Embed Size (px)
Citation preview

© The Author 2012. Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected]
Tectorigenin sensitizes paclitaxel-resistant human ovarian cancer cells through downregulation of the Akt and NFκB pathway
Yeong-In Yang1,2, Kyung-Tae Lee3, Hee-Juhn Park4, Tae Jin Kim5, Youn Seok Choi6, Ie-Ming Shih7 and Jung-Hye Choi1,2,*1Department of Life and Nanopharmaceutical Science, Kyung Hee University, Seoul 130-701, Republic of Korea, 2Department of Oriental Pharmacy, College of Pharmacy, Kyung Hee University, Seoul 130-701, Republic of Korea, 3Department of Biochemistry, College of Pharmacy, Kyung Hee University, Seoul 130-701, Republic of Korea, 4Department of Botanical Resources, Sangji University, Wonju 220-702, Republic of Korea, 5Cheil General Hospital, and Women’s Healthcare Center, Kwandong University College of Medicine, Seoul 100-380, Republic of Korea, 6Department of Obstetrics and Gynecology, School of Medicine, Catholic University of Daegu, Daegu 712-702, Republic of Korea and 7Department of Pathology and Oncology, Johns Hopkins Medical Institutions, Baltimore, MD 21231, USA
*To whom correspondence should be addressed. Tel: 82-2-961-2172; Fax: 82-2-962-0860; Email: [email protected]
Paclitaxel (Taxol) is currently used as the front-line chemothera-peutic agent for several cancers including ovarian carcinoma; how-ever, the drug frequently induces drug resistance through multiple mechanisms. The new strategy of using natural compounds in com-bination therapies is highly attractive because those compounds may enhance the efficacy of chemotherapy. In this study, we found that tectorigenin, an isoflavonoid isolated from flower of Pueraria thunbergiana, enhanced the growth-inhibitory effect of paclitaxel in paclitaxel-resistant ovarian cancer cells (MPSC1TR, A2780TR and SKOV3TR) as well as their naive counterparts. The combination of tectorigenin with paclitaxel resulted in a synergistic apoptosis com-pared with either agent alone through activation of caspases-3, -8 and -9. Treatment with tectorigenin inhibited the nuclear translo-cation of NFκB and the expression of NFκB-dependent genes such as FLIP, XIAP, Bcl-2, Bcl-xL and COX-2, which are known to be associated with chemoresistance. In addition, the tectorigenin–paclitaxel combination inhibited the phosphorylation of IκB and IKK and the activation of Akt in paclitaxel-resistant cancer cells. Moreover, tectorigenin–paclitaxel-induced cell growth inhibition was enhanced by pretreatment with the Akt inhibitor LY294002 or overexpression of the dominant negative Akt (Akt-DN), but reduced by overexpression of constitutively activated Akt (Akt-Myr). Furthermore, we found that Akt-Myr, at least in part, reversed tectorigenin–paclitaxel-induced nuclear translocation of NFκB and the phosphorylation of IκB and IKK. These data suggest that tectorigenin could sensitize paclitaxel-resistant human ovarian cancer cells through inactivation of the Akt/IKK/IκB/NFκB sign-aling pathway, and promise a new intervention to chemosensitize paclitaxel-induced cytotoxicity in ovarian cancer.
Introduction
Ovarian cancer is the sixth most common cancer and the fifth leading cause of cancer-related deaths among women in developed countries. It affects ~204 000 women a year worldwide and is responsible for about 125 000 deaths (1). Patients in most cases present in stage III or
IV with a 5-year survival of around 28 and 16%, respectively, whereas the 5-year survival rate is up to 80% for patients in stage I. Standard management for advanced ovarian cancer is cytoreductive surgery followed by paclitaxel-/platinum-based chemotherapy (2). Although ovarian cancer is considered a chemosensitive neoplasm with 80% of the tumors responding to conventional treatment in the early stage, patients with advanced or more aggressive tumors often experience chemoresistance and recurrence (3). Therefore, treatment failure is often attributed to primary or acquired resistance to chemotherapeutic agents, which remains a clinically formidable problem in managing most cancer patients (4). In this regard, overcoming chemoresistance is urgent for the treatment and survival of the patients.
Drug resistance is described as a multifactorial phenomenon, involv-ing the expression of defense factors and/or detoxification mechanisms, and alterations in drug–target interactions (e.g. target accessibility, target sensitivity, persistence of DNA cleavage and genomic localization of DNA damage) and the cellular response to specific cytotoxic lesions (5). However, recent studies on the chemoresistance of ovarian cancer sug-gest that a decreased susceptibility of the cancer to apoptosis is strongly associated with drug resistance (6). Bcl-2 and Bcl-xL are antiapoptotic Bcl-2 family proteins that prevent the formation of the mitochondrial apoptosis-induced channel (MAC), through which cytochrome c is released to the cytosol, triggering the intrinsic mitochondrial apoptosis pathway. The FLICE inhibitory protein (FLIP) and the X-linked inhibi-tor of apoptosis protein (XIAP) inhibit apoptosis by modulating death-signaling pathways at the death receptor/caspase-8 and the cytochrome c/apaf-1/caspase-9 pathway, respectively. Upregulation of the antiapop-totic protein Bcl-2 and/or downregulation of proapoptotic Bax have been correlated with increased chemoresistance (7–9). In addition, inhibitors of caspases such as FLIP and XIAP were implicated as determinants of the chemosensitivity of ovarian cancer (10–12). Thus, increasing the susceptibility of the cancer to apoptosis is a potential strategy to over-come drug resistance in ovarian cancer cells.
Phytoestrogens, a family of plant-derived polyphenolic compounds with a similar structure to endogenous estrogens, have been demonstrated to have various biological activities via complex mechanisms (13). Recently, phytoestrogens have been intensely investigated in the prevention and treatment of several hormone responsive cancers, including ovarian, breast and prostate cancer (14). In comparison with extensive literature evaluating the role of phytoestrogen on breast cancers, its role on ovarian cancer were less considered and investigated (15–18). Recent studies that have examined the link between phytoestrogen consumption and epithelial ovarian cancer risk tended to suggest an inverse association (19). In vitro studies using ovarian cancer cell lines, phytoestrogens showed anticancer activities by inducing apoptosis, autophagocytosis and cell cycle arrest (20–23). In addition, a phytoestrogen genistein and its synthetic derivative sensitized chemoresistant ovarian cancer cells to conventional ovarian cancer drugs such as cisplatin (24–26). However, the exact molecular mechanisms of action of the phytoestrogens in ovarian cancer have not been fully elucidated. Tectorigenin, a type of O-methylated isoflavone, has been demonstrated to possess estrogenic (27), antiinflammatory (28,29) and antitumor activities (30,31). A study to examine the estrogenic and antiproliferative activities of several herbs commonly used in Korean traditional medicine showed that Pueraria flower extract with tectorigenin as a major bioactive isoflavonoid had the highest estrogenic relative potency and potent cytotoxic effect against ovarian cancer cells (27,32). Tectorigenin inhibited the proliferation of hormone responsive prostate cancer cells, causing G1 arrest the induction of p21 (WAF1) or p27kip1 protein expression, and downregulation of PDEF, PSA, hTERT, and IGF-1 receptor gene expression (33,31). On the other hand, tectorigenin along with genistein showed estrogenic activities and regulated the
Abbreviations: CI, combination index; DMSO, dimethyl sulfoxide; ER, estrogen receptor; FLIP, FLICE inhibitory protein; IKK, IκB kinase; MAC, mitochondrial apoptosis-induced channel; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide; NFκB, nuclear factor-κB; PBS, phosphate buffered saline; PI, propidium iodide; PI3K, phosphoinositide 3-kinase; XIAP, X-linked inhibitor of apoptosis protein.
Carcinogenesis vol.33 no.12 pp.2488–2498, 2012doi:10.1093/carcin/bgs302Advance Access publication October 2, 2012
2488
Dow
nloaded from https://academ
ic.oup.com/carcin/article/33/12/2488/2464293 by guest on 06 January 2022

proliferation of MCF-7 and T47D human breast cancer cells (33). These observations suggested that the phytoestrogen tectorigenin may have some effects in hormone responsive ovarian cancers.
In this study, we investigated the effect of tectorigenin alone and in combination with paclitaxel on the growth of ovarian cancer cells. We found that tectorigenin alone does not induce potent cell death, but significantly sensitize human ovarian cancer cells to paclitaxel-induced cell death. We further investigated the cellular mechanisms underlying the sensitizing effect of tectorigenin on paclitaxel-induced cell death.
Materials and methods
MaterialsThe tectorigenin used in this study was isolated from the flower of Pueraria thunbergiana BENTH., as reported previously (34). Paclitaxel was pur-chased from A.G. Scientific, Inc (San Diego, CA). The RPMI 1640 medium, fetal bovine serum, penicillin and streptomycin were obtained from Life Technologies Inc (Grand Island, NY). The 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT), dimethyl sulfoxide (DMSO), RNase A, leupeptin, aprotinin, phenylmethylsulfonylfluoride, Triton X-100 and propidium iodide (PI) were purchased from Sigma–Aldrich Co. (St Louis, MO). The antibodies for anti-Bcl-2, anti-Bcl-xL, anti-FLIP, anti-COX-2, anti-Akt and anti-β-actin were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). The antibodies for the anti-XIAP, anti-caspase-3 and anti-caspase-8 were purchased from BD Biosciences, Pharmingen (San Diego, CA). The antibodies for anti-caspase-9 and anti-phospho-Akt were purchased from Cell Signaling Technology (Beverly, MA). The pcDNA3-Akt-Myr and pcDNA3-Akt-DN were obtained from Addgene (Cambridge, MA). The z-VAD-fmk, z-DEVD-fmk, PI3K/Akt inhibitor LY294002, IκB inhibitor Bay11-7082, IKK inhibitor BMS-34551 and proteasome inhibitor MG132 were obtained from Calbiochem (San Diego, CA).
Cell culturesOvarian cancer cells including SKOV3 and A2780 cells were purchased from American Type Culture Collection, and MPSC1 was previously established by Dr I.-M. Shih [Johns Hopkins School of Medicine, MD (36)]. The parental ovarian cancer cell lines (MPSC1, A2780 and SKOV3) (36) and the paclitaxel-resistant ovarian cancer cells (MPSC1TR, SKOV3TR and A2780TR) (37) were generated by the negative selection of cells in the presence of paclitaxel. The IOSE cell lines were generated by transfecting ovarian surface epithelial cells with SV40-T antigen and generously provided by Dr N. Auersperg (University of British Columbia, Vancouver, British Columbia, Canada) and Dr A. Godwin (Fox Chase Cancer Center, Philadelphia, PA) (37). The cells were cultured in RPMI 1640 supplemented with 5% fetal bovine serum, penicillin (100 U/ml) and streptomycin sulfate (100 µg/ml) (Life Technologies, Grand Island, NY). Tectorigenin and paclitaxel were dissolved in DMSO. Further dilutions were performed in cell culture media. DMSO was used as a vehicle control throughout the study.
MTT assayCytotoxicity was assessed using the MTT assay. Briefly, the cells (5 × 104) were seeded in each well containing 50 μl of RPMI medium in a 96-well plate. After 24 h, various concentrations of paclitaxel and/or tectorigenin were added. After 48 h, 50 μl of MTT (5 mg/ml stock solution) was added, and the plates were incubated for an additional 4 h. The medium was discarded, and the formazan blue that formed in the cells was dissolved in 50 μl of DMSO. The optical density was measured at 540 nm using a microplate spectropho-tometer (SpectraMax; Molecular Devices, Sunnyvale, CA).
Propidium iodide staining for cell cycle analysisOn the day of collection, the cells were harvested and washed twice with ice-cold phosphate buffered saline (PBS). The cells were fixed and permeabilized with 70% ice-cold ethanol at 4°C for 1 h. The cells were washed once with PBS and resuspended in a staining solution containing PI (50 μl/ml) and RNase A (250 μg/ml). The cell suspensions were incubated for 30 min at room tem-perature followed by fluorescence-activated cell sorting cater-plus flow cytom-etry (Becton Dickinson Co., Germany) using 10,000 cells per each group.
Annexin V and PI double staining for apoptosis analysisDuring apoptosis, the exposure of phosphatidylserine on the exterior surface of the plasma membrane can be detected by the binding of fluoresceinated annexin V (annexin V–fluorescein isothiocyanate). This assay is combined with an analysis of the exclusion of the plasma membrane integrity probe PI. For annexin V and PI double staining, cells were suspended in 100 μl of
binding buffer (10 mM N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid/NaOH, 140 mM NaCl, 2.5 mM CaCl2, pH 7.4) and stained with 5 μl of fluo-rescein isothiocyanate-conjugated annexin V and 5 μl of PI (50 μg/ml). The mixture was incubated for 15 min at room temperature in the dark, followed by fluorescence-activated cell sorting cater-plus flow cytometry analysis.
Immunoblot assayThe cells were washed with ice-cold PBS and extracted in protein lysis buffer (Intron, South Korea) and the protein concentration was determined by the Bradford assay. The samples of cell lysate were mixed with an equal volume of 5× sodium dodecyl sulfate sample buffer, boiled for 4 min and then separated on 10–12% sodium dodecyl sulfate–polyacrylamide gel elec-trophoresis gels. After electrophoresis, the proteins were transferred to poly-vinylidene difluoride membranes. The membrane was immunoblotted using specific primary antibodies at 4°C overnight. After washing, the signals were detected with horseradish peroxidase-conjugated secondary antibody for 1 h, and visualized using the ECL chemiluminescent system (Amersham Pharmacia Biotech, Oakville, Ontario, Canada). Following three washes in Tris-buffered saline-T, immunopositive bands were visualized by enhanced chemiluminescence and exposed to Image Quant LAS-4000 (Fujifilm Life Science, Japan).
Nuclear extractionThe cells were washed twice with ice-cold PBS, then scraped onto 1 ml of cold PBS and pelleted by centrifugation. The cell pellets were resuspended in hypo-tonic buffer [10 mM N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid (pH 7.9), 1.5 mM MgCl2, 10 mM KCl, 0.2 mM phenylmethylsulfonylfluoride, 0.5 mM dithiothreitol and 10 μg/ml aprotinin] and incubated on ice for 15 min. Cells were then lysed by adding 0.1% Nonidet P-40 and vortexed vigorously for 10 s. Nuclei were pelleted by centrifugation at 12,000g for 1 min at 4°C and resuspended in high salt buffer [20 mM N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid (pH 7.9), 25% glycerol, 400 mM KCl, 1.5 mM MgCl2, 0.2 mM ethylenediaminetetraacetic acid, 0.5 mM dithiothreitol, 1 mM NaF, 1 mM sodium orthovanadate].
Transient transfectionpcDNA3-Akt-Myr, pcDNA3-Akt-DN or empty vector were transfected by using polyethylenimine (Sigma–Aldrich Co.). Cells were plated in 6-well culture dishes and allowed to attach and grow 24 h before transfection. Each transfection mixture was prepared by mixing up the DNA (1 μg) and poly-ethylenimine (3 μl) in serum-free Opti-modified Eagle's medium (Invitrogen) and incubating for 15 min at room temperature. The transfection mixture was slowly added to the cells, which were allowed to recover for an additional 24 h before experimental treatments. To evaluate transfection efficiency, cotransfec-tion of the relevant plasmid and the GFP plasmid was performed. The fraction of transfected cells was evaluated using a fluorescent microscope. This trans-fection method routinely yielded 60–70% efficiency.
Statistical analysisStatistical analysis was carried out using Student’s t-test or one-way analysis of variance, and the level of significance was set at a P value of <0.05. Data are expressed as the mean ± SD.
Results
Tectorigenin enhances the sensitivity of ovarian cancer cells to paclitaxelTectorigenin has been demonstrated to inhibit cell growth in pros-tate cancer and leukemia cells (30,38). However, there are no reports on the effect of tectorigenin in ovarian cancer cells. The cytotoxic-ity of tectorigenin were measured in three pairs of human ovarian cancer cell lines, each pair consisting of a parental line and a stably paclitaxel-resistant subline, and one immortalized ovarian surface epithelial cell line (IOSE-80PC) (Supplementary Table 1, available at Carcinogenesis Online). The growth inhibitory effect of paclitaxel was markedly reduced in all three paclitaxel-resistant ovarian cancer cells compared with their parental cells. When compared with pacli-taxel, the direct cytotoxicity of tectorigenin in the seven cell lines tested (IC50 > 73 μM) was much less than that of paclitaxel. However, the growth inhibitory effect of tectorigenin was more significant in MPSC1TR, A2780TR and SKOV3TR (IC50 = 73, 78 and 89 μM, respec-tively) than in their parental cells (IC50 = 123, 108 and >200 μM, respectively). Since tectorigenin alone failed to show a potent cell growth inhibitory effect in ovarian cancer cells (Supplementary
Tectorigenin as a chemosensitizer for ovarian cancer
2489
Dow
nloaded from https://academ
ic.oup.com/carcin/article/33/12/2488/2464293 by guest on 06 January 2022
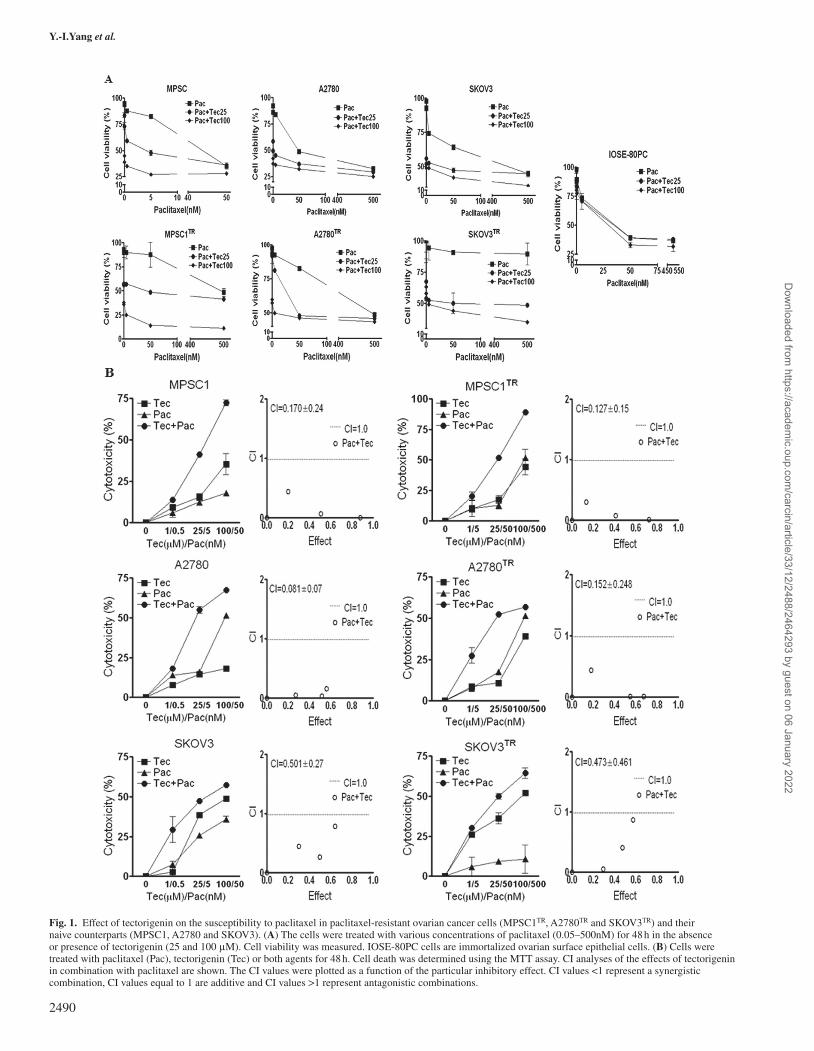
Fig. 1. Effect of tectorigenin on the susceptibility to paclitaxel in paclitaxel-resistant ovarian cancer cells (MPSC1TR, A2780TR and SKOV3TR) and their naive counterparts (MPSC1, A2780 and SKOV3). (A) The cells were treated with various concentrations of paclitaxel (0.05–500nM) for 48 h in the absence or presence of tectorigenin (25 and 100 μM). Cell viability was measured. IOSE-80PC cells are immortalized ovarian surface epithelial cells. (B) Cells were treated with paclitaxel (Pac), tectorigenin (Tec) or both agents for 48 h. Cell death was determined using the MTT assay. CI analyses of the effects of tectorigenin in combination with paclitaxel are shown. The CI values were plotted as a function of the particular inhibitory effect. CI values <1 represent a synergistic combination, CI values equal to 1 are additive and CI values >1 represent antagonistic combinations.
Y.-I.Yang et al.
2490
Dow
nloaded from https://academ
ic.oup.com/carcin/article/33/12/2488/2464293 by guest on 06 January 2022
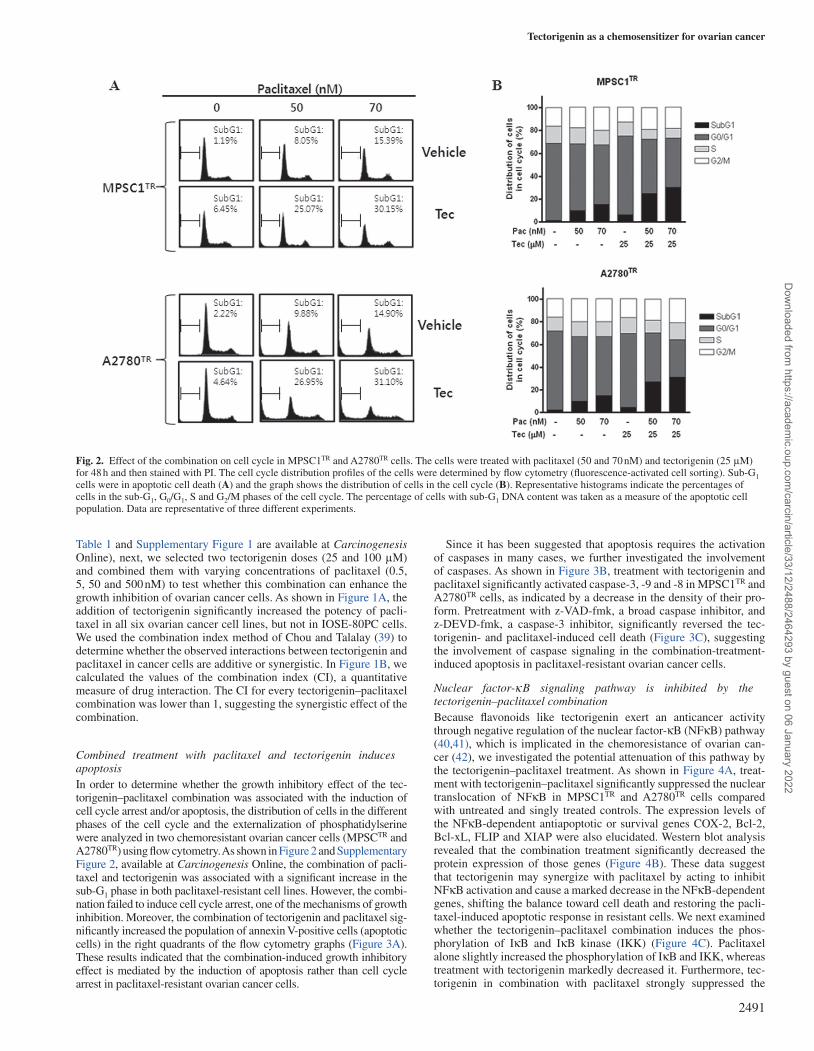
Table 1 and Supplementary Figure 1 are available at Carcinogenesis Online), next, we selected two tectorigenin doses (25 and 100 μM) and combined them with varying concentrations of paclitaxel (0.5, 5, 50 and 500 nM) to test whether this combination can enhance the growth inhibition of ovarian cancer cells. As shown in Figure 1A, the addition of tectorigenin significantly increased the potency of pacli-taxel in all six ovarian cancer cell lines, but not in IOSE-80PC cells. We used the combination index method of Chou and Talalay (39) to determine whether the observed interactions between tectorigenin and paclitaxel in cancer cells are additive or synergistic. In Figure 1B, we calculated the values of the combination index (CI), a quantitative measure of drug interaction. The CI for every tectorigenin–paclitaxel combination was lower than 1, suggesting the synergistic effect of the combination.
Combined treatment with paclitaxel and tectorigenin induces apoptosisIn order to determine whether the growth inhibitory effect of the tec-torigenin–paclitaxel combination was associated with the induction of cell cycle arrest and/or apoptosis, the distribution of cells in the different phases of the cell cycle and the externalization of phosphatidylserine were analyzed in two chemoresistant ovarian cancer cells (MPSCTR and A2780TR) using flow cytometry. As shown in Figure 2 and Supplementary Figure 2, available at Carcinogenesis Online, the combination of pacli-taxel and tectorigenin was associated with a significant increase in the sub-G1 phase in both paclitaxel-resistant cell lines. However, the combi-nation failed to induce cell cycle arrest, one of the mechanisms of growth inhibition. Moreover, the combination of tectorigenin and paclitaxel sig-nificantly increased the population of annexin V-positive cells (apoptotic cells) in the right quadrants of the flow cytometry graphs (Figure 3A). These results indicated that the combination-induced growth inhibitory effect is mediated by the induction of apoptosis rather than cell cycle arrest in paclitaxel-resistant ovarian cancer cells.
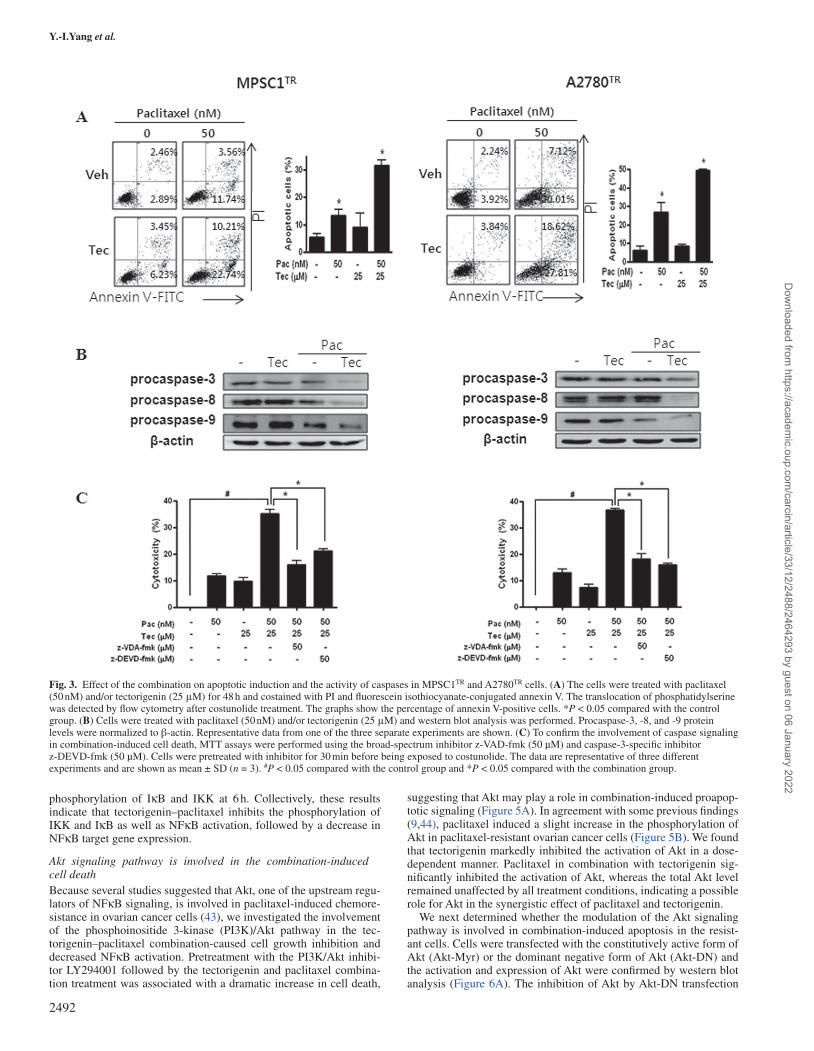
Since it has been suggested that apoptosis requires the activation of caspases in many cases, we further investigated the involvement of caspases. As shown in Figure 3B, treatment with tectorigenin and paclitaxel significantly activated caspase-3, -9 and -8 in MPSC1TR and A2780TR cells, as indicated by a decrease in the density of their pro-form. Pretreatment with z-VAD-fmk, a broad caspase inhibitor, and z-DEVD-fmk, a caspase-3 inhibitor, significantly reversed the tec-torigenin- and paclitaxel-induced cell death (Figure 3C), suggesting the involvement of caspase signaling in the combination-treatment-induced apoptosis in paclitaxel-resistant ovarian cancer cells.
Nuclear factor-κB signaling pathway is inhibited by the tectorigenin–paclitaxel combinationBecause flavonoids like tectorigenin exert an anticancer activity through negative regulation of the nuclear factor-κB (NFκB) pathway (40,41), which is implicated in the chemoresistance of ovarian can-cer (42), we investigated the potential attenuation of this pathway by the tectorigenin–paclitaxel treatment. As shown in Figure 4A, treat-ment with tectorigenin–paclitaxel significantly suppressed the nuclear translocation of NFκB in MPSC1TR and A2780TR cells compared with untreated and singly treated controls. The expression levels of the NFκB-dependent antiapoptotic or survival genes COX-2, Bcl-2, Bcl-xL, FLIP and XIAP were also elucidated. Western blot analysis revealed that the combination treatment significantly decreased the protein expression of those genes (Figure 4B). These data suggest that tectorigenin may synergize with paclitaxel by acting to inhibit NFκB activation and cause a marked decrease in the NFκB-dependent genes, shifting the balance toward cell death and restoring the pacli-taxel-induced apoptotic response in resistant cells. We next examined whether the tectorigenin–paclitaxel combination induces the phos-phorylation of IκB and IκB kinase (IKK) (Figure 4C). Paclitaxel alone slightly increased the phosphorylation of IκB and IKK, whereas treatment with tectorigenin markedly decreased it. Furthermore, tec-torigenin in combination with paclitaxel strongly suppressed the
Fig. 2. Effect of the combination on cell cycle in MPSC1TR and A2780TR cells. The cells were treated with paclitaxel (50 and 70 nM) and tectorigenin (25 μM) for 48 h and then stained with PI. The cell cycle distribution profiles of the cells were determined by flow cytometry (fluorescence-activated cell sorting). Sub-G1 cells were in apoptotic cell death (A) and the graph shows the distribution of cells in the cell cycle (B). Representative histograms indicate the percentages of cells in the sub-G1, G0/G1, S and G2/M phases of the cell cycle. The percentage of cells with sub-G1 DNA content was taken as a measure of the apoptotic cell population. Data are representative of three different experiments.
Tectorigenin as a chemosensitizer for ovarian cancer
2491
Dow
nloaded from https://academ
ic.oup.com/carcin/article/33/12/2488/2464293 by guest on 06 January 2022
phosphorylation of IκB and IKK at 6 h. Collectively, these results indicate that tectorigenin–paclitaxel inhibits the phosphorylation of IKK and IκB as well as NFκB activation, followed by a decrease in NFκB target gene expression.
Akt signaling pathway is involved in the combination-induced cell deathBecause several studies suggested that Akt, one of the upstream regu-lators of NFκB signaling, is involved in paclitaxel-induced chemore-sistance in ovarian cancer cells (43), we investigated the involvement of the phosphoinositide 3-kinase (PI3K)/Akt pathway in the tec-torigenin–paclitaxel combination-caused cell growth inhibition and decreased NFκB activation. Pretreatment with the PI3K/Akt inhibi-tor LY294001 followed by the tectorigenin and paclitaxel combina-tion treatment was associated with a dramatic increase in cell death,
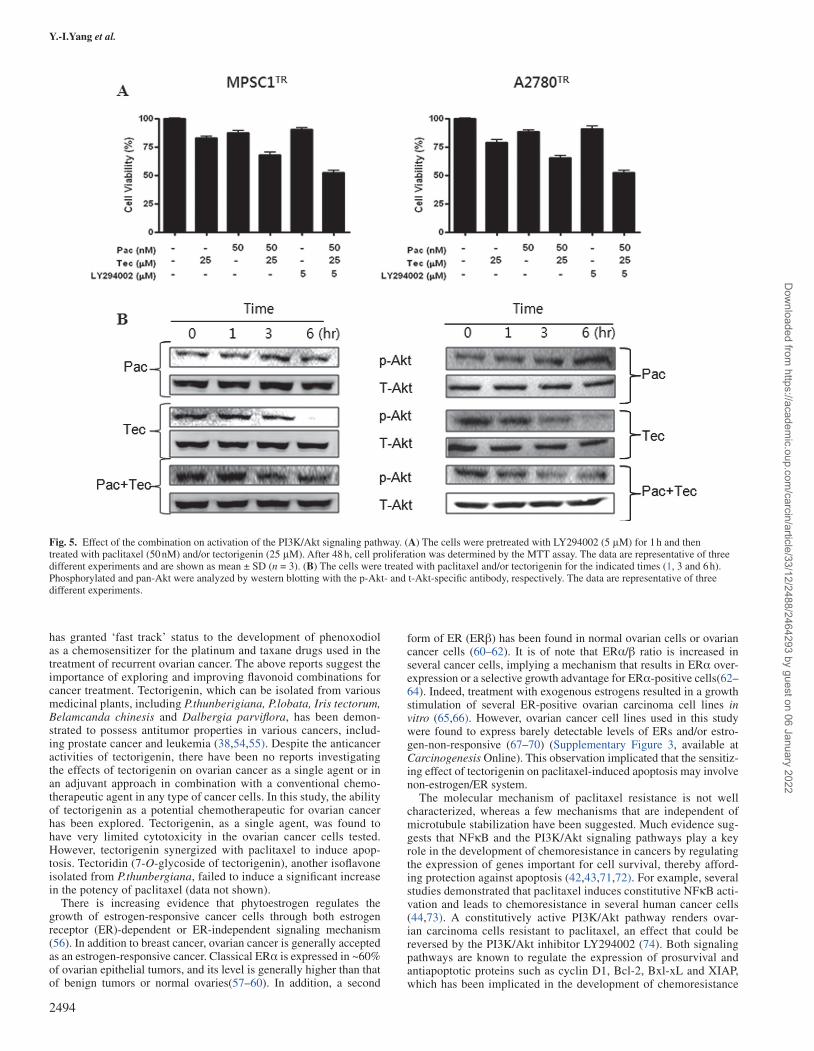
suggesting that Akt may play a role in combination-induced proapop-totic signaling (Figure 5A). In agreement with some previous findings (9,44), paclitaxel induced a slight increase in the phosphorylation of Akt in paclitaxel-resistant ovarian cancer cells (Figure 5B). We found that tectorigenin markedly inhibited the activation of Akt in a dose-dependent manner. Paclitaxel in combination with tectorigenin sig-nificantly inhibited the activation of Akt, whereas the total Akt level remained unaffected by all treatment conditions, indicating a possible role for Akt in the synergistic effect of paclitaxel and tectorigenin.
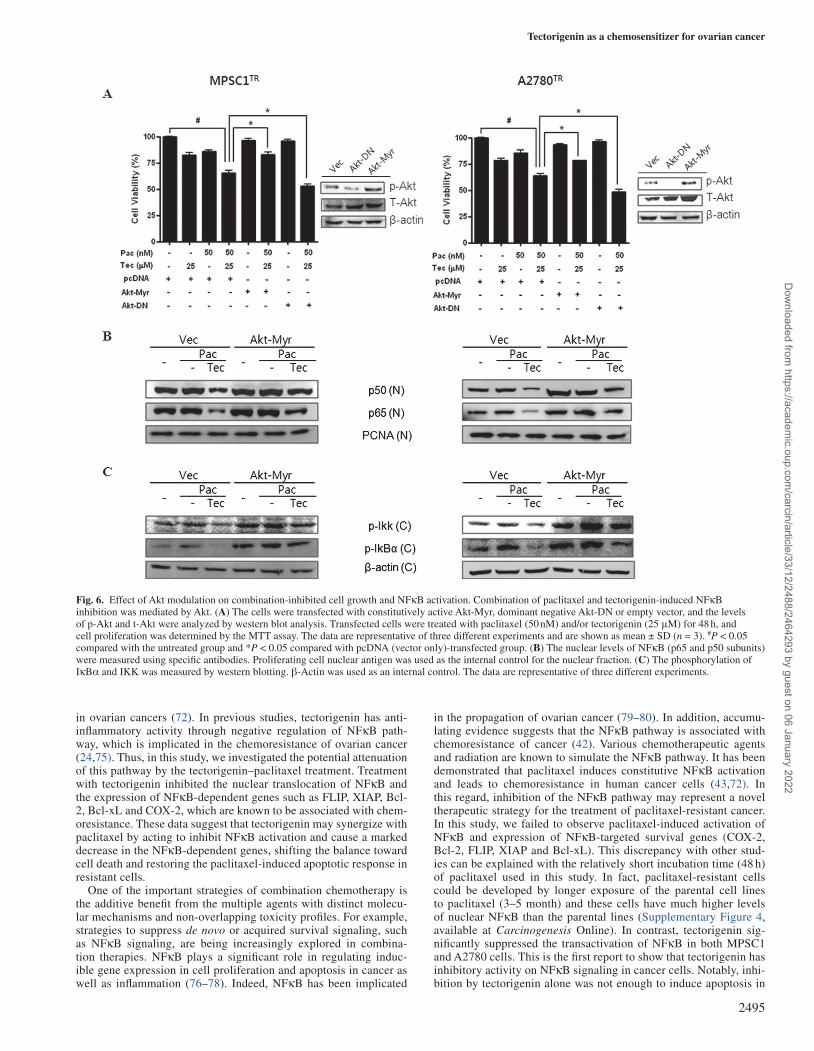
We next determined whether the modulation of the Akt signaling pathway is involved in combination-induced apoptosis in the resist-ant cells. Cells were transfected with the constitutively active form of Akt (Akt-Myr) or the dominant negative form of Akt (Akt-DN) and the activation and expression of Akt were confirmed by western blot analysis (Figure 6A). The inhibition of Akt by Akt-DN transfection
Fig. 3. Effect of the combination on apoptotic induction and the activity of caspases in MPSC1TR and A2780TR cells. (A) The cells were treated with paclitaxel (50 nM) and/or tectorigenin (25 μM) for 48 h and costained with PI and fluorescein isothiocyanate-conjugated annexin V. The translocation of phosphatidylserine was detected by flow cytometry after costunolide treatment. The graphs show the percentage of annexin V-positive cells. *P < 0.05 compared with the control group. (B) Cells were treated with paclitaxel (50 nM) and/or tectorigenin (25 μM) and western blot analysis was performed. Procaspase-3, -8, and -9 protein levels were normalized to β-actin. Representative data from one of the three separate experiments are shown. (C) To confirm the involvement of caspase signaling in combination-induced cell death, MTT assays were performed using the broad-spectrum inhibitor z-VAD-fmk (50 µM) and caspase-3-specific inhibitor z-DEVD-fmk (50 µM). Cells were pretreated with inhibitor for 30 min before being exposed to costunolide. The data are representative of three different experiments and are shown as mean ± SD (n = 3). #P < 0.05 compared with the control group and *P < 0.05 compared with the combination group.
Y.-I.Yang et al.
2492
Dow
nloaded from https://academ
ic.oup.com/carcin/article/33/12/2488/2464293 by guest on 06 January 2022

resulted in increased cell growth inhibition in tectorigenin-treated cells. Moreover, Akt-Myr significantly inhibited tectorigenin–pacli-taxel-induced apoptosis. These results showed that Akt expression overrode the effect of tectorigenin treatment and prevented the treated cells from undergoing cell death. These data clearly identified Akt as the upstream regulatory factor for tectorigenin augmentation of paclitaxel-induced cytotoxicity and apoptosis. We further investi-gated whether Akt inhibition by the combination acts downstream or upstream of NFκB inhibition. When Akt was activated by Akt-Myr overexpression, paclitaxel–tectorigenin-induced inhibition of NFκB nuclear translocation (Figure 6B) and IKK and IκB phosphorylation were markedly attenuated (Figure 6C). These data indicate that the inhibition of NFκB induced by the combination treatment is partly mediated by the inactivation of Akt through the inhibition of IKK and IκB in paclitaxel-resistant ovarian cancer cells.
Discussion
Paclitaxel (Taxol), a well-known microtubule stabilizer, has been used as the front-line chemotherapeutic agent for several types of human carcinomas, including ovarian cancer. However, the drug frequently induces drug resistance. The dose escalation required for overcoming the resistance of cancer cells can lead to severe
cytotoxicity in dose-limiting normal tissue. In this regard, develop-ing a therapeutic strategy using dual agents, rather than using single agents, has been evaluated to achieve higher therapeutic efficacy with less toxicity. Recently, natural compounds with high potential and relatively low toxicity have been explored for use in combi-nation with a conventional chemotherapeutic agent. For example, the curry spice curcumin, which is known to have antiinflammatory and chemopreventive activities, has been demonstrated to potentiate the antitumor activity of various chemotherapeutic agents, includ-ing taxane and platinum, in a wide range of cancer cells (45,46). Triptolide isolated from the Chinese herb, Tripterygium wilfordii, has been shown to enhance platinum-, doxorubicin-, CPT-11-, TNF-, and taxol-induced cytotoxicity in various types of cancer cells (47–51). In addition, flavonoids and their synthetic analogs have been intensely investigated in cancer treatment to increase the response to the chemotherapy. For example, the citrus flavonoid tangeretin sen-sitizes cisplatin-resistant human ovarian cancer cells through down-regulation of the PI3K/Akt pathway (52). Phenoxodiol, a synthetic derivative of the naturally occurring plant isoflavone genistein, has been demonstrated to have direct cytotoxic activity against different cancers. Additionally, phenoxodiol sensitizes chemoresistant ovar-ian cancer cells to platinum and taxane drugs, as well as gemcit-abine and topotecan (53). The U.S. Food and Drug Administration
Fig. 4. Effect of the combination on activation of the NFκB signaling pathway. The cells were treated with paclitaxel (50 nM) and/or tectorigenin (25 μM) for 48 h and then harvested for western blot analysis. (A) The nuclear levels of NFκB (p65 and p50 subunits) were measured using specific antibodies. Proliferating cell nuclear antigen was used as the internal control for the nuclear fraction. (B) The expression levels of NFκB-dependent genes (COX-2, Bcl-2, Bcl-xL, FLIP and XIAP) were analyzed by western blotting with their specific antibodies. (C) Cells were treated with paclitaxel and/or tectorigenin for the indicated times (1, 3 and 6 h). Phosphorylation of IκBα and IKK was measured by western blotting. β-Actin was used as a loading control. The data are representative of three different experiments.
Tectorigenin as a chemosensitizer for ovarian cancer
2493
Dow
nloaded from https://academ
ic.oup.com/carcin/article/33/12/2488/2464293 by guest on 06 January 2022
has granted ‘fast track’ status to the development of phenoxodiol as a chemosensitizer for the platinum and taxane drugs used in the treatment of recurrent ovarian cancer. The above reports suggest the importance of exploring and improving flavonoid combinations for cancer treatment. Tectorigenin, which can be isolated from various medicinal plants, including P.thunberigiana, P.lobata, Iris tectorum, Belamcanda chinesis and Dalbergia parviflora, has been demon-strated to possess antitumor properties in various cancers, includ-ing prostate cancer and leukemia (38,54,55). Despite the anticancer activities of tectorigenin, there have been no reports investigating the effects of tectorigenin on ovarian cancer as a single agent or in an adjuvant approach in combination with a conventional chemo-therapeutic agent in any type of cancer cells. In this study, the ability of tectorigenin as a potential chemotherapeutic for ovarian cancer has been explored. Tectorigenin, as a single agent, was found to have very limited cytotoxicity in the ovarian cancer cells tested. However, tectorigenin synergized with paclitaxel to induce apop-tosis. Tectoridin (7-O-glycoside of tectorigenin), another isoflavone isolated from P.thunbergiana, failed to induce a significant increase in the potency of paclitaxel (data not shown).
There is increasing evidence that phytoestrogen regulates the growth of estrogen-responsive cancer cells through both estrogen receptor (ER)-dependent or ER-independent signaling mechanism (56). In addition to breast cancer, ovarian cancer is generally accepted as an estrogen-responsive cancer. Classical ERα is expressed in ~60% of ovarian epithelial tumors, and its level is generally higher than that of benign tumors or normal ovaries(57–60). In addition, a second
form of ER (ERβ) has been found in normal ovarian cells or ovarian cancer cells (60–62). It is of note that ERα/β ratio is increased in several cancer cells, implying a mechanism that results in ERα over-expression or a selective growth advantage for ERα-positive cells(62–64). Indeed, treatment with exogenous estrogens resulted in a growth stimulation of several ER-positive ovarian carcinoma cell lines in vitro (65,66). However, ovarian cancer cell lines used in this study were found to express barely detectable levels of ERs and/or estro-gen-non-responsive (67–70) (Supplementary Figure 3, available at Carcinogenesis Online). This observation implicated that the sensitiz-ing effect of tectorigenin on paclitaxel-induced apoptosis may involve non-estrogen/ER system.
The molecular mechanism of paclitaxel resistance is not well characterized, whereas a few mechanisms that are independent of microtubule stabilization have been suggested. Much evidence sug-gests that NFκB and the PI3K/Akt signaling pathways play a key role in the development of chemoresistance in cancers by regulating the expression of genes important for cell survival, thereby afford-ing protection against apoptosis (42,43,71,72). For example, several studies demonstrated that paclitaxel induces constitutive NFκB acti-vation and leads to chemoresistance in several human cancer cells (44,73). A constitutively active PI3K/Akt pathway renders ovar-ian carcinoma cells resistant to paclitaxel, an effect that could be reversed by the PI3K/Akt inhibitor LY294002 (74). Both signaling pathways are known to regulate the expression of prosurvival and antiapoptotic proteins such as cyclin D1, Bcl-2, Bxl-xL and XIAP, which has been implicated in the development of chemoresistance
Fig. 5. Effect of the combination on activation of the PI3K/Akt signaling pathway. (A) The cells were pretreated with LY294002 (5 μM) for 1 h and then treated with paclitaxel (50 nM) and/or tectorigenin (25 μM). After 48 h, cell proliferation was determined by the MTT assay. The data are representative of three different experiments and are shown as mean ± SD (n = 3). (B) The cells were treated with paclitaxel and/or tectorigenin for the indicated times (1, 3 and 6 h). Phosphorylated and pan-Akt were analyzed by western blotting with the p-Akt- and t-Akt-specific antibody, respectively. The data are representative of three different experiments.
Y.-I.Yang et al.
2494
Dow
nloaded from https://academ
ic.oup.com/carcin/article/33/12/2488/2464293 by guest on 06 January 2022
in ovarian cancers (72). In previous studies, tectorigenin has anti-inflammatory activity through negative regulation of NFκB path-way, which is implicated in the chemoresistance of ovarian cancer (24,75). Thus, in this study, we investigated the potential attenuation of this pathway by the tectorigenin–paclitaxel treatment. Treatment with tectorigenin inhibited the nuclear translocation of NFκB and the expression of NFκB-dependent genes such as FLIP, XIAP, Bcl-2, Bcl-xL and COX-2, which are known to be associated with chem-oresistance. These data suggest that tectorigenin may synergize with paclitaxel by acting to inhibit NFκB activation and cause a marked decrease in the NFκB-dependent genes, shifting the balance toward cell death and restoring the paclitaxel-induced apoptotic response in resistant cells.
One of the important strategies of combination chemotherapy is the additive benefit from the multiple agents with distinct molecu-lar mechanisms and non-overlapping toxicity profiles. For example, strategies to suppress de novo or acquired survival signaling, such as NFκB signaling, are being increasingly explored in combina-tion therapies. NFκB plays a significant role in regulating induc-ible gene expression in cell proliferation and apoptosis in cancer as well as inflammation (76–78). Indeed, NFκB has been implicated
in the propagation of ovarian cancer (79–80). In addition, accumu-lating evidence suggests that the NFκB pathway is associated with chemoresistance of cancer (42). Various chemotherapeutic agents and radiation are known to simulate the NFκB pathway. It has been demonstrated that paclitaxel induces constitutive NFκB activation and leads to chemoresistance in human cancer cells (43,72). In this regard, inhibition of the NFκB pathway may represent a novel therapeutic strategy for the treatment of paclitaxel-resistant cancer. In this study, we failed to observe paclitaxel-induced activation of NFκB and expression of NFκB-targeted survival genes (COX-2, Bcl-2, FLIP, XIAP and Bcl-xL). This discrepancy with other stud-ies can be explained with the relatively short incubation time (48 h) of paclitaxel used in this study. In fact, paclitaxel-resistant cells could be developed by longer exposure of the parental cell lines to paclitaxel (3–5 month) and these cells have much higher levels of nuclear NFκB than the parental lines (Supplementary Figure 4, available at Carcinogenesis Online). In contrast, tectorigenin sig-nificantly suppressed the transactivation of NFκB in both MPSC1 and A2780 cells. This is the first report to show that tectorigenin has inhibitory activity on NFκB signaling in cancer cells. Notably, inhi-bition by tectorigenin alone was not enough to induce apoptosis in
Fig. 6. Effect of Akt modulation on combination-inhibited cell growth and NFκB activation. Combination of paclitaxel and tectorigenin-induced NFκB inhibition was mediated by Akt. (A) The cells were transfected with constitutively active Akt-Myr, dominant negative Akt-DN or empty vector, and the levels of p-Akt and t-Akt were analyzed by western blot analysis. Transfected cells were treated with paclitaxel (50 nM) and/or tectorigenin (25 μM) for 48 h, and cell proliferation was determined by the MTT assay. The data are representative of three different experiments and are shown as mean ± SD (n = 3). #P < 0.05 compared with the untreated group and *P < 0.05 compared with pcDNA (vector only)-transfected group. (B) The nuclear levels of NFκB (p65 and p50 subunits) were measured using specific antibodies. Proliferating cell nuclear antigen was used as the internal control for the nuclear fraction. (C) The phosphorylation of IκBα and IKK was measured by western blotting. β-Actin was used as an internal control. The data are representative of three different experiments.
Tectorigenin as a chemosensitizer for ovarian cancer
2495
Dow
nloaded from https://academ
ic.oup.com/carcin/article/33/12/2488/2464293 by guest on 06 January 2022

paclitaxel-resistant ovarian cancer cells. However, the combination of paclitaxel with tectorigenin significantly inhibited cell growth in paclitaxel-resistant ovarian cancer cells. NFκB is a dimer composed of p65 and p50 or p52 subunits and is located in the cytoplasm, where it is held by its endogenous inhibitor IκB. NFκB activation requires the degradation of its natural inhibitor IκB, which depends on IκB phosphorylation by the IKK complex and its subsequent ubiquitination and degradation by the proteasome, allowing NFκB nuclear translocation, binding to DNA and the activation of expres-sion of several genes required for cell survival and antiapoptosis. Paclitaxel alone did not change the phosphorylation of IκB but slightly increased the activation of IKK. In contrast, tectorigenin alone significantly inhibited the phosphorylation of both IκB and IKK. Thus, the combination-induced/decreased level of IκB and IKK phosphorylation may be due to the inhibitory effect of tecto-rigenin. Furthermore, these data indicate that the negative regulation of the NFκB pathway by the combination is mediated, at least partly, through the suppression of the IκB degradation and phosphorylation in paclitaxel-resistant ovarian cancer cells.
The PI3K/Akt signaling pathway is now accepted as being at least as important as the ras-mitogen-activated protein kinase pathway for cell survival and proliferation, and hence its potential role in car-cinogenesis is of immense interest. In ovarian cancer, it is becom-ing increasingly clear that the PI3K signaling pathway plays a major role in the regulation of cell proliferation, apoptosis, differentiation, tumorigenesis and angiogenesis (50,82,83). PI3K and its direct down-stream effector Akt are involved in the various aspects of cancer through the regulation of multiple downstream signals, among which the NFκB pathway is one of the most frequently mentioned targets. It is now well recognized that the PI3K/Akt pathway, regulating NFκB activation through an IKK-dependent mechanism, is heavily involved in keeping tumor cells alive by blocking apoptosis. In this study, the PI3K/Akt pathway was involved in the combination-induced cell death and the negative regulation of the IKK/IκB/NFκB pathway. In fact, the phosphorylation pattern of Akt by each single agent and their combination was very similar to that of IKK. These data suggest that Akt acts as an upstream regulatory factor of IKK, invoking the effect of tectorigenin in augmenting paclitaxel-induced cytotoxicity and apoptosis. It is of note that Akt-Myr overexpression only partially reversed the tectorigenin–paclitaxel-induced NFκB activation and cell death. These observations suggest the possibility that a signal-ing pathway other than Akt might be involved in the regulation of NFκB by the combination. In addition, the NFκB pathway might not be the only pathway for the action of paclitaxel, because the inhibitory effect of paclitaxel and tectorigenin was greater than that of paclitaxel and well-known NFκB inhibitors such as Bay11-7082, BMS-34551 and MG132 (Supplementary Figure 5, available at Carcinogenesis Online). Further molecular study is required.
Ovarian carcinoma remains one of the most lethal female malig-nancies and the fifth most common cause of cancer death in women. This high mortality is usually ascribed to the failure to detect ovar-ian cancer at an early stage and the lack of effective therapies for late-stage cancers. Today, the standard therapy for advanced ovarian carcinoma consists of extensive surgical resection followed by pacli-taxel/carboplatin combination chemotherapy. Although advances in surgery and chemotherapy have resulted in improved outcomes, 60–80% of patients with advanced disease relapse and die of com-plications related to the recurrent tumors (84). The development of resistance to the chemotherapeutic regimen of paclitaxel plus carbo-platin represents a major barrier to the long-term efficacy of ovarian cancer treatment. Therefore, it is urgent to understand the mecha-nisms underlying the development of resistance to chemotherapeu-tic agents and to develop novel drugs that re-sensitize tumor cells. Overall, the results presented here suggest that tectorigenin may enhance paclitaxel cytotoxicity against ovarian carcinoma cell lines by increasing the activation of caspase, leading to decreasing the NFκB and Akt pathway. The combination of paclitaxel and tecto-rigenin may provide a therapeutic benefit against paclitaxel-resistant ovarian cancers.
Supplementary material
Supplementary Table 1 and Figures 1–5 can be found at http://carcin.oxfordjournals.org/
Funding
Korea Science and Engineering Foundation (KOSEF) (to J.H.C.) funded by the Korea Government (MEST) (2010-0004306).
Acknowledgements
We are thankful to Dr Auersperg (University of British Columbia, Vancouver, British columbia) and Dr Andrew Godwin (Fox Chase Cancer Center, Philadelphia, PA) for providing IOSE-80 and IOSE-80PC cell lines.
Conflict of Interest Statement: None declared.
References
1. Parkin,D.M. et al. (2005) Global cancer statistics, 2002. CA. Cancer J. Clin., 55, 74–108.
2. Conte,P.F. et al. (1999) Up date in the management of advanced ovarian carcinoma. Crit. Rev. Oncol. Hematol., 32, 49–58.
3. Omura,G.A. et al. (1991) Long-term follow-up and prognostic factor analysis in advanced ovarian carcinoma: the Gynecologic Oncology Group experience. J. Clin. Oncol., 9, 1138–1150.
4. Agarwal,R. et al. (2003) Ovarian cancer: strategies for overcoming resist-ance to chemotherapy. Nat. Rev. Cancer, 3, 502–516.
5. Zunino,F. et al. (1997) Role of apoptotic response in cellular resistance to cytotoxic agents. Pharmacol. Ther., 76, 177–185.
6. Arts,H.J. et al. (2000) Options for modulation of drug resistance in ovarian cancer. Int. J. Gynecol. Cancer, 10(S1), 47–52.
7. Eliopoulos,A.G. et al. (1995) The control of apoptosis and drug resistance in ovarian cancer: influence of p53 and Bcl-2. Oncogene, 11, 1217–1228.
8. Herod,J.J. et al. (1996) The prognostic significance of Bcl-2 and p53 expression in ovarian carcinoma. Cancer Res., 56, 2178–2184.
9. Ling,X. et al. (2004) Induction of survivin expression by taxol (paclitaxel) is an early event, which is independent of taxol-mediated G2/M arrest. J. Biol. Chem., 279, 15196–15203.
10. Sasaki,H. et al. (2000) Down-regulation of X-linked inhibitor of apoptosis protein induces apoptosis in chemoresistant human ovarian cancer cells. Cancer Res., 60, 5659–5666.
11. Kirchhoff,S. et al. (2000) TCR-mediated up-regulation of c-FLIPshort correlates with resistance toward CD95-mediated apoptosis by block-ing death-inducing signaling complex activity. J. Immunol., 165, 6293–6300.
12. Krueger,A. et al. (2001) Cellular FLICE-inhibitory protein splice variants inhibit different steps of caspase-8 activation at the CD95 death-inducing signaling complex. J. Biol. Chem., 276, 20633–20640.
13. Havsteen,B. (1983) Flavonoids, a class of natural products of high pharma-cological potency. Biochem. Pharmacol., 32, 1141–1148.
14. Nishiumi,S. et al. (2011) Dietary flavonoids as cancer-preventive and ther-apeutic biofactors. Front. Biosci. (Schol. Ed)., 3, 1332–1362.
15. Shen,F. et al. (1997) Synergistic action of quercetin and genistein in human ovarian carcinoma cells. Oncol. Res., 9, 597–602.
16. Chen,M.H. et al. (2007) The proportional odds model for multivariate interval-censored failure time data. Stat. Med., 26, 5147–5161.
17. Tanaka,T. et al. (2002) Inhibitory effects of estrogenic compounds, 4-non-ylphenol and genistein, on 7,12-dimethylbenz[a]anthracene-induced ovar-ian carcinogenesis in rats. Ecotoxicol. Environ. Saf., 52, 38–45.
18. Klein,A. et al. (2006) Prolonged stabilization of platinum-resistant ovarian cancer in a single patient consuming a fermented soy therapy. Gynecol. Oncol., 100, 205–209.
19. McCann,S.E. et al. (2003) Risk of human ovarian cancer is related to die-tary intake of selected nutrients, phytochemicals and food groups. J. Nutr., 133, 1937–1942.
20. Ouyang,G. et al. (2009) Genistein induces G2/M cell cycle arrest and apop-tosis of human ovarian cancer cells via activation of DNA damage check-point pathways. Cell Biol. Int., 33, 1237–1244.
21. Thasni,K.A. et al. (2008) Genistein induces apoptosis in ovarian can-cer cells via different molecular pathways depending on Breast Cancer Susceptibility gene-1 (BRCA1) status. Eur. J. Pharmacol., 588, 158–164.
22. Choi,E.J. et al. (2007) Pro-apoptotic effect and cytotoxicity of genistein and genistin in human ovarian cancer SK-OV-3 cells. Life Sci., 80, 1403–1408.
Y.-I.Yang et al.
2496
Dow
nloaded from https://academ
ic.oup.com/carcin/article/33/12/2488/2464293 by guest on 06 January 2022

23. Gossner,G. et al. (2007) Genistein-induced apoptosis and autophagocytosis in ovarian cancer cells. Gynecol. Oncol., 105, 23–30.
24. Solomon,L.A. et al. (2008) Sensitization of ovarian cancer cells to cisplatin by genistein: the role of NF-kappaB. J. Ovarian Res., 1, 9.
25. Papazisis,K.T. et al. (2006) Synergistic effects of protein tyrosine kinase inhibitor genistein with camptothecins against three cell lines in vitro. Cancer Lett., 233, 255–264.
26. Silasi,D.A. et al. (2009) Phenoxodiol: pharmacology and clinical experi-ence in cancer monotherapy and in combination with chemotherapeutic drugs. Expert Opin. Pharmacother., 10, 1059–1067.
27. Shin,J.E. et al. (2006) Estrogenic effect of main components kakkalide and tectoridin of Puerariae Flos and their metabolites. Biol. Pharm. Bull., 29, 1202–1206.
28. Pan,C.H. et al. (2008) Tectorigenin inhibits IFN-gamma/LPS-induced inflammatory responses in murine macrophage RAW 264.7 cells. Arch. Pharm. Res., 31, 1447–1456.
29. Kim,Y.P. et al. (1999) Inhibition by tectorigenin and tectoridin of prosta-glandin E2 production and cyclooxygenase-2 induction in rat peritoneal macrophages. Biochim. Biophys. Acta, 1438, 399–407.
30. Morrissey,C. et al. (2004) Phytoestrogens derived from Belamcanda chin-ensis have an antiproliferative effect on prostate cancer cells in vitro. J. Urol., 172(6 Pt 1), 2426–2433.
31. Thelen,P. et al. (2005) Tectorigenin and other phytochemicals extracted from leopard lily Belamcanda chinensis affect new and established targets for therapies in prostate cancer. Carcinogenesis, 26, 1360–1367.
32. Kang,S.C. et al. (2006) Evaluation of oriental medicinal herbs for estro-genic and antiproliferative activities. Phytother. Res., 20, 1017–1019.
33. Umehara,K. et al. (2009) Flavonoids from the heartwood of the Thai medicinal plant Dalbergia parviflora and their effects on estrogenic-respon-sive human breast cancer cells. J. Nat. Prod., 72, 2163–2168.
34. Park,H.-J. et al. (1999) Isoflavone glycosides from the flowers of Pueraria thunbergiana. Phytochemistry, 51, 147–151.
35. Pohl,G. et al. (2005) Inactivation of the mitogen-activated protein kinase pathway as a potential target-based therapy in ovarian serous tumors with KRAS or BRAF mutations. Cancer Res., 65, 1994–2000.
36. Choi,J.H. et al. (2009) Functional analysis of 11q13.5 amplicon identifies Rsf-1 (HBXAP) as a gene involved in paclitaxel resistance in ovarian can-cer. Cancer Res., 69, 1407–1415.
37. Choi,J.H. et al. (2004) Overexpression of follicle-stimulating hormone receptor activates oncogenic pathways in preneoplastic ovarian surface epithelial cells. J. Clin. Endocrinol. Metab., 89, 5508–5516.
38. Lee,K.T. et al. (2001) Tectorigenin, an isoflavone of Pueraria thunbergiana Benth., induces differentiation and apoptosis in human promyelocytic leu-kemia HL-60 cells. Biol. Pharm. Bull., 24, 1117–1121.
39. Chou,T.C. et al. (1984) Quantitative analysis of dose-effect relationships: the combined effects of multiple drugs or enzyme inhibitors. Adv. Enzyme Regul., 22, 27–55.
40. Prasad,S. et al. (2010) Targeting inflammatory pathways by flavonoids for prevention and treatment of cancer. Planta Med., 76, 1044–1063.
41. Chen,S.S. et al. (2012) Advances in the treatment of ovarian cancer: a potential role of antiinflammatory phytochemicals. Discov. Med., 13, 7–17.
42. Nakanishi,C. et al. (2005) Nuclear factor-kappaB inhibitors as sensitizers to anticancer drugs. Nat. Rev. Cancer, 5, 297–309.
43. Kim,S.H. et al. (2007) Akt involvement in paclitaxel chemoresistance of human ovarian cancer cells. Ann. N. Y. Acad. Sci., 1095, 82–89.
44. Bava,S.V. et al. (2005) Sensitization of taxol-induced apoptosis by cur-cumin involves down-regulation of nuclear factor-kappaB and the serine/threonine kinase Akt and is independent of tubulin polymerization. J. Biol. Chem., 280, 6301–6308.
45. Goel,A. et al. (2010) Curcumin, the golden spice from Indian saffron, is a chemosensitizer and radiosensitizer for tumors and chemoprotector and radioprotector for normal organs. Nutr. Cancer, 62, 919–930.
46. Sreekanth,C.N. et al. (2011) Molecular evidences for the chemosensitiz-ing efficacy of liposomal curcumin in paclitaxel chemotherapy in mouse models of cervical cancer. Oncogene, 30, 3139–3152.
47. Matsui,Y. et al. (2008) Cancer-specific enhancement of cisplatin-induced cytotoxicity with triptolide through an interaction of inactivated glycogen synthase kinase-3beta with p53. Oncogene, 27, 4603–4614.
48. Westfall,S.D. et al. (2008) Role of triptolide as an adjunct chemotherapy for ovarian cancer. Chemotherapy, 54, 67–76.
49. Fidler,J.M. et al. (2003) PG490-88, a derivative of triptolide, causes tumor regression and sensitizes tumors to chemotherapy. Mol. Cancer Ther., 2, 855–862.
50. Chang,W.T. et al. (2001) Triptolide and chemotherapy cooperate in tumor cell apoptosis. A role for the p53 pathway. J. Biol. Chem., 276, 2221–2227.
51. Park,B. et al. (2011) Triptolide, histone acetyltransferase inhibitor, sup-presses growth and chemosensitizes leukemic cells through inhibition of gene expression regulated by TNF-TNFR1-TRADD-TRAF2-NIK-TAK1-IKK pathway. Biochem. Pharmacol., 82, 1134–1144.
52. Arafa,e.l.-.S.A. et al. (2009) Tangeretin sensitizes cisplatin-resistant human ovarian cancer cells through downregulation of phosphoinositide 3-kinase/Akt signaling pathway. Cancer Res., 69, 8910–8917.
53. Kelly,M.G. et al. (2011) Phase II evaluation of phenoxodiol in combina-tion with cisplatin or paclitaxel in women with platinum/taxane-refractory/resistant epithelial ovarian, fallopian tube, or primary peritoneal cancers. Int. J. Gynecol. Cancer, 21, 633–639.
54. Fang,R. et al. (2008) Cytotoxic effects of compounds from Iris tectorum on human cancer cell lines. J. Ethnopharmacol., 118, 257–263.
55. Thelen,P. et al. (2007) Phytoestrogens from Belamcanda chinensis regulate the expression of steroid receptors and related cofactors in LNCaP prostate cancer cells. BJU Int., 100, 199–203.
56. Zhao,E. et al. (2011) Phytoestrogen biological actions on Mammalian reproductive system and cancer growth. Sci. Pharm., 79, 1–20.
57. Risch,H.A. (1998) Hormonal etiology of epithelial ovarian cancer, with a hypothesis concerning the role of androgens and progesterone. J. Natl. Cancer Inst., 90, 1774–1786.
58. Vierikko,P. et al. (1983) Cytosol and nuclear estrogen and progestin recep-tors and 17 beta-hydroxysteroid dehydrogenase activity in non-diseased tis-sue and in benign and malignant tumors of the human ovary. Int. J. Cancer, 32, 413–422.
59. Willcocks,D. et al. (1983) Estrogen and progesterone receptors in human ovarian tumors. Gynecol. Oncol., 16, 246–253.
60. Rao,B.R. et al. (1991) Endocrine factors in common epithelial ovarian can-cer. Endocr. Rev., 12, 14–26.
61. Klaunig,J.E. et al. (2004) The role of oxidative stress in carcinogenesis. Annu. Rev. Pharmacol. Toxicol., 44, 239–267.
62. Brandenberger,A.W. et al. (1998) Estrogen receptor alpha (ER-alpha) and beta (ER-beta) mRNAs in normal ovary, ovarian serous cystadenocarci-noma and ovarian cancer cell lines: down-regulation of ER-beta in neoplas-tic tissues. J. Clin. Endocrinol. Metab., 83, 1025–1028.
63. Pujol,P. et al. (1998) Differential expression of estrogen receptor-alpha and -beta messenger RNAs as a potential marker of ovarian carcinogenesis. Cancer Res., 58, 5367–5373.
64. Rutherford,T. et al. (2000) Absence of estrogen receptor-beta expression in metastatic ovarian cancer. Obstet. Gynecol., 96, 417–421.
65. Chien,C.H. et al. (1994) Transcriptional activation of c-myc proto-onco-gene by estrogen in human ovarian cancer cells. Mol. Cell. Endocrinol., 99, 11–19.
66. Galtier,M. et al. (1992) Beta blockade prophylaxis of accessory pathway catecholamine sensitive circus movement tachycardia. Eur. J. Med., 1, 125–126.
67. Ercoli,A. et al. (2000) Activity of cisplatin and ICI 182,780 on estrogen receptor negative ovarian cancer cells: cell cycle and cell replication rate perturbation, chromatin texture alteration and apoptosis induction. Int. J. Cancer, 85, 98–103.
68. Lau,K.M. et al. (1999) Expression of human estrogen receptor-alpha and -beta, progesterone receptor, and androgen receptor mRNA in normal and malignant ovarian epithelial cells. Proc. Natl. Acad. Sci. U.S.A., 96, 5722–5727.
69. Brandenberger,A.W. et al. (1998) Estrogen receptor alpha (ER-alpha) and beta (ER-beta) mRNAs in normal ovary, ovarian serous cystadenocarci-noma and ovarian cancer cell lines: down-regulation of ER-beta in neoplas-tic tissues. J. Clin. Endocrinol. Metab., 83, 1025–1028.
70. Choi,J.H. et al. (2005) Expression of leptin receptors and potential effects of leptin on the cell growth and activation of mitogen-activated protein kinases in ovarian cancer cells. J. Clin. Endocrinol. Metab., 90, 207–210.
71. Mabuchi,S. et al. (2004) Inhibition of inhibitor of nuclear factor-kappaB phosphorylation increases the efficacy of paclitaxel in in vitro and in vivo ovarian cancer models. Clin. Cancer Res., 10, 7645–7654.
72. Fraser,M. et al. (2003) Chemoresistance in human ovarian cancer: the role of apoptotic regulators. Reprod. Biol. Endocrinol., 1, 66.
73. Huang,Y. et al. (2002) IkappaB kinase activation is involved in regulation of paclitaxel-induced apoptosis in human tumor cell lines. Mol. Pharmacol., 61, 105–113.
74. Hu,L. et al. (2002) Inhibition of phosphatidylinositol 3’-kinase increases efficacy of paclitaxel in in vitro and in vivo ovarian cancer models. Cancer Res., 62, 1087–1092.
75. Liu,G.H. et al. (2006) Inhibition of nuclear factor-kappaB by an antioxi-dant enhances paclitaxel sensitivity in ovarian carcinoma cell line. Int. J. Gynecol. Cancer, 16, 1777–1782.
Tectorigenin as a chemosensitizer for ovarian cancer
2497
Dow
nloaded from https://academ
ic.oup.com/carcin/article/33/12/2488/2464293 by guest on 06 January 2022

76. DiDonato,J.A. et al. (1997) A cytokine-responsive IkappaB kinase that activates the transcription factor NF-kappaB. Nature, 388, 548–554.
77. Huxford,T. et al. (1999) Structure and mechanism in NF-kappa B/I kappa B signaling. Cold Spring Harbor Symposia on Quantitative Biology, 64, 533–540.
78. Baldwin,A.S. Jr. (1996) The NF-kappa B and I kappa B proteins: new dis-coveries and insights. Annu. Rev. Immunol., 14, 649–683.
79. Hernandez,L. et al. (2010) Activation of NF-kappaB signaling by inhibi-tor of NF-kappaB kinase beta increases aggressiveness of ovarian cancer. Cancer Res., 70, 4005–4014.
80. Darb-Esfahani,S. et al. (2010) Expression of classical NF-kappaB pathway effectors in human ovarian carcinoma. Histopathology, 56, 727–739.
81. Annunziata,C.M. et al. (2010) Nuclear factor kappaB transcription factors are coexpressed and convey a poor outcome in ovarian cancer. Cancer, 116, 3276–3284.
82. Do,P.M. et al. (2012) Mutant p53 cooperates with ETS2 to promote etopo-side resistance. Genes Dev., 26, 830–845.
83. Brader,S. et al. (2004) Phosphoinositide 3-kinase signalling pathways in tumor progression, invasion and angiogenesis. Tumori, 90, 2–8.
84. Merritt,W.M. et al. (2009) Anti-angiogenic properties of metronomic topotecan in ovarian carcinoma. Cancer Biol. Ther., 8, 1596–1603.
Received May 17, 2012; revised September 11, 2012; accepted September 20, 2012
Y.-I.Yang et al.
2498
Dow
nloaded from https://academ
ic.oup.com/carcin/article/33/12/2488/2464293 by guest on 06 January 2022






















![[Third National Ovarian Consensus]](https://img.pdfslide.net/doc/110x75/6355648cf4b7d3d11c0c9f9a/third-national-ovarian-consensus.jpg)






