Embed Size (px)
Citation preview

Uncorrected
Proof
1The Androgen Receptor2Coactivator-Binding Interface
3Eva Estebanez-Perpina and Robert J. Fletterick
4Abstract When hormone binds to the androgen receptor (AR), the ligand-binding5domain (LBD) becomes ordered, displaying a new protein–protein interaction6surface called AF2 (coactivator-binding pocket), which is a hydrophobic groove7that fits AR coregulators. The association of coregulators with AR LBD is often a8critical step for its transcriptional function. Existing pharmaceuticals block AR9activity by disrupting AF2 surface’s ability to recruit coactivators. Such antagonists10bind to the hormone-binding site inside the LBD core and perturb the structure of11the most terminal helix of the LBD, distorting the AF2 surface. The AF2 pocket is12also a potential candidate for pharmaceutical intervention by surface-directed small13molecules that will directly block coactivator recruitment. Such molecules may be a14novel generation of antiandrogens for treating prostate cancer.
151 Introduction
16The science addressed in this chapter is one snapshot in the cinema verite of the17androgen receptor (AR) function: the nature of the interaction between its coregu-18latory partners and the coactivator-binding pocket present in its ligand-binding19domain (LBD).20The LBD is the heart of AR not only because it binds to steroid hormones, but21because it also determines interactions with chaperones and partner transcription22factors (Prescott and Coetzee 2006) (Fig. 1). It is believed that apo-AR (receptor in23the absence of hormone), like other steroid receptors, is partly folded, and hormone24binding is the pivotal folding key that triggers the formation of coregulator interac-25tion surfaces (Feng et al. 1998). When hormone binds to the receptor, unknown26conformational changes follow that drive disassembly of the AR-chaperone com-27plex (Aranda and Pascual 2001). One key change in structure, hypothesized by
R.J. Fletterick(*)Department of Biochemistry & Biophysics, University of California San Francisco, San Francisco,CA 94143-2240, USA, E-mail: [email protected]
D. Tindall and J. Mohler (eds.), Androgen Action in Prostate Cancer,DOI: 10.1007/978-0-387-69179_4, # Springer Science + Business Media, LLC 2009
BookID 142891_Tindall_ChapID 13_Proof# 1 - 6/2/09

Uncorrected
Proof
28 analogy with other NRs (Bourguet et al. 2000; Moras and Gronemeyer 1998; Wurtz29 et al. 1996), is a conformational change of the C-terminal helix 12 (H12) present in30 its LBD. Upon agonist binding, H12 and the C-terminal 20 amino acids (F domain)31 of AR fold against the body of the LBD (Nichols et al. 1998). In its new position,32 the F domain blocks the dimer assembly site known for other NRs, and H1233 completes the hydrophobic coactivator-binding pocket (also known as AF2 site,
Fig. 1 Hypothesized AR complexes. AR domains are represented as LBD (gray oval), DBD(green oval Au3), and NTD (brown oval). AR forms different macromolecular complexes dependingon what ligand is present in its LBP. (a) In the apo-AR absence of ligand is complexed tochaperones (larger pink sphere). (b) Upon DHT binding (blue diamond), AR undergoes a seriesof conformational changes and the chaperones (HSP, smaller pink sphere) are released; then AR istargeted to the nucleus and is (c) able to form productive complexes with coactivators (i.e., p160family, yellow oval). (d) After coactivator binding to AR AF2 pocket, recruitment of the tran-scriptional machinery (blue oval) occurs with transcription or AR-dependent genes (DNA helixshown in orange). (e) When AR LBP binds antiandrogens (red lightning), a series of conforma-tional changes disrupt AF2, impeding coactivator recruitment. (f) AR-coactivator recruitment canbe blocked by AF2- interacting compounds (red star). (g) Allosteric modulators of AF2 may alsoblock AR action
E. Estebanez-Perpina, R.J. Fletterick
BookID 142891_Tindall_ChapID 13_Proof# 1 - 6/2/09

Uncorrected
Proof
34for trans-activation function 2), which is a charge-capped hydrophobic groove35(Figs. 2–4) (Darimont 2003; Estebanez-Perpina et al. 2005). This process is differ-36ent from what happens when ligands bind to enzymes, which typically possess37preformed and solvent accessible active sites. Binding of an agonist to AR com-38pletes the hydrophobic core of the receptor, reshaping its surface, which in turn
Fig. 2 Structure of AR LBD. (a) AR LBD surface is represented in gray with the residues liningthe AF-2 pocket highlighted in blue Au4. (b) Secondary structure representation of AR LBD in graywith the buried DHT shown as a blue space-filling model. The position of helices 3, 5, and 12 isshown
Fig. 3 The AR AF2 pocket is a trifurcated groove formed by residues of helices H3 (Lys717,Val713), H5 (Asn733, Val730, Met734), and H12 (Glu897, Glu893, Met894). The floor of thepocket is formed by residues Phe725, Ile737, Asn738, Ile898, and Leu712
The Androgen Receptor Coactivator-Binding Interface
BookID 142891_Tindall_ChapID 13_Proof# 1 - 6/2/09

Uncorrected
Proof
39 allows assembly of coregulators on its surfaces (Estebanez-Perpina et al. 2005;40 Hur et al. 2004).41 Both the N-terminal domain (NTD) and LBD recruit partnering proteins (Bevan42 et al. 1999; Zhou et al. 2002; McKenna et al. 1999; Hodgson et al. 2005; Lonard and43 O’Malley 2006). A curious confusion relates to competition for coactivator binding44 to the AR AF2 pocket. The AR NTD itself binds coactivators but also engages in a45 hormone-dependent interaction with its LBD, and this intramolecular interaction is46 known to regulate AR transcriptional activity by competing with coactivator47 recruitment to the LBD (He and Wilson 2002; Hsu et al. 2005). Apparently, both48 the NTD and coactivators bind to the AF2 surface (He et al. 2002). Coactivators49 may alter the self-assembly of AR and compete with AR NTD for binding to the50 AF2 groove (He et al. 2001). The NTD-LBD interaction also affects AR binding to51 chromatin (Li et al. 2006).52 Coregulators comprise coactivators and corepressors, which are structurally and53 functionally diverse proteins (Lonard and O’Malley 2006, 2007; McKenna and54 O’Malley 2002; Wang et al. 2005). Corepressors are believed to bind to the55 unliganded receptor (apo-AR) or to the antagonist-bound state, but are released56 from the receptor upon hormone binding (agonist-bound AR) (Lazar 2003; Burd57 et al. 2006). Several members of the p160 family (SRC, steroid receptor coactiva-58 tors) and ARA family (androgen-receptor-associated proteins) of coactivators are
Fig. 4 LXXLL (SRC2-3) and FXXLL (ARA70) peptides bound to AR LBD AF2 pocket. ARLBD surface is colored in pale orange Au5. (a) AR LBD surface is shown in the absence of the LXXLLpeptide. The three subsites S1, S2, and S3 are indicated. (b) Three hydrophobic residues of thehydrophobic motif of the SRC2-2 peptide, Leu 1 (L1), Leu 4 (L4), and Leu 5 (L5), are shown asblue sticks and spheres. The three leucine side chains of this biomotif interact with the correspon-dent subpockets of the AF2 groove. (c) SRC2-2 peptide is shown as blue sticks and spheres. (d)SRC2-2 peptide is shown as a blue ribbon running along the AR AF2 pocket surface. (e) AR LBDsurface is shown in the absence of the FXXLF peptide. (f) The two phenylalanines and leucine sidechains of the FXXLF biomotif (shown as green sticks and spheres) interact with the correspondentsubsites of the AF2 groove. (g) ARA70 peptide is shown as green sticks and spheres. (h) ARA70peptide is shown as a green ribbon running along the AR AF2 pocket surface
E. Estebanez-Perpina, R.J. Fletterick
BookID 142891_Tindall_ChapID 13_Proof# 1 - 6/2/09

Uncorrected
Proof
59involved in normal transcription and in the development and progression of prostate60cancer (Chmelar et al. 2007; Yeh and Chang 1996; Agoulnik et al. 2005). Once61bound to AR, coactivators are part of multiprotein complexes that include the62transcriptional machinery (Shang et al. 2002; Choudhry et al. 2006) (Fig. 1).63These multisubunit complexes contain many different enzymatic activities such64as ubiquitylation, chromatin remodeling, or DNA methylation among others65(Lonard and O’Malley 2006, 2007).66Several lines of evidence support the idea that AR AF2 pocket can be a bona fide67pharmaceutical target (Arnold et al. 2005, 2006, 2007; Estebanez-Perpina and68Fletterick 2007; Chang and McDonnell 2005). Small molecules have been reported69to bind weakly to AR AF2 by Xray crystallography and fluorescent polarization70assays (Estebanez-Perpina et al. 2007a). Such compounds would serve as starting71point chemical scaffolds to design novel antiandrogens. Such antiandrogens will be72surface directed and would work as either steric blockers of the AF2 pocket or as73allosteric modulators, reshaping the AF2 surface or adjacent putative regulatory74surfaces (Estebanez-Perpina and Fletterick 2007; Estebanez-Perpina et al. 2007a).
752 AR Coactivator-Binding Pocket
76The LBD is the best understood of AR functional domains (Table 1, Fig. 2). The77elucidation of the crystal structure of AR LBD revealed the structural determinants78of AR agonist recognition (Matias et al. 2000; Sack et al. 2001; Askew et al. 2007),79AR antagonist resistance development (Bohl et al. 2005a, b), coactivator recogni-80tion and recruitment to AR surface (Estebanez-Perpina et al. 2005; Hur et al. 2004;81He et al. 2004, 2006) (Fig. 2). There is no structure of AR LBD in either the apo-82form, in the antagonist-bound conformation, or as a part of the multidomain/full-83length AR. All AR LDB structures deposited in the Protein Data Bank (PDB) for84either wild-type or mutant AR LBD are in the agonist-bound conformation (Fig. 2).85A structure of AR DBD domain in complex with its hormone-response element has86also been reported (Shaffer et al. 2004).87Two ligand-binding sites on the AR LBD contribute to successful AR function.88First is the ligand-binding pocket (LBP), which is formally defined as an allosteric89site since it ‘‘builds’’ the active site located some distance away. The LBP contains90embedded the hydrophobic hormone: testosterone or dihydrotestosterone (DHT).91Second, the coactivator-binding pocket (AF2) located at the surface, which is the92true active site (Fig. 1). Recruitment of coactivator proteins to the AF2 pocket is a93crucial step in AR function.94The AR AF2 groove is the most well-defined solvent-exposed protein–protein95interaction site on the AR surface (Figs. 2–4). Peptides derived from specific96coactivators represent good models for studying binding of coregulators to AF2.97Several crystal structures are known for coactivator peptides bound to AR AF298pocket at atomic resolution (Estebanez-Perpina et al. 2005; Hur et al. 2004;99Askew et al. 2007; He et al. 2004, 2006). These structures reveal the molecular
The Androgen Receptor Coactivator-Binding Interface
BookID 142891_Tindall_ChapID 13_Proof# 1 - 6/2/09
Uncorrected
Proof
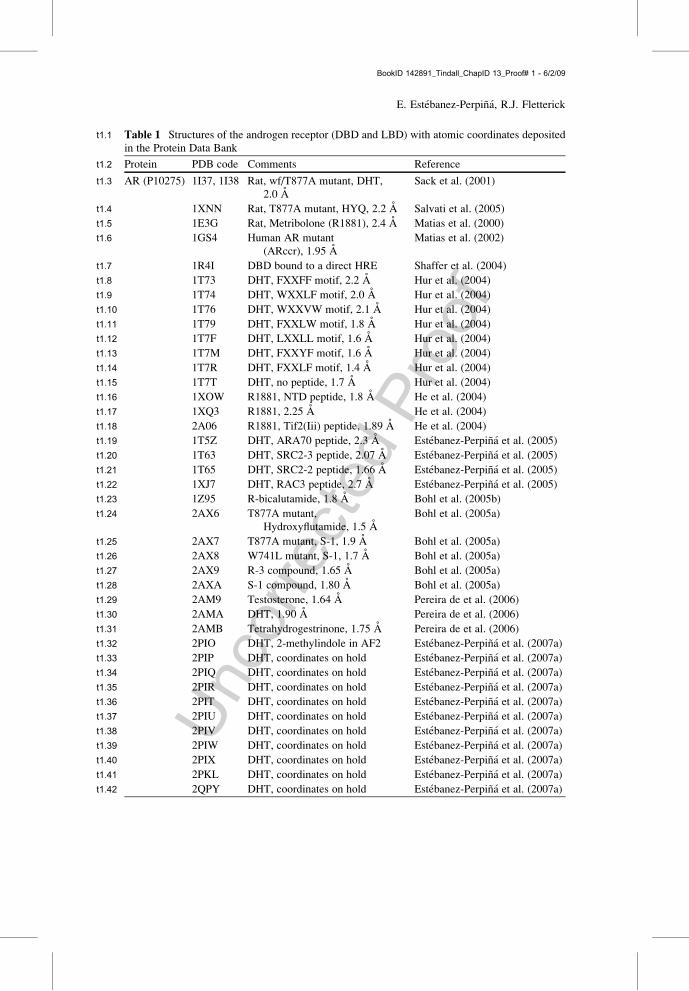
t1:1 Table 1 Structures of the androgen receptor (DBD and LBD) with atomic coordinates depositedin the Protein Data Bank
Protein PDB code Comments Referencet1:2
AR (P10275) 1I37, 1I38 Rat, wf/T877A mutant, DHT,2.0 A
Sack et al. (2001)t1:3
1XNN Rat, T877A mutant, HYQ, 2.2 A Salvati et al. (2005)t1:4
1E3G Rat, Metribolone (R1881), 2.4 A Matias et al. (2000)t1:5
1GS4 Human AR mutant(ARccr), 1.95 A
Matias et al. (2002)t1:6
1R4I DBD bound to a direct HRE Shaffer et al. (2004)t1:7
1T73 DHT, FXXFF motif, 2.2 A Hur et al. (2004)t1:8
1T74 DHT, WXXLF motif, 2.0 A Hur et al. (2004)t1:9
1T76 DHT, WXXVW motif, 2.1 A Hur et al. (2004)t1:10
1T79 DHT, FXXLW motif, 1.8 A Hur et al. (2004)t1:11
1T7F DHT, LXXLL motif, 1.6 A Hur et al. (2004)t1:12
1T7M DHT, FXXYF motif, 1.6 A Hur et al. (2004)t1:13
1T7R DHT, FXXLF motif, 1.4 A Hur et al. (2004)t1:14
1T7T DHT, no peptide, 1.7 A Hur et al. (2004)t1:15
1XOW R1881, NTD peptide, 1.8 A He et al. (2004)t1:16
1XQ3 R1881, 2.25 A He et al. (2004)t1:17
2A06 R1881, Tif2(Iii) peptide, 1.89 A He et al. (2004)t1:18
1T5Z DHT, ARA70 peptide, 2.3 A Estebanez-Perpina et al. (2005)t1:19
1T63 DHT, SRC2-3 peptide, 2.07 A Estebanez-Perpina et al. (2005)t1:20
1T65 DHT, SRC2-2 peptide, 1.66 A Estebanez-Perpina et al. (2005)t1:21
1XJ7 DHT, RAC3 peptide, 2.7 A Estebanez-Perpina et al. (2005)t1:22
1Z95 R-bicalutamide, 1.8 A Bohl et al. (2005b)t1:23
2AX6 T877A mutant,Hydroxyflutamide, 1.5 A
Bohl et al. (2005a)t1:24
2AX7 T877A mutant, S-1, 1.9 A Bohl et al. (2005a)t1:25
2AX8 W741L mutant, S-1, 1.7 A Bohl et al. (2005a)t1:26
2AX9 R-3 compound, 1.65 A Bohl et al. (2005a)t1:27
2AXA S-1 compound, 1.80 A Bohl et al. (2005a)t1:28
2AM9 Testosterone, 1.64 A Pereira de et al. (2006)t1:29
2AMA DHT, 1.90 A Pereira de et al. (2006)t1:30
2AMB Tetrahydrogestrinone, 1.75 A Pereira de et al. (2006)t1:31
2PIO DHT, 2-methylindole in AF2 Estebanez-Perpina et al. (2007a)t1:32
2PIP DHT, coordinates on hold Estebanez-Perpina et al. (2007a)t1:33
2PIQ DHT, coordinates on hold Estebanez-Perpina et al. (2007a)t1:34
2PIR DHT, coordinates on hold Estebanez-Perpina et al. (2007a)t1:35
2PIT DHT, coordinates on hold Estebanez-Perpina et al. (2007a)t1:36
2PIU DHT, coordinates on hold Estebanez-Perpina et al. (2007a)t1:37
2PIV DHT, coordinates on hold Estebanez-Perpina et al. (2007a)t1:38
2PIW DHT, coordinates on hold Estebanez-Perpina et al. (2007a)t1:39
2PIX DHT, coordinates on hold Estebanez-Perpina et al. (2007a)t1:40
2PKL DHT, coordinates on hold Estebanez-Perpina et al. (2007a)t1:41
2QPY DHT, coordinates on hold Estebanez-Perpina et al. (2007a)t1:42
E. Estebanez-Perpina, R.J. Fletterick
BookID 142891_Tindall_ChapID 13_Proof# 1 - 6/2/09

Uncorrected
Proof
100determinants of the regulatory complex formation and specificity. A crystal struc-101ture of AR LBD has also been reported with a corepressor peptide (SHP) bound to102its AF2 (Jouravel et al. 2007).103The coactivator peptides studied by Xray crystallography are of two types: the104first type comprised the actual domains or protein fragments from the family of105physiological coactivators known to interact with the AR LBD (p160 and ARA106family of coactivators) (Estebanez-Perpina et al. 2005), and the second type is107formed by libraries of peptides obtained by phage display that may mimic the108natural binding domains (Hur et al. 2004). Natural regulatory peptides usually109contain either the FxxLF motif (where F is phenylalanine, and X is any amino110acid) present in the coactivator ARA70, as well as the LxxLL motifs (where L is111leucine, and X is any amino acid) present in the p160 family member of coactiva-112tors. Peptides identified by phage display contain hydrophobic sequences with113homologous motifs have also been reported: FxxLF, FxxFF, FxxYF, FxxLW,114WxxLF, WxxVW, and LxxLL (Y and W are tyrosine and tryptophan, respectively)115(Hur et al. 2004; Hsu et al. 2003). These hydrophobic amino acid sequence motifs116are referred to as NR boxes (NR stands for Nuclear Receptor) (Fig. 4).117AR LBD is an a-helical protein with 12 helices arranged in three layers (Matias118et al. 2000; Sack et al. 2001). The AR AF2 surface is formed by residues of helices119H3, H5, and H12 (Fig. 2). The AF2 hydrophobic pocket is a trifurcated groove120formed by three moderately deep subpockets, each accommodating one of the three121hydrophobic side chains of the NR motif (F1XXL4F5 and L1XXL4L5, respectively)122(Figs. 2–4). The S1 subsite is the narrowest and deepest of the three, and it hosts the123first hydrophobic residue: Leu1/Phe1 is embedded within this subsite (Fig. 4). S1 is124lined by H5 residues M734, Q738, and H12 residues E897 and I898 (Figs. 3 and 4).125The S2 subsite is the broadest and shallowest of the three and hosts the second126hydrophobic residue: Leu4 is embedded within this subsite (Fig. 4). S2 is lined by127H3 residues K720 and V713, and H12 residue M894 (Figs. 3 and 4). The S3 subsite128is intermediate in size in comparison to the other two and quite deep. S3 is lined by129H5 residues M734 and H3 residue K720 (Figs. 3 and 4).130Both NR FXXLF and LXXLL motifs bind to the AR AF2 region as amphipathic131a-helices with hydrophobic surfaces complementary to AF2 (Fig. 4). There are two132clusters of oppositely charged residues on the AR LBD surface, at the opposite ends133of the AF2 hydrophobic groove, that assist in orienting and registering the bound134NR motifs in the AF2 groove. K720 and E897 are interesting because they function135as ‘‘helix-capping’’ residues, forming the electrostatic ‘‘charge clamp’’ residues and136stabilizing binding interactions of the FXXLF motif with the AF2 core via hydro-137gen bonding. At the start of the helix, Glu897 of AR forms hydrogen bonds to the138backbone amide (NH) of Phe1 and preceding Lys. At the end of the helix, Lys720 of139AR does not contact the carbonyl group of Leu4 or Phe5; instead, it forms hydrogen140bonds to the carbonyl groups flanking residues at positions 6 and 7. The alternative141NR-binding motif, LxxLL, interacts just like the FxxLF with Lys720, but its Leu1142residue does not contact Glu897 of AR. The structural basis of AR preference for143peptides containing FxxLF motif over the canonical LxxLL sequence has been144determined (Estebanez-Perpina et al. 2005; Hur et al. 2004; He et al. 2004).
The Androgen Receptor Coactivator-Binding Interface
BookID 142891_Tindall_ChapID 13_Proof# 1 - 6/2/09

Uncorrected
Proof
145 Structural comparisons of several AR LBDs bound by different regulatory peptides146 revealed that side chains forming the AF2 pocket rearrange to bind multiple147 variants of the NR boxes. There are specific structural features in the AF2 site of148 AR that allow accommodation of the bulkier FxxLF motif. For example, residues149 M434 and K720 of the AR LBD assume different conformations to make room for150 FxxLF peptide, allowing its interaction with I737 of AR helix H4. In general,151 coactivator peptides cause the reformable AF2 pocket to open upon binding,152 resulting in a remarkable complementary fit between the AF2 surface and the153 bound FxxLF motifs. Interestingly, both LxxLL and FxxLF containing helical154 NR peptides interact with the same set of residues on the surface of the AR LBD;155 however, these residues are engaged in different binding interactions. Furthermore,156 the major axes of the helices docked into the AF2 groove shift by more than 1 A157 between the two classes of regulatory peptides. The described studies focused on158 the central region of the interface between the regulatory peptides and the AR LBD.159 However, the interactions may be more extensive in vivo as neighboring features of160 coactivators find footholds on the AR surface. The sequences beyond the conserved161 hydrophobic motif hot spot help to confer specificity to the receptor–coactivator162 coupling (Estebanez-Perpina et al. 2005).163 Structural analysis of the corepressor peptide SHP bound to the AR AF2 region164 reveals that SHP peptide adopts a conformation similar to that observed for the165 bound LxxLL coactivator peptides (Jouravel et al. 2007). Several mutations in the166 AF2 pocket that affect Lys720, Ile737, Val730, Gln733 are implicated in prostate167 cancer and androgen-insensitivity syndromes (http://androgendb.mcgill.ca/)168 (Table 2). These mutations affect coactivator recruitment to AR. Specifically,169 mutations affecting residues Ile737 and Phe725 have been shown to disrupt the170 AR NTD/AF2 interaction without affecting hormone binding (Quigley et al. 2004).171 Many residues in the AF2 pocket are conserved among several members of the172 nuclear receptor family (Table 3). There are nuclear receptor coactivators that bind
t2:1 Table 2 Mutations at the binding pocket (LBP), and AF2 pocket associated with prostate cancerand androgen-insensitivity syndromes (AIS)
Residues Mutations in prostate cancer Mutations in AISt2:2AF2 pockett2:3
V713 Not described Not describedt2:4
V716 Not described Not describedt2:5
K717 Not described Not describedt2:6
K720 K720E Au6Not describedt2:7
R726 R726L R726Lt2:8
V730 V730M Not describedt2:9
Q733 Not described Q733Ht2:10
M734 Not described Not describedt2:11
I737 Not described I737It2:12
Q738 Not described Not describedt2:13
M894 Not described Not describedt2:14
E897 Not described Not describedt2:15
I898 Not described I898Tt2:16
E. Estebanez-Perpina, R.J. Fletterick
BookID 142891_Tindall_ChapID 13_Proof# 1 - 6/2/09
Uncorrected
Proof
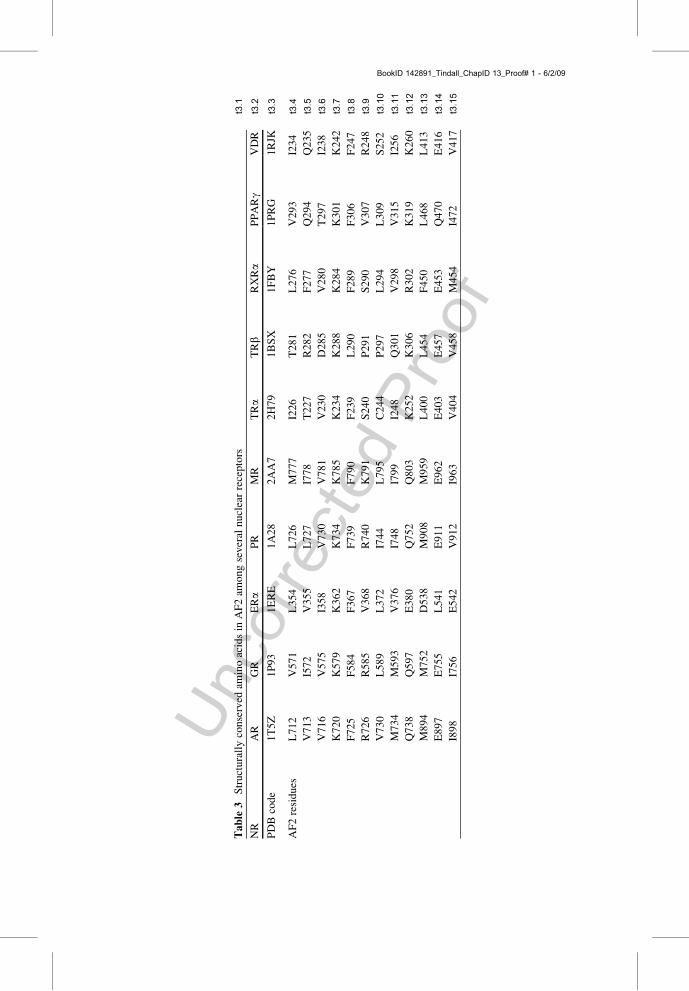
t3:1
Tab
le3
Structurallyconserved
aminoacidsin
AF2am
ongseveral
nuclearreceptors
NR
AR
GR
ERa
PR
MR
TRa
TRb
RXRa
PPARg
VDR
t3:2
PDBcode
1T5Z
1P93
1ERE
1A28
2AA7
2H79
1BSX
1FBY
1PRG
1RJK
t3:3
AF2residues
L712
V571
L354
L726
M777
I226
T281
L276
V293
I234
t3:4
V713
I572
V355
L727
I778
T227
R282
F277
Q294
Q235
t3:5
V716
V575
I358
V730
V781
V230
D285
V280
T297
I238
t3:6
K720
K579
K362
K734
K785
K234
K288
K284
K301
K242
t3:7
F725
F584
F367
F739
F790
F239
L290
F289
F306
F247
t3:8
R726
R585
V368
R740
K791
S240
P291
S290
V307
R248
t3:9
V730
L589
L372
I744
L795
C244
P297
L294
L309
S252
t3:10
M734
M593
V376
I748
I799
I248
Q301
V298
V315
I256
t3:11
Q738
Q597
E380
Q752
Q803
K252
K306
R302
K319
K260
t3:12
M894
M752
D538
M908
M959
L400
L454
F450
L468
L413
t3:13
E897
E755
L541
E911
E962
E403
E457
E453
Q470
E416
t3:14
I898
I756
E542
V912
I963
V404
V458
M454
I472
V417
t3:15
BookID 142891_Tindall_ChapID 13_Proof# 1 - 6/2/09

Uncorrected
Proof
173 to several nuclear receptors while others do not exhibit promiscuity and are174 selective to either one or another. The variety coactivator–nuclear receptor partner-175 ships increase the complexity of nuclear receptors transcriptional regulation176 (Lonard and O’Malley 2007).
177 3 The AF2 Pocket may be a Drugable Interaction Surface
178 Because the AF2 pocket plays a pivotal role in AR function, it may be a target for179 therapeutical intervention. In particular, blocking associations of AR with coregu-180 latory partners might lead to alternative treatments for prostate cancer. Compounds181 targeting the regulatory AR LBD surfaces have been proposed as alternative182 antiandrogens (Estebanez-Perpina et al. 2005, 2007a; Estebanez-Perpina and Flet-183 terick 2007; Chang and McDonnell 2005). Drugs that would bind to the AR-184 coregulator assembly site should be less susceptible to mutation of the AR protein185 by cancer cells because two proteins form the contact, and any mutation of the AF2186 surface to render the drug ineffective might also affect coregulator binding.187 Surface targeted antiandrogens directly blocking the AR coregulator assembly188 may be therapeutically useful alone or paired with the current LBP-antiandrogens.189 Small molecules would have to block AR associations by preferentially populating190 the surface normally filled by the amphipathic a-helices provided to AR by191 coregulator proteins (Estebanez-Perpina et al. 2005; Estebanez-Perpina and Fletter-192 ick 2007) (Fig. 4).193 The Au1first example of small molecules that block coactivator recruitment both in194 vitro and in vivo has been reported by Arnold et al. for the thyroid receptor b (TRb)195 (Arnold et al. 2005, 2006, 2007; Estebanez-Perpina et al. 2007b). These compounds196 (aromatic b-amino ketones (BAK)), identified in large-scale chemical screens, are197 representing a novel class of potent TRb antagonists. As the crystal structure198 revealed, these compounds bind irreversibly to one of the exposed cysteine residues199 located in TRb AF2 pocket (Estebanez-Perpina et al. 2007b). In addition, peptide200 antagonists for the AR-coactivator recruitment have been identified using phage201 display (Chang et al. 2005). The existence of peptides that target the ARAF2 pocket202 together with the discovery of TRb AF2-binding compounds set a precedent that203 could also be applied toAR for discovery of novel antiandrogens targeting its surface.204 The same research strategy that led to the discovery of TRb specific blockers205 also proved to be successful for AR, identifying its AF2 site as a pharmaceutical206 target. X ray crystallography combined with fluorescent polarization techniques207 have identified compounds that bind weakly to the AR AF2 surface. These com-208 pounds belong to different chemotypes including thyroid hormones (TRIAC and209 T3), purine analogs, and small heterocyclic molecules such as 2-methylindole.210 Most of these compounds exhibit weaker association at AF-2 site due to their211 poor fit. Subsite S1 of the AF2 that hosts F1 or L1 of the signature motif212 F1XXL4F5/L1XXL4L5 seems to be the hot spot for compounds that bind at AF2.
E. Estebanez-Perpina, R.J. Fletterick
BookID 142891_Tindall_ChapID 13_Proof# 1 - 6/2/09

Uncorrected
Proof
213Research strategies aiming at improving binding to the AF2 subsites S2 or S3 might214yield higher affinity compounds.
2154 AF2 is Allosterically Regulated by an Adjacent216Surface-Exposed Hydrophobic Pocket
217It has been observed that the AF2 pocket is reshaped not only by ligand binding to218the LBP but also by small molecules binding elsewhere on the LBD surface. Recent219structural and mutational studies have provided evidence that a novel hydrophobic220surface-exposed pocket termed BF-3 (Binding Function 3) is able to remodel the221AF2 surface and affect coactivator recruitment (Fig. 2). This surface pocket is222adjacent to the AF2 site and appears to modulate it allosterically (Estebanez-223Perpina et al. 2007a).224The BF-3 surface comprises H1 (Pro723, Phe673, and Ile672), H3–5 (Gly724225and Asn727) and H9 (Phe826, Glu829, Glu837, Arg840, and Asn833) (Fig. 2). The226crystal structures of ternary complexes of DHT-bound AR LBD with coactivator227peptides (containing either LXXLL or FXXLF motif) revealed that TRIAC (a228thyroid hormone) can bind to BF-3, causing coregulator peptides bound to AF-2292 to become disordered. Atomic resolution structural analysis revealed that binding230of TRIAC to BF-3 remodels the adjacent AF-2 pocket weakening coactivator231binding (Estebanez-Perpina et al. 2007a). Mutating residues that form BF-3 inhibits232AR function and its AF-2 activity in vitro and in vivo. Mutations at BF-3 can lead to233prostate cancer and androgen-insensitivity syndrome. Specifically, mutations of234Gln670, Ile672, and Leu830 are associated with prostate cancer (Buchanan et al.2352001a, b; Shi et al. 2002), and Leu830, Pro723, Gly724, and Arg840 are mutated in236androgen-insensitivity syndrome (http://www.androgendb.mcgill.ca/) (McPhaul2372002). Targeted mutagenesis of Asn727 and Arg840 in the BF-3 site abolished238the AR LBD activity, similar to inhibition observed with mutations in the AF-2392 region (Estebanez-Perpina et al. 2005). Curiously, the same mutations resulted in240a changed conformation of the bound TRIAC as revealed by Xray crystallography.241Similar to mutating Asn727 and Arg840, substitutions of Phe673, Pro723, Glu724,242Glu737 reduce AR activity. In contrast, mutations in the vicinity of the BF-3 site243(affecting residues Gln670, Ile672, Glu829, and Asn833) increase the AR AF-2442 activity up to fivefold (REF).245The BF-3 site could be present in other nuclear receptors. Consistent with this246speculation, part of the site, the H3–H4 loop, contains a signature sequence247(Brelivet et al. 2004). Superposition of available crystal structures reveals conser-248vation of BF-3 residues in the subfamily of steroid nuclear receptors. Mutations in249equivalent regions of estrogen and glucocorticoid receptor are shown to affect250coactivator binding (Tanenbaum et al. 1998; Milhon et al. 1997).251
252Acknowledgments We thank Elena Sablin, Debra Singer, and Leslie Cruz for their useful253comments on the manuscript Au2.
The Androgen Receptor Coactivator-Binding Interface
BookID 142891_Tindall_ChapID 13_Proof# 1 - 6/2/09

Uncorrected
Proof
254 References
255 Agoulnik I, Vaid A, Bingman WEIII, Erdeme H, Frolov A, Smith CL, Ayala G, Ittmann MM,256 Weigel NL: Role of SRC-1 in the promotion of prostate cancer cell growth and tumor257 progression. Cancer Res. 2005, 65:7976–7983.258 Aranda A, Pascual A: Nuclear hormone receptors and gene expression. Physiol Rev 2001,259 81:1269–1304.260 Arnold LA, Estebanez-Perpina E, Togashi M, Jouravel N, Shelat A, McReynolds AC, Mar E,261 Nguyen P, Baxter JD, Fletterick RJ, Webb P, Guy RK: Discovery of small molecule inhibitors262 of the interaction of thyroid hormone receptor with transcriptional coregulators. J Biol Chem263 2005, 280(52):43048–43055.264 Arnold LA, Estebanez-Perpina E, Togashi M, Shelat A, Ocasio CA, McReynolds AC, Nguyen P,265 Baxter JD, Fletterick RJ, Webb P, Guy RK: A high-throughput screening method to identify266 small molecule inhibitors of thyroid hormone receptor coactivator binding. Sci STKE 2006,267 341:13.268 Arnold L, Kosinski A, Estebanez-Perpi–a E, Robert J, Fletterick Guy RK: Inhibitors of the269 interaction of a thyroid hormone receptor and coactivators: preliminary structure-activity270 relationships. J Med Chem 2007, 50:5269–5280.271 Askew E, Gampe RT Jr, Stanley TB, Faggart JL, Wilson EM: Modulation of androgen receptor272 activation function 2 by testosterone and dihydrotestosterone. J Biol Chem 2007, 282:25801–273 25816.274 Bevan CL, Hoare S, Claessens F, Heery DM, Parker MG: The AF1 and AF2 domains of the275 androgen receptor interact with distinct regions of SRC1. Mol Cell Biol 1999, 19:8383–8392.276 Bohl CE, Miller DD, Chen J, Bell CE, Dalton JT: Structural basis for accommodation of277 nonsteroidal ligands in the androgen receptor. J Biol Chem 2005a, 280:37747–37754.278 Bohl CE, GaoW, Miller DD, Bell CE, Dalton JT: Structural basis for antagonism and resistance of279 bicalutamide in prostate cancer. Proc Natl Acad Sci USA 2005b, 102:6201–6206.280 Bourguet W, Germain P, Gronemeyer H: Nuclear receptor ligand-binding domains: three-dimen-281 sional structures, molecular interactions and pharmacological implications. Trends Pharmacol282 Sci 2000, 21:381–388.283 Brelivet Y, Kammerer S, Rochel N, Poch O, Moras D: Signature of the oligomeric behaviour of284 nuclear receptors at the sequence and structural level. EMBO Rep 2004, 5:423–429.285 Buchanan G, Yang M, Harris JM, Nahm HS, Han G, Moore N, Bentel JM, Matusik RJ, Horsfall286 DJ, Marshall VR, et al: Mutations at the boundary of the hinge and ligand binding domain287 of the androgen receptor confer increased transactivation function. Mol Endocrinol 2001a,288 15:46–56.289 Buchanan G, Greenberg NM, Scher HI, Harris JM, Marshall VR, Tilley WD: Collocation of290 androgen receptor gene mutations in prostate cancer. Clin Cancer Res 2001b, 7:1273–1281.291 Burd CJ, Morey LM, Knudsen KE: Androgen receptor corepressors and prostate cancer. Endocr292 Relat Cancer 2006, 13:979–994.293 Chang C, McDonnell DP: Androgen-receptor-cofactor interactions as targets for new drug dis-294 covery. Trends Pharmacol Sci 2005, 26:225–228.295 Chang CY, Abdo J, Hartney T, McDonnell DP: Development of peptide antagonists for the296 androgen receptor using combinatorial peptide phage display. Mol Endocrinol 2005,297 19:2478–2490.298 Chmelar R, Buchanan G, Need EF, Tilley W, Greenberg NM: Androgen receptor coregulators and299 their involvement in the development and progression of prostate cancer. Int J Cancer 2007,300 120:719–733.301 Choudhry M, Ball A, McEwan IJ: The role of the general transcription factor IIF in androgen302 receptor-dependent transcription. Mol Endocrinol 2006, 20:2052–2061.303 Darimont BD: Finding specificity within a conserved interaction site. Chem Biol 2003, 10:675–304 676.
E. Estebanez-Perpina, R.J. Fletterick
BookID 142891_Tindall_ChapID 13_Proof# 1 - 6/2/09

Uncorrected
Proof
305Estebanez-Perpina EJN, Fletterick RJ: Perspectives on designs of antiandrogens for prostate306cancer. Expert Opin Drug Discov 2007, 2:1341.307Estebanez-Perpina E, Moore JM, Mar E, Delgado-Rodrigues E, Nguyen P, Baxter JD, Buehrer308BM, Webb P, Fletterick RJ, Guy RK: The molecular mechanisms of coactivator utilization in309ligand-dependent transactivation by the androgen receptor. J Biol Chem 2005, 280:8060–8068.310Estebanez-Perpina E, Arnold AA, Nguyen P, Rodrigues ED, Mar E, Bateman R, Pallai P, Shokat311KM, Baxter JD, Guy RK, Webb P, Fletterick RJ: A surface on the androgen receptor that312allosterically regulates coactivator binding. Proc Natl Acad Sci USA 2007a, 104:16074–16079.313Estebanez-Perpina E, Arnold LA, Jouravel N, Togashi M, Blethrow J, Mar E, Nguyen P, Phillips314KJ, Baxter JD, Webb P, Guy RK, Fletterick RJ: Structural insight into the mode of action of a315direct inhibitor of coregulator binding to the thyroid hormone receptor. Mol Endocrinol.3162007b, 21:2919–2928.317Feng W, Ribeiro RC, Wagner RL, Nguyen H, Apriletti JW, Fletterick RJ, Baxter JD, Kushner PJ,318West BL: Hormone-dependent coactivator binding to a hydrophobic cleft on nuclear receptors.319Science 1998, 280:1747–1749.320He B, Wilson EM: The NH(2)-terminal and carboxyl-terminal interaction in the human androgen321receptor. Mol Genet Metab 2002, 75:293–298.322He B, Bowen NT, Minges JT, Wilson EM: Androgen-induced NH2- and COOH-terminal interac-323tion inhibits p160 coactivator recruitment by activation function 2. J Biol Chem 2001,324276:42293–42301.325He B, Lee LW, Minges JT, Wilson EM: Dependence of selective gene activation on the androgen326receptor NH2- and COOH-terminal interaction. J Biol Chem 2002, 277:25631–25639.327He B, Gampe RT Jr, Kole AJ, Hnat AT, Stanley TB, An G, Stewart EL, Kalman RI, Minges JT,328Wilson EM: Structural basis for androgen receptor interdomain and coactivator interactions329suggests a transition in nuclear receptor activation function dominance. Mol Cell 2004,33016:425–438.331He B, Gampe RT Jr, Hnat AT, Faggart JL, Minges JT, French FS, Wilson EM: Probing the332functional link between androgen receptor coactivator and ligand-binding sites in prostate333cancer and androgen insensitivity. J Biol Chem 2006, 281:6648–6663.334Hodgson M, Astapova I, Cheng S, Lee LJ, Verhoeven MC, Choi E, Balk SP, Hollenberg AN: The335androgen receptor recruits nuclear receptor CoRepressor (N-CoR) in the presence of mifepris-336tone via its N and C termini revealing a novel molecular mechanism for androgen receptor337antagonists. J Biol Chem 2005, 280:6511–6519.338Hsu CL, Yeh S, Chen YL, Ting HJ, Hu YC, Lin H, Wang X, Chang C: The use of phage display339technique for the isolation of androgen receptor interacting peptides with F/WXXLF/W and340FXXLY new signature motifs. J Biol Chem 2003, 278:23691–23698.341Hsu CL, Chen YL, Ting HJ, Lin WJ, Yang Z, Zhang Y, Wang L, Wu CT, Chang HC, Yeh S,342Pimplikar SW, Chang C: Androgen receptor (AR) NH2- and COOH-terminal interactions343result in the differential influences on the AR-mediated transactivation and cell growth. Mol344Endocrinol 2005, 19:350–361.345Hur E, Pfaff SJ, Sturgis PE, Hanne G, Buehrer BM, Fletterick RJ: Recognition and accommoda-346tion at the androgen receptor coactivator binding interface. PLoS 2004, 2:363.347Jouravel N, Sablin E, Arnold LA, Guy RK, Fletterick RJ: Interaction between the androgen348receptor and a segment of its corepressor SHP. Acta Crystallogr D Biol Crystallogr 2007,34963:1198–1200.350Lazar M: Nuclear receptor corepressors. Nucl Recept Signal. 2003, 1:e001.351Li J, Fu J, Toumazou C, Yoon HG, Wong J: A role of the amino-terminal (N) and carboxyl-352terminal (C) interaction in binding of androgen receptor to chromatin. Mol Endocrinol 2006,35320:776–785.354Lonard DM, O’Malley BW: The expanding cosmos of nuclear receptor coactivators. Cell 2006,355125:411–414.356Lonard DM, O’Malley BW: Nuclear receptor coregulators: judges, juries, and executioners of357cellular regulation. Mol Cell 2007, 27:691–700.
The Androgen Receptor Coactivator-Binding Interface
BookID 142891_Tindall_ChapID 13_Proof# 1 - 6/2/09

Uncorrected
Proof
358 Matias PM, Donner P, Coelho R, Thomaz M, Peixoto C, Macedo S, Otto N, Joschko S, Scholz P,359 Wegg A, et al: Structural evidence for ligand specificity in the binding domain of the human360 androgen receptor. Implications for pathogenic gene mutations. J Biol Chem 2000, 275:26164–361 26171.362 Matias PM, Carrondo MA, Coelho R, Thomaz M, Zhao XY,Wegg A, Crusius K, Egner U, Donner363 P: Structural basis for the glucocorticoid response in a mutant human androgen receptor (AR364 (ccr)) derived from an androgen-independent prostate cancer. J Med Chem 2002, 45:1439–365 1446.366 McKenna NJ, O’Malley BW: Minireview: nuclear receptor coactivators – an update. Endocrinol-367 ogy 2002, 143:2461–2465.368 McKenna N, Xu J, Nawaz Z, Tsai SY, Tsai MJ, O’Malley BW: Nuclear receptor coactivators:369 multiple enzymes, multiple complexes, multiple functions. J Steroid Biochem Mol Biol 1999,370 69:3–12.371 McPhaul MJ: Androgen receptor mutations and androgen insensitivity.Mol Cell Endocrinol 2002,372 198:61–67.373 Milhon J, Lee S, Kohli K, Chen D, Hong H, Stallcup MR: Identification of amino acids in the tau374 2-region of the mouse glucocorticoid receptor that contribute to hormone binding and tran-375 scriptional activation. Mol Endocrinol 1997, 11:1795–1805.376 Moras D, Gronemeyer H: The nuclear receptor ligand-binding domain: structure and function.377 Curr Opin Cell Biol 1998, 10:384–391.378 Nichols M, Rientjes JM, Stewart AF: Different positioning of the ligand-binding domain helix 12379 and the F domain of the estrogen receptor accounts for functional differences between agonists380 and antagonists. EMBO J 1998, 17:765–773.381 Pereira de J, Jesus-Tran K, Cote PL, Cantin L, Blanchet J, Labrie F, Breton R: Comparison of382 crystal structures of human androgen receptor ligand-binding domain complexed with various383 agonists reveals molecular determinants responsible for binding affinity. Protein Sci 2006,384 15:987–999.385 Prescott J, Coetzee GA: Molecular chaperones throughout the life cycle of the androgen receptor.386 Cancer Lett 2006, 231:12–19.387 Quigley C, Tan JA, He B, Zhou ZX, Mebarki F, Morel Y, Forest MG, Chatelain P, Ritzen EM,388 French FS, Wilson EM: Partial androgen insensitivity with phenotypic variation caused by389 androgen receptor mutations that disrupt activation function 2 and the NH(2)- and carboxyl-390 terminal interaction. Mech Ageing Dev 2004, 125:683–689.391 Sack JS, Kish KF, Wang C, Attar RM, Kiefer SE, An Y, Wu GY, Scheffler JE, Salvati ME,392 Krystek SR, Jr., et al: Crystallographic structures of the ligand-binding domains of the393 androgen receptor and its T877A mutant complexed with the natural agonist dihydrotestoster-394 one. Proc Natl Acad Sci USA 2001, 98:4904–4909.395 Salvati M, Balog A, Shan W, Wei DD, Pickering D, Attar RM, Geng J, Rizzo CA, Gottardis MM,396 Weinmann R, Krystek SR, Sack J, An Y, Kish K: Structure based approach to the design of397 bicyclic-1H-isoindole-1,3(2H)-dione based androgen receptor antagonists. Bioorg Med Chem398 Lett 2005, 15:271–276.399 Shaffer PL, Jivan A, Dollins DE, Claessens F, Gewirth DT: Structural basis of androgen receptor400 binding to selective androgen response elements. Proc Natl Acad Sci USA 2004, 101:4758–401 4763.402 Shang Y, Myers M, Brown M: Formation of the androgen receptor transcription complex. Mol403 Cell 2002, 9:601–610.404 Shi XB, Ma AH, Xia L, Kung HJ, de Vere White RW: Functional analysis of 44 mutant androgen405 receptors from human prostate cancer. Cancer Res 2002, 62:1496–1502.406 Tanenbaum DM, Wang Y, Williams SP, Sigler PB: Crystallographic comparison of the estrogen407 and progesterone receptor’s ligand binding domains. Proc Natl Acad Sci USA 1998, 95:5998–408 6003.409 Wang L, Hsu CL, Chang C: Androgen receptor corepressors: an overview. Prostate 2005, 63:117–410 130.
E. Estebanez-Perpina, R.J. Fletterick
BookID 142891_Tindall_ChapID 13_Proof# 1 - 6/2/09

Uncorrected
Proof
411Wurtz JM, Bourguet W, Renaud JP, Vivat V, Chambon P, Moras D, Gronemeyer H: A canonical412structure for the ligand-binding domain of nuclear receptors. Nat Struct Biol 1996, 3:206.413Yeh S, Chang C: Cloning and characterization of a specific coactivator, ARA70, for the androgen414receptor in human prostate cells. Proc Natl Acad Sci USA 1996, 93:5517–5521.415Zhou ZX, He B, Hall SH, Wilson EM, French FS: Domain interactions between coregulator ARA416(70) and the androgen receptor (AR). Mol Endocrinol 2002, 16:287–300.
The Androgen Receptor Coactivator-Binding Interface
BookID 142891_Tindall_ChapID 13_Proof# 1 - 6/2/09
Uncorrected
Proof
Author Query Form
Androgen Action in Prostate CancerChapter No.: 13
_______________________________________________________
Query Refs. Details Required Author’s response
AU1 Please check the change made to the sentence‘‘The first example elipse’’
AU2 In order to maintain consistency in this book, wehave deleted the ‘‘Abbreviations’’ list. Theexpanded forms of abbreviations are presentat the first occurrence in the text. Please check.
AU3 Colour figures are not allowed in this book.Please amend the caption of Figure 1 and itsartwork.
AU4 Colour figures are not allowed in this book.Please amend the caption of Figure 2 and itsartwork.
AU5 Colour figures are not allowed in this book.Please amend the caption of Figure 4 and itsartwork.
AU6 Please provide the significance of bold entries inTable 2.
BookID 142891_Tindall_ChapID 13_Proof# 1 - 6/2/09



![Evaluation of androgen-induced effects on the uptake of [18F]FDG, [11C]choline and [11C]acetate in an androgen-sensitive and androgen-independent prostate cancer xenograft model](https://img.pdfslide.net/doc/110x75/6361861c2fd302ae64094544/evaluation-of-androgen-induced-effects-on-the-uptake-of-18ffdg-11ccholine-and.jpg)

























