Embed Size (px)
Citation preview

lable at ScienceDirect
Cretaceous Research 32 (2011) 378e386
Contents lists avai
Cretaceous Research
journal homepage: www.elsevier .com/locate/CretRes
The chronology of dinosaur oospecies in south-western Europe: Refinementsfrom the Maastrichtian succession of the eastern Pyrenees
Bernat Vila a,*, Violeta Riera b, Ana María Bravo c, Oriol Oms b, Enric Vicens b, Rita Estrada b,Àngel Galobart a
a Institut Català de Paleontologia, Carrer Escola Industrial, 23, 08201 Sabadell, Barcelona, Catalonia, SpainbDepartament de Geologia, Universitat Autònoma de Barcelona, 08193 Bellaterra (Cerdanyola del Vallès) Barcelona, Catalonia, SpaincMuseo Geominero, Instituto Geológico y Minero de España, Calle Río Rosas, 23, 28003, Madrid, Spain
a r t i c l e i n f o
Article history:Received 19 May 2010Accepted in revised form 21 January 2011Available online 3 February 2011
Keywords:EggshellDinosaurMegaloolithidCretaceousPyreneesChronology
* Corresponding author. Tel.: þ34 937261769.E-mail addresses: [email protected] (B. Vil
(V. Riera), [email protected] (A.M. Bravo), [email protected] (E. Vicens), [email protected] (À. Galobart).
0195-6671/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.cretres.2011.01.009
a b s t r a c t
The Late Cretaceous deposits of the continental Tremp Formation in the Vallcebre Syncline (South-eastern Pyrenees) provide an extensive egg record of dinosaurs. The parataxonomical study andanalysis of multiple eggshell samples, the precise stratigraphical control of several sections and the timecalibration of the abundant egg levels enable the establishment of a robust oospecies succession. Thesuccessive occurrence of three megaloolithid oospecies (Megaloolithus sirugueieMegaloolithus mamil-lareeMegaloolithus sp.) in the Early and Late Maastrichtian is well correlated with the magnetic polaritytime scale throughout the chron 31. The replacement of Megaloolithus siruguei with Megaloolithusmamillare occurs around the reversal of chrons 31re31n. A comparison with oospecies successions fromArc basin localities (France) allows the age calibration for such oospecies replacement to be confirmed.This age refinement implies that some of the stated boundaries for the proposed oospecies assemblagesmay change and that an in-depth revision of the age and magnetic calibration of some south Europeanegg localities is required.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Dinosaur eggs and eggshells are some of the most abundantfossil evidence in continental deposits of Late Cretaceous of Laurasiaand Gondwana landmasses (Carpenter, 1999). They enable theassessment of the reproductive biology, diversity and palae-oenvironmental preferences of egg-laying dinosaurs by means ofthe study of clutch/nest taphonomy (e.g. Varricchio et al., 1997;Chiappe et al., 2004), microstructural features of eggshells (e.g.Deeming, 2006; Jackson et al., 2008), parataxonomy (e.g. Mikhaïlov,1991, 1997) or geochemical analysis (e.g. Sarkar et al., 1991; Cojanet al., 2003). Diversity studies using oospecies (Vianey-Liaud andGarcia, 2000; Vianey-Liaud et al., 2003; López-Martínez, 2003)may be significant in order to assess the true dinosaur diversity.Furthermore, some authors have stated that dinosaur eggshells canbe used as biochronological markers in continental deposits (Garciaand Vianey-Liaud, 2001a). For the latter objective it is crucial to
a), [email protected]@uab.cat (O. Oms),(R. Estrada), angel.galobart@
All rights reserved.
establish a solid chronology of ootaxa. However, the chronology ofthe continental palaeontological record of vertebrates has tradi-tionally been hampered for two reasons. The first is the correlationof the continental record with the standard stratigraphy, which isdifficult since the latter is based on marine biozonations. Second,the definition of a terrestrial biostratigraphy is more complex thana marine one due to various factors related to depositional settingsand the features of the organisms. Therefore, a succession of sites ina single section rarely occurs. This hampers the definition ofa vertical (time) succession of sites and palaeontological events.Difficulties in assigning a continental succession of palaeontologicalevents and their dating can be overcome if wework in long sectionswith a large number of sites wheremagnetostratigraphic dating canbe carried out. Such sections will generally be composite (goodlithostratigraphical control is required) and will generally needsome rough age estimation based on marine biostratigraphy.
All these favourable conditions occur in the Campanian andMaastrichtian terrestrial deposits with dinosaur eggshells of south-western Europe (Fig. 1). In France, oospecies of the MegaloolithidaeZhao, 1979 oofamily have long been identified and tentatively usedfor biochronological purposes in the Ariège, Hérault, Aude andBouches-du-Rhône departments (Vianey-Liaud et al., 1994, 1997;Garcia and Vianey-Liaud, 2001a, and references therein). In

Fig. 1. Geographic setting of the areas with Campanian-Maastrichtian terrestrial deposits at South-western Europe (South France and North Catalonia). With data from Garcia andVianey-Liaud, 2001a.
B. Vila et al. / Cretaceous Research 32 (2011) 378e386 379
Catalonia, some authors described several megaloolithid oospeciesin the south-central and south-eastern Pyrenean basins (Vianey-Liaud and López-Martínez, 1997; López-Martínez, 2000, 2003;Bravo et al., 2005). In these areas, there is goodmagnetostratigraphiccontrol, as shown by Westphal and Durand (1990), Galbrun et al.(1993), Cojan and Moreau (2006) and Oms et al. (2007). Such well-dated sections contain a significant succession of dinosaur oospecies.As a result, a biozonation has been proposed for the French andCatalan localities (Garcia and Vianey-Liaud, 2001a; López-Martínez,2003). However, they are not exempt from controversy since thedating dataset is not homogeneous throughout all the studiedsections. In the present paper, we present a magnetostratigraphic
Fig. 2. General geological map of the Vallcebre Syncline
calibration for the oospecies succession of the Vallcebre Syncline(South-easternPyrenees, Catalonia) and refine anddiscuss particularaspects of the previous French and Catalan biozonations.
2. Geological setting
The studied dinosaur eggshell localities are found in the Vall-cebre Syncline (Fig. 2). About 2,000 m of sediment crops out in thisarea, encompassing approximately 1,000 m of Upper Cretaceouscoastal calcarenites with rudist reefs (Terradets LimestonesFormation) and 900 m of Upper CretaceousePaleogene transitional
. Black lines indicate location of measured sections.
B. Vila et al. / Cretaceous Research 32 (2011) 378e386380
and continental strata (see Oms et al., 2007 and Riera et al., 2010and references therein).
The Tremp Formation consists of a succession of marine-to-continental transitional and continental strata that comprise theCretaceousePaleogene boundary. They were deposited followinga marine regression that began near the CampanianeMaastrichtianboundary. Four regional lithologic units can be recognized,which arefrom base to top (Rosell et al., 2001): (1) a marine-continental tran-sitional grey unit (grey marls with abundant invertebrates, lignites,limestone, and sandstone layers); (2) a lower fluvial red unit (redmudstones, sandstones, and paleosols); (3) the lacustrine VallcebreLimestones and laterally equivalent strata (with charophytes andMicrocodium); and (4) an upper fluvial red unit (red mudstones,sandstones, and conglomerates). The first two units are part of theMaastrichtian and contain a rich assemblage of fossil vertebratesincluding dinosaurs (titanosaurian sauropods, dromaeosauridtheropods, and hadrosaurian ornithopods), crocodiles, cheloniansand batoids (see references in Vila et al., 2005, 2008; Marmi et al.,2009, 2010); the latter two belong to the Danian. The Cretaceous-Paleogene boundary is placed at the base of the Vallcebre limestones(Oms et al., 2007). A good correlationwith equivalent strata from theArc Basin in France is observed (see Westphal and Durand, 1990).
3. Material and methods
Palaeontological and stratigraphical studies were conducted inseveral continental outcrops of the Tremp Formation along theVallcebre Syncline. Oological research consists of sampling, analysis,and parataxonomical identification and review of eggshell frag-ments. Stratigraphical work included the precise location of 29 egg-bearing levels in 8 sections. The correlation and calibration of thestratigraphical levels to the standard polarity scale was performedusing knownmagnetostratigraphic and biochronological constraints(Oms et al., 2007). The eggshell fragmentswere collected from in situ
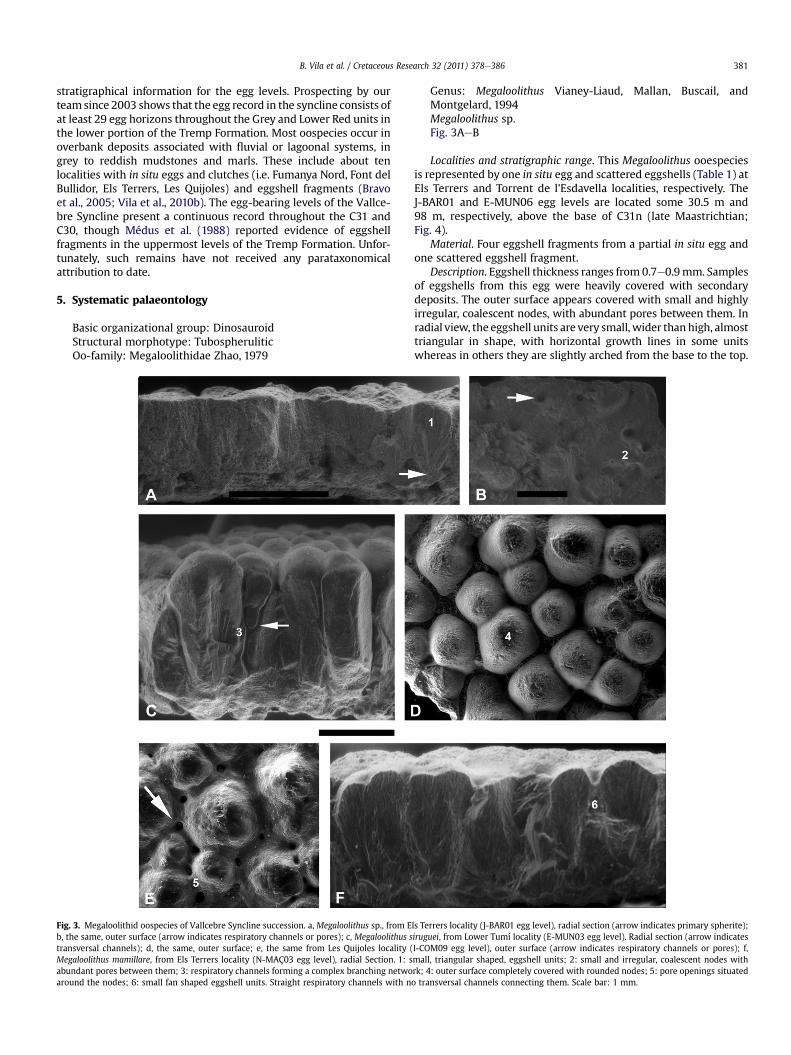
Table 1Listed samples and localities with Megaloolithus ooespecies in the Vallcebre Syncline. Eg
Locality Egg-bearing level (site) Source material Nu
Els Terrers J-BAR01 Eggs 4J-BAR03 Eggshells 1N-MAÇ03 Eggshells 5N-MAÇ17 Eggshells 1N-MAÇ18 Eggshells 3N-MAÇ16 Eggs 4N-MAÇ15 Eggshells 2N-MAÇ14 Eggshells 7N-MAÇ13 Eggshell 1N-MAÇ02 Egg debris 10N-MAÇ12 Eggs 1N-MAÇ07 Eggs 7N-MAÇ08 Eggs 13N-MAÇ11 Eggs 4N-MAÇ10 Eggshells 2N-MAÇ06 Eggs 2N-MAÇ09 Eggs 1N-MAÇ04 Eggs 4
Les Quijoles I-COM04 Eggs 7I-COM09 Eggs 2
Lower Tumí E-MUN02 Eggshells 4E-MUN01 Eggs 6E-MUN03 Eggs 4
Fumanya Nord E-MUN05 Eggs 5Fumanya Sud B-FUM10 Eggs 8Font del Bullidor A-FIG05 Eggs 42Torrent de l’Esdavella E-MUN06 Eggshells 1
1Peguera P2-COL01 Eggshells 4Coll de Jou L-ESP10 Eggshell 1
eggs at 16 levels and scattered in the sediment at 13 levels (Table 1).The material used for parataxonomical identification is part of thesamples listed in Bravo et al. (2005). Additional eggshell sampleswere collected at 18 new egg horizons and sections. They werecleaned with an ultrasonic bath or with potassium hydroxide, anddried at room temperature. All eggshells were first observed underbinocular microscope Nikkon SMZ-10 and stereomicroscope Leica�
MZ16A, in order to describe inner and outer eggshell surfaces, radialview and general preservation. Thickness was measured witha calliper; the mean of three repeated measurements was taken asthefinalvalue (Table 1). Some fresh broken fragmentsweremountedonaluminiumstubs, goldcoatedandphotographedundera scanningelectron microscope. Some other pieces were used to prepare thinslides, following standard petrographical techniques. Egg taxa wereidentified in terms of parataxonomy following Mikhaïlov (1997).
The studied material is housed in the collection of the InstitutCatalà de Paleontologia (IPS 23009, 27361e27375, 27378e27381,27386-27388, 27390, 27392, 27394, 27853, 46509, 46511e46527,53052, and 53053).
4. Results
4.1. Oospecies occurrence
Despite the fact thatmost of the oological reports traditionally arefrom the neighbouring Tremp and Coll de Nargó synclines (Sanzet al., 1995; Vila et al., 2010a, among others), the Vallcebre Synclinecontains an important record of dinosaur eggs and clutchesthroughout the Tremp Formation outcrops. Unpublishedworks fromthe sixties reported dinosaur fossils in the area (Aepler, 1967) andfirst publications by Ullastre and Masriera (1983), Médus et al.(1988), and Peitz (2000) provided evidence for eggshell fragmentsin a few stratigraphical levels of the area (L’Espà and Vallcebreoutcrops). None of these early works, however, provided precise
g levels are stratigraphically ordered in each locality.
mber of samples Thickness range (mm) Oospecies
0.7e0.9 Megaloolithus sp.1.46 Megaloolithus mamillare1.64e1.90 Megaloolithus mamillare2.09 Megaloolithus siruguei1.89e2.13 Megaloolithus siruguei2.21e2.41 Megaloolithus cf. siruguei2.01e2.92 Megaloolithus siruguei2.05e3.16 Megaloolithus siruguei3.30 Megaloolithus siruguei2.65e3.06 Megaloolithus siruguei2.81 Megaloolithus siruguei2.76e3.19 Megaloolithus siruguei2.18e2.99 Megaloolithus siruguei1.89e1.97 Megaloolithus siruguei2.23e2.27 Megaloolithus siruguei2.74 Megaloolithus siruguei2.80 Megaloolithus siruguei2.30e2.50 Megaloolithus siruguei1.93e2.76 Megaloolithus siruguei2.09e2.74 Megaloolithus siruguei1.79e1.95 Megaloolithus siruguei2.12e2.83 Megaloolithus siruguei2.69e2.95 Megaloolithus siruguei2.52e3.11 Megaloolithus siruguei2.80e3.30 Megaloolithus siruguei1.64e2.48 Megaloolithus siruguei0.73 Megaloolithus sp.1.04e1.14 Megaloolithus cf. mamillare1.04e1.13 Megaloolithus mamillare2.75 Megaloolithus siruguei
B. Vila et al. / Cretaceous Research 32 (2011) 378e386 381
stratigraphical information for the egg levels. Prospecting by ourteam since 2003 shows that the egg record in the syncline consists ofat least 29 egg horizons throughout the Grey and Lower Red units inthe lower portion of the Tremp Formation. Most oospecies occur inoverbank deposits associated with fluvial or lagoonal systems, ingrey to reddish mudstones and marls. These include about tenlocalities with in situ eggs and clutches (i.e. Fumanya Nord, Font delBullidor, Els Terrers, Les Quijoles) and eggshell fragments (Bravoet al., 2005; Vila et al., 2010b). The egg-bearing levels of the Vallce-bre Syncline present a continuous record throughout the C31 andC30, though Médus et al. (1988) reported evidence of eggshellfragments in the uppermost levels of the Tremp Formation. Unfor-tunately, such remains have not received any parataxonomicalattribution to date.
5. Systematic palaeontology
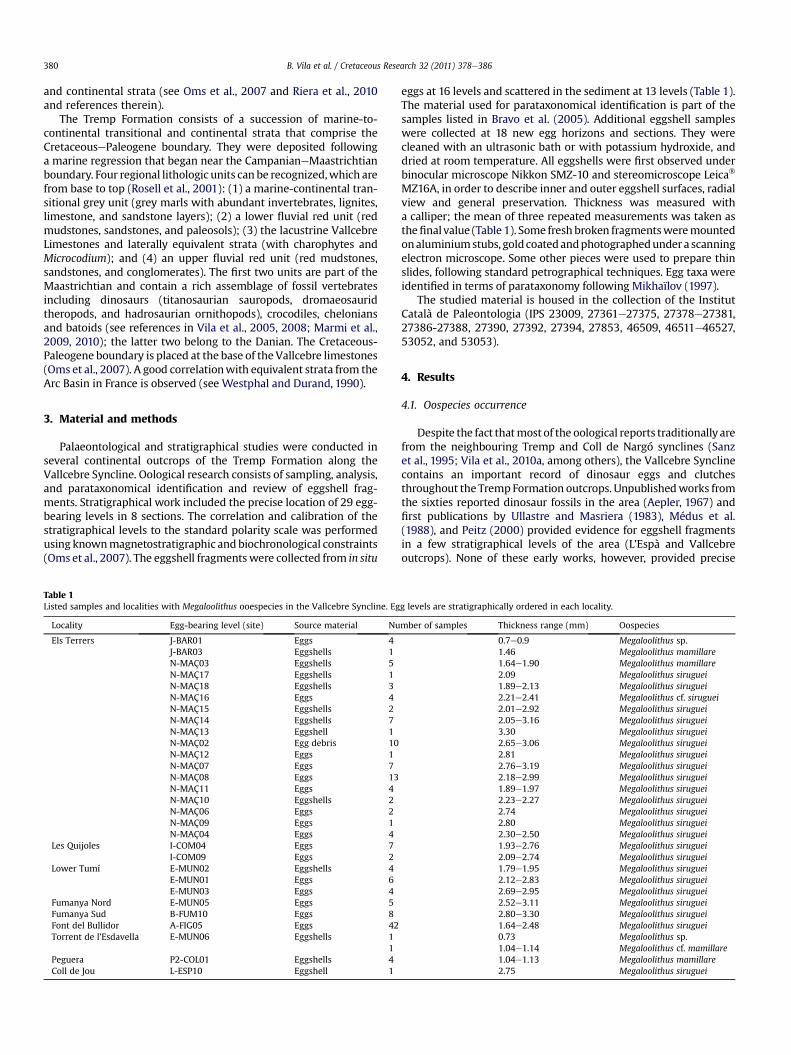
Basic organizational group: DinosauroidStructural morphotype: TubospheruliticOo-family: Megaloolithidae Zhao, 1979
Fig. 3. Megaloolithid oospecies of Vallcebre Syncline succession. a, Megaloolithus sp., from Eb, the same, outer surface (arrow indicates respiratory channels or pores); c, Megaloolithus stransversal channels); d, the same, outer surface; e, the same from Les Quijoles locality (Megaloolithus mamillare, from Els Terrers locality (N-MAÇ03 egg level), radial Section. 1: sabundant pores between them; 3: respiratory channels forming a complex branching netwoaround the nodes; 6: small fan shaped eggshell units. Straight respiratory channels with n
Genus: Megaloolithus Vianey-Liaud, Mallan, Buscail, andMontgelard, 1994Megaloolithus sp.Fig. 3AeB
Localities and stratigraphic range. This Megaloolithus ooespeciesis represented by one in situ egg and scattered eggshells (Table 1) atEls Terrers and Torrent de l’Esdavella localities, respectively. TheJ-BAR01 and E-MUN06 egg levels are located some 30.5 m and98 m, respectively, above the base of C31n (late Maastrichtian;Fig. 4).
Material. Four eggshell fragments from a partial in situ egg andone scattered eggshell fragment.
Description. Eggshell thickness ranges from 0.7e0.9mm. Samplesof eggshells from this egg were heavily covered with secondarydeposits. The outer surface appears covered with small and highlyirregular, coalescent nodes, with abundant pores between them. Inradial view, the eggshell units are very small, wider thanhigh, almosttriangular in shape, with horizontal growth lines in some unitswhereas in others they are slightly arched from the base to the top.
ls Terrers locality (J-BAR01 egg level), radial section (arrow indicates primary spherite);iruguei, from Lower Tumí locality (E-MUN03 egg level). Radial section (arrow indicatesI-COM09 egg level), outer surface (arrow indicates respiratory channels or pores); f,mall, triangular shaped, eggshell units; 2: small and irregular, coalescent nodes withrk; 4: outer surface completely covered with rounded nodes; 5: pore openings situatedo transversal channels connecting them. Scale bar: 1 mm.
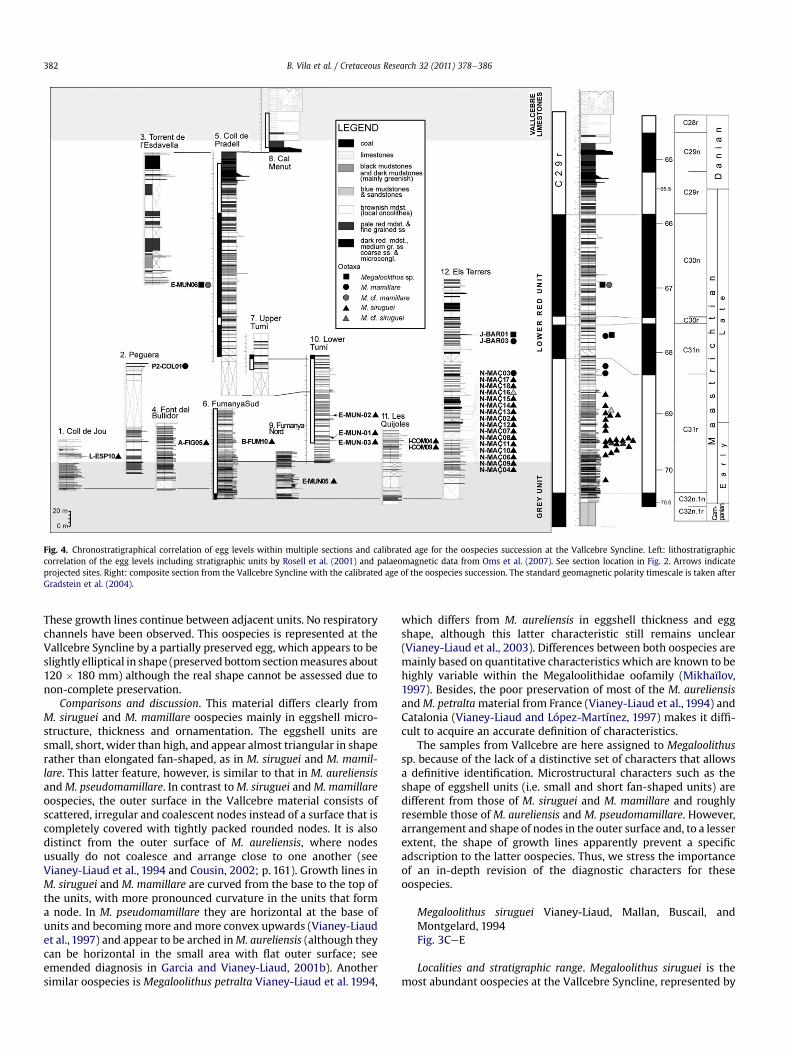
Fig. 4. Chronostratigraphical correlation of egg levels within multiple sections and calibrated age for the oospecies succession at the Vallcebre Syncline. Left: lithostratigraphiccorrelation of the egg levels including stratigraphic units by Rosell et al. (2001) and palaeomagnetic data from Oms et al. (2007). See section location in Fig. 2. Arrows indicateprojected sites. Right: composite section from the Vallcebre Syncline with the calibrated age of the oospecies succession. The standard geomagnetic polarity timescale is taken afterGradstein et al. (2004).
B. Vila et al. / Cretaceous Research 32 (2011) 378e386382
These growth lines continue between adjacent units. No respiratorychannels have been observed. This oospecies is represented at theVallcebre Syncline by a partially preserved egg, which appears to beslightly elliptical in shape (preserved bottomsectionmeasures about120 � 180 mm) although the real shape cannot be assessed due tonon-complete preservation.
Comparisons and discussion. This material differs clearly fromM. siruguei and M. mamillare oospecies mainly in eggshell micro-structure, thickness and ornamentation. The eggshell units aresmall, short, wider than high, and appear almost triangular in shaperather than elongated fan-shaped, as in M. siruguei and M. mamil-lare. This latter feature, however, is similar to that in M. aureliensisandM. pseudomamillare. In contrast toM. siruguei andM. mamillareoospecies, the outer surface in the Vallcebre material consists ofscattered, irregular and coalescent nodes instead of a surface that iscompletely covered with tightly packed rounded nodes. It is alsodistinct from the outer surface of M. aureliensis, where nodesusually do not coalesce and arrange close to one another (seeVianey-Liaud et al., 1994 and Cousin, 2002; p. 161). Growth lines inM. siruguei and M. mamillare are curved from the base to the top ofthe units, with more pronounced curvature in the units that forma node. In M. pseudomamillare they are horizontal at the base ofunits and becoming more and more convex upwards (Vianey-Liaudet al., 1997) and appear to be arched inM. aureliensis (although theycan be horizontal in the small area with flat outer surface; seeemended diagnosis in Garcia and Vianey-Liaud, 2001b). Anothersimilar oospecies is Megaloolithus petralta Vianey-Liaud et al. 1994,
which differs from M. aureliensis in eggshell thickness and eggshape, although this latter characteristic still remains unclear(Vianey-Liaud et al., 2003). Differences between both oospecies aremainly based on quantitative characteristics which are known to behighly variable within the Megaloolithidae oofamily (Mikhaïlov,1997). Besides, the poor preservation of most of the M. aureliensisandM. petraltamaterial from France (Vianey-Liaud et al., 1994) andCatalonia (Vianey-Liaud and López-Martínez, 1997) makes it diffi-cult to acquire an accurate definition of characteristics.
The samples from Vallcebre are here assigned to Megaloolithussp. because of the lack of a distinctive set of characters that allowsa definitive identification. Microstructural characters such as theshape of eggshell units (i.e. small and short fan-shaped units) aredifferent from those of M. siruguei and M. mamillare and roughlyresemble those of M. aureliensis and M. pseudomamillare. However,arrangement and shape of nodes in the outer surface and, to a lesserextent, the shape of growth lines apparently prevent a specificadscription to the latter oospecies. Thus, we stress the importanceof an in-depth revision of the diagnostic characters for theseoospecies.
Megaloolithus siruguei Vianey-Liaud, Mallan, Buscail, andMontgelard, 1994Fig. 3CeE
Localities and stratigraphic range. Megaloolithus siruguei is themost abundant oospecies at the Vallcebre Syncline, represented by

B. Vila et al. / Cretaceous Research 32 (2011) 378e386 383
scattered eggshells, and in situ eggs and clutches. The M. sirugueioospecies has been reported in 23 egg levels in seven localities(Table 1). There, samples have been taken from in situ eggs at 15sites. The basalmost egg horizon (E-MUN05 site) is located in thegrey unit, at 15 m above the base of C31r (early Maastrichtian). Theegg-bearing level contains in situ eggs of the oospecies Mega-loolithus siruguei in the Fumanya Nord locality (see details in Bravoet al., 2005). The M. siruguei and M. cf. siruguei oospecies areabundant in the overlaying egg levels up to the upper third of theC31r (Fig. 4). The latest egg level withM. siruguei is N-MAÇ17 in ElsTerrers locality (lateMaastrichtian). There, eggshell fragments occurscattered in fluvial deposits.
Material. 141 well-preserved eggshell fragments, 84 of themsampled from in situ eggs (Table 1)
Description. Eggshell thickness varies between 1.64e3.4 mm(average 2.50 mm). Inner surface covered with rounded and tightlypackedmammillae, which sometimes appear coveredwith a layer offibrous appearance. Outer surface completely covered with roundednodes,with no smooth spaces between them. Pore openings situatedaround the nodes within the irregular interstices between them.Eggshell units are spherulitic, formed by calcite crystals radiatingfrom a basal point, and generally more high than wide. Respiratorychannels are situated between the units, some of them connected bytransversal ones, forming a complex branching network. Eggshellsare in general well preserved, sometimes with secondary depositsalthough some samples show a carbonised appearance.
Comparisons and discussion. As stated above, the oospeciesM. siruguei shows a complex respiratory system structure, whichdoes not appear in other European oospecies (but see Vianey-Liaudet al., 2003). Other characters used in the diagnosis of the oospeciesof the Megaloolithidae such as eggshell units shape, thickness, poreand node diameters, are highly variable (Mikhaïlov, 1997) and thusshould be carefully analysed in order to differentiate variants ofspecimens from oospecies characters. We consider that thecomplex respiratory system is a diagnostic character of M. siruguei,enough to separate it from the other oospecies. This respiratorysystem is present in most of the eggshell samples from the Vall-cebre Syncline.
The samples from one egg-levels at Els Terrers locality (N-MAÇ16) exhibit close similarities to Megaloolithus siruguei. Theseeggshells (N ¼ 4) appear highly crystallised, with respiratorychannels filled with crystalline sediment, thus making difficult theidentification of transversal channels. Nevertheless, in one of theeggshells possible remains of such channels have been observed.Other characteristics are eggshell thickness, which ranges from2.21e2.41 mm (average 2.28 mm) and eggshell units shape, whicharemore high thanwide (fan-shaped units). Although all charactersobserved agree with those of M. siruguei, the small sample and thepreservation conditions, do not allow a precise assignation. For thisreason, we have ascribed this material to M. cf. siruguei.
Megaloolithus mamillare Vianey-Liaud, Mallan, Buscail, andMontgelard, 1994Fig. 3F
Localities and stratigraphic range. Megaloolithus mamillare isrepresented only by scattered eggshell fragments (Table 1). AbovetheM. siruguei egg horizons, theMegaloolithus mamillare oospeciesoccurs at two different stratigraphical levels in Els Terrers locality(N-MAÇ03 and J-BAR03) and in an unique egg level in Peguera (P2-COL01) and Torrent de l’Esdavella (E-MUN06) localities. N-MAÇ03and P2-COL01 are approximately located 22 m and 11 m, respec-tively, below the base of the C31n-C31r reversal, and J-BAR03 islocated some 29m above the base of C31n. The uppermost egg levelincluding Megaloolithus cf. mamillare is E-MUN06, which is located
about 98 m above the base of C31n (late Maastrichtian; Fig. 4). Bothsites are found in mudstones and sandstones of ancient overbankand channel facies, in a fluvial setting.
Material. Eleven eggshell fragments.Description. Thin eggshells (thickness 1.40e1.9 mm, average
1.65 mm) formed by small fan shaped units that are more high thanwide. External surface covered with rounded nodes sometimesfused forming short ridges. Growth lines continue between adja-cent units and appear slightly arched from the base to the top of theunits.
Comparisons and discussion. The eggshells differ fromM. sirugueiin their thickness (<2 mm), and frequent fusion of the nodes.Besides, the respiratory channels are more or less straight with notransversal channels connecting them, so they could be identifiedas a tubocanaliculate pore system (Mikhaïlov, 1991). The eggshellunits are spherulitic as in M. siruguei, but they differ in terms ofproportions (wider than high in M. mamillare). All the abovementioned characteristics correspond to those of Megaloolithusmamillare, Vianey-Liaud et al., 1994. Another similar oospecies isM. pseudomamillare, Vianey-Liaud et al., 1997 (now tentativelyconsidered a junior synonym of M. baghensis, Vianey-Liaud et al.,2003), which differs from M. mamillare only in the pattern ofgrowth lines, which appear horizontal at the base of the units andbecome arched at the top of the same in the former oospecies(Vianey-Liaud et al., 1997). Such a characteristic has not beenobserved in the Vallcebre material, so we consider these thineggshells to belong to the oospecies M. mamillare.
At El Torrent de l’Esdavella locality (E-MUN06) all charactersobserved in the collected samples (i.e. shape of the units, thickness)agreeswith those ofM.mamillare. However, the small sample (N¼ 1)and the preservation conditions, do not allow a precise assignment.For this reason, we have ascribed this material to M. cf. mamillare.
6. Discussion
The continental record of the Aix Basin (France) and thesouthern Pyrenees (Catalonia) contains a rich assemblage of dino-saur oospecies which enables the establishment of some distinctoospecies successions. Based on the French succession, Garcia andVianey-Liaud (2001a) proposed an eggshell biozonation for theEuropean Upper Cretaceous continental deposits. López-Martínez(2000, 2003) used the oospecies from the Tremp basin and Fontl-longa sections to tentatively establish similar successions. However,as López-Martínez (2003) noted, there are important biostrati-graphical conflicts between the Aix Basin and the South-centralPyrenees successions. These disagreements can be solved byrevising the correlation between the sections containing sites andthe magnetic polarity time scale, and contrasting with new egg-bearing calibrated sections.
At the Vallcebre Syncline the stratigraphical location of the 29above-mentioned egg levels builds up a solid oospecies successionthrough the lower Tremp Formation. This is characterized by thesuccessive occurrence of at least three Megaloolithus oospecies(M. sirugueieM. mamillareeM. sp.) which are well correlated withthe magnetic polarity time scale.
On the one hand, the presence of eggs assigned toMegaloolithussp. above M. siruguei and M. mamillare in late Maastrichtian levelsmay indicate that they correspond to M. pseudomamillare oospe-cies, as typically reported in French oospecies successions (Garciaand Vianey-Liaud, 2001a). However, some microstructuraleggshell characters seem not to concur with those in this oospecies(see parataxonomical discussion) and thus a thorough revision ofthis ootaxon is required.
The magnetic and age calibration of megaloolithid oospecies inthe Vallcebre Syncline together with comparison with other

B. Vila et al. / Cretaceous Research 32 (2011) 378e386384
successions enables three main refinements to the chronology ofdinosaur oospecies in southern Europe. First, our results clearlyindicate that the replacement of Megaloolithus siruguei with Mega-loolithus mamillare occurs in the upper part of C31r, in the lateMaastrichtian (Fig. 5A). This replacement is consistent with the datareported in the West Arc Basin (Garcia and Vianey-Liaud, 2001a; p.158; Fig. 4). There, the record ofMegaloolithus siruguei extends to thefirst appearance ofM.mamillare but they did not coexist at all. In theFrench basin, the replacement of the two oospecies most probablyfalls within or below C31n. On the contrary, these successions areapparently in contradiction with the data from the East Arc Basinwhere the authors indicate the coexistence of the two oospecieswithin C31n (Garcia and Vianey-Liaud, 2001a) (Fig. 5B). There, thisoverlap is the result of one egg horizon containing both oospecies inthe Rousset-Erben La Cairanne section plus an isolated locality inMaupague section (Garcia and Vianey-Liaud, 2001a; p. 156; Fig. 3a).However, there are no compelling reasons that this latter locality islocated above the coexistence level at the Rousset-Erben La Cairannesection (see correlation line in Fig. 3a of Garcia and Vianey-Liaud,2001a). Therefore, it is clearly observed that the purportedtemporal overlapofM. siruguei andM.mamillare shown in Figs. 4 and5 of Garcia and Vianey-Liaud (2001a) would be minimum or non-existent.
Second, the refinements and data from both West and East Arcbasins and Vallcebre Syncline reveal that the oospecies successionin the South-central Pyrenees has an inaccurate calibration,a possibility admitted by López-Martínez (2003; p.140). There, theauthor located the Megaloolithus siruguei-M. mamillare replace-ment within the C32n (late Campanian; Fig. 5C), a fact that stronglyconflicts with the abovementioned calibrations of Arc basins andthe Vallcebre Syncline. The disagreement may be due to the lack ofmagnetostratigraphic studies in the “Aren-2” eggshell assemblages(especially those from the Tremp Basin), which therefore leads toproblems with correct correlation. In consequence, in the South-central Pyrenean basins the age andmagnetic calibration of some ofthe egg localities (e.g. Basturs, Suterranya, Abella, Biscarri; Vianey-
Fig. 5. Alternative correlations of the South-European oospecies successions calibrated witgraphic ranges for the M. siruguei-M. mamillare replacement after (a) this study; (b) Garcoospecies assemblages based on a new calibration using the Vallcebre Syncline data, respe
Liaud and López-Martínez, 1997; López-Martínez, 2003) also needsto be revisited.
Third, the herein calibrated oospecies succession and thecomparison with the reviewed previous works have significantimplications for the general chronology of oospecies in southernEurope since it allows a more accurate establishment of the age ofsomeof theoospecies assemblages. Garcia andVianey-Liaud (2001a)reported the Assemblage 2 (containing M. siruguei, possibly the lastM. petralta, and the first M. mamillare) to be older than the Assem-blage 3 (characterized by the possible co-occurrence ofM.mamillareand M. pseudomamillare). However, assemblages were only definedby the occurrence of associated oospecies and not by the chro-nostratigraphical appearance or disappearance of particular ootaxa.Taking into account the minimum or likely non-existent temporaloverlapping of Megaloolithus siruguei and Megaloolithus mamillare(alleged coexistence reported in only one egg horizon), we proposethatMegaloolithusmamillare is not included in theAssemblage 2 andtherefore the Assemblage 3 is then featured by the first appearanceof this oospecies (later coexisting with Megaloolithus pseudoma-millare). The authors also stated that the boundary between theassemblages probably falls in the early Maastrichtian (see Fig. 5 inGarcia and Vianey-Liaud, 2001a; p. 159). However, the age calibra-tion for the Megaloolithus siruguei-Megaloolithus mamillare replace-ment in the Vallcebre Syncline and East and West Arc basinsdefinitively demonstrates that the alleged assemblage boundary fallsaround the C31reC31n polarity change or at the upper part of C31r,entirely in the late Maastrichtian (Fig. 5D).
Finally, diversity studies using oospecies may be useful inorder to assess the true dinosaur diversity. In the case of mega-loolithid oospecies, most authors agree in considering titanosauriansauropods as the most probable egg-layers (Chiappe et al., 2001).However, the correlation between parataxonomic and taxonomicforms ascribed to sauropods is yet tentative and the authors arecautious with their observations. Considering the oospecies assem-blages proposed by Garcia and Vianey-Liaud et al. (2001a) anddiversity works conducted on European Late Cretaceous sauropods
h the magnetic polarity scale. From left to right: standard scale and calibrated strati-ia and Vianey-Liaud (2001a); (c) López-Martínez (2003). d, new refinements on thectively. m ¼ M. mamillare; s ¼ M. siruguei

B. Vila et al. / Cretaceous Research 32 (2011) 378e386 385
(e.g. Vila et al., 2009) we observe that the number of oospeciesexceeds the taxonomic sauropod forms in the Upper Campanian-LowerMaastrichtian interval. This factmust be taken in caution sincean in-depth revision is required for most of the Late Campanian(Assemblage 1) oospecies. On the contrary, we observe that numberof sauropod oospecies roughly fits with the number of taxonomicforms in the chrons 31 and 30. Future works must also consider thelikely relationship between the occurrence of oospecies assemblagesand palaeoenvironmental changes through the Maastrichtian anddefinitively elucidate the most plausible explanation for such varia-tion in the south-western European basins.
7. Conclusions
The Late Cretaceous deposits of the continental Tremp Forma-tion in the Vallcebre Syncline yield a broad oological record ofdinosaurs. The analysis and parataxonomical study of numerouseggshell samples from abundant egg horizons together with anaccurate calibration of multiple stratigraphical sections enable us tobuild up a robust oospecies succession. This essentially features thesuccessive occurrence of three Megaloolithus oospecies (M.sirugueieM. mamillareeM. sp.) which are well correlated with themagnetic polarity time scale. Thus, the egg-bearing levels showa continuous record throughout the C31 and C30. Interestingly, theresults herein presented support data indicating that the replace-ment ofMegaloolithus sirugueiwithMegaloolithus mamillare occursaround the C31reC31n polarity change, in the Late Maastrichtian.As a result, the age of such replacement provides evidence that thealleged boundary for some proposed oospecies assemblages(Assemblage 2 and 3, after Garcia and Vianey-Liaud, 2003) falls inthe Late Maastrichtian. Finally, the revision of previous workssuggests that the age and magnetic calibration of some south-central Pyrenean egg localities need to be revisited.
Acknowledgments
This research was funded by the CGL2007-60054, CGL2008-06533-C03-01, 02/BTE, and CGL2009-09000 projects of the SpanishMinistry of Education and Culture. V.R. is supported by a scholar-ship from the Departament d’Innovació, Universitats i Empresa(Generalitat de Catalunya and European Social Fund). Fieldworkwas undertaken with grants from Direcció General del PatrimoniCultural (Departament de Cultura de la Generalitat de Catalunya).The facilities provided by Albert G. Sellés and Sandra Val (InstitutCatalà de Paleontologia), Serveis Cientifico-tècnics de la Universitatde Barcelona, and Consorci Ruta Minera are highly appreciated. Theauthors thank L. Salgado and one anonymous reviewer for helpfulcomments on the manuscript.
References
Aepler, R., 1967. Das Garumnium der Mulde von Vallcebre und ihre Tektonik(Spanien, Provinz Barcelona). Unpublished MSc Thesis. Freie Universität Berlin(Naturwissenschaftliche Fakultät), Germany, 101 pp.
Bravo, A.M., Vila, B., Galobart, À, Oms, O., 2005. Restos de huevos de dinosaurio en elsinclinal de Vallcebre (Berguedà, Provincia de Barcelona). Revista Española dePaleontología 10, 49e57.
Carpenter, K., 1999. Eggs, nests and baby dinosaurs. Indiana University Press,Bloomington, p. 335.
Chiappe, L.M., Salgado, L., Coria, R., 2001. Embryonic skulls of titanosaur sauropoddinosaurs. Science 293, 2444e2446.
Chiappe, L.M., Schmitt, J.G., Jackson, F.D., Garrido, A., Gingus, L., Grellet-Tinner, G.,2004. Nest structure for sauropods: Sedimentary criteria for recognition ofdinosaur nesting traces. Palaios 19, 89e95.
Cojan, I., Moreau, M.-G., 2006. Correlation of terrestrial climatic fluctuations withglobal signals during the upper Cretaceous-Danian in a compressive setting(Provence, France). Journal of Sedimentary Research 76, 589e604.
Cojan, I., Renard, M., Emmanuel, L., 2003. Palaeoenvironmental reconstruction ofdinosaur nesting sites based on geochemical approach to eggshells and
associated palaeosols (Maastrichtian, Provence basin, France). Palaeogeography,Palaeoclimatology, Palaeoecology 191, 111e138.
Cousin, R., 2002. Organisation des pontes de dinosauriens de la parafamille desMegaloolithidae Zhao, 1979. Bulletin trimestriel de la Société géologique deNormandie et des Amis du Muséum du Havre 89, 1e177.
Deeming, D.C., 2006. Ultrastructural and functional morphology of eggshellssupports the idea that dinosaur eggs were incubated buried in a substrate.Palaeontology 49, 171e185.
Galbrun, B., Feist, M., Colombo, F., Rocchia, R., Tambareau, Y., 1993. Magneto-stratigraphy of CretaceouseTertiary continental deposits, Ager Basin, Province ofLerida, Spain. Palaeogeography, Palaeoclimatology, Palaeoecology 102, 41e52.
Garcia, G., Vianey-Liaud, M., 2001a. Dinosaur eggshells as biochronological markersin Upper Cretaceous continental deposits. Palaeogeography, Palaeoclimatology,Palaeoecology 169, 153e164.
Garcia, G., Vianey-Liaud, M., 2001b. Nouvelles données sur les coquilles d’oeufs dedinosaures Megaloolithidae du Sud de la France: Systématique et variabilitéintraspécifique. Comptes Rendús de l’Academie des Sciences Paris. Earth andPlanetary Sciences 332, 185e191.
Gradstein, F.M., Ogg, J.G., Smith, A.G., 2004. A Geologic Timescale. CambridgeUniversity Press, Cambridge, p. 500.
Jackson, F., Varrichio, J., Jackson, R., Vila, B., Chiappe, L., 2008. Comparison of water-vapor conductance on a titanosaur egg from Argentina with a Megaloolithussiruguei from Spain. Paleobiology 34, 229e246.
López-Martínez, N., 2000. Eggshell sites from the Cretaceous-Tertiary transition inSouth-central Pyrenees (Spain). Extended Abstracts. In: Bravo, A.M., Reyes, T.(Eds.), First International Symposium on Dinosaur Eggs and Babies. Diputacióde Lleida, Lleida, pp. 95e115.
López-Martínez, N., 2003. Dating dinosaur oodiversity: chronostratigraphic controlof Late Cretaceous oospecies succession. Palaeovertebrata 32, 121e148.
Marmi, J., Vila, B., Galobart, À, 2009. Solemys (Chelonii, Solemydidae) remains fromthe Maastrichtian of Pyrenees: evidence for a semi-aquatic lifestyle. CretaceousResearch 30, 1307e1312.
Marmi, J., Vila, B., Oms, O., Galobart, À, Cappetta, H., 2010. Oldest records of stingrayspines (Chondrichthyes, Myliobatiformes). Journal of Vertebrate Paleontology30, 970e974.
Médus, J., Feist, M., Rocchia, R., Batten, D.J., Boclet, D., Colombo, F., Tambareau, Y.,Villatte, J., 1988. Prospects for recognition of the palynological Cretaceous/Tertiary boundary and iridium anomaly in nomarine facies of the easternSpanish Pyrenees: a preliminary report. Newsletter Stratigraphy 18, 123e138.
Mikhaïlov, K.E., 1991. Classification of fossil eggshells of amniotic vertebrates. ActaPalaeontologica Polonica 36, 193e238.
Mikhaïlov, K.E., 1997. Fossil and recent eggshell in amniotic vertebrates: finestructure, comparative morphology and classification. Special Papers in Palae-ontology 56, 1e80.
Oms, O., Dinarès-Turell, J., Vicens, E., Estrada, R., Vila, B., Galobart, À, Bravo, A.M., 2007.Integrated stratigraphy from the Vallcebre basin (southeastern Pyrenees, Spain):new insights on the continental Cretaceous-Tertiary transition in southwestEurope. Palaeogeography, Palaeoclimatology, Palaeoecology 255, 35e47.
Peitz, C., 2000. Megaloolithid dinosaur eggs from the Maastrichtian of Catalunya(NE-Spain) e Parataxonomic implications and stratigraphic utility. ExtendedAbstracts. In: Bravo, A.M., Reyes, T. (Eds.), First International Symposium onDinosaur Eggs and Babies. Diputació de Lleida, Lleida, pp. 155e159.
Riera, V., Marmi, J., Oms, O., Gomez, B., 2010. Orientated plant fragments revealingtidal palaeocurrents in the Fumanya mudflat (Maastrichtian, southern Pyr-enees): Insights in palaeogeographic reconstruction. Palaeogeography, Palae-oclimatology, Palaeoecology 288, 82e92.
Rosell, J., Linares, R., Llompart, C., 2001. El “Garumniense” prepirenaico. RevistaEspañola de Paleontología 14, 47e56.
Sarkar, A., Battacharya, S.K., Mohabey, D.M., 1991. Stable-isotope analysis of dino-saur eggshell paleoenvironmental implications. Geology 19, 1068e1071.
Sanz, J.L., Moratalla, J.J., Díaz-Molina, M., López-Martínez, N., Kälin, O., Vianey-Liaud, M., 1995. Dinosaur nests at the sea shore. Nature 376, 731e732.
Ullastre, J., Masriera, A., 1983. Le passage Crétacé-Tertiare dans les régions sud-pyrenénnes de la Catalogne: données nouvelles. Géologie Mediterranéenne 10,277e281.
Varricchio, D.J., Jackson, F., Borkowski, J.J., Horner, J.R., 1997. Nest and egg clutches ofthe dinosaur Troodon formosus and the evolution of the avian reproductivetraits. Nature 385, 247e250.
Vianey-Liaud, M., López-Martínez, N., 1997. Late Cretaceous dinosaur eggshells fromthe Tremp Basin, southern Pyrenees, Lleida, Spain. Journal of Paleontology 71,1157e1171.
Vianey-liaud, M., Garcia, G., 2000. The interest of French Late Cretaceous dinosaureggs and eggshells. Extended Abstracts. In: Bravo, A.M., Reyes, T. (Eds.), FirstInternational Symposium on Dinosaur Eggs and Babies. Diputació de Lleida,Lleida, pp. 165e175.
Vianey-Liaud, M., Hirsch, K., Sahni, A., Sigé, B., 1997. Late Cretaceous Peruvianeggshells and their relationships with Laurasian and eastern Gondwanianmaterial. Geobios 30, 75e90.
Vianey-Liaud, M., Khosla, A., Garcia, G., 2003. Relationships between European andIndian dinosaur eggs and eggshells of the oofamily Megaloolithidae. Journal ofVertebrate Paleontology 23, 575e585.
Vianey-Liaud, M., Mallan, P., Buscail, O., Montgelard, G., 1994. Review of Frenchdinosaur eggshells: morphology, structure, mineral and organic composition.In: Carpenter, K., Hirsch, K.F., Horner, J.R. (Eds.), Dinosaur Eggs and Babies.Cambridge University Press, Cambrige, pp. 151e183.

B. Vila et al. / Cretaceous Research 32 (2011) 378e386386
Vila, B., Oms, O., Galobart, À, 2005. Manus-only titanosaurid trackway fromFumanya (Maastrichtian, Pyrenees): further evidence for an underprint origin.Lethaia 38, 211e218.
Vila, B., Oms, O., Marmi, J., Galobart, À, 2008. Tracking Fumanya footprints (Maas-trichtian, Pyrenees): historical and ichnological overview. Oryctos 8, 115e130.
Vila, B., Galobart, À, Canudo, J.I, Le Loeuff, J., Oms, O., 2009. Late Cretaceous sauropoddiversity in Southern Europe. Journal of Vertebrate Paleontology 29, 196A.
Vila, B., Jackson, F.D., Fortuny, J., Sellés, A.G., Galobart, À, 2010a. 3-D modelling ofmegaloolithid clutches: insights about nest construction and dinosaur behav-iour. PLoS ONE 5 (5), e10362. doi:10.1371/journal.pone.0010362.
Vila, B., Galobart, À, Oms, O., Poza, B., Bravo, A.M., 2010b. Assessing the nestingstrategies of Late Cretaceous titanosaurs: 3-D clutch geometry from a newMegaloolithid eggsite. Lethaia 43, 197e208.
Westphal, M., Durand, J.P., 1990. Magnétostratigraphie des series continentalsfluvio-lacustres du Crétacé supérieur dans le sinclinal de l’Arc (regiónd’Aix-en-Provence, France). Bulletin Societe Geologique de France 8,609e620.
Zhao, Z., 1979. The advancement of researches on the dinosaurian eggs in China.South China Mesozoic and Cenosoic Red Formation. Science PublishingCompany, Beijing. 329e340.





























