor miR-145 inhibitscolon cancer cell growth by targeting oncogene friend leuke-mia virus integration 1 gene. Cancer 2011;117(1):86–95.
20. Shi B, Sepp-Lorenzino L, Prisco M, Linsley P, deAngelis T,Baserga R: Micro RNA 145 targets the insulin receptor sub-strate-1 and inhibits the growth of colon cancer cells. J BiolChem 2007;282(45):32582–32590.
21. Saarikangas J, Zhao H, Lappalainen P: Regulation of the actincytoskeleton-plasma membrane interplay by phosphoinositi-des. Physiol Rev 2010;90(1):259–289.
22. Heasman SJ, Ridley AJ: Mammalian Rho GTPases: Newinsights into their functions from in vivo studies. Nat Rev MolCell Biol 2008;9(9):690–701.
23. Bueno MJ, Perez de Castro I, Malumbres ML: Control of cellproliferation pathways by microRNAs. Cell Cycle 2008;7(20):3143–3148.
24. Sossey-Alaoui K, Bialkowska K, Plow EF: The miR200 familyof microRNAs regulates WAVE3-dependent cancer cell inva-sion. J Biol Chem 2009;284(48):33019–33029.
25. Szczyrba J, Loprich E, Wach S, Jung V, Unteregger G, Barth S,Grobholz R, Wieland W, Stohr R, Hartmann A, Wullich B,Grasser F: The microRNA profile of prostate carcinomaobtained by deep sequencing. Mol Cancer Res 2010;8(4):529–538.
26. Zhu S, Si ML, Wu H, Mo YY: MicroRNA-21 targets the tumorsuppressor gene tropomyosin 1 (TPM1). J Biol Chem 2007;282(19):14328–14336.
miR-145RegulatesSWAP70 inProstateCancer 1567
The Prostate














![Monograph Chapter: "Safavid army", pp. 145-158[In Azerbaijani Turkish]](https://img.pdfslide.net/doc/110x75/631eb8e956cbbb475005b2ac/monograph-chapter-safavid-army-pp-145-158in-azerbaijani-turkish.jpg)














