Embed Size (px)
Citation preview

TTr
Ja
b
c
d
a
ARR1AA
KACCDGS
1
hycaopeb
d(ri
BG
0h
Plant Science 215– 216 (2014) 39– 47
Contents lists available at ScienceDirect
Plant Science
jo ur nal home p age: www.elsev ier .com/ locate /p lantsc i
he cell-wall glycoproteins of the green alga Scenedesmus obliquus.he predominant cell-wall polypeptide of Scenedesmus obliquus iselated to the cell-wall glycoprotein gp3 of Chlamydomonas reinhardtii
ürgen Voigta,∗, Adam Stolarczykb, Maria Zychb, Przemysław Malecc, Jan Burczykb,d
Institute for Biochemistry, Charité – Universitätsmedizin Berlin, D-10117 Berlin, GermanyDepartment of Pharmacognosy and Phytochemistry, Silesian Medical University, 41-200 Sosnowiec, PolandDepartment of Plant Physiology and Biochemistry, Jagiellonian University, 30-387 Kraków, PolandLaboratory of Biotechnology, 43-400 Cieszyn, Poland
r t i c l e i n f o
rticle history:eceived 20 April 2013eceived in revised form2 September 2013ccepted 21 October 2013vailable online 29 October 2013
eywords:ntibodies
a b s t r a c t
The green alga Scenedesmus obliquus contains a multilayered cell wall, ultrastructurally similar to that ofChlamydomonas reinhardtii, although its proportion of hydroxyproline is considerably lower. Therefore,we have investigated the polypeptide composition of the insoluble and the chaotrope-soluble wall frac-tions of S. obliquus. The polypeptide pattern of the chaotrope-soluble wall fraction was strongly modifiedby chemical deglycosylation with anhydrous hydrogen fluoride (HF) in pyridine indicating that most ofthese polypeptides are glycosylated. Polypeptide constituents of the chaotrope-soluble cell-wall fractionwith apparent molecular masses of 240, 270, 265, and 135 kDa cross-reacted with a polyclonal anti-body raised against the 100 kDa deglycosylation product of the C. reinhardtii cell-wall glycoprotein GP3B.
ell wallhlamydomonas reinhardtiieglycosylationlycoproteinscenedesmus obliquus
Chemical deglycosylation of the chaotrope-soluble wall fraction resulted in a 135 kDa major polypep-tide and a 106 kDa minor component reacting with the same antibody. This antibody recognized specificpeptide epitopes of GP3B. When the insoluble wall fraction of S. obliquus was treated with anhydrousHF/pyridine, three polypeptides with apparent molecular masses of 144, 135, and 65 kDa were solubi-lized, which also occured in the deglycosylated chaotrope-soluble wall fraction. These findings indicatethat theses glycoproteins are cross-linked to the insoluble wall fraction via HF-sensitive bonds.
. Introduction
Microalgae, especially those belonging to the Chlorococcales,ave been traditionally used for aquatic feed, and in the recentears, they also have received renewed attention on account of theirapacity to offer numerous value-added products [1,2], and to acts cell factories for the production of biofuels [3,4]. Scenedesmusbliquus, like other members of the Chlorococcales, contains high
roportions of proteins and lipids [1]. S. obliquus also possessesxcellent ability of wastewater treatment, CO2 sequestration andiodiesel production [5–9].Abbreviations: dGP100, deglycosylated glycoprotein (final Mr 100 kDa); dICW,eglycosylated insoluble cell wall fraction; GP1, cell wall glycoprotein number 1Chlamydomonas reinhardtii); GP2, cell wall glycoprotein number 2 (Chlamydomonaseinhardtii); GP3, cell wall glycoprotein number 3 (Chlamydomonas reinhardtii); Ig,mmunoglobuline; IgG, immunoglobuline class G; PVDF, polyvinylidene fluoride.∗ Corresponding author at: Charité – Universitätsmedizin Berlin, Institute foriochemistry, Charité-Platz 1/Virchowweg 6 (CharitéCrossOver), D-10117 Berlin,ermany. Tel.: +49 30 450528045; fax: +49 30 450528942.
E-mail address: [email protected] (J. Voigt).
168-9452/$ – see front matter © 2013 Elsevier Ireland Ltd. All rights reserved.ttp://dx.doi.org/10.1016/j.plantsci.2013.10.011
© 2013 Elsevier Ireland Ltd. All rights reserved.
A strong barrier on the way of the utilization of the componentsproduced by S. obliquus and other members of the Chlorococcales isthe high resistance of their cell walls exhibiting a characteristic tril-aminar structure [10,11]. The trilaminar structure of the outer walllayers is resistant to treatment with a number of lytic enzymes likecellulases, hemicellulases, lysozyme, pronase and other hydrolasesas well as to drastic non-oxidative chemical treatments [10,12,13].The cell walls of S. obliquus strain 633 and many other membersof the Chlorococcales contain cellulose in the inner wall layersand insoluble, acetolysis-resistant, lipid-containing biopolymerstermed algaenans, which are localized in the trilaminar outer layersand contribute to the rigidity of these cell walls [11,14–17]. Otherconstituent of these rigid cell walls are glucosamine-containingbiopolymers and glycoproteins [18]. Insoluble hydroxyproline-containing glycoproteins are constituents of the cell walls of allhigher plants studied so far [19,20] and have been also found in theinsoluble cell-wall fraction of several green algae [21]. The insoluble
wall fraction of Chlamydomonas reinhardtii and other members ofthe Volvocales exclusively consist of cross-linked hydroxyproline-containing glycoproteins [22–24]. In the case of S. obliquus as forother Chlorococcales, however, there are no data concerning the cell
4 ce 21
wds3aomtt
2
2
mP2Ihw
2c
buhtct
2
mBog
2
dhw1dstw5sb
2
wwu2wc
0 J. Voigt et al. / Plant Scien
all glycoproteins apart from the amino acid composition of theifferent wall preparations [18]. Analysis of the amino acid compo-ition of cell wall of S. obliquus indicated a protein content of about0% [18]. Interestingly, differences were observed with respect tomino acid composition of cell walls prepared from homogenatesf living cells and maternal cell walls accumulated in the cultureedium [18]. Therefore, we have studied the cell-wall polypep-
ides of S. obliquus. The results of this investigation are shown inhe present communication.
. Materials and methods
.1. Algal strain and growth condition
The Scenedesmus obliquus var. alternas Christjuk, strain 633 isaintained in authors collection (Prof. Jan Burczyk, 43-400 Cieszyn,
oland). Algae were grown in medium I of [25] supplemented with.5 g/L each of glucose and sucrose as previously described [15,26].n order to isolate maternal cell walls and complete cell walls fromomogenates of mechanically disrupted cells, 30-day-old culturesere harvested by centrifugation.
.2. Cell harvesting and isolation of maternal cell walls from theulture medium
After centrifuging the culture for 10 min at 2400 × g, the greenottom layer of the sediment (algal cells) was covered with a pinkpper layer consisting of the maternal cell walls and could bearvested with a spatula [10]. Material collected from several cen-rifuge tubes was combined, suspended in deionized water andentrifuged again. This washing step was repeated several timeso obtain the pure maternal cell walls.
.3. Desintegration of algal cells
Algal cells, delivered from maternal cell wall material, wereechanically disrupted with a rotary desintegrator Type MSK
raun (Melsungen, Germany) cooled with a CO2 stream. Aliquotsf 10 g wet algal sediment were homogenized for 6 min with 50 glass microbeads (Ø 0.35 mm).
.4. Isolation of cell walls
Homogenization and all subsequent steps were performed inistilled water [26]. The microbeads were removed from theomogenates by a glass Schott-filter G-1 and washed with distilledater. The final volume of 100–150 ml filtrate was centrifuged for
0 min at 2400 × g. The sediment was washed several times withistilled water. If the sediment was still green, the desintegrationtep (see above) was repeated. For removing starch grains fromhe sample (visible as a white layer in the sediment), the sedimentas resuspended in distilled water and centrifuged for 10 min at
90 × g. Under these conditions, the starch grains were removed byedimentation. The cell walls were collected from the supernatanty centrifugation at 2400 × g for 10 min.
.5. Isolation of the chaotrope-soluble cell-wall fraction
The chaotrope-soluble cell-wall components were extractedith aqueous LiCl (3 M) from isolated cell walls. Purified cell wallsere suspended in deionized water, mixed with the same vol-
me of aqueous LiCl (6 M), stirred for 2 h at 4 ◦C and centrifuged at0,000 × g for 30 min. The sediment was resuspended in deionizedater and the treatment with aqueous LiCl was twice repeated. Theombined supernatants were desalted by dialysis for 2 days against
5– 216 (2014) 39– 47
deionized water containing 1 mM 2-mercaptoethanol with severalchanges and finally freeze-dried.
2.6. Purification of the insoluble cell-wall fraction
The insoluble cell-wall fraction was prepared from the LiCl-extracted cell walls by treatment with several detergent containingbuffers [27] to remove all SDS-soluble polypeptides and, finally,with deionized water to remove the detergents. To quantitativelyremove residual SDS-soluble polypeptides, the LiCl-extracted cellwalls were treated overnight at room temperature with urea-SDS buffer [28] containing 8 M urea, 2% (w/v) SDS, 200 mM2-mercatoethanol, 10 mM EDTA and 20 mM Tris–HCl (pH 7.5) fol-lowed by centrifugation at 20,000 × g for 60 min. This step wasrepeated at least twice and the supernatants checked for proteinby SDS-PAGE and subsequent silver-staining of the gels. After com-plete extraction of SDS-soluble polypeptides, the cell walls werewashed twice with 2 M NaCl–3% (w/v) sodium deoxycholate–3%(v/v) Triton X-100–20 mM Tris–HCl (pH 7.5), twice with deion-ized water containing 1% (v/v) Triton X-100 and several times withdeionized water. Finally, the purified insoluble wall fraction wasfreeze-dried.
2.7. Chemical deglycosylation
Deglycosylation of cell wall glycoproteins was performed bytreatment with hydrofluoride (HF)-saturated dry pyridine (Fluka,Neu-Ulm, Germany) as previously described [27,29].
2.8. Determination of protein
Protein was determined using the bicinchoninic acid (BCA)method [30]. Prior to the determination of protein, interferingsubstances like 2-mercaptoethanol and EDTA were eliminated byprecipitation with trichloroacetic acid (TCA) in the presence ofdeoxycholate [31]. The assay was performed in disposable micro-centrifuge tubes (1.5 ml size). Upon addition of protein samples,the final volume was adjusted to 1.0 ml with deionized water and0.1 ml of sodium deoxycholate (0.15%, w/v) was added. After stand-ing at room temperature for 10 min, 0.1 ml of trichloroacetic acid(72%, w/v) was added. Samples were vortexed and centrifuged at3000 × g for 15 min at room temperature. The supernatant wasaspired off and the pellet washed twice with deionized water. Pel-lets were solubilized by vortexing in 50 �l sodium dodecyl sulfate(SDS) containing 0.1 M NaOH. Immediately after adding the BCAreagent (1.0 ml), each sample was vortexed and incubated at 37 ◦Cfor 30 min. Finally, the optical density was measured at 562 nm.
2.9. SDS-PAGE and protein gel blot analyses
Polypeptides were precipitated by addition of trichloroaceticacid (final concentration 10%, w/v) in the cold and the precipitatescollected by centrifugation. The pellets were washed twice withdistilled water and redissolved in a small volume of urea-SDS buffer[28]. The polypeptides were separated by SDS-PAGE according toLaemmli [32]. After electrophoresis, the gels were either stainedwith Coomassie Brilliant Blue G250 or blotted electrophoreti-cally onto polyvinylidene fluoride (PVDF) membranes (Porablot,Macherey-Nagel, Düren, Germany) as described by Towbin et al.[33]. The protein gel blots were probed with polyclonal antibodiesraised in rabbits against different deglycosylated Chlamydomonas
cell-wall polypeptides and subsequent treatment with alkalinephosphatase-coupled goat anti-rabbit IgG antibodies (Amersham-Buchler, Braunschweig, Germany) as previously described [34].The indirectly bound alkaline phosphatase was detected by its
ce 215– 216 (2014) 39– 47 41
es
2
ttAswacGewide4(
2
sd1stmoabcasb5(bupfITtmmsoIU
3
3
ccPltwe
Fig. 1. SDS-PAGE analysis of the chaotrope-soluble cell wall polypeptides of S.obliquus. The chaotrope-soluble wall constituents extracted from purified cell wallsby treatment with 3 M LiCl were fractionated by SDS-PAGE on gel slabs containing12% (w/v) acrylamide and subsequently stained with Coomassie Brilliant Blue G250.Lane 1: untreated polypeptides extracted from purified cell walls of living cells cor-responding to 25 �g of protein; lanes 2 and 3: untreated polypeptides extracted frompurified maternal cell walls corresponding to 10 (lane 2) and 5 �g of protein (lane 3);
J. Voigt et al. / Plant Scien
nzymatic activity using 5-bromo-4-chloro-3-indolyl phosphate asubstrate in the presence of nitroblue-tetrazolium chloride [35].
.10. Epitope analysis
A total of 281 overlapping pentadeca peptides representinghe whole amino acid sequence of the Chlamydomonas GP3B pro-ein derived from the ORF of the GP3B cDNA (Accession No.M260992) were generated by spot synthesis using a celluloseheet as solid support [36]. These overlapping pentadeca peptidesere used to determine the epitope specificity of the polyclonal
nti-dGP100 antibody raised in rabbits against the 100 kDa degly-osylation product of the chaotrope-soluble cell-wall glycoproteinP3B of C. reinhardtii. After incubation with the antibody andxtensive washing, bound rabbit IgG were measured by incubationith alkaline phosphatase-coupled goat anti-rabbit IgG antibod-
es (Amersham-Buchler, Braunschweig, Germany) and subsequentetection of the indirectly bound alkaline phosphatase via itsnzyme activity as previously described [29,34] using 5-bromo--chloro-3-indolyl phosphate as substrate in the presence of MTT3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl-tetrazolium bromide).
.11. Immunofluorescence
100 �L of maternal cell wall preparation (S. obliquus strain 633)uspended initially in 20% ethanol were spun down in the Eppen-orf tube with a cooled centrifuge (Eppendorf 5418R; 10,000 × g,
min). The pellet was resuspended in 1 mL of PBS (pH 7.2) andubsequently washed 3 times with 1 mL of the fresh PBS solu-ion. The final, diluted cell wall suspension was dispersed on
icroscope slides coated with chrome-alum gelatin [37] and driedvernight at 4 ◦C. An indirect immunostaining was performed atmbient temperature (25 ◦C) according to the protocol describedy Heslop-Harrison and Heslop-Harrison [38] with some modifi-ations. Initially, the slides were washed 3 times for 10 min with
gentle shaking in PBS to remove an unbound material. Then, thepecimen was incubated for 1 h with 1% (w/v) of BSA (Sigma) tolock an unspecific antibody binding and subsequently washed 3×
min with PBS. The 45 min incubation with primary antibodiesanti-dGP100 or anti-dICW rabbit serum diluted 1:20) was followedy 3× 5 min PBS washing. Control preparations were performedsing a normal rabbit serum (IBSS Biomed, Kraków, Poland) inlace of the primary antibody. Then, the material was incubatedor 45 min with a secondary antibody (FITC-labelled anti-rabbitgG, Sigma F0382, dilution 1:50) and washed 3× 5 min with PBS.o avoid fluorescence quenching, the specimen was covered withhe mounting medium (90% (w/v) of PBS-buffered glycerol supple-
ented with p-phenylenodiamine in concentration of 0.1 mg/mL),ounted under cover slides and stored at 4 ◦C in darkness. The
lides were examined under Nikon Eclipse TE-200 inverted flu-rescence microscope equipped with an RGB digital camera andmage-Pro Plus software (Media Cybernetics Inc., Rockville MD,SA).
. Results
.1. The chaotrope-soluble cell-wall polypeptides of S. obliquus
Both cell walls prepared from homogenates of living S. obliquusells and mother-cell walls accumulated in the culture mediumontained chaotrope-soluble polypeptides as revealed by SDS-AGE analyses of extracts obtained by treatment with 3 M
ithium chloride (LiCl) (Fig. 1, lanes 1–3). The polypeptide pat-erns of the chaotrope-soluble wall fractions of these two cellall types were, however, rather different. Mother-cell wallsxclusively contained high-molecular-weight components with
lane 4: chemically deglycosylated polypeptides extracted from purified cell walls ofliving cells corresponding to 25 �g of protein; lanes 5 and 6: chemically deglyco-sylated polypeptides extracted from purified maternal cell walls corresponding to10 �g (lane 5) and 5 �g of protein (lane 6); lane 7: protein molecular mass markers.
apparent molecular masses >150 kDa (Fig. 1, lanes 2 and 3). Thesehigh-molecular-weight components also occurred in the cell wall ofliving cells, which additionally contained prominent band of about40 kDa and around 100 kDa (Fig. 1, lane 1).
Chemical deglycosylation with hydrogen fluoride (HF)-saturated dry pyridine increased the electrophoretic mobility ofthese chaotrope-soluble polypeptides (Fig. 1, lanes 4–6) indicatingthat these chaotrope-soluble polypeptides (Fig. 1, lanes 1–3) areglycoproteins. However, the 40 kDa component (Fig. 1, lane 1)was apparently lost during either chemical deglycosylation or thesubsequent purification of the deglycosylation products (Fig. 1,lane 4).
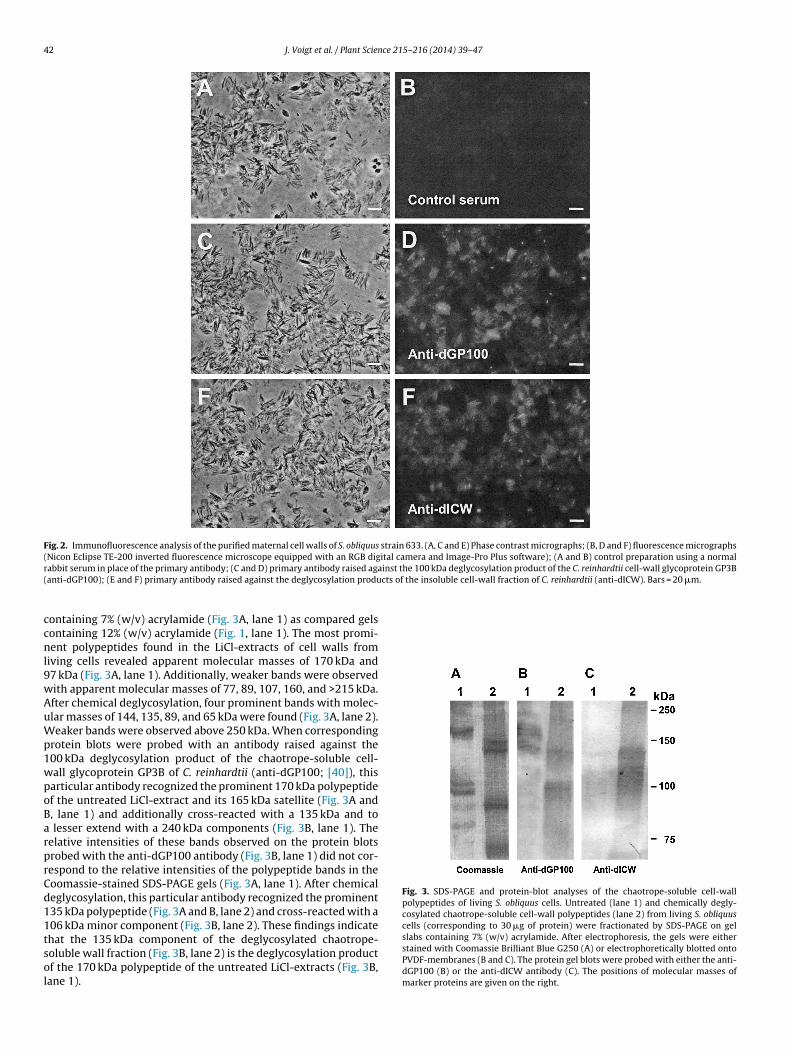
3.2. Immunofluorescence
As previously reported, polyclonal antibodies raised againstdeglycosylation products of C. reinhardtii cell wall polypeptidesreact with the cell surface of this particular green alga [39].Therefore, the finding that the cell wall of S. obliquus containschaotrope-soluble glycoproteins (Fig. 1) inspired us to analyze theisolated cell walls of S. obliquus for cross-reactivity with these par-ticular antibodies. As shown by immunofluorescence studies, theisolated Scenedesmus cell walls reacted specifically with antibodiesagainst the 100 kDa deglycosylation product of the chaotrope-soluble, cell-wall glycoprotein GP3B of C. reinhardtii (anti-dGP100;Fig. 2C and D) and with antibodies raised against the degly-cosylation products of the purified insoluble wall fraction of C.reinhardtii (Fig. 2E and F). Only rather faint fluorescence, however,was observed, when the primary antibodies were substituted byunspecific rabbit serum (control; Fig. 2A and B). These findingsindicate that the cell wall polypeptides of S. obliquus are relatedto the polypeptide backbones of the Chlamydomonas cell-wall con-stituents.
3.3. SDS-PAGE and protein-blot analyses of the chaotrope-soluble
cell wall polypeptidesA strongly improved separation of the high-molecular-weight,chaotrope-soluble polypeptides was obtained by SDS-PAGE on gels
42 J. Voigt et al. / Plant Science 215– 216 (2014) 39– 47
Fig. 2. Immunofluorescence analysis of the purified maternal cell walls of S. obliquus strain 633. (A, C and E) Phase contrast micrographs; (B, D and F) fluorescence micrographs(Nicon Eclipse TE-200 inverted fluorescence microscope equipped with an RGB digital camera and Image-Pro Plus software); (A and B) control preparation using a normalr inst the 100 kDa deglycosylation product of the C. reinhardtii cell-wall glycoprotein GP3B( cts of the insoluble cell-wall fraction of C. reinhardtii (anti-dICW). Bars = 20 �m.
ccnl9wAuWp1wpoBarprCd11tsol
Fig. 3. SDS-PAGE and protein-blot analyses of the chaotrope-soluble cell-wallpolypeptides of living S. obliquus cells. Untreated (lane 1) and chemically degly-cosylated chaotrope-soluble cell-wall polypeptides (lane 2) from living S. obliquuscells (corresponding to 30 �g of protein) were fractionated by SDS-PAGE on gel
abbit serum in place of the primary antibody; (C and D) primary antibody raised agaanti-dGP100); (E and F) primary antibody raised against the deglycosylation produ
ontaining 7% (w/v) acrylamide (Fig. 3A, lane 1) as compared gelsontaining 12% (w/v) acrylamide (Fig. 1, lane 1). The most promi-ent polypeptides found in the LiCl-extracts of cell walls from
iving cells revealed apparent molecular masses of 170 kDa and7 kDa (Fig. 3A, lane 1). Additionally, weaker bands were observedith apparent molecular masses of 77, 89, 107, 160, and >215 kDa.fter chemical deglycosylation, four prominent bands with molec-lar masses of 144, 135, 89, and 65 kDa were found (Fig. 3A, lane 2).eaker bands were observed above 250 kDa. When corresponding
rotein blots were probed with an antibody raised against the00 kDa deglycosylation product of the chaotrope-soluble cell-all glycoprotein GP3B of C. reinhardtii (anti-dGP100; [40]), thisarticular antibody recognized the prominent 170 kDa polypeptidef the untreated LiCl-extract and its 165 kDa satellite (Fig. 3A and, lane 1) and additionally cross-reacted with a 135 kDa and to
lesser extend with a 240 kDa components (Fig. 3B, lane 1). Theelative intensities of these bands observed on the protein blotsrobed with the anti-dGP100 antibody (Fig. 3B, lane 1) did not cor-espond to the relative intensities of the polypeptide bands in theoomassie-stained SDS-PAGE gels (Fig. 3A, lane 1). After chemicaleglycosylation, this particular antibody recognized the prominent35 kDa polypeptide (Fig. 3A and B, lane 2) and cross-reacted with a06 kDa minor component (Fig. 3B, lane 2). These findings indicate
hat the 135 kDa component of the deglycosylated chaotrope-oluble wall fraction (Fig. 3B, lane 2) is the deglycosylation productf the 170 kDa polypeptide of the untreated LiCl-extracts (Fig. 3B,ane 1).slabs containing 7% (w/v) acrylamide. After electrophoresis, the gels were eitherstained with Coomassie Brilliant Blue G250 (A) or electrophoretically blotted ontoPVDF-membranes (B and C). The protein gel blots were probed with either the anti-dGP100 (B) or the anti-dICW antibody (C). The positions of molecular masses ofmarker proteins are given on the right.

J. Voigt et al. / Plant Science 215– 216 (2014) 39– 47 43
Fig. 4. Determination of the epitope-specificity of the anti-dGP100 antibody by scanpeptide analysis. (A) Schematic representation of the amino acid sequence of thechaotrope-soluble Chlamydomonas cell-wall polypeptide GP3B. (Hydroxy)proline-rich domains are indicated by black boxes. The putative leader sequence isrepresented by a white box. Numbers indicate the positions of pentadeca pep-tides and amino acids from the N-terminus of the polypeptide. (B) 281 overlappingpentadeca peptides derived from the ORF of the GP3B cDNA (EMBL NucleotideSequence Database Accession No. AM260992) and representing the whole aminoacid sequence of GP3B were synthesized by spot synthesis using cellulose paperas solid support (Frank 1992). After treatment with blocking solution, the cellulose-bound scan peptides were incubated with the anti-dGP100 antibody. After extensivewap
brcaLwosCww2or
3
1gtdrwasyhecbppp
Table 1Amino acid sequences recognized by the anti-dGP100 antibody.
Peptide no.a Amino acid sequenceb
56 QITTLGYRRGRSPVA59–61 GRSPVAYWVPHDIVV65–67 DNPTGYVQLIDRIIL76–78 FTGLRPLLEGNS117 TNTLTFLKTSVITVS146–148 VAFPNGFQDYLNANPELSIL187 LNLGGVYTVRQQMPI208–210 AGAAAVSVYVGLNREPEAALF217 FNVTLPSGVSTPATA228–229 MNGFFSRIAFSGTPM242 FAVLPGAQFLPSFIP
a Cross-reacting pentadeca peptide identified by epitope analysis of the anti-
ashing, bound IgG were detected by incubation with alkaline phosphatase-couplednti-rabbit-IgG Igs and subsequent visualization of the indirectly bound alkalinehosphatase as described in Section 2.
When corresponding protein blots were probed with an anti-ody raised against the mixture of polypeptides, which wereeleased from the insoluble wall fraction of C. reinhardtii byhemical deglycosylation with HF/pyridine (anti-dICW; [29]), only
faint band of 135 kDa was detected in case of the untreatediCl-extracts (Fig. 3C, lane 1). After chemical deglycosylationith HF/pyridine, however, several cross-reacting bands were
bserved (Fig. 3C, lane 2) the prominent one obviously corre-ponding to the prominent 135 kDa polypeptides observed in theoomassie-stained gels (Fig. 3A, lane 2). Strong cross-reactivityith the anti-dICW antibody was also observed for several bandsith molecular masses between 98 and 120 kDa (Fig. 3C, lane
). In the corresponding Coomassie-stained gels (Fig. 3A, lane 2),nly minor components could be detected in this molecular massange.
.4. Specificities of the antibodies
The specificity of the anti-dGP100 antibody raised against the00 kDa deglycosylation product of the chaotrope-soluble cell-walllycoprotein GP3B of C. reinhardtii [40] was determined by epi-ope analysis using overlapping pentadeca peptides, which areerived from the amino acid sequence of GP3B and, together,epresent the whole amino acid sequence of this particular cell-all polypeptide (Fig. 4A and B). These pentadeca peptides with
n overlap of 3 amino acid residues were synthesized on a solidupport and exclusively contained proline instead of hydrox-proline residues occuring in the mature polypeptide due toydroxylation by prolyl 4-hydroxylase [40–42]. Primary sequencepitopes usually occur in several overlapping pentadeca peptidesontaining the same sequence motif, which correspond to neigh-
ouring spots on the solid support (Fig. 4B). Single pentadecaeptides binding the antibody without cross-reaction of overlap-ing peptides indicate that specific secondary structures of thesearticular peptides are recognized by the antibody. In the casedGP100 antibody (see Fig. 4).b The underlined sequences occur in two or more overlapping, crossreacting pen-
tadeca peptides and contain a primary structure epitope.
of anti-dGP100, a rather low number of epitopes are recognizedby this particular antibody (Fig. 4B), which are summarized inTable 1.
The anti-dICW antibody raised against the mixture of polypep-tides released from the insoluble wall fraction of C. reinhardtiiby chemical deglycosylation with HF/pyridine reacts with a largenumber of epitopes of the various constituents of the insolublewall fraction [29,34,40,43]. It also reacts with the hydroxyproline-rich domains of the deglycosylated chaotrope-soluble cell-wallglycoproteins of C. reinhardtii. In the case of the glycosylatedpolypeptides, these domains are not recognized by this antibodydue to glycosylation of the hydroxyproline residues (compareFig. 4A and B). Post-translational hydroxylation of proline residuesis catalyzed by prolyl 4-hydroxylase and occurs exclusively inproline-rich domains [41,42,44,45]. Prior to hydroxylation by pro-lyl 4-hydroxylase, these proline-rich sequences do not cross-reactwith the anti-dICW antibody [34,40,43]. The proline-rich sequencesof the chaotrope-soluble cell-wall glycoproteins GP1, GP2, and GP3of C. reinhardtii are shown in Table 2.
3.5. Polypeptides released from the insoluble wall fraction bychemical deglycosylation
When then freeze-dried aliquots of the insoluble fraction ofthe purified maternal cell wall of S. obliquus were subjected tochemical deglycosylation with HF/pyridine, at least some of itspolypeptide constituents became soluble in SDS-containing buffers(Fig. 5A, lane 2) as already reported for the insoluble glycopro-tein framework of the Chlamydomonas cell wall [27,29,34,40,43].Coomassie-stained SDS-PAGE gels revealed three prominent bandswith apparent molecular masses of 144, 135 and 65 kDa (Fig. 5A,lane 2), which were also found in the deglycosylation products ofthe chaotrope-soluble wall fraction (Fig. 5A, lane 1). The 89 kDacomponent, which was also prominent in the chemically deglyco-sylated, chaotrope-soluble wall fraction (Fig. 5A, lane 1), however,was completely missing in the deglycosylation products of theinsoluble wall fraction (Fig. 5A, lane 2). The latter findings clearlyshow that the polypeptides found in the deglycosylation productsof the insoluble wall fraction (Fig. 5A, lane 2) are not due to residualchaotrope-soluble polypeptides.
When corresponding protein blots were probed with the anti-dGP100 antibody (Fig. 5B), this particular antibody reacted withthe 144 and 135 kDa bands present in the deglycosylation prod-ucts of both the chaotrope-soluble and the insoluble wall fraction
(Fig. 5B, lanes 1 and 2). Additional cross-reacting component withapparent molecular masses between 106 and 120 kDa were alsoobserved in the deglycosylation products of both the chaotrope-soluble and the insoluble wall fraction (Fig. 5B, lanes 1 and 2),
44 J. Voigt et al. / Plant Science 215– 216 (2014) 39– 47
Table 2Amino acid sequences of the chaotrope-soluble cell-wall precursors presumably recognized by the anti-dICW antibody after hydroxylation of most of the proline residues.
Cell-wall precursora Positionb Amino acid sequencec
GP1 40–342 (SPXPP)nGP1 343–381 (SPSP)nGP2 286–338 RPPSPPLPPSPPPQPPSPLPPSPAPLPPSPPPSP LPPSPKPPTPPSPLPPAPPGP2 577–619 SPPPSPSPPPSPPQPPSPPPVPPSPPSPPPSPPS PANPSPPPPAPPGP2 775–818 CPPPPPAPSPPPKPPTPSPPPLPPQPNPPPAPPS PNPSPPPPPPGP3B 212–233 PPPPPSPLPPSPPPAPFPPPPFGP3B 413–438 QAQSPPPSPAPPLPPPSPPLPPPPVAGP3B 603–618 LPVPFPPPPPPPPPPG
a The chaotrope-soluble cell-wall glycoproteins were previously identified as precursors of the insoluble wall fraction of C. reinhardtii by mass spectroscopical sequencingo
or proe-rich
wSnlbpoadtCmIfwbAincowr
Fclsdmftew3p
f tryptic peptides and by immunochemical methods (Voigt et al. [34,43,50]).b Position of the proline-rich sequence in the amino acid sequence of the precursc Amino acid sequences derived from the ORF of the corresponding cDNA. Prolin
hich correspond to minor components of the correspondingDS-PAGE gels (Fig. 5, lanes 1 and 2). The prominent 89 kDa compo-ent of the deglycosylated, chaotrope-soluble wall fraction (Fig. 5A,
ane 1), however, did not cross-react with this particular anti-ody (Fig. 5B, lane 1). When the corresponding protein blots wererobed with anti-dICW antibody, different staining patterns werebtained for the deglycosylation products of the chaotrope-solublend the insoluble wall fractions (Fig. 5C, lanes 1 and 2). In theeglycosylation products of the chaotrope-soluble wall fraction,he 135 kDa component revealed the strongest signal (Fig. 5A and, lane 1). Additional cross-reacting material was found in theolecular mass range between 95 and 120 kDa (Fig. 5C, lane 1).
n the case of the deglycosylation products of the insoluble wallraction, however, the corresponding bands showed considerablyeaker reactivity towards this antibody, the most prominent band
eing observed at about 125, 65, and 45 kDa (Fig. 5C, lane 2).part from the 65 kDa polypeptide, which was also prominent
n the Coomassie-stained gels (Fig. 5A, lane 2), these compo-ents cross-reacting with the anti-dICW antibody were minoronstituents of this wall fraction. Due to the particular specificityf the anti-dICW antibody towards hydroxyproline-rich sequences,
e assume that these minor components contain hydroxyproline-ich domains.
ig. 5. Comparative SDS-PAGE and protein-blot analyses of the chemically degly-osylated polypeptides of the chaotrope-soluble and the insoluble wall fraction ofiving S. obliquus cells. Chemically deglycosylated polypeptides of the chaotrope-oluble wall fraction corresponding to 30 �g of protein (lane 1) and chemicallyeglycosylated polypeptides released from the insoluble wall fraction during treat-ent with hydrogen fluoride corresponding to 15 �g of protein (lane 2) were
ractionated by SDS-PAGE on gel slabs containing 7% (w/v) acrylamide. After elec-rophoresis, the gels were either stained with Coomassie Brilliant Blue G250 (A) orlectrophoretically blotted onto PVDF-membranes (B and C). The protein gel blotsere probed with either the anti-dGP100 (B) or the anti-dICW antibody (C). Lane
: protein molecular mass markers. The positions of molecular masses of markerroteins are given on the right.
lylpeptide. sequences of these types are known to be hydroxylated by proline hydroxylase.
3.6. The polypeptide constituents of the mother-cell walls
The LiCl-extracts of mother-cell walls contained two prominentpolypeptides with apparent molecular masses of 170 and 150 kDaand two weaker bands at higher molecular masses (Fig. 6A, lane1). All these polypeptides showed cross-reactivity with the anti-dICW antibody (Fig. 6B, lane 1), but not with the anti-dICW antibody(Fig. 6C, lane 1). After chemical deglycosylation, a single prominentband with an apparent molecular mass of 135 kDa was found inthe SDS-PAGE gels (Fig. 5A, lane 2), which cross-reacted with boththe anti-dGP100 antibody (Fig. 6B, lane 2) and the anti-dICW anti-body (Fig. 6C, lane 2). A predominant 135 kDa polypeptide was alsofound in the deglycosylation products of the insoluble wall fraction(Fig. 6A, lane 3), which also cross-reacted with the anti-dGP100antibody (Fig. 6B, lane 3), but only a rather weak signal with theanti-dICW antibody (Fig. 6C, lane 3), which revealed stronger bandswith components with apparent molecular masses between 95 and120 kDa not detected in the Coomassie-stained gels (Fig. 6A, lane3).
4. Discussion
Cell walls prepared from homogenates of living S. obliquus cells
and mother cells accumulated in the culture medium differ withrespect to their chemical composition including the proportionof glucosamine and the amino acid composition as previouslyreported [18]. In the present communication, we have shown thatFig. 6. SDS-PAGE and protein-blot analyses of the polypeptide constituentsof the maternal cell walls. Untreated (lane 1) and chemically deglycosylatedchaotrope-soluble cell-wall polypeptides (lane 2) from isolated maternal cell walls(corresponding to 15 �g of protein) and polypeptides released from the insolublefraction of the maternal cell walls during treatment with hydrogenfluoride (corre-sponding to 5 �g of protein; lane 3) were fractionated by SDS-PAGE on gel slabscontaining 7% (w/v) acrylamide. After electrophoresis, the gels were either stainedwith Coomassie Brilliant Blue G250 (A) or electrophoretically blotted onto PVDF-membranes (B and C). The protein gel blots were probed with either the anti-dGP100(B) or the anti-dICW antibody (C). Lane 4: protein molecular mass markers. Thepositions of molecular masses of marker proteins are given on the right.

J. Voigt et al. / Plant Science 215– 216 (2014) 39– 47 45
Table 3Presence of homologues to the Chlamydomonas chaotrope-soluble cell-wall glycoproteins in different green algae.
Species Order GP1 GP2 GP3 References
Chlamydomonas reinhardtii Volvocales + + + [46–49]Chlamydomonas eugametos Volvocales + + + [57,58]Chlamydomonas incerta Volvocales + + + [59]Volvox carteri Volvocales − + + [34,40,57,58]Gonium pectorale Volvocales − + + [60,61]Scendesmus obliquus Chlorococcales − − + This paperMicrasterias rotata Desmidiales ? ? (+)a [50]Gonatozygon brebissonii Zygnematales ? ? (+)a [50]Netrium digitus Zygnematales ? ? (+)a [50]Spirotaenia erythrocephala Zygnematales ? ? (+)a [50]
l-wallc
bcegeomwlptprt
bglclcrprppdwp
hp6mwpwAwmIpe2dslmtLl
a Putatively identified by a polyclonal antibody raised against the glycosylated celarbohydrate side chains.
oth cell types contain chaotrope-soluble glycoproteins, whichould be extracted by treatment with 3 M LiCl (Fig. 1). The LiCl-xtracts from both cell-wall types contain high-molecular-masslycoproteins with molecular masses ≥150 kDa (Fig. 1). In the LiCl-xtracts from cell wall prepared from homogenates of living S.bliquus cells, additional glycoproteins with apparent molecularasses of 40 and around 100 kDa were found (Fig. 1, lane 1), whichere missing in the LiCl-extracts from mother-cell walls (Fig. 1,
anes 2 and 3). Presumably, these 40 kDa and ≥100 kDa wall com-onents are degraded in the mother-cell walls during the release ofhe autospores. The lack of these 40 kDa and ≥100 kDa wall com-onents in the mother-cell walls provides an explanation for theeported differences in the amino acid composition of both cell-wallypes.
Chaotrope-soluble glycoproteins have been also reported toe cell-wall constituents of C. reinhardtii and several otherreen algae [46–50]. In the case of volvocine green algaeike Chlamydomonas and Volvox, the insoluble fraction of theell wall or extracellular matrix exclusively consists of cross-inked, hydroxyproline-containing glycoproteins [22–24,51]. Thehaotrope-soluble cell-wall glycoproteins GP1, GP2 and GP3 of C.einhardtii have been shown to be precursors of the insoluble glyco-rotein framework of the Chlamydomonas cell wall [34,40,43]. Theesults of our immunofluorescence studies (Fig. 2) revealed that theolyclonal antibodies raised against the 100 kDa deglycosylationroduct of the chaotrope-soluble cell-wall glycoprotein GP3B (anti-GP100) and against the deglycosylation products of the insolubleall fraction of C. reinhardtii (anti-dICW) both cross-reacted withurified cell walls of S. obliquus.
The deglycosylated LiCl-extracts from cell walls isolated fromomogenates of living S. obliquus cells contained four prominentolypeptides with apparent molecular masses of 144, 135, 89, and5 kDa and several minor components (Fig. 3A, lane 2). The 135 kDaajor constituent of the deglycosylated LiCl-extract cross-reactedith the anti-dGP100 antibody (Fig. 3B, lane 2) indicating that thisolypeptide is structurally related to the chaotrope-soluble cell-all glycoprotein GP3B of the volvocine green alga C. reinhardtii.dditionally, the anti-dGP100 antibody revealed cross-reactivityith a 106 kDa band (Fig. 3B, lane 2), which corresponded to ainor component in the Coomassie-stained gels (Fig. 3A, lane 2).
n the case of the untreated LiCl-extracts containing glycosylatedolypeptides, the anti-dGP100 antibody cross-reacted with sev-ral bands with apparent molecular masses of 135, 1765, 170, and40 kDa (Fig. 3B, lane 1).The relative intensities of these bandsiffered considerably from the relative intensities of the corre-ponding bands in the Coomassie-stained SDS-PAGE gels (Fig. 3A,ane 1). Similar results were obtained for the LiCl-extracts from the
aternal cell walls (Fig. 6A and B, lanes 1 and 2). We concludehat all the polypeptide constituents of the non-deglycosylatediCl-extracts cross-reacting with the anti-dGP100 antibody (Fig. 3B,ane 1, and Fig. 6B, lane 1) are differentially glycosylated
polypeptide GP3B of C. reinhardtii (anti-150 kDa), which strongly cross-reacts with
variants of the same 135 kDa polypeptide (Fig. 3A and B, lane2, and Fig. 6A and B, lane 2). This 135 kDa polypeptide must bestructurally related to the 100 kDa deglycosylation product of thechaotrope-soluble cell-wall glycoprotein GP3B of C. reinhardtii.Since the anti-dGP100 antibody was raised against the chemi-cally deglycosylated GP3B, it exclusively recognizes epitopes ofits polypeptide backbone (Fig. 4B and Table 1) and cross-reactswith non-glycosylated domains of the S. obliquus counterparts. Dueto considerable difference in size, we assume that the 100 kDapolypeptide backbone of the Chlamydomonas cell-wall polypeptideGP3B and its 135 kDa Scenedesmus counterpart are not very closelyrelated. However, a detailed comparative analysis of the amino acidsequences of both polypeptides is essential to estimate their evolu-tionary distance. The amino acid sequence of the Chlamydomonascell wall polypeptide GP3B is known [40], but not that of its S.obliquus homologue. The weak cross-reactivity of the deglycosy-lated 135 kDa polypeptide with the anti-dICW antibody (Fig. 3C,lane 2) indicates that it contains it least one hydroxyproline-richdomain. Three short (hydroxy)proline-rich domains have beenfound in the Chlamydomonas GP3B polypeptide (Table 1; [40]). Fur-thermore, both homologues also occur in the insoluble wall fraction(Fig. 5A–C, lane 2; [40]). Therefore, clarification of their significancefor the structure, generation and function of the insoluble wallfraction is of particular interest and must be studied in the future.
The insoluble wall fraction of C. reinhardtii and other vovocinegreen algae does not contain cellulose or other polysaccharides, butexclusively consists of a framework of hydroxyproline-containingglycoproteins [22–24]. Three types of cross-links between cell-wallglycoproteins have been described:
- peroxidase-catalyzed isodityrosine formation [52],- cross-linking via transglutaminase-dependent reactions [53], and- cross-linking via carbohydrate side-chains [54].
All these cross-links occur in the insoluble framework of theChlamydomonas cell wall [29,34,40]. Only those cross-links aredestroyed by HF-treatment, which occur between carbohydrateside-chains. Therefore, the polypeptide patterns of the deglyco-sylated chaotrope-soluble wall fraction and the deglycosylatedconstituent of the insoluble wall fraction of C. reinhardtii solubi-lized during HF-treatment were completely different [39]. In thecase of S. obliquus polypeptides of the same size were found inthe deglycosylation products of the LiCl-extracts and the insolu-ble wall fraction (Fig. 5; Fig. 6, lanes 2 and 3). indicating that thereis neither peroxidase-catalyzed nor transglutaminase-catalyzedcross-linking in the Scenedesmus cell wall, but only crosslinkingvia carbohydrate side-chains. Such kind of cross-links have been
recently also reported for the Arabidopsis cell wall [55].In the light of the evolution of the cell walls of the green algae,our finding that the cell wall of S. obliquus contains a homologueto the Chlamydomonas cell-wall glycoprotein GP3B but apparently

4 ce 21
ntToothfcpiitoredsfiohgrgiFotR
A
Cs
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
6 J. Voigt et al. / Plant Scien
o homologues to GP1 or GP2 is of particular interest with respecto the evolution of the diversity of the green alga cell walls [56].he chaotrope-soluble, cell-wall glycoproteins GP1, GP2, and GP3f C. reinhardtii differ considerably with respect to their proportionsf hydroxyproline, the size of their polypeptide backbones, andheir degree of glycosylation [34,40,43]. Homologues to the mostydroxyproline-rich cell-wall glycoprotein GP1 have been also
ound in other Chlamydomonas species, but are absent in the extra-ellular matrix of the colonial and multicellular Volvocaceae Goniumectorale and Volvox carteri (Table 3). GP2 and GP3 have been foundn all members of the Volvocaceae studied so far (Table 3). Specificnteractions between GP2 and GP3 have been reported [57]. Puta-ive homologues to GP3 have been also detected in the cell wallsf some members of the Zygnematales using a polyclonal antibodyaised against the glycosylated GP3B of C. reinhardtii [50]. How-ver, question arised whether or not this cross-reactivity is onlyue to similar glycosylation patterns or whether there are alsoimilarities in the polypeptide backbones. Taking together, thesendings indicate that comparative studies on both the occurencef GP3 homologues in various non-volvocalean green taxa and theomologies of their polypeptide backbones is a suitable tool toet a better insight into the evolution of these green algae. In thisespect, the GP3 homologues of S. obliquus seems to be a ratherood model because the size of its polypeptide backbone (135 kDa)s considerably larger than its C. reinhardtii counterpart (100 kDa).urthermore, cross-linking of the GP3 homologue of S. obliquusbviously takes place exclusively via HF-sensitive bonds in con-rast to the cell-wall glycoproteins of C. reinhardtii (Figs. 5 and 6;efs. [34,40,43]).)
cknowledgement
The authors wish to thank Dr. Ronald Frank (Department ofhemical Biology, Helmholtz Centre for Infection Research, Braun-chweig, Germany) for synthesis of the scan peptides.
eferences
[1] T.L. Chacon-Lee, G.E. Gonzales-Marino, Microalgae for “healthy” foods – possi-bilities and challenges, Compr. Rev. Food Sci. Food Safety 9 (2010) 655–675.
[2] R. Harun, M. Singh, G.M. Forde, M.K. Danquah, Bioprocess engineering ofmicroalgae to produce a variety of consumer products, Renew. Sust. EnergyRev. 14 (2010) 1037–1047.
[3] T.M. Mata, A.A. Martins, N.S. Caetano, Microalgae for biodiesel production andother applications: a review, Renew. Sust. Energy Rev. 14 (2010) 217–232.
[4] H.M. Amaro, A.C. Guedes, F.X. Malcata, Advances and perspectives in usingmicroalgae to produce biodiesel, Appl. Energy 88 (2011) 3402–3410.
[5] Y.L. Cheng, Y.C. Juang, G.Y. Liao, S.H. Ho, K.L. Yeh, C.Y. Chen, J.S. Chang, J.C. Liu,D.J. Lee, Dispersed ozone flotation of Chlorella vulgaris, Bioresour. Technol. 101(2010) 9092–9096.
[6] S.H. Ho, W.M. Chen, J.S. Chang, Scendesmus obliquus CNW-N as a potential can-didate for CO2 mitigation and biodiesel production, Bioresour. Technol. 101(2010) 8725–8730.
[7] S.H. Ho, W.B. Lu, J.S. Chang, Photobioreactor strategies for improving the CO2
fixation efficiency of indigenous Scenedesmus obliquus CNW-N: statistical opti-mization of CO2 feeding, illumination, and operation mode, Bioresour. Technol.105 (2012) 106–113.
[8] A. Kumar, S. Ergas, X. Yuan, A. Sahu, Q. Zhang, J. Dewulf, F.X. Malcata, H. van Lan-genhove, Enhance CO2 fixation and biofuel production via microalgae: recentdevelopments and future directions, Trends Biotechnol. 28 (2010) 371–380.
[9] S. Mandal, N. Mallick, Waste utilization and biodiesel production by the greenmicroalgae Scenedesmus obliquus, Appl. Environ. Microbiol. 77 (2011) 374–377.
10] J. Burczyk, The chemical composition of the cell wall of Scenedesmus obliquus.I. General chemical characterisation, Folia Histochem. Cytochem. 11 (1973)119–134.
11] B. Allard, J. Templier, High molecular weight lipids from the trilaminar outerwall (TLS)-containing microalgae Chlorella emersonii, Scenedesmus communisand Tetraedron minimum, Phytochemistry 57 (2001) 459–467.
12] A.W. Atkinson, B.E.S. Gunning, P.C.L. John, Sporopollenin in the cell wallof Chlorella and other algae: ultrastructure, chemistry and incorporation of[14C]acetate, studied in synchronous cultures, Planta 107 (1972) 1–32.
13] J.W. de Leeuw, C. Largeau, A review of macromolecular organic compoundsthat comprise living organisms and their role in keroge, coal and petroleum
[
[
5– 216 (2014) 39– 47
formation, in: M. M. Engel (Ed.), Organic Geochemistry. Principles and Appli-cations, Plenum Press, New York, 1993, pp. 23–72.
14] U. Brunner, R. Honegger, Chemical and ultrastructural studies on the distribu-tion of sporopollenin-like biopolymers in six genera of lichen phycobionts, Can.J. Bot. 63 (1985) 2221–2230.
15] J. Burczyk, Cell wall carotinoids in green algae which form sporopollenins,Phytochemistry 26 (1987) 121–128.
16] S. Derenne, C. Largeau, C. Berkaloff, B. Rousseau, C. Wilhem, P.G. Hatcher, Non-hydrolysable macromolecular constituents from outer walls of Chlorella fuscaand Nanochlorum eucaryotum, Phytochemistry 31 (1992) 1923–1929.
17] P. Blokker, S. Schouten, H. van den Ende, J.W. de Leeuw, P.G. Hatcher, J.S. Sin-ninghe Damsté, Chemical structure of algaenan from the fresh water Tetraedronminimum, Scenedesmus communis and Pediastrum boryanum, Organ. Geochem.29 (1998) 1453–1468.
18] J. Burczyk, B. Smietana, K. Terminska-Pabis, M. Zych, P. Kowalowski, Com-parison of nitrogen content amino acid composition and glucosaminecontent of cell walls of various chlorococcal algae, Phytochemistry 51 (1999)491–497.
19] G.I. Cassab, Plant cell wall proteins, Annu. Rev. Biochem. Plant Physiol. PlantMol. Biol. 49 (1998) 281–309.
20] L.S. Schnabelrauch, M.J. Kieliszewski, B.L. Upham, H. Alizedeh, D.T.A. Lamport,Isolation of pI 4.6 extensin peroxidase from tomato cell suspension cultures andidentification of Val-Tyr-Lys as putative intermolecular cross-link site, Plant J.9 (1996) 477–489.
21] J. Voigt, D. Wrann, H.P. Vogeler, W.A. König, M. Mix, Hydroxyproline-containingand glycine-rich cell-wall polypeptides are widespread in the green algae,Microbiol. Res. 149 (1994) 223–229.
22] K. Roberts, Crystalline glycoprotein cell walls of algae – their structure, compo-sition and assembly, Phil. Trans. R. Soc. Lond. B: Biol. Sci. 268 (1974) 129–146.
23] K. Roberts, C. Grief, G.J. Hills, P.J. Shaw, Cell wall glycoproteins: structure andfunction, J. Cell Sci. (Suppl. 2) (1985) 105–127.
24] S.H. Imam, M.J. Buchanan, H.C. Shin, W.J. Snell, The Chlamydomonas cell wall:characterization of the wall framework, J. Cell Biol. 101 (1985) 1599–1607.
25] E. Kessler, F.C. Czygan, Physiologische und biochemische Beiträge zurTaxonomie der Gattung Chlorella. IV. Verwertung organischer Stickstof-fverbindungen, Arch. Mikrobiol. 70 (1970) 211–216.
26] J. Burczyk, H. Szkawran, I. Zontek, F.C. Czygan, Carotinoids in the outer cell-walllayer of Scenedesmus (Chlorophyceae), Planta 151 (1981) 247–250.
27] H.P. Vogeler, J. Voigt, W.A. König, Polypeptide pattern of the insoluble wall com-ponent of Chlamydomonas reinhardtii and its variation during the vegetative cellcycle, Plant Sci. 71 (1990) 119–128.
28] J. Voigt, Extraction by lithium-chloride of hydroxyproline-rich glycoproteinsfrom intact cells of Chlamydomonas reinhardtii, Planta 164 (1985) 379–389.
29] J. Voigt, R. Frank, 14-3-3 proteins are constituents of the insoluble glyco-protein framework of the Chlamydomonas cell wall, Plant Cell 15 (2003)1399–1413.
30] P.F. Smith, R.I. Krohn, G.T. Hermanson, A.R. Mallis, F.H. Bartner, M.B. Proven-zano, E.K. Fujimoto, N.M. Goeke, B.J. Olson, D.C. Lenk, Measurement of proteinusing bicinchoninic acid, Anal. Biochem. 150 (1985) 76–85.
31] R.E. Brown, K.L. Jarvis, K.J. Hyland, Protein measurement using bicinchoninicacid: elimination of interfering substances, Anal. Biochem. 180 (1989) 136–139.
32] U.K. Laemmli, Cleavage of structural proteins during the assembly of the headof bacteriophage T4, Nature 227 (1970) 680–685.
33] H. Towbin, T. Staehelin, J. Gordon, Electrophoretic transfer of proteins frompolyacrylamide gels to nitrocellulose sheets: procedure and some applications,Proc. Natl. Acad. Sci. U.S.A. 76 (1979) 4350–4354.
34] J. Voigt, J. Wöstemeyer, R. Frank, The chaotrope-soluble glycoprotein GP2 is aprecursor of the insoluble glycoprotein framework of the Chlamydomonas cellwall, J. Biol. Chem. 282 (2007) 30381–30392.
35] R. Hawkes, E. Nidag, J. Gordon, A dot-immunobinding assay for monoclonal andother antibodies, Anal. Biochem. 119 (1982) 142–147.
36] R. Frank, Spot synthesis: an easy technique for the positionally addressable,parallel chemical synthesis on membrane support, Tetrahedron 48 (1992)9217–9232.
37] P.W. Pappas, The use of a chrome-alum gelatin (subbing) as a general adhesivefor paraffin sections, Stain Technol. 46 (1971) 121–124.
38] J. Heslop-Harrison, Y. Heslop-Harrison, Myosin associated with the surfaces oforganelles, vegetative nuclei and generative cells in angiosperm pollen grainsand tubes, J. Cell Sci. 94 (1989) 319–325.
39] J. Voigt, B. Hinkelmann, I. Liebich, M. Mix, Alteration of the cell surface duringthe vegetative cell cycle of the unicellular green alga Chlamydomonas rein-hardtii, Plant Cell Physiol. 37 (1996) 726–733.
40] J. Voigt, M. Kiess, R. Getzlaff, J. Wöstemeyer, R. Frank, Generation of the het-erodimeric precursor GP3 of the Chlamydomonas cell wall, Mol. Microbiol. 77(2010) 1512–1526.
41] K. Keskiaho, R. Hieta, R. Sormunen, J. Myllyharju, Chlamydomonas reinhardtiihas multiple prolyl 4-hydroxylases, one of which is essential for proper cellwall assembly, Plant Cell 19 (2007) 256–269.
42] J.H. Lee, S. Waffenschmidt, L. Small, U.W. Goodenough, Between-species anal-ysis of short-repeat modules in cell wall and sex-related hydroxyproline-richglycoproteins of Chlamydomonas, Plant Physiol. 144 (2007) 1813–1826.
43] J. Voigt, J. R: Frank, Wöstemeyer, The chaotrope-soluble glycoprotein GP1 isa constituent of the insoluble glycoprotein framework of the Chlamydomonascell wall, FEMS Microbiol. Lett. 291 (2009) 209–215.
44] M.J. Kieliszewski, D.T.A. Lamport, Extensin: repetitive motifs, functional sites,post-translational codes, and phylogeny, Plant J. 5 (1994) 157–172.

ce 21
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
J. Voigt et al. / Plant Scien
45] A.M. Showalter, B. Keppler, J. Lichtenberg, D. Gu, L.R. Welch, A bioinformaticsapproach to identification, classification, and analysis of hydroxyproline-richglycoproteins, Plant Physiol. 153 (2010) 485–513.
46] G.J. Hills, J.M. Phillips, M.R. Gray, K. Roberts, Self-assembly of a plant cell wallin vitro, J. Mol. Biol. 96 (1975) 431–434.
47] U.W. Goodenough, J.E. Heuser, The Chlamydomonas cell wall and its constituentglycoproteins analyzed by the quick-freeze, deep-etch technique, J. Cell Biol.101 (1985) 1550–1568.
48] U.W. Goodenough, J.E. Heuser, Molecular organization of cell-wall crystals fromChlamydomonas reinhardtii and Volvox carteri, J. Cell Sci. 90 (1988) 717–733.
49] U.W. Goodenough, B. Gebhart, R.P. Mecham, J.E. Heuser, Crystals of theChlamydomonas reinhardtii cell wall: polymerization, depolymerization andpurification of glycoprotein monomers, J. Cell Biol. 103 (1986) 924–942.
50] J. Voigt, H.P. Vogeler, W.A. König, D. Wrann, M. Mix, Chaotrope-soluble cellwall glycoproteins of Volvox and some members of the Zygnematophyceaeimmunologically related to the 150 kDa cell-wall glycoprotein of Chlamy-domonas reinhardtii, Microbiol. Res. 150 (1995) 129–137.
51] H. Ertl, R. Mengele, S. Wenzl, J. Engel, M. Sumper, The extracellular matrix ofVolvox carteri: molecular structure of the cellular compartment, J. Cell Biol. 109(1989) 3493–3501.
52] S. Waffenschmidt, J.P. Woessner, K. Beer, U.W. Goodenough, Isodityrosine
cross-linking mediates insolubilization of cell walls in Chlamydomonas, PlantCell 5 (1993) 809–820.53] S. Waffenschmidt, T. Kusch, J.P. Woessner, A transglutaminase immunologicallyrelated to tissue transglutaminase catalyzes cross-linking of cell wall proteinsin Chlamydomonas reinhardtii, Plant Physiol. 121 (1999) 1003–1015.
[
[
5– 216 (2014) 39– 47 47
54] O. Holst, V. Christoffel, R. Fründ, H. Moll, M. Sumper, A phosphodiester bridgebetween two arabinose residues as a structural element of an extracellularglycoprotein of Volvox carteri, Eur. J. Biochem. 181 (1989) 345–350.
55] L. Tan, S. Eberhard, S. Pattathil, C. Warder, J. Glushka, C. Yuan, X. Zhu, U. Avci, J.S.Miller, D. Baldwin, C. Pjam, R. Orlando, A. Darvill, M.G. Hahn, M.J. Kieliszewsji,D. Mohnen, An Arabidopsis cell wall proteoglycan consists of pectin and arabi-noxylan covalently linked to an arabinogalactan protein, Plant Cell 25 (2013)270–287.
56] D.S. Domozych, M. Ciancia, J.U. Fangel, M. Dalgaard Mikkelsen, P. Ulvskov,W.G.T. Willats, The cell walls of green algae: a journey through evolution anddiversity, Front. Plant Sci. 3 (2012) 1–7.
57] W.S. Adair, S.A. Steinmetz, D.M. Mattson, U.W. Goodenough, J.E. Heuser, Nucle-ated assembly of Chlamydomonas and Volvox cell walls, J. Cell Biol. 105 (1987)2373–2382.
58] J.P. Woessner, A.J. Molendijk, P. van Egmond, F.M. Klis, U.W. Goodenough, M.A.Haring, Domain conservation in several volvocalean cell wall proteins, PlantMol. Biol. 26 (1994) 947–960.
59] J.H. Lee, S. Waffenschmidt, L. Small, U.W. Goodenough, Between-speciesanalysis of short-repeat modules in cell wall and sex-related hydroxyproline-rich glycoproteins of Chlamydomonas, Plant Physiol. 144 (August) (2007)1813–1826.
60] W.S. Adair, H. Appel, Identification of a highly conserved hydroxyproline-richglycoprotein in the cell walls Chlamydomonas reinhardtii and two other Volvo-cales, Planta 179 (1989) 381–386.
61] J.P. Woessner, U.W. Goodenough, Volvocine cell walls and their constituentglycoproteins: an evolutionary perspective, Protoplasma 181 (1994) 245–258.





























