Embed Size (px)
Citation preview

The 3*,5*-Cyclic Adenosine Monophosphate ResponseElement Binding Protein (CREB) Is FunctionallyReduced in Human Toxic Thyroid Adenomas*
ANTONIO BRUNETTI, EUSEBIO CHIEFARI, SEBASTIANO FILETTI, AND
DIEGO RUSSO
Cattedra di Endocrinologia (A.B., E.C., S.F.), Dipartimento di Medicina Sperimentale e Clinica,Facolta di Medicina e Chirurgia; and Cattedra di Farmacologia (D.R.), Facolta di Farmacia,Universita degli Studi di Catanzaro, 88100 Catanzaro, Italy
ABSTRACTIn human normal thyrocytes, the cAMP-responsive signaling path-
way plays a central role in gene regulation, cell proliferation, anddifferentiation. Constitutive activation of the cAMP signal transduc-tion system has been documented in thyroid autonomously hyper-functioning adenomas in which activating mutations in either theTSH receptor gene or the Gsa protein gene (gsp oncogene) have beendescribed. The molecular mechanism whereby cAMP induces thyro-cyte proliferation is unknown, but recent evidence suggests that thetranscription factor cAMP response element binding protein (CREB)may serve as an important biochemical intermediate in this prolif-erative response. Herein we have investigated the expression ofCREB in normal and tumoral thyroid tissues from a series of ten
unrelated patients with autonomously hyperfunctioning adenomas,previously screened for mutations in the TSH receptor and Gsa genes.In all tumors examined, the expression of the activated, phosphory-lated form of CREB was markedly reduced compared with that of thecorresponding paired normal thyroid tissue, and this reduction wasindependent of the presence of mutations in the TSH receptor geneand Gsa gene. Moreover, no correlation was observed in these tissuesbetween CREB phosphorylation and either protein kinase A activityor protein phosphatase expression. Thus, these data suggest that inhuman hyperfunctioning thyroid adenomas, the PKA/CREB systemdoes not play a role in cell proliferation. (Endocrinology 141: 722–730,2000)
AUTONOMOUSLY hyperfunctioning thyroid adeno-mas are responsible for 5–30% of cases of thyrotoxi-
cosis and have a higher prevalence in iodine deficient areas.Histologically, they are well encapsulated benign tumorscharacterized by TSH-independent growth, iodine uptake,and function and by a large spectrum of clinical manifesta-tions, such as weight loss, sweating, nervousness, irritability,fine tremor, fatigue, and tachycardia (1).
Studies indicate that the cAMP signal transduction path-way is essential in mediating signaling in thyroid cells (2). Inthyrocytes, intracellular levels of cAMP are regulated pri-marily by adenylate cyclase, which is in turn modulated byextracellular stimuli mediated by TSH receptor (TSHR) andG proteins at the cell surface (2). While in most cell typescAMP inhibits cell proliferation by interfering with signalingthrough the mitogen-activated protein kinase pathway (3), innormal thyrocytes TSH activation of its receptor through anincrease of cAMP levels stimulates both cell proliferation anddifferentiation, as demonstrated by in vitro studies, mostlyusing nonhuman thyroid cell cultures (4). In human thyroidcells, a growth promoting effect of cAMP has not unequiv-ocally demonstrated. Moreover, in thyroid tumors the role ofTSHR-adenylate cyclase system is still unclear. Although
TSH is considered a growth promoter factor in thyroid tu-mors, studies indicate that cAMP may act as a growth in-hibitor in some human thyroid tumoral cell lines (5–9). Re-cently, activating mutations in either the TSHR gene or theGsa protein gene (the oncogene gsp), responsible for theconstitutive activation of the cAMP signal transduction sys-tem, have been documented in hyperfunctioning thyroidadenomas with different frequency (10) and have been pro-posed to play a role in the etiology of the disease. Consti-tutively activated cAMP pathway has also been implicatedin the formation of human pituitary adenomas, in which bothmutant gsp and overexpression of Gsa protein have beendescribed (11).
The molecular mechanism whereby cAMP stimulates bothproliferation and differentiation in human normal thyro-cytes, as well as in other endocrine systems, is still unknown.cAMP is known to stimulate the cAMP-dependent proteinkinase A (PKA), which in turn phosphorylates cytoplasmicand nuclear target proteins. One of the best characterizedPKA substrates is the nuclear transcription factor cAMP re-sponse element binding protein (CREB), which stimulatesthe transcription of cAMP-responsive genes after its phos-phorylation by PKA (12, 13). Recent evidence in vitro sug-gests that CREB is essential for a normal rate of growth of theFRTL-5 thyroid follicular cells (14). Therefore, examination ofCREB in thyroid adenomas may represent an importantpoint in the characterization of the molecular mechanismsthat are involved in the generation of thyroid tumors. Wehave investigated CREB phosphorylation in a series of hu-man toxic thyroid adenomas previously examined for the
Received July 15, 1999.Address all correspondence and requests for reprints to: Sebastiano
Filetti, M.D., Cattedra di Endocrinologie, Dipartimento di MedicinaSperimentale e Clinica, via T. Campanella, 88100 Catanzaro, Italy. E-mail: [email protected].
* This work was supported by a grant from the Associazione Italianaper la Ricerca sul Cancro and MURST (to S.F.).
0013-7227/00/$03.00/0 Vol. 141, No. 2Endocrinology Printed in U.S.A.Copyright © 2000 by The Endocrine Society
722

presence of either TSHR or gsp mutations (15, 16). We reportherein that CREB phosphorylation is markedly reduced in allthyroid adenomas relative to that in normal thyroid tissues,and this reduction appears to be independent by cAMP/PKApathway and phosphatase mediated dephosphorylation.
Materials and MethodsMaterials
BSA, HEPES, phenylmethylsulfonyl fluoride (PMSF), aprotinin, leu-peptin, Nonidet P-40, and T4 polynucleotide kinase were obtained fromSigma-Aldrich S.r.l. (Milan, Italy); radioisotopes, nitrocellulose mem-branes, autoradiography film and enhanced chemiluminescence (ECL)Western blotting detection reagents were from Amersham Italia S.r.l.(Milan, Italy); protein assay reagent and dithiothreitol (DTT) from Bio-Rad Laboratories, Inc. (Segrate, Milan, Italy); colorimetric protein kinaseA (PKA) assay kit from Pierce Chemical Co., Prodotti Gianni spa (Milan,Italy); poly(dI-dC) from Pharmacia Biotech Italia (Cologno Monzese,Milan, Italy); antibodies against CREB and CREB-PSer133, and 27-merdouble stranded consensus oligonucleotides containing DNA bindingsites for CREB transcription factor were purchased from Santa CruzBiotechnology, Inc. (Segrate, Milan, Italy); antihuman protein phospha-tase 1 (PP1) polyclonal antibody and antihuman protein phosphatase 2A(PP2A) monoclonal antibody were from Upstate Biotechnology, Inc.(Segrate, Milan, Italy); consensus oligonucleotide for OCT-1 transcrip-tion factor from Promega Corp. (Madison, WI). Basic laboratory pro-cedures were performed according to standard protocols (17) unlessotherwise stated.
Patients
Ten unrelated patients with hyperfunctioning thyroid adenomaswere studied (Table 1). Tissue specimens, obtained at the time of surgeryand frozen in liquid nitrogen, were taken from the tumor and from thehealthy surrounding tissue. The thyroid nodules were classified as au-tonomous toxic/hyperfunctioning adenomas on the basis of clinicalthyrotoxicosis, elevated serum levels of free thyroid hormones, andundetectable TSH levels, associated with a predominant 131I uptake inthe nodule as shown by 131I-scanning. All tumors were classified asadenomas according to conventional pathological criteria (18). Thisstudy was approved by the local ethical committee.
Nuclear protein extraction
Nuclei from 100 mg human thyroid were prepared by homogenizingthe tissue with a Tekmar homogenizer sd-45K homogenizer (19) in twovolumes of lysis buffer (10 mm HEPES pH 7.9, 25 mm KCl, 0.6 m sucrose,1 mm EDTA, 10% glycerol with 20 mg/ml aprotinin, 1 mm PMSF, 10 mmNa pyrophosphate, 1 mm Na3VO4, and 1 mm NaF). The homogenate wasfurther treated with a glass Dounce (type B), filtered through polypro-pylene mesh and centrifuged at 14,000 3 g at 4 C for 20 min. The crudenuclear pellet was washed twice with two volumes of lysis buffer,resuspended in two volumes of the same buffer (19) and nuclear extracts
were prepared following the procedure of Dignam et al., as previouslydescribed (20). Equal number of nuclei were homogenized and finalprotein concentrations in the extracts were determined using the col-orimetric assay of Bradford (19). The extraction of nuclear protein (1–2mg/ml) was the same for all of the samples analyzed. Nuclear extractswere normalized by use of the octamer (OCT-1) nuclear protein, amember of the OCT family that is apparently ubiquitous in mammaliancells (21). Consensus oligonucleotide for OCT-1 transcription factor was59 end labeled with (g-32P)ATP and T4 polynucleotide kinase and usedfor gel retardation assays under conditions suggested by the supplier(Promega Corp.).
Western blot analysis
Ten micrograms of nuclear protein derived from thyroid tissues wereseparated by SDS-PAGE (10% resolving gel) using a Minigel apparatus(Bio-Rad Laboratories, Inc.), and transferred to nitrocellulose mem-branes using transfer buffer containing 20% methanol, 25 mm Tris base,and 192 mm glycine and a Mini-transelectrophoretic transfer cell (Bio-Rad Laboratories, Inc.) (120 V, 1 h). After blocking the membranes for1.5 h at room temperature in PBS solution containing 5% BSA, themembranes were incubated at 4 C overnight in TBS buffer (20 mmTris-HCl, 150 mm NaCl, pH 7.5) containing 1:5000 dilution of polyclonalanti-CREB antibody specific for the total CREB-protein, or 1:2000 dilu-tion of polyclonal anti-CREB-PSer133 antibody specific for the 43-kDaphosphorylated CREB-protein and washed three times (10 min eachtime) in TBS with 0.2% Tween-20 (TBS-T). The membranes were thenincubated for 1 h at room temperature in TBS containing 1:5000 dilutionof goat antirabbit IgG antibody coupled to horseradish peroxidase, fol-lowed by two 30 min washings with TBS-T. Immunoreactive bands werevisualized by incubation with luminol and exposed to autoradiographyfilm. Quantification was achieved by densitometric scanning.
The same procedure was used for the immunodetection of PP1 andPP2A protein phosphatases in the nuclear extracts from normal andtumoral thyroid tissues.
Measurement of adenylate cyclase and PKA activities
Adenylate cyclase activity in normal and tumoral thyroid tissues wasassessed by measuring the amount of (32P)cAMP generated from(32P)ATP, following previously published procedures (22). For the PKAassay, thyroid tissues were washed with cold PBS, and homogenized ina buffer containing 20 mm HEPES pH 7.5, 10 mm EGTA, 40 mm b-glyc-erophosphate, 1% Nonidet P-40, 2.5 mm MgCl2, 1 mm dithiothreitol, 2mm sodium vanadate, 1 mm PMSF, 20 mg/ml aprotinin, and 20 mg/mlleupeptin. The activity of PKA was determined in each sample using acommercial nonradioactive PKA assay Kit (SpinZyme, Pierce ChemicalCo.) following the manufacturer’s instructions.
Gel retardation assay
Binding reactions were performed as previously described (20).Briefly, 10 mg of nuclear extracts from both normal and tumoral thyroidtissues were incubated with 2 ng of radiolabeled probe, in the presence
TABLE 1. Clinical and molecular characteristics of patientsa
Patient Sex Age (yr) FT3 FT4 TSH Gene mutation1 F 45 8.7 29.7 0.012 F 61 10.4 12.9 ND3 M 47 8.9 14.2 0.2 TSHR4 F 62 8.1 19.3 0.1 TSHR5 F 31 12.2 18.0 0.01 Gsa6 F 52 8.9 21.9 0.17 F 37 8.8 24.5 0.09 Gsa8 M 57 9.3 20.6 0.1 TSHR9 F 47 12.5 25.8 0.02
10 M 44 16.8 20.6 0.02
ND, Not detectable.Normal values: FT3 (3.4–7.2 pmol/liter); FT4 (10.3–25.7 pmol/liter); TSH (0.2–4.0 mU/liter).a Most of the patients presented only a T3-thyrotoxicosis, as expected as they came from an area with moderate/severe goitrogen endemia.
At diagnosis all of the patients presented weight loss, sweating, nervousness, irritability, fine tremor, fatigue and tachycardia.
CREB ACTIVITY IN HUMAN THYROID ADENOMAS 723
of 0.5 mg poly(dI-dC) which was used as competitor DNA for nonspecificDNA binding proteins in the nuclear extracts. After 30 min of incubationat 20 C, reaction products were separated by electrophoresis through anondenaturing 6% polyacrylamide gel, and free and bound DNA weredetected by autoradiography (20).
27-mer double stranded consensus oligonucleotides containing wild-type (59-AGAGATTGCCTGACGTCAGAGAGCTAG-39) and mutated(59-AGAGATTGCC TGTGGTCAGAGAGCTAG-39) cAMP response el-ement (CRE) motif were 59 end labeled with (g-32P)ATP and T4 polynu-cleotide kinase and used for gel retardation assays under conditionssuggested by the supplier (Santa Cruz Biotechnology, Inc.).
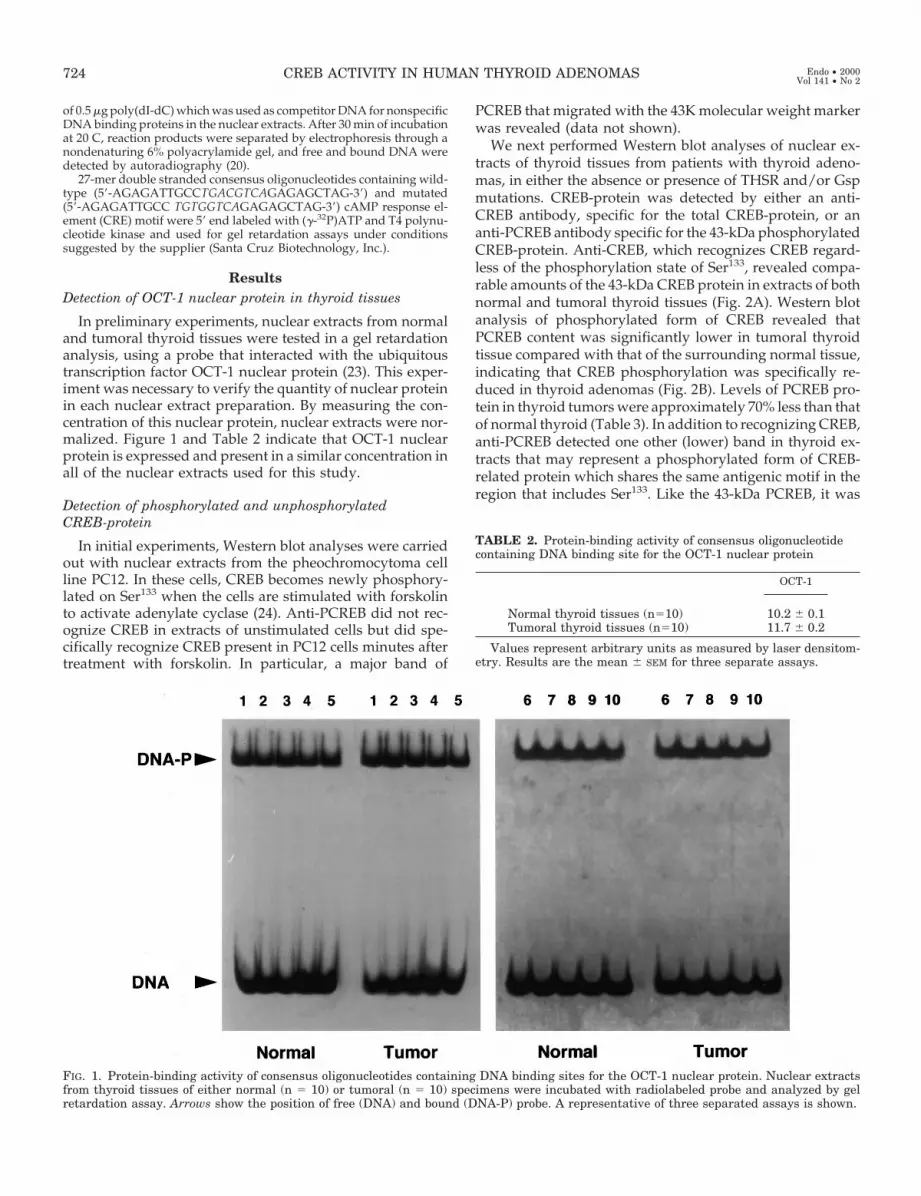
ResultsDetection of OCT-1 nuclear protein in thyroid tissues
In preliminary experiments, nuclear extracts from normaland tumoral thyroid tissues were tested in a gel retardationanalysis, using a probe that interacted with the ubiquitoustranscription factor OCT-1 nuclear protein (23). This exper-iment was necessary to verify the quantity of nuclear proteinin each nuclear extract preparation. By measuring the con-centration of this nuclear protein, nuclear extracts were nor-malized. Figure 1 and Table 2 indicate that OCT-1 nuclearprotein is expressed and present in a similar concentration inall of the nuclear extracts used for this study.
Detection of phosphorylated and unphosphorylatedCREB-protein
In initial experiments, Western blot analyses were carriedout with nuclear extracts from the pheochromocytoma cellline PC12. In these cells, CREB becomes newly phosphory-lated on Ser133 when the cells are stimulated with forskolinto activate adenylate cyclase (24). Anti-PCREB did not rec-ognize CREB in extracts of unstimulated cells but did spe-cifically recognize CREB present in PC12 cells minutes aftertreatment with forskolin. In particular, a major band of
PCREB that migrated with the 43K molecular weight markerwas revealed (data not shown).
We next performed Western blot analyses of nuclear ex-tracts of thyroid tissues from patients with thyroid adeno-mas, in either the absence or presence of THSR and/or Gspmutations. CREB-protein was detected by either an anti-CREB antibody, specific for the total CREB-protein, or ananti-PCREB antibody specific for the 43-kDa phosphorylatedCREB-protein. Anti-CREB, which recognizes CREB regard-less of the phosphorylation state of Ser133, revealed compa-rable amounts of the 43-kDa CREB protein in extracts of bothnormal and tumoral thyroid tissues (Fig. 2A). Western blotanalysis of phosphorylated form of CREB revealed thatPCREB content was significantly lower in tumoral thyroidtissue compared with that of the surrounding normal tissue,indicating that CREB phosphorylation was specifically re-duced in thyroid adenomas (Fig. 2B). Levels of PCREB pro-tein in thyroid tumors were approximately 70% less than thatof normal thyroid (Table 3). In addition to recognizing CREB,anti-PCREB detected one other (lower) band in thyroid ex-tracts that may represent a phosphorylated form of CREB-related protein which shares the same antigenic motif in theregion that includes Ser133. Like the 43-kDa PCREB, it was
TABLE 2. Protein-binding activity of consensus oligonucleotidecontaining DNA binding site for the OCT-1 nuclear protein
OCT-1
Normal thyroid tissues (n510) 10.2 6 0.1Tumoral thyroid tissues (n510) 11.7 6 0.2
Values represent arbitrary units as measured by laser densitom-etry. Results are the mean 6 SEM for three separate assays.
FIG. 1. Protein-binding activity of consensus oligonucleotides containing DNA binding sites for the OCT-1 nuclear protein. Nuclear extractsfrom thyroid tissues of either normal (n 5 10) or tumoral (n 5 10) specimens were incubated with radiolabeled probe and analyzed by gelretardation assay. Arrows show the position of free (DNA) and bound (DNA-P) probe. A representative of three separated assays is shown.
724 CREB ACTIVITY IN HUMAN THYROID ADENOMAS Endo • 2000Vol 141 • No 2

markedly reduced in nuclear extracts from thyroid adeno-mas. No correlation was observed between CREB proteinlevels and the presence of either TSHR or gsp mutations.
Detection of adenylate cyclase and PKA activities in normaland tumoral thyroid tissues
The importance of the adenylate cyclase in the regulationof intracellular levels of cAMP as well as the crucial roleplayed by PKA in the activation of CREB prompted us todetermine whether the reduced phosphorylation of CREBobserved in thyroid adenomas reflected a reduced adenylatecyclase and/or PKA activities in these tissues. In accordancewith the reports of others (25), adenylate cyclase activity wasonly slightly higher in all toxic adenomas when compared
with that of paired normal thyroid tissues (Fig. 3A), whereasthere were no measurable differences in PKA activity innormal and tumoral tissues (Fig. 3B).
Detection of PP2A protein phosphatase expression in normaland tumoral thyroid tissues
CREB is known to be dephosphorylated by PP1 and/orPP2A protein phosphatases (26, 27). We measured PP2Aprotein expression in nuclear extracts. As detected by West-ern blot analysis, comparable amounts of the 36-kDa PP2Aprotein were observed in both normal and tumoral thyroidtissues (Fig. 4, Table 4). Similar results were obtained with the34-kDa PP1 protein (data not shown).
FIG. 2. Levels of PCREB are reduced inhuman thyroid adenomas comparedwith normal thyroid. A, Western blotanalysis of nuclear extracts from nor-mal and tumoral tissues, using a poly-clonal anti-CREB antibody specific fortotal CREB protein. B, Western blots ofnuclear extracts using a polyclonal anti-PCREB antibody specific for phospho-CREB protein. The 43-kDa product rep-resents full-length PCREB protein. Arepresentative of three separated as-says is shown. N, Normal thyroid tissue(n 5 10); T, tumoral thyroid tissue (n 510).
TABLE 3. CREB protein levels and DNA-binding activity to CREB oligonucleotide in normal and tumoral thyroid tissues
Western blot analysis Gel retardation analysis
Normal tissues (n510) 10.0 6 1.2 9.2 6 0.16Tumoral tissues (n510) 2.97 6 1.2 2.5 6 0.1
Values represent arbitrary units as measured by laser densitometry. Results are the mean 6 SEM for three separate assays.
CREB ACTIVITY IN HUMAN THYROID ADENOMAS 725

Interaction of nuclear proteins from thyroid tissue withCREB consensus oligonucleotide
Nuclear extracts from thyroid tissue were incubated with32P-labeled CREB consensus oligonucleotide and tested forthe presence of specific DNA-binding factors using a gelretardation assay (20). In initial experiments using normalthyroid tissue, 10 mg of nuclear extracts were incubated withlabeled consensus oligonucleotide, in the presence of in-creasing amounts of the synthetic polynucleotide poly(dI-dC) (which was used as competitor DNA for non specificDNA binding proteins in the nuclear extracts). In vitro DNA-binding activity of nuclear extracts was supported by thedetection of a retarded DNA-protein complex (Fig. 5).
DNA binding activity to CREB consensus oligonucleotide innormal and tumoral thyroid tissues
We next determined binding of 32P-labeled CREB consen-sus oligonucleotide to nuclear proteins from normal andtumoral thyroid tissues. In tumoral tissue, DNA bindingactivity of nuclear extracts was significantly reduced (Figs. 6and 7). Laser densitometric scanning of the autoradiogramsrevealed that nuclear extracts from thyroid adenomas had
approximately 30% of DNA binding activity when comparedwith that of the surrounding normal thyroid tissue (Table 3).The decrease in DNA binding activity in nuclear extractsfrom thyroid adenomas paralleled the decrease in PCREBprotein abundance as detected by Western blot analysis. Thisreduction did not reflect the levels of adenylate cyclase ac-tivity observed in all tumoral thyroid tissues examined, sug-gesting that inhibition of DNA binding of CREB was unlikelydue to increased expression of ICERs (inducible cAMP earlyrepressors) that can be up-regulated, as part of feedback loop,by cAMP (28).
Specificity of DNA-protein binding
To determine specificity of DNA-protein binding, compe-tition assays were performed. The binding of thyrocyte nu-clear proteins to 32P-labeled CREB consensus oligonucleotidewas almost abolished when a 50-fold excess of unlabeledCREB consensus oligonucleotide was included in the bind-ing reaction (Fig. 7). Moreover, DNA binding activity ofnuclear extracts was prevented by using a 27-mer doublestranded synthetic oligonucleotide probe containing a mu-tated CRE motif (Fig. 7).
To confirm that PCREB protein was specifically reduced inthyroid adenomas, we next determined binding of a 27-merdouble stranded synthetic oligonucleotide containing DNAbinding site for TTF-1 transcription factor (29) to nuclearproteins from normal and tumoral thyroid tissues. As de-tected by gel retardation assays, nuclear extracts from tumortissue specimens demonstrated 5- to 10-fold enhanced DNA-binding activity when compared with that of the surround-ing normal tissue (Fig. 8).
Discussion
Based on a variety of in vitro data, mostly obtained withnonhuman thyroid cells, cAMP is considered to play a keyrole as a second messenger of TSH in stimulating both thy-rocyte proliferation and differentiation (2). Therefore, it hasbeen postulated that in thyroid tumors, at least in the moredifferentiated ones, an alteration in cAMP signal transduc-tion could participate in the development of the tumorigenicprocess. This alteration may be the consequence of mutationsor other defects present in some of the genes participating incAMP pathway. Somatic activating mutations of TSHR andGsa genes have been recently described in autonomouslyhyperfunctioning thyroid adenomas, in which a constitutiveactivation of the cAMP cascade has been demonstrated (10,31). However, although the constitutive activation of thecAMP cascade has been experimentally shown to cause in-creased proliferation of thyroid cells in vitro (32) and in vivo(33, 34), studies indicate that this mechanism may be alonenot sufficient to generate toxic thyroid adenomas (35), and farmore complex mechanisms may be effective in the patho-genesis of these tumors.
The importance of the nuclear regulatory protein CREB inmediating cAMP-dependent proliferative signals in thyroidcells has been recently documented, and recent evidencesuggests that CREB may serve as an important biochemicalintermediate in this proliferative response (14). In this study,we have investigated the expression of CREB in a series of
FIG. 3. Adenylate cyclase and PKA activities in normal and tumoralthyroid tissues. A, Adenylate cyclase activity. Activity of adenylatecyclase was performed by measuring the amount of (32P)cAMP gen-erated from (32P)ATP, following previously published procedures (22).Each value represents the mean of at least two determinations intriplicate. B, PKA activity. Activity of PKA was measured by spec-trophotometric analysis of the samples (see methods). Results are themean 6 SEM for three separated assays.3, Normal thyroid tissue (n 510); o, Tumoral thyroid tissue (n 5 10).
726 CREB ACTIVITY IN HUMAN THYROID ADENOMAS Endo • 2000Vol 141 • No 2

ten thyroid hyperfunctioning adenomas previously screenedfor the presence of TSHR and Gsa protein gene alterations.By using Western blot analysis, we measured the content ofCREB protein in thyroid tumors. We found that in all toxicadenomas examined the amounts of PCREB protein were 3-to 4-fold lower than that observed in nuclear extracts of thesurrounding normal tissue.
As a step toward understanding the molecular basis of theimpaired CREB phosphorylation observed in thyroid ade-nomas, we investigated whether this defect was accompa-nied by an altered adenylate cyclase and/or PKA activityand/or protein phosphatase expression in these tissues.
While the adenylate cyclase activity was significantly higherin all toxic adenomas compared with normal tissues, nosignificant differences in PKA activity were found. The ob-served discrepancy between adenylate cyclase activity andPKA activity may be related to the activation of cyclic nu-cleotide phosphodiesterases in tumoral tissues with persis-tent activation of adenylate cyclase. In this regard, increasein phosphodiesterase activity following the activation of theadenylate cyclase has been described either in vitro, in the ratthyroid cell line FRTL-5 expressing a constitutively activeGsa protein, or in vivo in human thyroid tissue (Clark, per-sonal communication) (32). It has been proposed that theoccurring phosphodiesterase feedback loop may counteractthe effects in the abnormal growth induced by the constitu-tive activation of adenylate cyclase (37, 38). Moreover, wehave data showing that in toxic thyroid adenomas phospho-
FIG. 4. Levels of PP2A protein phos-phatase in normal and tumoral thyroidtissues. Western blot analyses were car-ried out with nuclear extracts from thy-roid tissues using a mouse monoclonalantibody (1:2000 dilution) specific forthe human protein phosphatase 2A(PP2A). The 36-kDa PP2A protein isshown. A representative of three sepa-rated assays is shown. N, Normal thy-roid tissue (n 5 10); T, tumoral thyroidtissue (n 5 10).
TABLE 4. PP2A protein phosphatase levels in normal andtumoral thyroid tissues
PPSA
Normal tissues (n 5 10) 17.7 6 1.2Tumoral tissues (n 5 10) 18.1 6 0.9
Values represent arbitrary units as measured by laser densitom-etry. Results are the mean 6 SEM for three separate assays.
FIG. 5. Effect of the polynucleotide poly(dI-dC) on gel retardationassays with human thyroid nuclear extracts, using 27-bp CREB con-sensus oligonucleotide as probe. End-labeled CREB probe was incu-bated with 10 mg of nuclear extracts from normal thyroid tissues inthe presence of increasing amounts of poly(dI-dC) (0.1–20 mg) andDNA protein complexes were resolved on a nondenaturing 6% poly-acrylamide gel. Arrows show the position of the DNA protein com-plexes.
FIG. 6. Protein binding activity of CREB DNA to nuclear extractsfrom thyroid tissues. CREB DNA was labeled, incubated in the pres-ence of 0.5 mg poly(dI-dC) with nuclear extracts from normal andtumoral thyroid tissues, and DNA protein complexes were resolved asin Fig. 5. Lanes: C, probe alone; N, probe plus nuclear extract fromnormal thyroid tissue; T, probe plus nuclear extract from tumoralthyroid tissue. Arrows show the position of free (DNA) and bound(DNA-P) probe. A representative of three separated assays from eachpatient is shown.
CREB ACTIVITY IN HUMAN THYROID ADENOMAS 727

diesterase activity is higher in tumoral tissue compared withthat of the adjacent normal tissue. Discrepancy between ad-enylate cyclase and PKA activities has also been described inhuman brain tissue, where impaired G protein-stimulatedadenylate cyclase activity is not accompanied by reducedcAMP-dependent PKA activity (39). On the other hand, ourdata concerning protein phosphatase expression indicatethat the decreased phosphorylation of CREB in thyroid ad-enomas is not affected by dephosphorylation process. Thesefindings indicate, therefore, that the reduced phosphoryla-
tion of CREB in thyroid adenomas does not reflect a reducedlevel of adenylate cyclase and/or PKA activities and suggestthat CREB phosphorylation can be modulated by molecularmechanisms apparently independent by cAMP/PKA path-way (40). For example, phosphorylation of CREB throughmechanisms independent of cAMP and PKA has been re-ported in primary cell cultures from ovine pars tuberalis (41),and in the murine B lymphoma cell line BAL-17, in which ithas been shown that CREB Ser133 phosphorylation occursapparently without the intervention of PKA, and CREB re-sponds poorly to increased levels of cAMP (42). In addition,the existence of cAMP-dependent/PKA-independent tran-scriptional activation pathways, (with effects on differenti-ation and/or proliferation markers), has been postulated intwo recent studies, in which the exchange protein directlyactivated by cAMP (Epac) is able to activate Rap 1 protein ina cAMP-dependent but PKA-independent manner (43, 44),and the NIS upstream enhancer (NUE) in the 59 flankingregion of the rat sodium iodide symporter (NIS) gene, me-diates the cAMP-dependent transcription through a novelPKA-independent mechanism involving the thyroid specificfactor Pax-8 (45).
There is some controversy in the field concerning whetherphosphorylation at Ser133 is necessary for transcriptional ac-tivation of CREB and its DNA binding activity (26, 46). In ourstudy, we tested nuclear extracts from normal and tumoralthyroid tissues for their binding to consensus oligonucleo-tides containing DNA binding sites for the regulatory proteinCREB. Gel retardation analysis revealed that DNA bindingof PCREB was significantly reduced in nuclear extracts ob-tained from tumoral thyroid tissues, and this reduction par-alleled the decrease in PCREB protein levels as detected byWestern blot analysis. Therefore, these data indicate that inhuman hyperfunctioning thyroid adenomas cell prolifera-tion does not correlate with the activation of the cAMP/CREB pathway, suggesting the existence of other transform-ing events perhaps not dependent on the CREB family, asreported in BALB/c3T3 fibroblast cells, in which CREB hadno effect on cell growth, either in the presence or absence ofelevated cAMP (14). The possibility of cross-talk among in-
FIG. 7. Competition for binding between thyroid nuclear proteinsand CREB consensus oligonucleotide. CREB consensus oligonucleo-tide was 59 end-labeled and used as probe in gel retardation assayswith 10 mg of extracts from thyroid tissues under the same conditionsas in Fig. 6. Specificity of DNA-protein binding was determined byusing either a 50-fold molar excess of unlabeled CREB consensusoligonucleotide, or a labeled CREB mutant oligonucleotide. Lanes: 1,probe alone; 2, probe plus nuclear extract from tumoral thyroid tissue;3, probe plus nuclear extract from normal thyroid tissue; 4, probe plusnuclear extract from normal thyroid tissue in the presence of a 50-foldmolar excess of unlabeled CREB consensus oligonucleotide. In lane 5,nuclear extract from normal thyroid tissue were incubated with aCREB mutant oligonucleotide as probe. In lane 6, probe plus nuclearextract from normal thyroid tissue were incubated in the presence ofa specific anti-CREB antibody, able to induce a supershift of thecomplex. Arrows show the position of free (DNA) and bound (DNA-P)probes.
FIG. 8. Protein binding activity of TTF-1 DNA to nuclear extracts from thyroid tissues. TTF-1 DNA was labeled, incubated with nuclear extractsfrom normal (N) and tumoral (T) thyroid tissues, and DNA protein complexes were resolved as in Fig. 6. Arrows show the position of free (DNA)and bound (DNA-P) probe. The ubiquitous protein UFA (30) migrating below the TTF-1/DNA complex is visible. A representative of threeseparated assays from each patient is shown.
728 CREB ACTIVITY IN HUMAN THYROID ADENOMAS Endo • 2000Vol 141 • No 2

tracellular signaling pathways at a level before CREB phos-phorylation has been postulated (42). Reduced amounts ofCREB has been recently described also in proliferating, ac-tivated hepatic stellate cells (47).
Our data are in contrast with the results previously de-scribed in human GH-secreting tumors (11). In this study,CREB phosphorylation was elevated in pituitary adenomasexpressing the mutant gsp oncogene, and in tumors withoverexpression of Gsa protein relative to nonfunctioningadenomas. Instead, in our work, levels of CREB have beenstudied in thyroid samples obtained from the tumor andfrom the healthy surrounding tissue, and no correlationswere observed between CREB levels and the mutation ofeither TSHR or gsp.
We believe that this is the first report describing a quan-titative abnormality in CREB phosphorylation in thyroidtissues of human origin. The mechanism for this effect is stillunexplained, and it is possible that reduced levels of PCREBmay occur for differences in the cell cycle kinetics betweenthe adenoma and the normal tissue as observed in dog thy-roid cells (28). Studies aimed at better defining the mecha-nism(s) by which functionally reduced CREB is related to theprocess of thyroid tumorigenesis are in progress.
Acknowledgments
We thank Dr. F. Arturi for his collaboration in overviewing the clinicaldata of the patients and Prof. G. Damante for providing the consensusoligonucleotide containing DNA binding site for TTF-1 nuclear protein.
References
1. Hay ID, Morris JC 1991 Autonomously functioning thyroid nodules. In:Braverman LE, Utiger RD (eds) The Thyroid, A Fundamental and Clinical Text.JB Lippincott Company, Philadelphia, pp 698–704
2. Dumont JE, Lamy F, Roger P, Maenhaut C 1992 Physiological and patho-logical regulation of thyroid cell proliferation and differentiation by thyro-tropin and other factors. Physiol Rev 72:667–697
3. Wu J, Dent P, Jelinek T, Wolfman A, Weber M, Sturgill T 1993 Inhibition ofthe EGF-activated MAP kinase signalling pathway by adenosine 39,59 mono-phosphate. Science 262:1065–1069
4. Dremier S, Coppee F, Delange F, Vassart G, Dumont JE, Van Sande J 1996Thyroid autonomy: mechanism and clinical effects. J Clin Endocrinol Metab81:4187–4192
5. Helding NE, Cvejic D, Smeds S, Westermark B 1991 Expression of function-ally active receptors for thyrotropin and platelet-derived growth factor inhuman thyroid carcinoma cells. Endocrinology 129:2187–2193
6. Kimura H, Yamashita S, Namba H, Tominaga T, Tsuruta M, Yokoyama N,Izumi M, Nagataki S 1992 Interleukin-1 inhibits human thyroid carcinoma cellgrowth. J Clin Endocrinol Metab 75:596–602
7. Derwahl M, Kuemmel M, Goretzki P, Schatz H, Broecker M 1993 Expressionof the human thyroid carcinoma cell line that lacks an endogenous TSHreceptor: growth inhibition by cAMP. Biochem Biophys Res Commun191:1131–1138
8. Hishinuma A, Yamanaka T, Kasai T, So S, Bamba N, Shimoda S 1994 Growthregulation of the human thyroid cancer cell line by protein tyrosine kinase andcAMP-dependent protein kinase. Endocr J 41:399–407
9. Ohta K, Pang XP, Berg L, Hershman JM 1997 Growth inhibition of new humanthyroid carcinoma cell lines by activation of adenylate cyclase through theb-adrenergic receptor. J Clin Endocrinol Metab 82:2633–2638
10. Russo D, Arturi F, Chiefari E, Filetti S 1997 Molecular insights into TSHreceptor abnormality and thyroid disease. J Endocrinol Invest 20:36–47
11. Bertherat J, Chanson P, Montminy M 1995 The cyclic adenosine 39,59-mono-phosphate-responsive factor CREB is constitutively activated in human so-matotroph adenomas. Mol Endocrinol 7:777–783
12. Della Fazia MA, Servillo G, Sassone-Corsi P 1997 Cyclic AMP signalling andcellular proliferation: regulation of CREB and CREM. FEBS Lett 410:22–24
13. Gonzalez GA, Montminy MR 1989 Cyclic AMP stimulates somatostatin genetranscription by phosphorylation of CREB at serine 133. Cell 59:675–680
14. Woloshin PI, Walton KM, Rehfuss RP, Goodman RH, Cone RD 1992 39,59-cyclic adenosine monophosphate-regulated enhancer binding (CREB) activityis required for normal growth and differentiated phenotype in the FRTL5thyroid follicular cell line. Mol Endocrinol 6:1725–1733
15. Russo D, Arturi F, Wicker R, Chazenbalk GD, Schlumberger M, DuVillardJAD, Caillou B, Monier M, Rapoport B, Filetti S, Suarez HG 1995 Geneticalterations in thyroid hyperfunctioning adenomas. J Clin Endocrinol Metab80:1347–1351
16. Russo D, Arturi F, Suarez HG, Schlumberger M, DuVillard JAD, Crocetti U,Filetti S 1996 Thyrotropin receptor gene alterations in thyroid hyperfunction-ing adenomas. J Clin Endocrinol Metab 81:1548–1551
17. Sambrook J, Fritsch EF, Maniatis T 1989 Molecular Cloning: A LaboratoryManual. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
18. Hedinger C, Williams ED, Sabin LH 1989 The WHO histological classificationof thyroid tumors: a commentary on the second edition. Cancer 63:908–911
19. Yamamoto KK, Gonzalez GA, Biggs III WH, Montminy M 1988 Phospho-rylation-induced binding and transcriptional efficacy of nuclear factor CREB.Nature 334:494–498
20. Brunetti A, Foti D, Goldfine ID 1993 Identification of unique nuclear regu-latory proteins for the insulin receptor gene, which appear during myocyte andadipocyte differentiation. J Clin Invest 92:1288–1295
21. Brunetti A, Brunetti L, Foti D, Accili D, Goldfine ID 1996 Human diabetesassociated with defects in nuclear regulatory proteins for the insulin receptorgene. J Clin Invest 97:258–262
22. Derwahl M, Hamacher C, Russo D, Broecker M, Manole D, Schatz H, KoppP, Filetti S 1996 Constitutive activation of the Gs-a protein-adenylate cyclasepathway may not be sufficient to generate toxic thyroid adenomas. J ClinEndocrinol Metab 81:1898–1904
23. O’Neill EA, Fletcher C, Burrow CR, Hentz N, Roeder RG, Kelly TJ 1988Transcription factor OTF-1 is functionally identical to the DNA replicationfactor NF-III. Science 241:1210–1213
24. Ginty DD, Kornhauser JM, Thompson MA, Bading H, Mayo KE, TakahashiJS, Greenberg ME 1993 Regulation of CREB phosphorylation in the supra-chiasmatic nucleus by light and a circadian clock. Science 260:238–241
25. O’Sullivan C, Barton CM, Staddon SL, Brown CL, Lemoine NR 1991 Acti-vating point mutations of the gsp oncogene in human thyroid adenomas. MolCarcinog 4:345–349
26. Montminy M 1997 Transcriptional regulation by cyclic AMP. Annu Rev Bio-chem 66:807–822
27. Wadzinski BE, Wheat WH, Jaspers S, Peruski Jr LF, Lickteig LR, Johnson GL,Klemm DJ 1993 Nuclear protein phosphatase 2A dephosphorylates proteinkinase A-phosphorylated CREB and regulates CREB transcriptional stimula-tion. Mol Cell Biol 13:2822–2834
28. Uyttersprot N, Costagliola S, Dumont JE, Miot F 1999 Requirement for cAMP-response element (CRE) binding protein/CRE modulator transcription factorsin thyrotropin-induced proliferation of dog thyroid cells in primary culture.Eur J Biochem 259:370–378
29. Damante G, Di Lauro R 1994 Thyroid gene expression. Biochim Biophys Acta1218:255–266
30. Acebron A, Aza-Blanc P, Rossi DL, Lamas L, Santisteban P 1995 Congenitalhuman thyroglobulin defect due to low expression of the thyroid-specifictranscription factor TTF-1. J Clin Invest 96:781–785
31. Poertl S, Kirner J, Mann K, Hoermann R 1996 Enhanced functional activityin thyroid adenomas in vitro. Exp Clin Endocrinol Diabetes [Suppl 4] 104:39–40
32. Porcellini A, Ruggiano G, Pannain S, Ciullo I, Amabile G, Fenzi G, Av-vedimento EV 1997 Mutations of thyrotropin receptor isolated from thyroidautonomous functioning adenomas confer TSH-independent growth to thy-roid cells. Oncogene 15:781–789
33. Ledent C, Dumont JE, Vassart G, Parmentier M 1992 Thyroid expression ofan A2 adenosine receptor transgene induces thyroid hyperplasia and hyper-thyroidism. EMBO J 11:537–542
34. Michiels F, Caillou B, Talbot M, Dessarps-Freichey F, Maunaury M, Schlum-berger M, Merken L, Monier R 1994 Oncogenic potential of guanine nucle-otide stimulatory factor a subunit in thyroid gland of transgenic mice. ProcNatl Acad Sci USA 91:10488–10492
35. Derwahl M 1996 TSH receptor and Gs-a gene mutations in the pathogenesisof toxic thyroid adenomas - a note of caution. J Clin Endocrinol Metab81:2783–2785
36. Nemoz G, Sette C, Hess M, Muca C, Vallar L, Conti M 1995 Activation ofcyclic nucleotide phosphodiesterases in FRTL-5 thyroid cells expressing aconstitutively active Gsa. Mol Endocrinol 9:1279–1287
37. Weinstein LS, Shenker A, Gejman PV, Merino MJ, Friedman E, Spiegel AM1991 Activating mutations of the stimulatory G protein in the McCune-Al-bright syndrome. N Engl J Med 325:1688–1695
38. Schwindinger WF, Francomano CA, Levine MA 1992 Identification of amutation in the gene encoding the alpha subunit of the stimulatory G proteinof adenylyl cyclase in McCune-Albright syndrome. Proc Natl Acad Sci USA89:5152–5156
39. Bonkale WL, Fastbom J, Wiehager B, Ravid R, Winblad B, Cowburn RF 1996Impaired G-protein-stimulated adenylyl cyclase activity in Alzheimer’s dis-ease brain is not accompanied by reduced cyclic-AMP-dependent proteinkinase A activity. Brain Res 737:155–161
40. Armstrong R, Wen W, Meinkoth J, Taylor S, Montminy M 1995 A refractoryphase in cyclic AMP-responsive transcription requires down regulation ofprotein kinase A. Mol Cell Biol 15:1826–1832
CREB ACTIVITY IN HUMAN THYROID ADENOMAS 729

41. McNulty S, Ross AW, Shiu KY, Morgan PJ, Hastings MH 1996 Phosphor-ylation of CREB in ovine pars tuberalis is regulated both by cyclic AMP-dependent and cyclic AMP-independent mechanisms. J Neuroendocrinol8:635–645
42. Xie H, Wang Z, Rothstein TL 1996 Signaling pathways for antigen receptor-mediated induction of transcription factor CREB in B lymphocytes. Cell Im-munol 169:264–270
43. Kawasaki H, Springett GM, Mochizuki N, Toki S, Nakaya M, Matsuda M,Housman DE, Graybiel AM 1998 A family of cAMP-binding proteins thatdirectly activate Rap1. Science 282:2275–2279
44. deRooij J, Zwartkruis FJ, Verheijen MH, Cool RH, Nijman SM, Wittinghofer
A, Bos JL 1998 Epac is a Rap1 guanine-nucleotide-exchange factor directlyactivated by cyclic AMP. Nature 396:474–477
45. Ohno M, Zannini M, Levy O, Carrasco N, Di Lauro R 1999 The paired-domaintranscription factor Pax8 binds to the upstream enhancer of the rat sodium/iodide symporter gene and participates in both thyroid-specific and cyclic-AMP-dependent transcription. Mol Cell Biol 19:2051–2060
46. Schmid W, Nitsch D, Boshart M, Schutz G 1993 Role of cyclic AMP in thecontrol of cell-specific gene expression. Trends Endocrinol Metab 4:204–209
47. Houglum K, Kwan SL, Chojkier M 1997 Proliferation of hepatic stellate cellsis inhibited by phosphorylation of CREB on serine 133. J Clin Invest 99:1322–1328
730 CREB ACTIVITY IN HUMAN THYROID ADENOMAS Endo • 2000Vol 141 • No 2





























