Embed Size (px)
Citation preview

T A X O N O M I C A N D B I O S T R A T I G R A P H I C A L
R E - A S S E S S M E N T S O F S U B T E R R A N I P H Y L L U M E L L I O T T
( C O R A L L I N A L E S , R H O D O P H Y T A )
by D A V I D E B A S S I , W I L L I A M J . W O E L K E R L I N G and J A M E S H . N E B E L S I C K
ABSTRACT. A taxonomic and biostratigraphical re-assessment of Subterraniphyllum Elliott (Corallinales, Rhodophyta)is presented. Results from studies of the type collection and of newly collected material from north-eastern Italy andnorthern Slovenia have shown that this taxon is not a geniculate coralline red alga as originally suggested by Elliott andmost subsequent authors. Vegetatively, Subterraniphyllum most closely resembles certain living members of theCorallinales; however, the phenetic and phylogenetic relationships of Subterraniphyllum to other Corallinales cannotbe determined with greater certainty. The exclusion of Subterraniphyllum from any group of Corallinaceae withgenicula is based on unequivocal evidence that branch formation does not involve the occurrence of genicula.Subterraniphyllum seems to be restricted to the Oligocene. Reports of occurrences in Upper Eocene and LowerMiocene sediments cannot be substantiated. Subterraniphyllum, however, cannot be considered a useful stratigraphicalmarker until further data on its occurrence in well-dated carbonate sequences are acquired. This study illustrates theproblems associated with placing fossil coralline algal specimens into geniculate genera without the preservation ofrelevant morphological characters. This is especially true in the absence of the careful assessment of fossil materialwith respect to current taxonomic concepts of geniculate coralline genera, all of which are based on studies of livingspecies. According to the current concepts for geniculate coralline genera, the placing of fossil specimens intogeniculate genera without appropriate evidence must be avoided by grouping all potentially geniculate fragmentsunder the informal group `Geniculate sensu lato'. Furthermore, for all those many fossil specimens where unequivocalevidence is not present, it is possible to utilize `form genera' based on characters that are normally preserved. Thisleads to creating a consistent, workable system of applying names to most fossil corallines so that they can be reliablyused in relation to stratigraphical and palaeoecological studies.
E L L I O T T (1957) established Subterraniphyllum as a genus of geniculate coralline red algae (Corallinales,Rhodophyta) for specimens occurring mainly in Oligocene carbonate deposits in Iraq, Persia (Iran), Oman, andSicily. Elliott (1957) provided a brief account of the only included and newly described species, S. thomasii (asthomasi), chose a specimen from Iran as holotype, provided nine photographic illustrations of his material, andassigned the genus to the subfamily Corallinoideae within the family Corallinaceae. Elliott (1957, pp. 73±74)also suggested that the species was useful as a biostratigraphical dating tool because it appeared to be restrictedto Upper Eocene±Lower Miocene deposits.
Elliott (1957, p. 74) chose the speci®c epithet to honour A. N. Thomas, who (Thomas 1950, 1952) ®rstnoted the occurrence of the species in Asmari Limestone deposits in south-west Iran and who consideredthe alga to be stratigraphically useful.
Since 1957, Subterraniphyllum thomasii has been treated as a geniculate coralline red alga by mostauthors (but see Discussion), and it has been recorded from Oligocene deposits in Greece (Johnson 1965,p. 812), Borneo, Indonesia (Johnson 1966, p. 277), Italy (Mastrorilli 1968a, p. 1277; 1968b, p. 388), andMacedonia (Lemoine 1977, p. 34). S. thomasii has also been reported from the Upper Eocene andOligocene of Crete (Bonneau 1970, p. 1124), from the Upper Eocene of Karpathos, Crete and Rhodes(Aubouin et al. 1976, p. 389), and from Madagascar (Lemoine 1977, p. 36, citing an unpublished thesis ofKarche, 1972). An unidenti®ed species of Subterraniphyllum has also been reported from the Oligoceneand Lower Miocene of Sicily (Blondeau et al. 1972), and from the Oligocene of Cuba (Beckmann andBeckmann 1966, p. 30), but no further species have been described or formally transferred into the genus.
[Palaeontology, Vol. 43, Part 3, 2000, pp. 405±425, 3 pls] q The Palaeontological Association

Subterraniphyllum is also the only genus which has been referred to the Corallinales (subfamilyCorallinoideae) that appears to be con®ned to fossil occurrences (reported from the Upper Eocene,Oligocene and Lower Miocene deposits; there are no known living species. By contrast, the other sixgenera of geniculate Corallinales reported from the fossil record (Table 1) occur in various deposits fromthe Cretaceous onwards, and all six are also represented by species in the Recent.
There are three major problems associated with the taxonomy and potential biostratigraphical value ofSubterraniphyllum. Firstly, Elliott (1957) concluded that Subterraniphyllum was a geniculate corallinebased on the occurrence of `incipient bifurcation'. Fully developed genicula were absent from his material,
406 P A L A E O N T O L O G Y , V O L U M E 4 3
TABLE 1. Stratigraphical distribution of the geniculate coralline genera known as fossils. Johnson (1964, p. G31) andPoignant (1977, p. 180) recognized that only Calliarthron Manza, 1937, Jania Lamouroux, 1812, AmphiroaLamouroux, 1812, Arthrocardia Decaisne, 1842 emend. Areschoug, 1852, and Corallina Linnaeus, 1758 seem tobe represented as fossils. Later, Poignant (1979a, b, 1985) included in this list Metagoniolithon Weber-van Bosse,1904 and Subterraniphyllum Elliott, 1957. *Archaeoamphiroa nomen nudum; Late Devonian; Magathan (1985,p. 176).
StratigraphicalGenus distribution References
Amphiroa Lamouroux, 1812* Upper Cretaceous- Beckmann and Beckmann (1966);Recent Lemoine (1939); Conti (1950);
Johnson (1961), Poignant (1977,1979a)
Arthrocardia Decaisne emend. Upper Cretaceous- Raineri (1920); Conti (1950); JohnsonAreschoug, 1852 Recent (1961), Poignant (1977, 1979a)
Calliarthron Manza, 1937 Lower Miocene-Recent Johnson (1961); Beckmann andBeckmann (1966)
Corallina Linnaeus, 1758 Upper Cretaceous- Pfender (1924); Conti (1950); JohnsonRecent (1961, 1964); Poignant (1977, 1979a)
Jania Lamouroux, 1812 Upper Cretaceous- Lemoine (1927); Conti (1950);Recent Johnson (1961); Poignant (1977,
1979a)
Metagoniolithon Weber-van Bosse, Upper Paleocene-Recent Poignant (1977, 1979a, 1985)1904
Subterraniphyllum Elliott, 1957 Oligocene Elliott (1957); Johnson (1965, 1966);Beckmann and Beckmann (1966);Mastrorilli (1968a, b); Poignant(1977, 1979a, 1985)
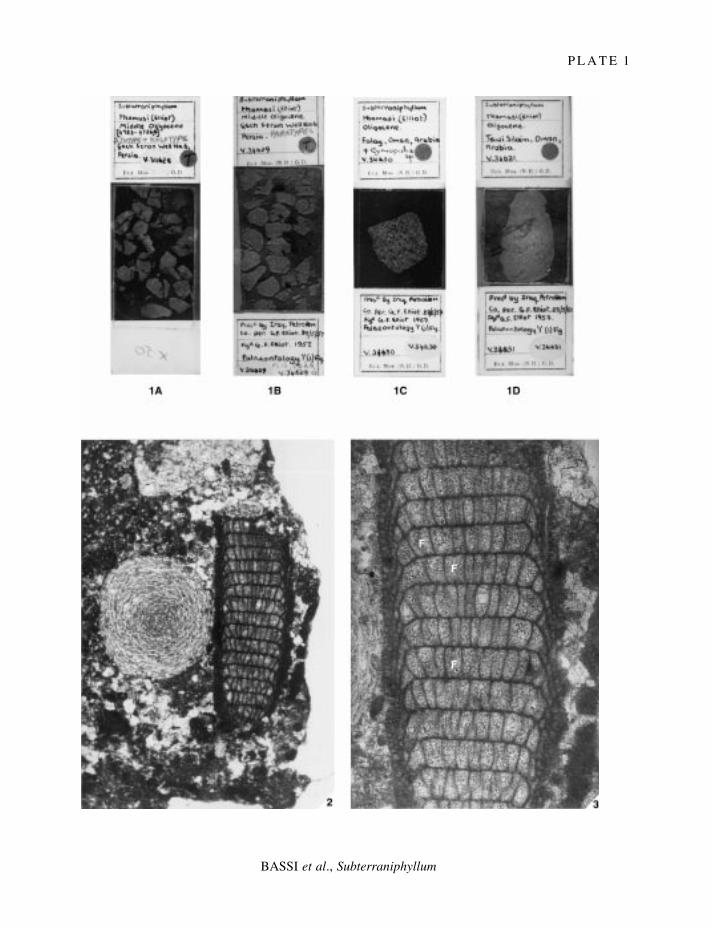
E X P L A N A T I O N O F P L A T E 1
Fig. 1. Subterraniphyllum thomasii Elliott, 1957; Elliott's Collection; Department of Palaeontology at The NaturalHistory Museum, London (UK); ´ 1. 1A, V34628; holotype and isotypes, Rupelian of Gach Saran, Persia (Iran). 1B,V34629; isotypes, Oligocene of Gach Saran, Persia (Iran). 1C, V34630; paratypes, Oligocene of Falaij, Oman. 1D,V34631; paratypes, Oligocene of Tawi Silaim, Oman.
Figs 2±3. Subterraniphyllum thomasii Elliott, 1957; holotype, sample V34628, Elliott's Collection; Rupelian of GachSaran, Persia (Iran). 2, medial longitudinal section of a branch together with Sphaerogypsina globula; ´ 40. 3,enlarged view of holotype showing the pseudoparenchymatous thallus with monomerous construction; noteprobable cell fusions (F) and the abrupt transition to much smaller cells near the periphery; ´ 90.
P L A T E 1
BASSI et al., Subterraniphyllum

and consequently, assignment of the genus to the subfamily Corallinoideae was presumptive. Moreover,Elliott did not see reproductive structures, especially conceptacles (which occur in all Corallinaceae), andthus placement of Subterraniphyllum in the family Corallinaceae was also presumptive. Secondly, therelationships of Subterraniphyllum to other Corallinales have not been examined in a modern context.Thirdly, the potential biostratigraphical value of Subterraniphyllum (see Elliott 1957, p. 73) has yet to befully assessed.
These problems became apparent after specimens referable to Subterraniphyllum thomasii wererecognised in Oligocene carbonate formations of northern Slovenia and northeastern Italy (Bassi andNebelsick 1997; Nebelsick and Bassi 1999). The present study was initiated to address the above problemsthrough a re-examination of the original collections upon which Subterraniphyllum was based and throughdetailed study of the newly collected material.
M A T E R I A L A N D M E T H O D S
Examinations were carried out on Elliott's original material, currently housed in the Department ofPalaeontology, The Natural History Museum (London), on thin sections of Ungaro (1978) coming fromthe Oligocene Calcareniti di Castelgomberto, outcropping at Lumignano (eastern Colli Berici, Veneto,north-eastern Italy); and on newly collected specimens from the Lower Oligocene Gornji Grad Beds,outcropping near Sv. Anton (Gornji Grad area, northern Slovenia; Bassi and Nebelsick 1997) (Text-®g. 1).In all cases, data were obtained from thin sections using light microscopy. Anatomical and morphologicalterms are those used by Woelkerling (1988), and Woelkerling et al. (1993). Ungaro's collection (1978) isdeposited at the Museo di Paleontologia of the Dipartimento di Scienze Geologiche e Paleontologiche,University of Ferrara (Italy); thin sections of Gornji Grad Limestones are deposited in the Institute andMuseum of Geology and Palaeontology of the University of TuÈbingen (Germany).
R E S U L T S
The Elliott collections: typi®cation
Elliott (1957) based the genus Subterraniphyllum and the species S. thomasii on specimens in thin sectionson four microscope slides (Pl. 1, ®g. 1) now housed in the Department of Palaeontology at The NaturalHistory Museum, London. Two of the slides (V.34628, V.34629) are based on material from Gach Saran,Persia (Iran); the third slide (V.34630) is based on material from Falaij, Oman; and the fourth slide(V. 34631) is based on material from Tawi Silaim, Oman. The whereabouts of the rock samples from
408 P A L A E O N T O L O G Y , V O L U M E 4 3
TEXT-FIG. 1. Geographical maps showing position of Lumignano in the Colli Berici (north-eastern Italy) and Sv. Antonin the Gornji Grad area (northern Slovenia).

which the thin sections were prepared is unknown. Elliott (1957) also referred material identi®ed by VanBellen (1956, pl. 2, ®gs B±C) as Dendrophyllum cf. gurgurdanensis from Iraq to Subterraniphyllumthomasii and mentioned further material from Sicily. The Van Bellen material has not been found, andthere are no specimens in the Elliott collections to substantiate the record from Sicily, although Elliott(1957, p. 74) acknowledged obtaining information from F. E. Eames on the occurrences of the alga.
Elliott (1957, p. 74, pl. 13, ®g. 5) designated one of the specimens on slide V.34628 as a holotype andconsidered certain other specimens (namely those on his pl. 13, ®gs 2±4, 9) from slides V.34628 andV.34629 to be paratypes. While Elliott`s selection of a holotype is in accord with the International Code ofBotanical Nomenclature (ICBN) (Greuter 1994), his designation of paratypes is not. All material on slidesV.34628 and V.34629 constitutes part of the same gathering (even though two different slide numberswere used), and thus in accord with ICBN Art. 9.3, all specimens on these two slides (other than thedesignated holotype) are isotypes (i.e. duplicates of the holotype) rather than paratypes. On the other hand,the specimens on slides V.34630 and V. 34631 from Oman constitute true paratypes in the context ofICBN Art. 9.5, where paratype is de®ned as a specimen cited in the protologue (i.e. the original account)that is neither the holotype nor an isotype. The Van Bellen (1956, pl. 2, ®gs B±C) specimens from Iraq citedby Elliott (1957, pp. 73±74) also qualify as paratypes, but the status of any material from Sicily isuncertain because Elliott (1957) neither mentioned particular specimens nor published illustrations, andvouchers have not been found.
The protologue (Elliott 1957, p. 74) cites both Well no. 10 (under the holotype) and Well no. 6 (in thelegend for ®gs 1±9) as the source of V.34628. The slide (i.e. V.34628) containing the holotype (i.e. Elliott1957, pl. 13, ®g. 5) is labelled as coming from Gach Saran Well no. 6; consequently it is assumed that thecitation of Well no. 10 in the protologue is an error. Neither Elliott (1957) nor Thomas (1950, 1952)provided information on Well numbers for the Gach Saran area.
Matters relating to typi®cation may be summarised as follows:
Genus SUBTERRANIPHYLLUM Elliott, 1957
Type species. Subterraniphyllum thomasii Elliott 1957, p. 73. According to the ICBN [Recommendation 60C.1(b)], asecond `i' is added in the speci®c epithet as a `stem augmentation'.
Holotype. Elliott 1957, pl. 13, ®g. 5 (on slide V.34628), from Well no. 6, Gach Saran, Persia (Iran). Department ofPalaeontology, The Natural History Museum (NHM), London.
Isotypes. Specimens on slides NHM V.34628 and NHM V.34629 other than the holotype.
Paratypes. Specimens on slides NHM V. 34630 (from Falaij, Oman) and NHM V.34631 (from Tawi Silaim, Oman);specimens illustrated by Van Bellen (1956, pl. 2, ®gs B±C) (repository of specimens uncertain).
Age of holotype and isotypes. Rupelian (Early Oligocene), in Asmari Limestone (south-west Iran) (Thomas 1952;Elliott 1957).
The Department of Palaeontology, The Natural History Museum, contains two additional slides (NHM V.56325,NHM V.56326) with material identi®ed by Elliott as Subterraniphyllum thomasii. These slides, like NHM V.34631,are based on samples from Tawi Silaim, Oman. They are not mentioned in the protologue (and thus do not qualify astypes) and were presented to the Museum by Elliott in 1971.
The Elliott collections: morphology and anatomy
The thin sections (V.34628±V.34631) comprising the original material contain fragments of Subterra-niphyllum thomasii in more or less median longitudinal view, tangential longitudinal view, transverseview, and oblique view. The holotype (Pl. 1, ®gs 2±3), which looks to be a more or less medianlongitudinal section, is 1?4 mm long and 0?55 mm in diameter. Other longitudinal fragments vary from
B A S S I E T A L . : T E R T I A R Y C O R A L L I N E A L G A E 409

0?72±1?48 mm in length and 0?31±0?55 mm in diameter (number of measurements, 12), while transversesections measure 0?31±0?78 mm in diameter (number of measurements, 14). No unequivocal branchapices were seen.
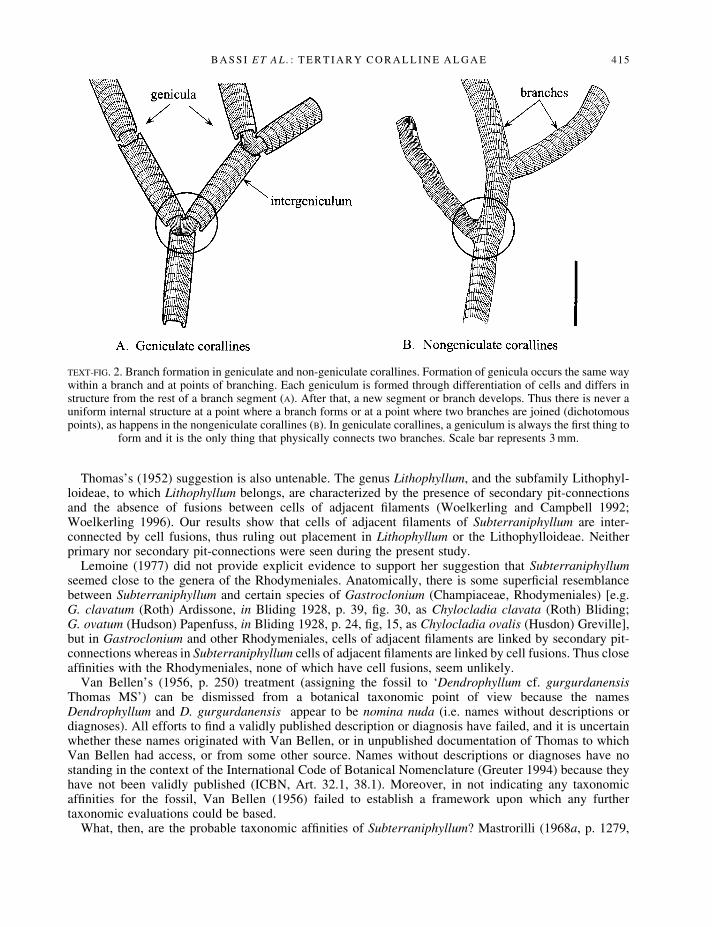
At least one fragment in V.34628 (Pl. 2, ®g. 1) and in V.34631 (Pl. 2, ®g. 2) show obvious points ofbranching, but in both cases, very little thallus is evident below the point of branching. The onset ofanother possible point of branching occurs in a fragment in V.34629 (Pl. 2, ®g. 3). These fragments show auniform internal structure with no changes in anatomy or cell arrangement at the points of branching. Thismeans that branch formation in S. thomasii does not involve genicula, because genicula always occur atpoints of branching and genicula always have an anatomy that is different from adjacent parts of a branch(Text-®g. 2; see also Woelkerling 1988, p. 60, ®gs 43±45, and Bassi 1998, p. 51, pl. 11, ®g. 1 forexamples).
In longitudinal view (Pl. 1, ®g. 3; Pl. 2, ®gs 2±3), Subterraniphyllum fragments appear pseudopar-enchymatous with a monomerous construction. In monomerous construction (see Woelkerling 1988,pp. 7±9, ®g. 16), there is a single system of ®laments in which some contribute to a core that runs more orless parallel to the surface of the thallus and others curve outwards and contribute collectively to a moreperipheral region. The relative proportions of ®laments in the core and peripheral regions can varysubstantially.
In Subterraniphyllum, the core region (commonly referred to as the medulla in the older literature; seeWoelkerling 1988, p. 17 for a discussion of this term) comprises most of the thallus, and individual core®laments can be traced for a number of cells. Cells are elongate (most commonly 2±3 times longer thanbroad), and the cells of adjacent ®laments are arranged more or less in tiers (i.e. coaxially). Sometimes thetiers appear arched (Pl. 2, ®g. 2), but more commonly they are planar except near the periphery, whereslight arching can occur (Pl. 1, ®g. 3; Pl. 2, ®g. 3). Cells in the core region were mostly 90±100 mm longand 30±40 mm in diameter as seen in longitudinal view. Thallus construction similar to that found inSubterraniphyllum occurs in some living corallines, including some species of Mesophyllum andSynarthrophyton (see Woelkerling and Harvey 1993; and Keats and Chamberlain 1994, 1997; Harveyet al. 1994; and Keats and Maneveldt 1997a for examples). It also occurs in some fossil species ofMesophyllum and Neogoniolithon (e.g. Bassi 1998, p. 35, pl. 3, ®g. 3; p. 41, pl. 6, ®g. 6). Near theperiphery, there is an abrupt transition to much smaller cells (Pl. 1, ®g. 3; Pl. 3, ®g. 2), mostly 10±15 mmlong and 10±15 mm in diameter as seen in longitudinal view. This kind of abrupt transition also occurs insome fossil corallines (e.g. Bassi 1998) and in some living corallines, including Synarthrophytonmunimentum Keats and Maneveldt (Keats and Maneveldt 1997a, p. 460, ®g. 32), S. patena (Hookerand Harvey, in Harvey 1849) Townsend (May and Woelkerling 1988, p. 59, ®gs 24±25; Harvey et al.1994, p. 334, ®gs 9±10), S. schielianum Woelkerling and Foster (Woelkerling and Foster 1989, pp. 46±47,®gs 12, 15±17), Mesophyllum lichenoides (Ellis) Lemoine (Woelkerling and Irvine 1986, p. 386, ®g. 16),and M. incisum (Foslie) Adey (Woelkerling and Harvey 1992, p. 387, ®gs 10±11; Keats and Maneveldt1997b, p. 204, ®gs 9, 11).
In transverse view (Pl. 3, ®g. 2), the thallus appears more or less circular in outline, suggesting that thebranches are more or less cylindrical. A comparatively narrow peripheral region composed of small cellssurrounds the large central core, where cells are much larger in sectional view. This re¯ects the structure asseen in longitudinal view and is consistent with the structure seen in some living corallines (see referencesabove).
410 P A L A E O N T O L O G Y , V O L U M E 4 3
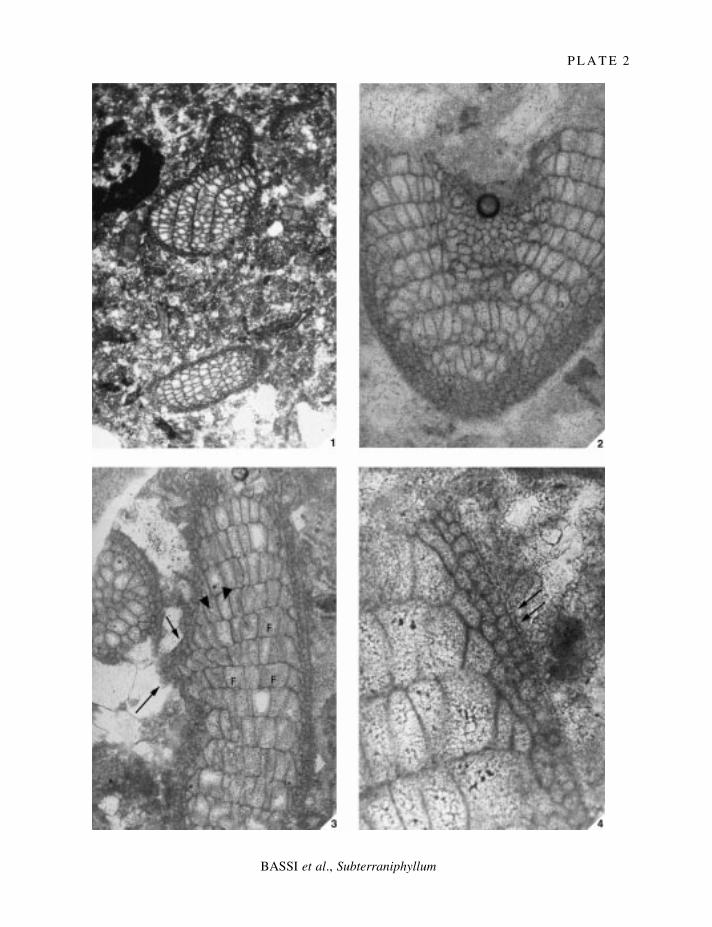
E X P L A N A T I O N O F P L A T E 2
Figs 1±4. Subterraniphyllum thomasii Elliott, 1957; Elliott's Collection. 1, isotype, V34628; oblique longitudinal sectionshowing a point of branching; ´ 40. 2, paratype, V34631; longitudinal section at a point of branching; note archedtiers of cells; ´ 90. 3, isotype, V34629; longitudinal view with slightly arched cells which are connected to oneanother by cell fusions (F); note possible onset of branching (arrows); arrowheads denote cell fusions in surfaceview; ´ 90. 4, isotype, V34629; sublongitudinal section showing possible epithallial cells (arrows); ´ 230.
P L A T E 2
BASSI et al., Subterraniphyllum

Cells resembling epithallial cells occur in several fragments (Pl. 2, ®g. 4) but are not well preserved.Outer walls look rounded rather than ¯attened and ¯ared; the former are characteristic of most genera ofliving corallines while the latter are characteristic of living species of Lithothamnion (e.g. see Woelkerling1988, p. 176, ®g. 192), Sporolithon (e.g. see Woelkerling 1988, p. 206, ®g. 246), and Exilicrusta(Chamberlain 1992, pp. 185±190, ®gs 7, 12).
Fusions between cells of adjacent ®laments occur (Pl. 2, ®g. 3; Pl. 3, ®gs 1±2), but apparently in smallnumbers, and in some fragments no cell fusions are obvious. Fusions involve the breakdown of part of thecommon walls between two cells (Pl. 3, ®gs 1±2), and when viewed from above (Pl. 2, ®g. 3, arrowheads),fusions may look like small circular holes in the cell wall. The pattern of cell fusions found inSubterraniphyllum thomasii also occurs in various species of Synarthrophyton [e.g. S. magellanicum(Foslie) Keats and Chamberlain: see Keats and Chamberlain 1997, p. 65, ®gs 32±33; S. patena: see Mayand Woelkerling 1988, p. 60, ®gs 27±28; Woelkerling 1988, p. 212, ®g. 256; Harvey et al. 1994, p. 335,®gs 9±11; S. schielianum: see Woelkerling and Foster 1989, p. 46, ®gs 12, 14]. Primary pit-connectionswere not evident in Elliott's material of Subterraniphyllum, probably owing to lack of preservation.Secondary pit-connections similarly were not evident, and no trichocytes or reproductive structures werefound.
The new material
Calcareniti di Castelgomberto. This formation crops out along the eastern margin of the Colli Berici(Vicenza, north-eastern Italy; Text-®g. 1) by showing steep morphological slopes with several karst-caves.It overlies the Marne di Priabona (Priabonian, Late Eocene; Ungaro 1969) and is overlain by a red clayhorizon followed by siliceous sands and sandstones (Ungaro 1978; `Arenarie di S. Urbano', EarlyMiocene, in Mietto 1988). The Calcareniti di Castelgomberto (Bosellini 1967), characterized by massiveor irregularly bedded limestones and marly limestones, range in the Colli Berici from about 80±200 m inthickness. The fossil content is mainly represented by corals, miliolids and coralline red algae.Nummulitids, soritids, molluscs, and echinoids are also present (Geister and Ungaro 1977; Ungaro1978; Frost 1981). The Eocene-Oligocene boundary in the Colli Berici has been recognized incorrespondence to the disappearance of the orthophragminids. The base of the Oligocene is characterizedby the occurrence of benthic foraminifera such as Pararotalia lithothamnica, Asterigerina rotulahaeringensis, Discorbis discoideas, and D. kiliani. This formation has been referred to the Early±Middle Oligocene by the occurrence of the foraminifera Nummulites ®chteli, N. vascus, Praerhapidioninadelicata, Spirolina cylindracea, Peneroplis glynnjonesi, and Asterigerina rotula haeringensis (Ungaro1978, pp. 245±246).
Subterraniphyllum thomasii occurred in sample nos. 74/10-11 (Pl. 3, ®g. 3) belonging to the Lumignanosection (Ungaro 1978, ®g. 5; Text-®g. 1). More or less transverse and oblique sections were present andwere largely concordant with fragments in the original Elliott collections. Cells in the core regionmeasured 60±70 mm long and 40±55 mm in diameter in the oblique sections; these are consistent with whatoccurs in the type material, although measurements on obliquely oriented cells are somewhat distortedrelative to those in longitudinal view. Reproductive structures did not occur.
412 P A L A E O N T O L O G Y , V O L U M E 4 3
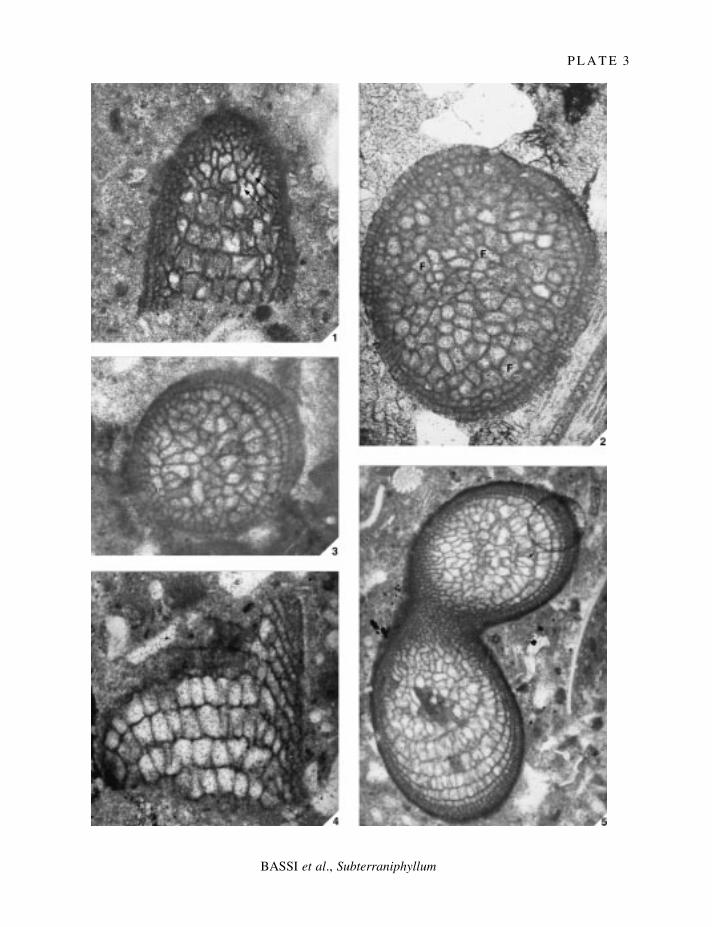
E X P L A N A T I O N O F P L A T E 3
Figs 1±2. Subterraniphyllum thomasii Elliott, 1957; Elliott's Collection. 1, isotype, V34628; oblique section, with cellfusions involving breakdown of part of the common walls between two cells (arrows); ´ 90. 2, isotype, V34630;transverse section showing the cell fusions (F); ´ 90.
Fig. 3. Subterraniphyllum thomasii Elliott, 1957; Ungaro's Collection (1978), Calcareniti di Castelgomberto,Oligocene of Lumignano, Colli Berici, north-eastern Italy; sample 74/10; oblique transverse section; ´ 90.
Figs 4±5. Subterraniphyllum thomasii Elliott, 1957; Gornji Grad Beds, Oligocene of Sv. Anton, Gornji Grad area,northern Slovenia; sample SLO94-044. 4, longitudinal section showing the peripheral cells; ´ 90. 5, oblique sectionat a branching point; note the uniform anatomy across the branch point; ´ 90.
P L A T E 3
BASSI et al., Subterraniphyllum

Gornji Grad Beds. The Gornji Grad area (northern Slovenia; Text-®g. 1) represents an important transitionzone between the northernmost extension of the Mediterranean Tethys to the south and the developingParatethys to the north and east. Although the rich fossil content and facies variations of the Gornji GradBeds are historically well documented (Reuss 1851, 1864; Morlot 1853; Rolle 1857; Stur 1871; Waters1874; Teller 1885, 1896, 1898), there have been few detailed modern examinations of its fauna, ¯ora aswell as microfacies (Hemleben 1964; Barta-Calmus 1973; Drobne et al. 1985; Bricl and Pavcic 1992). TheGornji Grad Beds (formally known as the `Oberburger Schichten', Auctorum) form part of a highly variedtransgressive sequence over a Triassic basement. This includes a basal series of terrigenous sediments andconglomerates followed by the Gornji Grad Beds, overlain by marls rich in foraminifera which are cappedby thick series of volcanoclastic tuffs. The Gornji Grad Beds themselves consist of both siliciclastics andcarbonates of marine origin and transgress above Triassic and Eocene limestones; the basal terrigenoussediments (see above) or conglomerates are of indeterminate age (possibly Eocene). The stratigraphicalframework of Gornji Grad is given by Drobne et al. (1985, p. 82) who recognised Nummulites ®chteli andN. germanicus as indicating an early Oligocene age. Furthermore, nannoplankton occurring in the marlybeds collected just above `the reef structures' (Auctorum; upper part of the Gornji Grad Beds) indicate theNP 23 zone (upper Lower Oligocene; Bricl and Pavcic 1992).
The carbonates consist predominantly of poorly sorted, biogenic rudstones with a wackestone matrix;pack- and grainstones are subordinate. The principal components are represented by coralline red algaetogether with a moderately diverse coral fauna including both solitary and colonial forms. Large and smallbenthic foraminifera as well as calcitic shelled bivalves are also common. Subordinate are gastropods,bryozoans, brachiopods, echinoderms, serpulids and green (dasycladacean and halimedacean) algae.
Subterraniphyllum thomasii was found only in sample SLO94-044 from `Sv. Anton' section (Text-®g. 1;Pl. 3, ®gs 4±5). Longitudinal, transverse and oblique fragments were present and were morphologicallyand anatomically concordant with fragments in the original Elliott collections except that in longitudinalview cells in the core region showed a somewhat greater range in length (65±95 mm) and were somewhatgreater in diameter (45±65 mm). In living corallines, such variation in cell size is of no taxonomicconsequence (e.g. Woelkerling and Campbell 1992; Woelkerling and Harvey 1993; Wilks and Woelk-erling 1994); and Johnson (1966, p. 277) reported similar variation in specimens of Subterraniphyllumthomasii from Borneo, Indonesia. Reproductive structures did not occur.
D I S C U S S I O N
The taxonomic placement of Subterraniphyllum
At least four suggestions on the taxonomic placement of Subterraniphyllum have appeared. Elliott (1957)and most subsequent authors (e.g. Johnson 1965, 1966; Beckmann and Beckmann 1966; Mastrorilli1968a, b) have regarded Subterraniphyllum to be a distinct genus of geniculate coralline red algae and tobelong to the Corallinaceae, subfamily Corallinoideae (sensu Lemoine 1939 and Mason 1953). Poignant(1985) similarly considered Subterraniphyllum to have genicula (`thalles articuleÂs') but did not indicatefamily or subfamily placement in his key. Lemoine (1977, pp. 34, 36), by contrast, suggested that thestructure of Subterraniphyllum did not resemble that of other genera of Corallinaceae but rather seemedcloser to the genera of the red algal order Rhodymeniales. Lemoine, however, did not formally assignSubterraniphyllum to the Rhodymeniales. Thomas (1952, p. 78) thought that it resembled Lithophyllum,while Van Bellen (1956) applied the name Dendrophyllum cf. gurgurdanensis Thomas MS withoutassigning it to a higher taxonomic group. None of the above suggestions is supported by the evidenceobtained during the present study.
Our results show that genicula do not occur (i.e. that Subterraniphyllum is not `segmented') because thefragments have a uniform anatomy across points of branching. If genicula were present, there would be anobvious, marked change in anatomy at points of branching (Text-®g. 2; Woelkerling 1988, p. 60, ®gs 43±45; Bassi 1998, p. 51, pl. 11, ®g. 1). Thus, Subterraniphyllum cannot be a segmented (geniculate) corallineas presumed by Elliott (1957, p. 73) and most subsequent authors, and it does not belong to theCorallinaceae, subfamily Corallinoideae or any other group characterized by the presence of genicula.
414 P A L A E O N T O L O G Y , V O L U M E 4 3
Thomas's (1952) suggestion is also untenable. The genus Lithophyllum, and the subfamily Lithophyl-loideae, to which Lithophyllum belongs, are characterized by the presence of secondary pit-connectionsand the absence of fusions between cells of adjacent ®laments (Woelkerling and Campbell 1992;Woelkerling 1996). Our results show that cells of adjacent ®laments of Subterraniphyllum are inter-connected by cell fusions, thus ruling out placement in Lithophyllum or the Lithophylloideae. Neitherprimary nor secondary pit-connections were seen during the present study.
Lemoine (1977) did not provide explicit evidence to support her suggestion that Subterraniphyllumseemed close to the genera of the Rhodymeniales. Anatomically, there is some super®cial resemblancebetween Subterraniphyllum and certain species of Gastroclonium (Champiaceae, Rhodymeniales) [e.g.G. clavatum (Roth) Ardissone, in Bliding 1928, p. 39, ®g. 30, as Chylocladia clavata (Roth) Bliding;G. ovatum (Hudson) Papenfuss, in Bliding 1928, p. 24, ®g, 15, as Chylocladia ovalis (Husdon) Greville],but in Gastroclonium and other Rhodymeniales, cells of adjacent ®laments are linked by secondary pit-connections whereas in Subterraniphyllum cells of adjacent ®laments are linked by cell fusions. Thus closeaf®nities with the Rhodymeniales, none of which have cell fusions, seem unlikely.
Van Bellen's (1956, p. 250) treatment (assigning the fossil to `Dendrophyllum cf. gurgurdanensisThomas MS') can be dismissed from a botanical taxonomic point of view because the namesDendrophyllum and D. gurgurdanensis appear to be nomina nuda (i.e. names without descriptions ordiagnoses). All efforts to ®nd a validly published description or diagnosis have failed, and it is uncertainwhether these names originated with Van Bellen, or in unpublished documentation of Thomas to whichVan Bellen had access, or from some other source. Names without descriptions or diagnoses have nostanding in the context of the International Code of Botanical Nomenclature (Greuter 1994) because theyhave not been validly published (ICBN, Art. 32.1, 38.1). Moreover, in not indicating any taxonomicaf®nities for the fossil, Van Bellen (1956) failed to establish a framework upon which any furthertaxonomic evaluations could be based.
What, then, are the probable taxonomic af®nities of Subterraniphyllum? Mastrorilli (1968a, p. 1279,
B A S S I E T A L . : T E R T I A R Y C O R A L L I N E A L G A E 415
TEXT-FIG. 2. Branch formation in geniculate and non-geniculate corallines. Formation of genicula occurs the same waywithin a branch and at points of branching. Each geniculum is formed through differentiation of cells and differs instructure from the rest of a branch segment (A). After that, a new segment or branch develops. Thus there is never auniform internal structure at a point where a branch forms or at a point where two branches are joined (dichotomouspoints), as happens in the nongeniculate corallines (B). In geniculate corallines, a geniculum is always the ®rst thing to
form and it is the only thing that physically connects two branches. Scale bar represents 3 mm.

pl. 102, ®gs 1±2), who placed the genus in the Corallinaceae, subfamily Corallinoideae, reported a cavityresembling a uniporate conceptacle at the surface of a transverse section of Subterraniphyllum. Poignant(1985) also considered that Subterraniphyllum had uniporate conceptacles, presumably on the basis of therecord of Mastrorilli. Unfortunately, it is not clear from the ®gures of Mastrorilli (1968a) whether thepossible uniporate conceptacle was actually produced by Subterraniphyllum or whether instead it belongsto an epiphyte on the surface of Subterraniphyllum. Until unequivocal, independent evidence ofconceptacles is found for Subterraniphyllum, this single report needs to be treated with caution.
The absence of reproductive structures in type material of Subterraniphyllum and the uncertaintyattending the report of Mastrorilli (1968a) mean that at present we cannot state unequivocally thatSubterraniphyllum possesses conceptacles. On vegetative grounds, however, there is nothing to excludeSubterraniphyllum from the Corallinales, and, as noted above, the vegetative anatomy of Subterrani-phyllum is similar to that of some other Corallinales, the view of Lemoine (1977) notwithstanding. Wesuggest, therefore, that Subterraniphyllum be retained in the Corallinales as a genus of uncertain position.
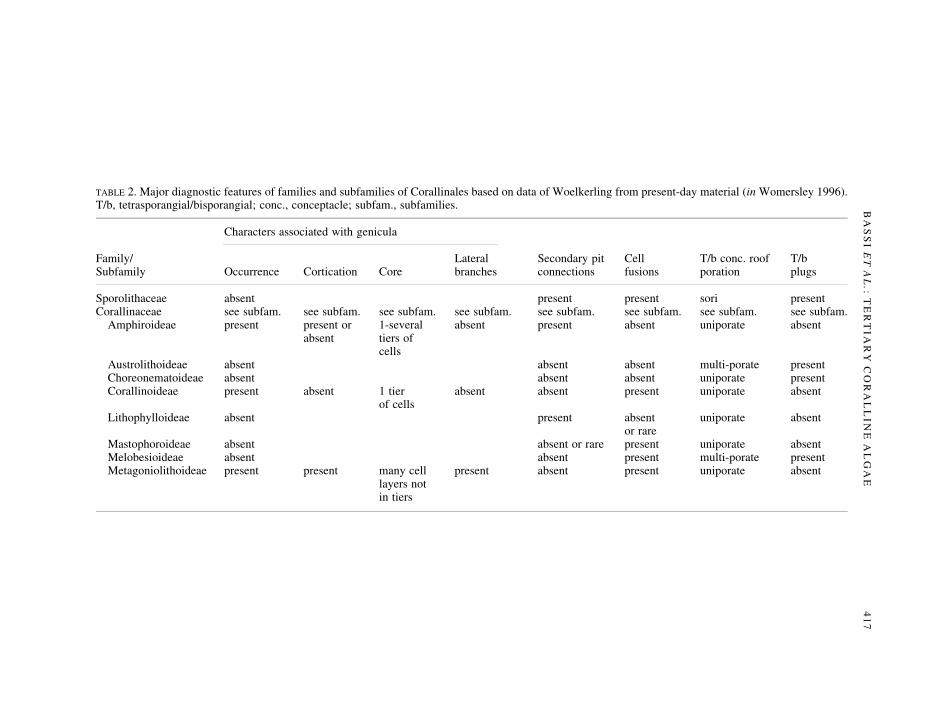
Using the Corallinales classi®cation system employed by Woelkerling (in Womersley 1996) (Table 2)as a framework, the possible af®nities of Subterraniphyllum can be somewhat more fully determined. Theabsence of genicula precludes possible af®nities with the subfamilies Amphiroideae, Corallinoideae andMetagoniolithoideae in the family Corallinaceae. All species in these subfamilies possess genicula. Thepresence in Subterraniphyllum of fusions between cells of adjacent ®laments precludes af®nities with theCorallinaceae, subfamilies Lithophylloideae, Choreonematoideae and Austrolithoideae, all of whichapparently lack cell fusions.
There is no ®rm evidence at present that would exclude Subterraniphyllum from the Sporolithaceae, orfrom the Corallinaceae, subfamily Mastophoroideae, or from the Corallinaceae, subfamily Melobesioi-deae. The possible but poorly preserved epithallial cells observed during the present study appear not tohave the ¯attened and ¯ared cells characteristic of species of Sporolithaceae and of the genera Exilicrustaand Lithothamnion in the subfamily Melobesioideae, but better preserved epithallial cells must be foundbefore any ®rm conclusions can be reached.
In terms of vegetative anatomy, Subterraniphyllum most closely resembles some living species ofMesophyllum and Synarthrophyton (see references cited above), although species of Mesophyllum andSynarthrophyton generally produce ¯attened branches rather than cylindrical branches. Amongst livingspecies of corallines, however, differences in cross-sectional shape of branches is not considered to betaxonomically signi®cant at generic level. Both Mesophyllum and Synarthrophyton belong to theCorallinaceae, subfamily Melobesioideae. All Melobesioideae produce tetrasporangia in multiporateconceptacles but produce gametangia within uniporate conceptacles. Thus the report of a uniporate cavityby Mastrorilli (1968a) does not preclude the possibility that Subterraniphyllum belongs to the subfamilyMelobesioideae. It is also possible, however, that Subterraniphyllum belongs to the subfamily Masto-phoroideae, which is characterized by uniporate tetrasporangial conceptacles. The matter cannot bedecided in the absence of further data on reproduction in Subterraniphyllum; consequently, Subterrani-phyllum is best treated as a genus of uncertain position within the Corallinales.
The possible relationships of Subterraniphyllum to other fossil genera ascribed at some stage to theCorallinales (see Woelkerling 1988, pp. 44±48 for a listing) awaits further study, including reinvestiga-tions of the original material on which these genera are based.
Age and stratigraphical distribution of Subterraniphyllum
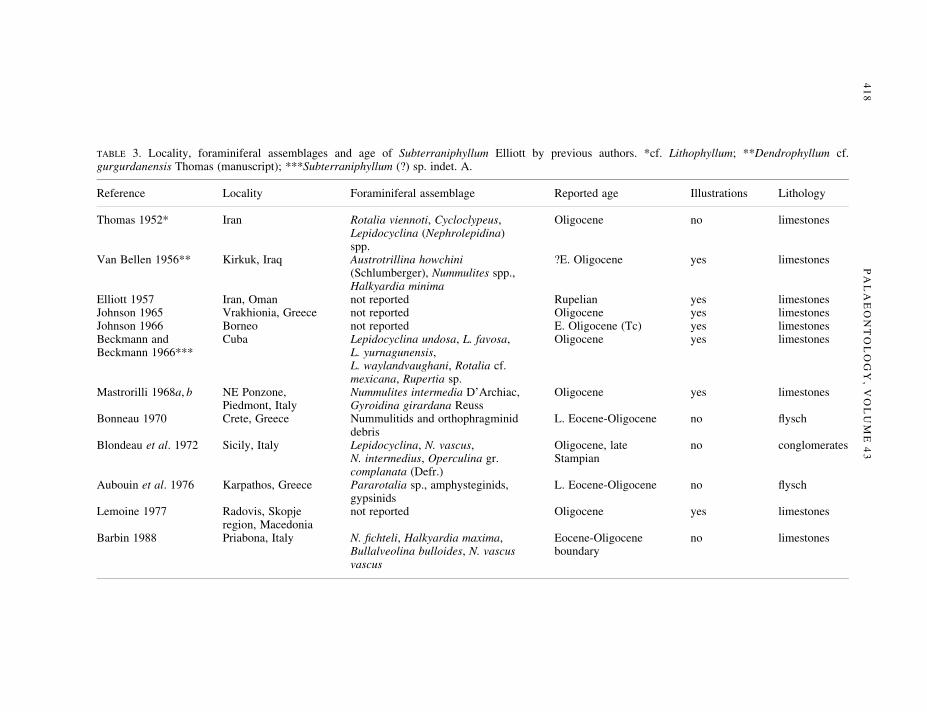
Elliott (1957) suggested that Subterraniphyllum could be used as a biostratigraphical marker because itappeared to be restricted to Upper Eocene, Oligocene and Lower Miocene carbonate successions. Thereported occurrences of this taxon in the Mediterranean and Paratethys are summarized in Table 3.
The ®rst record is that of Thomas (1952), who found Subterraniphyllum (reported as `cf. Lithophyllum',p. 78) at the bottom of the Asmari Limestones in the Gach Saran area. These beds are set between theEocene±Oligocene marls with Haplophragmina slingeri Thomas below, and the Oligocene limestoneswith Praerapydionina delicata Henson above. Moreover, Thomas af®rmed that the distinctive species`cf. Lithophyllum' also occurs in the middle part of the Khamir Limestone of Kuh-i-Khamir together with
416 P A L A E O N T O L O G Y , V O L U M E 4 3
BA
SS
IE
TA
L.:
TE
RT
IA
RY
CO
RA
LL
IN
EA
LG
AE
41
7
TABLE 2. Major diagnostic features of families and subfamilies of Corallinales based on data of Woelkerling from present-day material (in Womersley 1996).T/b, tetrasporangial/bisporangial; conc., conceptacle; subfam., subfamilies.
Characters associated with genicula
Family/ Lateral Secondary pit Cell T/b conc. roof T/bSubfamily Occurrence Cortication Core branches connections fusions poration plugs
Sporolithaceae absent present present sori presentCorallinaceae see subfam. see subfam. see subfam. see subfam. see subfam. see subfam. see subfam. see subfam.
Amphiroideae present present or 1-several absent present absent uniporate absentabsent tiers of
cellsAustrolithoideae absent absent absent multi-porate presentChoreonematoideae absent absent absent uniporate presentCorallinoideae present absent 1 tier absent absent present uniporate absent
of cellsLithophylloideae absent present absent uniporate absent
or rareMastophoroideae absent absent or rare present uniporate absentMelobesioideae absent absent present multi-porate presentMetagoniolithoideae present present many cell present absent present uniporate absent
layers notin tiers
41
8P
AL
AE
ON
TO
LO
GY
,V
OL
UM
E4
3
TABLE 3. Locality, foraminiferal assemblages and age of Subterraniphyllum Elliott by previous authors. *cf. Lithophyllum; **Dendrophyllum cf.gurgurdanensis Thomas (manuscript); ***Subterraniphyllum (?) sp. indet. A.
Reference Locality Foraminiferal assemblage Reported age Illustrations Lithology
Thomas 1952* Iran Rotalia viennoti, Cycloclypeus, Oligocene no limestonesLepidocyclina (Nephrolepidina)spp.
Van Bellen 1956** Kirkuk, Iraq Austrotrillina howchini ?E. Oligocene yes limestones(Schlumberger), Nummulites spp.,Halkyardia minima
Elliott 1957 Iran, Oman not reported Rupelian yes limestonesJohnson 1965 Vrakhionia, Greece not reported Oligocene yes limestonesJohnson 1966 Borneo not reported E. Oligocene (Tc) yes limestonesBeckmann and Cuba Lepidocyclina undosa, L. favosa, Oligocene yes limestonesBeckmann 1966*** L. yurnagunensis,
L. waylandvaughani, Rotalia cf.mexicana, Rupertia sp.
Mastrorilli 1968a, b NE Ponzone, Nummulites intermedia D'Archiac, Oligocene yes limestonesPiedmont, Italy Gyroidina girardana Reuss
Bonneau 1970 Crete, Greece Nummulitids and orthophragminid L. Eocene-Oligocene no ¯yschdebris
Blondeau et al. 1972 Sicily, Italy Lepidocyclina, N. vascus, Oligocene, late no conglomeratesN. intermedius, Operculina gr. Stampiancomplanata (Defr.)
Aubouin et al. 1976 Karpathos, Greece Pararotalia sp., amphysteginids, L. Eocene-Oligocene no ¯yschgypsinids
Lemoine 1977 Radovis, Skopje not reported Oligocene yes limestonesregion, Macedonia
Barbin 1988 Priabona, Italy N. ®chteli, Halkyardia maxima, Eocene-Oligocene no limestonesBullalveolina bulloides, N. vascus boundaryvascus

Nummulites ®chteli (p. 79). This latter is presently considered a marker for the Oligocene shallow benthicbiozones 21±22B (Rupelian±Early Chattian; Cahuzac and Poignant 1997).
Afterwards, Elliott (1957) assigned these specimens to Subterraniphyllum thomasii, but did not provideany information on the biostratigraphical setting of the type level. The record of this taxon from theAquitanian (Early Miocene) of Sicily and Iran reported by Elliott (1957) is not con®rmed by any referencenor by successive records. In the Late Eocene±Oligocene of Crete and Karpatos (Greece), Bonneau (1970)and Aubouin et al. (1976) recorded Subterraniphyllum thomasii in ¯ysch deposits, together with largerforaminifera such as nummulitids, orthophragminids, and amphisteginids. This stratigraphical assignment,however, cannot be veri®ed because no species was reported. The stratigraphical setting of Subterrani-phyllum ascribed to the Oligocene by Beckmann and Beckmann (1966), Blondeau et al. (1972), andBarbin (1988) seems to be con®rmed by the occurrence of biostratigraphical markers such as Nummulites®chteli, Nummulites vascus, Halkyardia maxima, and Lepidocyclina (Eulepidina) undosa (see Butterlin1981; Cahuzac and Poignant 1997). However, Blondeau et al. (1972) and Barbin (1988) did not illustratethe algal specimens. Other authors referred Subterraniphyllum to the Oligocene on the base of the corallinealgal assemblages (Johnson 1965, 1966; Lemoine 1977) or on foraminifera without biostratigraphicalmeaning (Mastrorilli 1968b).
The newly recorded specimens of Subterraniphyllum from northern Slovenia and north-eastern Italydescribed herein appear to originate from Oligocene rocks. The comparison of the well-documentedreports of Subterraniphyllum to biostratigraphically meaningful, large foraminiferal assemblages alsosuggests that its occurrence in Upper Eocene or Lower Miocene successions cannot be veri®ed.Subterraniphyllum cannot, however, be considered as a useful stratigraphical marker until further datawith respect to its occurrence in well-dated carbonate sequences are acquired.
C O N C L U D I N G R E M A R K S
The taxonomy of living species of coralline red algae (Corallinales) has undergone a marked change since1960 as a consequence of a number of major studies, many of which are cited in Johansen (1981),Woelkerling (1988), Irvine and Chamberlain (1994), Woelkerling (in Womersley 1996), and Woelkerlingand Lamy (1998). The group is now treated as a separate order of Rhodophyta (Silva and Johansen 1986),and is considered to represent a major evolutionary line within the red algae (Bailey and Chapman 1996,p. 364). Concepts of families, subfamilies, genera, and species have changed as a result of extensive newinformation which has shown that many characters relating to vegetative morphology and anatomy onceused to separate taxa vary too much to be taxonomically reliable. New genera have been described, andsome genera commonly used in the older literature have been relegated to synonymy. While there still isno consensus on which of several current proposals (e.g. see Cabioch 1972, 1988; Woelkerling, inWomersley 1996; Desikachary et al. 1998; Bailey 1999; older schemes listed by Woelkerling 1988,pp. 54±55) for the classi®cation of Corallinales at family, subfamily and tribe levels best re¯ects thephylogeny of the group, taxonomic concepts for many species and genera are far different now from whatthey were in 1960.
These developments have signi®cant implications for the taxonomy of fossil coralline red algae. Manyspecies and genera known only from the fossil record are based on characters that are now known to be ofno taxonomic value for delimiting living corallines, and thus by implication are of questionable value fordelimiting fossil taxa. This problem has been recognised by a number of people working on fossilcorallines (e.g. Bosence 1991, pp. 103±108; Braga et al. 1993; Iryu and Matsuda 1994, p. 444; Bassi 1995,p. 82; Basso 1995a, p. 576; Braga and Aguirre 1995; Rasser and Piller 1999). There is also an increasingrealization (see references above) that most fossil species and genera considered to be coralline red algaerequire taxonomic reassessment in the context of current knowledge of living corallines, and that re-examination of type collections and/or of material from type localities forms an essential part of thereassessment process, as exempli®ed by Aguirre and Braga (1998).
Our exclusion of Subterraniphyllum from any group of Corallinaceae with genicula is based onunequivocal evidence that branch formation does not involve the occurrence of genicula. By contrast, theinclusion of most fossil species in various geniculate genera of Corallinales (Table 1) by most authors is
B A S S I E T A L . : T E R T I A R Y C O R A L L I N E A L G A E 419

based on the supposition that genicula occur (both at points of branching and within branches), eventhough they have not been observed. In reality, genicula are unknown for most fossil species placed ingeniculate coralline genera, and indeed, genicula, as noted by Lemoine (1939), Johnson (1961), Bosence(1991) and Bassi (1998), are seldom preserved in fossil material.
In the absence of genicula, the placement of most fossil species into geniculate genera has been basedmainly on vegetative features observable in fragments presumed to be segments of geniculate branches.This is evident, for example, in keys to genera published by Lemoine (1970) and Poignant (1985) and inmany accounts of species placed in geniculate genera (e.g. see Ishijima 1954, pp. 53±75; Johnson 1964,pp. 30±36; Mastrorilli 1968b, pp. 380±388; Lemoine 1970, pp. 192±193). Such placements of fossilspecies in geniculate genera must be treated with caution, especially in the absence of further assessmentsof these fossils in relation to current concepts for geniculate coralline genera (e.g. see Johansen 1976;Womersley and Johansen 1996a±c), all of which are based on studies of living species. Bassi (1998, p. 21),recognising the dif®culties of placing specimens into geniculate genera without appropriate evidence, choseinstead to group all potentially geniculate fragments under the informal group `Geniculates sensu lato'. Thisapproach merits further application as it avoids the continued speculative use of names for fossil material.
The absence of critical characters does not mean that identi®cation is impossible, however, assometimes partial or even full identi®cation can be made from vegetative features that are preserved.For all those many fossil specimens where unequivocal evidence is not present, it is possible to recognisethat the specimens cannot be placed with certainty, and to utilise `form genera' to accommodate thesespecimens based on the characters which are normally preserved. This effort leads to a much betterunderstanding of fossil corallines in terms of which ones can or cannot be unequivocally associated withparticular geniculate genera while at the same time creating a consistent, workable system of applyingnames to most fossil corallines so that they can be reliably used in relation to stratigraphical andpalaeoecological studies.
Coralline red algae are more extensively represented in the fossil record than any other group ofRhodophyta, and as a consequence, it is theoretically possible to understand more about the geological andevolutionary history of this group than of any other group of red algae. The extent to which information inthe fossil record can contribute to our overall understanding of the Corallinales is dependent on continuedinteractions between those who work on living corallines and those who work on fossil corallines, andupon striving to interpret the taxonomy of this group using the same classi®cation system for both livingand fossil taxa. As taxonomic knowledge of living and fossil taxa improves, it will also be possible toassess more accurately, and place greater con®dence in, the stratigraphical value of particular fossilcorallines.
Acknowledgements. This study was supported by Italian MURST ex40% (C. Loriga) at the University of Ferrara, Italy;by the Department of Botany (W. J. Woelkerling) at La Trobe University, Australia; and by the DFG (German ScienceFoundation) SFB275 (Sonderforschungsbereich) at the University of TuÈbingen, Germany. We thank T. S. Foster,Curator of the Invertebrates and Plants Division (Department of Palaeontology, The Natural History Museum,London); S. Ungaro (University of Ferrara) for his kindness in facilitating the study of his original material sampled inthe Oligocene of the Colli Berici; P. Silva (University of California, Berkeley), who kindly con®rmed that he knew ofno validly published description or diagnosis for Dendrophyllum or for D. gurgurdanensis; C. Loriga Broglio(University of Ferrara), C. Hemleben, V. Mosbrugger (University of TuÈbingen), and K. Drobne (Slovenian Academyof Sciences and Arts, Ljubljana). We have sincerely appreciated the efforts of the referees of our original manuscriptfor their suggestions.
R E F E R E N C E S
AGUIRRE, J. and BRAGA, J. C. 1998. Redescription of Lemoine's (1939) types of coralline algal species from Algeria.Palaeontology, 41, 489±507.
ÐÐ ÐÐ and PILLER, W. E. 1996. Reassessment of Palaeothamnium Conti, 1946 (Corallinales, Rhodophyta). Reviewof Palaeobotany and Palynology, 94, 1±9.
ARESCHOUG, J. E. 1852. Ordo XII. Corallinaceae. 506±576. In AGARDH, J. G. (ed.). Species, Genera, et Ordines Algarum.Volume 2, Part 2. C. W. K. Gleerup, Lund, 750 pp.
420 P A L A E O N T O L O G Y , V O L U M E 4 3

AUBOUIN, J., BONNEAU, M. and DAVIDSON, J. 1976. Contribution aÁ la geÂologie de l'arc eÂgeÂen: l'õÃle de Karpathos. Bulletinde la Societe GeÂologique de France, 18, 385±401.
BAILEY, J. C. 1999. Phylogenetic positions of Lithophyllum incrustans and Titanoderma pustulatum (Corallinaceae,Rhodophyta) based on 18S rRNA gene sequence analyses, with a revised classi®cation of the Lithophylloideae.Phycologia, 38, 208±216.
ÐÐ and CHAPMAN, R. L. 1996. Evolutionary relationships among coralline red algae (Corallinaceae, Rhodophyta)inferred from 18S rRNA gene sequence analysis. 363±376. In CHAUDHARY, B. R. and AGRAWAL, S. B. (eds). Cytology,genetics and molecular biology of algae. Academic Publishing, Amsterdam, 439 pp.
BARBIN, V. 1988. The Eocene-Oligocene transition in shallow-water environment: the Priabonian stage type (Vicentin,northern Italy). 163±171. In PREMOLI SILVA, I., COCCIONI, R. and MONTANARI, A. (eds). The Eocene-Oligoceneboundary in the Marche-Umbria basin (Italy). International Subcommission on Paleogene Stratigraphy, AnconaMeeting, Italy, 268 pp.
BARTA-CALMUS, S. 1973. Revision de collections de madreÁporaires du Nummulitique du sud-est de la France, de l'Italieet de la Yougoslavie septentrionales. Unpublished dissertation thesis, University of Paris.
BASSI, D. 1995. Crustose coralline algal pavements from Late Eocene Colli Berici of northern Italy. Rivista Italiana diPaleontologia e Stratigra®a, 101, 81±92.
ÐÐ 1998. Coralline red algae (Corallinales, Rhodophyta) from the Upper Eocene Calcare di Nago (Lake Garda,northern Italy). Annali dell'UniversitaÁ di Ferrara, 7, Supplemento, 1±51.
ÐÐ and NEBELSICK, J. 1997. Coralline red algae from the lower Oligocene Gornji Grad formation, Slovenia: ®rstresults. GeoItalia ± 1st Forum, Italiano di Scienze della Terra, Rimini (Bellaria), Italy, Abstracts, 54.
BASSO, D. 1995a. Study of living calcareous algae by a paleontological approach: the non-geniculate Corallinaceae(Rhodophyta) of the soft bottoms of the Tyrrhenian Sea (Western Mediterranean). The genera Phymatolithon Foslieand Mesophyllum Lemoine. Rivista Italiana di Paleontologia e Stratigra®a, 100, 575±596.
ÐÐ 1995b. Living calcareous algae by a paleontological approach: the genus Lithothamnion Heydrich nom. cons.from the soft bottoms of the Tyrrhenian Sea (Mediterranean). Rivista Italiana di Paleontologia e Stratigra®a, 101, 349±366.
ÐÐ FRAVEGA, P. and VANNUCCI, G. 1997. The taxonomy of Lithothamnium ramosissimum (GuÈmbel non Reuss) Contiand Lithothamnium operculatum (Conti) Conti (Rhodophyta, Corallinaceae). Facies, 37, 167±182.
BECKMANN, J. P. and BECKMANN, R. 1966. Calcareous algae from the Cretaceous and Tertiary of Cuba. SchweizerischePalaontologische Abhandlungen, 85, 1±45.
BLIDING, C. 1928. Studien uÈber die Florideenordnung Rhodymeniales. Lunds Universitets ÊArsskrift, Ny FoÈljd,Avdelningen 2 (Medicin samt matematiska och naturvetenskapliga aÈmnen), 24 (3), 1-74.
BLONDEAU, A., LORENZ, C., MAGNE , J. and MASCLE, G. 1972. L'OligoceÁne et le MioceÁne infeÂrieur en Sicile centro-occidentale. Bulletin de la Societe GeÂologique de France, 14, 137±146.
BONNEAU, M. 1970. Les lambeaux allochtones du revers septentrional du massif des Psiloriti (CreÃte moyenne, GreÁce).Bulletin de la Societe GeÂologique de France, 12, 1124±1129.
BOSELLINI, A. 1967. Calcareniti di Castelgomberto. 25±26. In BOSELLINI, A., CARRARO, F., CORSI, M., DE VECCHI, G. P.,
GATTO, G. O., MALARODA, R., STURANI, C., UNGARO, S. and ZANNETTIN, B. (eds). Note illustrative della Carta geologicad'Italia, Foglio 49, Verona. Servizio Geologico d'Italia, Roma, 61 pp.
BOSENCE, D. W. J. 1991. Coralline algae: mineralization, taxonomy and palaeoecology. 98-113. In RIDING, R. (ed.).Calcareous algae and stromatolites. Springer-Verlag, Berlin, 571 pp.
BRAGA, J. C. and AGUIRRE, J. 1995. Taxonomy of fossil coralline algal species: Neogene Lithophylloideae (rhodophyta,Corallinaceae) from southern Spain. Review of Palaeobotany and Palynology, 86, 265±285.
ÐÐ BOSENCE, D. W. J. and STENECK, R. S. 1993. New anatomical characters in fossil coralline algae and their taxonomicimplications. Palaeontology, 36, 535±547.
BRICL, B. and PAVCIC, J. 1992. Frequency of nannoplankton in Oligocene marine clay in Slovenia. Slovenska AkademijaZnanosti in Umetnosti, Razprave IV Raureda, 32, 154±173.
BUTTERLIN, J. 1981. Claves para la determinacioÂn de macroforaminõÂferos de MeÂxico y del Caribe, del CretaÂcicoSuperior al Mioceno Medio. Instituto Mexicano del PetroÂleo, SubdireccioÂn de TecnologõÂa de ExploracioÂn, 1±219.
CABIOCH, J. 1972. EÂ tude sur les CorallinaceÂes. II. La morphogeneÁse: conseÂquences systeÂmatiques et phylogeÂneÂtiques.Cahiers de Biologie Marine, 13, 137±288.
ÐÐ 1988. Morphogenesis and generic concepts in coralline algae: a reappraisal. HelgolaÈnder Meeresuntersuchun-gen, 42, 493±509.
CAHUZAC, B. and POIGNANT, A. 1997. Essai de biozonation de l'Oligo-MioceÁne dans les bassins europeÂens aÁ l'aide desgrands foraminifeÁres neÂritiques. Bulletin de la SocieÂte GeÂologique de France, 168, 155±169.
CHAMBERLAIN, Y. M. 1992. Observations on two melobesioid crustose coralline red algal species from the British Isles:
B A S S I E T A L . : T E R T I A R Y C O R A L L I N E A L G A E 421

Exilicrusta parva, a new genus and species, and Lithothamnion sonderi Hauck. British Phycological Journal, 27,185±201.
CONTI, S. 1950. Alghe corallinacee fossili. Pubblicazioni dell'Istituto di Geologia dell'UniversitaÁ di Genova,Paleontologia, 4 (Serie A), 156 pp.
DECAISNE, J. 1842. Essais sur une classi®cation des Algues et des Polypiers calcifeÁres de Lamourous. Annales desSciences Naturelles, 2 SeÂries Botanique, 17, 297±380.
DESIKACHARY, T. V., KRISHNAMURTHY, V. and BALAKRISHNAN, M. S. 1998. Rhodophyta Vol. 2, Part 2B. Madras ScienceFoundation, Chennai, India, 359 pp.
DROBNE, K., PAVLOVEC, R., DROBNE, F., CIMERMAN, F. and SÏIKICÂ , L. 1985. Some larger foraminifera from the Upper Eoceneand the basal Oligocene beds in North Slovenia. GeolosÏki Glasnik, 28, 77±117.
ELLIOTT, G. F. 1957. Subterraniphyllum, a new Tertiary calcareous alga. Palaeontology, 1, 73±75.FROST, S. H. 1981. Oligocene reef coral biofacies of the Vicentin. 483±539. In TOOMEY, D. F. (ed.). European fossil reef
models. Special Publication of the Society of Economic Paleontologists and Mineralogists, 30, 1±546.GEISTER, J. and UNGARO, S. 1977. The Oligocene coral formations of the Colli Berici (Vicenza, northern Italy). Eclogae
Geologicae Helvetiae, 70, 811±823.GREUTER, W. 1994. International Code of Botanical Nomenclature (Tokyo Code) adopted by the Fifteenth International
Botanical Congress, Yokohama, August ± September 1993. Regnum Vegetabile 131. Koeltz Scienti®c Books,KoÈnigstein, Germany, xviii� 389 pp.
HARVEY, A. S., WOELKERLING, W. M. J. and WILKS, K. M. 1994. The genus Synarthrophyton (Corallinaceae, Rhodophyta) insouthern Australia. Phycologia, 33, 331±342.
HARVEY, W. H. 1849. Nereis Australis. Reeve, London, Part 2, 65±124 [For data on publication date, see Taxon (1968),17, 82].
HEMLEBEN, C. 1964. Geologisch-palaÈontologisch Untersuchung im Gebiet zwischen Gornji Grad (Oberburg) und NovaStifta (Neustift) in Nordslowenien/Jugoslawien. Unpublished Masters thesis, University of MuÈnich.
IRVINE, L. M. and CHAMBERLAIN, Y. M. 1994. Seaweeds of the British Isles. Volume 1 Rhodophyta, Part 2B, Corallinales,Hildenbrandiales. HMSO, London, vii� 276 pp.
IRYU, Y. and MATSUDA, S. 1994. Taxonomic studies of the Neogoniolithon fosliei complex (Corallinaceae, Rhodophyta) inthe Ryukyu Islands. Transactions and Proceedings of Palaeontological Society of Japan, New Series, 174, 426±448.
ISHIJIMA, W. 1954. Cenozoic coralline algae from the western Paci®c. Privately published, Tokyo, 87 pp.JOHANSEN, H. W. 1976. Current status of generic concepts in coralline algae. Phycologia, 21, 221±244.ÐÐ 1981. Coralline algae, a ®rst synthesis. CRC Press, Boca Raton, Florida, 239 pp.JOHNSON, J. H. 1961. Limestone-building algae and algal limestones. Colorado School of Mines, Boulder, 297 pp.ÐÐ 1964. Fossil and Recent calcareous algae from Guam. Professional Paper of the United States Geological
Survey, 403G, 1±40.ÐÐ 1965. Coralline algae from the Cretaceous and Early Tertiary of Greece. Journal of Paleontology, 39, 802±814.ÐÐ 1966. Tertiary red algae from Borneo. Bulletin of the British Museum (Natural History), Geology Series, 2,
257±280.KARCHE, J. P. 1972. Contribution aÁ l'eÂtude geÂologique de la Montagne d'Ambre. Unpublished Diploma thesis,
University of BesancËon.KEATS, D. W. and CHAMBERLAIN, Y. M. 1994. Two melobesioid coralline algae (Rhodophyta, Corallinales), Mesophyllum
erubescens (Foslie) Lemoine and Mesophyllum funafutiense (Foslie) Verheij from Sodwana Bay, South Africa.South African Journal of Botany, 60, 175±190.
ÐÐ ÐÐ 1997. The non-geniculate coralline algae Synarthrophyton eckloniae (Foslie) comb. nov. and S.magellanicum (Foslie) comb. nov. (Rhodophyta) in South Africa including a comparison with relevant types.European Journal of Phycology, 32, 55±79.
ÐÐ and MANEVELDT, G. 1997a. Two new melobesioid algae (Corallinales, Rhodophyta), Synarthrophyton robbenensesp. nov. and S. munimentum sp. nov., in South Africa and Namibia. Phycologia, 36, 447±467.
ÐÐ ÐÐ 1997b. First report of the melobesioid alga (Corallinales, Rhodophyta) Mesophyllum incisum (Foslie)Adey in South Africa. South African Journal of Botany, 63, 201±209.
LAMOUROUX, J. V. F. 1812. Exstrait d'une meÂmoire sur la classi®cation des polypiers coralligeÁnes non entieÂrementpierreux. Nouvelle Bulletin des Sciences de la Societe de Philomatematique, 3, 181±188.
LEMOINE, M. P. 1927. Sur quelques algues calcaires du nummulitique de la Haute Savoie. Bulletin du Museumd'Histoire Naturelle de Paris, 6, 545±551.
ÐÐ 1939. Algues calcaires fossiles de l'AlgeÂrie. MateÂriaux pour la Carte GeÂologique de l'AlgeÂrie. 1re SeÂriePaleÂontologie, 9, 1±128.
LEMOINE, M. 1970. Les algues ¯orideÂes calcaires du CreÂtace du Sud de la France. Archives du Museum d'HistoireNaturelle, SeÂrie 7, 10 (for 1968), 129±240.
422 P A L A E O N T O L O G Y , V O L U M E 4 3

ÐÐ 1977. EÂ tude d'une collection d'Algues CorallinaceÂes de la reÂgion de Skopje (Yougoslavie). Revue deMicropaleÂontologie, 20, 10±43.
LINNAEUS, C. 1758. Systema Naturae. Salvii, Stockholm, 10th edition, vol. 1, ii� 824 pp.MAGATHAN, E. R. 1985. Devonian reef-associated articulate red algae from Western Canada. 170±178. In TOOMEY, D. F.
and NITECKI, M. H. (eds). Palaeoalgology: contemporary research and applications. Springer-Verlag, Berlin, 376 pp.MANZA, A. V. 1937. The genera of the articulated Corallines. Proceedings of the National Academy of Sciences, 23,
44±48.MASON, L. R. 1953. The crustaceous coralline algae of the Paci®c Coast of the United States, Canada and Alaska.
University of California Publications in Botany, 26, 313±390.MASTRORILLI, V. I. 1968a. Caratteri morfologici e strutturali di un esemplare fertile di Subterraniphyllum Elliott
rinvenuto nell'Oligocene di Ponzone (Piemonte). Rivista Italiana di Paleontologia e Stratigra®a, 74, 1275±1283.ÐÐ 1968b. Nuovo contributo allo studio delle Corallinacee dell'Oligocene Ligure-Piemontese: i reperti della
tavoletta Ponzone. Atti dell'Istituto di Geologia, UniversitaÁ di Genova, 5 (2), 153±406.MAY, D. I. and WOELKERLING, W. J. 1988. Studies on the genus Synarthrophyton (Corallinaceae, Rhodophyta) and its type
species, S. patena (J. D. Hooker et W. H. Harvey) Townsend. Phycologia, 26, 50±71.MIETTO, P. 1988. Aspetti geologici dei Monti Berici. 13±23. In: I Colli Berici. Natura e civiltaÁ . Edizioni Signum,
Padova, 251 pp.MORLOT, A. V. 1853. Einige Bemerkungen uÈber geologische VerhaÈltnisse der Untersteiermark- 2. Bericht der
Monatliche Vereinigung der Steiermark, 1±25.NEBELSICK, J. and BASSI, D. 1999. Facies and fabric of coralline algal dominated shelf carbonates from the Lower
Oligocene of Slovenia. The Lyell Meeting 1999, `Organism-environment feedbacks in carbonate platforms andreefs', London, Abstracts, 20±21.
PFENDER, J. 1924. Sur une formation quaternaire marine des coÃtes de Provence. Bulletin de la Societe GeÂologique deFrance, 4, 1±193.
PHILIPPI, R. 1837. Beweis dass die Nulliporen P¯anzen sind. Archiv fuÈr Naturgeschichte, 3, 387±393.POIGNANT, A. F. 1977. The Mesozoic red algae: a general survey. 177±189. In FLUÈ GEL, E. (ed.). Fossil algae: recent
results and developments. Springer-Verlag, Berlin, 375 pp.ÐÐ 1979a. Les CorallinaceÂes meÂsozoõÈques et ceÂnozoiques: hypotheÁses phylogeÂneÂtiques. Bulletin du Centre de
Recherche, Exploration-Production Elf Aquitaine, 3, 753±755.ÐÐ 1979b. DeÂtermination geÂneÂrique des CorallinaceÂes MeÂsozoõÈques et CeÂnozoõÈques. Bulletin du Centre de
Recherche, Exploration-Production Elf Aquitaine, 3, 757±765.ÐÐ 1985. PreÂsentation simpli®eÂe d'une nouvelle classi®cation geÂneÂrique des RhodophyceÂes meÂsozoõÈques et
ceÂnozoõÈques. Informatisation. Bulletin de la Societe GeÂologique de France, (8) 1, 603±605.RAINERI, R. 1920. Alghe fossili corallinacee della Libia. Atti della SocietaÁ Italiana di Scienze Naturali, 59, 137±148.RASSER, M. and PILLER, W. E. 1997. Depth distribution of calcareous encrusting associations in the northern Red Sea
(Safaga, Egypt) and their geological implications. Proceedings of the Eighth International Coral Reef Symposium,1, 743±748.
ÐÐ ÐÐ 1999. Application of neontological taxonomic concepts to Late Eocene coralline algae (Rhodophyta) of theAustrian Molasse Zone. Journal of Micropalaeontology, 18, 67±80.
REUSS, A. 1851. VorlaÈu®ge Faunenliste der Oberburger Schichten. Jahrbuch der Geologische Reichsanstalt, 1, 1±162.ÐÐ 1864. Die fossilen Foraminifera, Anthozoen, und Bryozoen von Oberburg in der Steiermark. Kaiserliche
Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Classe Denkschriften, 23, 1±36.ROLLE, F. 1857. Geologische Untersuchungen in der Gegend zwischen Weitenstein, Windisch-GraÈtz, Cilli und
Oberburg in der Untersteiermark. Jahrbuch der Geologische Reichsanstalt, 3, 403±465.SILVA, P. C. and JOHANSEN, H. W. 1986. A reappraisal of the order Corallinales (Rhodophyceae). British Phycological
Journal, 21, 245±254.STUR, D. 1871. Geologie der Steiermark. Geognostischer-montanischer Verein der Steiermark, Graz, 654 pp.TELLER, F. 1885. OligozaÈnbildungen im Fiestritzthal bei Stein in Krain. Verhandlungen der Geologische Reichsanstalt,
1885, 193±200.ÐÐ 1896. ErlaÈuterungen zur geologischen Karte der im Reichsrathe vertretenen KoÈnigreiche und LaÈnder der
OÈ sterreich-ungarischen Monarchie, SW-Gruppe. Eisenkappel und Kanker. Nr. 83, 1:75,000, Vienna, 142 pp.ÐÐ 1898. ErlaÈuterungen zur geologischen Karte der im Reichsrathe vertretenen KoÈnigreiche und LaÈnder der
OÈ sterreich-ungarischen Monarchie, SW-Gruppe. Eisenkappel und Kanker. Nr. 84, 1:75,000, Vienna, 170 pp.THOMAS, A. N. 1950. The Asmari Limestone of south-west Iran. International Geological Congress, Report of the 18th
Session, UK 1948, 6, 35±44.ÐÐ 1952. Facies variations in the Asmari Limestone. International Geological Congress, Report of the 18th Session,
UK 1948, 10, 74±82.
B A S S I E T A L . : T E R T I A R Y C O R A L L I N E A L G A E 423

UNGARO, S. 1969. EÂ tude micropaleÂontologique et stratigraphique de l'EÂ oceÁne supeÂrieur (Priabonien) de Mossano (ColliBerici). MeÂmoires du Bureau de Recherches GeÂologiques et MinieÁres, 69, 267±280.
ÐÐ 1978. L'Oligocene dei Colli Berici. Rivista Italiana di Paleontologia, 84, 199±278.VAN BELLEN, R. C. 1956. The stratigraphy of the ``Main Limestone'' of the Kirkuk, Bai Hassan, and Qarah Chauq Dagh
structures in north Iraq. Journal of the Institute of Petroleum, 42, 233±263.WATERS, H. 1874. Remarks on the fossils from Oberburg, Styria. Quarterly Journal of the Geological Society, London,
30, 339±341.WEBER-VAN BOSSE, A. 1904. Corallineae verae of the Malay Archipelago. iii, 78-110. In WEBER-VAN BOSSE, A. and
FOSLIE, M. 1904. The Corallinaceae of the Siboga expedition. Siboge Expeditie, 61, 1±110 pp.WILKS, K. M. and WOELKERLING, W. J. 1994. An account of southern Australian species of Phymatolithon (Corallinaceae,
Rhodophyta) with comments on Leptophytum. Australian Systematic Botany, 7, 183±223.WOELKERLING, W. J. 1988. The coralline red algae: an analysis of the genera and subfamilies of nongeniculate
Corallinaceae. Oxford University Press, London and Oxford, 268 pp.ÐÐ 1996. Subfamily Lithophylloideae. 214±237. In WOMERSLEY, H. B. S. (ed.). The marine benthic ¯ora of southern
Australia. Part IIIB. Gracilariales, Rhodymeniales, Corallinales and Bonnemaisoniales. Australian BiologicalResources Study, Canberra, 392 pp.
ÐÐ and CAMPBELL, S. J. 1992. An account of southern Australian species of Lithophyllum (Corallinaceae,Rhodophyta). Bulletin of the British Museum (Natural History), Botany Series, 22, 1±107.
ÐÐ and FOSTER, M. S. 1989. A systematic and ecographic account of Synarthrophyton schielianum sp. nov.(Corallinaceae, Rhodophyta) from the Chatham Islands. Phycologia, 28, 39±60.
ÐÐ and HARVEY, A. 1992. Mesophyllum incisum (Corallinaceae, Rhodophyta) in southern Australia: implications forgeneric and speci®c delimitation in the Melobesioideae. British Phycological Journal, 27, 381±399.
ÐÐ ÐÐ 1993. An account of southern Australian species of Mesophyllum (Corallinaceae, Rhodophyta). AustralianSystematic Botany, 6, 571±637.
ÐÐ and IRVINE, L. M. 1986. The neotypi®cation and status of Mesophyllum (Corallinaceae, Rhodophyta). Phycologia,25, 379±396.
ÐÐ ÐÐ and HARVEY, A. S. 1993. Growth-forms in non-geniculate coralline red algae (Corallinales, Rhodophyta).Australian Systematic Botany, 6, 277±293.
ÐÐ and LAMY, D. 1998. Non-geniculate coralline red algae and the Paris MuseÂum: systematics and scienti®c history.Publications Scienti®que du MuseÂum/ADAC, Paris, viii� 767 pp.
WOMERSLEY, H. B. S. 1996. The marine benthic ¯ora of southern Australia. Part IIIB. Gracilariales, Rhodymeniales,Corallinales and Bonnemaisoniales. Australian Biological Resources Study, Canberra, 392 pp.
ÐÐ and JOHANSEN, H. W. 1996a. Subfamily Amphiroideae. 283±288. In WOMERSLEY, H. B. S. (ed.). The marine benthic¯ora of southern Australia. Part IIIB. Gracilariales, Rhodymeniales, Corallinales and Bonnemaisoniales.Australian Biological Resources Study, Canberra, 392 pp.
ÐÐ ÐÐ 1996b. Subfamily Corallinoideae. 288±317. In WOMERSLEY, H. B. S. (ed.). The marine benthic ¯ora ofsouthern Australia. Part IIIB. Gracilariales, Rhodymeniales, Corallinales and Bonnemaisoniales. Canberra,Australian Biological Resources Study, 392 pp.
ÐÐ ÐÐ 1996c. Subfamily Metagoniolithoideae. 317±323. In WOMERSLEY, H. B. S. (ed.). The marine benthic ¯ora ofsouthern Australia. Part IIIB. Gracilariales, Rhodymeniales, Corallinales and Bonnemaisoniales. AustralianBiological Resources Study, Canberra, 392 pp.
DAVIDE BASSI
Dipartimento di Scienze Geologiche e PaleontologicheUniversitaÁ di Ferrara
Corso Ercole I d'Este 32I-44100 Ferrara, Italy
e-mail [email protected]
WILLIAM J. WOELKERLING
School of BotanyLa Trobe University
Bundoora,Victoria 3083, Australia
e-mail [email protected]
424 P A L A E O N T O L O G Y , V O L U M E 4 3

JAMES H. NEBELSICK
Institute and Museum of Geology and PalaeontologyUniversity of TuÈbingen
Sigwartstrasse 10D-72076 TuÈbingen, Germany
e-mail [email protected] received 14 June 1999Revised typescript received 6 December 1999
B A S S I E T A L . : T E R T I A R Y C O R A L L I N E A L G A E 425





























