Embed Size (px)
Citation preview

www.elsevier.com/locate/brainres
Brain Research 999 (2004) 9–19
Research report
The epithalamus of the developing and adult frog: calretinin expression
and habenular asymmetry in Rana esculenta
Vittorio Guglielmottia,*, Luigia Cristinoa, Errico Sadaa, Marina Bentivogliob
a Institute of Cybernetics, CNR, ‘‘E. Caianiello’’, I-80078 Pozzuoli, Naples, ItalybDepartment of Morphological and Biomedical Sciences, University of Verona, Verona, Italy
Accepted 14 October 2003
Abstract
Expression of the calcium binding protein (CaBP) calretinin (CR) was studied with immunohistochemistry in the pineal complex and
habenular nuclei (HN) of the developing and adult frog Rana esculenta. The frog pineal complex is a medial structure formed by two
interconnected components, the frontal organ and the pineal organ or epiphysis; the habenular nuclei are bilateral and are asymmetric due
to subdivision of the left dorsal nucleus into medial and lateral components. In the pineal complex, calretinin immunostaining of cells and
fibers was consistently observed in developing and adult frogs. In the habenulae, calretinin immunoreactivity exhibited instead marked
variations during development, and was expressed only in cells of the medial subnucleus of the left dorsal habenula. In particular,
calretinin was detected at larval stages, peaked during metamorphosis, was markedly downregulated at the end of metamorphosis, and was
evident again in adulthood. This sequence of calretinin expression was confirmed by quantitative analysis of immunoreactive cells in the
left habenula. In tadpoles, calretinin-positive cells exhibited a dorsoventral gradient of density, while in adulthood, they were distributed
throughout the dorsoventral extent of the medial subnucleus. The study demonstrates a peculiar developmental pattern, with transient
downregulation, of asymmetric calretinin expression in the frog epithalamus. The findings indicate that calcium and calcium buffering
systems may play critical roles in neurogenetic and neuronal migration processes implicated in the formation of the asymmetric habenular
portion in amphibians. In addition, the reappearance of calretinin expression in the adult frog supports a distinct functional role of the
asymmetric habenular component in amphibians.
D 2004 Elsevier B.V. All rights reserved.
Theme: Development and regeneration
Topic: Cell differentiation and migration
Keywords: Asymmetry; Calretinin; Development; Epithalamus; Frog; Immunocytochemistry
1. Introduction circuits [10]. The organization of the epiphysis exhibits
The habenular nuclei (HN) and the epiphysis are dien-
cephalic structures located medially in the epithalamus of all
vertebrates. The HN are connected with the interpeduncular
nucleus forming the habenulo-interpeduncular system,
which is one of the circuits most conserved in phylogenesis.
In the vertebrate brain, the epithalamus plays a main
functional role in the regulation of cyclic behavior, such
as circadian and reproductive functions, as well as in the
modulation of other systems, and in particular limbic
0006-8993/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.brainres.2003.10.053
* Corresponding author. Tel.: +39-81-8675134; fax: +39-81-8675128.
E-mail address: [email protected] (V. Guglielmotti).
marked variations in different taxa. Thus, in cyclostomes,
the epiphysis evolves as pineal complex, e.g. pineal organ
proper and parapineal organ, to turn into a single epiphyseal
outgrowth, the pineal gland, in mammals. The HN include
bilaterally two nuclei protruding into the third ventricle and
interconnected by the habenular commissure.
Marked structural asymmetry of the HN represents a
striking feature of the epithalamus in several non-mamma-
lian species [9,16,27,30,31,34,49], and of the pineal com-
plex in most anamniotes and reptiles [17,18,59,60]. In
particular, the comparison between the left and right HN
in the brain of many vertebrates pointed out structural
differences in size, neuronal organization, neurochemistry
and connectivity between the two sides, raising a debate

V. Guglielmotti et al. / Brain Research 999 (2004) 9–1910
about the evolutionary origin of epithalamic structures (see
Ref. [13] for review). Recent data have demonstrated that in
the zebrafish diencephalon, the left–right identity is influ-
enced by the parapineal organ [22].
In the frog, the HN are subdivided into dorsal and ventral
nuclei, homologous to the medial and lateral nuclei, respec-
tively, in mammals. Morphological asymmetries have been
reported in the left dorsal habenular nucleus of Rana
esculenta [32] and they correlate with unilateral projections
of the left habenula to the frontal organ [33], which is the
extracranial component of the pineal complex. The func-
tional significance of such asymmetric organization of the
frog HN is, however, still unclear.
Asymmetric neurochemical features have also been de-
scribed in the frog epithalamus. In particular, selective left–
right differences in the expression of nitric oxide synthase
(NOS), the synthetic enzyme of the free radical nitric oxide,
have been reported in the HN and pineal complex of R.
esculenta during development [26]. Asymmetry of calreti-
nin (CR) immunoreactivity was also mentioned in the HN of
adult frogs [41]. This latter feature indicates differences in
calcium buffering systems between the two sides of the frog
brain, because CR is a calcium binding protein (CaBP)
which acts in the control of the fluctuation of cytosolic
calcium ions. Genes coding for CR are well preserved
during vertebrate evolution [43] and CR is the main EF-
hand CaBP expressed in regions of the frog nervous system
[21]. Fluctuations of calcium ions are involved in many
neuronal functions (see Ref. [39] for review). In particular,
intracellular calcium buffering levels represent a key pa-
rameter during neural development, and a direct relationship
between calcium and neuronal differentiation has been
suggested [2,3,25,40].
Transient expression of CR during development has been
reported to play a primary role in the reshaping of the
habenulo-interpeduncular system in the trout [44]. However,
no data are available on the distribution of CR in the
developing HN of other vertebrates, and, in particular in
the developing and mature pineal complex of amphibians.
On this basis, the present work was aimed at investigating
the ontogenesis of CR expression in the frog epithalamus,
and at a correlation of such data with previous findings
obtained in this brain region.
2. Materials and methods
2.1. Animals
Tadpoles and adult frogs of the species R. esculenta
were used. Fertilized spawn (available during spring) and
adult animals (25–30 g body weight, corresponding to
about 3 years of age) were collected from the countryside
around Naples. The spawn were bred at room temperature
according to a previously described protocol [24]. The
collection of the material was authorized by the competent
Italian authorities, and the experiments were conducted
under institutional approval. All efforts were made to
minimize animal suffering and the number of animals
used.
Following the staging criteria of Manelli and Margar-
itora [38] for R. esculenta, we used tadpoles of stages 31,
34, 38, 40, 44, 48, and 50 (stage 50 is the last one).
These tables [38] subdivide the development of the frog
in three periods: the embryonic and larval periods, and
metamorphosis. Briefly: (i) the embryonic period includes
stages 1–30, related to the use of deutoplasm as nutritive;
(ii) the larval period, including stages 31 and 32, is
characterized by autonomous feeding and appearance of
the hindlimb buds; (iii) metamorphosis includes stages
33–50: tadpoles develop mainly the body and hindlimbs
until stage 44, followed by the appearance of forelimbs,
atrophy of the tail, and capability also of terrestrial life in
stages 45–50.
Due to difficulties in the supply and breeding of post-
metamorphic frogs, juvenile animals were not available.
2.2. Tissue preparation for immunohistochemistry
For CR imunohistochemistry, tadpoles sampled at the
above-mentioned stages (n = 4 per stage) were anesthetized
in a 0.2 g/l solution of tricaine methanesulfonate (MS 222;
Sigma, St. Louis, MO) and fixed intracardially with 4%
paraformaldehyde and 2% picric acid (salt) in 0.1 M
phosphate buffer, pH 7.4 (PB). The perfusion was per-
formed by injecting the fixative through a glass micropipette
(tip diameter: 60–70 Am), connected with a plastic tube to a
syringe. The viscera were removed after perfusion, and
tadpoles at stages 31–34 were processed as wholemounts.
In animals at stages 38–50, undesired tissue was removed
by transverse cuts through the caudal tip of the spinal cord
and then longitudinally at level of the eyes; the head,
including the frontal organ was thus dissected. Adult frogs
(n = 4) were anesthetized in 0.35 g/l solution of MS 222 and
perfused through the aorta with 10 ml of 0.64% NaCl
followed by 25 ml of the same fixative used for tadpoles.
The brain and the flap of skin containing the frontal organ
were then dissected out.
The tissue samples were postfixed for 4 h at room
temperature in the same fixative used for perfusion. The
excess of picric acid was washed out by several changes in
PB, and the tissue samples were then soaked in 30% sucrose
in PB overnight at 4 jC for cryoprotection. They were
finally embedded in a solution of 6% agar and 30% sucrose
in PB for cutting. Horizontal (n = 3 animals per age group)
and transverse serial sections (n = 1 animal per age group)
were cut on a cryostat at a 14-Am thickness. All sections
were collected, mounted in two adjacent series on glass
slides pretreated with Vectabond (Vector, Burlingame, CA),
and processed for CR immunohistochemistry. After the
immunoreaction, one of the series of horizontal sections
was counterstained with cresyl violet.

V. Guglielmotti et al. / Brain Research 999 (2004) 9–19 11
Due to the anatomical features of the pineal organ, which
is flattened on top of the habenular commissure, the hori-
zontal sections provided the largest surface of epithalamic
structures, allowing also a detailed study of the most dorsal
levels of the HN.
2.3. Tissue preparation for Nissl staining
Adult frogs (n = 2) and tadpoles at the same stages used
for immunohistochemistry (n= 2 per stage) were destined
to the cytoarchitectonic study in cresyl violet-stained
sections. These animals were fixed and dissected using
the same procedure described above. The tissue samples
were postfixed for 3 h at room temperature, washed in PB,
dehydrated, embedded in paraffin and cut at 15 Am in
serial sections, that were collected on slides pretreated with
Vectabond. One animal per age group was cut in the
horizontal plane and the other in the transverse plane.
2.4. CR immunohistochemistry
The sections destined to immunohistochemistry,
mounted on slides, were immersed in 0.75% H2O2 in Tris
buffer saline at pH 7.3 (TBS) to block endogenous
peroxidase activity. After rinsing in TBS, they were
preincubated in 10% normal goat serum (NGS) and
0.4% Triton-X100 in TBS for 1 h, followed by incubation
for 3 days at 4 jC in rabbit anti-CR (Swant, Bellinzona,
Switzerland), diluted 1:2000 in the preincubation solution.
The specificity of this antibody for the frog brain was
previously assessed by Western blot analysis [41].
After incubation in primary antibody, the sections were
rinsed and incubated for 4 h at room temperature in
secondary biotinylated goat anti-rabbit IgG (Vector), dilut-
ed 1:200 in the preincubation solution. After several rinses
in TBS, the sections were incubated for 2 h in avidin–
biotin complex (Vector), diluted in TBS according to the
supplier’s indications, followed by rinses in TBS and by
reaction with 3–3Vdiaminobenzidine (Sigma) and H2O2 for
4–5 min. The sections were then rinsed and, as men-
tioned above, one of the horizontal series per animal was
counterstained with Nissl staining. Finally, the sections
were dehydrated, cleared in xylene and coverslipped with
DPX.
For control, transverse sections randomly collected dur-
ing the cutting procedure were treated with the same
protocol but omitting the primary antibody. No immunopo-
sitivity was found in these sections.
2.5. Nissl staining
For Nissl staining, the sections were stained with 0.1%
cresyl violet (Merck) in 0.1 M acetate–acetic acid buffer,
pH 3.7. The sections were then differentiated in alcohol,
dehydrated, cleared in xylene and coverslipped with
DPX.
3. Data analysis
All the material was examined at the microscope under
bright-field illumination. The proportion of CR-immuno-
positive cells was evaluated in the left dorsal habenula in
all the immunoreacted and counterstained sections of
developing and adult animals (n = 3 per age group). All
CR-immunoreactive neurons containing a darkly stained
nucleolus visible in the focal plane, as well as immuno-
negative cells exhibiting darkly stained nucleoli, were
counted, using a 40� objective, in the medial subnucleus
of the left dorsal habenula, in which CR-immunostained
cells were concentrated (see below). In addition, to make
sure that cell counting was not affected by bias in the
delimitation of the subnuclei of the left habenula in
different sections and animals, cells were counted in both
the components (the medial and lateral subnuclei) of the
left dorsal habenula.
4. Results
4.1. Asymmetry of the mature and developing habenulae
The data on habenular asymmetry observed in the
present investigation in cresyl violet-stained sections were
consistent with those described in the adult and developing
frog in previous studies [9,26], and will be briefly recalled
here for clarity. The asymmetry of the adult frog HN is
due to peculiarities of the left dorsal nucleus (Fig. 1A),
which is subdivided into medial and lateral subnuclei,
represented by two fused shells of cells surrounding a
central neuropil, while a single dorsal nucleus is located on
the right side of the HN. In R. esculenta, the rostromedial
part of the medial subnucleus is further subdivided by a
septum in two distinct compartments, named medial and
lateral compartment or neuropil (Fig. 1B). As illustrated in
Fig. 1A, the mediocaudal portion of the medial subnucleus
is formed by a neuropil wrapped in a shell of cells.
A similar cytoarchitectonic organization was observed
in the dorsal HN of developing animals. In the left
nucleus, the different compartments developed sequentially
(Fig. 1C–E). The left medial habenular subnucleus and the
right nucleus were the first structures appearing in the
epithalamus during development. The left lateral subnu-
cleus appeared at stage 30 but became prominent only
from stage 34 onward (Fig. 1C), and developed further
until the end of metamorphosis (Fig. 1E).
4.2. CR immunoreactivity in the epithalamus
As it will be presented in detail below, throughout the
examined developmental stages and in adulthood, CR
immunoreactivity was observed in cells and fibers of the
pineal complex. CR exhibited instead a peculiar pattern of
expression in the HN, since it was detected only in cells of

Fig. 1. Photomicrographs of Nissl-stained transverse (A–D) and horizontal (E) sections through the habenular nuclei at different rostrocaudal levels of adult
(A,B) and developing (C–E) frogs. (A) Section through the middle portion, showing the features of habenular asymmetry in the adult frog: the left dorsal
habenular nucleus comprises the medial (M) and lateral (L) subnuclei, whereas a single nucleus (R) is seen on the right side. (B) Section through the
rostromedial level, showing that a septum subdivides further the medial subnucleus into medial (m) and lateral (l) compartments. C–E, in which the circled
numbers indicate the developmental stage, show the sequence of cytoarchitectural arrangement of the medial subnucleus into medial and lateral compartments,
as well as the development of the lateral subnucleus (arrowhead in C; L in D and E) which starts from stage 31. III, third ventricle; hc, habenular commissure.
Scale bars: 100 Am.
V. Guglielmotti et al. / Brain Research 999 (2004) 9–1912
the left dorsal habenula, and mainly in those of the medial
compartment, with temporal variations.
4.2.1. Stages 31, 34
CR immunoreactivity was found in cells of the medial
subnucleus of the left dorsal habenula since the first
examined stage (Fig. 2A), when the lateral subnucleus
was not yet recognizable. At stage 31, intense CR
immunostaining of cells and fibers was observed in the
pineal organ and in the frontal organ (Fig. 2B, C). In the
latter structure, CR positivity was evident in cells of
different sizes, including relatively large elements (6.5–
11.5 Am in diameter) that exhibited intense immunostain-
ing of cell bodies and their processes (Fig. 2C). The
pineal tract also appeared intensely stained in the epiph-
ysis (Fig. 2B).
At subsequent stages, e.g. stage 34, the organization of
the HN progressed towards a mature configuration, and CR
immunoreactivity was evident in clusters of cells located in
the left habenula and in the adjacent pineal region, where
labeling of cells and of pineal tract fibers was observed in
the epiphysis (Fig. 2D). At stage 34, the percentage
(F standard deviation) of CR-immunoreactive neurons was
4.1F 0.4% of the cells of the medial subnucleus, and
3.6F 0.2% of the total population of cells of the left dorsal
habenula (Fig. 3). It should be reminded that at this stage,
the medial subnucleus occupied most of the extent of the
dorsal habenula.
4.2.2. Stage 38
Compartmentalization of the left dorsal habenula was
well evident at this stage (Fig. 2F). CR immunoreactivity,
clearly asymmetric, was evident in cells of the medial
compartment of the medial subnucleus close to the third
ventricle (Fig. 2E, F). Cells exhibiting various degrees of
intensity of CR immunoreactivity were observed (Fig. 2E,
F). At this stage, the proportion of CR-immunoreactive
neurons was higher than at stage 34, accounting for
10.0F 0.8% of the cells of the medial subnucleus and
for 8.7F 0.6% of the cells of the entire left dorsal
habenula (Fig. 3).
Intense CR immunopositivity of cells and fibers was still
observed in the extra- and intracranial portions of the pineal
complex, including the pineal tract.

Fig. 2. CR immunoreactivity in the left dorsal habenula and pineal complex during development in horizontal sections at different stages (indicated by
circled numbers). (A) At stage 31, a few labeled cells (arrow) are clustered in the dorsal region of the medial habenular subnucleus (M). (B) CR-
immunostained cells and fibers are evident in the pineal organ (arrows), from which fibers of the pineal tract depart (arrowhead). (C) CR-
immunostained cells bodies and their processes are also evident in the frontal organ (FO). (D) At stage 34, the pineal organ (arrow), pineal tract
(arrowhead) and cells in the dorsal portion of the medial habenular subnucleus (M) are the only CR-immunopositive structures of the epithalamic
region. (E) At stage 38, cells at the most dorsal level of the medial habenular compartment (m) are intensely immunolabeled. (F) At the ventral level
of the same sample, cells of the medial neuropil (m) lining the third ventricle (III) exhibit moderate immunoreactivity. (G) At stage 40, CR
immunolabeling of cells (arrowhead) is evident in the medial subnucleus (M), grouped near the third ventricle (III), and in the pineal organ (arrow).
(H) Higher power image of intensely (white arrows) and moderately (arrowheads) CR-positive cells in the medial compartment (m). (I) At stage 44,
the low power view of the epithalamus shows CR immunoreactivity on the left side (arrowhead), while the right habenula (R) appears devoid of
labeling; the arrows point to the pineal organ. (J): Intense immunostaining of cells at the most dorsal level of the medial subnucleus. (K) Intense
immunolabeling of cells and fibers in both the medial subnucleus (M; arrowhead) and pineal organ (arrows). Scale bars: 100 Am in A–F; 50 Am in
G–K.
V. Guglielmotti et al. / Brain Research 999 (2004) 9–19 13
4.2.3. Stages 40, 44
The pattern of CR immunoreactivity observed at stage
40 in the HN and pineal organ was similar to that
described above. In the habenular medial compartment,
immunostained cells were still mainly grouped near the
third ventricle (Fig. 2G, H), exhibiting clear asymmetry. In
the pineal complex, immunolabeling was observed in cells
and fibers.
CR labeling of both the left dorsal habenula (Fig. 2J)
and pineal complex (Fig. 2I, K) was also evident at stage
44, so that the features of immunoreactivity of these two
structures, very close one to the other, were still very
similar (Fig. 2I, K).
The relative proportion of CR-immunoreactive neurons
in the left habenula peaked at stage 40 (17.8F 0.5% in the
medial subnucleus, and 9.4F 0.6% in the entire left dorsal

Fig. 3. Bar diagram illustrating the percentage (F standard deviation) of
CR-immunoreactive cells with respect to the total cells population of the
medial subnucleus (black bars) or the entire left dorsal habenula (white
bars) at different stages of development and in adulthood (A).
V. Guglielmotti et al. / Brain Research 999 (2004) 9–1914
habenula), and decreased at stage 44 (12.9F 0.6%, and
6.5F 0.3%, respectively) (Fig. 3).
4.2.4. Stages 48, 50
At stage 48, the HN exhibited mature cytoarchitectonic
features (Fig. 4C). Interestingly, CR immunoreactivity was
found to be drastically downregulated in the left habenula at
this stage (Fig. 4A, C), while it was still present in the pineal
complex (Fig. 4B). In the HN, CR labeling was detected
only in a few cells located in the dorsal region of the medial
III cp
A
M
III
B
CPO
48
Fig. 4. (A–C) CR immunoreactivity at stage 48 in horizontal sections. (A) Sectio
positive cells (arrowheads) in the medial habenular subnucleus (M). Labeled fibe
several cells are intensely labeled. Immunostained fibers of probable commissural
through the pineal organ (large arrow) showing immunostaining of the frontal ne
along the choroid plexus (cp) of the third ventricle (III), and enters the brain at th
indicated by arrowhead). (C) CR-immunostained and Nissl-counterstained section
at the border of the third ventricle (III) are evident in the medial subnucleus (M). No
the boxed regions. Scale bars: 100 Am.
subnucleus (Fig. 4A, C). Fibers located at the border of the
left dorsal habenula, of probable commissural origin, were
also CR-stained (Fig. 4A). The proportion of immunoreac-
tive neurons had decreased further with respect to the
previous stages, accounting at stage 48 for 10.5F 0.7% of
the cells of the medial subnucleus and 6.1F 0.4% of those
of the left habenula (Fig. 3).
At stage 50, a few CR-positive cells were observed in the
most dorsal part of the left habenula (Fig. 5A, B), while no
CR labeling was observed in the other portions of the HN.
CR immunoreactivity was still observed in neurons and
fibers of the pineal complex (Fig. 5A, C). In the frontal
organ (Fig. 5C), some cells were devoid of labeling, while
stained fibers of epiphyseal neurons located at the border of
this structure were oriented toward the adjacent left dorsal
habenula (Fig. 5B). The percentage of CR-immunoreactive
neurons dropped at this stage to 6.8F 0.6% in the medial
subnucleus and to 3.6F 0.3% with respect to the total cell
population of the left dorsal habenula (Fig. 3).
4.2.5. Adulthood
Asymmetric expression of CR was detected in the adult
animals in the medial subnucleus of the left dorsal habenula
(Fig. 5D, F). Most of the immunoreactive cells were intensely
stained. The CR-immunostained cells were mainly clustered
in the habenular region bordering the third ventricle (Fig. 5F),
and they accounted for 14.3F 0.8% of cells of the medial
subnucleus and for 7.8F 0.6% of the total cell population of
the left habenula (Fig. 3). No CR imunoreactivity was found
in the lateral subnucleus and in the right habenula (Fig. 5D,
E). Intensely stained cells and their processes were also
observed in the pineal complex (Fig. 5G, H).
M
LIII
cp
n through the left dorsal habenula and pineal organ showing the paucity of
rs of the frontal nerve (large arrow) enter the pineal organ (PO), in which
origin (small arrows) surround the habenular subnucleus. (B) Dorsal section
rve (small arrow) that runs rostrocaudally (from left to right in the image)
e level of the habenular commissure (which lies under the black skin strip
through the left dorsal habenula: a few positive cells (arrows) located mainly
te the difference in the size of cells of the medial and lateral (L) subnuclei in

Fig. 5. CR-labeling in horizontal (A–G) and transverse (H) sections through the left dorsal habenula and pineal complex at the end of metamorphosis (stage 50;
A–C) and in adulthood (indicated by the letter A; D–H). (A) Dorsal level of the habenula showing a few immunostained cells (white arrows) in the medial
subnucleus (M). The pineal organ (PO) and the frontal nerve (arrow) are intensely positive. (B) Higher magnification of the area boxed in A, showing lightly
stained but well evident processes of pineal cells (arrowheads) oriented toward the left habenula at the border between the two regions. (C) Labeling of cells
and their processes in the frontal organ (FO). (D) Intensely labeled cells and processes in the medial subnucleus (M). (D, E) The lateral subnucleus (L) and the
right nucleus (R) are devoid of labeling. (F) Ventral level of the medial subnucleus (M) of the dorsal left habenula illustrating the intense staining of cells
mainly lining the third ventricle (III). Both D and F show habenular sections at more ventral levels than those shown for tadpoles. (G, H) CR immunoreactivity
in the frontal organ and pineal organ, respectively; the arrowhead in H points to an immunolabeled pineal cell and its process. Scale bars: 100 Am in A, D–F;
20 Am in B; 50 Am in C, G, H.
V. Guglielmotti et al. / Brain Research 999 (2004) 9–19 15
4.2.6. Overall cytoarchitectonic arrangement of CR-immu-
noreactive neurons in the left dorsal habenula
In addition to the developmental sequence of expression
described above, it is noteworthy that the topographical
distribution of the asymmetric CR positivity was different in
the left habenula of developing and adult animals (Fig. 6).
During development, intensely CR-immunostained cells
were packed in the left dorsal habenula along a dorso-
ventral gradient of density and, as described above, they
were mainly grouped close to the third ventricle (Fig.
2D, E, H, J). In the adult frog, intensely CR-immuno-
reactive neurons, and their processes oriented toward the
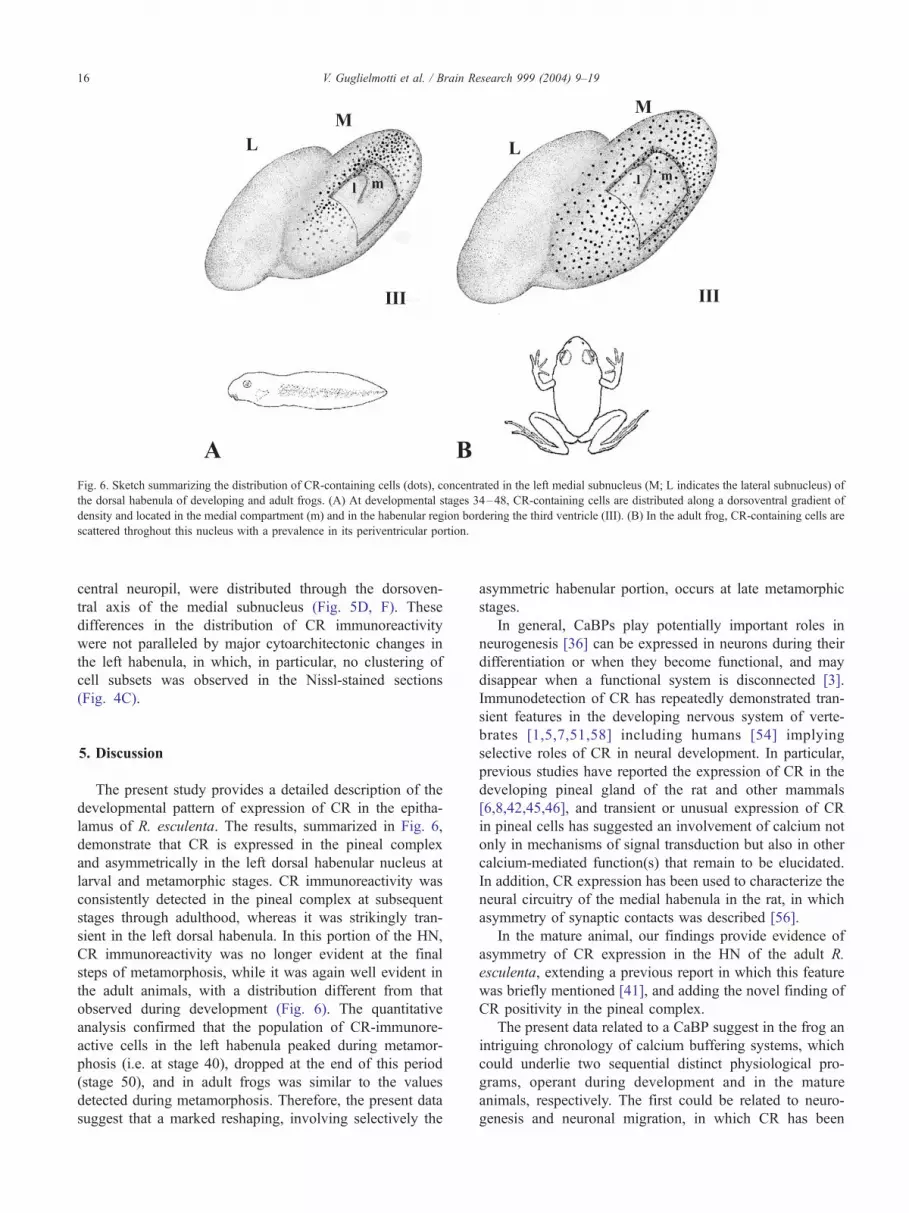
Fig. 6. Sketch summarizing the distribution of CR-containing cells (dots), concentrated in the left medial subnucleus (M; L indicates the lateral subnucleus) of
the dorsal habenula of developing and adult frogs. (A) At developmental stages 34–48, CR-containing cells are distributed along a dorsoventral gradient of
density and located in the medial compartment (m) and in the habenular region bordering the third ventricle (III). (B) In the adult frog, CR-containing cells are
scattered throghout this nucleus with a prevalence in its periventricular portion.
V. Guglielmotti et al. / Brain Research 999 (2004) 9–1916
central neuropil, were distributed through the dorsoven-
tral axis of the medial subnucleus (Fig. 5D, F). These
differences in the distribution of CR immunoreactivity
were not paralleled by major cytoarchitectonic changes in
the left habenula, in which, in particular, no clustering of
cell subsets was observed in the Nissl-stained sections
(Fig. 4C).
5. Discussion
The present study provides a detailed description of the
developmental pattern of expression of CR in the epitha-
lamus of R. esculenta. The results, summarized in Fig. 6,
demonstrate that CR is expressed in the pineal complex
and asymmetrically in the left dorsal habenular nucleus at
larval and metamorphic stages. CR immunoreactivity was
consistently detected in the pineal complex at subsequent
stages through adulthood, whereas it was strikingly tran-
sient in the left dorsal habenula. In this portion of the HN,
CR immunoreactivity was no longer evident at the final
steps of metamorphosis, while it was again well evident in
the adult animals, with a distribution different from that
observed during development (Fig. 6). The quantitative
analysis confirmed that the population of CR-immunore-
active cells in the left habenula peaked during metamor-
phosis (i.e. at stage 40), dropped at the end of this period
(stage 50), and in adult frogs was similar to the values
detected during metamorphosis. Therefore, the present data
suggest that a marked reshaping, involving selectively the
asymmetric habenular portion, occurs at late metamorphic
stages.
In general, CaBPs play potentially important roles in
neurogenesis [36] can be expressed in neurons during their
differentiation or when they become functional, and may
disappear when a functional system is disconnected [3].
Immunodetection of CR has repeatedly demonstrated tran-
sient features in the developing nervous system of verte-
brates [1,5,7,51,58] including humans [54] implying
selective roles of CR in neural development. In particular,
previous studies have reported the expression of CR in the
developing pineal gland of the rat and other mammals
[6,8,42,45,46], and transient or unusual expression of CR
in pineal cells has suggested an involvement of calcium not
only in mechanisms of signal transduction but also in other
calcium-mediated function(s) that remain to be elucidated.
In addition, CR expression has been used to characterize the
neural circuitry of the medial habenula in the rat, in which
asymmetry of synaptic contacts was described [56].
In the mature animal, our findings provide evidence of
asymmetry of CR expression in the HN of the adult R.
esculenta, extending a previous report in which this feature
was briefly mentioned [41], and adding the novel finding of
CR positivity in the pineal complex.
The present data related to a CaBP suggest in the frog an
intriguing chronology of calcium buffering systems, which
could underlie two sequential distinct physiological pro-
grams, operant during development and in the mature
animals, respectively. The first could be related to neuro-
genesis and neuronal migration, in which CR has been

V. Guglielmotti et al. / Brain Research 999 (2004) 9–19 17
repeatedly implicated [19,20,52,53]. These processes estab-
lish an intimate histogenetic and functional relationship
between the epiphysis and a part of it that migrates into
the habenular region to form the asymmetric portion of the
HN. The second program could be triggered after complete
differentiation of the epithalamus, when the asymmetric
portion of the HN has acquired functional independence.
Altogether the present results in the developing frog
suggest transient functional activity in the left HN, in
contrast with the persistence of parameters related to CaBP
expression in the pineal complex. These findings parallel
those observed in a previous study on NOS positivity in the
HN and pineal complex [26], indicating that the develop-
mental program of the frog epithalamus involves an array of
different molecules. In agreement with the previous results
obtained in the study of NOS, CR was here found to be
expressed during development in the left dorsal habenula
and mainly located in the medial neuropil, a distinct region
of the left medial subnucleus bordering the third ventricle.
Such variations in protein expression are likely to be
involved in the establishment of asymmetrical activity in
the diencephalon of amphibians, and could represent a
phylogenetic step leading to more complex lateralization
phenomena during brain evolution.
Very few data are available on the expression of CaBPs
in the adult and developing epithalamic structures of verte-
brates. In the HN of adult vertebrates, contradictory findings
have been reported on CaBP expression in the different
components of the HN, i.e. in the lateral and/or medial
habenula [11,23,29], but habenular asymmetry was not
reported in these studies. Asymmetric distribution of the
CaBP calbindin was described in the HN of the dogfish by
Rodriguez-Moldes et al. [47] who, however, did not inves-
tigate other epithalamic structures, such as the epiphysis.
Transient CR expression was reported in the habenular
complex of the developing trout, in which habenular asym-
metry was not described [44].
On the other hand, asymmetric transient expression of
neuroactive molecules in the HN is not an event occurring
only during development. Asymmetry of serotonin-like
immunoreactivity was described in the right habenula of
coho salmon during smolt transformation, a life period
linked to naturally occurring endocrine changes [15]. To-
gether with the present findings, these results indicate that
life conditions characterized by major behavioral and phys-
iological changes, e.g. the amphibian metamorphosis or the
smolt transformation in coho salmon, induce unilateral
variations in the habenular complex. It is also interesting
to note that in coho salmon a persistent asymmetric expres-
sion of serotonin was reported in the dorsal subdivision of
the left habenular nucleus and in the pineal organ [16]. The
present findings strikingly parallel such data. Thus, also in
the frog, our neurochemical data support a strict correlation
of the HN with the pineal complex, previously pointed out
on the basis of cytological similarities and topographical
proximity [16]. Similar conclusions have also been reached
in the lizard [17], in which HN asymmetry was hypothe-
sized to represent a specialization associated with unilateral
projection from the parietal eye.
A growing body of anatomical and functional evidence
supports close interactions between the habenular complex
and the pineal gland also in mammals [50]. The region
implicated in such relationship is the medial habenular
nucleus [12,35,48]. Although habenular asymmetry was
not reported in the above studies, it is worth recalling that,
as mentioned above, the medial habenula of mammals is
homologous to the dorsal habenula of the frog [28]. It is
also noteworthy that asymmetry is not an exclusive pecu-
liarity of lower vertebrates, because habenular left–right
differences have been demonstrated in birds and mammals
[27,30,57].
Studies on the developing diencephalon have reported
that the laterality of the asymmetric development of the
HN and pineal complex is regulated by Nodal signaling
pathway genes and by factors released from midline tissue
[14], whose role in left–right axis formation appears to be
conserved from fish to human [4]. In zebrafish, the genes
implicated in such mechanism are expressed on the left
side of the embryonic dorsal diencephalon, in a region
corresponding to the presumptive epiphysis or pineal organ
[14,37]. In zebrafish, the habenular asymmetry is lost
following selective ablation of the parapineal organ [22].
In the sea lamprey, proliferating cell nuclear antigen
immunoreactivity was found in the parapineal organ and
in the asymmetric habenular primordium; moreover, pro-
liferating cells were observed in this species in cells of the
asymmetric right habenula, mostly confined to its ventro-
caudal area [55].
Taken together with the present findings in amphibia, the
above studies indicate that diencephalic asymmetries, very
marked in many species of cyclostomes, fish and reptiles,
can derive from an interplay between the pineal and
habenular primordia. Furthermore, the data recall attention
on the study of cerebral asymmetries in vertebrates.
Acknowledgements
The preparation of this manuscript was supported by
grants from the Italian National Research Council (CNR).
References
[1] A. Ambrus, R. Kraftsik, I. Barakat-Walter, Ontogeny of calretinin
expression in rat dorsal root ganglia, Dev. Brain Res. 106 (1998)
101–108.
[2] L. Anglister, I.C. Farber, A. Shahar, A. Grinvald, Localization of
voltage-sensitive calcium channels along developing neurites: their
possible role in regulating neurite elongation, Dev. Biol. 94 (1982)
351–365.
[3] K.G. Baimbridge, M.R. Celio, J.H. Rogers, Calcium-binding proteins
in the nervous system, Trends Neurosci. 8 (1992) 303–308.

V. Guglielmotti et al. / Brain Research 999 (2004) 9–1918
[4] R.N. Bamford, E. Roessler, R.D. Burdine, U. Saplakoglu, J. de la Cruz,
M. Splitt, J. Towbin, P. Bowers, B. Marino, A.F. Schier, M.M. Shen,
M. Muenke, B. Casey, Loss of function mutations in the EGF–CFC
gene CFC1 are associated with human left – right laterality defects,
Nat. Genet. 26 (2000) 365–369.
[5] E. Bastianelli, R. Pochet, Transient expression of calretinin during
development of chick cerebellum. Comparison with calbindin-D28k,
Neurosci. Res. 17 (1993) 53–61.
[6] E. Bastianelli, R. Pochet, Calbindin-D28k, calretinin, and recoverin
immunoreactivities in developing chick pineal gland, J. Pineal Res. 17
(1994) 103–111.
[7] E. Bastianelli, R. Pochet, Calbindin-D28k, calretinin, and S-100 im-
munoreactivities in rat pineal gland during postnatal development,
J. Pineal Res. 18 (1995) 127–134.
[8] E. Bastianelli, K. Moutairou, M.T. Akele-Akpo, R. Darboux, R. Po-
chet, Calcium binding proteins immunohistochemistry and identifica-
tion of neurons in the mammalian pineal gland of the African giant
rat: Cricetomys gambianus, Gen. Physiol. Biophys. 18 (1999) 5–17.
[9] V. Braitenberg, M. Kemali, Exceptions to bilateral symmetry in
the epithalamus of lower vertebrates, J. Comp. Neurol. 138 (1970)
137–146.
[10] A.B. Butler, W. Hodos, Comparative Vertebrate Neuroatomy. Evolu-
tion and Adaptation, Wiley-Liss, 1996.
[11] M.R. Celio, Calbindin D-28k and parvalbumin in the rat nervous
system, Neuroscience 35 (1990) 375–475.
[12] M.D. Chafetz, F.H. Gage, Pineal and habenula innervation altered by
septal lesions, Brain Res. Bull. 10 (1983) 27–31.
[13] M.L. Concha, S.W. Wilson, Asymmetry in the epithalamus of verte-
brates, J. Anat. 199 (2001) 63–84.
[14] M.L. Concha, R.D. Burdine, C. Russell, A.F. Schier, S.W. Wilson, A
Nodal signaling pathway regulates the laterality of neuroanatomical
asymmetries in the zebrafish forebrain, Neuron 28 (2000) 399–409.
[15] L.O.E. Ebbesson, B. Holmqvist, T. Ostholm, P. Ekstrom, Transient
serotonin-immunoreactive neurons coincide with a critical period of
neural development in coho salmon (Oncorhynchus kisutch), Cell
Tissue Res. 268 (1992) 389–392.
[16] P. Ekstrom, S.O.E. Ebbesson, The left habenular nucleus contains a
discrete serotonin-immunoreactive subnucleus in the coho salmon
(Oncorhynchus kisutch), Neurosci. Lett. 91 (1988) 121–125.
[17] G.A. Engbretson, A. Reiner, N. Brecha, Habenular asymmetry and
the central connections of the parietal eye of the lizard, J. Comp.
Neurol. 198 (1981) 155–165.
[18] W.D. Eldred, T.E. Finger, J. Nolte, Central projections of the frontal
organ of Rana pipiens, as demonstrated by the anterograde transport
of horseradish peroxidase, Cell Tissue Res. 211 (1980) 215–222.
[19] P.S. Eriksson, E. Perfilieva, T. Bjork-Eriksson, A.M. Alborn, C. Nord-
borg, D.A. Peterson, F.H. Gage, Neurogenesis in the adult human
hippocampus, Nat. Med. 4 (1998) 1313–1317.
[20] A.V. Faure, D. Grunwald, M.J. Mountin, M. Hilly, J.P. Mauger, I.
Marty, M. De Waard, M. Villaz, M. Albrieux, Developmental ex-
pression of the calcium release channels during early neurogenesis of
the mouse cerebral cortex, Eur. J. Neurosci. 14 (2001) 1613–1622.
[21] R. Gabriel, B. Volgyi, E. Pollak, Calretinin-immunoreactive elements
in the retina and optic tectum of the frog, Rana esculenta, Brain Res.
782 (1998) 53–62.
[22] T.J. Gamse, C. Thisse, B. Thisse, M.E. Halpern, The parapineal me-
diates left – right asymmetry in zebrafish diencephalon, Development
130 (2003) 1059–1068.
[23] L.M. Garcia-Segura, D. Baetens, J. Roth, A.W. Norman, L. Orci,
Immunohistochemical mapping of calcium-binding protein immu-
noreactivity in the rat central nervous system, Brain Res. 296 (1984)
75–86.
[24] D. Gioffre, Un acquario per la Rana, Aquarium 6 (1976) 393–396.
[25] R. Guglielmone, G. Corvetti, First appearance and distribution of cal-
retinin-immunoreactive neurons in the early development of the chick
central nervous system, Cell Tissue Res. 300 (2000) 21–28.
[26] V. Guglielmotti, L. Fiorino, Nitric oxide synthase activity reveals an
asymmetrical organization of the frog habenulae during development:
a histochemical and cytoarchitectonic study from tadpoles to the ma-
ture Rana esculenta, with notes on the pineal complex, J. Comp.
Neurol. 411 (1999) 441–454.
[27] C.J. Gurusinghe, D. Erhlich, Sex-dependent structural asymmetry of
the medial habenular nucleus of the chicken brain, Cell Tissue Res.
240 (1985) 149–152.
[28] J.A. Harris, V. Guglielmotti, M. Bentivoglio, Diencephalic asymme-
tries, Neurosci. Biobehav. Rev. 20 (1996) 637–643.
[29] S.S. Jande, L. Maler, D.E.M. Lawson, Immunohistochemical map-
ping of vitamin D-dependent calcium-binding protein in brain, Nature
294 (1981) 765–767.
[30] M. Kemali, Morphological asymmetry of the habenulae of a macros-
matic mammal, the mole, Z. Mikrosk.-Anat. Forsch. 98 (1984)
951–954.
[31] M. Kemali, I. Agrelli, The habenulo-interpeduncular nuclear system
of a reptilian representative Lacerta sicula, Z. Mikrosk.-Anat. Forsch.
85 (1972) 325–333.
[32] M. Kemali, V. Braitenberg, Atlas of the Frog’s Brain, Springer, Hei-
delberg, 1969.
[33] M. Kemali, A. De Santis, The extracranial portion of the pineal com-
plex of the frog (frontal organ) is connected to the pineal, the hypo-
thalamus, the brain stem and the retina, Exp. Brain Res. 53 (1983)
193–196.
[34] M. Kemali, A. Miralto, E. Sada, Asymmetry of the habenulae in the
elasmobranch Scyllium stellare. Light microscopy, Z. Mikrosk.-Anat.
Forsch. 94 (1980) 794–800.
[35] H.W. Korf, T. Sato, A. Oksche, Complex relationships between the
pineal organ and the medial habenular nucleus-pretectal region of the
mouse as revealed by S-antigen immunocytochemistry, Cell Tissue
Res. 261 (1990) 493–500.
[36] E.D. Lephart, H. Taylor, N.A. Jacobson, M.A. Watson, Calretinin and
calbindin-D28K in male rats during postnatal development, Neuro-
biol. Aging 19 (1998) 253–257.
[37] J.O. Liang, A. Etheridge, L. Hantsoo, A.L. Rubinstein, S.J. Nowak,
J.C. Izpisua Belmonte, M.E. Halpern, Asymmetric Nodal signaling in
the zebrafish diencefhalon positions the pineal organ, Development
127 (2000) 5101–5112.
[38] H. Manelli, F. Margaritora, Tavole cronologiche dello sviluppo di
Rana esculenta, Rendiconti Accademia Nazionale dei XL. XII
(1961) 1–15 + 12 Tables.
[39] M.P. Mattson, Calcium as sculptor and destroyer of neural circuitry,
Exp. Gerontol. 27 (1992) 29–49.
[40] M.P. Mattson, A. Taylor-Hunter, S.B. Kater, Neurite outgrowth in
individual neurons of a neuronal population is differentially regulated
by calcium and cyclic AMP, J. Neurosci. 8 (1988) 1704–1711.
[41] D. Necchi, C. Soldani, M.B. Pisu, G. Bernocchi, E. Scherini, Distri-
bution of calretinin-like immunoreactivity in the brain of Rana es-
culenta, J. Chem. Neuroanat. 16 (1999) 233–243.
[42] A. Novier, D. Nicolas, R. Krstic, Calretinin immunoreactivity in pineal
gland of different mammals including man, J. Pineal Res. 21 (1996)
121–130.
[43] M. Parmentier, Structure of the human cDNAs and genes coding for
calbindin D28k and calretinin, in: R. Pochet, D.E.M. Lawson, C.W.
Heizmann (Eds.), Calcium Binding Proteins in Normal and Trans-
formed Cells, Plenum, New York, 1990, pp. 27–34.
[44] A. Porteros, J.G. Brinon, R. Arevalo, C. Crespo, J. Aijon, J.R.
Alonso, Transient expression of calretinin in the trout habenulo-in-
terpeducular system during development, Neurosci. Lett. 254 (1998)
9–12.
[45] P. Redecker, Developmental pattern of cell type-specific calretinin
immunoreactivity in the postnatal gerbil pineal gland, Brain Res.
Dev. Brain Res. 105 (1998) 43–50.
[46] P. Redecker, Y. Cetin, H.W. Korf, Differential immunocytochemical
localization of calretinin in the pineal gland of three mammalian
species 25 (1996) 9–18.
[47] I. Rodriguez-Moldes, J.P. Timmermans, D. Adriaensen, M.H.A. De

V. Guglielmotti et al. / Brain Research 999 (2004) 9–19 19
Groodt-Lasseel, D.W. Scheuermann, R. Anadon, Asymmetric distri-
bution of calbindin-D28k in the ganglia habenulae of an elasmobranch
fish, Anat. Embryol. 181 (1990) 389–391.
[48] O.K. Ronnekleiv, M. Moller, Brain–pineal nervous connections in
the rat: an ultrastructure study following habenular lesion, Exp. Brain
Res. 37 (1979) 551–562.
[49] C.M. Rovainen, Neurobiology of lampreys, Physiol. Rev. 59 (1979)
1007–1077.
[50] R. Sandyk, Relevance of the habenular complex to neuropsychiatry: a
review and hypothesis, Int. J. Neurosci. 61 (1991) 189–219.
[51] B. Schlosser, G. Klausa, G. Prime, G. Ten Bruggencate, Postnatal
development of calretinin- and parvalbumin-positive interneurons in
the rat neostriatum: an immunohistochemical study, J. Comp. Neurol.
405 (1999) 185–198.
[52] M. Setzer, N. Ulfig, Differential expression of calbindin and calretinin
in the human fetal amygdala, Microsc. Res. Tech. 46 (1999) 1–17.
[53] Y. Toba, K. Ajiki, M. Horie, K. Sango, H. Kawano, Immunohistochem-
ical localization of calbindin D-28k in the migratory pathway from the
rat olfactory placode, J. Neuroendocrinol. 13 (2001) 683–694.
[54] N. Ulfig, Calcium-binding proteins in the human developing brain,
Adv. Anat. Embryol. Cell Biol. 165 (2002) 1–92.
[55] B. Villar-Cheda, E. Perez-Costas, M. Melendez-Ferro, X.M. Abalo,
R. Rodrıguez-Munoz, R. Anadon, M.C. Rodicio, Proliferating cell
nuclear antigen (PCNA) immunoreactivity and development of the
pineal complex and habenula of sea lamprey, Brain Res. Bull. 57
(2002) 285–287.
[56] J.A. Wilson, M.D. Kawaja, Distribution of calretinin-immunoreactive
septal axons in the normal and deafferented medial habenula of adult
rats, J. Comp. Neurol. 374 (1996) 593–606.
[57] A. Wree, K. Zilles, A. Schleicher, Growth of fresh volumes and
spontaneous cell death in the nuclei habenulae of albino rats during
ontogenesis, Anat. Embryol. 161 (1981) 419–431.
[58] Y.H. Yan, J.F. Van Brederode, A.E. Hendrickson, Transient co-local-
ization of calretinin, parvalbumin, and calbindin-D28K in developing
visual cortex of monkey, J. Neurocytol. 24 (1995) 825–837.
[59] J. Yanez, H. Meissl, R. Anadon, Central projections of the parapineal
organ of the adult rainbow trout (Oncorhynchus mykiss), Cell Tissue
Res. 285 (1996) 69–74.
[60] J. Yanez, M.A. Pombal, R. Anadon, Afferent and efferent connections
of the parapineal organ in lampreys: a tract tracing and immunocy-
tochemical study, J. Comp. Neurol. 403 (1999) 171–189.





























