Embed Size (px)
Citation preview

The double life of Ceratobasidium: orchid mycorrhizal fungi and their potentialfor biocontrol of Rhizoctonia solani sheath blight of rice
Ana Teresa Mosquera-Espinosa1
Department of Agricultural Sciences, NationalUniversity of Colombia at Palmira
Research Group in Orchids, Ecology, and PlantSystematics. National University of Colombia atPalmira
Paul BaymanDepartment of Biology, University of Puerto Rico at RıoPiedras, PO Box 23360, San Juan, Puerto Rico 00931-3360
Gustavo A. PradoRice Pathology Program, CIAT-Colombia of Palmira
Arnulfo Gomez-CarabalıDepartment of Animal Sciences, National University ofColombia, Palmira
J. Tupac OteroDepartment of Biological Sciences, National Universityof Colombia, Palmira and Institute of EnvironmentalStudies, IDEA National University of Colombia atPalmira
Research Group in Orchids, Ecology and PlantSystematics. National University of Colombia atPalmira
Abstract: Ceratobasidium includes orchid mycorrhi-zal symbionts, plant pathogens and biocontrol agentsof soilborne plant pathogens. It is not known to whatextent members of the first guild also can participatein the others. Ceratobasidium spp. were isolated fromroots of Colombian orchids and identified by phylog-eny based on nrITS sequences. Phylogenetic group-ing of Ceratobasidium spp. isolates correspondedto orchid host substrate (epiphytic vs. terrestrial).Isolates were tested for virulence on rice and forbiocontrol of Rhizoctonia solani, causal agent ofsheath blight of rice. All Ceratobasidium spp. isolatescaused some signs of sheath blight but significantlyless than a pathogenic R. solani used as a positivecontrol. When Ceratobasidium spp. isolates wereinoculated on rice seedlings 3 d before R. solani,they significantly reduced disease expression com-pared to controls inoculated with R. solani alone. Theuse of Ceratobasidium spp. from orchids for biologicalcontrol is novel, and biodiverse countries such asColombia are promising places to look for newbiocontrol agents.
Key words: biological control, integrated pestmanagement, pathogenic fungi, soilborne pathogen
INTRODUCTION
The closely related genera Ceratobasidium andThanatephorus (Agaricomycotina, Cantharellales) in-clude fungi with a wide range of trophic strategies:plant pathogens, saprotrophs and mycorrhizal symbi-onts of orchids (Agrios 2002, Montalvo et al. 2006).The best known pathogens are Rhizoctonia solani (5
Thanatephorus cucumeris), which cause diverse diseas-es of crops. Among these, R. solani causes sheathblight of rice, considered the second most importantfungal disease of rice and the cause of large economiclosses worldwide (Correa et al. 2001, Chaudhary et al.2003).
Rhizoctonia also includes species with teleomorphsin Ceratobasidium, which have characteristic binucle-ate hyphal cells (often called ‘‘binucleate Rhizocto-nia’’ or BNR, as opposed to the multinucleate cellsof Thanatephorus). Some Ceratobasidium species aremycorrhizal symbionts of orchids, whose seeds re-quire an association with a fungus to obtain sufficientnutrition for germination (Arditti 1992, Otero et al.2002, Rasmussen 2002). Orchid mycorrhizae areunlike other types of mycorrhizae in that the fungusprobably receives little or nothing from the plant inmost cases (Rasmussen and Rasmussen 2009, Selosseand Roy 2009). It is not clear how these fungi get thecarbon they pass to orchids; they could be parasites,saprotrophs or mycorrhizal associates of other plants.Understanding nutritional strategies of orchid my-corrhizal fungi may lead to new strategies forconservation of species of Orchidaceae, the largestfamily of plants and one with many species that arerare and becoming rarer (Flanagan et al. 2006,Clements et al. 2011).
Other Ceratobasidium species are plant pathogens,and some have been used for biocontrol of soilborneplant pathogens (Burns and Benson 2000, Sneh et al.2004). Ceratobasidium spp. isolates from soil also havebeen used to control R. solani in crops such as cotton,soybean, vegetable seedlings and grasses (Burpee andGoulty 1984, Villajuan-Abgona et al. 1996, Jabaji-Hareand Neate 2005, Khan et al. 2005, Wen et al. 2005),but there are no reports of its use to control sheathblight of rice. Sheath blight of rice is controlledmainly with synthetic fungicides and resistant rice
Submitted 13 Mar 2012; accepted for publication 11 Jul 2012.1 Corresponding author. E-mail: [email protected]
Mycologia, 105(1), 2013, pp. 141–150. DOI: 10.3852/12-079# 2013 by The Mycological Society of America, Lawrence, KS 66044-8897
141

varieties (Correa et al. 2001). Therefore, to helpdevelop an integrated pest management (IPM)strategy for sheath blight of rice, we tested thebiocontrol ability of Ceratobasidium spp. isolates fromorchid roots.
Colombia is an ideal country in which to studythese issues; it has one of the richest orchid florasin the world (Arditti 1992, Dressler 1993, Calderon2006). However, its orchid mycorrhizal fungi arealmost entirely unexplored (Mosquera-Espinosa et al.2010). Also, in some areas of Colombia Rhizoctoniasheath blight is a serious problem for rice production,causing up to 40% yield loss in susceptible varieties(Correa et al. 2001).
In this study we combined these themes: aphylogenetic analysis of Ceratobasidium spp. isolatesfrom roots of orchids, assays of virulence of theseisolates on rice and their potential for biocontrol ofsheath blight. The objectives were: (i) IdentifyCeratobasidium spp. isolates from roots of orchids indifferent habitats in Colombia and use phylogeneticanalysis to determine whether habitat preferencesreflect phylogenetic relationships. (ii) Test theirvirulence on rice plants. (iii) Test the potential ofCeratobasidium spp. isolates from orchids for biocon-trol of R. solani on rice. We predicted that the leastvirulent isolates of Ceratobasidium spp. would bestprotect rice plants from R. solani, and that Ceratoba-sidium spp. isolates from terrestrial orchids wouldbetter protect rice plants from a soilborne pathogenthan isolates from epiphytic orchids. (iv) Determinethe effect of different doses of N and pasteurized vs.unpasteurized soil on virulence and biocontrol byCeratobasidium spp. High N was reported to increasethe incidence and severity of R. solani on rice (Savaryet al. 1995, Slaton et al. 2003, Cooke et al. 2006). Weexpected pasteurization of soil would decrease thebiocontrol efficiency of Ceratobasidium spp. becausebeneficial microorganisms might help in biocontrol(Alexander 1981, Giri et al. 2005).
MATERIALS AND METHODS
Phylogenetic analysis of Ceratobasidium spp. isolated fromColombian orchids.—Collection of orchids from two regionsof Colombia, isolation of mycorrhizal fungi and amplifica-tion and sequencing of the nuclear ribosomal ITS regionwere reported by Mosquera-Espinosa et al. (2010). Sectionsof root cortex with pelotons were surface-sterilized in 70%
EtOH, 2.5% NaClO and sterile distilled water 1 min eachand plated on potato dextrose agar (PDA) acidified with0.2% lactic acid to inhibit growth of bacteria. Fungalcolonies were isolated in pure culture on PDA and isolatesof form-genus Rhizoctonia were selected for sequencing onbasis of hyphal morphology (Sneh et al. 1991). DNA wasisolated by a miniprep method and the nuclear ribosomal
ITS was amplified and sequenced (Otero et al. 2002).Orchids sampled and fungi isolated are summarized(TABLE I).
ITS sequences were aligned with the multiple sequencealignment program Muscle (www.ebi.ac.uk/Tools/msa/muscle/) and alignments were adjusted manually in BioEdit5.0. The closest hits from BLAST queries in GenBank wereincluded in the dataset for phylogenetic analysis, along withrepresentative ITS sequences from several anastomosisgroups (AGs) of Ceratobasidium. Trees were rooted withsequences of two Tulasnella isolates from the orchidsSpathoglottis plicata (Singapore, AJ313458, Ma et al. 2003)and Vanilla poitaei (Puerto Rico, GenBank DQ834391,Porras-Alfaro and Bayman 2007). Tulasnella was chosen asan outgroup based on recent phylogenetic work on theCantharellales, which includes Ceratobasidium and Thana-tephorus (Moncalvo et al. 2006).
Phylogenetic relationships were estimated with maximumparsimony (MP) and maximum likelihood (ML) withheuristic searches in PAUP 4.0b10 (Swofford 2001). TheML tree was based on the HKY85 model of sequenceevolution as estimated by FindModel. Bootstrap support wascalculated for the MP tree using 1000 repetitions.
Virulence of Ceratobasidium spp. on rice plants.—Nine ofthe 12 Ceratobasidium spp. isolates from Colombian orchidswere tested for virulence on rice in vitro and in vivo. Diseaseincidence was tested on cut leaves (FAO 1985) and diseaseseverity on plants in the greenhouse (Liu et al. 2009).
Disease incidence on rice leaves in vitro.—Mature leaves from40 d old plants of Oryza sativa var. japonica cv. Koshihikari(Kitazawa Seed Co., Oakland, California) were harvested.Koshihikari is moderately susceptible to sheath blight(Cartwright and Lee 2001). Seven centimeter-long pieceswere placed on filter paper in Petri dishes, with four piecesper plate and 10 replicate plates per treatment. Each leafpiece was inoculated with one 3 mm agar plug taken from5 d old cultures on PDA. Plates were incubated at 25 C and100% humidity, suitable for development of sheath blight(Correa et al. 2001, Ferrero and Nguyen 2003). Percentincidence was recorded 4 d after inoculation based on howmany of the four leaf pieces developed signs of sheathblight (i.e. 0, 25, 50, 75 or 100%) (FAO 1985). Rhizoctoniasheath blight of rice first appears as irregular, elliptical, red-brown lesions. In time the center of the lesions turns white-gray, leaving a red-brown outer ring (Correa et al. 2001,Agrios 2002). Sixteen isolates were tested: nine Ceratobasi-dium spp. isolates described above, one Ceratobasidium sp.and one Tulasnella sp. isolated from orchid mycorrhizae inPuerto Rico, two R. solani potato pathogens (Rhs1AP andBS69, both AG-3, Charlton et al. 2008), one R. solani ricepathogen (AG1-1A) included as a positive control and twonegative controls (uninoculated leaf pieces and leaf piecesinoculated with PDA only). The experiment had acompletely randomized design. Data were analyzed byANOVA. The large coefficient of variation was addressedby transforming the data as !(X + 0.5) (Gomez and Gomez1984). A post-hoc comparison of means was done withDuncan’s multiple range test (P 5 0.05) using SAS (2002).
142 MYCOLOGIA
Disease severity on rice plants in the greenhouse.—Green-house tests followed a standardized methodology (Liu et al.2009): Twenty-one day-old plants of the variety Fedearroz50, one of the most widely planted in Colombia, were used(Jaramillo et al. 2003, Cuevas 2005). Fungi were inoculatedat the crown, simulating natural infection by the pathogenin the soil (Agrios 2002). Each plant was inoculated withone 3 mm diam plug taken from a 5 d old culture on PDA.Once inoculated, each pot was covered with a microcham-ber (made from a 2 L plastic soft drink bottle) to maintain100% relative humidity and 30 C, conditions required forinfection by the fungus (Agrios 2002, Liu et al. 2009). Tomaintain constant soil humidity, pots were partially sub-merged in trays of water. Ten days after inoculation diseaseseverity was scored as percent leaf with lesions for eachplant, following the methodology of Liu et al. (2009); the
proportion of leaf area with lesions was estimated for thelowest leaf affected, the two leaves above it, and the stem,and the three leaves and stem were weighted 30%, 25%,25% and 20% respectively. For severity assays the R. solaniAG-3 isolates from potato, Ceratobasidium spp. from PuertoRico and Tulasnella spp. were not included.
The experiment used five plants per pot, three pots pertreatment and a randomized block design in a factorialexperiment. A total of 72 combinations were tested: two soiltypes (pasteurized vs. unpasteurized) 3 three doses of N(described below) 3 12 treatments (inoculation with nineCeratobasidium spp. isolates and the three controls de-scribed above). The experiment was repeated 1 mo after thefirst run. Analyses by ANOVA combined data from the tworuns because no significant difference was observedbetween them. Because the coefficient of variation was
TABLE I. Isolates of Ceratobasidium spp. from orchids
Isolate GenBank accession Orchid Habitat Locality % similarity Highest BLAST hit
Q1M13 GU206536 Notylia sp. Epiphyte Valle del Cauca 93% CeratobasidiumAG-B(o),AB219143
Q1.1 M14 GU206537 Habenaria sp. 1 Terrestrial Valle del Cauca 94% Ceratobasidium YACuevas, A. 2005,DQ279043
Q1.2 M14 GU206538 Habenaria sp. 2 Terrestrial Valle del Cauca 96% Ceratobasidium YACuevas, A. 2005,DQ279043
Q2.1 M14 GU206539 Cranichis sp. 1 Terrestrial Valle del Cauca 100% CeratobasidiumAG-G, AB478783
100% Ceratobasidium,EU605732
Q2.2 M14 GU206540 Cranichis sp. 2 Terrestrial Valle del Cauca 100% Ceratobasidium,DQ279043
Q1M15 GU206541 Vanilla sp. Hemiepiphyte Valle del Cauca 99% Endophyte,FJ613838
Q1 M16 1.1 GU206542 Epidendrummelinanthum 1
Lithophyte Valle del Cauca 98% Thanatephoruscucumeris,AJ000202
97% CeratobasidiumAG-F, DQ102436
Q1 M16 1.2 GU206543 Epidendrummelinanthum 2
Lithophyte Valle del Cauca 100% UnculturedCeratobasidium,EU002954
Q3 M17 1.1 GU206544 Trizeuxis falcata 1 Epiphyte Antioquia 97% CeratobasidiumJTO091,AF472295
Q3 M17 1.2 GU206545 Trizeuxis falcata 2 Epiphyte Antioquia 97% CeratobasidiumJTO031,AF503970
Q1M19 GU206547 Maxillaria sp. Epiphyte Valle del Cauca 97% CeratobasidiumJTO031,AF503970
Q2M19 GU206546 Dichaea sp. Epiphyte Valle del Cauca 97% CeratobasidiumJTO075,AF472291
R. solani (Tolima2399-1 oryzica 1)
GU206548 Oryza sativa Pathogen CIAT, Colombia 100% T. cucumeris,FJ667267
MOSQUERA-ESPINOSA ET AL.: ORCHID MYCORRHIZAE AND DISEASE BIOCONTROL 143

high, data were transformed as !(X + 0.5) (Gomez andGomez 1984). Post-hoc tests to compare means usedDuncan’s multiple range test with significance consideredP 5 0.05, using the statistical package SAS (2002).
Biocontrol of R. solani by Ceratobasidium spp. on rice plantsin greenhouse conditions.—Experimental design and mea-surement of disease severity were as described for thevirulence experiments described above. However, the timeof inoculation was modified as follows: Ceratobasidium spp.isolates were inoculated 2 d before the virulent isolate of R.solani. During these days the plants were incubated in a60 m2 chamber, with plastic curtains and three humidifiersthat ran 1 h twice a day to maintain an environmentconducive to development of disease. After inoculation,plants were incubated in microchambers as describedabove. The experiment was repeated 1 mo after the firstrun.
Effects of soil N and pasteurization.—In the greenhouseexperiments described above, the effect of N and ofpasteurized vs. unpasteurized soil on virulence of Ceratoba-sidium spp. and biocontrol of R. solani was tested. N wasapplied in three doses: 0, 150 and 250 kg N /ha, in the formof ammonium sulfate (26% N) as reported Savary et al.(1995), Slaton et al. (2003), Cooke et al. (2006). Soil wascollected from a rice field in Jamundı, Valle del Cauca,Colombia, to ensure that its physicochemical propertieswere suitable for rice production (IGAC 2006).
RESULTS
Phylogeny of Ceratobasidium spp. isolates fromorchid roots.—All isolates from orchids were assignedto Ceratobasidium (in one case, Thanatephorus) basedon BLAST results and the binucleate condition ofyoung hyphae (in one case uninucleate: Q1M13 fromNotylia sp., TABLE I) (Mosquera-Espinosa et al. 2010);phylogenetic analysis agreed with these assignments.Identification to species was not possible because theisolates were divergent from named species andbecause species-level taxonomy of Ceratobasidium ispoorly resolved. The isolate from a rice plant withsheath blight (used here as a positive control) had100% ITS homology with R. solani AG1-1A, the AG towhich the sheath blight pathogen belongs (Sneh et al.1991, Agrios 2002, Bernardes de Assis et al. 2008).
Phylogenetic analyses grouped the isolates intothree clades, labeled ‘‘epiphytic’’, ‘‘Thanatephorus’’and ‘‘terrestrial’’ (FIG. 1), which corresponded to thehabitats of the host orchids, but also to some extentwith host phylogeny. Isolates from the epiphyticorchids Dichaea, Maxillaria, Notylia and Trizeuxis(all in subfamily Epidendroideae) formed a cladewith 73% bootstrap support. This epiphytic clade alsoincluded reference sequences from CeratobasidiumAG-Q and mycorrhizal fungi of epiphytic orchids inPuerto Rico (clades A, B and C, Otero et al. 2002;
Otero et al. 2007), as well as from an epiphytic orchidfrom Singapore (AJ318430) and an isolate fromstrawberry (Japan, AB219143, pathogenicity not stated).
The ‘‘Thanatephorus’’ clade, with 69% bootstrapsupport, included a Ceratobasidium spp. isolate fromthe epiphytic orchid Epidendrum melinanthum 1 (alsoin subfamily Epidendroideae) and a reference se-quence from an orchid from Puerto Rico (JTO048).It also included reference sequences from patho-gens listed as Rhizoctonia, Thanatephorus and Cerato-basidium with a global distribution: Thanatephorus
FIG. 1. Phylogeny of Ceratobasidium isolates fromorchids and related fungi based on ITS sequences. Themaximum likelihood tree was based on a 580 bp alignmentwith 195 informative characters. Two ITS sequences fromTulasnella were defined as the outgroup. Dotted lines markclades discussed in the text. Numbers at the nodes showbootstrap values from the corresponding maximum parsi-mony (MP) tree. The asterisk marks the only branch notshared with the MP tree. 2Ln likelihood 5 3773.74826. Thecorresponding maximum parsimony analysis generated32 equally parsimonious trees, 606 steps long, CI 5 0.650,RI 5 0.788.
144 MYCOLOGIA
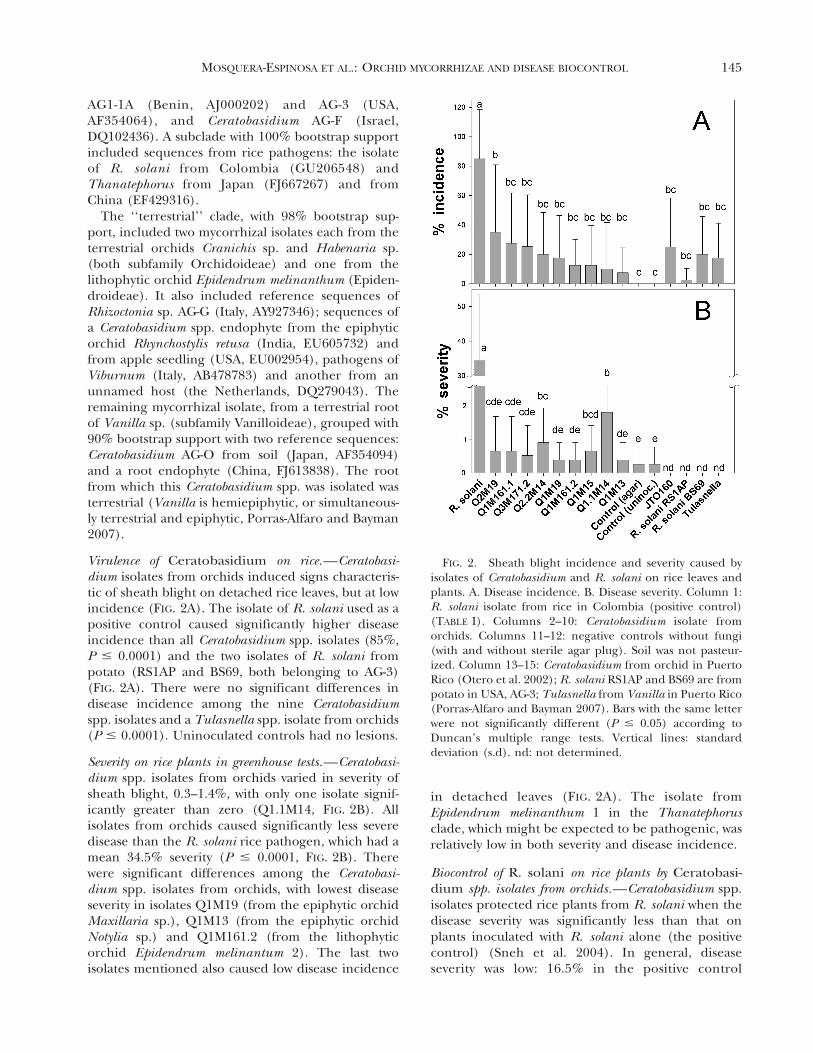
AG1-1A (Benin, AJ000202) and AG-3 (USA,AF354064), and Ceratobasidium AG-F (Israel,DQ102436). A subclade with 100% bootstrap supportincluded sequences from rice pathogens: the isolateof R. solani from Colombia (GU206548) andThanatephorus from Japan (FJ667267) and fromChina (EF429316).
The ‘‘terrestrial’’ clade, with 98% bootstrap sup-port, included two mycorrhizal isolates each from theterrestrial orchids Cranichis sp. and Habenaria sp.(both subfamily Orchidoideae) and one from thelithophytic orchid Epidendrum melinanthum (Epiden-droideae). It also included reference sequences ofRhizoctonia sp. AG-G (Italy, AY927346); sequences ofa Ceratobasidium spp. endophyte from the epiphyticorchid Rhynchostylis retusa (India, EU605732) andfrom apple seedling (USA, EU002954), pathogens ofViburnum (Italy, AB478783) and another from anunnamed host (the Netherlands, DQ279043). Theremaining mycorrhizal isolate, from a terrestrial rootof Vanilla sp. (subfamily Vanilloideae), grouped with90% bootstrap support with two reference sequences:Ceratobasidium AG-O from soil (Japan, AF354094)and a root endophyte (China, FJ613838). The rootfrom which this Ceratobasidium spp. was isolated wasterrestrial (Vanilla is hemiepiphytic, or simultaneous-ly terrestrial and epiphytic, Porras-Alfaro and Bayman2007).
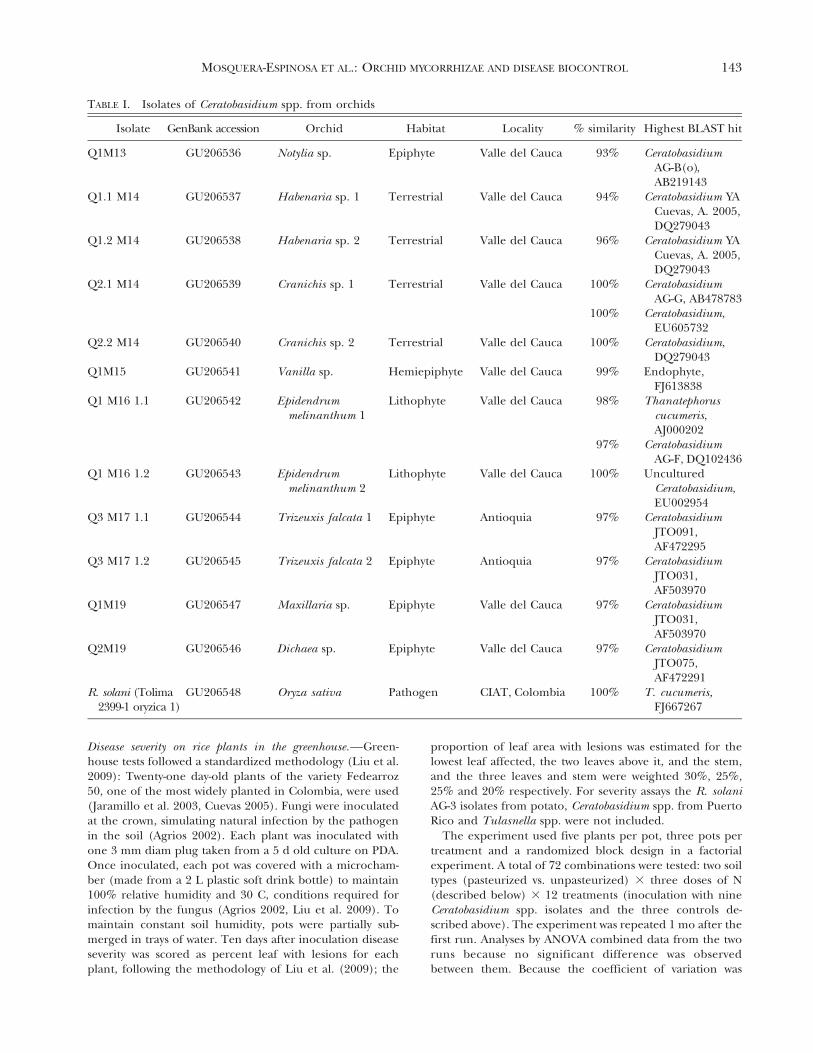
Virulence of Ceratobasidium on rice.—Ceratobasi-dium isolates from orchids induced signs characteris-tic of sheath blight on detached rice leaves, but at lowincidence (FIG. 2A). The isolate of R. solani used as apositive control caused significantly higher diseaseincidence than all Ceratobasidium spp. isolates (85%,P # 0.0001) and the two isolates of R. solani frompotato (RS1AP and BS69, both belonging to AG-3)(FIG. 2A). There were no significant differences indisease incidence among the nine Ceratobasidiumspp. isolates and a Tulasnella spp. isolate from orchids(P # 0.0001). Uninoculated controls had no lesions.
Severity on rice plants in greenhouse tests.—Ceratobasi-dium spp. isolates from orchids varied in severity ofsheath blight, 0.3–1.4%, with only one isolate signif-icantly greater than zero (Q1.1M14, FIG. 2B). Allisolates from orchids caused significantly less severedisease than the R. solani rice pathogen, which had amean 34.5% severity (P # 0.0001, FIG. 2B). Therewere significant differences among the Ceratobasi-dium spp. isolates from orchids, with lowest diseaseseverity in isolates Q1M19 (from the epiphytic orchidMaxillaria sp.), Q1M13 (from the epiphytic orchidNotylia sp.) and Q1M161.2 (from the lithophyticorchid Epidendrum melinantum 2). The last twoisolates mentioned also caused low disease incidence
in detached leaves (FIG. 2A). The isolate fromEpidendrum melinanthum 1 in the Thanatephorusclade, which might be expected to be pathogenic, wasrelatively low in both severity and disease incidence.
Biocontrol of R. solani on rice plants by Ceratobasi-dium spp. isolates from orchids.—Ceratobasidium spp.isolates protected rice plants from R. solani when thedisease severity was significantly less than that onplants inoculated with R. solani alone (the positivecontrol) (Sneh et al. 2004). In general, diseaseseverity was low: 16.5% in the positive control
FIG. 2. Sheath blight incidence and severity caused byisolates of Ceratobasidium and R. solani on rice leaves andplants. A. Disease incidence. B. Disease severity. Column 1:R. solani isolate from rice in Colombia (positive control)(TABLE I). Columns 2–10: Ceratobasidium isolate fromorchids. Columns 11–12: negative controls without fungi(with and without sterile agar plug). Soil was not pasteur-ized. Column 13–15: Ceratobasidium from orchid in PuertoRico (Otero et al. 2002); R. solani RS1AP and BS69 are frompotato in USA, AG-3; Tulasnella from Vanilla in Puerto Rico(Porras-Alfaro and Bayman 2007). Bars with the same letterwere not significantly different (P # 0.05) according toDuncan’s multiple range tests. Vertical lines: standarddeviation (s.d). nd: not determined.
MOSQUERA-ESPINOSA ET AL.: ORCHID MYCORRHIZAE AND DISEASE BIOCONTROL 145
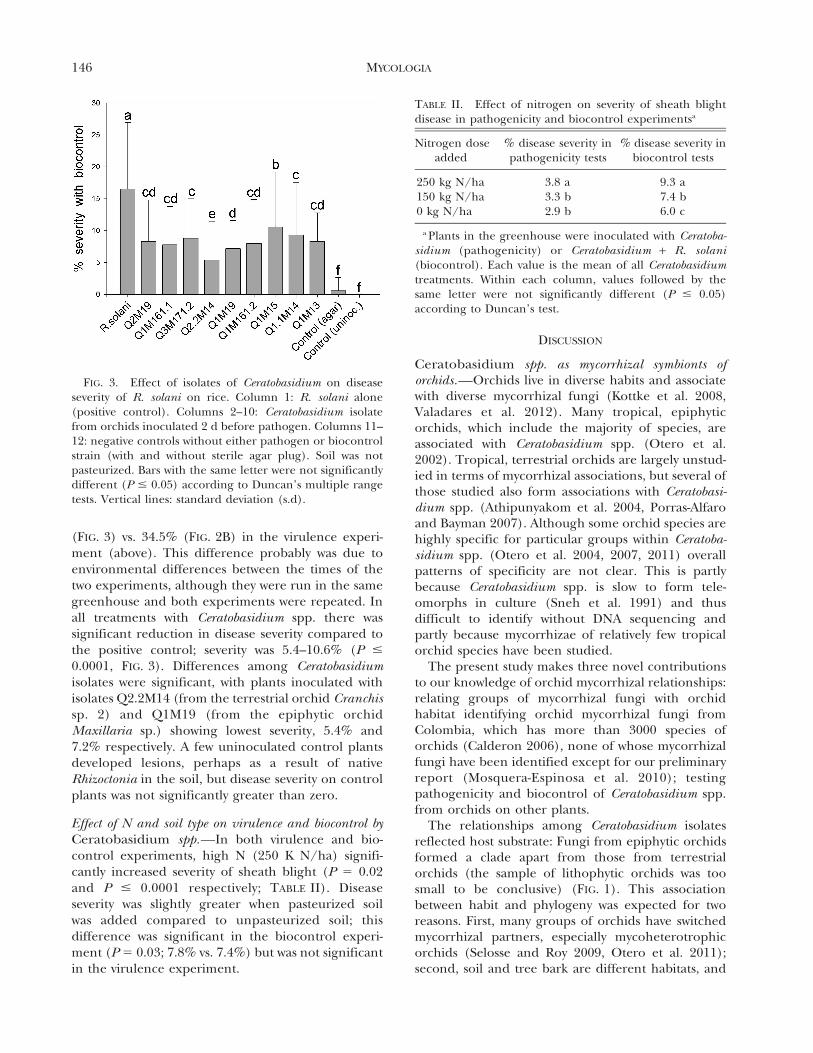
(FIG. 3) vs. 34.5% (FIG. 2B) in the virulence experi-ment (above). This difference probably was due toenvironmental differences between the times of thetwo experiments, although they were run in the samegreenhouse and both experiments were repeated. Inall treatments with Ceratobasidium spp. there wassignificant reduction in disease severity compared tothe positive control; severity was 5.4–10.6% (P #
0.0001, FIG. 3). Differences among Ceratobasidiumisolates were significant, with plants inoculated withisolates Q2.2M14 (from the terrestrial orchid Cranchissp. 2) and Q1M19 (from the epiphytic orchidMaxillaria sp.) showing lowest severity, 5.4% and7.2% respectively. A few uninoculated control plantsdeveloped lesions, perhaps as a result of nativeRhizoctonia in the soil, but disease severity on controlplants was not significantly greater than zero.
Effect of N and soil type on virulence and biocontrol byCeratobasidium spp.—In both virulence and bio-control experiments, high N (250 K N/ha) signifi-cantly increased severity of sheath blight (P 5 0.02and P # 0.0001 respectively; TABLE II). Diseaseseverity was slightly greater when pasteurized soilwas added compared to unpasteurized soil; thisdifference was significant in the biocontrol experi-ment (P 5 0.03; 7.8% vs. 7.4%) but was not significantin the virulence experiment.
DISCUSSION
Ceratobasidium spp. as mycorrhizal symbionts oforchids.—Orchids live in diverse habits and associatewith diverse mycorrhizal fungi (Kottke et al. 2008,Valadares et al. 2012). Many tropical, epiphyticorchids, which include the majority of species, areassociated with Ceratobasidium spp. (Otero et al.2002). Tropical, terrestrial orchids are largely unstud-ied in terms of mycorrhizal associations, but several ofthose studied also form associations with Ceratobasi-dium spp. (Athipunyakom et al. 2004, Porras-Alfaroand Bayman 2007). Although some orchid species arehighly specific for particular groups within Ceratoba-sidium spp. (Otero et al. 2004, 2007, 2011) overallpatterns of specificity are not clear. This is partlybecause Ceratobasidium spp. is slow to form tele-omorphs in culture (Sneh et al. 1991) and thusdifficult to identify without DNA sequencing andpartly because mycorrhizae of relatively few tropicalorchid species have been studied.
The present study makes three novel contributionsto our knowledge of orchid mycorrhizal relationships:relating groups of mycorrhizal fungi with orchidhabitat identifying orchid mycorrhizal fungi fromColombia, which has more than 3000 species oforchids (Calderon 2006), none of whose mycorrhizalfungi have been identified except for our preliminaryreport (Mosquera-Espinosa et al. 2010); testingpathogenicity and biocontrol of Ceratobasidium spp.from orchids on other plants.
The relationships among Ceratobasidium isolatesreflected host substrate: Fungi from epiphytic orchidsformed a clade apart from those from terrestrialorchids (the sample of lithophytic orchids was toosmall to be conclusive) (FIG. 1). This associationbetween habit and phylogeny was expected for tworeasons. First, many groups of orchids have switchedmycorrhizal partners, especially mycoheterotrophicorchids (Selosse and Roy 2009, Otero et al. 2011);second, soil and tree bark are different habitats, and
FIG. 3. Effect of isolates of Ceratobasidium on diseaseseverity of R. solani on rice. Column 1: R. solani alone(positive control). Columns 2–10: Ceratobasidium isolatefrom orchids inoculated 2 d before pathogen. Columns 11–12: negative controls without either pathogen or biocontrolstrain (with and without sterile agar plug). Soil was notpasteurized. Bars with the same letter were not significantlydifferent (P # 0.05) according to Duncan’s multiple rangetests. Vertical lines: standard deviation (s.d).
TABLE II. Effect of nitrogen on severity of sheath blightdisease in pathogenicity and biocontrol experimentsa
Nitrogen doseadded
% disease severity inpathogenicity tests
% disease severity inbiocontrol tests
250 kg N/ha 3.8 a 9.3 a150 kg N/ha 3.3 b 7.4 b0 kg N/ha 2.9 b 6.0 c
a Plants in the greenhouse were inoculated with Ceratoba-sidium (pathogenicity) or Ceratobasidium + R. solani(biocontrol). Each value is the mean of all Ceratobasidiumtreatments. Within each column, values followed by thesame letter were not significantly different (P # 0.05)according to Duncan’s test.
146 MYCOLOGIA

different groups of fungi are likely to dominate. Forexample, fungi in roots of Vanilla differed significantlybetween substrates, with Ceratobasidium spp. morecommon in roots in soil and Tulasnella spp. morecommon in roots on tree bark (Porras-Alfaro andBayman 2007). However, the phylogenetic relation-ships among Ceratobasidium spp. isolates also reflectedhost phylogeny, at least at subfamily rank (FIG. 1). Thiswas due to the association between orchid habitat andsystematics; all the terrestrial orchids sampled be-longed to the relatively primitive subfamily Orchidoi-deae, whereas the epiphytic orchids belonged to thederived subfamily Epidendroideae.
Thanatephorus formed a clade within Ceratobasi-dium (FIG. 1) implying that Ceratobasidium as cur-rently defined is paraphyletic, as has been reported inGonzalez et al. (2001), Moncalvo et al. (2006), Porras-Alfaro and Bayman (2007). This clade was composedmostly of pathogens, including a rice pathogen fromColombia but also included mycorrhizal fungi oforchids in Colombia and Puerto Rico.
The ability of Ceratobasidium spp. isolates tostimulate germination and growth of orchid seedswas predicted by phylogenetic relationships, suggest-ing a genetic basis for this behavior (Otero et al. 2004,2005). We therefore expected that closely relatedisolates would be very similar in disease severity andbiocontrol ability on rice, but there was no evidenceof this (FIG. 2). The fact that most of the isolatesshowed pathogenicity on rice (FIG. 2), althoughslight, suggests that they might be at least partlyparasitic in nutritional mode. Although a number ofCeratobasidium species are plant pathogens, thenutrition of orchid mycorrhizal species is unknown.Understanding their nutrition might be useful inconservation of the orchid species which depend onthem for seed germination (Otero et al. 2007).
The Ceratobasidium spp. isolates described herecame from the cortical cells of orchid roots withvisible pelotons (Mosquera-Espinosa et al. 2010),which suggests that the fungi had a functionalmycorrhizal relationship with their orchid hosts(Hadley and Williamson 1972, Suarez et al. 2006).However, not all fungi in orchid roots are mycorrhizaland we did not test functional aspects of theassociation. Symbiotic seed germination experimentsdemonstrated a functional mycorrhizal relationshipwith fungi closely related to the ‘‘epiphytic’’ clade(FIG. 1) (Otero et al. 2004, 2005).
Ceratobasidium species as plant pathogens andbiocontrol agents.—In this study Ceratobasidium spp.isolated from orchids caused lower incidence andseverity of disease on rice than R. solani AG1-1Aisolated from rice (FIG. 2). Studies have shown that
some Rhizoctonia AGs are more virulent to the hostfrom which they were isolated than on other hosts(Bolkan and Ribeiro 1985, Bernardes de Assis et al.2008). Some authors have recommended that viru-lence tests should score disease from the lowest levelto the highest to identify hypovirulent Ceratobasidiumspp. isolates with biocontrol potential against soil-borne pathogens (Sneh et al. 2004), as we do here.
Nonpathogenic Ceratobasidium spp. have beenisolated from soil and some show biocontrol activityagainst soilborne plant pathogens. They have beenused against R. solani causing damping-off in a varietyof crops: beans (Xue et al. 1998, Wen et al. 2005),corn (Pascual et al. 2000), soybeans (Khan et al.2005), cotton and radish (Ichielevich-Auster et al.1985, Sneh et al. 2004, Jabaji-Hare and Neate, 2005),cucumber (Villajuan-Abgona et al. 1996) and grasses(Burpee and Goulty 1984). The relationship of thesegroups to the Ceratobasidium species that formmycorrhizae with orchids is not clear, but in somecases they are closely related (FIG. 1; Otero et al. 2002,2004, 2007; Porras-Alfaro and Bayman 2007). Biocon-trol is sometimes more effective when the biocontrolagent is related to the pathogen, because theirrequirements are more likely to be similar.
However, as with any potential biocontrol agent,the potential pathogenicity of these isolates on thehost plant must be considered carefully. Someanastomosis groups (AGs) of Thanatephorus andCeratobasidium are pathogenic on a narrow range ofhosts (e.g. AG-3 on potato) whereas others attack arange of hosts (e.g. AG4 in different crops) (Bolkanand Ribeiro 1985, Agrios 2002). Although all theCeratobasidium spp. isolates tested were significantlyless virulent on rice than an isolate of R. solani thatgrouped with AG1-1A, they were still capable ofcausing lesions typical of sheath blight (FIG. 4). Manybiocontrol agents have a certain level of virulence tothe host (Burns and Benson 2000). Low infectiondoes not necessarily imply a decrease in yield, and insome cases may increase yield and plant success,mainly by the production of plant growth-promotingsubstances (Sneh 1998, Sneh et al. 2004).
Some Ceratobasidium spp. isolates produced abun-dant mycelium on rice plant tissues and surroundingsoil (FIG. 5A, B), as well as sclerotia (FIG. 5C);extensive colonization is usually considered a favor-able sign for biocontrol. Studies have not describedthis colonization in BNR used for biocontrol; on thecontrary, they have been described as slow-growingcompared to soilborne pathogens (Sneh et al. 2004).
Relative efficacy of Ceratobasidium spp. isolates.—Differences in biocontrol among Ceratobasidium spp.isolates were significant in some cases (FIG. 3). It is
MOSQUERA-ESPINOSA ET AL.: ORCHID MYCORRHIZAE AND DISEASE BIOCONTROL 147

interesting that those most effective at biocontrolwere not those that were least virulent (FIGS. 2, 3).The most effective biocontrol occurred with aCeratobasidium spp. isolate from the terrestrial orchidCranichis sp. 2 (Q2.2M14) (FIG. 3). This isolate hadlow disease severity in pathogenicity tests, althoughnot as low as some other isolates (FIG. 2). Thecombination of high protection in the biocontrolexperiment and low severity in the pathogenicityexperiments suggests that this isolate is the mostpromising candidate for biocontrol of rice sheathblight. A further advantage of Q2.2M14 is that it is afungus isolated from roots of a terrestrial orchid andis more likely to survive and provide effectivebiocontrol in the soil environment than an isolatefrom an epiphytic orchid adapted to live on tree bark.
Effect of doses of N and soil on virulence and biocontrolby Ceratobasidium spp.—The relationship betweenhigh N and increased disease severity agreed with Savaryet al. (1995), Slaton et al. (2003), Cooke et al. (2006).Increased severity in pasteurized soil also was expected,because pasteurization (as well as killing pathogens)destroys beneficial microorganisms that compete withpathogens (Alexander 1981, Giri et al. 2005). Chemicalanalysis showed that K and Zn occurred at low levels (K5 0.34 cmol/kg, Zn 5 3.6 mg/kg), which could havehad a negative effect on the response of plants toinfection by R. solani. These two elements affect rice’sresistance to soilborne pathogens (Guerrero 1991,Marschner 1995). However, disease severity in thisstudy never reached 40%, the threshold at which sheath
blight begins to cause economically significant losses inrice (Correa et al. 2001, Liu et al. 2009).
These results suggest that where rice is fertilized with Nsheath blight is more likely to cause a significantreduction in yield. N affects physiological processesinvolved in rice yield (leaf growth, photosynthesis,tillering etc., Jaramillo et al. 2003). Growers who applyN fertilizers to increase yield may not realize that they arepredisposing the plant to attack by fungal pathogenssuch as R. solani (Fedearroz 2000, Slaton et al. 2003,Cooke et al. 2006). Rice growers who fertilize aretherefore those most in need of effective biocontrolstrategies. Inducing suppressive soils by inoculation withbiocontrol organisms is an established strategy forcontrol of pathogens (Sneh 1998). Such a strategy couldbe used with Ceratobasidium to protect rice fields fromattack by R. solani, minimizing its adverse effects on yield.
This study provides evidence that Ceratobasidiumisolates from orchids may be effective at biocontrolof Rhizoctonia sheath blight of rice and have lowvirulence on rice (under greenhouse conditions) withpotential for application as part of an IPM program.However, pathogenicity on other hosts, particularlyon crop plants that are grown with (or in rotationwith) rice, and mode of action should be testedbefore field trials can be considered.
ACKNOWLEDGMENTS
We thank the Rice Pathology Group at CIAT, Palmira,Colombia, for assistance, the NSF-sponsored CREST-CATEC program at UPR Rıo Piedras (HRD 0734826) forsupport, and Marc Cubeta for the R. solani AG-3 isolates.The first author was supported by a fellowship fromCOLCIENCIAS Doctorados Nacionales 2006. The authorsdeclare no conflicts of interest.
LITERATURE CITED
Agrios GN. 2002. Fitopatologıa. 2nd ed. Mexico, DF:Editorial Limusa S.A. 830 p.
FIG. 5. Growth and colonization by Ceratobasidiumisolates in biocontrol experiments on rice. A. growth onthe surface of the plant (black arrows); B. on soil surfacearound the plant (black and white arrows); C. production ofsclerotia on soil (red arrows). Bars 5 1 cm.
FIG. 4. Differences in severity of sheath blight on rice plantsinoculated with: A. R. solani isolate from rice in Colombia(positive control) (TABLE I) (35%); B. Ceratobasidium Q1M161.2, from the orchid Epidendrum melinanthum 2 (1%); C.Ceratobasidium Q1M13, from the orchid Notylia sp. (2%).
148 MYCOLOGIA

Alexander M. 1981. Introduccion a la microbiologıa del suelo.1st ed. Mexico, DF: AGT Editor S.A. 491 p.
Arditti J. 1992. Fundamentals of orchid biology. New York:John Wiley & Sons. 483 p.
Athipunyakom P, Manoch L, Pileuk C. 2004. Isolation andidentification of mycorrhizal fungi from 11 terrestrialorchids. Kasetsart J Nat Sci 36:216–228.
Bernardes de Assis J, Peyer P, Rush MC, Zala M, McDonaldBA, Ceresini P. 2008. Divergence between sympatricrice and soybean-infecting populations of Rhizoctoniasolani anastomosis group-1 IA. Phytopathology 98:1326–1333, doi:10.1094/PHYTO-98-12-1326
Bolkan HA, Ribeiro WRC. 1985. Anastomosis groups andpathogenicity of Rhizoctonia solani isolates from Brazil.Plant Dis 69:599–601, doi:10.1094/PD-69-599
Burns J, Benson M. 2000. Biocontrol of damping-off ofCatharanthus roseus caused by Pythium ultimum withTrichoderma virens and binucleate Rhizoctonia fungi.Plant Dis 84:644–648, doi:10.1094/PDIS.2000.84.6.644
Burpee L, Goulty G. 1984. Suppression of brown patchdisease of creeping bentgrass by isolates of nonpatho-genic Rhizoctonia spp. Phythopathology 74:692–694,doi:10.1094/Phyto-74-692
Calderon Saenz E. 2006. Libro rojo de plantas de Colombia.Vol. 6. Orquıdeas, primera parte. Bogota: Instituto vonHumboldt. 828 p.
Cartwright R, Lee F. 2001. Management of rice diseases. In:Slaton NA, ed. Rice production handbook. Misc. Publ.192. Little Rock, Arkansas Coop. Ext. Serv. p 87–100.
Clements MA, Otero JT, Miller JT. 2011. Phylogeneticrelationships in Pterostylidinae (Cranichideae: Orchi-daceae): combined evidence from nuclear and plastidDNA sequences. Aust J Bot 59:99–117, doi:10.1071/BT10190
Cooke B, Gareth D, Kaye B. 2006. The epidemiology ofplant diseases, 2nd ed. The Netherlands: Springer.576 p.
Correa F, Meneses R, Gutierrez A, Garcıa A, Antigua G,Gomez J, Calvert L. 2001. Guıa para el trabajo decampo en el manejo integrado de plagas del arroz. Cali,Colombia: CIAT, IIA, FLAR. 72 p.
Cuevas A. 2005. Cincuenta razones para sembrar Fedearroz50. Fedearroz 53:32–39.
Charlton ND, Carbone I, Tavantzis SM, Cubeta MA. 2008.Phylogenetic relatedness of the M2 double-strandedRNA in Rhizoctonia fungi. Mycologia 100:555–564,doi:10.3852/07-108R
Chaudhary RC, Nanda JS, Tran DV. 2003. Guıa paraidentificar las limitaciones de campo en la produccionde arroz. Published online at: ftp://ftp.fao.org/docrep/fao/006/y2778s/y2778s00.pdf. [last accessed20 Jun 2012].
Dressler RL. 1993. Phylogeny and classification of theorchid family. London: Cambridge Univ. Press. 313 p.
FAO. 1985. Manual para patologos vegetales. 2nd ed. Chile:FAO. 438 p.
FEDEARROZ. 2000. Manejo y conservacion de suelos para laproduccion de arroz en Colombia. Bogota: Fedearoz. 78 p.
Ferrero A, Nguyen N. 2003. The sustainable development ofrice-based production systems in the world. FAO, Italy.
Published on line at: http://www.fao.org/docrep/008/y5682e/y5682e0g.htm. [last accessed 20 Jun 2012].
Flanagan NS, Peakall R, Clements MA, Otero JT. 2006.Conservation of difficult taxonomic groups: the case ofthe Australian orchid, Microtis angusii. Conserv Genet7:847–859, doi:10.1007/s10592-006-9119-8
Giri B, Giang PH, Kumari R, Prasad R, Varma A. 2005.Microorganisms in soils: roles in genesis and functions.In: Buscot F, Varma A, eds. Soil biology. Vol. 3. Berlin:Springer-Verlag. 419 p.
Gomez K, Gomez A. 1984. Statistical procedures foragricultural research. 2nd ed. New York: John Wiley& Sons. 608 p.
Gonzalez D, Carling DE, Kuninaga S, Vilgalys R, Cubeta MA.2001. Ribosomal DNA systematics of Ceratobasidiumand Thanatephorus with Rhizoctonia anamorphs. Myco-logia 93:1138–1150, doi:10.2307/3761674
Guerrero R. 1991. Fertilizacion de cultivos en clima calido.2nd ed. Bogota: Monomeros Colombo-Venezolanos.312 p.
Hadley G, Williamson B. 1972. Features of mycorrhizalinfection in some Malayan orchids. New Phytol 71:1111–1118, doi:10.1111/j.1469-8137.1972.tb01989.x
Ichielevich-Auster M, Sneh B, Koltin Y, Barash I. 1985.Suppression of damping-off caused by Rhizoctonia speciesby a nonpathogenic isolate of R. solani. Phytopathology75:1080–1084, doi:10.1094/Phyto-75-1080
Jabaji-Hare S, Neate SM. 2005. Nonpathogenic binucleateRhizoctonia spp. and benzothiadiazole protect cottonseedlings against Rhizoctonia damping-off and Alter-naria leaf spot in cotton. Phytopathology 95:1030–1036, doi:10.1094/PHYTO-95-1030
Jaramillo S, Pulver E, Duque MC. 2003. Efecto del manejode la fertilizacion nitrogenada en arroz de riego, sobrela expresion del potencial de rendimiento de lıneaselites y cultivares comerciales. Palmira, Colombia:Fondo Latinoamericano de Arroz de Riego (FLAR) yCentro Internacional de Agricultura Tropical (CIAT).12 p.
Khan FU, Nelson BD, Helms TC. 2005. Greenhouseevaluation of binucleate Rhizoctonia for control of R.solani in soybean. Plant Dis 89:373–379, doi:10.1094/PD-89-0373
Kottke I, Haug I, Setaro S, Suarez JP, Weiß M, Preusing M,Nebelb M, Oberwinkler F. 2008. Guilds of mycorrhizalfungi and their relation to trees, ericads, orchids andliverworts in a Neotropical mountain rain forest. BasicAppl Ecol 9:13–23, doi:10.1016/j.baae.2007.03.007
Liu G, Jia Y, Correa-Victoria F, Prado G, Yeater K, McClung A,Correll J. 2009. Mapping quantitative trait loci respon-sible for resistance to sheath blight in rice. Phytopathol-ogy 99:1078–1084, doi:10.1094/PHYTO-99-9-1078
Marschner H. 1995. Mineral nutrition of higher plants. 2nded. New York: Academic Press. 889 p.
Moncalvo JM, Nilsson RH, Koster B, Dunham SM, BernauerT, Matheny PB, Porter TM, Margaritescu S, Weiß M,Garnica S, Danell E, Langer G, Langer E, Larsson E,Larsson KH, Vilgalys R. 2006. The cantharelloid clade:dealing with incongruent gene trees and phylogenetic
MOSQUERA-ESPINOSA ET AL.: ORCHID MYCORRHIZAE AND DISEASE BIOCONTROL 149

reconstruction methods. Mycologia 98:937–948, doi:10.3852/mycologia.98.6.937
Mosquera-Espinosa AT, Bayman P, Otero JT. 2010. Cerato-basidium como hongo micorrızico de orquıdeas enColombia. Acta Agron 59:316–326.
Otero JT, Ackerman JD, Bayman P. 2002. Diversity and hostspecificity of endophytic Rhizoctonia-like fungi fromtropical orchids. Am J Bot 89:1852–1858, doi:10.3732/ajb.89.11.1852
———, ———, ———. 2004. Differences in mycorrhizalpreferences between two tropical orchids. Molec Ecol10:207–212.
———, Bayman P, Ackerman JD. 2005. Variation inmycorrhizal performance in the epiphytic orchid Tolum-nia variegata in vitro: the potential for natural selection.Evol Ecol 19:29–43, doi:10.1007/s10682-004-5441-0
———, Flanagan N, Herre A, Ackerman JD, Bayman P.2007. Widespread mycorrhizal specificity correlates tomycorrhizal function in the Neotropical, epiphyticorchid Ionopsis utricularioides (Orchidaceae). Am JBot 94:1944–1950, doi:10.3732/ajb.94.12.1944
———, Thrall PH, Clements M, Burdon JJ, Miller JT. 2011.Codiversification of orchids (Pterostylidinae) and theirassociated mycorrhizal fungi. Aust J Bot 59:480–497,doi:10.1071/BT11053
Pascual C, Raymundo A, Hyakumachi M. 2000. Efficacy ofhipovirulent binucleate Rhizoctonia spp. to controlbanded leaf and sheath blight in corn. J Gen PlantPathol 66:95–102, doi:10.1007/PL00012928
Porras-Alfaro A, Bayman P. 2007. Mycorrhizal fungi ofVanilla: diversity, specificity and effects on seedgermination and plant growth. Mycologia 99:510–525,doi:10.3852/mycologia.99.4.510
Rasmussen HN. 2002. Recent developments in the studyof orchid mycorrhiza. Plant Soil 244:149–163, doi:10.1023/A:1020246715436
———, Rasmussen FN. 2009. Orchid mycorrhiza: implica-tions of a mycophagous life style. Oikos 118:334–345,doi:10.1111/j.1600-0706.2008.17116.x
Savary S, Castilla P, Elazegui A, McLaren G, Ynalvez M, TengS. 1995. Direct and indirect effects of nitrogen supplyand disease source structure on rice sheath blight spread.Phytopathology 85:959–965, doi:10.1094/Phyto-85-959
Selosse MA, Roy M. 2009. Green plants that feed on fungi:facts and questions about mixotrophy. Trends Plant Sci14:64–70, doi:10.1016/j.tplants.2008.11.004
Slaton N, Cartwright R, Meng J, Gbur E, Norman R. 2003.Sheath blight severity and rice yield as affected bynitrogen fertilizer rate, application method and fungi-cide. Agron J 95:1489–1496, doi:10.2134/agronj2003.1489
Sneh B. 1998. Use of non-pathogenic or hypovirulent fungalstrains to protect plants against closely related fungalpathogens. Biotechnol Adv 16:1–2, doi:10.1016/S0734-9750(97)00044-X
———, Burpee L, Ogoshi A. 1991. Identification ofRhizoctonia species. St Paul, Minnesota: AmericanPhytopathological Society. 133 p.
———, Yamoah E, Stewart A. 2004. Hypovirulent Rhizocto-nia spp. isolates from New Zealand soils protect radishseedlings against Damping-off caused by R. solani. NZPlant Protect 57:54–58.
Suarez JP, Michael W, Abele A, Garnica S, Oberwinkler F,Kottke I. 2006. Diverse tulasnelloid fungi form mycor-rhizas with epiphytic orchids in an Andean cloudforest. Mycol Res 110:1257–1270, doi:10.1016/j.mycres.2006.08.004
Valadares RB, Pereira MC, Otero JT, Cardoso EJ. 2012.Narrow fungal mycorrhizal diversity in a population ofthe orchid Coppensia doniana. Biotropica 44:114–122,doi:10.1111/j.1744-7429.2011.00769.x
Villajuan-Abgona R, Kageyama K, Hyakurnachi M. 1996.Biocontrol of Rhizoctonia damping-off of cucumber bynon-pathogenic binucleate Rhizoctonia. Eur J PlantPathol 102:227–235, doi:10.1007/BF01877961
Wen K, Seguin P, St-Arnaud M, Jabaji-Hare S. 2005. Real-time quantitative RT-PCR of defense-associated genetranscripts of Rhizoctonia solani-infected bean seedlingsin response to inoculation with a nonpathogenicbinucleate Rhizoctonia isolate. Phytopathology 95:345–353, doi:10.1094/PHYTO-95-0345
Xue L, Charest PM, Jabaji-Hare SH. 1998. Systemicinduction of peroxidases, 1,3-b-glucanases, chitinasesand resistance in bean plants by binucleate Rhizoctoniaspecies. Phytopathology 88:359–365, doi:10.1094/PHYTO.1998.88.4.359
150 MYCOLOGIA





























