Embed Size (px)
Citation preview

The coat morphogenetic protein SpoVID is necessary for sporeencasement in Bacillus subtilis
Katherine H. Wang1, Anabela L. Isidro2,#, Lia Domingues2,#, Haig A. Eskandarian1, Peter T.McKenney1, Kevin Drew1,3, Paul Grabowski1,4, Ming-Hsiu Chua1, Samantha N. Barry1,Michelle Guan1, Richard Bonneau1,3, Adriano O. Henriques2, and Patrick Eichenberger1,*
1Center for Genomics and Systems Biology, Department of Biology, New York University, 1009Silver Center, 100 Washington Square East, New York, NY10003, USA2Instituto de Tecnologia Química e Biológica, Universidade Nova de Lisboa, Avenida daRepública, Apartado 127, 2781-901 Oeiras, Portugal3Courant Institute of Mathematical Sciences, Department of Computer Science, New YorkUniversity, New York, NY 10003, USA
SUMMARYEndospores formed by Bacillus subtilis are encased in a tough protein shell known as the coat,which consists of at least 70 different proteins. We investigated the process of spore coatmorphogenesis using a library of 40 coat proteins fused to GFP and demonstrate that twosuccessive steps can be distinguished in coat assembly. The first step, initial localization ofproteins to the spore surface, is dependent on the coat morphogenetic proteins SpoIVA andSpoVM. The second step, spore encasement, requires a third protein, SpoVID. We show that inspoVID mutant cells, most coat proteins assembled into a cap at one side of the developing sporebut failed to migrate around and encase it. We also found that SpoIVA directly interacts withSpoVID. A domain analysis revealed that the N-terminus of SpoVID is required for encasementand is a structural homolog of a virion protein, whereas the C-terminus is necessary for theinteraction with SpoIVA. Thus, SpoVM, SpoIVA and SpoVID are recruited to the spore surface ina concerted manner and form a tripartite machine that drives coat formation and spore encasement.
KeywordsBacillus subtilis; morphogenesis; spore coat; sporulation; subcellular localization
INTRODUCTIONElucidating the molecular mechanisms governing the formation of supramolecular structuresis a fundamental objective in developmental biology. The challenge of controlling theassembly of dozens of proteins into large organized functional structures is shared by allorganisms. Examples include cellular machines as diverse as the bacterial flagellum(McCarter, 2006), the divisome (Goehring and Beckwith, 2005), eukaryotic nuclear porecomplexes (Alber et al., 2007), clathrin-coated vesicles (Kaksonen et al., 2005) and variousenvelopes, including exoskeletons (Frand et al., 2005) and viral capsids (Dokland et al.,1997). Though diverse in structure and function, all of the machines above must be
4present address, Department of Ecology and Evolution, University of Chicago, 1101 E 57th Street, Chicago, IL60637, USA.*corresponding author#these authors contributed equally to this work.
NIH Public AccessAuthor ManuscriptMol Microbiol. Author manuscript; available in PMC 2010 November 1.
Published in final edited form as:Mol Microbiol. 2009 November ; 74(3): 634–649. doi:10.1111/j.1365-2958.2009.06886.x.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

assembled in a defined order and their assembly must be co-ordinated with various geneexpression programs, including development, cell cycle and stress responses. Here, we areinterested in the morphogenesis of the B. subtilis spore coat, where more than 70 differentproteins are assembled during spore development into a complex multi-layered structure thatencases the spore (Driks, 1999; Driks, 2002a; Henriques et al., 2004; Henriques and Moran,2007; Kim et al., 2006; Kuwana et al., 2002; Lai et al., 2003).
Endospores (hereafter spores) formed by Bacillus and Clostridium species are metabolicallydormant cells characterized by their extreme resistance to heat, radiation and chemicals(Nicholson et al., 2000). In Bacillus species, spore formation (sporulation) is initiated inresponse to nutrient limitation. However, since nutrients are required for sporulation ofClostridium species, the mechanisms of sporulation initiation differ between the two genera(Paredes et al., 2005; Stephenson and Hoch, 2002; Woods and Jones, 1986). Althoughspores can remain dormant for extended periods of time (Nicholson, 2003), sporegermination in Bacillus species occurs as soon as a sufficient amount of nutrients isavailable to sustain cell growth (Setlow, 2003). An impaired coat structure underminesseveral of the spore resistance and germination properties (Henriques et al., 2004). Bycreating a physical barrier, the coat protects the spore against digestion by eukaryoticpredators (Klobutcher et al., 2006; Laaberki and Dworkin, 2008) and large molecules, suchas lysozyme. A second function of the coat is to mediate proper recognition and access ofnutrients to specific receptors found on the inner spore membrane (Setlow, 2003). The coatfurther facilitates germination by housing cell wall hydrolases that are required duringgermination for degradation of the spore peptidoglycan (Lambert and Popham, 2008;Ragkousi et al., 2003).
Sporulation has been studied in a great amount of detail in the model organism B. subtilis(Eichenberger, 2007; Errington, 2003; Piggot and Coote, 1976; Piggot and Losick, 2002;Stragier and Losick, 1996). Shortly after initiation of sporulation, an asymmetric divisionseparates the sporulating cell, generating two cellular compartments: a large mother cell anda smaller forespore. Asymmetric division is followed by a phagocytosis-like process, wherethe forespore is engulfed by the mother cell, creating a protoplast surrounded by a doublemembrane. The spore peptidoglycan (cortex) is synthesized between the two membranes.Spore coat assembly begins shortly after asymmetric division and continues until the coatencases the entire spore. This event marks the completion of the maturation process and themother cell lyses, releasing the spore in the environment.
Spore coat morphogenesis is a complex process in B. subtilis (Driks, 1999; Henriques andMoran, 2007). Coat proteins are all produced in the mother cell under the control of the σE
and σK factors and their identification was first achieved by reverse genetics (Bourne et al.,1991; Cutting et al., 1991; Donovan et al., 1987; Naclerio et al., 1996; Sacco et al., 1995;Zhang et al., 1993; Zheng et al., 1988). A second group of coat proteins was identified morerecently based on gene expression profiles and proteomics-based approaches (Eichenbergeret al., 2003; Henriques et al., 1995; Henriques et al., 1997; Kuwana et al., 2002; Lai et al.,2003; McPherson et al., 2005). Since the determination of the σE and σK regulons bytranscriptional profiling (Eichenberger et al., 2004; Steil et al., 2005), it has become possibleto systematically characterize mother cell genes of unknown function and identify spore coatproteins fused to the green fluorescent protein (GFP) based on their characteristicsubcellular localization pattern (Eichenberger et al., 2003; Kim et al., 2006).
Spore coat assembly is initiated by genes under the control of σE, shortly after asymmetricdivision and continues after engulfment of the forespore by the mother cell, with late geneexpression under the control of σK. Assembly of spore coat proteins into a supramolecularstructure is controlled by a small group of proteins, the coat morphogenetic proteins (Driks,
Wang et al. Page 2
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

1999; Henriques and Moran, 2007). Two morphogenetic proteins sit at the top of theassembly pathway: SpoVM (hereafter VM) and SpoIVA (hereafter IVA). VM, a shortamphipatic α helix, has the ability to bind to positively curved membranes, including thosethat are formed at the beginning of the engulfment process (Ramamurthi et al., 2009), and isessential for spore coat assembly (Levin et al., 1993). IVA is required for attachment of thespore coat (Piggot and Coote, 1976; Roels et al., 1992) and restriction of VM to the outerforespore membrane (Ramamurthi et al., 2006). Conversely, the pattern of subcellularlocalization of a GFP fusion to IVA is impaired in VM cells (Price and Losick, 1999). IVAcan self-assemble into higher order structures in an ATP-dependent manner (Ramamurthiand Losick, 2008). In addition to IVA and VM, the main coat morphogenetic proteins in B.subtilis also include SpoVID (Beall et al., 1993; Driks et al., 1994), SafA (Ozin et al., 2000;Takamatsu et al., 1999) and CotE (Zheng et al., 1988).
In this report, we are primarily concerned with SpoVID (hereafter VID), its interaction withIVA and VM and their concerted roles in directing the deposition of every coat protein tothe spore surface. Some aspects of the VID phenotype are reminiscent of IVA mutants,which led the original studies to conclude that VID played a role similar to IVA inanchoring the coat to the spore surface (Beall et al., 1993). However, IVA mutants differfrom VID mutants in the sense that IVA spores are devoid of cortex. We show here that theroles of IVA and VID in coat assembly are also distinct. In contrast to IVA, VID isdispensable for initial localization of coat proteins to the spore surface, but essential for alate stage in coat morphogenesis that we define as spore encasement. Our conclusion isbased on fluorescence microscopy experiments, where we recorded the patterns ofsubcellular localization for 39 coat protein-GFP fusions in VID cells. We also engineered aseries of targeted VID in-frame deletion mutants and demonstrated that spore encasement isdependent on the N-terminal domain of VID that, according to a consensus fold-recognitionstructure prediction method, could be a structural homolog of a phage coat protein. Ourdeletion analysis also identified two adjacent regions at the C-terminus of VID, including ashort conserved region that directly interacts with IVA, that are involved in the recruitmentof VID itself to the forespore surface. Finally, we propose a two-step model of spore coatassembly. The first step, the initial localization of coat proteins to the spore suface isprimarily controlled by IVA. The second step, which we name encasement, is the transitionfrom an asymmetric cap of fluorescence on one side of the developing spore to a symmetricdistribution of individual coat proteins around the circumference of the spore. Encasement isdriven by the concerted action of VM and VID. Hence, the main stages in coat assembly arespecified by the assembly of a tripartite molecular machine, formed by VM, IVA and VID atthe surface of the developing spore.
RESULTSGenetic interactions among coat morphogenetic proteins: IVA, VM and VID
To examine genetic interactions in the subcellular localization of IVA, VM and VID to thesurface of the forespore, we imaged IVA-GFP, VM-GFP and VID-GFP fusions insporulating cells in all combinations of fusions and deletions (Fig. 1). A table listing allstrains is supplied in the Supporting Information (Table S1), as well as a table providing adetailed quantification of the localization patterns (Table S2). Our IVA-GFP and VID-GFPfusions were functional in spore germination assays that were used as an indirectmeasurement of spore coat integrity (Fig. S1). We did not assay the functionality of the VM-GFP fusion, as it had been previously characterized (van Ooij and Losick, 2003).
In otherwise wild type sporulating cells, green fluorescent signals corresponding to VID-GFP (Fig. 1A, first row), VM-GFP (Fig. 1B, first row), IVA-GFP (Fig. 1C, first row)appeared shortly after asymmetric division of the sporangium, 2 hours after initiation of
Wang et al. Page 3
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

sporulation by suspension in Sterlini-Mandelstam (SM) medium (Sterlini and Mandelstam,1969). During the engulfment process, the VID-GFP and VM-GFP signals overlapped withthe curved septum revealed by membrane staining (FM4-64, red). By contrast, IVA-GFP didnot immediately track along the engulfing membranes. After engulfment was completed(hour 4), the red signal around the forespore was lost because the impermeability of theplasma membrane prevented staining of the forespore membranes (Pogliano et al., 1999).By that time, VID-GFP and VM-GFP had formed continuous rings of uniform intensity atthe surface of the forespore (shells in three dimensions). However, the IVA-GFP signalconsisted of two strongly staining polar caps that did not appear to be connected. In electronmicrographs of thin-sectioned spores (Driks, 1999), the coat often appears to be thicker atthe poles. It is possible that these differences in thickness may be a consequence of a non-uniform distribution of IVA around the spore. Nevertheless, we also note that in merodiploidstrains (i.e cells expressing the native copy of spoIVA in addition to the GFP fusion), a moreuniform ring is obtained (Price and Losick, 1999;Ramamurthi and Losick, 2008) indicatingthat small differences in protein levels and/or functionality of GFP fusions can directlyimpact the completeness of the fluorescent rings.
We found that both IVA and VM were essential for normal VID-GFP subcellularlocalization. However, their respective contributions to the VID-GFP localization processwere distinct. In IVA cells (Fig. 1A, middle row), recruitment of VID-GFP to the foresporesurface was severely impaired. These cells exhibited a diffuse fluorescent signal in themother cell cytoplasm with some enrichment of the signal near the septum (hour 2). At thelater time point (hour 4), additional regions of signal enrichment were sometimes observedat various locations in the mother cell cytoplasm. These regions may correspond toaggregated coat proteins. Our observations are consistent with the previously characterizedrole of IVA in anchoring coat proteins to the spore surface (Piggot and Coote, 1976;Roels etal., 1992). Furthermore, the dependency on IVA for VID subcellular localization had alreadybeen recognized in immunofluorescence microscopy experiments carried out by Ozin et al.(2001).
The VID-GFP localization pattern was also modified in VM cells (Fig. 1A, last row), but in alargely different manner than in IVA cells. In comparison to IVA cells, a brighter, crescent-shaped, VID-GFP signal was noted at the surface of the forespore in VM cells, with minimaldiffusion into the mother cell cytoplasm. However, no complete VID-GFP fluorescent ringswere ever observed. These observations imply that the initial recruitment of VID-GFP to theforespore surface is more directly correlated with the presence of IVA than it is with that ofVM. Nevertheless, VM is necessary for the completion of encirclement of the forespore byVID-GFP.
Next, we determined if VID was required for VM-GFP and IVA-GFP subcellularlocalization. VM-GFP localization was unaffected in VID cells (Fig. 1B, last row), a resultthat was anticipated considering that VM can localize to curved membranes in vitro(Ramamurthi et al., 2009). However, IVA-GFP localization was incomplete in VID cells(Fig. 1C, last row). Similarly to the localization of VID-GFP in VM cells reported above, acrescent-shaped fluorescent signal was observed on the mother cell proximal side of theforespore, but no fluorescence was detected on the mother cell distal side. An equivalentpattern has been described for GFP-IVA in VM cells (Ramamurthi et al., 2006) and wasobserved under our experimental conditions as well (Fig. 1C, middle row). Thus, VM andVID are both necessary for the transition of IVA-GFP from a single cap to a full ring offluorescence. Considering that VM-GFP subcellular localization is independent of VID, weinfer that VM is located upstream of VID in the genetic hierarchy controlling ring formation.However, since IVA is necessary for the proper localization of both VID-GFP (see above)and VM-GFP (Fig. 1B, middle row), we placed it at the top of the genetic hierarchy
Wang et al. Page 4
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

controlling the targeting of coat proteins to the forespore surface. As previouslydemonstrated, IVA is involved in restricting VM-GFP to the outer forespore membrane(Ramamurthi et al., 2006).
Taken together, we interpret our data and previously published experiments to indicate thattwo successive steps can be distinguished in the coat morphogenesis process. The first stepis the targeting of coat proteins, including VID and VM, to the forespore surface, a processprimarily controlled by IVA (Fig. 1D, first row). The second step, that we have named sporeencasement, is the formation of a full shell of spore coat proteins around the circumferenceof the spore (see below). This later morphogenetic transition is controlled by VM and VID,which act in part by promoting assembly of IVA around the spore (Fig. 1D, second row).Our results indicate that these two processes are distinct phenomena that can be geneticallyuncoupled.
VID and VM are necessary for spore encasementIn order to investigate further the process of spore encasement, we analyzed the subcellularlocalization of individual spore coat proteins fused to GFP in VID, VM and IVA cells. Ourlibrary of coat protein-GFP fusions consists, in addition to VID-GFP, IVA-GFP and VM-GFP, of 37 constructs for a total of 40 fusions. These experiments allowed us to identifywhich proteins required VID, VM or both genes for spore encasement. The complete resultsof our analysis are displayed in Figs. S2 (for VID dependency) and S3 (for VM dependency).Selected examples are presented in Fig. 2 (which also includes data for IVA dependency),with a detailed quantification provided in Table S3.
As observed above for IVA-GFP, we noticed that most coat protein-GFP fusions were stillable to anchor at the forespore surface in VID and VM cells. However, in most cases, thefinal morphogenetic transition was not completed, in the sense that no complete fluorescentrings were ever obtained. For a majority of fusions, a single fluorescent polar cap wasvisible on the mother cell proximal side of the forespore, whereas a fluorescent signal rarely(if ever) appeared on the distal side of the forespore. In VID cells, this characteristic patternof a spore encasement defect was observed for IVA-GFP (Fig. 1C), CotE-GFP (Fig. 2A),CotO-GFP (Fig. 2B) and YaaH-GFP (Fig. 2C), as well as CotA-GFP, CotB-GFP, CotD-GFP, CotG-GFP, CotM-GFP, CotS-GFP, CotT-GFP, CotU-GFP, CotW-GFP, CotZ-GFP,Tgl-GFP, YheD-GFP, YhjR-GFP, YisY-GFP, YknT-GFP, YmaG-GFP, YncD-GFP, YtxO-GFP, YutH-GFP, YuzC-GFP and YybI-GFP (Fig. S2). An identical pattern was observed inVM cells (Figs. 2A-C and S3). By contrast, in IVA cells (Fig. 2A-C), morphogenesis wasblocked at an earlier stage, as exemplified by the presence of single dots of fluorescence atvarious locations in the mother cell cytoplasm. Thus, these data suggest that, for this largegroup of coat proteins, VID and VM are both necessary for mediating spore encasement,whereas IVA is required for initial localization to the surface of the forespore.
A second group of fusions also presented distinct alterations of their localization pattern inVID cells. In contrast to the previous category, a fluorescent signal was usually observed onboth sides of the forespore. However, similarly to the first group, ring morphogenesis wasincomplete, as exemplified by the presence of several unconnected small dots offluorescence encircling the maturing forespore. No strong decrease of the fluorescencesignals was noted in VID cells, thus arguing against degradation of unassembled fusionproteins. Furthermore, the absence of diffuse fluorescence in the mother cell of VID cellssuggests that the initial targeting of these fusions to the forespore surface was independentof VID. This group comprises SafA-GFP (Fig. 2D), CotP-GFP, CotQ-GFP, OxdD-GFP,YeeK-GFP, YjqC-GFP, YjzB-GFP, YlbD-GFP, YppG-GFP and YxeE-GFP (Fig. S2). Bycontrast, in VM cells (Figs. 2D and S3), assembly was, once again, blocked at the single capstage, whereas in IVA cells, morphogenesis was blocked at an earlier stage (Fig. 2D). These
Wang et al. Page 5
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

results suggest that for these fusions VM is the main factor driving encasement, even thoughthe contribution from VID remains necessary.
Finally, a third group is composed of fusions to VM (Fig. 1B), YhaX (Fig. 2E), LipC(Masayama et al., 2007) and YsnD (Kim et al., 2006) that form complete fluorescent ringsin VID cells (Fig. S2). However, in VM cells (Figs. 2E and S3), ring formation of YhaX-GFP, LipC-GFP and YsnD-GFP was impaired, suggesting that, for these fusions, VM isdriving encasement independently of VID (and possibly with the help of another coatprotein that remains to be identified). In IVA cells, YhaX-GFP produced a diffusefluorescent signal that contrasts with the punctuate pattern that was observed for most coatprotein fusions in that background (Fig. 2E). Taken together, these data suggest that VM isat the top of a hierarchy regulating encasement, while for 35 out of 39 coat proteins VID isalso contributing to encasement.
Time course analysis of CotE-GFP localization to the forespore surfaceIn order to examine in more detail the role of VID in the encasement step, we carried out atime course analysis of the subcellular localization of CotE-GFP in wild type, VID and IVAcells (Fig. 3 and Table S4). CotE is a morphogenetic protein located at the interface betweenthe inner and outer coat layers (Driks et al., 1994) and required for assembly of the outerspore coat layer (Zheng et al., 1988). Previous studies using immunoelectron microscopyhave suggested that VID was required for the maintenance of the CotE ring around theforespore (Driks et al., 1994). We observed that recruitment of CotE-GFP to the surface ofthe forespore begins as soon as the protein is produced. The CotE-GFP fluorescent signalcan be detected initially as a single dot of fluorescence, at or near the center of the curvedseptum (hour 1.5). Importantly, and in contrast to VID-GFP and VM-GFP, the CotE-GFPprotein does not seem to track along the engulfing membranes. Even in otherwise wild typecells, there is a post-engulfment stage when the assembly of CotE-GFP stalls at the singlemother cell proximal polar cap stage (hour 2). Up to that stage, the spatio-temporal patternof subcellular localization of CotE-GFP was identical in wild type (Fig. 3A) and VID cells(Fig. 3B). In wild type cells, a fluorescent signal appeared at the distal pole at hour 2.5. Thissecond polar cap was fully extended at the next time point (hour 3) and the ring began toclose at hour 4. However, encasement did not occur in VID cells and we never observed acomplete ring of CotE-GFP fluorescence, even after prolonged incubation of thecorresponding culture. By contrast, in IVA cells (Fig. 3C), the fluorescence pattern neverprogressed beyond the single dot stage, again illustrating the distinct roles that IVA and VIDplay in coat assembly. Thus, we propose that the main function of VID in CotE-GFPlocalization, as well as the localization of every other coat protein with a similar dependenceon VID, is to drive the transition from initial localization of the protein to encasement, theformation of a complete ring. In the experiments presented below, we will use this propertyas a morphogenetic marker to assess the functionality of VID in-frame deletion mutants.
The conserved N-terminal domain of VID is predicted to be a structural homolog of aphage capsid protein
Next, we sought to determine which parts of VID were required for its morphogeneticproperties. Amino acid (a.a.) sequence comparisons of VID orthologs in endospore-formingbacteria revealed regions of conserved sequence at the N- and C-termini of the protein,whereas the central region of the protein is highly variable in length and sequencecomposition (Fig. S4).
Using FFAS-03 (Jaroszewski et al., 2005), a consensus fold-recognition structure predictionmethod, we obtained two strong hits (FFAS > 200) for the N-terminal sequence of VID(hereafter N-domain, a.a. 1 to 137): to the coat protein for the bacteriophage PP7 (Fig. S5A)
Wang et al. Page 6
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

(Tars et al., 2000) and to the lipocalin-like fold (Fig. S5B) (Grzyb et al., 2006). The maindifference between these fold predictions is the last secondary structure element of the N-domain, which would be a helix in the phage protein prediction and a strand in the lipocalinprediction.
The C-terminal region of VID includes a LysM domain between a.a. 525 and 575 (Costa etal., 2006). LysM domains have the ability to bind peptidoglycan and have been identified inseveral other bacterial proteins, including coat proteins SafA and YaaH (Buist et al., 2008;Ozin et al., 2000; Takamatsu et al., 1999). It has been suggested that the LysM domainfacilitates targeting of VID by binding to cell wall material located in the space between thetwo spore membranes or to cell wall precursors, even before the cortex is formed (Costa etal., 2006; Henriques and Moran, 2007; Ozin et al., 2000; Ozin et al., 2001; but see theDiscussion). However, the region of sequence conservation at the C-terminus extendsbeyond the LysM domain and includes residues 500 to 524. Hereafter, we refer to thissecond region of conservation as region A (because we show below that it is essential to theinteraction with IVA).
The conserved N-terminal domain is necessary for coat morphogenesisIn order to investigate the contribution of the N-domain to coat morphogenesis, we designedan experimental system where encasement by a CotE-YFP fusion was used as a reporter forVID morphogenetic activity. We generated a series of in-frame deletions in VID whereevery construct (which were all driven by the native VID promoter) was fused to the codingsequence of cfp and integrated at the non-essential amyE locus in a VID null strain. Tocontrol for proper expression and localization of the resulting VID-CFP fusions, we carriedout fluorescence microscopy and Western blotting experiments at hourly intervals afterinitiation of sporulation. For each construct, we determined the localization of the CotE-YFPfusion, which was expressed from the native cotE locus. On Western blots using anti-GFPantibodies, bands of the expected sizes were obtained for all of the constructs with minimaldifferences in signal intensities (Fig. S6A).
All of the deletions that were generated in the N-domain (Δ24-136, Δ86-136 and Δ125-136)recapitulated the phenotype of the null mutant strain, i.e. CotE-YFP localization wasblocked at the single cap stage (Fig. 4). Importantly, the initial localization to the foresporesurface of the corresponding VID-CFP fusions was not affected, either temporally orspatially, implying that the recruitment of VID to the forespore surface is independent ofresidues 24 to 136. We also determined the localization of a few other YFP fusions from ourlibrary (to IVA, YtxO, CotP and SafA) in the strain that expresses the SpoVIDΔ86-136-CFPfusion (Fig. S7) and observed disrupted localization patterns that are consistent with thosereported in Figs. 2 and S2, confirming that the morphogenetic function of the N-domain wasnot limited to CotE.
In addition, we analyzed the function of the central region of VID (hereafter region M,where M stands for middle region). We observed that the morphogenetic activity of theprotein is largely maintained when extended fragments of region M (Δ140-424 andΔ140-484) are missing (Figs. S6B and S8). Some minor changes in activity were noted forthe largest deletion construct (VIDΔ140-484-CFP). The shape of both the VID-CFP and CotE-YFP rings was perturbed and the time necessary to complete ring formation extended byapproximately an hour. Although further studies will be necessary to clarify the role of thecentral region in spore coat assembly, our data show that most of region M is dispensable forspore encasement.
Wang et al. Page 7
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

The conserved C-terminal region is required for anchoring at the forespore surfaceNext, we attempted to determine the respective roles of the two conserved regions at the C-terminus of the protein: the LysM domain (from a.a. 525 to 575) and region A (from a.a. 500to 524). Three C-terminally truncated VID-CFP constructs were generated: a VID1-525-CFPconstruct, which only removes the LysM domain; and two constructs (VID1-500-CFP andVID1-484-CFP) that remove both the LysM domain and region A. In contrast to the N-terminus, we find that the C-terminus appears to be necessary for the initial localization ofVID-CFP itself to the spore surface.
The localization of each construct to the forespore surface was impaired, as indicated by thepresence of a strong and diffuse fluorescence signal in the mother cell cytoplasm (Fig. 5).Western blot analyses revealed fusions of the expected sizes (Fig. S6C), confirming that theobserved fluorescence pattern was due to the deletion within the fusion proteins and not tothe inadvertent cleavage of the CFP moiety. When the deletion was limited to the LysMdomain, the mislocalization was less pronounced and a significant enrichment of the signalwas still noted at the forespore surface (on both sides of the forespore, although the mothercell distal cap is less extended than the mother cell proximal cap). By contrast, noenrichment at the forespore surface was noted for either VID1-500-CFP or VID1-484-CFP,suggesting that all of the necessary residues involved in localization to the forespore surfacehad been lost.
In the case of the VID1-525-CFP fusion, it seems that a sufficient amount of themorphogenetic protein localized properly to permit some signal from CotE-YFP to appear atthe mother cell distal forespore pole, suggesting a residual amount of encasement activity(Fig. 5). By contrast, in the cases of VID1-500-CFP and VID1-484-CFP, encasement wasprevented to a level comparable to what was observed in the VID null strain. Thus, similarlyto the N-domain, the C-terminal region of VID is necessary for spore encasement. However,unlike the N-domain, this may be an indirect effect caused by the failure of themorphogenetic protein to reach its proper subcellular location. Altogether, results from Fig.5 imply that the C-terminal region is essential for localizing VID to the forespore surfacewith a more significant contribution of region A, than from the LysM domain.
In order to investigate the contribution of the 484-525 region independently of the LysMdomain, we generated two additional in-frame deletion constructs. The first construct(VIDΔ500-525-CFP) removes region A, whereas the second construct (VIDΔ484-500-CFP)leaves region A intact. Western blots for each construct are displayed in Fig. S6D. TheVIDΔ500-525-CFP construct showed a diffuse fluorescent signal in the mother cell cytoplasmand some preferential localization at the surface of the forespore (Fig. 5). However, contraryto the truncation of the LysM domain, no localization of VIDΔ500-525-CFP was observed onthe mother cell distal side of the forespore. As a consequence of this incomplete localizationpattern, CotE-YFP ring formation was stalled at the initial localization stage. In contrast, thedeletion of a.a. 484 to 500 had no observable effect, demonstrating that this region wasdispensable for VID function, at least under our experimental conditions.
Taken together, our data indicate that the C-terminal region of VID constitutes a targetingsignal, which comprises two components: the LysM domain and region A. Both elementscontribute to the targeting of VID.
The C-terminus of VID is required for the interaction with IVASince subcellular localization of VID is strictly dependent on IVA in fluorescencemicroscopy experiments (Fig. 1A), we decided to determine whether VID and IVAinteracted with one another in biochemical assays. In a first series of experiments, we usedGST fusions to VID produced in E. coli (Figs. 6A and S9A) and measured their ability to
Wang et al. Page 8
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

pull down IVA. The results presented in Fig. 6B showed the retention of purified IVA by afusion to GST to full length VID. In order to more precisely map the interacting region, weused a series of truncated forms of VID (Costa et al., 2006) and observed that IVA waspulled down by a L201-A575 fragment that is lacking the entire N-domain. By contrast, it wasnot recovered by the T499, R202, A302, N399 and L201-N399 fragments, suggesting thatneither the N-domain, nor the central region of VID were able to interact with IVA in theabsence of the C-terminal region. From these experiments, we conclude that VID interactsdirectly with IVA and that the region delimited by residues 499 to 575 is required for theinteraction.
To confirm this interaction using a different assay, we performed affinity blottingexperiments (Einarson and Orlinick, 2002). We generated a series of VID constructscontaining a His6 tag (Fig. 6C). The protein fragments (including His6-mCherry used asnegative control) were produced in E. coli, purified on Ni2+-NTA agarose columns, resolvedby SDS-PAGE and stained with Coomassie (Fig. S9B). The proteins were transferred to anitrocellulose membrane and hybridized with purified His6-IVA. Next, the binding of IVAto the membrane was detected by immunoblotting with an anti-IVA antiserum. We observedbinding of IVA to full-length VID, but not to the purified LysM domain or to the first 120residues of VID (Fig. 6D). As a control, we showed that the anti-IVA antibody did notcross-react with VID or mCherry. Next, we tested the binding of IVA to the proteinfragments VIDΔLysM, VID121-575 and VID121-521, which all contain region A. AlthoughVIDΔLysM and VID121-521 did not contain the LysM domain, every fragment was able torecruit IVA (Fig. 6E). Therefore, our data are consistent with the interpretation that region Ais necessary for the interaction with IVA, even in the absence of the the LysM domain.
DISCUSSIONThe assembly of 70 proteins into a functional spore coat constitutes a paradigm for theformation of a supramolecular structure in a cell undergoing development (Driks, 1999;Henriques and Moran, 2007). Coat assembly is a dynamic and inherently asymmetricprocess, as exemplified by the patterns of subcellular localization previously reported for thecoat proteins IVA, SafA, CotE, YheD and YutH (Ozin et al., 2001; Price and Losick, 1999;van Ooij et al., 2004; Webb et al., 1995). We began this work with a characterization of theeffects of the deletion of VID on coat assembly. We have presented evidence that VID is acoat protein whose morphogenetic activity is required during a later stage of coat assembly.Specifically, we propose that VID is governing a morphogenetic transition that we namedspore encasement. Our conclusion is supported by the analysis of the patterns of subcellularlocalization for 39 coat protein-GFP fusions in VID cells. We have shown that all of thesefusions initially localized to the forespore surface independently of VID and 90% (35 out of39 fusions) failed to complete formation of the ring pattern typical of normal coat proteinlocalization. In a majority of cases (70%, 25 out of 35 fusions), the assembly process wasblocked at the stage just after initial localization as a single cap of fluorescence on themother cell proximal side of the forespore. Given that the spore coat phenotype of the VIDmutant strain is markedly different from that of IVA cells, we propose that encasement isindeed a distinct second step of spore coat morphogenesis.
Our findings confirm and expand the conclusions of (Ozin et al., 2001) who observed byimmunofluorescence microscopy that VID was required for SafA to fully encircle theforespore. In addition, our fluorescence microscopy data are reminiscent of previouslypublished transmission electron microscopy images of VID cells (Beall et al., 1993).Although the principal conclusion of that original study was that the main role of VID wasto anchor the spore coat to the forespore surface, we note that in some of these images, thespore coat material found in the center of the mother cell had a horseshoe shape, which
Wang et al. Page 9
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

would be consistent with incomplete spore encasement, followed by detachment of the sporecoat from the spore surface. This phenomenon can also be observed in the VID mutantstrains analyzed by Driks et al. (1994). We rarely observed detached caps of fluorescence inthe mother cell of VID cells. However, we attribute this difference to the fact thatfluorescence microscopy is a less disruptive method than electron microscopy and does notrequire fixing, staining and sectioning of sporulating cells. Taken together, theseobservations and our data suggest that, in coat assembly, VID facilitates the morphogenetictransition from single polar cap to a complete shell.
We also draw a parallel between B. subtilis spore coat assembly and B. anthracisexosporium formation. The exosporium constitutes the outermost spore structure in B.anthracis, B. cereus and several other spore forming bacteria, although B. subtilis spores aredevoid of it (Driks, 2002b; Henriques and Moran, 2007). In the absence of the exosporiumprotein ExsY, assembly of the B. anthracis exosporium is blocked at the single cap stage(Boydston et al., 2006; Steichen et al., 2007). Thus, although different morphogeneticproteins regulate spore encasement during exosporium and spore coat formation, bothprocesses include at least two successive steps (Giorno et al., 2009).
Our second major finding is that the N-domain of VID is essential for the morphogeneticprocess of spore encasement. We predicted that the first 150 residues of VID fold into asingle protein domain with two likely folds. The first fold corresponds to the coat protein ofthe PP7 phage (Tars et al., 2000). Although hypothetical, this previously unsuspectedstructural homology between two proteins that belong respectively to spore and phageenvelopes suggests that similar mechanisms may be operating during the assembly of sporeand phage external protective structures. Specifically, the PP7 phage capsid proteinundergoes multimerization during phage morphogenesis, and our preliminary results alsoindicate that both the full-length VID protein and the N-domain alone can form multimericstructures (data not shown). The second prediction is a lipocalin-like fold. The β-barrelstructure characteristic of lipocalins is usually a binding site for small hydrophobicmolecules, such as lipids (Campanacci et al., 2006). It remains to be tested whether VID canbind lipids in the manner of some lipocalins. However, since lipocalins can also bind otherproteins (Skerra, 2008), the similarity to the lipocalin fold suggests that the N-domain maybe involved in protein-protein interactions.
Finally, our analysis showed that the C-terminus of VID is essential for targeting VID itselfto the surface of the spore. This region has two main components, the previouslycharacterized LysM domain (a.a. 525-575) and the directly adjacent region A (a.a. 500-524).Both are involved in recruitment to the forespore surface, although the contribution ofregion A seems more important. A parallel can be drawn to the morphogenetic protein SafA,which also has a LysM domain (at its N-terminal end) followed by a conserved stretch ofamino acids. In that case as well, both components are necessary for the subcellularlocalization of the protein to the surface of the forespore (Costa et al., 2006). Given thatLysM domains have been shown to interact with peptidoglycan, it has been suggested thatthe LysM domain of VID may facilitate the initial targeting of the protein to the outerforespore membrane by recognizing peptidoglycan or peptidoglycan precursors, even beforesynthesis of the spore cortex (Ozin et al., 2000; Ozin et al., 2001; Costa et al., 2006;Henriques and Moran, 2007). However, unlike other proteins with LysM domains, VID doesnot contain a signal sequence for secretion, suggesting that an atypical mechanism wouldhave to be used to translocate the LysM domain across the outer forespore membrane. Analternative possibility is that the LysM domain contributes to the initial targeting of VID byfacilitating its interaction with IVA, presumably through region A, and that it only binds tothe cortex at a later stage of development, when the forespore outer membrane becomes
Wang et al. Page 10
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

more permeable, or even during spore germination, when the cell wall hydrolases that werestored in the spore coat are released to carry out cortex degradation.
Our model of spore coat morphogenesis distinguishes between two successive steps: initiallocalization of coat proteins to the spore surface and spore encasement (Fig. 7). In thismodel, we propose specific roles for the three principal coat morphogenetic proteins: IVA,VM and VID. IVA is essential to the initial localization step of every coat protein. Thisconclusion is supported by the observation that in electron micrographs of IVA cells, the coatappears to be completely detached from the spore, as swirls of aggregated proteins floatingin the middle of the cell (Piggot and Coote, 1976;Roels et al., 1992). Similarly, IVA isrequired for recruitment of VID through a direct interaction, presumably involving region A.Although purified VM-GFP is able to localize to positively curved membranes in vitro andwhen heterologously expressed in Escherichia coli and Saccharomyces cerevisiae, it alsophysically interacts with IVA. This interaction appears to be necessary to restrict VM to theouter spore membrane in vivo (Ramamurthi et al., 2006).
We propose that the next step, encasement, is driven by the combined actions of VID, andVM. IVA has been shown to form polymer cables upon the addition of ATP to purified IVAin vitro (Ramamurthi and Losick, 2008). Subsequently, VM and VID may be necessary tophysically bend cables of IVA around the spore surface. The structural homology of the N-domain of VID to a phage coat protein, as well as our preliminary observations (data notshown), suggests that VID may also be able to multimerize. Therefore, one could envision amechanism whereby VID, IVA and VM form a tripartite machine that polymerizes on top ofthe positively curved outer spore membrane and drives encasement of the spore by the sporecoat. Once the components are properly localized, VID multimerizes to form a phageenvelope-like structure around the surface of the spore while simultaneously binding to IVAand bending IVA cables around the outer spore surface. VM, via its interaction with IVAand binding to the outer spore membrane, localizes the continually polymerizing machine tothe outer spore membrane as engulfment proceeds. The molecular details of how themachine assembles remain to be discovered.
Our model does not currently provide a detailed mechanism for the formation of the secondfocus of VID-GFP on the mother cell distal side of the spore. This second focus wouldpresumably act as a nucleation site to promote the formation of the second polar cap. Furtherstudies will be necessary to distinguish between alternative mechanisms. For instance,continuous expansion in a single plane of the first single focus could lead to the formation ofthe second focus on the opposite side of the spore. This mechanism would imply anintermediate structure connecting the two foci in the form of a thin fluorescent ring aroundthe spore. Similar rings have been observed before for the sporulation protein YabP-GFP(van Ooij et al., 2004), but we did not detect such an intermediate in time courseexperiments with VID-GFP. However, it remains possible that this structure is short-livedand difficult to capture with traditional approaches. Alternatively, the formation of thesecond focus might be conditioned by specific positional cues on the distal site of theforespore. Such a cue could be provided by the sporulation protein SpoIIIE (or a protein thatinteracts with it). It has indeed been shown that the single fluorescent focus formed by aSpoIIIE-GFP fusion migrates from the mother cell-proximal to the distal side of theforespore during engulfment (Sharp and Pogliano, 1999).
Considering that every coat protein requires IVA for localization to the spore, anexplanation of the encasement of IVA itself may explain the mechanism of encasement forthe remainder of the coat. However, we favor a model with contributions from additionalmorphogenetic proteins, such as SafA and CotE, to relay the effects of VID and IVA todownstream components of the coat protein network. This hierarchical organization of the
Wang et al. Page 11
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

assembly process is supported in part by the observation that the N-domain of VID interactsdirectly with SafA (Costa et al., 2006). The existence of the second and third classes ofVID-dependent coat proteins, those that form an incomplete punctuate ring and those thatare unaffected by a VID deletion respectively, also suggest the contribution of other coatproteins to promote optimal spore encasement. However, each class of coat proteins is fullydependent on VM for encasement implying a branched hierarcy of encasement regulation:class 1 includes 70% of the coat proteins (i.e. those that are strictly dependent on VM andVID for encasement), class 2 includes 20% of the coat proteins (strictly dependent on VMand partially dependent on VID) and class 3 includes 10% of the coat proteins (strictlydependent on VM and potentially dependent on yet unidentified coat proteins).
In conclusion, we have shown that VID and VM play critical roles in spore coat assembly bymediating the process of spore encasement and that encasement is a process of spore coatmorphogenesis distinct from initial localization under the control of IVA. Both the N-terminal and C-terminal conserved regions of VID are required for full morphogeneticactivity. However, the role of the C-terminal region may be limited to the targeting andanchoring of the protein to the surface of the spore via either a direct interaction with IVA orby facilitating this interaction, whereas the N-domain is likely to constitute the activemorphogenetic component necessary for encasement. These three proteins appear to be theminimal requirements to form a shell of IVA protein around the spore and thus, to recruit therest of the coat. The predicted structural homology to a phage coat protein essential forcapsid formation raises the intriguing possibility that parts of the molecular mechanism offormation of protective envelopes are conserved from bacteriophages to bacterial spores.
EXPERIMENTAL PROCEDURESStrains, plasmids and primers
All strains used here were derivatives of the wild-type strain PY79 (Youngman et al., 1984)and are listed in Table S1. Strains construction (including VID mutagenesis, production ofGST fusions to VID, production and purification of full-length and truncated forms of His6-VID and His6-IVA) is detailed in the Supporting Information. Every plasmid andoligonuleotide primer used is listed in Tables S5 and S6, respectively.
Growth and sporulation conditionsCells were incubated at 37°C in hydrolyzed casein medium to A600nm of 0.6, pelleted bycentrifugation at 4000 g for 5 min and resuspended in SM medium (Sterlini andMandelstam, 1969). After resuspension, cells were returned to 37°C and imaged at theindicated times by fluorescence microscopy.
Fluorescence microscopyFluorescence microscopy was performed as described before (Kim et al., 2006). Briefly,1ml aliquots of the sporulating cultures were transferred into microcentrifuge tubes and spundown at 8000RPM for 2 minutes in a microcentrifuge. Pellets were resuspended in 100μLPBS supplemented with the membrane dye FM4-64 (Invitrogen) at 1.5μg ml-1 finalconcentration. Two μl of the concentrated sample were placed on a microscope slide andcovered by a poly-L-lysine-treated coverslip for analysis. Control experiments wereperformed (Figures S10 and S11, Table S4) to confirm that neither the centrifugationconditions nor the poly-L-lysine treatment influenced the localization patterns of the GFPfusions. Images were taken using a Nikon 90i motorized fluorescent microscope equippedwith a Roper 1K monochrome digital camera and driven by the NIS-Elements AR 3.0software. Images were processed with Adobe Photoshop 8.0 for minor adjustments ofbrightness and contrast.
Wang et al. Page 12
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

GST-pull down assaysPull down assays using GST fused to the full-length VID protein or to various deletionforms of the protein were conducted essentially as described before (Costa et al., 2006),except that purified IVA (10 μg) was used as the prey protein. IVA was detected byimmunoblot analysis using an anti-His6 antibody (Novagen) at a 1:10000 dilution.
Affinity blottingThe protocol described by Einarson and Orlinick (2002) was followed with minormodifications. The partially purified full-length and truncated forms of His6-VID and His6-mCherry (5 μM final concentration) were resolved by SDS-PAGE (10% gels for theVID1-120 fragment and VID-LysM domain, and 15% gels for all the other forms) andtransferred to a nitrocellulose membrane. The membranes were incubated for 10 min inbasic buffer (20mM HEPES pH 7.9; 50mM KCl; 10mM MgCl2; 1mM DTT; 0,1 NonidetP-40) and then for 1 hour with blocking buffer (5% non-fat dry milk in basic buffer). Next,the membranes were incubated overnight with gentle agitation at 4°C in interaction buffer(1% nonfat dry milk; 5% glycerol in basic buffer) containing 5 nM of purified His6-IVA.The membrane was then washed three times with PBS 0.2% Triton X-100 for 10 minutesand twice with PBS containing 0.2% Triton X-100 and 100 mM KCl. Finally, bound IVAwas detected by immunoblotting (Costa et al., 2006) with an anti-IVA antibody at a 1:500dilution.
Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
AcknowledgmentsWe are grateful to Adam Driks for discussion, comments on the manuscript, sharing of unpublished data andstrains, Richard Losick for comments on the manuscript and strains, Jonathan Dworkin, Alan Grossman and PatrickStragier for strains. We thank Teresa Costa and Ana M. Almeida for participating at an earlier stage of the project,Uschi Auguste, Alex LoPinto, Aparna Modi, Lara Winterkorn and Kevin Wu for technical assistance. Weacknowledge the financial support of NIH grant GM081571 to PE and training grant in Developmental Genetics5T32HD007520 to PTM, Department of the Army award number W81XWH-04-1-0307 to PE and RB, NSFDBI-0820757 to RB, grant POCTI/BIA-BCM/60855/2004 and ERA-Net Pathogenomics CDIFFGEN fromFundação para a Ciência e a Tecnologia (F.C.T) to AOH and a Collegiate Research Scholar fellowship from NewYork University to SB. The content of this material does not necessarily reflect the position or the policy of theGovernment and no official endorsement should be inferred.
REFERENCESAlber F, Dokudovskaya S, Veenhoff LM, Zhang W, Kipper J, Devos D, Suprapto A, Karni-Schmidt
O, Williams R, Chait BT, Rout MP, Sali A. Determining the architectures of macromolecularassemblies. Nature. 2007; 450:683–694. [PubMed: 18046405]
Beall B, Driks A, Losick R, Moran CP Jr. Cloning and characterization of a gene required forassembly of the Bacillus subtilis spore coat. J Bacteriol. 1993; 175:1705–1716. [PubMed: 8449878]
Bourne N, FitzJames PC, Aronson AI. Structural and germination defects of Bacillus subtilis sporeswith altered contents of a spore coat protein. J Bacteriol. 1991; 173:6618–6625. [PubMed: 1917883]
Boydston JA, Yue L, Kearney JF, Turnbough CL Jr. The ExsY protein is required for completeformation of the exosporium of Bacillus anthracis. J Bacteriol. 2006; 188:7440–7448. [PubMed:16936017]
Buist G, Steen A, Kok J, Kuipers OP. LysM, a widely distributed protein motif for binding to(peptido)glycans. Mol Microbiol. 2008; 68:838–847. [PubMed: 18430080]
Campanacci V, Bishop RE, Blangy S, Tegoni M, Cambillau C. The membrane bound bacteriallipocalin Blc is a functional dimer with binding preference for lysophospholipids. FEBS Lett. 2006;580:4877–4883. [PubMed: 16920109]
Wang et al. Page 13
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Costa T, Isidro AL, Moran CP Jr. Henriques AO. Interaction between coat morphogenetic proteinsSafA and SpoVID. J Bacteriol. 2006; 188:7731–7741. [PubMed: 16950916]
Cutting S, Zheng LB, Losick R. Gene encoding two alkali-soluble components of the spore coat fromBacillus subtilis. J Bacteriol. 1991; 173:2915–2919. [PubMed: 1708381]
Dokland T, McKenna R, Ilag LL, Bowman BR, Incardona NL, Fane BA, Rossmann MG. Structure ofa viral procapsid with molecular scaffolding. Nature. 1997; 389:308–313. [PubMed: 9305849]
Donovan W, Zheng LB, Sandman K, Losick R. Genes encoding spore coat polypeptides from Bacillussubtilis. J Mol Biol. 1987; 196:1–10. [PubMed: 2821284]
Driks A, Roels S, Beall B, Moran CP Jr. Losick R. Subcellular localization of proteins involved in theassembly of the spore coat of Bacillus subtilis. Genes Dev. 1994; 8:234–244. [PubMed: 8299942]
Driks A. Bacillus subtilis spore coat. Microbiol Mol Biol Rev. 1999; 63:1–20. [PubMed: 10066829]Driks, A. Proteins of the spore core and coat. In: Sonenshein, AL.; Hoch, JA.; Losick, R., editors.
Bacillus subtilis and its closest relatives: from genes to cells. American Society for Microbiology;Washington, DC: 2002a. p. 527-536.
Driks A. Maximum shields: the assembly and function of the bacterial spore coat. Trends Microbiol.2002b; 10:251–254. [PubMed: 12088650]
Eichenberger P, Jensen ST, Conlon EM, van Ooij C, Silvaggi J, Gonzalez-Pastor JE, Fujita M, Ben-Yehuda S, Stragier P, Liu JS, Losick R. The sigmaE regulon and the identification of additionalsporulation genes in Bacillus subtilis. J Mol Biol. 2003; 327:945–972. [PubMed: 12662922]
Eichenberger P, Fujita M, Jensen ST, Conlon EM, Rudner DZ, Wang ST, Ferguson C, Haga K, Sato T,Liu JS, Losick R. The program of gene transcription for a single differentiating cell type duringsporulation in Bacillus subtilis. PLoS Biol. 2004; 2:e328. [PubMed: 15383836]
Eichenberger, P. Genomics and cellular biology of endospore formation. In: Graumann, P., editor.Bacillus Cellular and Molecular Biology. Caister Academic Press; Norfolk, UK: 2007. p. 375-418.
Einarson, MB.; Orlinick, JR. Identification of protein-protein interactions with gluthatione-S-transferase fusion proteins. In: Golemis, E., editor. Protein-Protein Interactions. Cold SpringHarbor Laboratory Press; Cold Spring Harbor, NY: 2002. p. 375-418.
Errington J. Regulation of endospore formation in Bacillus subtilis. Nat Rev Microbiol. 2003; 1:117–126. [PubMed: 15035041]
Frand AR, Russel S, Ruvkun G. Functional genomic analysis of C. elegans molting. PLoS Biol. 2005;3:e312. [PubMed: 16122351]
Giorno R, Mallozzi M, Bozue J, Moody KS, Slack A, Qiu D, Wang R, Friedlander A, Welkos S, DriksA. Localization and assembly of proteins comprising the outer structures of the Bacillus anthracisspore. Microbiology. 2009; 155:1133–1145. [PubMed: 19332815]
Goehring NW, Beckwith J. Diverse paths to midcell: assembly of the bacterial cell division machinery.Curr Biol. 2005; 15:R514–526. [PubMed: 16005287]
Grzyb J, Latowski D, Strzalka K. Lipocalins - a family portrait. J Plant Physiol. 2006; 163:895–915.[PubMed: 16504339]
Henriques AO, Beall BW, Roland K, Moran CP Jr. Characterization of cotJ, a sigma E-controlledoperon affecting the polypeptide composition of the coat of Bacillus subtilis spores. J Bacteriol.1995; 177:3394–3406. [PubMed: 7768848]
Henriques AO, Beall BW, Moran CP Jr. CotM of Bacillus subtilis, a member of the alpha-crystallinfamily of stress proteins, is induced during development and participates in spore outer coatformation. J Bacteriol. 1997; 179:1887–1897. [PubMed: 9068633]
Henriques, AO.; Costa, TV.; Martins, LO.; Zilhao, R. The functional architecture and assembly of thespore coat. In: Ricca, E.; Henriques, AO.; Cutting, SM., editors. Bacterial Spore Formers.Probiotics and Emerging Applications. Horizon Bioscience; Norfolk, UK: 2004. p. 65-85.
Henriques AO, Moran CP Jr. Structure, assembly, and function of the spore surface layers. Annu Revof Microbiol. 2007; 61:555–588. [PubMed: 18035610]
Jaroszewski L, Rychlewski L, Li Z, Li W, Godzik A. FFAS03: a server for profile--profile sequencealignments. Nucleic Acids Res. 2005; 33:W284–288. [PubMed: 15980471]
Kaksonen M, Toret CP, Drubin DG. A modular design for the clathrin- and actin-mediatedendocytosis machinery. Cell. 2005; 123:305–320. [PubMed: 16239147]
Wang et al. Page 14
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Kim H, Hahn M, Grabowski P, McPherson DC, Otte MM, Wang R, Ferguson CC, Eichenberger P,Driks A. The Bacillus subtilis spore coat protein interaction network. Mol Microbiol. 2006;59:487–502. [PubMed: 16390444]
Klobutcher LA, Ragkousi K, Setlow P. The Bacillus subtilis spore coat provides “eat resistance”during phagocytic predation by the protozoan Tetrahymena thermophila. Proc Natl Acad Sci U SA. 2006; 103:165–170. [PubMed: 16371471]
Kuwana R, Kasahara Y, Fujibayashi M, Takamatsu H, Ogasawara N, Watabe K. Proteomicscharacterization of novel spore proteins of Bacillus subtilis. Microbiology. 2002; 148:3971–3982.[PubMed: 12480901]
Laaberki MH, Dworkin J. Role of spore coat proteins in the resistance of Bacillus subtilis spores toCaenorhabditis elegans predation. J Bacteriol. 2008; 190:6197–6203. [PubMed: 18586932]
Lai EM, Phadke ND, Kachman MT, Giorno R, Vazquez S, Vazquez JA, Maddock JR, Driks A.Proteomic analysis of the spore coats of Bacillus subtilis and Bacillus anthracis. J Bacteriol. 2003;185:1443–1454. [PubMed: 12562816]
Lambert EA, Popham DL. The Bacillus anthracis SleL (YaaH) protein is an N-acetylglucosaminidaseinvolved in spore cortex depolymerization. J Bacteriol. 2008; 190:7601–7607. [PubMed:18835992]
Levin PA, Fan N, Ricca E, Driks A, Losick R, Cutting S. An unusually small gene required forsporulation by Bacillus subtilis. Mol Microbiol. 1993; 9:761–771. [PubMed: 8231808]
Masayama A, Kuwana R, Takamatsu H, Hemmi H, Yoshimura T, Watabe K, Moriyama R. A novellipolytic enzyme, YcsK (LipC), located in the spore coat of Bacillus subtilis, is involved in sporegermination. J Bacteriol. 2007; 189:2369–2375. [PubMed: 17220230]
McCarter LL. Regulation of flagella. Curr Opin Microbiol. 2006; 9:180–186. [PubMed: 16487743]McPherson DC, Kim H, Hahn M, Wang R, Grabowski P, Eichenberger P, Driks A. Characterization of
the Bacillus subtilis spore morphogenetic coat protein CotO. J Bacteriol. 2005; 187:8278–8290.[PubMed: 16321932]
Naclerio G, Baccigalupi L, Zilhao R, De Felice M, Ricca E. Bacillus subtilis spore coat assemblyrequires cotH gene expression. J Bacteriol. 1996; 178:4375–4380. [PubMed: 8755863]
Nicholson WL, Munakata N, Horneck G, Melosh HJ, Setlow P. Resistance of Bacillus endospores toextreme terrestrial and extraterrestrial environments. Microbiol Mol Biol Rev. 2000; 64:548–572.[PubMed: 10974126]
Nicholson WL. Using thermal inactivation kinetics to calculate the probability of extreme sporelongevity: implications for paleomicrobiology and lithopanspermia. Orig Life Evol Biosph. 2003;33:621–631. [PubMed: 14601931]
Ozin AJ, Henriques AO, Yi H, Moran CP Jr. Morphogenetic proteins SpoVID and SafA form acomplex during assembly of the Bacillus subtilis spore coat. J Bacteriol. 2000; 182:1828–1833.[PubMed: 10714986]
Ozin AJ, Samford CS, Henriques AO, Moran CP Jr. SpoVID guides SafA to the spore coat in Bacillussubtilis. J Bacteriol. 2001; 183:3041–3049. [PubMed: 11325931]
Paredes CJ, Alsaker KV, Papoutsakis ET. A comparative genomic view of clostridial sporulation andphysiology. Nature Rev Microbiol. 2005; 3:969–978. [PubMed: 16261177]
Piggot PJ, Coote JG. Genetic aspects of bacterial endospore formation. Bacteriol Rev. 1976; 40:908–962. [PubMed: 12736]
Piggot, PJ.; Losick, R. Sporulation genes and intercompartmental regulation. In: Sonenshein, AL.;Hoch, JA.; Losick, R., editors. Bacillus subtilis and its closest relatives: from genes to cells.American Society for Microbiology; Washington, DC: 2002. p. 483-518.
Pogliano J, Osborne N, Sharp MD, Abanes-De Mello A, Perez A, Sun YL, Pogliano K. A vital stainfor studying membrane dynamics in bacteria: a novel mechanism controlling septation duringBacillus subtilis sporulation. Mol Microbiol. 1999; 31:1149–1159. [PubMed: 10096082]
Price KD, Losick R. A four-dimensional view of assembly of a morphogenetic protein duringsporulation in Bacillus subtilis. J Bacteriol. 1999; 181:781–790. [PubMed: 9922240]
Ragkousi K, Eichenberger P, van Ooij C, Setlow P. Identification of a new gene essential forgermination of Bacillus subtilis spores with Ca2+-dipicolinate. J Bacteriol. 2003; 185:2315–2329.[PubMed: 12644503]
Wang et al. Page 15
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Ramamurthi KS, Clapham KR, Losick R. Peptide anchoring spore coat assembly to the outer foresporemembrane in Bacillus subtilis. Mol Microbiol. 2006; 62:1547–1557. [PubMed: 17427285]
Ramamurthi KS, Losick R. ATP-driven self-assembly of a morphogenetic protein in Bacillus subtilis.Mol Cell. 2008; 31:406–414. [PubMed: 18691972]
Ramamurthi KS, Lecuyer S, Stone HA, Losick R. Geometric cue for protein localization in abacterium. Science. 2009; 323:1354–1357. [PubMed: 19265022]
Roels S, Driks A, Losick R. Characterization of spoIVA, a sporulation gene involved in coatmorphogenesis in Bacillus subtilis. J Bacteriol. 1992; 174:575–585. [PubMed: 1729246]
Sacco M, Ricca E, Losick R, Cutting S. An additional GerE-controlled gene encoding an abundantspore coat protein from Bacillus subtilis. J Bacteriol. 1995; 177:372–377. [PubMed: 7814326]
Setlow P. Spore germination. Curr Opin Microbiol. 2003; 6:550–556. [PubMed: 14662349]Sharp MD, Pogliano K. An in vivo membrane fusion assay implicates SpoIIIE in the final stages of
engulfment during Bacillus subtilis sporulation. Proc Natl Acad Sci U S A. 1999; 96:14553–14558. [PubMed: 10588743]
Skerra A. Alternative binding proteins: anticalins - harnessing the structural plasticity of the lipocalinligand pocket to engineer novel binding activities. FEBS J. 2008; 275:2677–2683. [PubMed:18435758]
Steichen CT, Kearney JF, Turnbough CL Jr. Non-uniform assembly of the Bacillus anthracisexosporium and a bottle cap model for spore germination and outgrowth. Mol Microbiol. 2007;64:359–367. [PubMed: 17493122]
Steil L, Serrano M, Henriques AO, Volker U. Genome-wide analysis of temporally regulated andcompartment-specific gene expression in sporulating cells of Bacillus subtilis. Microbiology.2005; 151:399–420. [PubMed: 15699190]
Stephenson K, Hoch JA. Evolution of signalling in the sporulation phosphorelay. Mol Microbiol.2002; 46:297–304. [PubMed: 12406209]
Sterlini JM, Mandelstam J. Commitment to sporulation in Bacillus subtilis and its relationship todevelopment of actinomycin resistance. Biochem J. 1969; 113:29–37. [PubMed: 4185146]
Stragier P, Losick R. Molecular genetics of sporulation in Bacillus subtilis. Annu Rev Genet. 1996;30:297–241. [PubMed: 8982457]
Takamatsu H, Kodama T, Nakayama T, Watabe K. Characterization of the yrbA gene of Bacillussubtilis, involved in resistance and germination of spores. J Bacteriol. 1999; 181:4986–4994.[PubMed: 10438771]
Tars K, Fridborg K, Bundule M, Liljas L. The three-dimensional structure of bacteriophage PP7 fromPseudomonas aeruginosa at 3.7-A resolution. Virology. 2000; 272:331–337. [PubMed: 10873776]
van Ooij C, Losick R. Subcellular localization of a small sporulation protein in Bacillus subtilis. JBacteriol. 2003; 185:1391–1398. [PubMed: 12562810]
van Ooij C, Eichenberger P, Losick R. Dynamic patterns of subcellular protein localization duringspore coat morphogenesis in Bacillus subtilis. J Bacteriol. 2004; 186:4441–4448. [PubMed:15231775]
Webb CD, Decatur A, Teleman A, Losick R. Use of green fluorescent protein for visualization of cell-specific gene expression and subcellular protein localization during sporulation in Bacillus subtilis.J Bacteriol. 1995; 177:5906–5911. [PubMed: 7592342]
Woods DR, Jones DT. Physiological responses of Bacteroides and Clostridium strains toenvironmental stress factors. Adv Microb Physiol. 1986; 27:1–64. [PubMed: 3532715]
Youngman P, Perkins JB, Losick R. Construction of a cloning site near one end of Tn917 into whichforeign DNA may be inserted without affecting transposition in Bacillus subtilis or expression ofthe transposon-borne erm gene. Plasmid. 1984; 12:1–9. [PubMed: 6093169]
Zhang J, Fitz-James PC, Aronson AI. Cloning and characterization of a cluster of genes encodingpolypeptides present in the insoluble fraction of the spore coat of Bacillus subtilis. J Bacteriol.1993; 175:3757–3766. [PubMed: 8509331]
Zheng LB, Donovan WP, Fitz-James PC, Losick R. Gene encoding a morphogenic protein required inthe assembly of the outer coat of the Bacillus subtilis endospore. Genes Dev. 1988; 2:1047–1054.[PubMed: 3139490]
Wang et al. Page 16
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1. Genetic requirements for the subcellular localization of VID-GFP, VM-GFP and IVA-GFPCells were sporulated at 37 °C and imaged at the indicated times after suspension in SMmedium. Cells were stained with membrane stain (FM4-64) and analyzed by fluorescencemicroscopy. A group of representative cells is shown for each fusion. Images consist ofoverlays of GFP (green) on membranes (red) (A) Subcellular localization of VID-GFP insporulating cells. First row: VID-GFP in an otherwise wild type strain (PE654). Second row:VID-GFP in a IVA::neo strain (PE710). Third row: VID-GFP in a VM::erm strain (KW254)(B) Subcellular localization of VM-GFP in sporulating cells. First row: VM-GFP in anotherwise wild type strain (CVO1000). Second row: VM-GFP in a IVA::neo strain(PE2198). Third row: VM-GFP in a VID::kan strain (PE1469) (C) Subcellular localizationof IVA-GFP in sporulating cells. First row: IVA-GFP in an otherwise wild type strain(PE1022). Second row: IVA-GFP in a VM::erm strain (PE1026). Third row: IVA-GFP in aVID::kan strain (PE1166). (D) Schematic representation of the genetic interactions betweenIVA, VM and VID. The first step of the subcellular localization process, the targeting to theforespore surface is controlled by IVA. The second step, spore encasement, is controlled byVM and VID.
Wang et al. Page 17
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
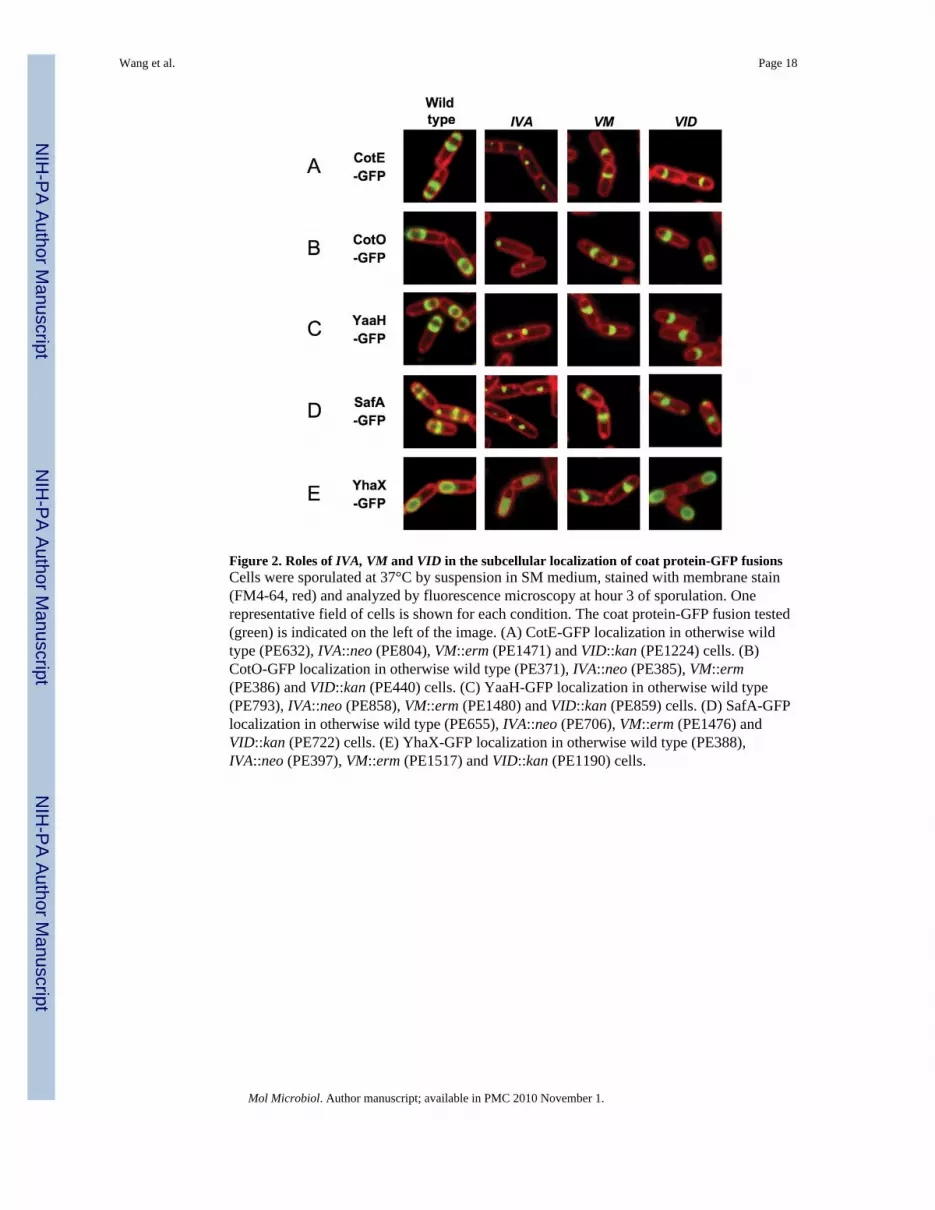
Figure 2. Roles of IVA, VM and VID in the subcellular localization of coat protein-GFP fusionsCells were sporulated at 37°C by suspension in SM medium, stained with membrane stain(FM4-64, red) and analyzed by fluorescence microscopy at hour 3 of sporulation. Onerepresentative field of cells is shown for each condition. The coat protein-GFP fusion tested(green) is indicated on the left of the image. (A) CotE-GFP localization in otherwise wildtype (PE632), IVA::neo (PE804), VM::erm (PE1471) and VID::kan (PE1224) cells. (B)CotO-GFP localization in otherwise wild type (PE371), IVA::neo (PE385), VM::erm(PE386) and VID::kan (PE440) cells. (C) YaaH-GFP localization in otherwise wild type(PE793), IVA::neo (PE858), VM::erm (PE1480) and VID::kan (PE859) cells. (D) SafA-GFPlocalization in otherwise wild type (PE655), IVA::neo (PE706), VM::erm (PE1476) andVID::kan (PE722) cells. (E) YhaX-GFP localization in otherwise wild type (PE388),IVA::neo (PE397), VM::erm (PE1517) and VID::kan (PE1190) cells.
Wang et al. Page 18
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3. CotE-GFP fluorescent ring formation is dependent on VIDCells were sporulated at 37°C and imaged at the indicated times after suspension in SMmedium. Cells were stained with membrane stain (FM4-64) and analyzed by fluorescencemicroscopy. A few representative cells are shown for each fusion. First row: CotE-GFPfluorescence. Second row: fluorescence of membranes. Third row: overlay of CotE-GFP(green) and membranes (red). (A) CotE-GFP fusion in an otherwise wild type strain(PE632). (B) CotE-GFP fusion in a VID::kan strain (PE1224). (C) CotE-GFP fusion in aIVA::neo strain (PE804).
Wang et al. Page 19
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 4. The N-domain of VID-CFP is necessary for subcellular localization of CotE-YFPCells were sporulated at 37°C and imaged four hours after suspension in SM medium.Samples were stained with membrane stain (FM4-64) and analyzed by fluorescencemicroscopy. The VID-CFP deletion constructs are described on the left panel. The N-domain (N, dark blue) includes residues 1 to 137; the middle region (M, light blue) isdelimited by residues 138 to 499; the C-terminal region has two components, region A (lightpurple, a.a. 500 to 524) and the previously identified LysM domain (dark purple, a.a. 525 to575) (Costa et al., 2006). The first column shows the VID-CFP fluorescence, the secondcolumn the fluorescence of membranes. The third column corresponds to the CotE-YFPfluorescence in the same cells. The fourth column is an overlay of the CotE-YFPfluorescence (green) on the cells stained with FM4-64 (red). The first row is KW363(VID::kan amyE::VID-cfp cat cotEΩcotE-yfp spc), the second row is PE1964 (VID::kanamyE::VIDΔ24-136-cfp cat cotEΩcotE-yfp spc), the third row is PE1925 (VID::kanamyE::VIDΔ86-136-cfp cat cotEΩcotE-yfp spc), the fourth row is PE2094 (VID::kanamyE::VIDΔ125-136-cfp cat cotEΩcotE-yfp spc).
Wang et al. Page 20
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 5. The C-terminal region of VID-CFP is necessary for initial localization to the foresporesurfaceCells were sporulated at 37°C and imaged four hours after suspension in SM medium.Samples were stained with membrane stain (FM4-64) and analyzed by fluorescencemicroscopy. The VID-CFP constructs are described on the left panel. Each region issymbolized by a different color according to the code established in Fig. 4. The VID-CFPfluorescence is reported in the first column. The second column shows the fluorescence ofthe membranes. The third column corresponds to the CotE-YFP fluorescence in the samecells. The fourth column is an overlay of the CotE-YFP fluorescence (green) on the cellsstained with FM4-64 (red). The first row is KW363 (VID::kan amyE::VID-cfp catcotEΩcotE-yfp spc), the second row is PE2088 (VID::kan amyE::VID1-525-cfp catcotEΩcotE-yfp spc), the third row is PE2087 (VID::kan amyE::VID1-500-cfp cat cotEΩcotE-yfp spc), the fourth row is PE1981 (VID::kan amyE::VID1-484-cfp cat cotEΩcotE-yfp spc),the fifth row is PE2093 (VID::kan amyE::VIDΔ500-525-cfp cat cotEΩcotE-yfp spc), the sixthrow is PE2004 (VID::kan amyE::VIDΔ484-500-cfp cat cotEΩcotE-yfp spc).
Wang et al. Page 21
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
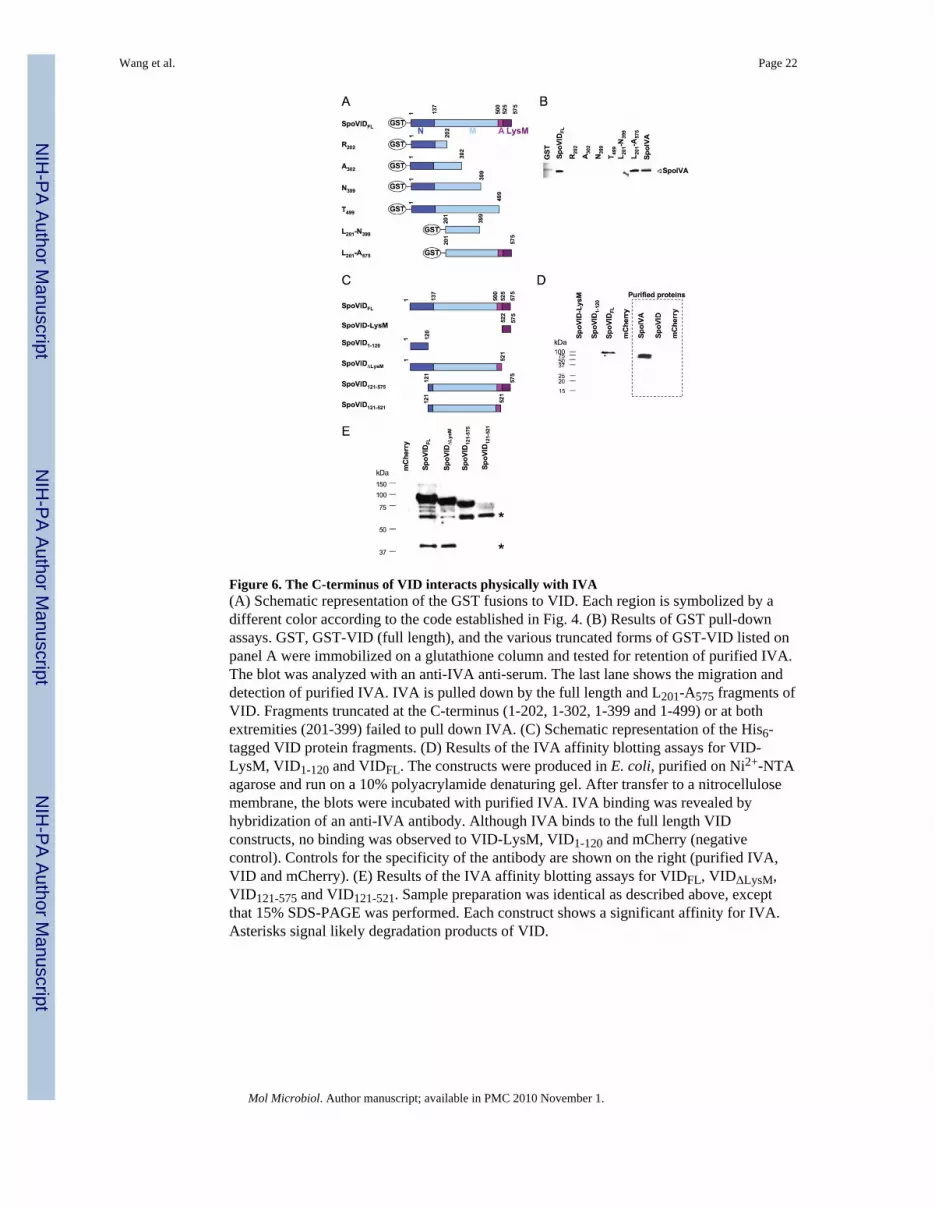
Figure 6. The C-terminus of VID interacts physically with IVA(A) Schematic representation of the GST fusions to VID. Each region is symbolized by adifferent color according to the code established in Fig. 4. (B) Results of GST pull-downassays. GST, GST-VID (full length), and the various truncated forms of GST-VID listed onpanel A were immobilized on a glutathione column and tested for retention of purified IVA.The blot was analyzed with an anti-IVA anti-serum. The last lane shows the migration anddetection of purified IVA. IVA is pulled down by the full length and L201-A575 fragments ofVID. Fragments truncated at the C-terminus (1-202, 1-302, 1-399 and 1-499) or at bothextremities (201-399) failed to pull down IVA. (C) Schematic representation of the His6-tagged VID protein fragments. (D) Results of the IVA affinity blotting assays for VID-LysM, VID1-120 and VIDFL. The constructs were produced in E. coli, purified on Ni2+-NTAagarose and run on a 10% polyacrylamide denaturing gel. After transfer to a nitrocellulosemembrane, the blots were incubated with purified IVA. IVA binding was revealed byhybridization of an anti-IVA antibody. Although IVA binds to the full length VIDconstructs, no binding was observed to VID-LysM, VID1-120 and mCherry (negativecontrol). Controls for the specificity of the antibody are shown on the right (purified IVA,VID and mCherry). (E) Results of the IVA affinity blotting assays for VIDFL, VIDΔLysM,VID121-575 and VID121-521. Sample preparation was identical as described above, exceptthat 15% SDS-PAGE was performed. Each construct shows a significant affinity for IVA.Asterisks signal likely degradation products of VID.
Wang et al. Page 22
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 7. Model for spore coat assembly and spore encasementSpore coat assembly can be subdivided into two consecutive stages: initial localization ofspore coat proteins to the forespore surface and spore encasement. The amphipathic α helicalVM (green) recognizes positively curved membranes and interacts directly with the C-terminus of the coat morphogenetic protein IVA (red). This complex is responsible for therecruitment to the spore surface of every coat protein (orange), as well as VID (blue andpurple). Two fragments of VID are necessary for recruitment, region A (light purple), whichinteracts directly with IVA, and the LysM domain (dark purple). In the encasement stage,the morphogenetic N-terminal domain of VID (dark blue) promotes multimerization of coatproteins around the nascent spore, either by a direct interaction or via IVA and othermorphogenetic proteins. The multimerization process continues until the spore has beenfully encased.
Wang et al. Page 23
Mol Microbiol. Author manuscript; available in PMC 2010 November 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript





























