Embed Size (px)
Citation preview

Neurochemistry International 42 (2003) 239–249
The altered expression of GABA shunt enzymes in the gerbilhippocampus before and after seizure generation
Tae-Cheon Kanga,∗, Seung-Kook Parka, In Koo Hwanga, Sung-Jin Ana, Soo Young Choib,Oh-Shin Kwonc, Nam-In Baekd, Hyeon Yong Leee, Moo Ho Wona,1
a Department of Anatomy, College of Medicine, Hallym University, Chunchon 200-702, Kangwon-Do, South Koreab Department of Genetic Engineering, College of Life Science, Hallym University, Chunchon 200-702, Kangwon-Do, South Korea
c Department of Biochemistry, College of Natural Sciences, Kyungpook National University, Taegu 702-701, South Koread Department of Life Science and Plant Metabolism Research Center, Kyunghee University, Suwon 449-701, South Korea
e Division of Food and Biotechnology, Kangwon National University, Chunchon 200-701, Kangwon-Do, South Korea
Received 26 February 2002; received in revised form 6 May 2002; accepted 8 May 2002
Abstract
In the present study, the distribution of succinic semialdehyde dehydrogenase (SSADH) and succinic semialdehyde reductase (SSAR)in the hippocampus of the Mongolian gerbil and its association with various sequelae of spontaneous seizure were investigated in orderto identify the roles of GABA shunt in the epileptogenesis and the recovery mechanisms in these animals. Both SSADH and SSARimmunoreactivities in the GABAergic neurons were significantly higher in the pre-seizure groups of seizure sensitive (SS) gerbil ascompared to those seen in the seizure resistant (SR) gerbils. The distributions of both SSADH and SSAR immunoreactivities in thehippocampus showed significant differences after the on-set of seizure. At 3 h postictal, when compared to the pre-seizure group of SSgerbils, a decline in the immunoreactivities in the perikarya was observed. At 12 h after seizure on-set, the densities of both SSADH andSSAR immunoreactivities were begun to recover to the pre-seizure level of SS gerbils. These results suggest that the GABAergic neurons inthe hippocampal complex of the SS gerbil may be highly activated. In addition, the imbalance of GABA shunt expressions in the GABAergicneurons may imply a malfunction of the metabolism of GABAergic neurons in the SS gerbils, and this defect may trigger seizure on-set.Therefore, the initiation of seizure, at least in gerbils, may be the result of a malfunction in GABA shunt in the GABAergic neurons.© 2003 Elsevier Science Ltd. All rights reserved.
Keywords: Epilepsy; Seizure; GABA shunt; Hippocampus; Gerbil; Succinic semialdehyde dehydrogenase; Succinic semialdehyde reductase
1. Introduction
Although the hippocampal formation is a region thatfrequently exhibits hyperexcitability and seizure activity inchronic models of epilepsy, the functional state of GABAin the hippocampus continues to be debated. Some inves-tigators have suggested that GABAergic neurons in thehippocampal formation may be less active in seizure-proneanimals (Bekenstein and Lothman, 1993; During et al.,
Abbreviations: GABA, �-aminobutyric acid; GABA-T, GABA transa-minase; GAD, glutamic acid decarboxylase; SSAR, succinic semialde-hyde reductase; SSADH, succinic semialdehyde dehydrogenase; SR,seizure resistant; SS, seizure sensitive; PBS, phosphate-buffered saline;PB, phosphate-buffer
∗ Corresponding author. Tel.:+82-33-240-1613-4;fax: +82-33-256-1614.
E-mail addresses: [email protected] (T.-C. Kang),[email protected] (M.H. Won).
1 Co-corresponding author.
1995; Gulyas et al., 1999). Thus, decreased GABAergic in-hibition has been suggested as one cause of hyperexcitabil-ity (De Lanerolle et al., 1989; Obenaus et al., 1993; Viziand Sperlagh, 1999), and a reduction in GABA-mediatedinhibition is thought to underlie seizure susceptibility(Brooks-Kayal et al., 1998; Buhl et al., 1996).
On the other hand, other data has suggested that there isan increase rather than a decrease in GABAergic function.Many investigators have recently demonstrated that many ofthe GABAergic neurons that remain in these animals receiveafferent input and are electrophysiologically active duringthe chronic stage (Esclapez et al., 1997; Rempe et al., 1997).Increased GAD expression has been observed in GABAergicinterneurons of the hippocampus in patients with temporallobe epilepsy and in rats following kainate induced seizures(Babb et al., 1989; Feldblum et al., 1990; Marksteinerand Sperk, 1988). Electrical kindling also appears to in-crease recurrent inhibition in the dentate gyrus (Maru andGoddard, 1987). According to these hypotheses, seizures
0197-0186/03/$ – see front matter © 2003 Elsevier Science Ltd. All rights reserved.PII: S0197-0186(02)00079-7

240 T.-C. Kang et al. / Neurochemistry International 42 (2003) 239–249
begin because there are too many GABAergic interneu-rons and too many inhibitory synaptic contacts between theGABAergic interneurons (disinhibition hypothesis).
The Mongolian gerbil exhibits spontaneous seizures inresponse to a variety of stimuli without neuron degen-eration that would be induced by neurotoxins includingkainate. Moreover, epileptic and non-epileptic animals canbe directly compared to detect differences in brain anatomyand electrophysiology that correlate with seizure behavior(Armijo et al., 1992; Buchhalter, 1993; Kang et al., 2000a,b).Interestingly, epileptogenesis in this animal has been linkedto abnormalities of the GABAergic system in the hippocam-pus. For example, seizure susceptibility in SS gerbils isreduced by the administration of GABA agonist (Loscher,1984), in spite of an excessive population of GABAergicneurons in the dentate gyrus, as compared to SR gerbils(Peterson et al., 1985; Peterson and Ribak, 1987). Further-more, our recent report demonstrated that the mismatchedGAD65/GAD67 ratio may closely related to the epilepto-genesis in the gerbil (Kang et al., 2001a). Therefore, thesereports have provided a background for the hypothesis thatGABA degradation may play a key role in epileptogenesisin this animal. However, in our previous study (Kang et al.,2001b), there was no difference observed in GABA-T ex-pression between the SR and pre-seizure group of SS gerbils.
Therefore: (1) whether the enzymes associated to thedownstream of GABA shunt may change chronologicallyafter seizure on-set; and (2) whether these alterations maycorrelate with the attenuation of seizure activity, needs tobe clarified. In the present study, both SSADH and SSARexpressions in the gerbil hippocampus associated with vari-ous sequelae of spontaneous seizures as well as the roles ofGABA shunt enzymes in epilepsy were investigated.
2. Materials and methods
2.1. Experimental animals
This study utilized the progeny of Mongolian gerbils(Meriones unguiculatus) obtained from Experimental An-imal Center, Hallym University, Chunchon, South Korea.The animals were provided with a commercial diet andwater ad libitum under controlled temperature, humidityand lighting conditions (22± 2 ◦C, 55± 5% and a 12:12light/dark cycle with lights). Procedures involving ani-mals and their care were conducted in conformity with theinstitutional guidelines that are in compliance with inter-national laws and policies (NIH Guide for the Care andUse of Laboratory Animals, NIH Publication No. 85-23,1985).
Each animal was tested a minimum of three times as de-scribed byPaul et al. (1981). Only animals with a consistentstage 4 or 5 seizure score, according to the seizure severityrating scale ofLoskota et al. (1974), were included in thepresent study as SS gerbils. SR gerbils never demonstrated
the seizure activity; thus they were assigned seizure severityscores of 0.
Seventy-five SS and 15 SR gerbils (about 8-month old)were used in the present experiment. To examine the tem-poral changes of both SSADH and SSAR expressions fol-lowing seizure, SS gerbils were divided into five groups;pre-seizure group, post-seizure groups I–IV (n = 15, re-spectively) that recovered normally at 30 min, 3 , 12 or 24 hafter the onset of tonic–clonic generalized seizure, respec-tively (Kang et al., 2000a,b).
2.2. Tissue processing and immunohistochemistry
The gerbils were anesthetized with pentobarbital sodium,and perfused via the ascending aorta with 200 ml of4% paraformaldehyde in PB. The brains were removed,post-fixed in the same fixative for 4 h and rinsed in PBcontaining 30% sucrose at 4◦C for 2 days. Thereafter,the tissues were frozen and sectioned with a cryostat at30�m and consecutive sections were collected in six-wellplates containing PBS. These free-floating sections werefirst incubated with 10% normal horse serum for 30 min atroom temperature. They were then incubated in the mouseanti-SSADH or SSAR antibody (diluted 1:500–1000;Choet al., 1996; Choi et al., 1995, 2001) in PBS containing0.3% Triton X-100 and 2% normal horse serum overnightat room temperature. After washing three times for 10 minwith PBS, sections were incubated sequentially, in horseanti-mouse IgG (Vector, USA) and streptavidin (Vector,USA), diluted 1:200 in the same solution as the primary an-tiserum. Between the incubations, the tissues were washedwith PBS three times for 10 min each. The sections werevisualized with 3,3′-diaminobenzidine (DAB) in 0.1 MTris buffer and mounted on the gelatin-coated slides. Theimmunoreactions were observed under the Axioscope mi-croscope (Carl Zeiss, Germany). In order to establish thespecificity of the immunostaining, a negative control testwas carried out with pre-immune serum instead of primaryantibody. The negative control resulted in the absence ofimmunoreactivity in any structures.
2.3. Enzymatic assays
Three animals in each group were used in this immunoblotstudy. After sacrifice and removal of hippocampus, the tis-sues were homogenized in 10 mM potassium PB containing0.1 mM EDTA, 1 mM 2-mercaptoethanol and 1 mM PMSF.The individual 10% (w/v) homogenates were centrifuged at10,000× g for 1 h. Protein concentration was determinedby Bio-rad dye binding protein assay, using bovine serumalbumin as a standard. Five hundred micrograms of eachsupernatant were used for enzymatic assays.
To analyze SSADH activity, the formation of NADH wasmeasured by the increase in absorbance at 340 nm. All assayswere performed in duplicate and the initial velocity data wascorrelated with a standard assay mixture containing 30�M

T.-C. Kang et al. / Neurochemistry International 42 (2003) 239–249 241
succinic semialdehyde and 1 mM NAD+ in 0. 1 M sodiumpyrophosphate (pH 8.4) at 25◦C.
In addition, to assay SSAR activity, the oxidation ofNADPH to NADP+ was measured by following the de-crease in absorbance at 340 nm. All assays were performedin duplicate, and initial velocity data were correlated witha standard assay mixture containing succinic semialde-hyde (120 M) and NADPH (50 M) in 0.1 M potassium PB(pH 7.2) at 25◦C. One unit of enzyme activity was de-fined as the amount of enzyme required to oxidize 1 M ofNADPH/min at 25◦C. And then, the spectrophotometricmeasurements were carried out using a Kontron UVIKON930 double beam spectrophotometer. Fluorescence spectrawere recorded on a Kontron SFM 25 spectrofluorimeter.
2.4. Quantitation of data and statistical analysis
Fifteen sections per one animal were viewed through amicroscope connected via CCD camera to a PC monitor. Ata magnification of 25–50×, the regions were outlined onthe monitor and measured their areas. Images of GAD im-munoreactivity in hippocampal complex were captured withan Applescanner. The brightness and contrast of each imagefile were calibrated by Adobe Photoshop version 2.4.1, fol-lowed by analysis using NIH Image 1.59 software. Valuesof background staining were obtained and subtracted fromthe immunoreactive intensities. All data obtained from the
Fig. 1. Showing coronal sections of the dentate gyrus of SR and SS gerbils. Compared to the case of SR (A), neurons in the dentate hilus are stronglystained for SSADH in the pre-seizure group of SS gerbils (B). At 3 h postictal, the number of neurons is significantly decreased due to the loss ofthe SSADH immunoreactivity in the neurons (C). The density of SSADH immunoreactivity at 12 h after the on-set of seizure is slightly elevated (D).Bar = 50�m.
quantitative data were analyzed using one-way ANOVA testto determine statistical significance. Bonferroni’s test wasused for post-hoc comparisons.P value below 0.01 or 0.05was considered statistically significant.
3. Results
3.1. The dentate gyrus
In the present study, both SSADH and SSAR immunore-activities were weakly detected in the granule cell layer andin the hilar neurons of SR gerbils. A similar distributionof these immunoreactivities in this region was observed inthe pre-seizure group of SS gerbils, however, both SSADHand SSAR immunoreactivities were significantly higher inthe pre-seizure groups of SS gerbils than in the SR gerbils(Figs. 1A and B, and 2A and B). Interestingly, the distribu-tions of both SSADH and SSAR immunoreactivities in thedentate gyrus showed significant differences after the on-setof seizure. In post-seizure group II (3 h postictal), as com-pared to the pre-seizure group of SS gerbils, the declinesof their immunoreactivities in the perikarya was observed(Figs. 1C and 2C). However, there were no differences ineither SSADH or SSAR immunoreactivities at 30 min pos-tictal (data not shown). At 12 h after seizure on-set, the den-sities of both SSADH and SSAR immunoreactivities wereslightly increased (Figs. 1D and 2D).

242 T.-C. Kang et al. / Neurochemistry International 42 (2003) 239–249
Fig. 2. Showing coronal sections of the dentate gyrus of SR and SS gerbils. Compared to SR gerbils (A), the SSAR immunoreactivity is strongly detectedin the hilar neurons of the pre-seizure group of SS gerbils (B). At 3 h postictal, the SSADH immunoreactivity in the neurons is significantly decreases(C). The SSAR immunoreactivity begins to enhance at 12 h after the on-set of seizure (D). Bar= 50�m.
Fig. 3. Showing coronal sections of the CA2–3 regions of SR (A) and SS gerbils (B–D). The SSADH immunodensity in the pre-seizure group (B)is higher than that in SR gerbil (A). At 3 h postictal (C), a decline in SSADH immunoreactivity in the neurons is observed. At 12 h postictal (D), itsimmunoreactivity in neurons is re-elevated. Bar= 100�m.

T.-C. Kang et al. / Neurochemistry International 42 (2003) 239–249 243
Fig. 4. Showing coronal sections of the CA2–3 regions of SR (A) and SS gerbils (B–D). Similar to the SSADH, the decline of SSAR immunoreactivityis observed at 3 h after the onset of seizure (C), and its immunoreactivity is re-enhanced until 12 h (D). (A) SR, (B) pre-seizure group. Bar= 100�m.
Fig. 5. Immunohistochemical staining for SSADH in the subiculum. The SSADH immunoreactivity in the pre-seizure groups is strongly observed (B), ascompared to the SR gerbils (A). A noticeable decrease in the SSADH immunoreactivity is observed by 3 h after seizure-onset (C). Its immunoreactivityin the subiculum is re-elevated at 12 h postictal (D). Bar= 50�m.
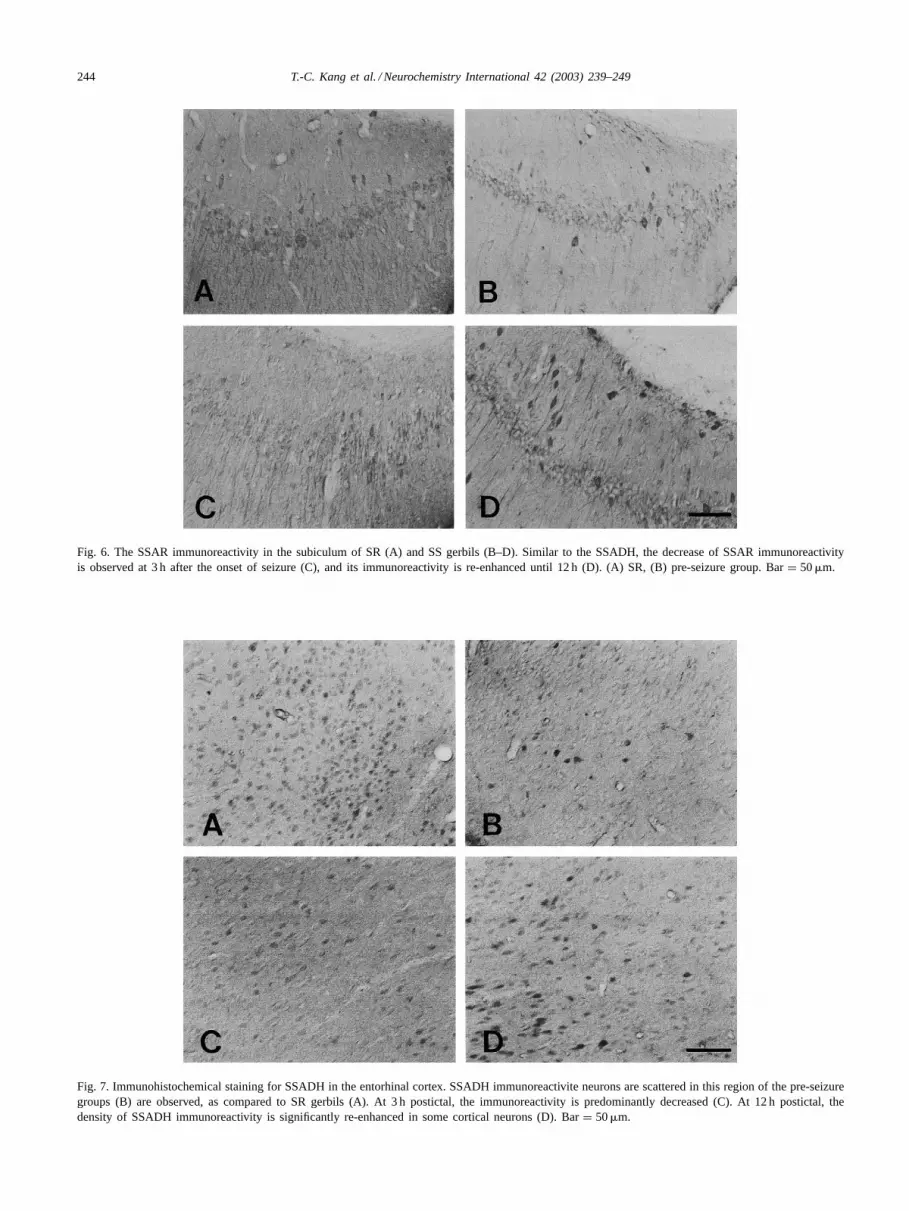
244 T.-C. Kang et al. / Neurochemistry International 42 (2003) 239–249
Fig. 6. The SSAR immunoreactivity in the subiculum of SR (A) and SS gerbils (B–D). Similar to the SSADH, the decrease of SSAR immunoreactivityis observed at 3 h after the onset of seizure (C), and its immunoreactivity is re-enhanced until 12 h (D). (A) SR, (B) pre-seizure group. Bar= 50�m.
Fig. 7. Immunohistochemical staining for SSADH in the entorhinal cortex. SSADH immunoreactivite neurons are scattered in this region of the pre-seizuregroups (B) are observed, as compared to SR gerbils (A). At 3 h postictal, the immunoreactivity is predominantly decreased (C). At 12 h postictal, thedensity of SSADH immunoreactivity is significantly re-enhanced in some cortical neurons (D). Bar= 50�m.
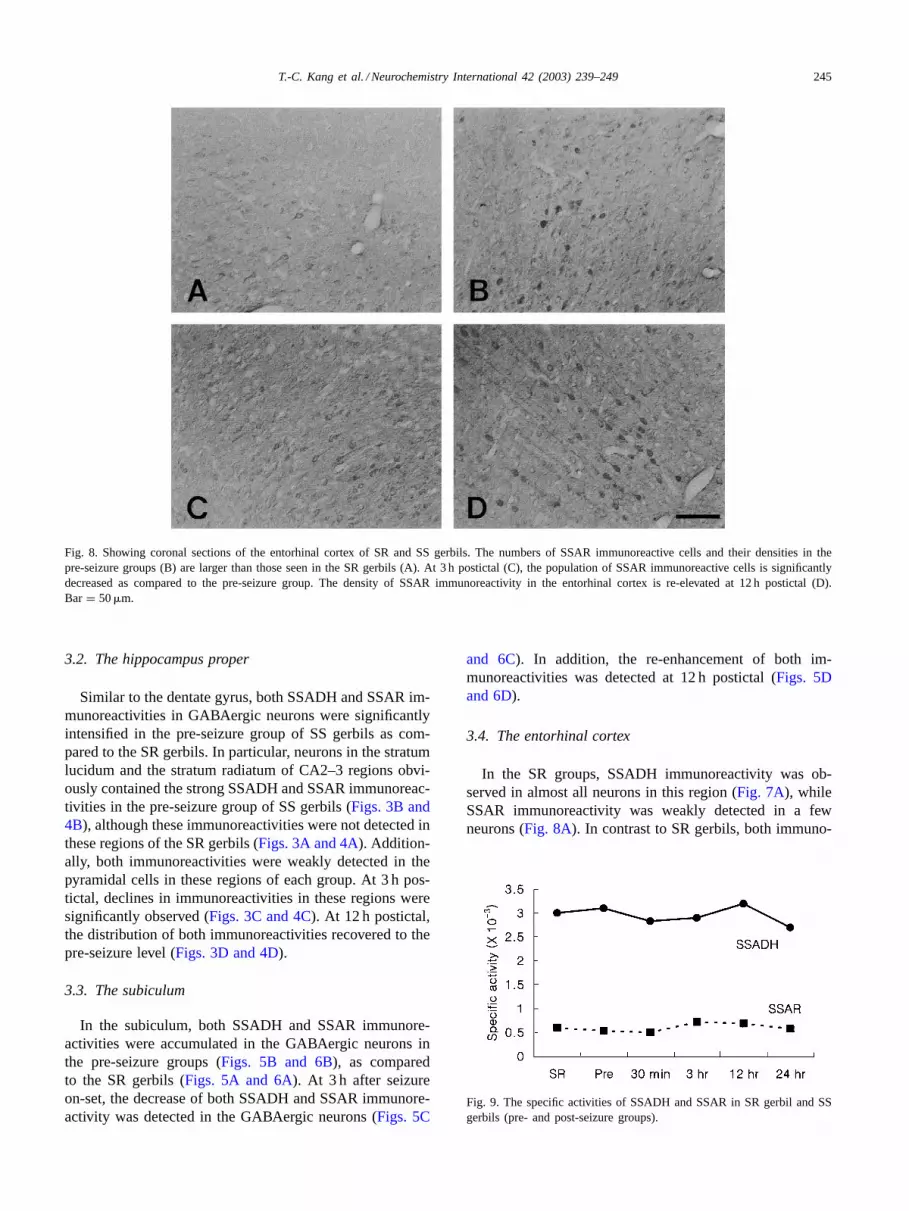
T.-C. Kang et al. / Neurochemistry International 42 (2003) 239–249 245
Fig. 8. Showing coronal sections of the entorhinal cortex of SR and SS gerbils. The numbers of SSAR immunoreactive cells and their densities in thepre-seizure groups (B) are larger than those seen in the SR gerbils (A). At 3 h postictal (C), the population of SSAR immunoreactive cells is significantlydecreased as compared to the pre-seizure group. The density of SSAR immunoreactivity in the entorhinal cortex is re-elevated at 12 h postictal (D).Bar = 50�m.
3.2. The hippocampus proper
Similar to the dentate gyrus, both SSADH and SSAR im-munoreactivities in GABAergic neurons were significantlyintensified in the pre-seizure group of SS gerbils as com-pared to the SR gerbils. In particular, neurons in the stratumlucidum and the stratum radiatum of CA2–3 regions obvi-ously contained the strong SSADH and SSAR immunoreac-tivities in the pre-seizure group of SS gerbils (Figs. 3B and4B), although these immunoreactivities were not detected inthese regions of the SR gerbils (Figs. 3A and 4A). Addition-ally, both immunoreactivities were weakly detected in thepyramidal cells in these regions of each group. At 3 h pos-tictal, declines in immunoreactivities in these regions weresignificantly observed (Figs. 3C and 4C). At 12 h postictal,the distribution of both immunoreactivities recovered to thepre-seizure level (Figs. 3D and 4D).
3.3. The subiculum
In the subiculum, both SSADH and SSAR immunore-activities were accumulated in the GABAergic neurons inthe pre-seizure groups (Figs. 5B and 6B), as comparedto the SR gerbils (Figs. 5A and 6A). At 3 h after seizureon-set, the decrease of both SSADH and SSAR immunore-activity was detected in the GABAergic neurons (Figs. 5C
and 6C). In addition, the re-enhancement of both im-munoreactivities was detected at 12 h postictal (Figs. 5Dand 6D).
3.4. The entorhinal cortex
In the SR groups, SSADH immunoreactivity was ob-served in almost all neurons in this region (Fig. 7A), whileSSAR immunoreactivity was weakly detected in a fewneurons (Fig. 8A). In contrast to SR gerbils, both immuno-
Fig. 9. The specific activities of SSADH and SSAR in SR gerbil and SSgerbils (pre- and post-seizure groups).

246 T.-C. Kang et al. / Neurochemistry International 42 (2003) 239–249
Fig. 10. The intensities of SSADH and SSAR immunoreactivity in the hippocampal complex after the on-set of seizure. Den, dentate gyrus; Hp,hippocampus proper; Sub, subiculum; Ent, entorhinal cortex (∗P < 0.05, ∗∗P < 0.01, significant differences from seizure-resistant gerbils).
reactivities in the pre-seizure group were accumulated ina few neurons, and their densities were more intensified(Figs. 7B and 8B). At 3 h postictal, both SSADH andSSAR immunoreactivities were significantly decreasedin this region (Figs. 7C and 8C). At 12 h postintal, bothimmunoreactivities began to enhance and recover to the SSlevel (Figs. 7D and 8D).
3.5. The analyses of enzyme activities andquantitative data
As for the results of the enzyme activity assay, the specificactivities of both SSADH and SSAR in the pre-seizure groupof SS gerbils was slightly higher than those in the SR gerbils.However, there was no statistical difference between thesegroups. In addition, the specific activities of these enzymeswere not altered following seizure, versus the pre-seizuregroup. The specific activities of SSADH and SSAR in theSR gerbil were 0.003 and 0.0006 units/mg protein, respec-tively, and those in the SS gerbils (pre- and post-seizuregroups) were 0.0026–0.0032 and 0.0005–0.00072 units/mgprotein, respectively (Fig. 9). On the other hand, the re-sults of the quantitative data obtained using image anal-ysis were consistent with the immunohistochemical study(Fig. 10).
4. Discussion
The major findings of the present study are as follows: (1)both SSADH and SSAR immunoreactivies were accumu-lated in the GABAergic neurons in the hippocampal complexof pre-seizure SS gerbils as compared to the SR gerbils; (2)there was no difference in the enzyme activities between SRgerbil and pre-seizure groups; (3) the immunoreactivities,but nor activities, were altered in the hippocampal complexfollowing seizure on-set.
4.1. The difference of GABA shunt expression in thehippocampal complex between SR and SS gerbils
It is well established that GABA metabolism plays an im-portant role in the epileptogenesis of human patients andexperimental animal models. However, one of the questionsthat have eluded researchers in the past is whether there is adifference in the GABAergic system, synthesis or degrada-tion, in the hippocampus (seeSection 1). Particularly in thegerbils, excessive GABAergic neurons in the dentate gyrusof SS gerbils have been reported (Peterson et al., 1985;Peterson and Ribak, 1987). Additionally, in our previousstudy, the expression of GABA-T in the hippocampus ofthe pre-seizure group of SS gerbils showed no difference ascompared to the SR gerbils. Therefore, the roles of GABAmetabolism in the epileptogenesis of this animal remain anenigma.
In the present study, we observed the differential distri-bution of SSADH and SSAR in the hippocampal complexbetween the SR and the pre-seizure groups of SS gerbils.Both enzymes were strongly expressed in the hippocampalcomplex of the pre-seizure SS gerbils, as compared to SRgerbils, although GABA-T expression did not differ betweenthe SR gerbil and the pre-seizure group (Kang et al., 2001b).These results are consistent with previous study demonstrat-ing the administration of GABA-T inhibitor induces the el-evation of SSADH expression (for review,Maitre, 1997).Therefore, these mismatches in the expressions of GABAshunt enzymes may be a compensatory reaction to regulatethe seizure activity in the gerbil. In addition, we could notdetect the alteration of both SSADH and SSAR specific ac-tivities in the SS gerbil hippocampus before/after seizureon-set, as compared with the SR gerbil one. These resultsindicate that the alteration of both SSADH and SSAR ex-pressions, in the hippocampal complex of the SS gerbil maydirectly control the GABA shunt ratio without altered spe-cific enzyme activities.

T.-C. Kang et al. / Neurochemistry International 42 (2003) 239–249 247
However, these results may be crucial evidence that couldexplain the paradoxical increases in the GABAergic neuronsin the pre-seizure group of SS gerbils (Peterson et al., 1985;Peterson and Ribak, 1987) and the mechanism of epilepto-genesis in this animal. This is because the elevations of bothSSADH and SSAR expressions may demonstrate that theGABA shunt in the GABAergic neurons is enhanced, andas a result the concentration of GABA used as a metabolitemay be higher than that used as a neurotransmitter. Thus,epileptogenesis in the SS gerbil may result from less GABAsecretion in the nerve terminals and lower concentration ofthis amino acid in the synaptic clefts, as compared to the SRgerbils. This hypothesis is supported by the lower number ofsynaptic vesicles in the GABAergic nerve terminals of theSS gerbil than in the SR gerbils, in spite of increase in thenumber of hippocampal GABAergic neurons and terminalsin the SS gerbil (Peterson et al., 1985; Peterson and Ribak,1987). Therefore, our findings indicate the hyper-activityand the high-energy consumption of GABAergic neurons inthe hippocampus of the SS gerbil. This is because a directrelationship between GABA metabolism and the averagefiring rate of GABAergic neurons is present in the epilep-tic hippocampus (Erlander et al., 1991; Erlander and Tobin,1991; Feldblum et al., 1993; Hendry, 1991; Najlerahim et al.,1992). Furthermore,Sierra-Paredes et al. (2000)have re-ported that extracellular glutamate and aspartate levels weredecreased during seizure, and suggested that extra-synapticamino acid pools may play an important role in seizure ac-tivity. Therefore, in the present study, alterations of GABAshunt enzymes in the GABAergic neurons also suggest thatchanged non-synaptic GABA level, similar to other neuro-transmitter, may be one of the epileptogenic factors in thegerbil.
4.2. Change in the expressions, but not activities, ofSSADH and SSAR following seizure on-set
In the present study, 30 min after the on-set of seizure,the expressions of both SSADH and SSAR were similarlydetected in the hippocampal complex, as compared withpre-seizure group. In addition, our previous study (Kanget al., 2001b) reported an accumulation of GABA-T im-munoreactivity in GABAergic neurons at 30 min postictalseizure. These changes are temporally similar to the re-sults of another our previous studies, which demonstratedincreases in neuropeptide Y biosynthesis, somatostatin re-lease in the dentate hilar neurons 30 min postictal (Kanget al., 2000a,b). Therefore, energy metabolisms, particularlyGABA shunt (Gerfen and Wilson, 1996; Schmid et al., 1999)in GABAergic neurons may be increased due to the releaseor synthesis of the neurotransmitters.
However, expressions of these enzymes were lower at 3 hpostictal. This data suggests that GABA degradation maybe decreased, but GABA concentration in the nerve ter-minal and synaptic cleft may be elevated to regulate neu-ronal excitability in the hippocampal complex. These results
were consistent with previous studies (Gale et al., 1984;Giorgi and Meek, 1984; Le Gal La Salle et al., 1984; Yunget al., 1998) reporting that administrations of kainate, aseizure-inductive compound, result in a marked reductionof GABA-T. Furthermore, the inhibition of GABA shunt byvigabatrin, an irreversible antagonist of GABA-T, enhancesGABA efflux via reversal of the GABA transporter (Wuet al., 2001).
Therefore, these decreases indicate that the GABA con-centration or activity in the nerve terminal and synaptic cleftmay be elevated. Moreover, our previous study has demon-strated that the expressions of both GAD65 and GAD67 issignificantly elevated in the gerbil hippocampus followingseizure (Kang et al., 2001a). Therefore, the decreases of bothSSADH and SSAR expressions in the gerbil hippocampalcomplex following seizure may be related to the attenuationof seizure activity via the increase of GABA turnover. Thisis because in synaptosomes, GABA shunt enzymes, particu-larly GABA-T, are primary factors in regulating the turnoverof GABA (Choi et al., 1996; Erecinska et al., 1996). Thishypothesis is also supported by our previous study report-ing that the expression of GABA-T was reduced in the ger-bil dentate gyrus at 3 h postictal in order to regulate seizureactivity (Kang et al., 2001b).
4.3. Differential patterns of SSADH and SSARimmunoreactivities in the hippocampal complex
In the present study, the temporal alteration in bothSSADH and SSAR immunoreactivities was constant beforeor after seizure on-set. However, the spatial patterns ofthese enzyme immunoreactivities were different from neu-ronal types. In the hippocampal complex, both SSADH andSSAR immunoreactivities were strongly accumulated inthe GABAergic neurons. However, in the entorhinal cortex,SSADH immunoreactivity was observed in most neurons inthe SR gerbils, while SSAR immunoreactivity was weaklydetected in only a few neurons. In contrast to SR gerbils,both immunoreactivities in the pre-seizure group were ac-cumulated in a small number of neurons. These resultssuggests that the imbalance of GABA shunt in the entorhi-nal cortex may also play an important role in regulatingneuronal activity into and out of the hippocampal com-plex during the seizure stage in the gerbil. This is becausehippocampal output fibers from CA1 and the subiculumstrongly activate deep layers of the entorhinal cortex. Thiscan then drive synchronous discharges to the superficiallayers (II and III). Layer II, which sends the major excita-tory input to the granular cell layer in the hippocampus, isunder strong local inhibitory control and layer III projectsinto the subiculum and the CA1 neurons where it evokespredominant inhibitory effects by presumably stimulatingGABA interneurons (Jones and Lambert, 1990; Empson andHeinemann, 1995). In addition, previous studies reportedthat the afferent and efferent pathways of hippocampalconnection in the SS gerbil were different from those of

248 T.-C. Kang et al. / Neurochemistry International 42 (2003) 239–249
the SR gerbil (Nitsch et al., 1994; Scotti et al., 1997). Inparticular, in the perforant path of the SS gerbil, the alteredGABAergic system may affect their functional propertiesand be instrumental for the maintenance of behavioralseizures (Scotti et al., 1997). Furthermore,Ribak and Khan(1987) demonstrated that surgical lesions of hippocampalpathway could terminate seizure activity in the SS gerbils.Therefore, our findings postulate a hypothesis that, in theentorhinal cortex, the decrease in GABA shunt enzymeimmunoreactivities may serve a malfunction to regulateneuronal activity and input signals to the hippocampus.
4.4. The roles of GABA shunt in the epileptogenesis inthe gerbil
The considered hypothesis concerning epileptogenesis,particularly in the Mongolian gerbils, is the “disinhibition”(Peterson and Ribak, 1987). This is because SS gerbils arereported to have excess GABAergic interneurons in the den-tate gyrus (Peterson et al., 1985; Peterson and Ribak, 1987)and have excess inhibitory synapses contacting GABAer-gic interneurons in the dentate gyrus of the hippocampus(Peterson et al., 1985; Farias et al., 1992). In the presentstudy, however, the alterations of GABA shunt enzymesimmunoreactivities in the GABAergic neurons followingseizure on-set suggests that the epileptogenesis in the gerbilmay not be evoked by the excessive GABA-mediated inhi-bition, but by the enhancement of GABA consumption usedas a metabolite. This hypothesis is supported by previousreports demonstrating the reduced inhibition was found inthe gerbil hippocampus after seizure on-set (Buckmasteret al., 2000), and during tetanic stimulation the efficacyof GABA-mediated inhibition of the hippocampus wasdecreased (Thompson, 1994). Therefore, the initiation ofseizure in the case of the gerbil may result from the abnor-mal GABA shunt pathway in the GABAergic neurons, butnot from hyper-inhibition of GABAergic neurons.
Acknowledgements
The authors would like to thank Mr. Suek Han for histechnical helps on this study. This work was supportedby Grant No. R01-2000-00157 from the Basic ResearchProgram of the Korea Science and Engineering Foundation,and the Hallym Academy of Sciences at Hallym University,Korea, 2002-3.
References
Armijo, J.A., Arteaga, R., Valdizan, E.M., Herranz, J.L., 1992.Co-administration of vigabatrin and valproate in children withrefractory epilepsy. Clin. Neuropharmacol. 15, 459–469.
Babb, T.L., Pretorius, J.K., Kupfer, W.R., Crandall, P.H., 1989.Glutamate decarboxylase-immunoreactive neurons are preserved inhuman epileptic hippocampus. J. Neurosci. 7, 2562–2574.
Bekenstein, J.W., Lothman, E.W., 1993. Dormancy of inhibitoryinterneurons in a model of temporal lobe epilepsy. Science 259, 97–100.
Brooks-Kayal, A.R., Shumate, M.D., Jin, H., Rikhter, T.Y., Coulter, D.A.,1998. Selective changes in single cell GABA(A) receptor subunitexpression and function in temporal lobe epilepsy. Nat. Med. 4, 1166–1172.
Buchhalter, J.R., 1993. Animal models of inherited epilepsy. Epilepsia34 (Suppl. 3), S31–41.
Buckmaster, P.S., Jongen-Relo, A.L., Davari, S.B., Wong, E.H., 2000.Testing the disinhibition hypothesis of epileptogenesis in vivo andduring spontaneous seizures. J. Neurosci. 20, 6232–6240.
Buhl, E.H., Otis, T.S., Mody, I., 1996. Zinc-induced collapse of augmentedinhibition by GABA in a temporal lobe epilepsy model. Science 271,369–373.
Cho, S.W., Hong, J.W., Lee, S.J., Choi, S.Y., 1996. Inactivation ofan NADPH-dependent succinic semialdehyde reductase byo-phthal-aldehyde. FEBS Lett. 382, 179–182.
Choi, E.Y., Park, S.Y., Jang, S.H., Song, M.S., Cho, S.W., Choi, S.Y., 1995.Production and characterization of monoclonal antibodies to bovinebrain succinic semialdehyde reductase. J. Neurochem. 64, 371–377.
Choi, E.Y., Jang, S.H., Choi, S.Y., 1996. Human brain GABA transaminaseis immunologically distinct from those of other mammalian brains.Neurochem. Int. 28, 597–600.
Choi, S.Y., Bahn, J.H., Lee, B.R., Jeon, S.G., Jang, J.S., Kim, C.K., Jin,L.H., Kim, K.H., Park, J.S., Park, J., Cho, S.W., 2001. Brain succinicsemialdehyde dehydrogenase: identification of reactive lysyl residueslabeled with pyridoxal-5′-phosphate. J. Neurochem. 76, 919–925.
De Lanerolle, N.C., Kim, J.H., Robbins, R.J., Spencer, D.D., 1989.Hippocampal interneuron loss and plasticity in human temporal lobeepilepsy. Brain Res. 495, 387–395.
During, M.J., Ryder, K.M., Spencer, D.D., 1995. Hippocampal GABAtransporter function in temporal-lobe epilepsy. Nature 376, 174–177.
Empson, R., Heinemann, U., 1995. The perforant path projection tohippocampal area CA1 in the rat hippocampal–entorhinal cortexcombined slice. J. Physiol. 4843, 707–720.
Erecinska, M., Nelson, D., Daikhin, Y., Yudkoff, M., 1996. Regulation ofGABA level in rat brain synaptosomes: fluxes through enzymes of theGABA shunt and effects of glutamate, calcium, and ketone bodies. J.Neurochem. 67, 2325–2334.
Erlander, M.G., Tobin, A.J., 1991. The structural and functionalheterogeneity of glutamic acid decarboxylase: a review. Neurochem.Res. 16, 215–226.
Erlander, M.G., Tillakaratne, N.J., Feldblum, S., Patel, N., Tobin, A.J.,1991. Two genes encode distinct glutamate decarboxylases. Neuron 7,91–100.
Esclapez, M., Hirsch, J.C., Khazipov, R., Ben-Ari, Y., Bernard, C.,1997. Operative GABAergic inhibition in hippocampal CA1 pyramidalneurons in experimental epilepsy. Proc. Natl. Acad. Sci. U.S.A. 94,12151–12156.
Farias, P.A., Low, S.Q., Peterson, G.M., Ribak, C.E., 1992. Morphologicalevidence for altered synaptic organization and structure in thehippocampal formation of seizure-sensitive gerbils. Hippocampus 2,229–245.
Feldblum, S., Ackermann, R.F., Tobin, A.J., 1990. Long-term increaseof glutamate decarboxylase mRNA in a rat model of temporal lobeepilepsy. Neuron 5, 361–371.
Feldblum, S., Erlander, M.G., Tobin, A.J., 1993. Different distributionsof GAD65 and GAD67 mRNAs suggest that the two glutamatedecarboxylases play distinctive functional roles. J. Neurosci. Res. 34,689–706.
Gale, K., Sarvey, C., Stock, J., Childs, J.A., Yalisove, B., Day, R.E.,1984. Quantitative histochemical measurement of GABA transaminase:method for evaluation of intracellular lesions produced by excitotoxicagents. Brain Res. 307, 255–262.
Gerfen, C.R., Wilson, C.J., 1996. The basal ganglia. In: Björklund,A., Hökfelt, T., Swanson, L.W. (Eds.), Handbook of Chemical

T.-C. Kang et al. / Neurochemistry International 42 (2003) 239–249 249
Neuroanatomy: Integrated Systems of the CNS. Elsevier, Amsterdam,pp. 369–466.
Giorgi, O., Meek, J.L., 1984.�-Aminobutyric acid turnover in rat striatum:effects of glutamate and kainic acid. J. Neurochem. 42, 215–220.
Gulyas, A.I., Acsady, L., Freund, T.F., 1999. Structural basis of thecholinergic and serotonergic modulation of GABAergic neurons in thehippocampus. Neurochem. Int. 34, 359–372.
Hendry, S.H.C., 1991. Delayed reduction in GABA and GAD immuno-reactivity of neurons in the adult monkey dorsal lateral geniculatenucleus following monocular deprivation or enucleation. Exp. Brain.Res. 86, 47–59.
Jones, R.S.G., Lambert, J.D.C., 1990. Entorhinal–hippocampal connec-tions: a speculative view of their function. Trends Neurosci. 16, 58–64.
Kang, T.-C., Park, S.K., Do, S.G., Suh, J.G., Jo, S.M., Oh, Y.S., Jeong,Y.G., Won, M.H., 2000a. The over-expression of somatostatin in thegerbil entorhinal cortex induced by seizure. Brain Res. 882, 55–61.
Kang, T.-C., Park, S.H., Park, S.K., Lee, J.C., Jo, S.M., Do, S.G., Suh,J.G., Oh, Y.S., Lee, J.Y., Won, M.H., 2000b. The temporal and spatialexpressions of neuropeptide Y induced by seizure in the hippocampalcomplex of gerbil. Brain Res. 870, 179–184.
Kang, T.-C., Kim, H.S., Seo, M.O., Choi, S.Y., Kwon, O.S., Baek,N.I., Lee, H.Y., Won, M.H., 2001a. The temporal alteration ofGAD67/GAD65 ratio in the gerbil hippocampal complex followingseizure. Brain Res. 920, 159–169.
Kang, T.-C., Park, S.K., Bahn, J.H., Jeon, S.G., Jo, S.M., Cho, S.W.,Choi, S.Y., Won, M.H., 2001b. The alteration of�-aminobutyric acidtransaminase expression in the gerbil hippocampus induced by seizure.Neurochem. Int. 38, 609–614.
Le Gal La Salle, G., Shen, K.F., Feldblum, S., 1984. Role of thehippocampus, amygdala and the substantia nigra in the evolution ofstatus epilepticus induced by systemic injection of kainic acid in therat. Rev. Electroencephalogr. Neurophysiol. Clin. 14, 235–240.
Loscher, W., 1984. Evidence for abnormal sensitivity of the GABA systemin gerbil with genetically determined epilepsy. In: Fariello, R.G., Lloyd,K.G., Morselli, P.L., Quesney, L.F., Eagle, J. (Eds.), Neurotransmitters,Seizures and Epilepsy. Raven Press, New York, pp. 179–188.
Loskota, W.J., Lomax, P., Rich, S.T., 1974. The gerbil as a model for thestudy of the epilepsies: seizure patterns and ontogenesis. Epilepsia 15,109–119.
Maitre, M., 1997. The�-hydroxybutyrate signalling system in brain:organization and functional implications. Prog. Neurobiol. 51, 337–361.
Marksteiner, J., Sperk, G., 1988. Concomitant increase of somatostatin,neuropeptide Y and glutamate decarboxylase in the frontal cortex ofrats with decreased seizure threshold. Neuroscience 26, 379–385.
Maru, E., Goddard, G.V., 1987. Alteration in dentate neuronal activitiesassociated with perforant path kindling. Part III. Enhancement ofsynaptic inhibition. Exp. Neurol. 96, 46–60.
Najlerahim, A., Williams, S.F., Pearson, R.C.A., Jefferys, J.G.R., 1992.Increased expression of GAD mRNA during the chronic epileptic
syndrome due to intrahippocampal tetanus toxin. Exp. Brain Res. 90,332–342.
Nitsch, C., Maly, I.P., Mori, D., Scotti, A.L., 1994. Evidence forthe co-localization of parvalbumin and glutamate, but not GABA,in the perforant path of the gerbil hippocampal formation: acombined immunocytochemical and microquantitative analysis, in theperforant path of the gerbil hippocampal formation: a combinedimmunocytochemical and microquantitative analysis. J. Neurochem.62, 1276–1284.
Obenaus, A., Esclapez, M., Houser, C.R., 1993. Loss of glutamatedecarboxylase mRNA-containing neurons in the rat dentate gyrusfollowing pilocarpine-induced seizures. J. Neurosci. 13, 4470–4485.
Paul, L.A., Fried, I., Watanabe, K., Forsythe, A.B., Scheibel, A.B., 1981.Structural correlates of seizure behavior in the Mongolian gerbil.Science 213, 924–926.
Peterson, G.M., Ribak, C.E., 1987. Hippocampus of the seizure-sensitivegerbil is a specific site for anatomical changes in the GABAergicsystem. J. Comp. Neurol. 261, 405–422.
Peterson, G.M., Ribak, C.E., Oertel, W.H., 1985. A regional increase inthe number of hippocampal GABAergic neurons and terminals in theseizure-sensitive gerbil. Brain Res. 340, 384–389.
Rempe, D.A., Bertram, E.H., Williamson, J.M., Lothman, E.W., 1997.Interneurons in area CA1 stratum radiatum and stratum oriens remainfunctionally connected to excitatory synaptic input in chronicallyepileptic animals. J. Neurophysiol. 78, 1504–1515.
Ribak, C.E., Khan, S.U., 1987. The effects of knife cuts of hippocampalpathways on epileptic activity in the seizure-sensitive gerbil. BrainRes. 418, 146–151.
Scotti, A.L., Kalt, G., Bollag, O., Nitsch, C., 1997. Parvalbumin disappearsfrom GABAergic CA1 neurons of the gerbil hippocampus with seizureonset while its presence persists in the perforant path. Brain Res. 760,109–117.
Schmid, G., Chittolini, R., Raiteri, L., Bonanno, G., 1999. Differentialeffects of zinc on native GABA(A) receptor function in rathippocampus and cerebellum. Neurochem. Int. 34, 399–405.
Sierra-Paredes, G., Galan-Valiente, J., Vazquez-Illanes, M.D., Aguilar-Veiga, E., Sierra-Marcuno, G., 2000. Effect of ionotropic glutamatereceptors antagonists on the modifications in extracellular glutamateand aspartate levels during picrotoxin seizures: a microdialysis studyin freely moving rats. Neurochem. Int. 37, 377–386.
Thompson, S.M., 1994. Modulation of inhibitory synaptic transmissionin the hippocampus. Prog. Neurobiol. 42, 575–609.
Vizi, E.S., Sperlagh, B., 1999. Separation of carrier mediated and vesicularrelease of GABA from rat brain slices. Neurochem. Int. 34, 407–413.
Wu, Y., Wang, W., Richerson, G.B., 2001. GABA transaminase inhibitioninduces spontaneous and enhances depolarization-evoked GABA effluxvia reversal of the GABA transporter. J. Neurosci. 21, 2630–2639.
Yung, K.K., Kwok, K.H., Gao, Z.G., Choi, S.Y., Kwok, F.S., 1998.Expression of GABA transaminase immunoreactivity in interneuronsof the rat neostriatum. Neurochem. Int. 33, 567–572.





























