Embed Size (px)
Citation preview

HORMONES AND BEHAVIOR 7, 59-74 (1976)
The Behavioral and Somatic Effects of Ovariectomy and Replacement Therapy in Female Collared Lemmings
(Dicrostonyx groenhndicus)
JOHN F. HASLERl and E. M. BANKS
Department of Ecology, Ethology, and Evolution, University of Illiflois, Urbana, lllinois 61801
Reproductive behavior and organ weights in intact estrous and diestrous, and in ovariectomized treated and untreated female collared lemmings (Dicrostorzyr groenlmdicus) were examined. Sexual behavior of intact diestrous and untreated ovariectomized females was similar, and females in both groups behaved differently from intact estrous females in scores for lordosis and mounting the male. Receptivity was observed in some females within 3 days after the first daily dose of as little as 0.1 ug of estradiol benzoate (EB) and in 83% of all females treated with 0.1 ug or more of EB for 5 days. Vaginal perforation and cornification were related to the dose of EB and length of treatment. The doses of EB used were not related to the lordosis quotient. Ovariectomy and subsequent EB treatment influenced uterine but not preputial gland weight. Adrenal weight was not influenced by ovariectomy or EB treatment, but the increased adrenal weights observed may have been related to the daily pairing of females with males.
The endocrine control of sexual behavior in female mammals has been extensively studied (see Young, 1961; Davidson, 1972; Lisk, 1973). Most of the research in this area has focused on rodents, especially those such as the laboratory rat, guinea pig, and golden hamster, that exhibit regular estrous cycles and ovulate spontaneously. Few mammals besides the domestic rabbit and cat with induced ovulation have served as subjects for the study of the hormonal control of receptivity.
Work in our laboratory has centered on various aspects of the behavior and reproductive physiology of the collared lemming (Dicrostonyx groenlandi- CUS), an arctic microtine rodent. Females of this species are induced ovulators and exhibit long, continuous periods of estrus (Hasler, Dziuk and Banks,
lJohn F. Hasler’s present address is: Animal Reproduction Laboratory, Colorado State University, Ft. Collins, Colorado 80523.
59
Copyright @ 1976 by Academic Press, Inc. Ah rights of reproduction in any form reserved.

60 HASLER AND BANKS
1974). Banks (1968) described mating behavior in intact estrous and diestrous females of this species.
The purpose of this study was to investigate some aspects of the relationship between gonadal hormones and reproductive state in the female collared lemming. To achieve this, the influence of ovariectomy and subse- quent replacement injections of varying doses of estrogen and progesterone on both reproductive behavior and the weights of certain organs was investigated.
MATERIALS AND METHODS
The lemmings used in this study were laboratory-born descendants of animals trapped near Ft. Churchill, Manitoba in 1967. The animals were maintained at a temperature of 12 _+ 3°C on a photoperiod of 18 hr L:6 hr D. Commercial rabbit chow (Purina) was fed ad Zib. and water was supplied in suspended bottles. Further details on the laboratory maintenance of this species are available elsewhere (Hasler and Banks, 1975a).
AU females used in this study were adult virgins, 4 to 7 months of age, and weighing approximately 60 g. Ovariectomies were performed under Meto- fane inhalant anesthesia (Pitman Moore, Inc.). Ovariectomized females were not used in experiments for a minimum of 5 weeks after surgery. Behavioral data were recorded on a keyboard, magnetic tape, PDP/8 system (White, 1971).
Eight sexually experienced males 6 to 10 months of age were used in the experiments. Previous work by Banks (1968) indicated a large degree of individual variation in male sexual behavior. Therefore, the same eight proven males were used in behavior tests with females in all experimental groups. Tests were conducted between 0900 and 1200 hr in light.
Dyadic encounters between males and females were staged in a semi- circular arena constructed of galvanized tin, a plywood floor, and a front wall of glass. The arena was 50 cm in diameter. One pair of lemmings was tested at a time. Prior to testing, a male and female were introduced into the arena and were kept separated for 5 minutes by a cardboard partition. Data recording commenced with the removal of the cardboard separator. A record was made of the frequency, duration and latency (to the nearest 0.1 set) of six male and female behaviors.
Behavior of Males
Autogroom-Self-licking or self-scratching with either the hindlegs or forelegs. Areas groomed included the perineal or genital region. Allogroom-Licking or scratching any part of the female’s body. Attack-Attempt to bite females.

LEMMING SEXUAL BEHAVIOR 61
Chase-Followed female when she moved away. Mount-Approached female, raised forelegs, and gripped pelvic region. Ejaculation-A vigorous series of thrusts performed while mounted. After ejaculation the male often rolled partly over on his side as the female moved away. Evidence for ejaculation was provided by the presence of the copulation plug.
Behavior of Females
Autogroom-Same as for male. Allogroom-Same as for male. Attack-Same as for male. Sparring-Forefeet pushed against the male’s neck and shoulders, as he extended his forefeet against female’s neck and shoulders. This upright behavior was performed simultaneously by both members of a pair and is listed as a female behavior for the sake of convenience. Mount-Approached male, raised forelegs, and placed her chest and forelegs on any portion of the male’s trunk. Lordosis-Female stationary, head raised at an angle of approximately 45” from vertical, back arched concavely, and vulvar region exposed. Lordosis sometimes occurred when the female was approached or touched, but not mounted by the male. Prior to behavioral testing, a vaginal smear was obtained by the lavage
method from all females with a perforate vagina. Smears were examined wet, unstained at 100X and an estimate was made of the relative abundance of the different cell types.
After the final behavioral test, females were killed by cervical disloca- tion, weighed to the nearest 0.1 g and fixed in acetic-formol-alcohol (AFA) for 48 hr and stored in 70% ethanol. Later, ovaries (in intact females), uteri, preputial glands, and adrenals were dissected, cleaned of fat, blotted on paper, and weighed to the nearest 0.1 mg.
Specific details of three experiments comprising this study follow.
Experiment I
Procedures. The study was designed to provide baseline information on behavior and organ weights for comparison with hormonally-treated females. Three groups of eight females were used: (1) natural estrus; (2) natural diestrus; and (3) ovariectomized. Each estrous female was paired with a male, and behavioral recordings were made until ejaculation occurred (usually within 10 minutes). Behavioral interactions were recorded for 5 minutes for each female in groups 2 an d 3. Each group was tested once with the same males, on a different day.

62 HASLER AND BANKS
Results. The scores for the behavioral recordings of estrous, diestrous, and untreated ovariectomized females paired with males are shown in Table 1. The duration of the behavioral tests of estrous females varied because testing continued until ejaculation occurred. Consequently, scores during each test period for all behaviors except ejaculation were calculated on the basis of a
TABLE 1
Mean Scores (+ SE) of Behaviors Performed during 5-Min Tests by Estrous (E), Diestrous (D), and Untreated Ovariectomized (Ovx) Females and Their Mates
Estrus Pa
(n = 8) (E vs. D)
Female behaviors
Autogroom Fb 0.3 r 0.1 D 1.5 j: 1.1
Allogroom F 0.7 f. 0.4 D 1.0 ?r 0.1
Attack F 0.7 * 0.3 D 0.5 t 0.2
Spar F 19.0 2 3.2 D 29.3 + 4.5
Mount F 2.5 r 1.1 ** D 2.6 + 1.1 **
Lordosis F 14.3 + 2.6 *** D 22.4 ? 4.1 ***
Male behaviors
Autogroom F 3.1 f 1.0 ** D 8.7 -t 2.2 **
Allogroom F 2.8 t 1.6 D 6.3 f 4.0
Attack F 0.2 r 0.2 D 0.2 I! 0.1
Chase F 6.6 k 2.4 D 7.9 f 1.4 *
Mount F 15.4 i 2.8 ** D 24.5 + 4.1 ***
Ejaculate F 1.0 + 0.0 ***
D 3.4 A 0.2 ***
-
State of female
Diestrus P ovx P
(n = 8) (D vs. Ovx) (n = 8) (Ovx vs. E)
0.5 2 0.4 0.4 + 0.3 0.5 t 0.2 1.1 * 0.6 0.5 -f 0.2 0.3 I! 0.2
16.9 ? 2.7 51.8 r 11.1
0 0 0 0
0.7? 0.3 1.9 + 0.8 3.6 + 1.6 8.2 k 4.3 1.4 t 0.8 0.7 f 0.4
14.0 k 4.5 18.2 + 6.1 *
6.9 + 2.5 5.0 f 1.4 0 0
0.3? 0.1 0.9 t 0.3 0.6 ie 0.1 1.2 f 0.4 0.8 f 0.3 0.7 t 0.2
17.7 + 3.8 43.6 i 12.0
0.2 2 0.1 0.1 f 0.1 0 0
1.6 i- 0.5 7.0 f 2.9 1.2 i: 0.6 3.6 i 2.0 0.4 5 0.4 0.4 f 0.4 8.0 2 2.0 7.9 + 2.4 6.1 * 3.7 6.1 t 3.7 0 0
* * *** ***
*
*** *** *** ***
a(p is based on Mann-Whitney U test. Siegel, 1956). bF = frequency, D = duration (set). *p < .05.
**p < .Ol. ***p < .OOl.

LEMMING SEXUAL BEHAVIOR 63
single 5-min period in order to compare them to the scores of the diestrous and ovariectomized groups (i.e., scores longer or shorter than 5 minutes were calculated on a per 5-min basis). The Kruskal Wallis one-way analysis of variance test (Siegel, 1956) was performed on all behaviors among the three treatments. Mann-Whitney U tests were performed when significant differ- ences were found in any behaviors among the treatments.
There were no significant differences among the three groups in the following behaviors recorded for the females: autogrooming, allogrooming, attack, and sparring. However, estrous females mounted males significantly more often and for a longer total duration than did either diestrous or ovariectomized females. In addition, lordosis behavior was observed only in estrous females.
As indicated in Table 1, there was more frequent autogrooming by males when paired with estrous females than when the same males were paired with either diestrous or ovariectomized females. The duration of autogrooming was longer when the males were paired with estrous females compared to diestrous females, but not significantly longer than with ovariectomized females. Although the site of autogrooming was not categorized, males paired with estrous females more frequently groomed their genitalia than other body areas. Some males licked the genital area after almost every mount or intromission. Genital grooming was rarely observed when males were paired with females not in estrus. Males mounted estrous females significantly more often and for a longer duration than either diestrous or ovariectomized females. However, males spent significantly more time chasing diestrous females than either of the other two categories of females. Ejaculation occurred only when males were paired with estrous females.
Experiment 2
Procedure. This experiment was designed to show the influence of various doses of estrogen and progesterone on behavior and on certain organ weights of ovariectomized females. Ovariectomized females were randomly divided into six groups, and each group was tested separately during a different week with the same males. Each female in the six groups listed in Table 2 was paired with a male for 5 min on the first day of testing (Day 0): immediately thereafter she received her first injection. Behavioral testing followed by an injection was repeated on 5 successive days and, on the sixth day after behavioral testing, the females were killed and their organs prepared as described above. Each female was paired with the same male every day. Hormones were administered subcutaneously in 0.05 ml corn oil. Nonreceptive females were tested behaviorally with a male for 5 min. If during the first 5 min of a test, lordosis occurred when a female was mounted, the test was continued until ejaculation took place.

64 HASLER AND BANKS
TABLE 2
Percentage of Females Receptive on Different Days of Hormone Treatment
Percentage of females receptive
Number of days of treatment
States of females Treatment0 NO12 3 4 5
Estrus Diestrus Ovariectomized Ovariectomized Ovariectomized Ovariectomized Ovariectomized Ovariectomized
Ovariectomtied
None None None
10.0 pg EBb 1.0 p.g EBb 0.1 /qg EBb 0.01 pg EBb 0.1 pg EBb plus
250.0 pg ProgsC corn oil (0.05 ml)
8 100 8 0 8 0 8 0 0 38 38 SO 75 8 0 0 25 50 75 75 8 0 0 25 75 88 100 8 00 0 0 0 0
5 0 0 20 60 80 100 8 00 0 0 0 0
aFemales were injected once daily for 5 days (Days O-4). The group injected with progesterone was injected with EB daily for 5 days and progesterone on Days 4 and 5, 4 hr prior to behavioral testing.
bEB = estradiol benzoate, Sigma Chemical Co. CProg. = progesterone, Eli Lilly and Co.
Results. The major goal of the behavioral recordings was to determine how closely hormonally-induced receptivity compared to natural estrous behavior. Statistical comparisons (Mann-Whitney U test, Siegel, 1956) were made of the behaviors of all females achieving receptivity and their mates at the end of 5 days of the various treatments. All behavioral scores were calculated on a per 5-min basis, as explained above. Females were defined as receptive if they exhibited lordosis any time during a S-min test when mounted by the male. Exogenous estrogen produced an effect on female behavior. Table 2 shows the effectiveness of the different hormone treatments on the induction of receptivity. Receptivity was first noted in some females within 48 hr after the first treatment with doses of 0.1, 1.0, or 10.0 pg estradiol benzoate (EB). A daily dose of 0.1 pg EB for 5 days or the same dose of EB for 5 days with 25Opg progesterone injected on Days 4 and 5 were the two treatments resulting in the highest percentage (100%) of receptive females.
A lordosis quotient (LQ) was calculated for each receptive female. (LQ = number of times lordosis exhibited per number of mounts by male X 100). The LQ was calculated for each test period from the time of pairing to ejaculation. The LQs of individual females in all treatment groups ranged from 46 to 126. An LQ of more than 100 indicated that a female exhibited

LEMMING SEXUAL BEHAVIOR 65
lordosis more times than she was mounted. Some of these females were found in every treatment group and in the natural estrous group. Comparisons were made among treatment groups by a one-way analysis of variance of the LQs of the receptive females on Days 3 and 5 of the injection regime. The mean i SE LQs of receptive females on Day 5 were as follows: natural estrus, 89.6 + 7.6; 10.0 pg EB, 76.1 + 6.7; 1.0 /~g EB, 87.5 + 5.4; 0.1 pg EB, 91.2 + 6.8; 0.1 pg EB + Prog., 77.7 + 6.4. There were no significant differences among the LQs for any of the treatments on Days 3 or 5, or the group in natural estrus.
Nine of the 12 behaviors recorded for receptive females in all treatment groups and their mates were not significantly different in frequency or duration from those of females in natural estrus observed in Experiment 1. However, there were significant differences in the frequency and duration of sparring, male allogrooming, and male chasing. These differences were similar to those shown by the same females when they were nonreceptive, prior to the first hormone injection. There was a decrease in sparring and an increase in allogrooming by the males (Table 3). In addition, there was a decrease in
TABLE 3
Mean (+ SE) Scores of Sparring, Allogrooming, and Chasing by Males When Paired with Females in Natural Estrus or
Hormonally-Treated, Receptive Ovariectomized Females
Behaviors
Treatment of females IV Sparring Allogrooming Chasing
Estrus 8 Fa 19.0 + 3.2 2.8 i 1.6 6.6 i 2.4 D 29.3 + 4.5 6.3 i 4.0 7.9 It 1.4
Ovariectomized Pb P P 10.0 & EBC 6F 7.8 f 0.9 ** 4.1 c 1.3 2.6 + 0.7 +**
D 15.7 A 3.0 * 20.5 -t 8.2 ** 3.6 + 1.1 ** 1.0 pg EBc 6F 5.9 i 1.7 *** 4.3 + 0.5 0.7 i- 0.2 ***
D 18.9 f 2.9 * 29.0 + 4.9 ** 5.0 + 2.5 0.1 pg EBc 8F 5.9 k 0.8 *** 9.5 f 2.9 * 1.4 + 0.5 ***
D 9.1 + 2.4 *** 27.3 z 9.5 * 2.1 2 0.7 *** 0.1 ng EBc plus 5F 6.0 f 1.4 * 7.2t2.1 * 3.2 k 1.4 **
250.0 pg Prog. D 11.4r2.3 * 22.0 f 5.4 ** 4.7 + 1.8
OF = frequency, D = duration (set). bsignificance levels of hormone treated groups vs. estrous group based on Mann-
Whitney U test, Siegel, 1956. CTreatment as described in Table 2. *p < .025.
**p < .Ol. ***p < .OOl.

66 HASLER AND BANKS
chasing by the males. The behaviors of males and females in the groups in which no females became receptive (0.01 E.cg EB and corn oil) were not significantly different from scores of the same animals prior to the first injection. Each of these groups showed a decrease in sparring and an increase in male allogrooming when compared to the first group of untreated ovariectomized females in Experiment t.
Table 4 shows daily changes for the different treatment groups in the percentage of females with perforate vaginae and cornified vaginal smears. Females from Experiment 1 are included for comparison. All females in natural estrus were perforate and had highly cornified vaginal smears. AU diestrous females selected for the experiment were imperforate. Previous work (Hasler and Banks, 1975b), showed that both imperforate and perforate females with a vaginal smear composed of leucocytes were nonreceptive and in a diestrous state.
A total of 9% (5/53) of the untreated ovariectomized females listed in Table 4 had perforate vaginae. Daily injections of between 0.01 and l.O,ug EB induced vaginal perforation in some females 24 to 48 hr after the first injection. All females injected with 0.1 fig EB or more exhibited vaginal cornification 96 hr after the first injection. There was a latency of approxi- mately 24 hr between the first injection of EB and the appearance of vaginal cornification. Although daily injections of 0.01 1.18 EB induced vaginal perforation in 75% of females, there was no apparent effect on vaginal cornification.
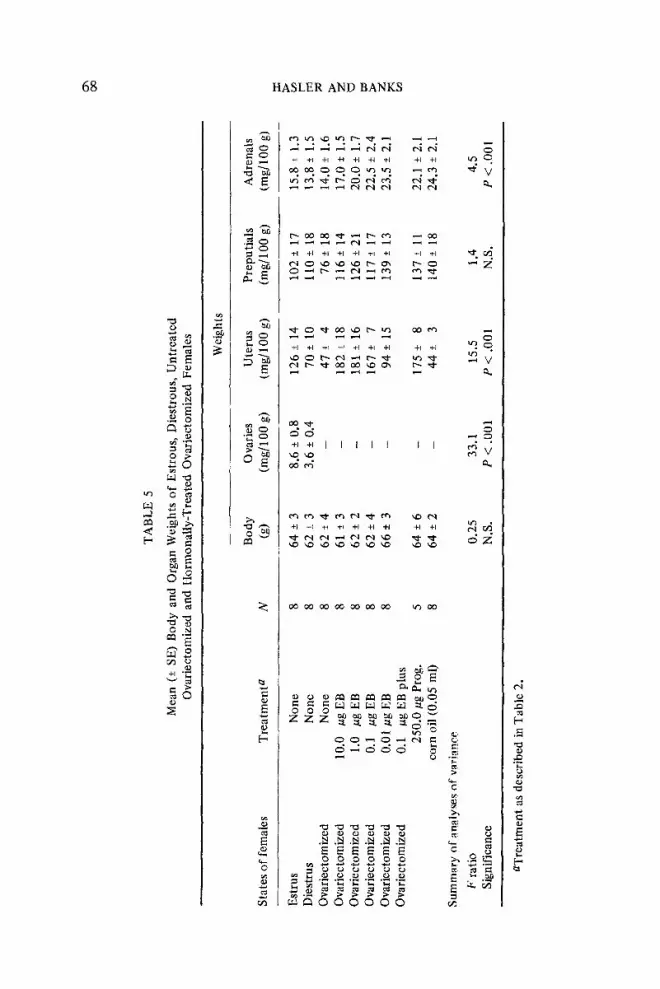
The body and organ weights of females from Experiments 1 and 2 are shown in Table 5. One-way analyses of variance were performed on the body and organ weights to determine if there were significant differences between experimental groups. Student’s t tests were performed when significant differences were found by one-way analysis of variance.
There were no significant differences in body weights among any of the groups. However, since there was a wide range of variation in body weight within each group, organ weights were calculated on a mg/lOO g of body weight basis. Ovaries from estrous females were significantly heavier (p < .OOl) than ovaries from diestrous females. The uteri of estrous females were significantly heavier @< -00.5) than the uteri of diestrous females, which, in turn, were significantly heavier (p < .05) than the uteri of untreated ovariecto- mized females. Females treated with five daily doses of 10.0, 1.0, or 0.1 pg EB or 0.1 E.cg EB plus 250 pg progesterone had significantly heavier uteri (p < .025) than females in natural estrus. There was no significant difference in the uterine weights of estrous females and females treated with 0.01 pg EB.
There were no significant differences in the weights of the preputial glands of the females in any of the groups. In addition, there were no significant differences in adrenal weights among estrous, diestrous, untreated ovariectomized females and females injected with lO.Opg EB. However,

TABL
E 4
Perc
enta
ge o
f Fe
mal
es w
ith C
orni
fied
Vagi
nal
Smea
rs a
nd P
erfo
rate
Vag
inae
on
Diff
eren
t D
ays
of H
orm
one
Trea
tmen
t0
Perc
ent
of fe
mal
es:
corn
ified
/per
fora
te
Num
ber
of d
ays
of t
reat
men
t E
Stat
e of
fem
ales
2
Trea
tmen
t b
N
0 1
2 3
4 5
z n E
stru
s N
one
8 lo
o/lo
o K
Die
stru
s N
one
8 o/
o O
varie
ctom
ized
N
one
8 01
0 E
Ova
riect
omiz
ed
10.0
~(
g EB
8 O
/25
o/75
88
/88
loo/
loo
Ova
riect
omiz
ed
loo/
loo
100/
100
; 1.
0 pg
EB
8 o/
13
O/3
8 50
175
loo/
loo
loo/
loo
loo/
loo
g O
varie
ctom
ized
0.
1 pg
EB
8 o/
o O
/25
25/2
5 63
175
Ova
riect
omiz
ed
100/
100
100/
100
5 0.
01 p
g EB
8
O/l
3 o/
25
1313
8 13
150
13/5
0 O
varie
ctom
ized
0.
1 pg
EB
plus
13
175
* 2 25
0.0
pg P
rog.
5
o/o
O/2
0 60
160
SO/S
O
Ova
riect
omiz
ed
loo/
loo
~00/
100
corn
oil
(0.0
5 m
l) 8
O/l
3 o/
13
o/13
o/
13
o/13
o/
13
aVag
insl
sm
ear c
ompo
sed
of >
50%
~ cor
nifie
d ce
lls.
bTre
atm
ent
as d
escr
ibed
in T
able
2.
TABL
E 5
Mea
n (+
SE)
Bod
y an
d O
rgan
Wei
ghts
of
Est
rous
, D
iest
rous
, U
ntre
ated
O
varie
ctom
ized
an
d H
arm
onal
ly-T
reat
ed
Ova
riect
omiz
ed
Fem
ales
Wei
ghts
Stat
es o
f fe
mal
es
Trea
tmen
t0
Body
C
d -
Est
rus
Die
stru
s O
varie
ctom
ized
O
varie
ctom
ized
O
varie
ctom
ized
O
varie
ctom
ized
O
varie
ctom
ized
O
varie
ctom
ized
Non
e N
one
Non
e 10
.0
fig E
B 1.
0 jig
EB
0.1
fig E
B 0.
01 p
g EB
0.
1 jg
EB
plus
25
0.0
pg P
rog.
co
rn o
il (0
.05
ml)
Sum
mar
y of
ana
lyse
s of
var
ianc
e F
ratio
Si
gnifi
canc
e
5 64
f 6
8
64 f
2
64 f
3
62 +
3
62 +
4
61 i
3 62
t 2
62
f 4
66
k 3
0.25
N
.S.
Ova
ries
G-4
1 00
d
[Jte
rus
(w/l0
0 8)
Pr
eput
ials
(m
g/lO
O 9
)
8.6
* 0.
8 12
6 +
14
3.6
t 0.
4 70
+ 1
0 -
47%
4
- 18
2 +
18
- 18
1 +
16
- 16
7 +
7 -
94?
15
- 17
5+
8 -
44+
3
33.1
P
< .O
Ol
15.5
P
< .O
Ol
102
f. 17
11
0 f
18
76 +
18
116~
14
12
6+21
11
7 I!
17
139
t 13
137
+ 11
14
0 r
18
1.4
N.S
.
Adre
nals
(w
/l 00
s)
15.8
f 1
.3
F
13.8
+_ 1
.5
F m
14.0
+ 1
.6
P
17.0
f 1
.5
>
20.0
t
1.7
3
22.5
f 2
.4
$ 23
.5 I
+ 2.1
22.1
f 2
.1
24.3
k 2
.1
4.5
P<.O
Ol
aTre
atm
ent
as d
escr
ibed
in T
able
2.

LEMMING SEXUAL BEHAVIOR 69
females in all of the other treatment groups had adrenals which were significantly heavier Cp < .Ol) than the adrenals of the estrous, diestrous, and untreated ovariectomized females. Females which received 10.0 pg EB had significantly lighter adrenals (p < .02) than all other injected females except those which received l.Opg EB.
Experiment 3
Procedure. In an attempt to clarify further some of the changes in organ weights observed in Experiment 2, another group of virgin females was randomly divided into four groups. One group was sham-ovariectomized, and the other three groups were ovariectomized. Ten weeks after surgery the four groups were treated once daily for 5 consecutive days as shown in Table 6. After the fifth day of treatment, vaginal smears were obtained from all perforate females and they were killed, weighed and their organs prepared as described above.
Results. The mean body and organ weights of the females in Experi- ment 3 are listed in Table 6. One-way analyses of variance revealed no significant differences among groups for any organs except uterine weights. Uterine weights of intact, sham-ovariectomized females and estrogen-injected females were not significantly different but both groups had significantly heavier uteri 0, < .02) according to Student’s t test than did the oil-injected or the untreated ovariectomized females.
TABLE 6
Mean (? SE) Body and Organ Weights of Sham-Ovariectomized, Untreated Ovariectomized and Hormonally-Treated Ovariectomized Females
Weights
Treatment Body Uterus Preputials Adrenals
N (9) (w/l00 g) (mg/lOO d (mg/lOO g)
Sham-ovariectomized 10 59 f 5 102 i- 20 84i 12 14.3 i 1.1 Ovariect. 1.0 EBQ pg 7 65 f 5 144 t 25 126 ?r 20 15.4 f. 1.8 Ovariect. corn oil (.05 ml) 7 62 +_ 4 46~ 7 109 i 14 14.6 + 1.2 Ovariect. no treatment 6 59 f 6 51i 8 96+- 17 17.7 f 1.8
Summary of analyses of variance
F ratio 0.75 8.2 2.3 0.5 Significance N.S. P < ,001 N.S. N.S.
QEB and oil-injected females were injected once daily for 5 days and all females were killed on the sbth day, 10 weeks postovariectomy.

70 HASLER AND BANKS
DISCUSSION
Most of the reproductive behaviors of lemming pairs in which the females were in natural estrus or diestrus were comparable in temporal frequency and duration to those reported by Banks (1968) for the same species. However, there were several noteworthy differences. We observed that males spent significantly more time chasing diestrous than estrous females. Banks (1968) did not record chasing by the male, but he did report that avoidance of the male was significantly more frequent and of longer duration by diestrous than estrous females. He also reported more male perineal investigation of diestrous than of estrous females. In the present study, we did not distinguish perineal investigation from allogrooming of any other part of the female’s body, and we found no significant difference in the frequency and duration of male allogrooming of estrous and diestrous females. Banks (1968) observed more frequent and longer bouts of aggression by both estrous and diestrous females than were observed in the present study. The difference in the two studies may be due to the fact that Banks used sexually experienced females, whereas females in the present study were virgins.
Many studies of the influence of hormones on reproductive behavior in the female laboratory rat have not mentioned aggression by females. Hardy (1972), however, reported that diestrous females of the Sprague-Dawley strain often attacked the male with their teeth. Estrous females were not aggressive. The female golden hamster has been reported to be very aggressive toward males while in a diestrous state, but not while in estrus (Kislak and Beach, 1955). Although it is difficult to compare two species in different studies, it appears that the female collared lemming is considerably less aggressive toward the male than is the nonreceptive female golden hamster. It should be noted that female lemmings in this study were smaller than males. This is in contrast to the hamster where the female is the larger of the sexes. Females in all of the groups in the present study averaged less than one aggressive interaction per 5-min test. Observations on other females in our colony, however, revealed that even during estrus, older, multiparous females tended to be far more aggressive than the virgin females used in this study. Some of the possible implications of the similar levels of aggression in estrous and diestrous collared lemmings were discussed by Banks (1968).
As was shown for the collared lemming in the present study, ovariecto- my of the female hamster (Vandenbergh, 1971) was reported as not lowering the level of aggression. However, another laboratory (Payne and Swanson, 1971, 1972) reported that ovariectomy in the hamster significantly reduced aggression.
Rarely has the influence of hormones on sexual behavior in rodents been characterized by any index other than the lordosis quotient. However, Hardy and DeBold (1971) showed that laboratory rats, hormonally induced

LEhfMING SEXUAL BEHAVIOR 71
into receptivity, differed significantly from females in natural estrus in some behaviors other than the LQ score. Hardy and DeBold measured the duration and intensity of lordosis, in addition to ear-wiggling and hopping by the female. In the present study, with a few exceptions, mounting of the male was only observed in females which also exhibited lordosis. In the guinea pig, mounting of the male is also an index of receptivity and has been shown to be exhibited earlier in the estrous cycle than lordosis (Young and Roundlett, 1939).
In the present study we found -significant differences in some of the behaviors recorded for the hormonally-treated females compared to females in natural estrus. There are several possible explanations for the observed differences. The behaviors which differed temporally from the natural estrous group were all probably initiated by the males. Behaviors that differed were sparring, male allogrooming, and male chasing. Week by week scores for male allogrooming and sparring had also changed in the different groups of ovariectomized females prior to hormone treatment. Since the hormone- treated groups were tested as much as 6 weeks after the estrous group, a change in male behavior may have occurred because of the frequency with which they were paired with females. Thus it appears that male allogrooming and sparring changed with time, independent of the treatment of the females. However, the possibility that the observed changes in male behavior were induced by some kind of physiological or behavioral change in the ovariecto- mized females with time and independent of treatment cannot be ruled out. The data on adrenal weights in the hormonally-treated females indicated that a change occurred in these females independent of treatment.
We suggest that the receptivity observed in females injected with between 0.1 and 10.0 pg EB or 0.1 pg EB plus 250 pg progesterone was comparable to that of females in natural estrus. There were no significant differences in LQ, mean lordosis duration, or in the duration and frequency of the following female behaviors: mounting, aggression, autogrooming and allogrooming. Uterine weights from ovariectomized females in this study reflected only roughly the dose of EB administered.
Adrenal weights of females in most of the hormonally and oil-injected, ovariectomized groups in Experiment 2 were surprisingly high. These weights were independent of treatment. In contrast, exogenous estrogen increased adrenal weights in ovariectomized female laboratory rats (Kitay, 1963) and golden hamsters (Gaskin and Kitay, 1970). The data from Experiment 2 of the present study suggest the possibility of adrenal hypertrophy with time following ovariectomy. Experiment 3 was designed to clarify the cause of the increased adrenal weights observed in Experiment 2. However, we found no significant differences between adrenal weights from intact and ovariectomized females which were either EB-injected, oil-injected, or not treated daily for 5 days 10 weeks after ovariectomy. The only planned difference in the design of

72 HASLER AND BANKS
Experiment 2 and 3 was that females in Experiment 2 were exposed to males daily for 6 days. This exposure may have affected the observed adrenal hypertrophy. However, this hypothesis has not been tested.
The most sensitive response to EB treatment was vaginal perforation. In most female lemmings, ovariectomy resulted in the closing of the vagina with an epithelial plate. Subsequent EB treatment resulted in a rapid opening of the plate. This response occurred more rapidly and in response to a lower dose of EB than did vaginal cornification. Nevertheless, vaginal cornification occurred more quickly in response to EB than did behavioral receptivity, a condition similar to that reported for the laboratory rat (Davidson et al., 1968) and mouse (Mirskaia and Crew, 1930). In addition, the minimal dose of EB that induced vaginal cornification in the lemming was comparable (on a body weight basis) to that reported for the laboratory rat (Davidson, et al., 1968; Whalen and Hardy, 1970).
Behavioral receptivity (in the collared lemming) as measured by the LQ seemed to be an all or none phenomenon in the present study. However, it is possible that a different dosage range of EB might have revealed changes in LQ. Hormonally-treated females either never exhibited a lordosis response to male mounting or they showed a frequency of response that was not significantly different from females in natural estrus. In contrast, a graded series of estrogen doses in laboratory rats has been shown to result in a graded LQ response (Boling and Blandau, 1939; Davidson et al., 1968). In the domestic rabbit (McDonald, Vidal, and Beyer, 1970), estrogen dosage influenced the percentage of females becoming receptive and the duration of receptivity, but not the latency to receptivity or the LQ.
Pilot studies in our laboratory indicated that progesterone injected after a dose of estrogen which was, itself, insufficient to induce receptivity would not induce receptivity. In addition, progesterone did not facilitate the receptivity observed in females treated with higher doses of EB. Therefore, the present study concentrated on investigating the influence of various doses of EB administered alone.
Some studies have indicated that estrogen alone will induce receptivity in female laboratory rats (Davidson et al., 1968; Edwards, Whalen, and Nadler, 1968). However, with physiological doses of estrogen, an extended period of treatment is usually necessary, and most investigators agree that progesterone injected after pretreatment with estrogen facilitates the induction of receptivi- ty. Measurement of circulating hormone concentrations has shown that progesterone rises in response to the preovulatory peak of LH just prior to the onset of receptivity in the laboratory rat (Uchida, Kadowaki, and Miyake, 1969) and in the golden hamster (Leavitt and Blaha, 1970; Ciaccio and Lisk, 1971). Since the collared lemming is an induced ovulator and exhibits long periods of constant estrus (Hasler et al., 1974), it is unlikely that receptivity is triggered by a cyclic change in the level of progesterone. The LH peak which

LEMMING SEXUAL BEHAVIOR 73
instigates a rise in progesterone in rodents with an estrous cycle would not be expected to occur in the lemming until after copulation. Furthermore, except in female lemmings in postpartum estrus, there would be no corpora lutea present as a possible source of progesterone. We suggest that a constant level of estrogen, with little or no influence of progesterone, maintains receptivity in this species. It has been demonstrated that physiological doses of estrogen alone will induce receptivity in ovariectomized cats (Whalen and Hardy, 1970) and domestic rabbits (McDonald et al., 1970).
ACKNOWLEDGMENTS
We thank Margaret Tyler for technical assistance and Dr. A. W. Ghent for statistical advice. We are grateful to Dr. Aron Averbuch for modifying the magnetic tape, PDP/& system used in this study. Support was provided by USPHSMH 10175 and NSFGB 20505. This research represents part of a thesis submitted to the University of Illinois by J. F. Hasler in partial fulfillment of the Ph.D. degree.
REFERENCES
Banks, E. M. (1968). Behavioural biology of the collared lemming Dicrostonyx groen- lundicus (TrailI): 11. Sexual behaviour. Anim. Behzv. 16, 263-270.
Boling, J. L., and Blandau, R. J. (1939). The estrogen-progesterone induction of mating responses in the spayed female rat. Endocrinology 25, 359-364.
Ciaccio, L. A., and Lisk, R. D. (1971). Hormonal control of cyclic estrus in the female hamster. Amer. J. physiol. 221, 936-942.
Davidson, J. M. (1972). Hormones and reproductive behavior. In S. Levine, (Ed.), Hormones and Behavior. Academic Press, New York.
Davidson, J. M., Smith, E R., Rodgers, C. H., and Bloch, G. J. (1968). Relative thresholds of behavioral and somatic responses to estrogen. physiol. B&m. 3, 227-229.
Edwards, D. A., Whalen, R. E., and Hadler, R. D. (1968). Induction of estrus: estrogen-progesterone interactions. Physiol. Behav. 3, 29-33.
Gaskin, J. H., and Kitay, J. I. (1970). Adrenocortical function in the hamster: Sex differences and effects of gonadal hormones. Endocrinology 87, 779-786.
Hardy, D. F. (1972). Sexual behavior in continuously cycling rats. Behaviour 41, 288-297.
Hardy, D. F., and DeBold, J. F. (1971). The relationship between levels of exogenous hormones and the display of lordosis by the female rat. Horm. Behav. 2, 287-297.
Hasler, J. F., and Banks, E. M. (1975a). Reproductive performance and growth in captive collared lemmings (Dicrostonyx groenlandicus). Canad. J. Zool., 53, 777-787.
Hasler, J. F., and Banks, E. M. (1975b). The influence of exteroceptive factors on the estrous cycle of the collared lemming (Dfuostonyx gvoenlandicus). Biol, Reprod. 12, 647-656.
Hasler, J. F., Dziuk, P. J., and Banks, E. M. (1974). Ovulation and related phenomena in the collared lemming (Dicrostorzyx groenkzndicus). J. Reprod. Fert. 38, 21-28.
Kislak, J. W., and Beach, F. A. (1955). Inhibition of aggressiveness by ovarian hormones. Endccrinology 56, 684-692.

74 HASLER AND BANKS
Kitay, J. 1. (1963). Pituitary-adrenal function in the rat after gonadectomy and gonadal hormone replacement. Endocrinology 73, 253-260.
Leavitt, W. W., and Blaha, G. L. (1970). Circulating progesterone levels in the golden hamster during the estrous cycle, pregnancy and lactation. Biol. Reprod. 3, 353-361.
Lisk, R. D. (1973). Hormonal regulation of sexual behavior in polyestrous mammals common to the laboratory. In Handbook of Physiology. Physiology of the female reproductive tract. Amer. Physiol. Sot., Washington, D. C.
McDonald, P. G., Vidal, N., and Beyer, C. (1970). Sexual behavior in the ovariectomized rabbit after treatment with different amounts of gonadal hormones. Horm. Behav. 1, 161-172.
Mirskaia, L., and Crew, F. A. E. (1930). Maturity in the female mouse. Proc. Roy. Sot. Edinb. 50, 179-186.
Payne, A. P., and Swanson, H. H. (1971). Hormonal control of aggressive dominance in the female hamster. Physiol. Behav. 6, 355-3.57.
Payne, A. P., and Swanson, H. H. (1972). The effect of sex hormones on the aggressive behaviour of the female golden hamster @fesocricetus auratus waterhouse). Anim. Behav. 20, 782-787.
Siegel, S. (1956). Nonparametric statistics for the Behavioral Sciences. McGraw-Hill, New York.
Uchida, K., Kadowaki, M., and Miyake, T. (1969). Ovarian secretion of progesterone and 2Oa-hydroxypregn4ex-3one during rat estrous cycle in chronological relation to pituitary release of luteinizing hormone. Endocrinol. Jap. 16, 227-237.
Vandenbergh, J. G. (1971). The effects of gonadal hormones on the aggressive behaviour of adult golden hamsters (Mesocricetus auratus). Anim. Behav. 19, 589-594.
Whalen, R. E., and Hardy, D. F. (1970). Induction of receptivity in female rats and cats with estrogen and testosterone. Physiol. Behav. 5, 529-533.
White, R. E. C. (1971). Wrats: A computer compatible system for automatically recording and transcribing behavioural data, Behaviour 40, 135-161.
Young, W. C. (1961). The hormones and mating behavior. In W. C. Young, (Ed.), Sex and Internal Secretions. Williams and Wilkins, Baltimore.
Young, W. C., and Rundlett, B. (1939). The hormonal induction of homosexual behavior in the spayed female guinea pig. Psychosom. Med. 1, 449-460.






















