Embed Size (px)
Citation preview

doi:10.1016/j.jmb.2008.02.001 J. Mol. Biol. (2008) 378, 31–43
Available online at www.sciencedirect.com
The C-terminal Region of the Meiosis-specific ProteinKinase Ime2 Mediates Protein Instability and is Requiredfor Normal Spore Formation in Budding Yeast
Fatih Sari1, Melanie Heinrich1, Wibke Meyer1, Gerhard H. Braus1
and Stefan Irniger1,2⁎
1Institute of Microbiology andGenetics, Georg-August-University, Grisebachstr. 8,D-37077 Göttingen, Germany2Institute of Microbiology,Leibniz University Hannover,Schneiderberg 50, D-30167Hannover, Germany
Received 20 October 2007;received in revised form24 January 2008;accepted 1 February 2008Available online12 February 2008
*Corresponding author. Institute ofGenetics, Georg-August-University,D-37077 Göttingen, Germany. [email protected] addresses: M. Heinrich, GE
Josef-Engert-Str.11, D-93053 RegenMeyer, Institute of Genetics, HeinUniversitätsstr.1, D-40225 Düsseldorf,Abbreviations used: CDK, cyclin-
MAK, male germ cell-associated kinkinase; APC/C, anaphase promotincyclosome; DAPI, 4′,6-diamidino-2-pspindle pole body.
0022-2836/$ - see front matter © 2008 E
The cyclin-dependent kinase Cdk1 and the related kinase Ime2 act in concertto trigger progression of the meiotic cell cycle in the yeast Saccharomycescerevisiae. These kinases share several functions and substrates duringmeiosis, but their regulation seems to be clearly different. In contrast toCdk1, no cyclin seems to be involved in the regulation of Ime2 activity. Ime2is a highly unstable protein, andwe aimed to elucidate the relevance of Ime2instability. We first determined the sequence elements required for Ime2instability by constructing a set of deletions in the IME2 gene. None of thesmall deletions in Ime2 affected its instability, but deletion of a 241 aminoacid C-terminal region resulted in a highly stabilized protein. Thus, the C-terminal domain of Ime2 is important for mediating protein instability. Thestabilized, truncated Ime2 protein is highly active in vivo. Replacement of theIME2 genewith the truncated IME2ΔC241 in diploid strains did not interferewith meiotic nuclear divisions, but caused abnormalities in spore formation,as manifested by the appearance of many asci with a reduced spore numbersuch as triads and dyads. The truncated Ime2 caused a reduction of sporenumber in a dominant manner. We conclude that downregulation of Ime2kinase activity mediated by the C-terminal domain is required for theefficient production of normal four-spore asci. Our data suggest a role forIme2 in spore number control in S. cerevisiae.
© 2008 Elsevier Ltd. All rights reserved.
Keywords: meiosis; protein kinase; proteolysis; Saccharomyces cerevisiae;sporulation
Edited by J. KarnIntroduction
Cyclin-dependent kinases (CDKs) have fundamen-tal roles for triggering progression of the eukaryotic
Microbiology andGrisebachstr. 8,l address:
NEART AG, BioPark,sburg, Germany; W.rich-Heine-University,Germany.dependent kinase;ase; ICK, intestine cellg complex/henylindole; SPB,
lsevier Ltd. All rights reserve
cell cycle. The activity of these kinases depends onthe association with unstable regulatory proteinsknown as cyclins, which mediate oscillation of thekinase activity during the cell cycle. In the buddingyeast Saccharomyces cerevisiae, the kinase Cdk1 (alsoknown as Cdc28) is the sole essential Cdk for cell-cycle progression.1,2 Different cyclins associate withCdk1: the G1 cyclins Cln1-3, the S-phase cyclinsClb5 and Clb6 and the G2/mitotic cyclins Clb1-4.Cdk1 activity is also required for the meiotic cell
cycle.3–5 Meiosis is a specialized cell division allow-ing diploid cells the exchange of genetic material andthe generation of haploid offspring. In the meioticcell cycle, one round of DNA replication is followedby two rounds of chromosome segregation. Follow-ing the meiotic divisions of S. cerevisiae, four haploidspores are produced within an ascus.In addition to Cdk1, the related protein kinase Ime2
is also essential for progression through meiosis.6
d.

32 Ime2 Instability and Spore Formation in Yeast
Ime2 is a conserved protein kinase among eukar-yotes. Homologs of Ime2 have been identified inother fungi,7–10 and this kinase has significant simi-larities to mammalian male germ cell-associatedkinase (MAK) and intestine cell kinase (ICK).11,12
In yeast, expression of the IME2 gene is restrictedto meiosis.13 The function of this protein kinase isessential for various processes during meiosis, suchas the timely execution of DNA replication, the effi-cient expression of early and middle meiotic genesand for meiotic nuclear divisions.4,14,15 It was pro-posed that Ime2 assists Cdk1 and that it can take oversome functions of Cdk1 during meiosis.3,5 Consis-tent with this idea, it was found that Ime2 fulfills thefunctions of the G1-specific Cdk1 before premeioticDNA replication, rendering the G1 cyclins Cln1-3dispensable for the meiotic cell cycle.16Recently, it was demonstrated that Cdk1 and Ime2
phosphorylate their substrates on different consen-sus sequences. Whereas Cdk1 phosphorylates pro-teins within the S/T-P-x-K/R sequence, Ime2 motifswere identified as R-P-x-S/T-R/A.17,18 Despite thesedifferences, Cdk1 and Ime2 have a number of com-mon substrates. Like Cdk1, Ime2 is involved inphosphorylation of the CDK inhibitor Sic1, but phos-phorylates this factor on different sites.19 Remark-ably, Ime2 activity is essential, but not sufficient fortriggering Sic1 proteolysis.16,20
Cdh1, a coactivator of the anaphase promotingcomplex/cyclosome (APC/C), is a further commontarget of both Cdk1 and Ime2. In vivo studies showedthat ectopic expression of IME2 caused phosphor-ylation of Cdh1,21 and recently it was demonstratedthat Ime2 phosphorylates Cdh1 in vitro.18 Thus, Ime2seems to be involved in APC/C inactivation beforepre-meiotic S-phase and it is likely that this kinasecontributes to the highly versatile regulation ofAPC/C activity during meiotic divisions.22 Furthersubstrates of Ime2 are the transcription factors Ime1andNdt80, regulating the expression of earlymeioticgenes or of middle and late meiotic genes, res-pectively.5 Ime2 is also important for DNA replica-tion by phosphorylating the replication protein A,23
and contributes to prevent DNA replication duringmeiosis by phosphorylating components of the pre-replicative complex.18
The recent data reported byHolt et al.18 provided aclue as to why the regulation of meiosis may benefitfrom the presence of two different kinases withdifferent phosphorylation motifs in their substrates.The accessibility of Cdk1 and Ime2 phosphorylationsites by phosphatases is apparently different. Cdc14,a phosphatase required for exit from mitosis andmeiosis I, efficiently removed phosphates fromCdk1sites, but poorly from Ime2 sites. It was proposedthat Ime2 restricts the activity of Cdc14 and helps tomaintain sufficient kinase activity to prevent anintervening S-phase between the meiotic divisions.Little is known about regulation of Ime2 kinase
activity during meiosis. Ime2 kinase activity in-creases around premeiotic S-phase, then decreasesand subsequently rises to its highest levels duringmeiotic divisions.4 In contrast to Cdk1, no regula-
tory subunits corresponding to cyclins have beenidentified for Ime2.A mechanism for regulating Ime2 activity is prob-
ably phosphorylation. Residues Thr242 and Tyr244in the activation loop are phosphorylated, and this isdependent on the kinase Cak1 and on autophos-phorylation.24,25 This modification is required forthe function of Ime2 in meiosis. Phosphorylation ofIme2 was also detected in the C-terminal region,25
which is a regulatory domain having apparentlyinhibitory roles.26 Mutations of phosphorylationsites Ser520 and Ser625 had moderate effects onsporulation, but displayed synthetic phenotypeswith mutations in genes impaired in the exit formmeiosis I.25
The C-terminal region was identified as a bindingsite for regulatory proteins, at least under specificnutritional conditions. It was found that in mediacontaining nitrogen in combination with a non-fer-mentable carbon source, the GTPase Gpa2 binds thisregion and thereby inhibits the kinase activity.27
Modulation of protein stability may represent afurther possible mechanism for the regulation ofIme2 activity. Ime2 was found to be an unstable pro-tein. Pulse labeling experiments showed that Ime2 isdegraded rapidly in cells undergoing meiosis.28 Afurther study showed that Ime2 was degraded ra-pidly in response to small amounts of glucose, andthat this degradation is mediated by the ubiquitinligase Skp1/Cullin/F-box protein (SCF), associatedwith the F-box protein Grr1.29 Ime2 was shown to behighly unstable when expressed in dividing cells.21
It is not known whether the intrinsic instability ofIme2 has any role during the sporulation program ofyeast. To address this question, we were interestedto determine the consequence of the expression of astabilized version of Ime2. To enable such tests, wefirst identified Ime2 elements responsible for itsinstability. By constructing a set of deletions, wefound that only upon removal of a large regioncomprising most of the C-terminal regulatorydomain, Ime2 was distinctly stabilized. Expressionof this stable, highly active version of Ime2 indiploid cells did not interfere with meiotic divisions,but resulted in the accumulation of asci withreduced spore number, such as triads and dyads.Thus, the C-terminal region has an important rolefor Ime2 instability and for the efficient formation offour-spore asci.
Results
Ime2 instability in dividing cells is independentof the PEST-rich region and the F-box proteinGrr1
Ime2 is an unstable protein in meiotic cells.28 Totest the relevance of Ime2 instability for sporulation,we aimed to obtain a stabilized and active version ofIme2. For this purpose, we first wanted to identifythe regions in Ime2 responsible for protein instabi-

33Ime2 Instability and Spore Formation in Yeast
lity. Since Ime2 is a highly unstable protein whenexpressed in dividing cells,21 this allowed us toanalyze Ime2 stability by simple promoter shut-offexperiments.We first testedwhether Ime2 instability in dividing
cells is dependent on the PEST-rich region in itsC-terminal region. This region was implicated inmediating Ime2 proteolysis under conditions oflow concentrations of glucose in meiosis.29 To testwhether the PEST-rich element is needed for theinstability of Ime2 in dividing cells, we constructeda plasmid containing an HA6-tagged version ofIme2 lacking the PEST-rich region, corresponding toamino acids 479-499. This truncated IME2 genewas expressed from the inducible GAL1 promoter.Protein stability was analyzed in strains containingeither GAL-IME2ΔPEST-HA6 or, as a control, inGAL-IME2-HA6 strains by promoter shut-off experi-ments. Strains were incubated in the presence ofgalactose for 1 h and then transferred to glucosemedium. We found that Ime2 without the PESTregion is as unstable as the full-length Ime2 (Fig. 1a).Thus, other sequences than the PEST-rich region areresponsible for the rapid proteolysis of Ime2 in di-viding cells.Previously, the F-box protein Grr1, a component of
the ubiquitin-ligase SCF-Grr1, has been implicated inIme2 degradation in response to low concentrationsof glucose.29 To test whether the instability of Ime2 individing cells depends also on Grr1, we tested thestability of Ime2 in grr1 deletion strains by promoter
Fig. 1. Ime2 instability in dividing cells is independentof the PEST-rich region and of the F-box protein Grr1. (a)Strains expressing either GAL-IME2-HA6 or GAL-IME2Δ-PEST-HA6 (encoding Ime2 with a deletion of the PEST-rich region 479-499) were pre-grown in YEP+Rafmedium. Galactose (2% (w/v) final concentration) wasadded and the mixture was incubated for 1 h. Then, cellswere filtered and transferred to glucose medium (timezero) to shut off the GAL1 promoter. Samples taken at theindicated time-points after the transfer were analyzed byimmunoblotting using the anti-HA-antibody. Anti-Cdc28antibody was used as a loading control (LC). (b) Wildtype and the grr1 deletion strain, both containing theGAL-IME2-HA6 construct, were treated as described forstrains in (a).
shut-off experiments. The expression of GAL-IME2-HA6 was induced transiently by the addition ofgalactose.We found that Ime2was highly unstable ingrr1 mutants (Fig. 1b).We conclude that Grr1 and the PEST-rich re-
gion are dispensable for Ime2 instability in dividingcells.
Small deletions have little influence on stability,but a large C-terminal deletion distinctlystabilizes Ime2
Since deletion of the PEST-rich element did notcause Ime2 stabilization, we next aimed to identifythe regions in Ime2 responsible for protein instabil-ity. We created a set of deletions each comprisingabout 200 base pairs of the IME2 coding sequence(Fig. 2a, upper ten constructs). Each deletion cons-truct lacking 60-70 amino acids was expressed fromthe GAL1 promoter and fused at its C-terminalregion with a HA6 tag. The resulting ten deletionconstructs, each cloned in a centromeric plasmid,were transformed into yeast cells and stability wastested by promoter shut-off experiments.We found that none of the truncated Ime2 proteins
was distinctly stabilized (Fig. 2b). Most of the pro-teins were as unstable as the full-length Ime2. Aweak stabilization was observed for deletion of theC-terminal 45 amino acids (Ime2Δ601-645).To identify regions required for Ime2 instability,
we created constructs with larger deletions in theregulatory C-terminal region (Fig. 2a, lower ten con-structs). A deletion of the 241 amino acid C-terminalregion (amino acids 405-645) resulted in a highlystabilized protein, indicating that degradation ele-ments are localized in this region (Fig. 2c). A 179amino acid deletion of the C-terminal region (aminoacids 467-645) also significantly stabilized Ime2 pro-tein. Deletion of a smaller C-terminal region, 112amino acids (534-645) resulted in a partially unstableIme2 protein. This indicates that the region of aminoacids 467-534, encompassing the PEST-rich element,contains a degradation motif with moderate acti-vity. Other deletions in the C-terminal region hadonly weak or no influence on Ime2 instability. Thepresence of region 534-600 as the sole C-terminalelement results in a significant Ime2 instability (Fig.2c, construct Ime2Δ405-533/Δ 601-645), suggestingthat this element contains potent degradationmotifs. The combination of regions 467-533 or534-607 with the very C-terminal region (607-645)leads to a highly efficient proteolysis of Ime2 proteinand, therefore, the 45 amino acids at the very Cterminus may also contain sequences for proteininstability.Our results imply that the regulatory C-terminal
domain of Ime2 mediates instability of Ime2and that this region contains multiple degradationsignals, which can act alone with different efficien-cies, but have the most pronounced activity whenpresent in combination. Since no sequence elementis essential for Ime2 instability, different elementsobviously have overlapping functions.

Fig. 2. Large deletions in the C-terminal region cause stabilization of Ime2. (a) A map of the deletion constructs of theIME2 gene (K97: catalytic site). (b) Strains carrying different CEN-plasmids with GAL-IME2-HA6 deletion constructs weregrown in minimal medium without tryptophan and containing raffinose. After galactose addition, protein stability wasanalyzed by promoter shut-off experiments. For this purpose, strains were filtered and transferred into minimal mediumcontaining glucose. Ime2 protein was detected using the anti-HA antibody. Cdc28 used as a loading control (LC). TheIme2Δ202-264 protein was not detectable in this assay, for unknown reasons. (c) Strains containing six larger deletions inthe C-terminal region of Ime2 were analyzed as in (b).
34 Ime2 Instability and Spore Formation in Yeast
Truncated, stabilized Ime2 is highly activein vivo
We next tested whether the C-terminally trun-cated, highly stabilized Ime2 is functional. For thispurpose, we tested whether expression of the highlystable IME2Δ405-645 version, referred to asIME2ΔC241, influences the growth of yeast cells.Expression of this gene from the GAL1 promoteron a centromeric plasmid had significant effects oncell division in galactose medium. The truncatedIme2 was more toxic than the full-length version,suggesting that it was highly active (Fig. 3a). Plas-mids with shorter deletions in Ime2 did not causea similar decrease in the viability of yeast strains(data not shown). To exclude the possibility thatthe distinct phenoytpe upon IME2ΔC241 expres-sion is not caused artificially by the HA6 tag,we constructed a truncated Ime2 lacking theHA6 tag. Expression of this variant was also highlytoxic for yeast strains (data not shown). Theincreased activity of truncated Ime2 may be due
to an accelerated accumulation of the protein com-pared to full-length Ime2 protein. Indeed, theIme2ΔC241 protein accumulated to higher levelsthan full-length Ime2 (Fig. 3b). Even a strain con-taining five copies of GAL-IME2-HA produced lessIme2 protein than strains containing the truncatedIme2.Upon incubation in liquid medium containing
galactose, the expression of IME2ΔC241 resulted in auniform cell cycle arrest (Fig. 3c). Most cells wereunbudded, reminiscent of the arrest described forstrong over-expression of full-length IME2 in multi-copy integration strains.21 Inspection of cells expres-sing IME2ΔC241 revealed that most cells arrested asunbudded cells with short mitotic spindles (data notshown). This phenotype is similar to the expressionof five copies of GAL-IME2.21 This arrest in ametaphase-like state was dependent on the mor-phogenesis checkpoint protein Swe1. In swe1 mu-tants, GAL-IME2ΔC241 frequently arrested withelongated spindles, which is seen also in cells ex-pressing full-length IME2 (Figs. 3d and e). We con-
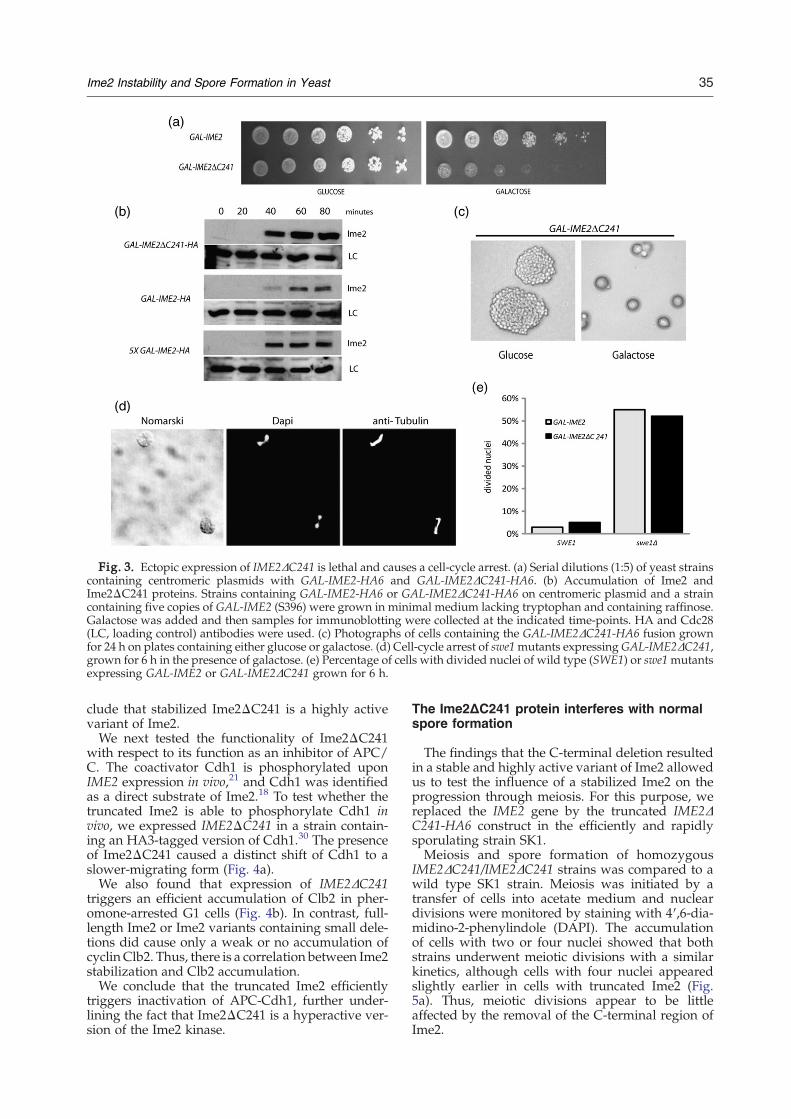
Fig. 3. Ectopic expression of IME2ΔC241 is lethal and causes a cell-cycle arrest. (a) Serial dilutions (1:5) of yeast strainscontaining centromeric plasmids with GAL-IME2-HA6 and GAL-IME2ΔC241-HA6. (b) Accumulation of Ime2 andIme2ΔC241 proteins. Strains containing GAL-IME2-HA6 or GAL-IME2ΔC241-HA6 on centromeric plasmid and a straincontaining five copies of GAL-IME2 (S396) were grown in minimal medium lacking tryptophan and containing raffinose.Galactose was added and then samples for immunoblotting were collected at the indicated time-points. HA and Cdc28(LC, loading control) antibodies were used. (c) Photographs of cells containing the GAL-IME2ΔC241-HA6 fusion grownfor 24 h on plates containing either glucose or galactose. (d) Cell-cycle arrest of swe1mutants expressingGAL-IME2ΔC241,grown for 6 h in the presence of galactose. (e) Percentage of cells with divided nuclei of wild type (SWE1) or swe1mutantsexpressing GAL-IME2 or GAL-IME2ΔC241 grown for 6 h.
35Ime2 Instability and Spore Formation in Yeast
clude that stabilized Ime2ΔC241 is a highly activevariant of Ime2.We next tested the functionality of Ime2ΔC241
with respect to its function as an inhibitor of APC/C. The coactivator Cdh1 is phosphorylated uponIME2 expression in vivo,21 and Cdh1 was identifiedas a direct substrate of Ime2.18 To test whether thetruncated Ime2 is able to phosphorylate Cdh1 invivo, we expressed IME2ΔC241 in a strain contain-ing an HA3-tagged version of Cdh1.30 The presenceof Ime2ΔC241 caused a distinct shift of Cdh1 to aslower-migrating form (Fig. 4a).We also found that expression of IME2ΔC241
triggers an efficient accumulation of Clb2 in pher-omone-arrested G1 cells (Fig. 4b). In contrast, full-length Ime2 or Ime2 variants containing small dele-tions did cause only a weak or no accumulation ofcyclin Clb2. Thus, there is a correlation between Ime2stabilization and Clb2 accumulation.We conclude that the truncated Ime2 efficiently
triggers inactivation of APC-Cdh1, further under-lining the fact that Ime2ΔC241 is a hyperactive ver-sion of the Ime2 kinase.
The Ime2ΔC241 protein interferes with normalspore formation
The findings that the C-terminal deletion resultedin a stable and highly active variant of Ime2 allowedus to test the influence of a stabilized Ime2 on theprogression through meiosis. For this purpose, wereplaced the IME2 gene by the truncated IME2ΔC241-HA6 construct in the efficiently and rapidlysporulating strain SK1.Meiosis and spore formation of homozygous
IME2ΔC241/IME2ΔC241 strains was compared to awild type SK1 strain. Meiosis was initiated by atransfer of cells into acetate medium and nucleardivisions were monitored by staining with 4′,6-dia-midino-2-phenylindole (DAPI). The accumulationof cells with two or four nuclei showed that bothstrains underwent meiotic divisions with a similarkinetics, although cells with four nuclei appearedslightly earlier in cells with truncated Ime2 (Fig.5a). Thus, meiotic divisions appear to be littleaffected by the removal of the C-terminal region ofIme2.

†http://www.yeastgenome.org
Fig. 4. Ime2ΔC241 causes Cdh1 phosphorylation andClb2 stabilization in G1 cells. (a) A strain containingGALL-HA-CDH1 as well as a strain containing GALL-HA-CDH1 and GAL-IME2, both bar1 mutants, were arrestedin G1 phase with the pheromone α-factor. Galactose wasadded to induce expression from the GAL1 promoter.Samples for immunoblotting were collected at the indi-cated time-points after galactose addition. Anti-HA anti-bodies and Cdc28 antibodies as a loading control (LC)were used. (b) A strain containing the GAL-CLB2 cons-truct was transformed with centromeric plasmids con-taining different GAL-IME2-HA6 constructs. Transfor-mants, all bar1mutants, were grown on minimal mediumlacking tryptophan and arrested with the α-factor. Ga-lactose was added and samples for immunoblotting werecollected at the indicated time-points after the additionof galactose. Anti-Clb2 and Cdc28 antibodies (LC, loadingcontrol) were used.
36 Ime2 Instability and Spore Formation in Yeast
However, the expression of the stabilized Ime2interfered with the formation of four-spore asci.Cells contained predominantly two or three sporesinstead of four spores in the majority of asci of wildtype strains (Fig. 5b). Microscopic examination oftwo-spore asci showed that two nuclei were packedinto spores, whereas the other two remained nakedinside the cell (Fig. 5c).We found similar effects of the truncated Ime2 in
another strain background, W303. This strain spo-rulates more slowly and less efficiently than SK1. Byreplacing the IME2 gene with the truncated version,we observed that four-spore asci were producedonly rarely in homozygous mutants. In contrast,dyads and triads were often produced (not shown).Thus, the defect in spore formation is not restrictedto the SK1 strain background.To provide further evidence that the dyad forma-
tion of strains containing the truncated Ime2 cor-relates with stabilization of Ime2, we used strainscarrying a short deletion in the Ime2 C-terminalregion. An Ime2 version with the Δ467-533 deletion(including the PEST-rich region), which does notcause Ime2 stabilization (Fig. 2b), was introducedinto the SK1 strain. A homozygous strain lacking
this short sequence displayed no apparent abnorm-alities in either meiotic divisions or in spore form-ation compared to wild type strains (data notshown). No increase in dyad formation comparedto wild type strains was observed.
Spores in dyads are viable and haploid
The appearance of four nuclei in sporulatingstrains with truncated Ime2 suggests that meioticdivisions occur normally in the presence of this Ime2version. This should result in the production ofhaploid spores. To test whether indeed haploidspores are produced in dyads of IME2ΔC241/IME2ΔC241 homozygous diploid strains, we ana-lyzed 88 dyads of this strain by dissection with amicromanipulator. Ten of these dissected dyads areshown in Fig. 6. This analysis revealed that morethan 80% of spores in dyads were viable. Each of theviable segregants was able to mate with only one ofthe two mating type tester strains, either MATa orMATα, indicating that all viable spores are haploid.These results indicate that cells with a truncatedIme2 frequently produce dyads containing haploidspores. We found that of 68 dyads with two viablespores, 55 contained one MATa and one MATαspore. Thirteen contained either twoMATa orMATαspores. Since the MAT locus is centromere-linked,this high frequency (81%) of a/α cells suggests thatnon-sister dyads are produced. This correlates withthe expected value (84%) calculated from theexpected recombination frequency between theMAT locus and the centromere (genetic distance30 cM†).31 Random dyad formation would result in67% of MATa/MATα asci.
Ime2ΔC241 causes spore number reduction in adominant manner
We next tested whether the increased formationof dyads and triads may be a dominant effect oftruncated Ime2 or rather caused by loss of functiondue to the removal of the C-terminal region. Forthis purpose, sporulation experiments with hete-rozygous strains containing both a normal and atruncated IME2 allele were performed. The IME2/IME2ΔC241 strain displayed a reduction of tetrads,whereas the production of dyads and triads in-creased (Fig. 7). However, the effect on sporenumber reduction was less distinct than that inthe homozygous IME2ΔC241/IME2ΔC241 diploidstrain.It could be assumed that the reduction in spore
number of the heterozygous strain may be rather aloss of function because both copies of IME2 maybe required for normal spore formation. To excludethis possibility, we tested sporulation of a diploidstrain containing only one copy of IME2. We foundthat an IME2/Δime2 heterozygous strain sporu-
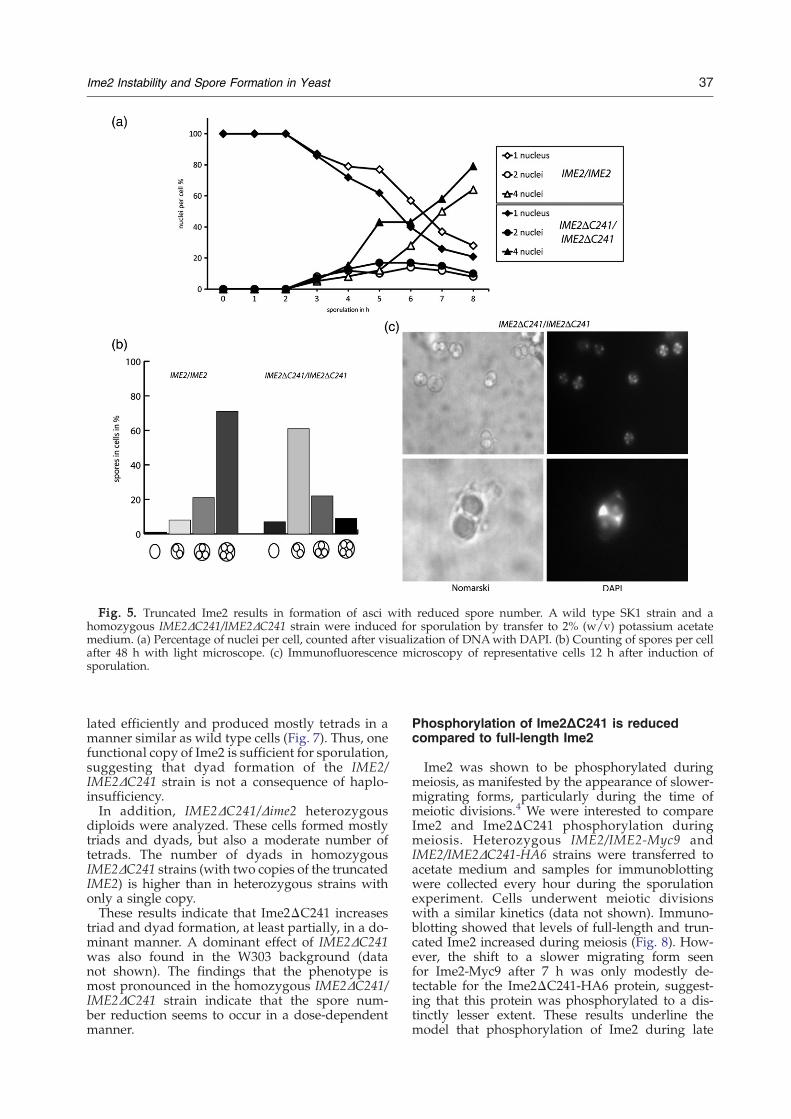
Fig. 5. Truncated Ime2 results in formation of asci with reduced spore number. A wild type SK1 strain and ahomozygous IME2ΔC241/IME2ΔC241 strain were induced for sporulation by transfer to 2% (w/v) potassium acetatemedium. (a) Percentage of nuclei per cell, counted after visualization of DNAwith DAPI. (b) Counting of spores per cellafter 48 h with light microscope. (c) Immunofluorescence microscopy of representative cells 12 h after induction ofsporulation.
37Ime2 Instability and Spore Formation in Yeast
lated efficiently and produced mostly tetrads in amanner similar as wild type cells (Fig. 7). Thus, onefunctional copy of Ime2 is sufficient for sporulation,suggesting that dyad formation of the IME2/IME2ΔC241 strain is not a consequence of haplo-insufficiency.In addition, IME2ΔC241/Δime2 heterozygous
diploids were analyzed. These cells formed mostlytriads and dyads, but also a moderate number oftetrads. The number of dyads in homozygousIME2ΔC241 strains (with two copies of the truncatedIME2) is higher than in heterozygous strains withonly a single copy.These results indicate that Ime2ΔC241 increases
triad and dyad formation, at least partially, in a do-minant manner. A dominant effect of IME2ΔC241was also found in the W303 background (datanot shown). The findings that the phenotype ismost pronounced in the homozygous IME2ΔC241/IME2ΔC241 strain indicate that the spore num-ber reduction seems to occur in a dose-dependentmanner.
Phosphorylation of Ime2ΔC241 is reducedcompared to full-length Ime2
Ime2 was shown to be phosphorylated duringmeiosis, as manifested by the appearance of slower-migrating forms, particularly during the time ofmeiotic divisions.4 We were interested to compareIme2 and Ime2ΔC241 phosphorylation duringmeiosis. Heterozygous IME2/IME2-Myc9 andIME2/IME2ΔC241-HA6 strains were transferred toacetate medium and samples for immunoblottingwere collected every hour during the sporulationexperiment. Cells underwent meiotic divisionswith a similar kinetics (data not shown). Immuno-blotting showed that levels of full-length and trun-cated Ime2 increased during meiosis (Fig. 8). How-ever, the shift to a slower migrating form seenfor Ime2-Myc9 after 7 h was only modestly de-tectable for the Ime2ΔC241-HA6 protein, suggest-ing that this protein was phosphorylated to a dis-tinctly lesser extent. These results underline themodel that phosphorylation of Ime2 during late
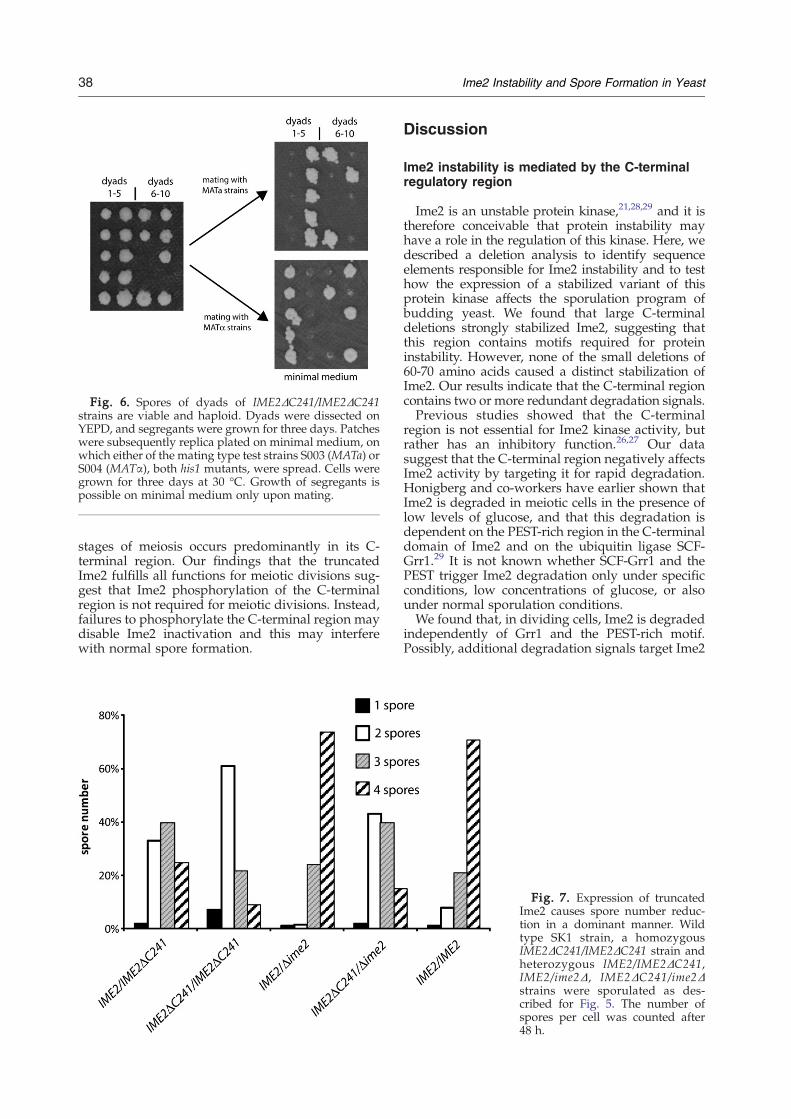
Fig. 6. Spores of dyads of IME2ΔC241/IME2ΔC241strains are viable and haploid. Dyads were dissected onYEPD, and segregants were grown for three days. Patcheswere subsequently replica plated on minimal medium, onwhich either of the mating type test strains S003 (MATa) orS004 (MATα), both his1 mutants, were spread. Cells weregrown for three days at 30 °C. Growth of segregants ispossible on minimal medium only upon mating.
38 Ime2 Instability and Spore Formation in Yeast
stages of meiosis occurs predominantly in its C-terminal region. Our findings that the truncatedIme2 fulfills all functions for meiotic divisions sug-gest that Ime2 phosphorylation of the C-terminalregion is not required for meiotic divisions. Instead,failures to phosphorylate the C-terminal region maydisable Ime2 inactivation and this may interferewith normal spore formation.
Discussion
Ime2 instability is mediated by the C-terminalregulatory region
Ime2 is an unstable protein kinase,21,28,29 and it istherefore conceivable that protein instability mayhave a role in the regulation of this kinase. Here, wedescribed a deletion analysis to identify sequenceelements responsible for Ime2 instability and to testhow the expression of a stabilized variant of thisprotein kinase affects the sporulation program ofbudding yeast. We found that large C-terminaldeletions strongly stabilized Ime2, suggesting thatthis region contains motifs required for proteininstability. However, none of the small deletions of60-70 amino acids caused a distinct stabilization ofIme2. Our results indicate that the C-terminal regioncontains two or more redundant degradation signals.Previous studies showed that the C-terminal
region is not essential for Ime2 kinase activity, butrather has an inhibitory function.26,27 Our datasuggest that the C-terminal region negatively affectsIme2 activity by targeting it for rapid degradation.Honigberg and co-workers have earlier shown thatIme2 is degraded in meiotic cells in the presence oflow levels of glucose, and that this degradation isdependent on the PEST-rich region in the C-terminaldomain of Ime2 and on the ubiquitin ligase SCF-Grr1.29 It is not known whether SCF-Grr1 and thePEST trigger Ime2 degradation only under specificconditions, low concentrations of glucose, or alsounder normal sporulation conditions.We found that, in dividing cells, Ime2 is degraded
independently of Grr1 and the PEST-rich motif.Possibly, additional degradation signals target Ime2
Fig. 7. Expression of truncatedIme2 causes spore number reduc-tion in a dominant manner. Wildtype SK1 strain, a homozygousIME2ΔC241/IME2ΔC241 strain andheterozygous IME2/IME2ΔC241,IME2/ime2Δ, IME2ΔC241/ime2Δstrains were sporulated as des-cribed for Fig. 5. The number ofspores per cell was counted after48 h.

Fig. 8. Phosphorylation of Ime2ΔC241 is reduced compared to the full-length protein. Heterozygous IME2/IME2-MYC9 and IME2/IME2ΔC241-HA6 strains were induced to enter meiosis by transfer to acetate medium. Samples werecollected at the indicated time-points after transfer. Immunoblotting was performed with anti-Myc or anti-HA antibodies,respectively.
39Ime2 Instability and Spore Formation in Yeast
for proteolysis in dividing cells. It is conceivable thatIme2 instability is of particular importance in cellsundergoing normal cell divisions, thereby helping toprevent accumulation of this kinase. Abnormal Ime2accumulation is known to cause severe defects incell cycle progression (Fig. 3c).21 To prevent such adisastrous outcome, cells may use multiple mechan-isms, including Ime2 instability, to prevent Ime2accumulation in dividing cells.
Role for Ime2 in the regulation of spore number
The finding that the truncated, active Ime2 versiondoes not display discernible defects in the progres-sion throughmeiosis implies that downregulation ofIme2 by the regulatory C-terminal domain is dis-pensable for the completion of nuclear divisions.Instead, the C-terminal region appears to have arole in regulating spore formation. Strains with atruncated Ime2 version formed few asci with fourspores, but produced many triads and dyads. Im-portantly, deletions of short C-terminal elementsneither resulted in protein stabilization nor in anincrease of asci with reduced spore numbers. Thisreflects a correlation between Ime2 stability andspore formation.By analyzing heterozygous strains, we found
that the C-terminally truncated Ime2 had, at leastpartially, a dominant effect. We found also thathomozygous strains with two alleles of IME2ΔC241caused repeatedly a more pronounced reduction ofspore number than heterozygous IME2/IME2ΔC241and IME2ΔC241/ime2Δ strains suggesting that thestabilized Ime2 affects spore number reduction in adose-dependent manner. Interestingly, over-expres-sion of IME1 also leads to a distinct increase in dyadformation.5 Ime1 is a transcriptional activator of theIME2 gene and it is possible that this effect is due tothe production of increased Ime2 protein.Our data suggest that Ime2 inhibits spore produc-
tion, and that its inactivation is required for thepacking of all nuclei into spores. Despite the cor-relation between Ime2 stabilization and defects innormal asci formation, it is possible that Ime2 inac-tivation may be induced by mechanisms other thanIme2 instability. Post-translational modifications orbinding of inhibitory proteins could also result inIme2 inactivation. The C-terminal part of Ime2 isknown to be phosphorylated and mutations of twoserine residues, Ser520 and Ser625, resulted in a
moderate accumulation of cells containing twospores.25 It was not tested in this study whetherthese spores were haploid.Possibly, phosphorylation may serve as a degrada-
tion signal, as found for targets of the SCF ubiquitin-ligase.32 Although SCF-Grr1 has been proposed to beinvolved in Ime2 proteolysis,29 the role of thisubiquitin ligase in mediating Ime2 instability duringa normal meiosis remains to be shown. Interestingly,a grr1 deletion strain also displayed defects in sporepacking resembling the phenotype of cells contain-ing stabilized Ime2.29
A key issue in the future will be to elucidate themechanism, how Ime2 contributes to regulate thespore numberwithin asci. Spore formation in yeast isinitiated by assembling a prospore membrane atspindle pole bodies (SPBs).33,34 Meiotic SPBs arecharacterized by a specific structure, the meioticplaque that is composed of the three proteinsMpc54,Mpc70 and Spo74.34–36 A low abundance of one ofthese proteins interferes with formation of the pro-spore membrane on one of the SPBs of the meiosis IIspindle. Heterozygousmutants of any of these genesresults in dyad formation, showing that levels ofthese proteins are rate limiting.37 There is compellingevidence that spore number control is a mechanismfor yeast cells to adjust spore production to theavailability of nutrients. Decreasing the levels ofacetate promotes the formation of asci with fewerspores.36 Dyad formation was also observed foryeast cells sporulating in the presence of glycerol assole carbon source.35 Intermediates of the glyoxylatepathway required for the utilization of acetate ascarbon source were suggested as signaling mole-cules, but it is not known how nutritional signalsmay be transmitted to the SPBs. It will be interestingto test whether Ime2 is involved in such a pathwayand whether a stabilized Ime2 affects the abundanceor SPB association of one of the Mpc54, Mpc70 orSpo74 proteins.
Ime2-related proteins in other eukaryotes
Ime2 is a conserved protein kinase among euk-aryotes. Homologs of Ime2 have been identified inother fungi, such as Pit1 and Mde3 in Schizosacchar-omyces pombe,7 and Crk1 in both Candida albicans8
and Ustilago maydis.9,10 Ime2 displays similarity tomammalian kinases, such as male germ cell asso-ciated kinase MAK and ICK.11,12 There is evidence

40 Ime2 Instability and Spore Formation in Yeast
that each of these Ime2-related kinases is involved inregulating sexual development.Interestingly, S. pombe mde3 pit1 double mutants
produced asci with reduced spores, implying that aninactivation of these genes has a phenotype similarto the hyperactivation of Ime2. Thus, the S. pombegenes are involved in regulating spore production,but obviously have roles opposite to those of Ime2 inbudding yeast. Whereas the S. pombe genes are re-quired for proper spore formation, budding yeastIme2 negatively affects this process. A further diffe-rence between budding and fission proteins is thatIme2 is essential for meiotic divisions, but Mde3 andPit1 are dispensable.To clarify the precise function of Ime2 and its
related protein kinases in regulating spore produc-tion will be an interesting task for the future, as wellas to unravel the role of mammalian homologs ingametogenesis.
Materials and Methods
Yeast strains
Yeast strains used in this study are derivates of eitherthe W303 strain or, for sporulation experiments, derivatesof the SK1 strains. All strains used in this study are givenin Table 1. For constructing a SK1 strain carrying thetruncated IME2ΔC241-HA6 gene instead of the normalIME2 gene, a DNA fragment containing the IME2ΔC241-HA6 sequence was amplified from the plasmid FS112using the primers FAT1 and FAT2. The PCR fragmentdigested with Acc65I and SalI was subcloned into anAcc65I-SalI-digested pBluescript II KS vector. A kanMXmodule (kanamycin-resistance gene) was amplified fromplasmid pYM138 using the primers FAT3 and FAT4 andsubsequently introduced behind the IME2ΔC241 gene. Forhomologous integration, the IME2ΔC-HA-KanMXmodulewas amplified with primers FAT5 and FAT6, andtransformed into SK1 strains. Integration was verified byPCR and immunoblotting. Removal of the C-terminal 241amino acids of Ime2 was also performed by a directtagging method,38 adding the HA6 tag after amino acid
Table 1. Saccharomyces cerevisiae strains used in this study
Strain Relevant genotype
S001 W303-1A wild type strain MATa, ade2trp1-1. leu2-3,112, his3-11,15, ura3, can1-100, G
S003 MATa, his1S004 MATα, his1S396 MATa, GAL-IME2-HA6-TRP1 (5×)S436 MATa, swe1:LEU2S474 MATa, GALL-HA-CDH1-TRP1, bar1∷HS497 SK1 wild-type strain, diploidS510 MATa, grr1∷URA3, GAL-IME2-HA6-TRS577 MATa, GAL-IME2ΔPEST-HA6-TRP1S608 MATa/MATα, IME2/IME2ΔC241-HA6-kaS610 MATa/MATα, IME2ΔC241-HA6-kanMX/ime2S611 MATa/MATα, ime2∷kanMX /ime2∷kanMS613 MATa/MATα, IME2 /ime2∷kanMXS617 MATa, GALL-HA-CDH1-TRP1, GAL-IME2ΔC241-URS635 MATa/MATα, IME2ΔC241-HA6-kanMX/IME2ΔC24S718 MATa/MATα, IME2/IME2−MYC9−TR
403. For this purpose, the HA6 tag was PCR amplifiedfrom plasmid pYM3.Full-length IME2-HA6 and IME2Δ467-533-HA6 were
cloned into an integrative plasmid essentially as describedfor the IME2ΔC241-HA6 plasmid.The ime2 deletion strain in the SK1 background was
constructed by amplification of the disruption cassettefrom the EUROSCARF ime2 deletion strain, transforma-tion into the SK1 strain, selection on kanamycin plates andverification of transformants by PCR.
Plasmid construction
Plasmids used in this study are given in Table 2.Construction of the GAL-IME2-HA6 fusion was asdescribed.21 The gene fusion was cloned as a KpnI-HindIIIfragment into the centromeric plasmid YCplac22.39 Theversion lacking the PEST-rich sequence was constructedby PCR amplification of the GAL promoter and the N-terminal region of IME2 using primers Mel1 and Mel5.The received PCR product was cut with KpnI and PstI andcloned into the KpnI-PstI-digested vector YIplac204. In asecond step, the newly generated vector was reopenedwith PstI-HindIII and a PstI-HindIII fragment includingthe C-terminal part of IME2 and a HA tag, amplified withthe primers Mel3 and Mel4, was cloned into it.A set of deletions in the IME2 gene, each lacking about
201 base pairs (60-70 amino acids, Fig. 2a) was constructedusing a plasmid containing the GAL-IME2-HA6 genefusion (pME2116). Fragments for the deletion set wereamplified from plasmid pME2116 and subsequentlycloned into the vector YCplac22. Ten PCR fragmentswith different lengths comprised the GAL1-promoter andN-terminal region of IME2. These fragments were ampli-fied by the use of the oligonucleotide Mel1 and differentoligonucleotides (Ime2del 1-10; Table 3) from the IME2open reading frame. The PCR products were digestedwithKpnI and PstI, and cloned into YCplac22. Ten additionalfragments comprised variable lengths of the C-terminalpart of the IME2 open reading frame and the HA6 epitope.These fragments were amplified by the use of theoligonucleotide Mel3 and ten different oligonucleotide(Ime2del 11-20; Table 3) from the IME2 open readingframe. These PCR products were digested with PstI andHindIII, and cloned into YCplac22. On the basis of the 20IME2 fragments, a deletion was gained by fusion of one of
Background Source
-1,AL, psi+
W303 R. Rothstein
W303 R. RothsteinW303 R. RothsteinW303 21W303 21
isG W303 30SK1 M. Knop
P1 W303 This workW303 This work
nMX SK1 This work∷kanMX SK1 This workX SK1 This work
SK1 This workA3, bar1∷HisG W303 This work1-HA6-kanMX SK1 This workP1 SK1 This work

Table 2. Plasmids used in his study
Plasmid Description Source
YCplac22 Centromeric plasmid, TRP1 marker 40YIplac204 Integrative plasmid, TRP1 marker 40YIplac211 Integrative plasmid, URA3 marker 40
pBluescript II KS FermentaspYM1 Tagging plasmid 39pYM3 Tagging plasmid 39pME2116 GAL-IME2-HA6 in YCplac22 21FS100 GAL-IME2ΔPEST-HA6 in YIplac204 This workFS101 GAL-IME2Δ2-67-HA6 in YCplac22 This workFS102 GAL-IME2Δ68-135-HA6 in YCplac22 This workFS103 GAL-IME2Δ136-201-HA6 in YCplac22 This workFS104 GAL-IME2Δ202-264-HA6 in YCplac22 This workFS105 GAL-IME2Δ265-337-HA6 in YCplac22 This workFS106 GAL-IME2Δ338-404-HA6 in YCplac22 This workFS107 GAL-IME2Δ405-466-HA6 in YCplac22 This workFS108 GAL-IME2Δ467-533-HA6 in YCplac22 This workFS109 GAL-IME2Δ534-600-HA6 in YCplac22 This workFS110 GAL-IME2Δ601-645-HA6 in YCplac22 This workFS111 GAL-IME2Δ405-533-HA6 in YCplac22 This workFS112 GAL-IME2Δ405-645-HA6 in YCplac22
(termed GAL-IME2ΔC241-HA6 in the text)This work
FS113 GAL-IME2Δ405-645 in YIplac211 This workFS114 IME2ΔC241-HA6 and kanMX in pBluescript II KS This workFS115 IME2-HA6 and kanMX in pBluescript II KS This workFS116 IME2Δ467-533-HA6 and kanMX in pBluescript II KS This workFS117 GAL-IME2Δ467-645-HA6 in YCplac22 This workFS118 GAL-IME2Δ534-645-HA6 in YCplac22 This workFS119 GAL-IME2Δ405-466/Δ601-645-HA6 in YCplac22 This workFS120 GAL-IME2Δ405-533/Δ601-645-HA6 in YCplac22 This work
41Ime2 Instability and Spore Formation in Yeast
the N-terminal fragment with one of the C-terminalfragment at the PstI site (see Fig. 2a). The combination ofthe fragments resulted in a deletion of approximately 200
Table 3. Oligonucleotides used in this study
Oligonucleotide
Mel1Mel3Mel4 CMel5IME2 del 1IME2 del 2IME2 del 3 CIME2 del 4 CIME2 del 5 CIME2 del 6 CIME2 del 7IME2 del 8 CIME2 del 9 CIME2 del 10IME2 del 11IME2 del 12IME2 del 13IME2 del 14IME2 del 15IME2 del 16IME2 del 17 CIME2 del 18 CIME2 del 19IME2 del 20FAT1FAT2FAT3FAT4FAT5 ATACA
FAT6 ATATAA
base pairs. With this procedure, ten plasmids, FS101-110,were obtained. Furthermore, six additional constructs(FS111, FS112 and FS117-120) with larger deletions of the
Sequence 5′to 3′
CCGGTCACTTAAGCTCGAGCGGCGGAAGCTTTATGACCATGATTACGCC
CCGGGCTGCAGAAAGAAATTCGTAATAATTTGGCCCCCGGGCTGCAGATCATCATTATCTTCTTCAACG
CCCCCCCTGCAGGAATAAGAAGTAATACAAACCGCCCCCCCTGCAGTTTACCCAATATGTTCGATAAAGGCCCCCCTGCAGAATAAAGACCTCAAAAATTTGTATCACCCCCCTGCAGGATAAGAATATTTTCAGGTTTTAAATCCCCCCTGCAGTTTGGAATAGTAACCACTTCTTAATCCCCCCTGCAGCAAGTTTAATTTATGGACCAAATTACCCCCCCTGCAGTTCTGTATTAGTCACATTGCCTCCCCCCCTGCAGAAATTCTACCAAAGTTAGTTTTTCTTCCCCCCTGCAGAATTTTCATTATCATTTGTCAACTGCCCCCCCCTGCAGGGCATCACCCAAAACATTATCA
CCCCCCCTGCAGCAACATGATATAAGGGGTACTTTACCCCCCCTGCAGTCTGAAAATTACCAATTACATA
CCCCCCCTGCAGACACCAAGTACCCAGTATTTTGACCCCCCCTGCAGCCTTTGGATATTTGGGCATTCG
CCCCCCCTGCAGAAATTGCCTTATGTAGAAGGTTCACCCCCCCTGCAGCAAGCATTGATTTTTGCAGGTATACCCCCCTGCAGTTAAACGAATTCGTTGAAGAAGATACCCCCCTGCAGATAAATAAAGATGAAGCAGACAACACCCCCCCTGCAGTCTTTAGGCGATTCGTTTAATTCACCCCCCCTGCAGGGCCGCATCTTTTACCCATACCCCCCCGGTACCATGGTTGAAAAACGTAGTAGCCCCCCGTCGACCATGATTACGCCAAGCTTCCCCCCGTCGACGACATGGAGGCCCAGAAT
CCCCCCGCGGCCGCTGGATGGCGGCGTTAGTAGACGTAATAAAAGAAGAGTACTAAGCTTAACTTAATAGGTCATTTGGTACCATGGTTGAAAAACGTAGTAG
CTATATAACTGTGCCTAACGATTCGTCAGTCAAATGACAAATTAGCGGCCGCTGGATGGCGGCGTTAGTA

42 Ime2 Instability and Spore Formation in Yeast
IME2 open reading frame, deletions of various length inthe 3′ region of the IME2 open reading frame,were created.These constructs were cloned into the vector YCplac22 asdescribed above.Plasmid FS113 (GAL-IME2ΔC241 with no HA tag) was
constructed by removing the HA6 from plasmid bydigestion with the enzymes XbaI and KpnI, blunt-endingwith Klenow enzyme, religation and subcloning of theconstruct into YIplac211.
Media and growth conditions
Cells were grown either in YEP (1% yeast extract, 2%peptone) complete medium or minimal medium lackingthe appropriate amino acid for plasmid selection. Mediacontained either 2% (w/v) glucose or 2% (w/v) raffinoseas carbon source. For induction of the GAL1 promoter,cells were pregrown in medium containing raffinose, andthen galactose was added to a final concentration of 2%(w/v). For promoter shut-off experiments, cells werefiltered and transferred into glucose-containing medium.Cells were arrested in the G1 phase by the addition of α-factor to a final concentration of 5 μg/ml (w/v). Cells wereincubated for at least 2.5 h, until more than 90% of cellswere unbudded.
Sporulation experiments
Sporulation of yeast SK1 strains was performed asdescribed.40 In brief, single colonies were picked from YEPplates containing 2% (v/v) glycerol and streaked to YEPDplates. Patches were grown for 48 h and then inoculated inYEP containing 2% (w/v) potassium acetate. After 13.5 h,cells were filtered, washed and transferred to sporulationmedium (2% (w/v) potassium acetate). Sporulation wasmonitored by counting nuclei of DAPI-stained cells.
Immunoblotting and immunofluorescence
Cell extracts were prepared as described.41 Yeast cellswere broken with glass beads. The enhanced chemilumi-nescence detection system (Amersham) was used for thedetection of specific proteins. Antibodies were used in1:1000 (anti-HA and anti-Myc) or 1:2000 (anti-Cdc28 andanti-Clb2) concentrations. For indirect immunofluores-cence, cells were fixed in 3.7% (v/v) formaldehyde. Sphe-roplasts were prepared as described.42 DAPI staining andanti-tubulin antibodies (YOL 1/34) were used for visua-lisation of nuclei and spindles, respectively.
Acknowledgements
We are grateful to Tülin Ersahin and Sevim Dalvafor help in some of the experiments. We thank ÖzgürBayram for critical comments on the manuscript,Kim Nasmyth and Michael Knop for providingyeast strains and plasmids, as well as Marta Galovaand Michael Knop for technical advice for thesporulation experiments. This work was supportedby the Fonds der Chemischen Industrie und theVolkswagen-Stiftung.
References
1. Mendenhall, M. D. & Hodge, A. E. (1998). Regulationof Cdc28 cyclin-dependent protein kinase activityduring the cell cycle of the yeast Saccharomyces cerevi-siae. Microbiol. Mol. Biol. Rev. 62, 1191–1243.
2. Bloom, J. & Cross, F. R. (2007). Multiple levels of cyclinspecificity in cell-cycle control. Nature Rev. Mol. CellBiol. 8, 149–160.
3. Guttmann-Raviv, N., Boger-Nadjar, E., Edri, I. &Kassir, Y. (2001). Cdc28 and Ime2 possess redundantfunctions in promoting entry into premeiotic DNAreplication in Saccharomyces cerevisiae. Genetics, 159,1547–1558.
4. Benjamin, K. R., Zhang, C., Shokat, K. M. &Herskowitz, I. (2003). Control of landmark events inmeiosis by the CDK Cdc28 and the meiosis-specifickinase Ime2. Genes Dev. 17, 1524–1539.
5. Honigberg, S. M. (2004). Ime2p and Cdc28p: co-pilotsdriving meiotic development. J. Cell Biochem. 92,1025–1033.
6. Yoshida, M., Kawaguchi, H., Sakata, Y., Kominami, K.,Hirano, M., Shima, H. et al. (1990). Initiation of meiosisand sporulation in Saccharomyces cerevisiae requires anovel protein kinase homologue. Mol. Gen. Genet. 221,176–186.
7. Abe, H. & Shimoda, C. (2000). Autoregulated expres-sion of Schizosaccharomyces pombe meiosis-specifictranscription factor Mei4 and a genome-wide searchfor its target genes. Genetics, 154, 1497–1508.
8. Chen, J., Zhou, S., Wang, Q., Chen, X., Pan, T. & Liu,H. (2000). Crk1, a novel Cdc2-related protein kinase, isrequired for hyphal development and virulence inCandida albicans. Mol. Cell Biol. 20, 8696–8708.
9. Garrido, E. & Perez-Martin, J. (2003). The crk1 geneencodes an Ime2-related protein that is required formorphogenesis in the plant pathogen Ustilago may-dis. Mol. Microbiol. 47, 729–743.
10. Garrido, E., Voss, U., Muller, P., Castillo-Lluva, S.,Kahmann, R. & Perez-Martin, J. (2004). The inductionof sexual development and virulence in the smutfungus Ustilago maydis depends on Crk1, a novelMAPK protein. Genes Dev. 18, 3117–3130.
11. Fu, Z., Schroeder, M. J., Shabanowitz, J., Kaldis, P.,Togawa, K., Rustgi, A. K. et al. (2005). Activation of anuclear Cdc2-related kinase within a mitogen-acti-vated protein kinase-like TDY motif by autopho-sphorylation and cyclin-dependent protein kinase-activating kinase. Mol. Cell Biol. 25, 6047–6064.
12. Fu, Z., Larson, K. A., Chitta, R. K., Parker, S. A., Turk,B. E., Lawrence, M. W. et al. (2006). Identification ofyin-yang regulators and a phosphorylation consensusfor male germ cell-associated kinase (MAK)-relatedkinase. Mol. Cell Biol. 26, 8639–8654.
13. Smith, H. E. & Mitchell, A. P. (1989). A transcriptionalcascade governs entry into meiosis in Saccharomycescerevisiae. Mol. Cell Biol. 9, 2142–2152.
14. Mitchell, A. P., Driscoll, S. E. & Smith, H. E. (1990).Positive control of sporulation-specific genes by theIME1 and IME2 products in Saccharomyces cerevisiae.Mol. Cell Biol. 10, 2104–2110.
15. Kassir, Y., Adir, N., Boger-Nadjar, E., Raviv, N. G.,Rubin-Bejerano, I., Sagee, S. & Shenhar, G. (2003).Transcriptional regulation of meiosis in buddingyeast. Int. Rev. Cytol. 224, 111–171.
16. Dirick, L., Goetsch, L., Ammerer, G. & Byers, B. (1998).Regulation of meiotic S phase by Ime2 and a Clb5,6-associated kinase in Saccharomyces cerevisiae. Science,281, 1854–1857.

43Ime2 Instability and Spore Formation in Yeast
17. Moore, M., Shin, M. E., Bruning, A., Schindler, K.,Vershon, A. & Winter, E. (2007). Arg-Pro-X-Ser/Thr isa consensus phosphoacceptor sequence for the meio-sis-specific Ime2 protein kinase in Saccharomycescerevisiae. Biochemistry, 46, 271–278.
18. Holt, L. J., Hutti, J. E., Cantley, L. C. & Morgan, D. O.(2007). Evolution of Ime2 phosphorylation sites onCdk1 substrates provides a mechanism to limit theeffects of the phosphatase Cdc14 in meiosis. Mol. Cell,25, 689–702.
19. Sawarynski, K. E., Kaplun, A., Tzivion, G. & Brush, G.S. (2007). Distinct activities of the related proteinkinases Cdk1 and Ime2. Biochim. Biophys. Acta, 1773,450–456.
20. Sedgwick, C., Rawluk, M., Decesare, J., Raithatha, S.,Wohlschlegel, J., Semchuk, P. et al. (2006). Saccharo-myces cerevisiae Ime2 phosphorylates Sic1 at multiplePXS/T sites but is insufficient to trigger Sic1 degrada-tion. Biochem. J. 399, 151–160.
21. Bolte, M., Steigemann, P., Braus, G. H. & Irniger, S.(2002). Inhibition of APC-mediated proteolysis by themeiosis-specific protein kinase Ime2. Proc. Natl Acad.Sci. USA, 99, 4385–4390.
22. Irniger, S. (2006). Preventing fatal destruction: inhibi-tors of the anaphase-promoting complex in meiosis.Cell Cycle, 5, 405–415.
23. Clifford, D. M., Marinco, S. M. & Brush, G. S. (2004).The meiosis-specific protein kinase Ime2 directsphosphorylation of replication protein A. J. Biol.Chem. 279, 6163–6170.
24. Schindler, K., Benjamin, K. R., Martin, A., Boglioli, A.,Herskowitz, I. &Winter, E. (2003). The Cdk-activatingkinase Cak1p promotes meiotic S phase throughIme2p. Mol. Cell Biol. 23, 8718–8728.
25. Schindler, K. & Winter, E. (2006). Phosphorylation ofIme2 regulates meiotic progression in Saccharomycescerevisiae. J. Biol. Chem. 281, 18307–18316.
26. Kominami, K., Sakata, Y., Sakai, M. & Yamashita, I.(1993). Protein kinase activity associated with theIME2 gene product, a meiotic inducer in the yeastSaccharomyces cerevisiae. Biosci. Biotechnol. Biochem. 57,1731–1735.
27. Donzeau, M. & Bandlow, W. (1999). The yeast tri-meric guanine nucleotide-binding protein alphasubunit, Gpa2p, controls the meiosis-specific kinaseIme2p activity in response to nutrients. Mol. Cell Biol.19, 6110–6119.
28. Guttmann-Raviv, N., Martin, S. & Kassir, Y. (2002).Ime2, a meiosis-specific kinase in yeast, is required fordestabilization of its transcriptional activator, Ime1.Mol. Cell Biol. 22, 2047–2056.
29. Purnapatre, K., Gray, M., Piccirillo, S. & Honigberg, S.M. (2005). Glucose inhibits meiotic DNA replication
through SCFGrr1p-dependent destruction of Ime2pkinase. Mol. Cell Biol. 25, 440–450.
30. Zachariae, W., Schwab, M., Nasmyth, K. & Seufert, W.(1998). Control of cyclin ubiquitination by CDK-regulated binding of Hct1 to the anaphase promotingcomplex. Science, 282, 1721–1724.
31. Davidow, L. S., Goetsch, L. & Byers, B. (1980).Preferential occurrence of nonsister spores in two-spored asci of Saccharomyces cerevisiae: evidence forregulation of spore-wall formation by the spindle polebody. Genetics, 94, 581–595.
32. Cardozo, T. & Pagano, M. (2004). The SCF ubiquitinligase: insights into a molecular machine. Nature Rev.Mol. Cell Biol. 5, 739–751.
33. Moreno-Borchart, A. C. & Knop, M. (2003). Prosporemembrane formation: how budding yeast gets shapedin meiosis. Microbiol. Res. 158, 83–90.
34. Neiman, A. M. (2005). Ascospore formation in theyeast Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev.69, 565–584.
35. Nickas, M. E., Diamond, A. E., Yang, M. J. & Neiman,A. M. (2004). Regulation of spindle pole function by anintermediary metabolite. Mol. Biol. Cell, 15, 2606–2616.
36. Taxis, C., Keller, P., Kavagiou, Z., Jensen, L. J.,Colombelli, J., Bork, P. et al. (2005). Spore numbercontrol and breeding in Saccharomyces cerevisiae: a keyrole for a self-organizing system. J. Cell Biol. 171,627–640.
37. Bajgier, B. K., Malzone, M., Nickas, M. & Neiman, A.M. (2001). SPO21 is required for meiosis-specificmodification of the spindle pole body in yeast. Mol.Biol. Cell, 12, 1611–1621.
38. Knop, M., Siegers, K., Pereira, G., Zachariae, W., Winsor,B., Nasmyth, K. & Schiebel, E. (1999). Epitope taggingof yeast genes using a PCR-based strategy: more tagsand improved practical routines. Yeast, 15, 963–972.
39. Gietz, R. D. & Sugino, A. (1988). New yeast-Escherichia coli shuttle vectors constructed with invitro mutagenized yeast genes lacking six-base pairrestriction sites. Gene, 74, 527–534.
40. Buonomo, S. B., Clyne, R. K., Fuchs, J., Loidl, J.,Uhlmann, F. & Nasmyth, K. (2000). Disjunction ofhomologous chromosomes in meiosis I depends onproteolytic cleavage of the meiotic cohesin Rec8 byseparin. Cell, 103, 387–398.
41. Surana, U., Amon, A., Dowzer, C., McGrew, J., Byers,B. & Nasmyth, K. (1993). Destruction of the CDC28/CLB mitotic kinase is not required for the metaphaseto anaphase transition in budding yeast. EMBO J. 12,1969–1978.
42. Pringle, J. R., Adams, A. E., Drubin, D. G. & Haarer, B.K. (1991). Immunofluorescence methods for yeast.Methods Enzymol. 194, 565–602.





























