Embed Size (px)
Citation preview

Fungal Genetics and Biology 43 (2006) 261–272
www.elsevier.com/locate/yfgbi
The crtS gene of Xanthophyllomyces dendrorhous encodes a novel cytochrome-P450 hydroxylase involved in the conversion of �-carotene
into astaxanthin and other xanthophylls
Vanessa Álvarez a, Marta Rodríguez-Sáiz a, Juan Luis de la Fuente a, Eduardo J. Gudiña b, Ramiro P. Godio b, Juan F. Martín b, José Luis Barredo a,¤
a R&D Biology, Antibióticos S.A., Avenida de Antibióticos 59-61, 24009 León, Spainb Institute of Biotechnology of León (INBIOTEC) and University of León, Science Park, Av. Real 1, 24006 Leon, Spain
Received 26 September 2005; accepted 19 December 2005Available online 7 February 2006
Abstract
The conversion of �-carotene into xanthophylls is a subject of great scientiWc and industrial interest. We cloned the crtS gene involvedin astaxanthin biosynthesis from two astaxanthin producing strains of Xanthophyllomyces dendrorhous: VKPM Y2410, an astaxanthinoverproducing strain, and the wild type ATCC 24203. In both cases, the ORF has a length of 3166 bp, including 17 introns, and codes fora protein of 62.6 kDa with similarity to cytochrome-P450 hydroxylases. crtS gene sequences from strains VKPM Y2410, ATCC 24203,ATCC 96594, and ATCC 96815 show several nucleotide changes, but none of them causes any amino acid substitution, except a G2268
insertion in the 13th exon of ATCC 96815 which causes a change in the reading frame. A G1470 !A change in the 5� splicing region ofintron 8 was also found in ATCC 96815. Both point mutations explain astaxanthin idiotrophy and �-carotene accumulation in ATCC96815. Mutants accumulating precursors of the astaxanthin biosynthetic pathway were selected from the parental strain VKPM Y2410(red) showing diVerent colors depending on the compound accumulated. Two of them were blocked in the biosynthesis of astaxanthin,M6 (orange; 1% astaxanthin, 71 times more �-carotene) and M7 (orange; 1% astaxanthin, 58 times more �-carotene, 135% canthaxan-thin), whereas the rest produced lower levels of astaxanthin (5–66%) than the parental strain. When the crtS gene was expressed in M7,canthaxanthin accumulation disappeared and astaxanthin production was partially restored. Moreover, astaxanthin biosynthesis wasrestored when X. dendrorhous ATCC 96815 was transformed with the crtS gene. The crtS gene was heterologously expressed in Mucorcircinelloides conferring to this fungus an improved capacity to synthesize �-cryptoxanthin and zeaxanthin, two hydroxylated compoundsfrom �-carotene. These results show that the crtS gene is involved in the conversion of �-carotene into xanthophylls, being potentiallyuseful to engineer carotenoid pathways.© 2005 Elsevier Inc. All rights reserved.
Keywords: �-Carotene; Hydroxylase; Ketolase; Astaxanthin; Canthaxanthin; Zeaxanthin; �-Cryptoxanthin
1. Introduction
Carotenoids are a family of yellow to orange-red terpe-noid pigments synthesized by photosynthetic organismsand by many bacteria and fungi (Britton et al., 1998) usedas colorants, feed supplements, and nutraceuticals in the
* Corresponding author. Fax: +34 987 895986.E-mail address: [email protected] (J.L. Barredo).
1087-1845/$ - see front matter © 2005 Elsevier Inc. All rights reserved.doi:10.1016/j.fgb.2005.12.004
food, medical, and cosmetic industries (Bauernfeind,1981). Although dietary carotenoids inhibit the onset ofmany diseases in which free radicals are involved, only afew of the more than 600 identiWed carotenoids are pro-duced industrially (Bauernfeind, 1981), including �-caro-tene, lycopene, astaxanthin, canthaxanthin, lutein, andcapxanthin (Bhosale, 2004). Commercial production ofcarotenoids from microorganisms is a new approach morefavorable environmentally than synthetic manufactureby chemical procedures. Astaxanthin (3,3�-dihydroxy-�,

262 V. Álvarez et al. / Fungal Genetics and Biology 43 (2006) 261–272
�-carotene-4-4�-dione) is a xanthophyll used as a feedadditive to pigment Xesh in salmon and trout aquaculture(Johnson et al., 1977). Additionally, several positive eVectson degenerative diseases have been reported when asta-xanthin was used as a nutraceutical (Chew et al., 1999;Wang et al., 2000). Due to its increasing importance, bio-technological methods of astaxanthin production havebeen developed with the green alga Haematococcus pluvi-alis and the heterobasidiomycetous yeast Xanthophyllomy-ces dendrorhous, the teleomorphic state of PhaYarhodozyma (Golubev, 1995; Johnson and Lewis, 1979;PhaV et al., 1972).
The biosynthetic pathway for astaxanthin has been stud-ied in X. dendrorhous (Andrewes et al., 1976; Ducrey San-pietro and Kula, 1998; Verdoes et al., 2003; Visser et al.,2003). The conversion of common isoprenoid precursorsinto �-carotene is catalyzed by four enzymatic activities: (i)geranylgeranyl pyrophosphate (GGPP) synthase (encodedby the crtE gene), which converts farnesyl pyrophosphateand isopentenyl pyrophosphate into GGPP; (ii) phytoenesynthase (encoded by the crtYB gene), which links two mol-ecules of GGPP to form phytoene; (iii) phytoene desaturase(encoded by the crtI gene), which introduces four doublebonds in the phytoene molecule to yield lycopene; and (iv)lycopene cyclase (encoded also by the crtYB gene), whichsequentially converts the � acyclic ends of lycopene to �rings to form �-carotene and �-carotene.
Two additional enzymatic activities convert �-caroteneinto astaxanthin through several biosynthetic intermediates(Fig. 1): a ketolase which incorporates two 4-keto groups inthe molecule of �-carotene, and a hydroxylase which intro-
duces two 3-hydroxy groups. The existence of a monocyclicpathway diverging from the dicyclic pathway at neurospo-rene and proceeding through �-zeacarotene, 3,4-dide-hydrolycopene, torulene, 3-hydroxy-3�,4�-didehydro-�,�-carotene-4-one (HDCO) to the end product 3,3�-dihy-droxy-�,�-carotene-4,4�-dione (DCD) has also been pro-posed (An et al., 1999).
The astaxanthin biosynthetic genes crtE (Lodato et al.,2004; Visser et al., 2003), crtI (Verdoes et al., 1999a), andcrtYB (Verdoes et al., 1999b) involved in the steps to �-carotene have been cloned from X. dendrorhous althoughno linear relationship between their transcript levels andcarotenoid biosynthesis was found (Lodato et al., 2004).Additionally, a gene named ast from X. dendrorhousATCC 96594 coding for astaxanthin synthase, a cyto-chrome-P450 type enzyme involved in the conversion of�-carotene, has been subject of a patent application(Hoshino et al., 2000). However, it is unclear whether theenzyme encoded by ast is able to add both the keto andthe hydroxyl groups to the two rings of the �-carotenemolecule, as an in vitro enzymatic reaction has not beendescribed yet (Visser et al., 2003). X. dendrorhous mutantstrains blocked in the step after �-carotene have been iso-lated (Chun et al., 1992; Cifuentes et al., 1997; Girardet al., 1994), but the biochemistry of the blocked reac-tion(s) has not been elucidated.
Therefore, it was of great interest to clone and character-ize the astaxanthin biosynthetic gene(s) from X. dendror-hous involved in the conversion of �-carotene intoastaxanthin, and to determine whether this gene(s) could beused to engineer xanthophyll biosynthetic pathways.
Fig. 1. Potential biosynthetic steps for the conversion of �-carotene and �-carotene into xanthophylls in X. dendrorhous. The ketolase (K) activity incorpo-
β-Carotene
HK
β-Cryptoxanthin(3-hydroxy-β,β-carotene)
Echinenone (β,β-carotene-4-one)
Canthaxanthin(β,β-carotene-4-4’-dione)
Phoenicoxanthin
K H
γ-Carotene
Torulene
HDCO (3-hydroxy-3’,4’-didehydro-β,-ψ-carotene-4-one)
Cyclase (crtYB)
Desaturase (crtI)
O OH
O
O
O
OHO
O
OH
Zeaxanthin(3,3’-dihydroxy-β,β-carotene)
AdonixanthinO OH
OH
Astaxanthin(3,3’-dihydroxy-β,β-carotene-4-4’-dione)
OHO
O OH
3 4
3’
3’-HydroxyechinenoneO
O 3-Hydroxyechinenone
K H
H
K
KH
H
K
KH
H K
OH
OH
OH
OH
4’
rates two keto groups in the molecule of �-carotene and the hydroxylase (H) activity introduces two hydroxyl groups.

V. Álvarez et al. / Fungal Genetics and Biology 43 (2006) 261–272 263
2. Materials and methods
2.1. Microorganisms, culture conditions, and cloning vectors
The astaxanthin producing strains X. dendrorhous VKPMY2410, ATCC 24203 (CECT 11028), and ATCC 96815 (alsodeposited as ATCC 74486) were cultured by Xask fermenta-tion in YEPD broth (10g/L yeast extract, 20 g/L bactopep-tone, and 20g/L glucose). Flasks were inoculated with acolony from a fresh agar plate and incubated at 250 rpm and21 °C for 96 h (de la Fuente et al., 2004; Verdoes et al., 2003).N-Methyl-N�-nitro-N-nitrosoguanidine-induced mutants ofX. dendrorhous were selected in YEPDA medium asdescribed previously (de la Fuente et al., 2004). Mucor circi-nelloides carotenoid production was tested following growthon YNB plates for 4 days (Velayos et al., 2000). Escherichiacoli SURE (Stratagene) and E. coli LE392 (Sambrook et al.,1989) were the hosts for �GEM12 (Promega) phage deriva-tives. Phage �GEM12 was used to construct genomiclibraries of X. dendrorhous VKPM Y2410 and ATCC 24203.E. coli DH5� (Sambrook et al., 1989) was the recipient strainfor high-frequency plasmid transformation. pBluescriptphagemids (Stratagene) and pGEM-T (Promega) were usedfor subcloning and sequencing. General procedures for plas-mid DNA puriWcation, cloning, and transformation of E. coliwere those of Sambrook et al. (1989).
2.2. Construction and screening of X. dendrorhous genomic libraries
The DNAs from X. dendrorhous VKPM Y2410 andATCC 24203 were puriWed as previously described (Spechtet al., 1982). Fragments of genomic DNA (17–22 kb) wereselected from Sau3AI partially digested DNA, ligated to�GEM12, and packaged with the “Packagene” kit(Promega) to yield about 2£ 104 pfus. For screening, thelibraries were ampliWed in E. coli LE392, plated to obtainabout 1.5£ 104 pfus, and hybridized with [32P]dCTP-labeled probes according to standard methods (Sambrooket al., 1989). Recombinant phages were ampliWed in liquidmedium to purify their DNA (Sambrook et al., 1989).
2.3. PCR ampliWcation
Four oligonucleotides were designed according to con-served sequences from crtW or crtZ genes of Arabidopsis tha-liana, Lycopersicon esculentum, Capsicum annuum, Pantoeaananas, Pantoea agglomerans, Flavobacterium sp., Erwiniaherbicola, Agrobacterium aurantiacum, Paracoccus marcusii,Alcaligenes sp., and H. pluvialis: P65 (forward; 5�-GGWTGGCACGAGTCYCACCAC-3�) and P66 (reverse; 5�-CTGGTGRACRAGTTCGTCGTG-3�) to amplify a fragment ofthe crtZ gene, and P68 (forward; 5�-TTCATCATCGCYCACGAYGCCATG-3�) and P69 (reverse; 5�-GGGRAGCCARGTTCCGAAGACGAA-3�) for the crtW gene. To clonethe ast gene (Hoshino et al., 2000), two probes of 1.6 and1.3 kb were ampliWed with primers P102 (forward; 5�-CGCC
GCCAACCTGCCTGAAT-3�) and P97 (reverse; 5�-GGGTAAGAAGAGAGTCCAGAGCGACGATAG-3�), and withprimers P102 (forward) and P103 (reverse; 5�-GGACCCGACGGAGAGTGACAAACA-3�), respectively. OptimizedampliWcation reactions (20�l in a DNA Thermal Cycler 480,Perkin-Elmer, Norwalk, CT) contained about 100ng ofgenomic DNA, 20mM Tris–HCl, pH 8.8, 2mM MgSO4,10mM KCl, 10 mM (NH4)2SO4, 0.1% Triton X-100, 0.1 mg/ml nuclease-free BSA, 0.5�M of each primer, 1.0U of TurboPfu DNA polymerase (Stratagene), and dNTPs 200�M each.The reaction mixtures were overlaid with mineral oil andsubjected to 35 PCR cycles with the following programs: (i)primers P65 and P66 [94°C, 30 s (120 s for the Wrst cycle);45°C, 60 s; 72 °C, 60 s]; (ii) P68 and P69 [94 °C, 60s (120s forthe Wrst cycle); 51 °C, 60 s; 72 °C, 60 s]; (iii) P97 and P102[94°C, 10s (120 s for the Wrst cycle); 48 °C, 30 s; 72 °C, 45s];(iv) P102 and P103 [94°C, 10 s (120 s for the Wrst cycle); 48 °C,30s; 72 °C, 45s]. Total RNA from X. dendrorhous VKPMY2410 was incubated at 50°C for 30 min, and subjected to 35RT-PCR cycles [94°C, 10 s; 58°C, 30 s; 68 °C, 60 s (extending5s per cycle after 11th)] with primers P126 (forward; 5�-CTCCCCATGGTCATCTTGGTCTTGCTC-3�) and P127(reverse; 5�-GAATCAACTCATTCGACCGGC-3�), usingthe “Titan One Tube RT-PCR System” (Roche MolecularBiochemicals), to amplify the cDNA from the crtS gene as a1674bp fragment. AmpliWed fragments were puriWed fromagarose gels using the “Qiaex II Gel Extraction kit” (Qiagen)and subcloned into pGEM-T (Promega).
2.4. Nucleic acid hybridization and sequencing
Southern hybridizations were made with digoxigenin-labeled probes (“Non-radioactive labeling and immunologi-cal detection kit,” Roche Molecular Biochemicals) accordingto the recommendations of the manufacturer. Cells after 72and 96h from a fed-batch fermentation for astaxanthin pro-duction were frozen in liquid nitrogen, and total RNA wasrecovered, resolved by agarose-formaldehyde gel electropho-resis, and subjected to RT-PCR. Clones to be sequenced wereconstructed with the “Erase a base” system (Promega) andsequenced with an ABI PRISM 377 DNA sequencer(Applied Biosystems). DNA sequences were analyzed withthe Dnastar and Winstar packages (DNASTAR), and com-pared to known sequences by using the BLASTP andBLASTX programs (Altschul et al., 1997). The codon-prefer-ence algorithm (Geneplot, Winstar) was used to detect thepresence of ORFs. Multiple protein alignments were madewith the Clustal V algorithm and pair wise alignments withthe Lipman–Pearson method (MegAlign, Winstar).
2.5. crtS gene expression
Transformation of X. dendrorhous �-carotene producingmutants M7 and ATCC 96815 was mediated byAgrobacterium tumefaciens AGL-1 (Lazo et al., 1991) usingthe integrative plasmids pALPR200 and pBGcrtS5, respec-tively. pALPR200 was constructed by excising a HindIII–

264 V. Álvarez et al. / Fungal Genetics and Biology 43 (2006) 261–272
SacI fragment including the GUS gene from pBI121 (Clon-tech) and inserting a 6.0kb PvuII fragment including the crtSgene from X. dendrorhous VKPM Y2410 and the hygromycinresistance gene. pBGcrtS5 was constructed by excising aPvuII fragment including the EGFP (enhanced green Xuores-cent protein) encoding gene from pBGgHg (Chen et al., 2000)and inserting a 4.0kb PstI fragment containing the crtS genefrom X. dendrorhous ATCC 24203. Plasmid pALMC11 wasconstructed, to express the crtS gene in M. circinelloidesMS12 (leuA1, pyrG4), by inserting into pLeu4 (Roncero et al.,1989) the cDNA from the crtS gene (1.6kb NcoI–NotI)expressed under the control of the carRA promoter fromBlakeslea trispora (611pb NcoI; Rodríguez-Sáiz et al., 2004).M. circinelloides MS12 was transformed as previouslydescribed (Ruiz-Hidalgo et al., 1999) with a mix of 1.5mg/mlCaylase (Cayla, Toulouse, France) and 4–12U/ml of Strepto-zyme for protoplast preparation. Several transformants ofeach plasmid, prototrophic for leucine, were selected on YNBminimal medium (Lampila et al., 1985) supplemented withuridine and osmotically stabilized with 0.55M sorbitol.
2.6. Carotenoid analysis
Carotenoids were extracted with methanol/dichlorometh-ane (50/50) and re-dissolved in acetone. Thin layer chroma-tography (TLC) analyses were done with dichloromethane/
ethyl acetate (80/20) as the mobile phase and aluminum sheetsilica gel 60 (DC-alufolien Kielselgel 60, Merck) as the sta-tionary phase. High-performance liquid chromatography(HPLC) analyses were performed at 474 nm with a Nucleosil100 NH2 column (resin particle diameter, 5�m; 250 by4.6 mm) and hexane/ethyl acetate (50/50 ratio) with a Xowrate of 1 ml/min as the mobile phase.
3. Results
3.1. Cloning and characterization of the crtS gene from X. dendrorhous
The DNA probes ampliWed by PCR using primer pairsP65–P66 (crtZ) and P68–P69 (crtW) (see Materials andmethods) were sequenced and compared to SwissProt andPIR databases, but unfortunately none of them showed sig-niWcant similarity to bacterial crtW or crtZ genes. However,the 1.6 and 1.3 kb probes ampliWed from X. dendrorhousVKPM Y2410 with primer pairs P97–P102 and P102–P103matched the ast sequence from X. dendrorhous ATCC96594 (Hoshino et al., 2000). Two genomic libraries from X.dendrorhous VKPM Y2410 and ATCC 24203 werescreened with the 1.6 and 1.3 kb probes, and three positivephages were isolated from VKPM Y2410 (fALPR2,fALPR3, and fALPR4) and two from ATCC 24203
Fig. 2. (A) Restriction map of the genomic DNA region of X. dendrorhous including the crtS gene (big arrow). Exons encoding the deduced protein(62.6 kDa) are shown as boxes from E-1 to E-18. Little arrows show putative glycosylation sites located in amino acids 42, 86, 219, and 449 of CrtS. Pro-moter region: pyrimidine-rich stretches, TATA box, and CAAT boxes are bold and underlined; CreA-binding motifs are boxed. (B) Comparison of crtSgene sequences of X. dendrorhous strains ATCC 24203 (DQ202402), ATCC 96815 (AY946023), ATCC 96594 (AX034666), and VKPM Y2410(DQ201828). The number of the intron (I-) or exon (E-) where the change was found and their positions with reference to the ATG translation start codonare shown. ATCC 96815 has a G1470 ! A change in the 5� splicing region of I-8 and a G2268 insertion in E-13 which cause astaxanthin idiotrophy and�-carotene accumulation.
Intron / Exon I-1 I-4 I-6 E-7 I-7 I-8 E-9 E-11 E-13 I-15 I-17
Position (bp) 158
695
1108
1110
1120
1134
1137
1153
1191
1245
1257
1287
1347
1354
1356
1377
1378
1470
1480
1488
1493
1523
1538
1540
1541
1601
1607
1619
1628
1952
2247
2268
2677
2931
ATCC 24203 T G T C T A T T G T T A C A C C T G A C T C T C G T T T G T T - G T
ATCC 96815 T G C T C G C C G C C G T C T T C A G T C T C A A C C C A T T G G T
ATCC 96594 A A T C T A T T A T T G C A C C T G A C T C T C G T T T G C G - T G
VKPM Y2410 A A T C T A T T A T T G C A C C T G A C T C T C G T T T G C G - T G
B
A
Eco
RI
Xba
I
Sal
I
Xba
I
Sm
aI
Sal
I
Nco
I
GTGCGGCTTTAGGCTCTGCAACTGCGTCTGGAGCAAGTGCTGCCACTGAGGCGGCTGTGGCATCACCGGCGATTAAAGGCGCGGCCGGCGGACCATCCATCAGAGGTGCAGCCGCTGCTGGACGAGGTGGGGCCTCAACCAGGGGTGCTAGAGGTGGACGGGCGCCGACGACATTATTCGCAGGTGCACTGGGTAAAGCCCAGGCAAGTGGAACGGGCACAAAACGAGCTCGACCGGAAGATGATGCGGCGGCAACTGGTTCTGGGCTGGGGAACACCCTATTCGACCGGTTGAATGAGTCACAATCGGCCGGCAGCCCTGCTGCAGGCACCGGCGCAAGTGGCTCCGGACCGAAGCCTCGCCAGCAGTTGATCAAGCGAACCAAGCCGAACAATCCTGGCGCGCCTGGAGGAGCGGGAGCGGGAGGAGCAGCAGGTGATGCATCGGGTGGACAGAATCAGTAGTGTGTGTGTATGTGTGTAGTGTAGTTGGGTTGTCCCATGTGCTTCTTCTTATCATCATCATTTCTTTAAAATCTCTACATTGAATGTTTACCGGAACGGGCTTTGATGATACTACGGACCACGTTGTGTAACCAGTTCGATTGAGATTACGATTAGATAGCCGATCCGTCGATCAGATCTCGATCTAGAGCGACATCTGGCTCGATCGGTCCTTGCCGAAAATCAGGGCACCGATCAGGGCAGAGGAACGCCGAGGCCGAACGAGACAGACACACCATCATCATCAGCCATGTCTTTTTTGTGATCGTTTTTACATACTACCCGTCGATTCTAACCTTCTTTCTTCTTCTCTTGCCATCTTTGCATTCTCTATCTCGTGTAACATCGATCCGATTCTTGCCACCTACTTTCTCCATATG
crtS
Pst
I
Pst
I
Sm
aI
E-1 E-2 E-3 E-4 E-5 E-6 E-7 E-8 E-9 E-10 E-11 E-12 E-13 E-14 E-15E-16 E-17 E-18

V. Álvarez et al. / Fungal Genetics and Biology 43 (2006) 261–272 265
(fcrtS58 and fcrtS160). All of them were isolated andmapped, showing the hybridization signals in fragments of4.2 kb SmaI, 4.0 kb PstI, and 2.5 kb SalI (Fig. 2A).
A 6.0 kb fragment from VKPM Y2410 including 2.0 kbupstream of crtS, the complete ORF, and 1.0 kb downstreamwas cloned into pBluescript II KS (+) to give pALPR105.This fragment was sequenced in both strands (GenBankAccession no. DQ201828) and analyzed with the codon-pref-erence algorithm (Geneplot) showing a 3166bp ORFconsisting of eighteen exons (at positions 1–118, 201–249,365–481, 554–645, 769–983, 1067–1100, 1190–1332, 1406–1465, 1548–1647, 1724–1820, 1898–2008, 2085–2118, 2209–2280, 2429–2516, 2580–2637, 2714–2787, 2865–2889, and2980–3166) and 17 intervening sequences, as deduced fromthe cDNA sequence (Fig. 2A). The nucleotide sequence ofstrain ATCC 24203 was also determined (GenBank Acces-sion No. DQ202402) and encoded the same protein.
Sequences known to be important in transcription initia-tion were detected at the 5�-Xanking region (Fig. 2A), includ-ing pyrimidine-rich regions between positions ¡47 to ¡82and ¡105 to ¡124 (with reference to the ATG translationstart codon at position 1905), a TATA-like box (TTTAAAAat position ¡352; consensus TATAAA), and two potentialCAAT boxes at positions ¡489 and ¡578. Additionally, fourCreA-binding motifs [(G/C)(C/T)GG(A/G)G] (Cubero andScazzocchio, 1994) were found at ¡475, ¡614, ¡754, and¡853. The consensus sequence AATAAA, potentiallyinvolved in the polyadenylation of the 3� terminus, wasfound 434 bp downstream the translation end codon TGA.
crtS gene sequence from VKPM Y2410 showed severalbase changes when compared to the sequences from the cor-responding gene from ATCC 24203, ATCC 96815 (Gen-Bank Accession No. AY946023) and ATCC 96594(GenBank Accession no. AX034666). Changes are located inexons 7, 9, 11, and 13, and in introns 1, 4, 6, 7, 8, 15, and 17,but none of them causes any amino acid substitution, excepta G2268 insertion in the 13th exon of ATCC 96815 whichcauses a change in the reading frame. A G1470!A change inthe 5� splicing region of intron 8 was also found in ATCC96815. Both point mutations explain astaxanthin idiotrophyand �-carotene accumulation in ATCC 96815 (Fig. 2B).
3.2. Characterization of the deduced polypeptide
The deduced polypeptide has a predicted molecularmass of 62,630 Da and a pI of 5.75, and showed similarityto cytochrome-P450 monooxygenases. Comparison tocytochrome P450 from diVerent families allowed to identifythe oxygen-binding site (339AGYETS344) including theinvariant G340 and T343 residues common to all cytochromeP450 proteins (Teutsch et al., 1993), the heme-bindingdomain (488FISGPRACFG497) thought to be involved inthe folding of the heme-binding pocket (Poulos et al., 1987),and a domain (394ESLR397) found in all cytochrome P450proteins involved in the maintenance of the three-dimen-sional structure (Yu and Strobel, 1997) (Fig. 3A). Conser-vation at the Cys495, which provides the thiolate ligand to
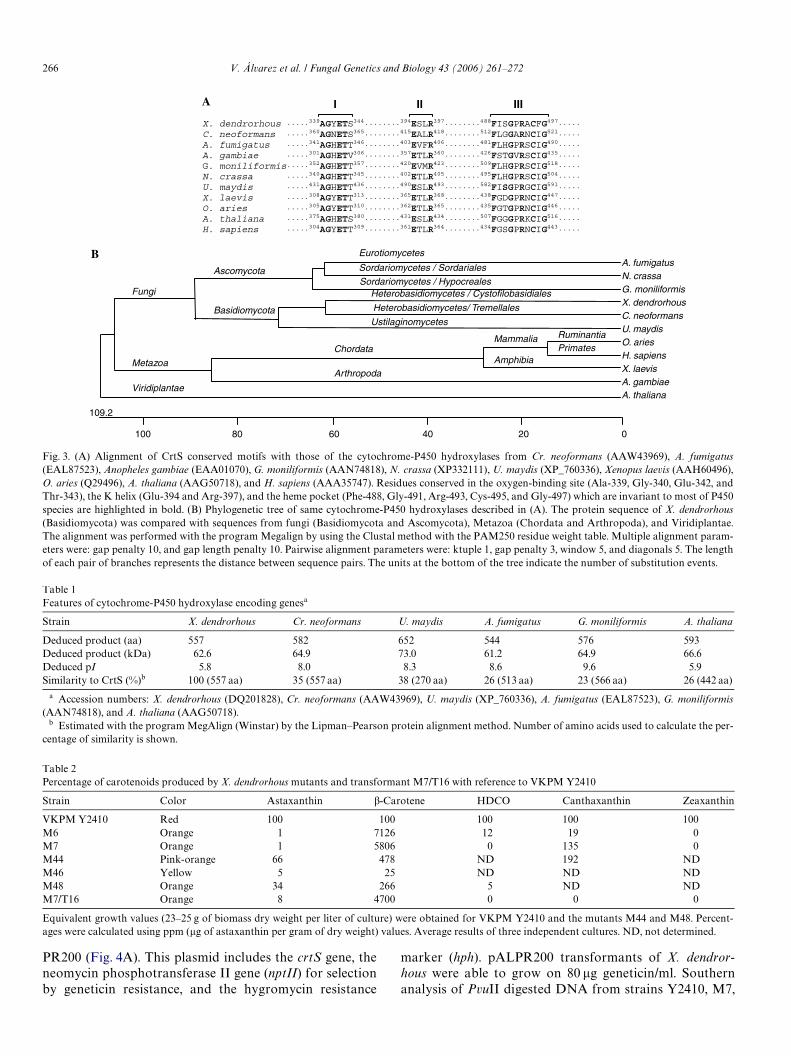
the heme group (Shimizu et al., 1988), was also observed.The deduced amino acid sequence includes four potentialN-glycosylation motifs [NX(T/S)] (Marshall, 1972) at posi-tions 42, 86, 219, and 449 (Fig. 2A).
A phylogenetic tree was constructed showing the evolu-tive relationship of CrtS with cytochrome-P450 hydroxy-lases from bacterial, fungal, and higher eukaryote origins(Fig. 3B). The CrtS protein was closely related to cyto-chrome-P450 hydroxylases of fungi, especially to thosefrom the Basidiomycota Cryptococcus neoformans andUstilago maydis, and the Ascomycota Aspergillus fumiga-tus, Giberella moniliformis, and Neurospora crassa (Table 1).However, it showed insigniWcant identity to the crtZ geneproduct (�-carotene hydroxylase) involved in carotenoidbiosynthesis in bacteria (Ag. aurantiacum, Pa. marcusii,Alcaligenes sp., P. ananas, P. agglomerans, Flavobacteriumsp., E. herbicola, and Erwinia uredovora), the green micro-alga H. pluvialis, and plants (A. thaliana, L. esculentum, andC. annuum). Furthermore, it lacks identity to the crtW geneproducts (�-carotene ketolase) in the databases.
3.3. Isolation of astaxanthin non-producing and/or �-carotene overproducing mutants of X. dendrorhous by classical mutagenesis
In order to obtain X. dendrorhous mutants blocked in theconversion of �-carotene into astaxanthin, classical randommutation and screening were done. A total of 60 N-methyl-N�-nitro-N-nitrosoguanidine-induced mutants were selectedbecause of their diVerent visual color pattern (orange, intenseyellow, pale yellow, or white) with reference to the parentalstrain X. dendrorhous VKPM Y2410 (red). After TLC analy-sis of carotenoids, Wve of them (M6, M7, M44, M46, andM48) were selected as potential astaxanthin non-producingand/or �-carotene overproducing mutants. SubsequentHPLC analyses and comparison to the parental VKPMY2410 strain (Table 2) showed that mutants M6 (orange)and M7 (orange) almost did not produce astaxanthin (1%)and overproduced �-carotene (71 and 58 times more,respectively); M7 also overproduced canthaxanthin (�,�-car-otene-4-4�-dione; 135%), whereas only 19% canthaxanthinwas produced by M6; mutants M44 (pink-orange) and M48(orange) produced lower quantity of astaxanthin (66 and34%, respectively) but overproduced �-carotene (478 and266%, respectively). Astaxanthin (5%) and �-carotene (25%)production was poorer in mutant M46 (yellow). Accordingto these results, M7 was selected to express the crtS gene.Additionally, X. dendrorhous ATCC 96815, an astaxanthinnon-producing and �-carotene accumulating mutant, wasused for crtS gene expression.
3.4. crtS gene expression in X. dendrorhous M7 partially restored astaxanthin production
The ability of crtS gene to complement the astaxanthinidiotrophy of M7 mutant was tested by Ag. tumefaciensAGL-1 mediated transformation with plasmid pAL-
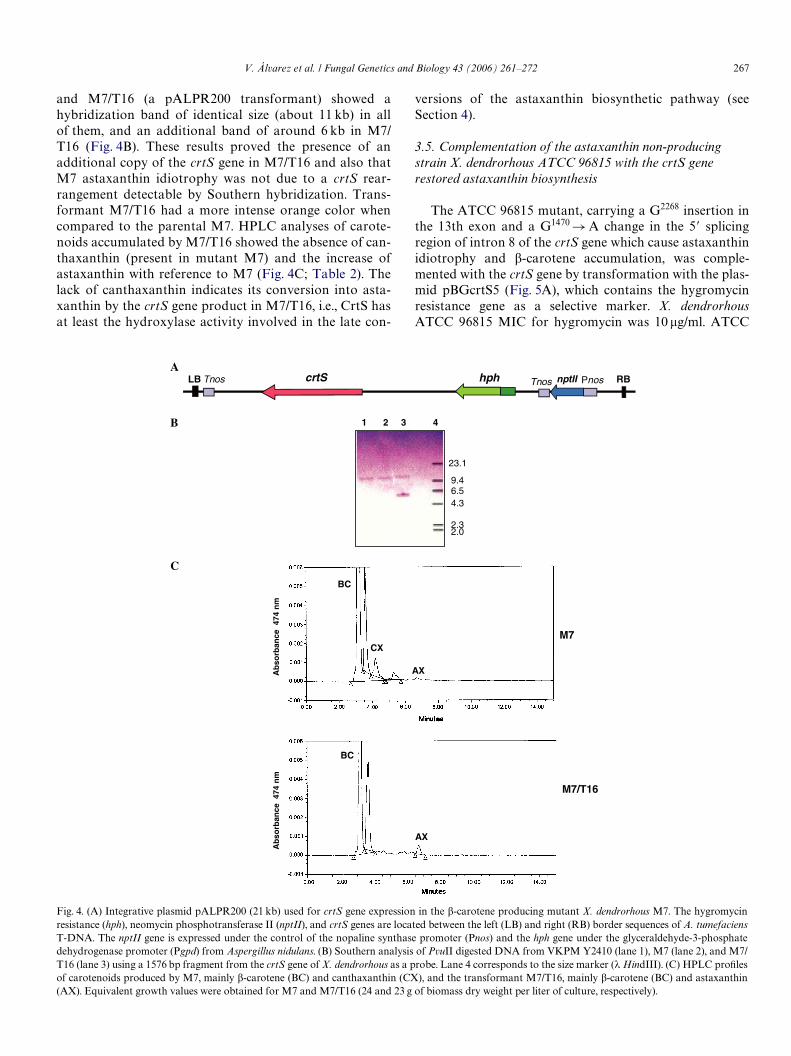
266 V. Álvarez et al. / Fungal Genetics and Biology 43 (2006) 261–272
PR200 (Fig. 4A). This plasmid includes the crtS gene, theneomycin phosphotransferase II gene (nptII) for selectionby geneticin resistance, and the hygromycin resistance
marker (hph). pALPR200 transformants of X. dendror-hous were able to grow on 80 �g geneticin/ml. Southernanalysis of PvuII digested DNA from strains Y2410, M7,
Fig. 3. (A) Alignment of CrtS conserved motifs with those of the cytochrome-P450 hydroxylases from Cr. neoformans (AAW43969), A. fumigatus(EAL87523), Anopheles gambiae (EAA01070), G. moniliformis (AAN74818), N. crassa (XP332111), U. maydis (XP_760336), Xenopus laevis (AAH60496),O. aries (Q29496), A. thaliana (AAG50718), and H. sapiens (AAA35747). Residues conserved in the oxygen-binding site (Ala-339, Gly-340, Glu-342, andThr-343), the K helix (Glu-394 and Arg-397), and the heme pocket (Phe-488, Gly-491, Arg-493, Cys-495, and Gly-497) which are invariant to most of P450species are highlighted in bold. (B) Phylogenetic tree of same cytochrome-P450 hydroxylases described in (A). The protein sequence of X. dendrorhous(Basidiomycota) was compared with sequences from fungi (Basidiomycota and Ascomycota), Metazoa (Chordata and Arthropoda), and Viridiplantae.The alignment was performed with the program Megalign by using the Clustal method with the PAM250 residue weight table. Multiple alignment param-eters were: gap penalty 10, and gap length penalty 10. Pairwise alignment parameters were: ktuple 1, gap penalty 3, window 5, and diagonals 5. The lengthof each pair of branches represents the distance between sequence pairs. The units at the bottom of the tree indicate the number of substitution events.
X. dendrorhous ·····339AGYETS344········394ESLR397········488FISGPRACFG497····· C. neoformans ·····360AGNETS365········415EALR418········512FLGGARNCIG521····· A. fumigatus ·····341AGHETT346········403EVFR406········481FLHGPRSCIG490·····A. gambiae ·····301AGHETV306········357ETLR360········426FSTGVRSCIG435·····
G. moniliformis·····352AGHETT357········420EVMR423········509FLHGPRSCIG518····· N. crassa ·····340AGHETT345········402ETLR405········495FLHGPRSCIG504····· U. maydis ·····431AGHETT436········490ESLR493········582FISGPRGCIG591·····X. laevis ·····308AGYETT313········365ETLR368········438FGDGPRNCIG447·····
O. aries ·····305AGYETT310········362ETLR365········435FGTGPRNCIG446····· A. thaliana ·····375AGHETS380········431ESLR434········507FGGGPRKCIG516····· H. sapiens ·····304AGYETT309········361ETLR364········434FGSGPRNCIG443·····
III III
A. fumigatus N. crassa G. moniliformis X. dendrorhous C. neoformans U. maydis O. aries H. sapiens X. laevis A. gambiae A. thaliana
Eurotiomycetes
Sordariomycetes / Hypocreales Heterobasidiomycetes / Cystofilobasidiales
Heterobasidiomycetes/ Tremellales
Ustilaginomycetes
Chordata
Fungi
Metazoa
Viridiplantae
Arthropoda
Mammalia
Amphibia
Basidiomycota
Ascomycota Sordariomycetes / Sordariales
Ruminantia Primates
100 80 60 40 20 0
109.2
A
B
Table 1Features of cytochrome-P450 hydroxylase encoding genesa
a Accession numbers: X. dendrorhous (DQ201828), Cr. neoformans (AAW43969), U. maydis (XP_760336), A. fumigatus (EAL87523), G. moniliformis(AAN74818), and A. thaliana (AAG50718).
b Estimated with the program MegAlign (Winstar) by the Lipman–Pearson protein alignment method. Number of amino acids used to calculate the per-centage of similarity is shown.
Strain X. dendrorhous Cr. neoformans U. maydis A. fumigatus G. moniliformis A. thaliana
Deduced product (aa) 557 582 652 544 576 593Deduced product (kDa) 62.6 64.9 73.0 61.2 64.9 66.6Deduced pI 5.8 8.0 8.3 8.6 9.6 5.9Similarity to CrtS (%)b 100 (557 aa) 35 (557 aa) 38 (270 aa) 26 (513 aa) 23 (566 aa) 26 (442 aa)
Table 2Percentage of carotenoids produced by X. dendrorhous mutants and transformant M7/T16 with reference to VKPM Y2410
Equivalent growth values (23–25 g of biomass dry weight per liter of culture) were obtained for VKPM Y2410 and the mutants M44 and M48. Percent-ages were calculated using ppm (�g of astaxanthin per gram of dry weight) values. Average results of three independent cultures. ND, not determined.
Strain Color Astaxanthin �-Carotene HDCO Canthaxanthin Zeaxanthin
VKPM Y2410 Red 100 100 100 100 100M6 Orange 1 7126 12 19 0M7 Orange 1 5806 0 135 0M44 Pink-orange 66 478 ND 192 NDM46 Yellow 5 25 ND ND NDM48 Orange 34 266 5 ND NDM7/T16 Orange 8 4700 0 0 0
V. Álvarez et al. / Fungal Genetics and Biology 43 (2006) 261–272 267
and M7/T16 (a pALPR200 transformant) showed ahybridization band of identical size (about 11 kb) in allof them, and an additional band of around 6 kb in M7/T16 (Fig. 4B). These results proved the presence of anadditional copy of the crtS gene in M7/T16 and also thatM7 astaxanthin idiotrophy was not due to a crtS rear-rangement detectable by Southern hybridization. Trans-formant M7/T16 had a more intense orange color whencompared to the parental M7. HPLC analyses of carote-noids accumulated by M7/T16 showed the absence of can-thaxanthin (present in mutant M7) and the increase ofastaxanthin with reference to M7 (Fig. 4C; Table 2). Thelack of canthaxanthin indicates its conversion into asta-xanthin by the crtS gene product in M7/T16, i.e., CrtS hasat least the hydroxylase activity involved in the late con-
versions of the astaxanthin biosynthetic pathway (seeSection 4).
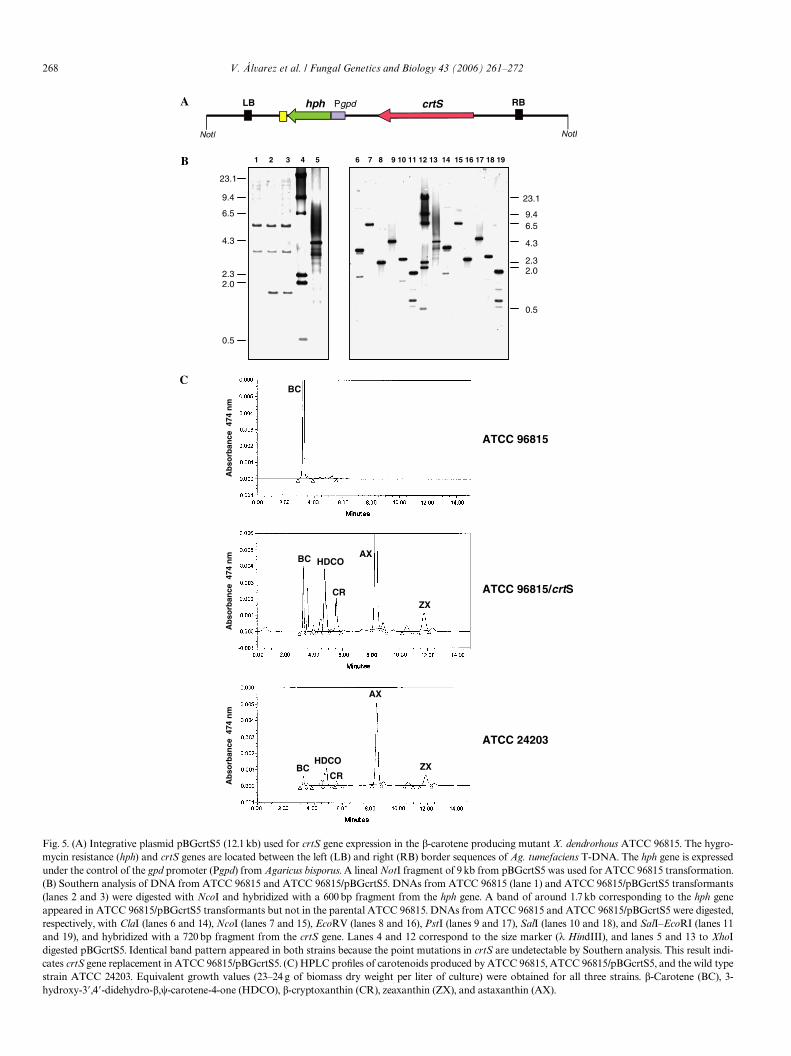
3.5. Complementation of the astaxanthin non-producing strain X. dendrorhous ATCC 96815 with the crtS gene restored astaxanthin biosynthesis
The ATCC 96815 mutant, carrying a G2268 insertion inthe 13th exon and a G1470 !A change in the 5� splicingregion of intron 8 of the crtS gene which cause astaxanthinidiotrophy and �-carotene accumulation, was comple-mented with the crtS gene by transformation with the plas-mid pBGcrtS5 (Fig. 5A), which contains the hygromycinresistance gene as a selective marker. X. dendrorhousATCC 96815 MIC for hygromycin was 10 �g/ml. ATCC
Fig. 4. (A) Integrative plasmid pALPR200 (21 kb) used for crtS gene expression in the �-carotene producing mutant X. dendrorhous M7. The hygromycinresistance (hph), neomycin phosphotransferase II (nptII), and crtS genes are located between the left (LB) and right (RB) border sequences of A. tumefaciensT-DNA. The nptII gene is expressed under the control of the nopaline synthase promoter (Pnos) and the hph gene under the glyceraldehyde-3-phosphatedehydrogenase promoter (Pgpd) from Aspergillus nidulans. (B) Southern analysis of PvuII digested DNA from VKPM Y2410 (lane 1), M7 (lane 2), and M7/T16 (lane 3) using a 1576 bp fragment from the crtS gene of X. dendrorhous as a probe. Lane 4 corresponds to the size marker (� HindIII). (C) HPLC proWlesof carotenoids produced by M7, mainly �-carotene (BC) and canthaxanthin (CX), and the transformant M7/T16, mainly �-carotene (BC) and astaxanthin(AX). Equivalent growth values were obtained for M7 and M7/T16 (24 and 23 g of biomass dry weight per liter of culture, respectively).
A
B
C
LB RBPnosTnosTnos hph nptIIcrtS
23.1
9.46.54.3
2.32.0
1 2 3 4
M7
AX
BC
Ab
sorb
ance
474
nm
CX
BC
Ab
sorb
ance
474
nm
AX
M7/T16
268 V. Álvarez et al. / Fungal Genetics and Biology 43 (2006) 261–272
Fig. 5. (A) Integrative plasmid pBGcrtS5 (12.1 kb) used for crtS gene expression in the �-carotene producing mutant X. dendrorhous ATCC 96815. The hygro-mycin resistance (hph) and crtS genes are located between the left (LB) and right (RB) border sequences of Ag. tumefaciens T-DNA. The hph gene is expressedunder the control of the gpd promoter (Pgpd) from Agaricus bisporus. A lineal NotI fragment of 9 kb from pBGcrtS5 was used for ATCC 96815 transformation.(B) Southern analysis of DNA from ATCC 96815 and ATCC 96815/pBGcrtS5. DNAs from ATCC 96815 (lane 1) and ATCC 96815/pBGcrtS5 transformants(lanes 2 and 3) were digested with NcoI and hybridized with a 600 bp fragment from the hph gene. A band of around 1.7 kb corresponding to the hph geneappeared in ATCC 96815/pBGcrtS5 transformants but not in the parental ATCC 96815. DNAs from ATCC 96815 and ATCC 96815/pBGcrtS5 were digested,respectively, with ClaI (lanes 6 and 14), NcoI (lanes 7 and 15), EcoRV (lanes 8 and 16), PstI (lanes 9 and 17), SalI (lanes 10 and 18), and SalI–EcoRI (lanes 11and 19), and hybridized with a 720 bp fragment from the crtS gene. Lanes 4 and 12 correspond to the size marker (� HindIII), and lanes 5 and 13 to XhoIdigested pBGcrtS5. Identical band pattern appeared in both strains because the point mutations in crtS are undetectable by Southern analysis. This result indi-cates crtS gene replacement in ATCC 96815/pBGcrtS5. (C) HPLC proWles of carotenoids produced by ATCC 96815, ATCC 96815/pBGcrtS5, and the wild typestrain ATCC 24203. Equivalent growth values (23–24 g of biomass dry weight per liter of culture) were obtained for all three strains. �-Carotene (BC), 3-hydroxy-3�,4�-didehydro-�,�-carotene-4-one (HDCO), �-cryptoxanthin (CR), zeaxanthin (ZX), and astaxanthin (AX).
A
B
C
Pgpdhph crtS RBLB
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
ATCC 96815/crtS
ATCC 96815
ATCC 24203
BC HDCOAX
CRZX
Ab
sorb
ance
474
nm
BCHDCO
AX
CRZX
Ab
sorb
ance
474
nm
BC
Ab
sorb
ance
474
nm
23.1
9.4
6.5
4.3
2.3 2.0
0.5
23.1
9.4 6.5
4.3
2.3 2.0
0.5

V. Álvarez et al. / Fungal Genetics and Biology 43 (2006) 261–272 269
96815/pBGcrtS5 transformants resistant to 30 �g hygro-mycin/ml were selected and tested by Southern hybridiza-tion with probes from the crtS and hph genes, showing thepresence of the hph gene and an undamaged copy of thecrtS gene (Fig. 5B). One of the transformants was tested byTLC and HPLC analyses showing, as well as �-carotene,HDCO(3-hydroxy-3�,4�-didehydro-�,�-carotene-4-one),�-cryptoxanthin (3-hydroxy-�,�-carotene), astaxanthin,and zeaxanthin (3,3�-dihydroxy-�,�-carotene) accumula-tion, similar xanthophylls proWle as the wild type ATCC24203 (Fig. 5C). These results indicate that crtS indeedcomplements the mutation of ATCC 96815 and conWrmthe involvement of this gene in astaxanthin biosynthesis.The simultaneous restoration of astaxanthin and zeaxan-thin biosynthesis could indicate that the crtS gene productis involved in both (ketolase and hydroxylase) enzymaticactivities involved in these late conversions, or thatthis strain has an additional enzyme(s) able to biosynthe-size the keto-derivatives HDCO and astaxanthin (seeSection 4).
3.6. Expression of crtS cDNA in M. circinelloides caused increased �-cryptoxanthin and zeaxanthin production
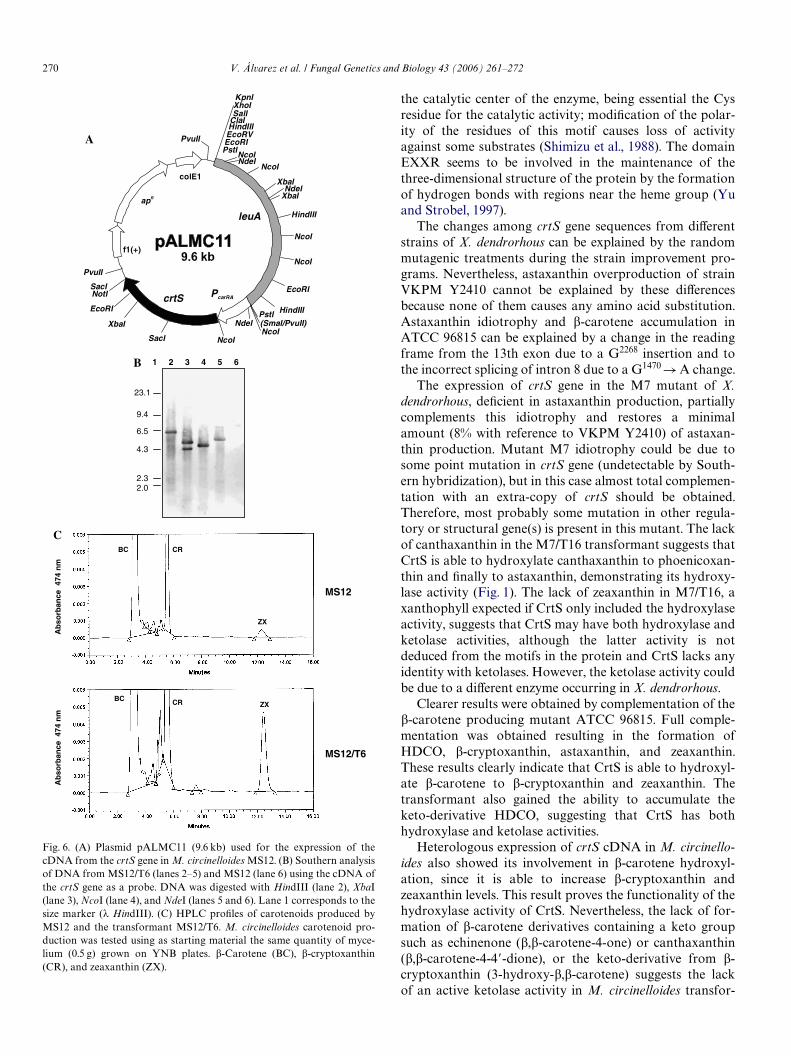
The crtS cDNA was heterologously expressed inM. circinelloides MS12 (leuA1, pyrG4) using the autono-mous replicating plasmid pALMC11 (Fig. 6A). This plas-mid includes the cDNA expressed under the control of thecarRA promoter of B. trispora (Rodríguez-Sáiz et al.,2004) and the leuA leucine encoding gene as an auxotro-phic marker. Several pALMC11 transformants prototro-phic for leucine were obtained and carotenoid productionwas analyzed by TLC and HPLC in six transformants(named MS12/T1 to MS12/T6) showing diVerent shadesof yellow, and in the parental MS12 strain. Southernanalysis of HindIII, XbaI, NcoI, and NdeI digestedDNA from MS12 and MS12/T6 showed hybridizationpatterns agreeing with pALMC11 replicating autono-mously in the transformant, indicating that this plasmidhad not been rearranged (Fig. 6B). Ampicillin-resistantcolonies were selected by transforming E. coli DH5�with total DNA from MS12/T6. Fifty independentclones were analyzed, and most of them (>98%) hadthe expected restriction pattern for pALMC11, conWrm-ing the extra-chromosomal location of the plasmid inM. circinelloides and an almost total lack of rearrange-ments.
The parental strain MS12 produced �-carotene asmain carotenoid, and also �-cryptoxanthin (3-hydroxy-�,�-carotene) and zeaxanthin (3,3�-dihydroxy-�,�-caro-tene). Transformants showed similar �-caroteneproduction (90–100%), and increased �-cryptoxanthin(140–160%) and zeaxanthin (190–330%) (Fig. 6C).These results indicate that the crtS gene product hasthe hydroxylase activity involved in the late conver-sions of the astaxanthin biosynthetic pathway (seeSection 4).
4. Discussion
The crtS gene cloned from the astaxanthin producingstrains X. dendrorhous VKPM Y2410 and ATCC 24203encodes a polypeptide similar to cytochrome-P450 hydrox-ylases, especially to those from fungi, but it showed insig-niWcant identity with the protein encoded by the crtZ geneinvolved in the hydroxylation of �-carotene in bacteria,algae, and plants. The presence of 17 introns has not beenreported in any other crtW or crtZ genes previouslydescribed, explaining the diYculty of the cloning processusing probes from bacterial, algae or plant genes in the dat-abases. Nevertheless, numerous introns have beendescribed in other genes from X. dendrorhous (Verdoeset al., 1999a,b). These results clearly indicate that the evolu-tive origin of the crtS gene is diVerent from that of the crtWand crtZ that catalyze similar reactions in other microor-ganisms. According to the phylogenetic tree, the CrtS cyto-chrome-P450 hydroxylase of X. dendrorhous is more closelyrelated to the homologous protein from fungi (Cr. neofor-mans and U. maydis) than to the corresponding enzymefrom plant (A. thaliana) and higher eukaryotes (Ovis ariesand Homo sapiens), and probably has evolved from anancestral P450 common to X. dendrorhous and other fungi.
The motifs described in the promoter region of the crtSgene involve regulatory functions. The pyrimidine-richstretches suggest a putative transcription start site, since itappears in highly expressed genes immediately upstream ofthis site (Davis and Hynes, 1991). Most of the fungal geneshave a TATA box located 30–50 bp upstream of the tran-scription start point (Davis and Hynes, 1991), which agreeswith the supposed functional TATA box located 352 bpupstream to the translation start point. The CAAT box ispresent in the promoter region of about 30% of knowneukaryotic genes located between 50 and 200 bp upstreamof the transcription start site (Davis and Hynes, 1991). Thepresence in the crtS promoter region of four consensusmotifs for CreA binding, a negative regulator mediatingcarbon catabolism (Cubero and Scazzocchio, 1994), indi-cates a potential carbon catabolite repression. In fact, whenX. dendrorhous is fermented in the presence of high concen-trations of glucose, the astaxanthin production ratedecreased remarkably (Yamane et al., 1997), being the car-bon/nitrogen ratio a critical factor for maximum astaxan-thin production (Vustin et al., 2004).
The structural features identiWed in CrtS (secondarystructures, oxygen-binding site, and heme pocket) andthose key amino acids involved in either stabilizing thepolypeptide conformation or providing ligands for hemeand substrate interactions are coincident with the reportedcrystal structure for the cytochrome P450cam from Pseudo-monas putida, a camphor 5-monooxygenase (Poulos et al.,1987). The oxygen-binding domain GXET is very con-served in cytochrome P450 from eukaryotes; oxygen bindsto the Gly residue obtaining two electrons from NAD(P)Hoxidation through the acidic residues E or D in the motif(Teutsch et al., 1993). The heme-binding domain represents
270 V. Álvarez et al. / Fungal Genetics and Biology 43 (2006) 261–272
Fig. 6. (A) Plasmid pALMC11 (9.6 kb) used for the expression of thecDNA from the crtS gene in M. circinelloides MS12. (B) Southern analysisof DNA from MS12/T6 (lanes 2–5) and MS12 (lane 6) using the cDNA ofthe crtS gene as a probe. DNA was digested with HindIII (lane 2), XbaI(lane 3), NcoI (lane 4), and NdeI (lanes 5 and 6). Lane 1 corresponds to thesize marker (� HindIII). (C) HPLC proWles of carotenoids produced byMS12 and the transformant MS12/T6. M. circinelloides carotenoid pro-duction was tested using as starting material the same quantity of myce-lium (0.5 g) grown on YNB plates. �-Carotene (BC), �-cryptoxanthin(CR), and zeaxanthin (ZX).
B
C
ZX
BC
ZXBC
CR
CR
Ab
sorb
ance
474
nm
A
bso
rban
ce 4
74 n
m
MS12
MS12/T6
1 2 3 4 5 6
23.1
9.4
6.5
4.3
2.3 2.0
the catalytic center of the enzyme, being essential the Cysresidue for the catalytic activity; modiWcation of the polar-ity of the residues of this motif causes loss of activityagainst some substrates (Shimizu et al., 1988). The domainEXXR seems to be involved in the maintenance of thethree-dimensional structure of the protein by the formationof hydrogen bonds with regions near the heme group (Yuand Strobel, 1997).
The changes among crtS gene sequences from diVerentstrains of X. dendrorhous can be explained by the randommutagenic treatments during the strain improvement pro-grams. Nevertheless, astaxanthin overproduction of strainVKPM Y2410 cannot be explained by these diVerencesbecause none of them causes any amino acid substitution.Astaxanthin idiotrophy and �-carotene accumulation inATCC 96815 can be explained by a change in the readingframe from the 13th exon due to a G2268 insertion and tothe incorrect splicing of intron 8 due to a G1470!A change.
The expression of crtS gene in the M7 mutant of X.dendrorhous, deWcient in astaxanthin production, partiallycomplements this idiotrophy and restores a minimalamount (8% with reference to VKPM Y2410) of astaxan-thin production. Mutant M7 idiotrophy could be due tosome point mutation in crtS gene (undetectable by South-ern hybridization), but in this case almost total complemen-tation with an extra-copy of crtS should be obtained.Therefore, most probably some mutation in other regula-tory or structural gene(s) is present in this mutant. The lackof canthaxanthin in the M7/T16 transformant suggests thatCrtS is able to hydroxylate canthaxanthin to phoenicoxan-thin and Wnally to astaxanthin, demonstrating its hydroxy-lase activity (Fig. 1). The lack of zeaxanthin in M7/T16, axanthophyll expected if CrtS only included the hydroxylaseactivity, suggests that CrtS may have both hydroxylase andketolase activities, although the latter activity is notdeduced from the motifs in the protein and CrtS lacks anyidentity with ketolases. However, the ketolase activity couldbe due to a diVerent enzyme occurring in X. dendrorhous.
Clearer results were obtained by complementation of the�-carotene producing mutant ATCC 96815. Full comple-mentation was obtained resulting in the formation ofHDCO, �-cryptoxanthin, astaxanthin, and zeaxanthin.These results clearly indicate that CrtS is able to hydroxyl-ate �-carotene to �-cryptoxanthin and zeaxanthin. Thetransformant also gained the ability to accumulate theketo-derivative HDCO, suggesting that CrtS has bothhydroxylase and ketolase activities.
Heterologous expression of crtS cDNA in M. circinello-ides also showed its involvement in �-carotene hydroxyl-ation, since it is able to increase �-cryptoxanthin andzeaxanthin levels. This result proves the functionality of thehydroxylase activity of CrtS. Nevertheless, the lack of for-mation of �-carotene derivatives containing a keto groupsuch as echinenone (�,�-carotene-4-one) or canthaxanthin(�,�-carotene-4-4�-dione), or the keto-derivative from �-cryptoxanthin (3-hydroxy-�,�-carotene) suggests the lackof an active ketolase activity in M. circinelloides transfor-

V. Álvarez et al. / Fungal Genetics and Biology 43 (2006) 261–272 271
mants. The heterologous expression of crtZ (hydroxylase)and crtW (ketolase) astaxanthin biosynthetic genes fromAg. aurantiacum in M. circinelloides under the control ofthe homologous glyceraldehyde-3-phosphate dehydroge-nase gene promoter has been recently described (Pappet al., 2006). In this case, crtZ is able to increase �-crypto-xanthin and zeaxanthin levels, whereas �-cryptoxanthin,echinenone, canthaxanthin, and a small amount of astaxan-thin are produced when crtW is expressed. These resultsagree with the presence of a �-carotene hydroxylase activityin the enzyme encoded by crtS and the lack (or lack offunctionality in M. circinelloides) of the �-carotene ketolaseactivity. Furthermore, the zeaxanthin found in MS12 sug-gests the existence of a �-carotene hydroxylase activity inM. circinelloides, an enzyme that remains to be character-ized.
At present, astaxanthin is produced commercially bychemical synthesis or biotechnologically with the alga H.pluvialis and the basidiomycete X. dendrorhous. The crtSgene described here can be used to engineer the astaxanthinbiosynthetic pathway of X. dendrorhous to develop newstrains with improved astaxanthin content or for the pro-duction of new xanthophylls.
Acknowledgments
This work was supported by a grant of the CICYT toboth Antibióticos S.A. (BIO2000-0157-P4-03) and J.F.M.(INBIOTEC; BIO2000-0157-P4-02). V.A. was supported bya fellowship, Acción MIT-F2, from the Spanish Ministry ofScience and Technology. E.J.G. and R.P.G. received fellow-ships from the FPU Programme of the Spanish Ministry ofEducation and Culture. The authors thank P. Romaine forpBGgHg, and P. Merino, M. Sandoval, P. Oricheta, C.Flórez, E.G. Quiñones, and M. Álvarez for technical assis-tance.
References
Altschul, S.F., Madden, T.L., SchäVer, A.A., Zhang, J., Zhang, Z., Miller,W., Lipman, D.J., 1997. Gapped BLAST and PSI-BLAST: a new gen-eration of protein database search programs. Nucleic Acids Res. 25,3389–3402.
An, G.H., Cho, M.H., Johnson, E.A., 1999. Monocyclic carotenoid biosyn-thetic pathway in the yeast PhaYa rhodozyma (Xanthophyllomycesdendrorhous). J. Biosci. Bioeng. 88, 189–193.
Andrewes, A.G., PhaV, H.J., Starr, M.P., 1976. Carotenoids of P. rhodo-zyma, a red pigmented fermenting yeast. Phytochemistry 15, 1003–1007.
Bauernfeind, J.C., 1981. Carotenoids as Colorants and Vitamin A Precur-sors: Technical and Nutritional Applications. Academic Press, NewYork.
Bhosale, P., 2004. Environmental and cultural stimulants in the productionof carotenoids from microorganisms. Appl. Microbiol. Biotechnol. 63,351–361.
Britton, G., Liaaen-Jensen, S., Pfander, H., 1998. Carotenoids. BirkhäuserVerlag, Basel.
Chen, X., Stone, M., Schlagnhaufer, C., Romaine, P., 2000. A fruiting bodytissue method for eYcient Agrobacterium-mediated transformation ofAgaricus bisporus. Appl. Environ. Microbiol. 66, 4510–4513.
Chew, B.P., Park, J.S., Wong, M.W., Wong, T.S., 1999. A comparison of theanticancer activities of dietary beta-carotene, canthaxanthin and asta-xanthin in mice in vivo. Anticancer Res. 19, 1849–1853.
Chun, S.B., Chin, J.E., Bai, S., An, G.H., 1992. Strain improvement ofPhaYa rhodozyma by protoplast fusion. FEMS Microbiol. Lett. 93,221–226.
Cifuentes, V., Hermosilla, G., Martínez, C., Leon, R., Pincheira, G., Jimé-nez, A., 1997. Genetics and electrophoretic karyotyping of wild-typeand astaxanthin mutant strains of PhaYa rhodozyma. Antonie VanLeeuwenhoek 72, 111–117.
Cubero, B., Scazzocchio, C., 1994. Two diVerent, adjacent and divergentzinc Wnger binding sites are necessary for CREA-mediated carboncatabolite repression in the proline gene cluster of Aspergillus nidulans.EMBO J. 13, 407–415.
Davis, M.A., Hynes, M.J., 1991. Regulatory circuits in Aspergillus nidulans.In: Bennett, J.W., Lasure, L.L. (Eds.), More Gene Manipulations inFungi. Academic Press, San Diego, CA, pp. 151–189.
de la Fuente, J.L., Peiro, E., Díez, B., Marcos, A.T., Schleissner, C., Rodrí-guez-Sáiz, M., Rodríguez, C., Cabri, W., Barredo, J.L., 2004. Methodfor producing astaxanthin by fermenting selected strains of Xantho-phyllomyces dendrorhous. European Patent Application EP 1479777.
Ducrey Sanpietro, L.M., Kula, M.R., 1998. Studies of astaxanthin biosyn-thesis in Xanthophyllomyces dendrorhous (PhaYa rhodozyma). EVectsof inhibitors and low temperature. Yeast 14, 1007–1016.
Girard, P., Falconnier, B., Bricout, J., Vladescu, B., 1994. �-Carotene pro-ducing mutants of PhaYa rhodozyma. Appl. Microbiol. Biotechnol. 41,183–191.
Golubev, W.I., 1995. Perfect state of Rhodomyces dendrorhous (PhaYa rho-dozyma). Yeast 11, 101–110.
Hoshino, T., Ojiva, K., Stoguchi, Y., 2000. Astaxanthin synthase. Euro-pean Patent Application EP 1035206.
Johnson, E.A., Conklin, D., Lewis, M.J., 1977. The yeast PhaYa rhodozymaas a dietary pigment source for salmonids and crustaceans. J. Fish.Beast. Board Can. 34, 2417–2421.
Johnson, E.A., Lewis, M.J., 1979. Astaxanthin formation in the yeastPhaYa rhodozyma. J. Gen. Microbiol. 115, 173–183.
Lampila, L.E., Wallen, S.E., Bullerman, L.B., 1985. A review of factorsaVecting biosynthesis of carotenoids by the order Mucorales. Myco-pathologia 90, 65–80.
Lazo, G.R., Stein, P.A., Ludwig, R.A., 1991. A DNA transformation-com-petent Arabidopsis genomic library in Agrobacterium. Biotechnology 9,963–967.
Lodato, P., Alcaino, J., Barahona, S., Retamales, P., Jiménez, A., Cifuentes,V., 2004. Study of the expression of carotenoid biosynthesis genes inwild-type and deregulated strains of Xanthophyllomyces dendrorhous(Ex.: PhaYa rhodozyma). Biol. Res. 37, 83–93.
Marshall, R.D., 1972. Glycoproteins. Annu. Rev. Biochem. 41, 673–702.Papp, T., Velayos, A., Bartok, T., Eslava, A.P., Vagvolgyi, C., Iturriaga,
E.A., 2006. Heterologous expression of astaxanthin biosynthesisgenes in Mucor circinelloides. Appl. Microbiol. Biotechnol. 69,526–531.
PhaV, H.J., Miller, M.W., Yoneyama, M., Soneda, M., 1972. A comparativestudy of the yeast Xorae associated with trees on the Japanese Islandsand on the west coast of North America. In: Terui, G. (Ed.), Fermenta-tion Technology Today. Society of Fermentation Technology, Osaka,Japan, pp. 759–774.
Poulos, T.L., Finzel, B.C., Howard, A.J., 1987. High-resolution crystalstructure of cytochrome P450cam. J. Mol. Biol. 195, 687–700.
Rodríguez-Sáiz, M., Paz, B., de La Fuente, J.L., López-Nieto, M.J., Cabri,W., Barredo, J.L., 2004. Blakeslea trispora genes for carotene biosyn-thesis. Appl. Environ. Microbiol. 70, 5589–5594.
Roncero, M.I.G., Jepsen, L.P., Strøman, P., van Heeswijck, R., 1989. Char-acterization of a leuA gene and an ARS element from Mucor circinello-ides. Gene 84, 335–343.
Ruiz-Hidalgo, M.J., Eslava, A.P., Álvarez, M.I., Benito, E.P., 1999. Heter-ologous expression of the Phycomyces blakesleeanus phytoene dehy-drogenase gene (carB) in Mucor circinelloides. Curr. Microbiol. 39,259–264.

272 V. Álvarez et al. / Fungal Genetics and Biology 43 (2006) 261–272
Sambrook, J., Fritsch, E.F., Maniatis, T., 1989. Molecular Cloning. A Lab-oratory Manual, second ed. Cold Spring Harbor Laboratory Press,Cold Spring Harbor, NY.
Shimizu, T., Hirano, K., Takahashi, M., Hatano, M., Fujii-Kuriyama, Y.,1988. Site-directed mutagenesis of rat liver cytochrome P-450d: axialligand and heme incorporation. Biochemistry 27, 4138–4141.
Specht, C.A., DiRusso, C.C., Novotny, C.P., Ullrich, R.C., 1982. A methodfor extracting high-molecular-weight deoxyribonucleic acid from fungi.Anal. Biochem. 119, 158–163.
Teutsch, H.G., Hasenfratz, M.P., Lesot, A., Stoltz, C., Garnier, J.M., Jel-tsch, J.M., Durst, F., Werck-Reichhart, D., 1993. Isolation andsequence of a cDNA encoding the Jerusalem artichoke cinnamate 4-hydroxylase, a major plant cytochrome P450 involved in the generalphenylpropanoid pathway. Proc. Natl. Acad. Sci. USA 90, 4102–4106.
Velayos, A., Eslava, A.P., Iturriaga, E.A., 2000. A bifunctional enzyme withlycopene cyclase and phytoene synthase activities is encoded by thecarRP gene of Mucor circinelloides. Eur. J. Biochem. 267, 5509–5519.
Verdoes, J.C., Misawa, N., van Ooyen, A.J.J., 1999a. Cloning and charac-terisation of the astaxanthin biosynthetic gene encoding phytoenedesaturase of Xanthophyllomyces dendrorhous. Biotechnol. Bioeng. 63,750–755.
Verdoes, J.C., Krubasik, K.P., Sandmann, G., van Ooyen, A.J., 1999b. Iso-lation and functional characterisation of a novel type of carotenoid
biosynthetic gene from Xanthophyllomyces dendrorhous. Mol. Gen.Genet. 262, 453–461.
Verdoes, J.C., Sandmann, G., Visser, H., Díaz, M., van Mossel, M., vanOoyen, A.J.J., 2003. Metabolic engineering of the carotenoid biosyn-thetic pathway in the yeast Xanthophyllomyces dendrorhous (PhaYarhodozyma). Appl. Environ. Microbiol. 69, 3728–3738.
Visser, H., van Ooyen, A.J.J., Verdoes, J.C., 2003. Metabolic engineering ofthe astaxanthin-biosynthetic pathway of Xanthophyllomyces dendror-hous. FEMS Yeast Res. 4, 221–231.
Vustin, M.M., Belykh, E.N., Kishilova, S.A., 2004. Relationship betweenastaxanthin production and the intensity of anabolic processes in theyeast PhaYa rhodozyma. Mikrobiologiia 73, 751–757.
Wang, X., Willen, R., Wadstrom, T., 2000. Astaxanthin-rich algal meal andvitamin C inhibit Helicobacter pylori infection in BALB/cA mice. Anti-microb. Agents Chemother. 44, 2452–2457.
Yamane, Y., Higashida, K., Nakashimada, Y., Kakizono, T., Nishio, N.,1997. InXuence of oxygen and glucose on primary metabolism andastaxanthin production by PhaYa rhodozyma in batch and fed-batchcultures: kinetic and stoichiometric analysis. Appl. Environ. Microbiol.63, 4471–4478.
Yu, X.C., Strobel, H.W., 1997. Hydroperoxide-mediated cytochrome P450-dependent 8-anilino-1-naphthalenesulfonic acid destruction, productformation and P450 modiWcation. Mol. Cell. Biochem. 167, 159–168.








