Embed Size (px)
Citation preview

The evolution of foliar terpene diversity in Myrtaceae
Amanda Padovan • Andras Keszei •
Carsten Kulheim • William J. Foley
Received: 4 February 2013 / Accepted: 7 October 2013 / Published online: 13 October 2013
� Springer Science+Business Media Dordrecht 2013
Abstract Plant terpenes play many roles in natural
systems, from altering plant–animal interactions, to
altering the local abiotic environment. Additionally,
many industries depend on terpenes. For example,
commercially used essential oils, including tea tree oil
and lavender oil, are a mixture of terpenes. Many
species of the family Myrtaceae form a key resource
for these industries due to the high concentration of
terpenes found predominately in their leaves. The
frequency of chemotypic differences within many
species and populations can lead to costly errors in
industry. Terpene diversity in Myrtaceae is driven by
variation in the terpene synthase enzymes, which
catalyse the conversion a few common substrates into
thousands of terpene structures. We review terpene
diversity within and between species of Myrtaceae and
relate this to variation in the terpene synthase enzymes
to reconstruct the evolution of foliar terpene diversity
in Myrtaceae. We found that (1) high inter- and intra-
species variation exists in terpene profile and that a-
pinene the most likely ancestral foliar terpene, and (2)
that high concentration of 1,8-cineole (a compound
which is regarded as the signature compound of
Myrtaceae) is limited to just four Myrtaceae sub-
families. We suggest that the terpene synthase
enzymes do not limit terpene diversity in this family
and variation in these enzymes suggests a mode of
enzymatic evolution that could lead to high 1,8-
cineole production. Our analysis highlights the need to
standardise methods for collecting and reporting foliar
terpene data, and we discuss some methods and issues
here. Although there are many gaps in the published
data, our large scale analysis using the results of many
studies, shows the value of a family wide analysis for
understanding both the evolution and industrial
potential of terpene-producing plants.
Keywords Monoterpene � Sesquiterpene �Terpene synthase � Eucalyptus � Melaleuca �Cineole � Evolution
Terpenes are arguably one of the most important
groups of compounds in the plant kingdom because
they form such a large classes of plant secondary
metabolites and play many roles in the lives of plants.
Terpenes facilitate interactions between a plant and its
biotic environment, for example terpenes are involved
in direct and indirect defence against herbivory in the
terrestrial and marine worlds (Andrew et al. 2007;
Maida et al. 1993; Matsuki et al. 2011; Van Poecke
et al. 2001), defence against pathogens (Huang et al.
Electronic supplementary material The online version ofthis article (doi:10.1007/s11101-013-9331-3) contains supple-mentary material, which is available to authorized users.
A. Padovan (&) � A. Keszei � C. Kulheim � W. J. Foley
Research School of Biology, Australian National
University, Building 116 Daley Road, Canberra,
ACT 0200, Australia
e-mail: [email protected]
123
Phytochem Rev (2014) 13:695–716
DOI 10.1007/s11101-013-9331-3

2012; Levin 1976) and in pollinator attraction (Pi-
chersky and Gershenzon 2002). They slow down
decomposition of leaf litter on the forest floor,
resulting in slower nutrient cycling within the ecosys-
tem (Horner et al. 1988). Terpenes can also alter the
local abiotic environment, through air quality and thus
local climate; for example Australian eucalypt forests
are known to release terpenes, particularly during the
day, changing the ionic composition of the air
resulting in higher local temperatures and lower local
humidity and rainfall (Suni et al. 2008; Vickers et al.
2009).
Terpenes in Myrtaceae
Terpenes are a defining feature of Myrtaceae, which is
renowned for having some of the highest concentra-
tions of foliar terpenes in the plant kingdom (Keszei
et al. 2008). The recent completion of the Eucalyptus
grandis genome, the first genome from Myrtaceae to
be fully sequenced (Grattapaglia et al. 2012), has
revealed the presence of many more terpene synthase
enzymes than in other terpene-rich plant species.
These enzymes catalyse the synthesis of terpene
structures from a relatively small pool of substrates.
Approximately 112 terpene synthases were identified
in the E. grandis genome, with most (*80) being
expressed in young and mature leaves (Kulheim et al.
2013). The next largest terpene synthase family is
from grape (Vitis vinifera), which contains 53 terpene
synthase sequences, however the fruits and leaves of
grape contain neither a diverse profile nor high
concentrations of terpenes (Martin et al. 2010). The
discovery of such a large family of terpene synthases
in the E. grandis genome prompted us to examine the
patterns of terpene diversity in Myrtaceae.
There are 17 tribes of Myrtaceae comprising
approximately 140 genera and 5,500 species (Biffin
et al. 2010; Thornhill and Crisp 2012). Myrtaceae are
predominately found in the southern hemisphere with
species diversity hotspots in Australia and south east
Asia (Thornhill and Crisp 2012). Originally, two sub-
families of Myrtaceae were recognised; the fleshy-
fruited Myrtoideae and the dry capsular-fruited Le-
ptospermoideae (reviewed by (Thornhill and Crisp
2012). However, more recent molecular phylogenies
have shown there are only two distantly related tribes
containing fleshy-fruited species, but these tribes also
contain dry-fruited species: Myrteae and Syzygieae
(Biffin et al. 2010; Thornhill et al. 2012; Wilson et al.
2005). Species from the Xanthostemoneae, Myrteae
and Syzygieae are commonly found in the tropical
rainforests of Australia and south-east Asia (Brophy
et al. 2006), whereas species of Eucalypteae, Mela-
leuceae and Leptospermeae are common in forest
woodlands from the dry tropics through to the
temperate zones (Brophy and Southwell 2002). With
leaf morphology closely related to habitat (Wilson
2011), woodland plants are predicted to defend their
leaves much more strongly than do rainforest plants
(Grubb et al. 1998). Therefore, we might expect the
leaves of rainforest species have fewer terpenes
compared with those of open forest and woodland
species notwithstanding the many other roles terpenes
can play in the plant.
Not surprisingly, people have exploited plants from
this family for many industrial purposes, for example
tea tree oil is extracted from Melaleuca alternifolia
(Butcher et al. 1994; Carson et al. 2006), eucalyptus
oil is extracted from the 1,8-cineole rich leaves of
several Eucalyptus species (Boland et al. 1991) and
guava (the fruit of Psidium guajava) is a common food
crop and important in traditional south east Asian
medicines (Chen et al. 2007).
Terpenes play a number of roles in the interaction
between a plant and its environment, however to date
the terpenes in Myrtaceae have only been implicated
in defensive roles against herbivores and pathogens. If
terpenes are predominately employed in defensive
roles in Myrtaceae, there may have been a co-
evolutionary arms race with herbivores, which would
drive diversification of terpene profiles within and
between species (Kant and Baldwin 2007) and would
explain the significant qualitative and quantitative
variation in terpene profiles within species, popula-
tions and even individuals (Andrew et al. 2010;
Padovan et al. 2012; Wallis et al. 2011). While intra-
specific diversity of terpene profiles present a viable
evolutionary response for survival in a large range of
environmental conditions (for example: Crankshaw
and Langenheim 1981; Langenheim et al. 1980;
Linhart and Thompson 1995), it also presents a
significant problem for those industries that rely on a
specific terpene profile and both quality and quantity
of terpenes affect of the profitability of the essential oil
industries (Homer et al. 2000).
696 Phytochem Rev (2014) 13:695–716
123

How is qualitative and quantitative variation
in foliar terpenes affected by the terpene
biosynthesis pathway?
The terpene biosynthetic pathway is well understood
(Dudareva et al. 2005; Kulheim et al. 2011; Lichtent-
haler 1999). Briefly, the mevalontate (MVA) and
methylerythritol phosphate (MEP) pathways result in
the production of isopentenyl pyrophosphate (IPP),
the universal precursor to all terpenes (Dudareva et al.
2005, Lichtenthaler 1999). IPP is isomerised to
dimethyl allyl pyrophsphate (DMAPP). Subsequently,
DMAPP and IPP connect to form a small number of
prenyl pyrophosphates. The MVA pathway is local-
ised in the cytosol and produces IPP that is used in the
synthesis of farnesyl pyrophosphate (which is used by
sesqui- and triterpene synthases) and is localised in the
cytosol (Dudareva et al. 2005). The MEP pathway,
localised in the chloroplast, produces IPP that is used
in the synthesis of geranyl and geranylgeranyl pyro-
phosphate (which are used by mono- and di- and
tetraterpene synthases, respectively) and is localised in
the chloroplast (Lichtenthaler 1999). The prenyl
pyrophosphates formed by the connection of DMAPP
and IPP are the substrates for terpene synthase
enzymes. The terpene synthases are a large family,
and catalyse the conversion of a small set of substrates
into thousands of terpene products (Wise et al. 1998).
There are two points in the biosynthetic pathway that
influence the qualitative profile of terpenes. Firstly, the
ratio of monoterpenes to sesquiterpenes is influenced
by the synthesis of geranyl pyrophosphate (GPP) and
farnesyl pyrophosphate (FPP), the precursors for
monoterpene and sesquiterpenes, respectively. Both
are synthesised from the same precursors but by
unique enzymes in different cellular compartments
(Dudareva et al. 2005; Kulheim et al. 2011; Lichtent-
haler 1999). Variation in the efficiency of these
enzymes can influence the monoterpene:sesquiterpene
ratio (Degenhardt et al. 2009; McCaskill and Croteau
1995).
Secondly, the qualitative terpene profile can be
influenced via the action of terpene synthases. Melal-
euca quinquenervia provides a good example of this.
The foliar terpene chemotypes of this species correlate
with differential expression of at least two terpene
synthases (Padovan et al. 2010). Terpene synthases are
often multi-product enzymes, with some accepting
several prenyl pyrophosphate substrates (e.g. GPP and
FPP, for mono and sesquiterpenes, respectively), to
produce terpene skeletons (Keszei et al. 2008; Wise
et al. 1998). The overall amino acid sequence of
terpene synthases is highly conserved across species
(Bohlmann et al. 1998), however these enzymes have
variable catalytic pockets to facilitate the production
of a range of terpene skeletons, and some enzymes
have two areas in the catalytic pocket that are involved
in distinct steps in the reaction cascade (Kollner et al.
2006).
Although many terpenes are direct products of
terpene synthases, some terpenes are formed by the
actions of one or more cytochrome P450 (CYP)
enzymes acting on the product of a terpene synthase-
catalysed reaction. For example, limonene can be
converted into trans-isopiperitenol, trans-carveol,
carvone, and menthol (Bohlmann and Keeling 2008;
Bouwmeester et al. 1999; Croteau et al. 2005) and a-
pinene can be converted to myrtenol (Aharoni et al.
2004) through a variety of CYP enzymes. Whereas
several economically important terpenes require a
CYP step in their biosynthesis, there are few data
available on cytochrome P450 enzymes known to play
a role in terpene biosynthesis in Myrtaceae and the
terpenes that are known to require a CYP for their
biosynthesis are not common in the leaves of Myrt-
aceae species. Therefore we will focus on terpene
synthase enzymes and how these relate to foliar
terpene variation in Myrtaceae.
Understanding terpene variation in plants addresses
many ecological and evolutionary questions, includ-
ing differential herbivory in thyme (Thymus vulgaris)
(Linhart and Thompson 1995) and origins of intra-
specific chemical variation (Butcher et al. 1994).
Because terpene variation is a direct result of enzy-
matic variation, we can explore the evolution of
chemical variation. Comparison of terpene data from
large groups of species will contribute to our under-
standing of their evolution and ecological interactions,
and may elucidate the origin of terpene production in
related species. Foliar terpene data and terpene
synthase sequence data have been published for
thousands of species worldwide and contain a wealth
of ecological and evolutionary information. Yet, it
remains remarkably under explored. One recent
exception for the Myrtaceae was a study of latitudinal
variation of Eucalyptus foliar terpenes in Australia
(Steinbauer 2010), although that only sampled 66 out
of the more than 900 Eucalyptus taxa. Here we collate
Phytochem Rev (2014) 13:695–716 697
123

foliar terpene data from 1,362 species of Myrtaceae
and describe the patterns of foliar terpene production
and the developments of chemotypes across the
family, in order to:
1. identify evolutionary trends in foliar terpene
diversity within and between species of
Myrtaceae
2. identify constraints on terpene production with
and between species of Myrtaceae
3. identify species that may be of commercial
importance and highlight species where caution
should be taken
Search methods and data processing
Terpene diversity
We compiled a database of the foliar terpene diversity of
all species of Myrtaceae for which published data were
available. We included the year of the study, the species,
the number of chemotypes and the foliar terpene
profile(s). We recorded the five most abundant terpenes
but only if these contributed more than 7 % to the total
oil. We documented the order of most abundant
compounds, but did not record the concentration or
proportion of each compound as these were measured
and recorded differently by various authors. We used the
molecular phylogeny described by Thornhill and Crisp
(2012) to determine which sub-family each species
belonged to and used EUCLID (an electronic identifi-
cation tool for Eucalyptus: Slee et al. 2006) to determine
which section each Eucalyptus species belongs to. For
this study, chemotypes are defined by their single most
abundant compound, called dominant terpenes. Due to
differences in methods between publications, it was
difficult to determine if differences in minor compounds
(those that were not the most abundant) are the result of
chemotypic differences or due to differences in methods
of extraction and analysis.
We have collated data from a number from
publications dated between 1920 and 2012. On top
of the technological changes that have influenced the
data quality in that time, the aim of many publications
is different and thus the number of samples collected is
highly variable. To account for this variation in
experimental design and still look at patterns of foliar
terpene variation across Myrtaceae, we used an
artificial weighting system based on the order of
abundance of compounds in each species. We gave the
most abundant compound in a profile a value of 100,
the second most abundant compound a value of 50, the
third most abundant compound a value of 25 and so on
to the sixth most abundant compound which has a
value of 3. This allowed us to calculate the mean of
these values for all species within a tribe (Thornhill
and Crisp 2012) and we presented this in a heat map
(Table 1—all species are included in Supp. Table 1).
The heat map allows us to visualise patterns in large
data sets, in this case we can easily see the distribution
of specific terpenes across the family Myrtaceae. We
have also used this data to investigate the distribution
of chemotypes across the family. However the original
experiments were not designed for this purpose, which
means there is no consistency in the age or develop-
mental stage of the tissues collected. Whilst this is a
limitation, we were able to identify some interesting
patterns that warrant further investigation.
Terpene synthase enzymes
We compiled a separate database containing data from
site-directed mutagenesis studies on angiosperm ter-
pene synthase enzymes. We chose site-directed muta-
genesis studies only as these provide a good system for
investigating enzyme structure and function. We
recorded the year of the study, the species, the name
of the enzyme, all sequence variations of that enzyme
investigated in the study, the product profile and relative
activity of each enzyme and variant and the x-ray
crystallographic structure used to generate a 3D model
of this enzyme. We generated our own 3D models of the
10 proteins, using the crystallographic structure of
bornyl diphosphate synthase from Salvia officinalis
(1n20A—sourced from the ExPASy database through
SPBDV (Guex et al. 1995–2011). The monoterpene
synthase sequences were truncated before the RRX8W
motif to correspond to the truncation of the model
(1n20A). This motif signifies the end of the plastid-
targeting sequence in monoterpene synthases (Wise
et al. 1998). We used the SPBDV program (Guex et al.
1995–2011) to generate a 3D homology model of the
enzymes and submitted this to the ExPASy sever for
optimisation using default parameters (Arnold et al.
2006). We then generated 2D ‘maps’ of the genes from
the start to stop codons. On these we plotted the amino
acids that contribute to the internal surface of the
698 Phytochem Rev (2014) 13:695–716
123

catalytic pocket, those that were altered in the site-
directed mutagenesis study and the highly conserved
DDXXD motif (Fig. 1).
A brief description of the data
We collected data from 1,393 species of Myrtaceae
from 67 genera of 13 tribes (Tables 1, 2). For some
species, there was more than one chemical profile
reported and we have designated these as putative
chemotypes. This means we have terpene data for
1,694 samples across the family. These samples are not
evenly distributed across all tribes (Table 1). Mela-
leuceae, Eucalypteae and Syncarpieae are well sam-
pled for foliar terpenes, with data available for more
than 75 % of species in each tribe. Despite being some
of the most specious tribes, very few species of
Chamelaucieae (3 %), Myrteae (6 %) and Syzygieae
(1 %) have been sampled for foliar terpene analysis
(Table 1). Some species of Myrtaceae are much better
investigated than others and this unavoidably intro-
duces bias into our analysis. Irrespective of the number
of species in a tribe, those species whose leaves are
industrially important are more extensively sampled
for foliar terpenes than those that contain species
whose leaves are less industrially important.
What patterns are there in foliar terpene
chemotypic development across the family
Myrtaceae (Tables 2, 3)?
There are 175 species of Myrtaceae with more than
one chemical form or chemotype in this study (14 %
Table 1 A heat map of the most abundant foliar terpenes in all species of Myrtaceae
4 4 4 4 4 4 4 4 5 5 5 6 6 6
- - - - - - - - + - + - - -
- - - - - - - - - - - - - -
1.7 0.3 1.9 - 4.1 - 0.4 0.2 0.3 - 0.0 - - 0.6
20.0 - - - - - - - - - - - - -
1.4 0.1 8.4 0.0 0.7 - - - 2.0 0.6 0.2 0.4 1.9 4.7
1.6 - 1.6 - 3.6 - - - - - 0.6 - - -
7.1 - 14.3 - - - - - - - - - - -
- - - - - - - - - - - - - -
0.4 0.0 0.6 0.7 1.3 0.8 0.9 0.7 - - 0.4 - - 0.0
- - - - - - - - - - - - - -
- - - - - - - - - - - - - -
- - - - - - - - - - - - - -
- - - - - 13.3 - - - - - - - 3.3
- - - - - - - - - - - - - -
Carbocation 0 0 1 1 1 1 1 1 1 2 2 2 3 3 3 3 3 3 3 3 3 3 3 3 4
P450 + + - + - - + - + - - - - - + - + - - + - - - + -
Xanthostemoneae 2 7 - - - - - - - - - - 12.5 - 25.0 - - - - - - - - - - - -
Melaleuceae 3 404 - 0.5 - 0.9 0.0 0.2 0.3 0.5 - 4.5 0.2 0.2 37.3 3.5 0.0 4.9 0.6 0.2 9.2 - 58.9 6.7 0.2 - 2.0
Syncarpieae 1 4 - - - - - - - - - - - - 120.0 - - - - - - - - - - - -
Eucalypteae 3 797 - 0.1 0.2 0.3 0.4 0.1 0.3 - 0.4 0.2 0.3 0.1 37.5 2.9 0.3 3.7 0.1 - 3.2 0.3 50.2 3.3 0.4 0.6 0.1
Leptospermeaea 4 77 - 1.3 1.3 0.6 1.5 - 0.2 0.3 - 1.3 0.6 0.6 40.3 0.3 - 10.9 1.3 - 3.2 - 15.6 0.6 - - -
Chamelaucieae 5 13 - - - - - - 14.3 - - 7.1 - - 92.9 3.6 - - - 21.4 21.4 - 35.7 - - - -
Lindsayomyrteae 1 1 - - - - - - - - - - 25.0 - - - - - - - - - - - - - -
Myrteae 35 355 1.8 - 0.7 0.6 - - - 0.6 - 2.6 1.4 2.0 24.8 0.6 - 5.2 0.1 - 9.0 0.1 9.8 1.7 0.6 - 0.4
Kanieae 5 12 - - - - - - - - - - - 15.4 76.9 - - 15.4 - - 23.1 - - - - - 7.7
Tristanieae 2 2 - - - - - - - - - - - - 75.0 - - - - - - - - - - - -
Backhousieae 1 3 - - - 33.3 - - 33.3 16.7 - - - - - - - - - - - - - - - - -
Syzygieae 4 15 - - 6.7 - - 0.8 - 3.3 - 1.7 - - 15.0 - - 10.0 - - - - - 7.5 - - -
Metrosidereae 1 1 - - - - - - - - - - - - - - - - - - - - - - - - -
CarbocationP450 - - - - - - - - + - - - - - - - - - - - - - - - +
Xanthostemoneae 2 7 - - - 6.3 - 12.5 - 37.5 - 6.3 - 37.5 - - - 25.0 - - 25.0 25.0 - - - - 18.8
Melaleuceae 3 404 - 0.0 0.3 - 0.5 - - 3.9 0.9 0.3 - 0.2 - - 0.6 0.1 - - 1.2 0.6 4.6 0.2 2.3 - 5.2
Syncarpieae 1 4 - - - - - - - - - - - - - - - - - - - - - - - - -
Eucalypteae 3 797 0.7 - 0.6 - 0.6 - - 0.9 0.2 0.2 - 0.3 - - 0.1 0.1 - - - - 8.0 0.3 6.6 - 4.3
Leptospermeaea 4 77 1.9 - 1.3 1.3 2.9 - - 7.5 - 3.7 - 0.3 - - - 0.3 - 0.3 0.2 0.6 12.7 - 3.9 - 3.2
Chamelaucieae 5 13 - - - - - - - - - - - - - - - - - - - - - - - - -
Lindsayomyrteae 1 1 - - - - - - - 100.0 - 50.0 - - - - - - - - - - - - - - -
Myrteae 35 355 0.3 0.8 0.7 0.7 2.4 1.7 0.6 17.8 5.3 1.8 0.6 2.1 0.7 0.8 2.1 1.2 0.7 2.7 1.8 1.2 1.0 - 7.4 0.7 7.5
Kanieae 5 12 - - - - 19.2 - 15.4 46.2 - - 7.7 7.7 - - - - - - - - - 7.7 7.7 - 15.4
Tristanieae 2 2 - - - - - - - - - - - - - - - - - - - - 50.0 - - - -
Backhousieae 1 3 - - - - - - - - - - - - - - - - - - - - - - - - -
Syzygieae 4 15 - - - 3.3 - - - 5.0 13.3 0.8 - - - - - 3.3 - 10.0 - - - - 25.0 - 10.0
Metrosidereae 1 1 - - - - - - - - - - - - - - - - - - - - - - - - -
0 1 1 1 2 2 2 2 2 2 3 3 3 3 3 3 3 4 4 4 5 5 6 6 6 6 6 6 6 6 6 6 6- - - - - - - -
- - - - - - - -
5.0 0.8 0.2 0.1 2.5 - - -
20.0 20.0 - - - - - -
5.0 5.9 0.3 0.4 0.6 0.3 - 0.7
5.5 1.3 0.6 - 3.4 - - -
10.7 - - - - - - -
- - - - - - - -
4.4 3.0 1.4 0.8 0.6 - 1.0 0.9
23.1 - - - - - - -
- - - - - - - -
- - - - - - - -
4.1 - - - - - - -
- - - 100.0 - - - -
On the x-axis are the foliar terpenes and on the y-axis are the tribes of species and a phylogeny defined by Thornhill et al. (2012). The
first table contains monoterpenes and the second table contains sesquiterpenes. The most abundant terpenes are shown in a darker grey
and the least abundant terpenes are shown in a lighter grey. Genera refers to the number of genera represented in each tribe and
samples refers to the number of samples represented in each tribe. Carbocation refers to which carbocation is acted on by a terpene
synthase to result in a particular terpene starting from the FPP or GPP in Figure 3 from Keszei et al. (2008). P450 refers to whether a
cytochrome P450 enzyme is involved in the production of each compound: ? = P450 involved, - = P450 not involved
Phytochem Rev (2014) 13:695–716 699
123

Fig. 1 Gene maps of the terpene synthases from angiosperms
investigated through site directed mutagenesis. The black solid
boxes represent the gene from start codon to stop codon. The
conserved DDXXD domain is represented by a raised black bar
in about the middle of the genes. The raised grey bars represent
sites that have been altered in that enzyme during site-directed
mutagenesis studies. The white bars represent amino acids that
contribute to the internal surface of the catalytic pocket
Table 2 The proportion of species captured in the 13 tribes of Myrtaceae in this study
The phylogeny is a simplified version of that reported by Thornhill and Crisp (2012). The column ‘‘Species’’ refers to the number of
species identified in each tribe (Govaerts 2008). The column ‘‘Sampled’’ shows the number of species for which we collected foliar
terpene data and this as a proportion of the total number of species. The columns ‘‘1,8-cineole’’ and ‘‘a-pinene’’ indicate if these
compounds are common and abundant compounds in the tribe: ? symbolises this and - symbolises these are not common or
abundant
700 Phytochem Rev (2014) 13:695–716
123

of species), some of which have not previously been
reported as chemotypes but rather two independent
studies have identified different chemical forms.
Species that are better characterised, either through
multiple studies or heavy sampling within a single
study, have more chemotypic variation. The chemo-
type is defined by the most abundant compound and
most often there are just two chemotypes of each
species, with an average of 2.4 chemotypes per
species. 73 species have chemotypic patterns involv-
ing both monoterpenes and sesquiterpenes, 90 species
have chemotypic patterns involving only monoter-
penes and 11 species have chemotypic patterns
involving only sesquiterpenes. Only 35 (*20 %)
species from this list have neither an a-pinene nor a
1,8-cineole chemotype, despite these commonly being
the most abundant foliar terpene.
In about half of the species, there is a major
remodelling of the terpene biosynthetic pathway
among the two chemotypes, switching between a
monoterpene-dominated profile and a sesquiterpene-
dominated profile. This would involve repartitioning
substrates between the cytosol and the plastid, possible
through unidirectional transport of IPP across the
chloroplast membrane (Webb et al. 2013), and a
change between high expression of (1) FPPS ? ses-
quiterpene synthases and (2) GPPS ? monoterpene
synthases. The majority of the genes involved in
terpene biosynthesis should be regulated differently
between the two chemotypes, possibly with the
exception of some MEP pathway genes, since there
is evidence of substrate sharing across the plastidal
membrane model plants (Dudareva et al. 2005) and as
well in Melaleuca alternifolia (Webb et al. 2013).
Of the 13 tribes, Meleuceae and Eucalypteae have
the largest number of species with foliar terpene
chemotypes and most often there are just two chem-
otypes in a species. In the vast majority of species
(80 %) there is an a-pinene or 1,8-cineole foliar
terpene chemotype. It is less common to see both
chemotypes in a single species, with this scenario
happening in only 20 % of species with chemotypes
(Table 2). High concentrations of foliar a-pinene is
likely to have appeared first in the common ancestor to
all Myrtaceae, whilst high concentrations of foliar 1,8-
cineole is likely to have appeared in the common
ancestor to Melaleuceae and Eucalypteae.
Although this data set is not ideal for exploring
chemotypic diversity across Myrtaceae (the data were
collected using different methods and none of the
experiments were designed for chemotype detection,
with most sampling from one or a few individuals within
a population), we have identified several species with
multiple chemical forms. In many cases, the chemical
differences indicate changes in how terpene biosynthe-
sis proceeds amongst chemotypes. This poses the
question: what benefits does a species get from vastly
different chemical forms? How is terpene biosynthesis
regulated given such a large remodelling has occurred
within a species? Although these questions have been
pursued in other systems, there are not enough data on
Myrtaceae to allow answering the question with any
confidence (see a recent review of this topic by Moore
et al. (2013)). The first step would be to determine the
chemical profile of entire species of Myrtaceae, with
many hundreds of samples from natural populations in
different climates, as has only been done in a few species
for example: Eucalyptus globulus (Wallis et al. 2011),
Eucalyptus tricarpa (Andrew et al. 2013), M. alterni-
folia (Butcher et al. 1994; Homer et al. 2000) and M.
quinquenervia (Ireland et al. 2002).
What evolutionary trends exist in foliar terpene
diversity within the family Myrtaceae (Table 2)?
a-Pinene and 1,8-cineole are the most common
terpenes amongst the Myrtaceae, with more than
50 % of the samples and species having a-pinene or
1,8-cineole as a dominant compound in their foliar
terpene profile. These compounds are also the most
abundant, with approximately 25 and 35 % having a-
pinene and 1,8-cineole, respectively, as the most
abundant compound in the foliar terpene profile of
species and samples. 1,8-cineole is not found as a
dominant compound in any sample in eight tribes
(Xanthostemoneae, Syncarpieae, Lindsayomyrteae,
Kanieae, Tristanieae, Backhousieae, Syzygieae and
Metrosidereae), whereas a-pinene is not a dominant
compound in samples of just three tribes (Lindsayo-
myrteae, Backhousieae and Metrosidereae). The foliar
terpene profiles of species from the latter three tribes is
often dominated by sesquiterpenes, particularly spath-
ulenol, globulol and bicyclogermacrene. The tribes
Melaleuceae, Eucalypteae and Myrteae show the
greatest monoterpene and sesquiterpene diversity in
terms of the number of unique dominant compounds in
the foliar terpene profile.
Phytochem Rev (2014) 13:695–716 701
123

Ta
ble
3T
he
do
min
ant
com
po
un
do
fch
emo
typ
esfr
om
17
5sp
ecie
so
fM
yrt
acea
e
Sp
ecie
s(*
den
ote
sfi
rst
rep
ort
of
po
ssib
lech
emo
typ
icv
aria
tio
n)
Mo
no
terp
ene
chem
oty
pes
Ses
qu
iter
pen
ech
emo
typ
es
Ch
1C
h2
Ch
3C
h4
Ch
1C
h2
Ch
3
(A)
Au
stro
myr
tus
hil
lii
b-P
inen
eD
Cad
inen
e
Ca
mp
om
an
esia
rho
mb
ea*
Lin
alo
ol
Bic
ycl
og
erm
acre
ne
Co
rym
bia
citr
iod
ora
*C
itro
nel
lal
Ele
mo
l
Eu
caly
ptu
sb
au
eria
na*
Cin
eole
Sp
ath
ule
no
l
Eu
caly
ptu
sb
eyer
i*C
ineo
leE
ud
esm
ol
Eu
caly
ptu
sca
lig
ino
sa*
a-P
inen
eE
ud
esm
ol
Eu
caly
ptu
sca
lop
hyl
la*
a-P
inen
eF
arn
eso
l
Eu
caly
ptu
sca
ma
ldu
len
sis
(sy
nE
uca
lyp
tus
rost
rata
,E
uca
lyp
tus
rost
rata
var
bo
rea
lis)
*P
hel
lan
dre
nes
Cin
eole
a- P
inen
e
Sp
ath
ule
no
l
Eu
caly
ptu
scl
ad
oca
lyx
(sy
nE
uca
lyp
tus
cory
no
caly
x)*
a-P
inen
eC
ary
op
hy
llen
eO
xid
e
Eu
caly
ptu
sco
ccif
era
*a
- Ph
ella
nd
ren
e
Cin
eole
Eu
des
mo
l
Eu
caly
ptu
sco
ng
lom
era
ta*
a- P
hel
lan
dre
ne
Glo
bu
lol
Eu
caly
ptu
sco
nic
a*
Cin
eole
a- P
inen
e
Sp
ath
ule
no
l
Eu
caly
ptu
sd
alr
ymp
lea
na
*a
-Pin
ene
Cin
eole
Sp
ath
ule
no
l
Eu
caly
ptu
sd
aw
son
ii*
a- P
hel
lan
dre
ne
Iso
bic
ycl
og
erm
acra
lE
ud
esm
ol
Eu
caly
ptu
sd
eglu
pta
(sy
nE
uca
lyp
tus
na
ud
inia
na
)*A
pp
leO
ilN
ero
lid
ol
Eu
caly
ptu
sd
eleg
ate
nsi
s(s
yn
Eu
caly
ptu
sg
iga
nte
a)
sub
tasm
an
ien
sis*
a- P
hel
lan
dre
ne
Eu
des
mo
l
Eu
caly
ptu
sd
um
osa
*a
-Pin
ene
Sp
ath
ule
no
l
Eu
caly
ptu
sd
wye
ri*
Cin
eole
Sp
ath
ule
no
l
Eu
caly
ptu
sfa
stig
ata
*a
-Pin
ene
Eu
des
mo
l
Eu
caly
ptu
sfi
cifo
lia
*a
-Pin
ene
Bic
ycl
og
erm
acre
ne
Eu
caly
ptu
sg
un
nii
*a
-Pin
ene
Cin
eole
Sp
ath
ule
no
l
Eu
caly
ptu
sla
evo
pin
eaa
-Pin
ene
Eu
des
mo
l
Eu
caly
ptu
sle
uco
ph
loia
*C
ineo
leG
lob
ulo
l
Eu
caly
ptu
sm
acu
lata
*a
-Pin
ene
Cin
eole
Eu
des
mo
lG
uai
ol
Eu
caly
ptu
sm
ela
no
ph
loia
*a
-Pin
ene
Bic
ycl
og
erm
acre
ne
Eu
caly
ptu
sn
eso
ph
ila
*a
-Pin
ene
Vir
idifl
oro
lG
lob
ulo
l
702 Phytochem Rev (2014) 13:695–716
123

Ta
ble
3co
nti
nu
ed
Sp
ecie
s(*
den
ote
sfi
rst
rep
ort
of
po
ssib
lech
emo
typ
ic
var
iati
on
)
Mo
no
terp
ene
chem
oty
pes
Ses
qu
iter
pen
ech
emo
typ
es
Ch
1C
h2
Ch
3C
h4
Ch
1C
h2
Ch
3
Eu
caly
ptu
sn
itid
a(s
yn
Eu
caly
ptu
sa
myg
da
lin
asu
b
nit
ida
,E
uca
lyp
tus
rad
iata
Var
D)*
a-P
hel
lan
dre
ne
Eu
des
mo
l
Eu
caly
ptu
so
bli
qu
a*
Cy
men
eb
- Ph
ella
nd
ren
e
Bic
ycl
og
erm
acre
ne
Eu
caly
ptu
so
btu
sifl
ora
*C
ineo
leB
icy
clo
ger
mac
ren
e
Eu
caly
ptu
sp
au
cifl
ora
(sy
nE
uca
lyp
tus
cori
ace
a,
Eu
caly
ptu
sp
hle
bo
ph
ylla
)*
Cy
men
eE
ud
esm
ol
Eu
caly
ptu
sp
laty
pu
ssu
bh
eter
op
hyl
la*
a-P
inen
eA
rom
aden
dre
ne
Eu
caly
ptu
sp
oly
carp
ab-
Pin
ene
Vir
idifl
oro
lG
lob
ulo
l
Eu
caly
ptu
sp
ruin
osa
*C
ineo
leG
lob
ulo
l
Eu
caly
ptu
sq
ua
dra
ng
ula
taC
ym
ene
Cin
eole
Eu
des
mo
l
Eu
caly
ptu
sru
bid
a*
Cin
eole
Sp
ath
ule
no
l
Eu
caly
ptu
sru
dis
*a-
Pin
ene
Bic
ycl
og
erm
acre
ne
Eu
caly
ptu
ssi
eber
i(s
yn
sieb
eria
na
)*b-
Ph
ella
nd
ren
e
Eu
des
mo
l
Eu
caly
ptu
ssp
ars
a*
a-P
inen
eB
icy
clo
ger
mac
ren
e
Eu
caly
ptu
ste
ssel
lari
s*L
imo
nen
eA
rom
aden
dre
ne
Eu
caly
ptu
sw
ats
on
ian
asu
bw
ast
on
ian
a*
b-P
inen
eB
icy
clo
ger
mac
ren
e
Eu
caly
ptu
sw
ilco
xii*
Cin
eole
Eu
des
mo
l
Eu
caly
ptu
sya
rra
ensi
s*C
ineo
leb
-Car
yo
ph
yll
ene
Eu
gen
iaa
xill
ari
sa-
Pin
ene
Ger
mac
ren
eD
Eu
gen
iab
rasi
lien
sis
Lim
on
ene
a-P
inen
ea
- Th
uje
ne
b-C
ary
op
hy
llen
eS
pat
hu
len
ol
Eu
gen
iab
rasi
lien
sis
-pu
rple
var
iety
a-P
inen
eC
ary
op
hy
llen
eOx
ide
Eu
gen
iap
un
icif
oli
aL
inal
oo
lb
-Car
yo
ph
yll
ene
Eu
gen
iap
yrif
orm
isa-
Pin
ene
Lim
on
ene
Bic
ycl
og
erm
acre
ne
b-C
ary
op
hy
llen
e
Eu
gen
iau
nifl
ora
Lim
on
ene
Ner
oli
do
l
Ku
nze
aa
mb
igu
aa-
Pin
ene
Vir
idifl
oro
lG
lob
ulo
l
Lep
tosp
erm
um
mic
roca
rpu
ma-
Pin
ene
Sp
ath
ule
no
l
Lep
tosp
erm
um
sco
pa
riu
mG
eran
yl
acet
ate
Lin
alo
ol
a-P
inen
eM
eth
ylc
inn
imat
eb
-Car
yo
ph
yll
ene
Ele
men
eC
op
aen
e
Lo
ph
om
yrtu
sco
nfe
rtu
sa-
Pin
ene
Aro
mad
end
ren
e
Phytochem Rev (2014) 13:695–716 703
123

Ta
ble
3co
nti
nu
ed
Sp
ecie
s(*
den
ote
sfi
rst
rep
ort
of
po
ssib
lech
emo
typ
icv
aria
tio
n)
Mo
no
terp
ene
chem
oty
pes
Ses
qu
iter
pen
ech
emo
typ
es
Ch
1C
h2
Ch
3C
h4
Ch
1C
h2
Ch
3
Lo
ph
om
yrtu
sla
cifl
uu
sa
-Pin
ene
Aro
mad
end
ren
e
Lo
ph
om
yrtu
ssu
ave
ole
ns
a-P
inen
eb-
Car
yo
ph
yll
ene
Mel
ale
uca
cord
ata
Cin
eole
Eu
des
mo
l
Mel
ale
uca
inca
na
sub
inca
na
Cin
eole
Glo
bu
lol
Mel
aleu
cale
iop
yx
isL
inal
oo
lC
ineo
leE
ud
esm
ol
Mel
ale
uca
ner
vosa
*C
ineo
leS
pat
hu
len
ol
M.
qu
inq
uen
ervi
a*
Cin
eole
Ner
oli
do
la
-Sel
inen
eV
irid
iflo
rol
Mel
ale
uca
thyo
ides
b-P
inen
eS
pat
hu
len
ol
Mel
ale
uca
tub
ercu
lata
var
.T
ub
ercu
lata
Lin
alo
ol
Eu
des
mo
l
Mel
ale
uca
vim
inea
sub
vim
inea
a-P
inen
eE
ud
esm
ol
Myr
ceu
gen
iacu
cull
ata
a-P
inen
eN
ero
lid
ol
a-S
elin
ene
Myr
ceu
gen
iam
yrci
oid
es*
a-P
inen
eS
pat
hu
len
ol
Myr
cia
cup
rea*
My
rcen
eb-
Car
yo
ph
yll
ene
Myr
cia
ga
le*
My
rcen
eE
ud
esm
ol
Myr
cia
nth
esfr
ag
an
sa
-Pin
ene
Lim
on
ene
a-C
adin
ol
M.
pu
ng
ens
Cin
eole
b-C
ary
op
hy
llen
e
Myr
cia
ria
ten
ella
*a
-Pin
ene
b-C
ary
op
hy
llen
e
Neo
fab
rica
myr
tifo
lia
a-P
inen
eb-
Car
yo
ph
yll
ene
Pim
enta
psu
eud
oca
ryo
ph
yllu
sE
ug
eno
lM
eth
yl
iso
eug
eno
lG
eran
iol
b-C
ary
op
hy
llen
e
Psi
diu
mg
ua
java
*a
-Pin
ene
Lim
on
ene
Cin
eole
Hex
2en
alB
isab
ole
ne
b-C
ary
op
hy
llen
e
Wa
terh
ou
sia
mu
lgra
vea
na
b-P
inen
eB
icy
clo
ger
mac
ren
e
Sp
ecie
s(*
den
ote
sfi
rst
rep
ort
of
po
ssib
lech
emo
typ
icv
aria
tio
n)
Ch
1C
h2
Ch
3C
h4
Ch
5C
h6
(B)
Ba
ckh
ou
sia
citr
iod
ora
*C
itra
lC
itro
nel
lal
Ger
ania
l
Ble
ph
aro
caly
xtw
eed
iei
Cin
eole
a-P
inen
e
Ca
llis
tem
on
vim
ina
lis*
Cin
eole
a-P
inen
e
Ca
lyp
tra
nth
essp
ruce
an
a*
Lim
on
ene
a-P
inen
e
Ca
mp
om
an
esia
ad
am
an
tiu
ma
-Pin
ene
Ger
anio
lL
imo
nen
e
Ch
am
ela
uci
uu
nu
nci
na
tum
Cit
ron
ella
la-
Pin
ene
Lim
on
ene
704 Phytochem Rev (2014) 13:695–716
123

Ta
ble
3co
nti
nu
ed
Sp
ecie
s(*
den
ote
sfi
rst
rep
ort
of
po
ssib
lech
emo
typ
icv
aria
tio
n)
Mo
no
terp
ene
chem
oty
pes
Ses
qu
iter
pen
ech
emo
typ
es
Ch
1C
h2
Ch
3C
h4
Ch
1C
h2
Ch
3
Da
rwin
iaci
trio
do
raM
eth
ylg
eran
ate
Met
hy
lmy
rten
ate
Dec
asp
erm
um
hu
mil
ea
-Pin
ene
b-P
inen
e
Eu
caly
ptu
sa
caci
ifo
rmis
*C
ineo
lea-
Pin
ene
Eu
caly
ptu
sa
cced
ens*
Cin
eole
a-P
inen
e
Eu
caly
ptu
sa
mp
lifo
lia
*C
ineo
lea-
Pin
ene
Eu
caly
ptu
sa
myg
da
lin
a(s
yn
Eu
caly
ptu
ssa
lici
foli
a)*
Cin
eole
a-P
inen
eP
iper
ito
ne
Eu
caly
ptu
sa
nd
rew
sii
sub
an
dre
wsi
i*a
-Pin
ene
a-P
hel
lan
dre
ne
Eu
caly
ptu
sa
ng
op
ho
roid
es*
Cin
eole
a-P
inen
e
Eu
caly
ptu
sa
po
do
ph
ylla
a-P
inen
eL
imo
nen
e
E.
aro
ma
ph
loia
*C
ineo
leA
pp
leO
il
Eu
caly
ptu
sa
stri
ng
ens*
Cy
men
ea-
Pin
ene
Cin
eole
Eu
caly
ptu
sb
osi
sto
an
a*
a-P
inen
eC
ineo
le
Eu
caly
ptu
sb
otr
yoid
es*
a-P
inen
eC
ym
ene
Eu
caly
ptu
sca
lcic
ola
*C
ineo
lea-
Ter
pin
eol
Eu
caly
ptu
sca
leyi
(sy
nE
uca
lyp
tus
coer
ule
a)*
a-P
inen
eC
ineo
le
Eu
caly
ptu
sci
trio
do
raC
itro
nel
lol
Cit
ron
ella
l
Eu
caly
ptu
scn
eori
foli
a*
Cin
eole
a-P
hel
lan
dre
ne
Eu
caly
ptu
sco
nsi
den
ian
a*
a-P
inen
eC
ineo
le
Eu
caly
ptu
sco
rnu
ta*
Cin
eole
a-P
inen
e
Eu
caly
ptu
sco
smo
ph
ylla
*C
ineo
lea-
Pin
ene
Eu
caly
ptu
scr
ebra
Cin
eole
a-P
inen
eb-
Pin
ene
Eu
caly
ptu
scr
enu
lata
Cy
men
ea-
Pin
ene
c-T
erp
inen
e
Eu
caly
ptu
scu
rtis
ii*
Oci
men
eb -
Pin
ene
Eu
caly
ptu
sd
ives
Cin
eole
a-P
hel
lan
dre
ne
Pip
erit
on
e
Eu
caly
ptu
sel
ata
(sy
nE
uca
lyp
tus
an
dre
an
a,
Eu
caly
ptu
sn
um
ero
sa)*
a-P
hel
lan
dre
ne
Pip
erit
ol
Pip
erit
on
e
Eu
caly
ptu
ser
yth
roco
rys*
a-T
erp
ineo
lC
ineo
le
E.
glo
bu
lus
sub
glo
bu
lus*
a-P
inen
eC
ineo
le
Eu
caly
ptu
sg
om
ph
oce
ph
ala
*a
-Pin
ene
Car
vo
ne
Eu
caly
ptu
sim
laye
nsi
s*C
ineo
lea-
Pin
ene
Eu
caly
ptu
sjo
hn
sto
nii
(sy
nE
uca
lyp
tus
mu
elle
ri)*
Cin
eole
a-P
inen
e
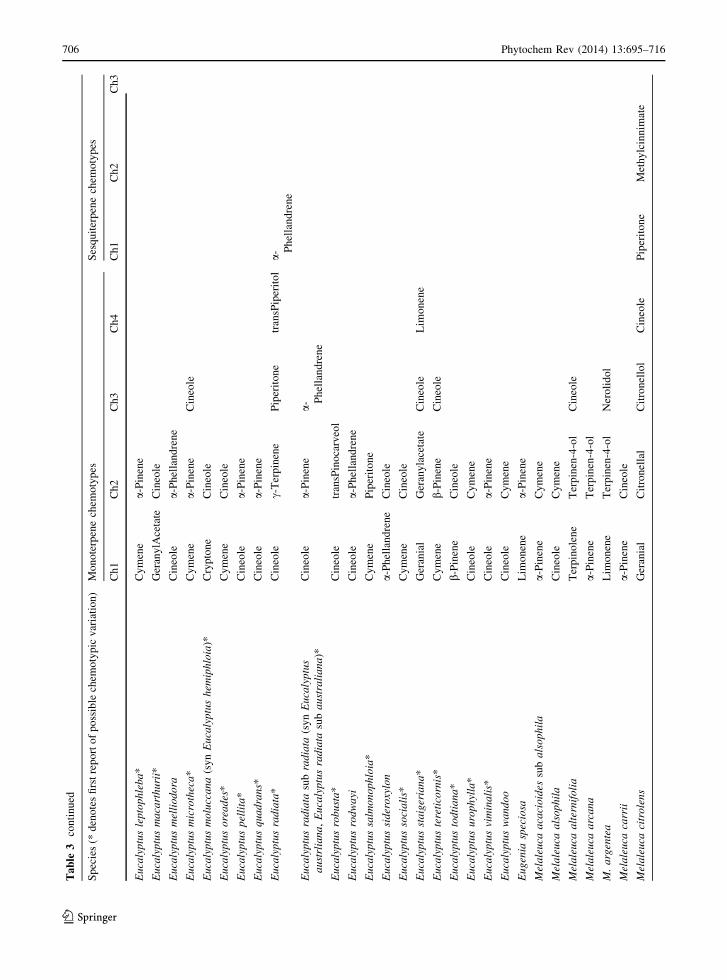
Phytochem Rev (2014) 13:695–716 705
123
Ta
ble
3co
nti
nu
ed
Sp
ecie
s(*
den
ote
sfi
rst
rep
ort
of
po
ssib
lech
emo
typ
icv
aria
tio
n)
Mo
no
terp
ene
chem
oty
pes
Ses
qu
iter
pen
ech
emo
typ
es
Ch
1C
h2
Ch
3C
h4
Ch
1C
h2
Ch
3
Eu
caly
ptu
sle
pto
ph
leb
a*
Cy
men
ea-
Pin
ene
Eu
caly
ptu
sm
aca
rth
uri
i*G
eran
ylA
ceta
teC
ineo
le
Eu
caly
ptu
sm
elli
od
ora
Cin
eole
a-P
hel
lan
dre
ne
Eu
caly
ptu
sm
icro
thec
a*
Cy
men
ea-
Pin
ene
Cin
eole
Eu
caly
ptu
sm
olu
cca
na
(sy
nE
uca
lyp
tus
hem
iph
loia
)*C
ryp
ton
eC
ineo
le
Eu
caly
ptu
so
rea
des
*C
ym
ene
Cin
eole
Eu
caly
ptu
sp
elli
ta*
Cin
eole
a-P
inen
e
Eu
caly
ptu
sq
ua
dra
ns*
Cin
eole
a-P
inen
e
Eu
caly
ptu
sra
dia
ta*
Cin
eole
c-T
erp
inen
eP
iper
ito
ne
tran
sPip
erit
ol
a-P
hel
lan
dre
ne
Eu
caly
ptu
sra
dia
tasu
bra
dia
ta(s
yn
Eu
caly
ptu
s
au
strl
ian
a,
Eu
caly
ptu
sra
dia
tasu
ba
ust
rali
an
a)*
Cin
eole
a-P
inen
ea
- Ph
ella
nd
ren
e
Eu
caly
ptu
sro
bu
sta
*C
ineo
letr
ansP
ino
carv
eol
Eu
caly
ptu
sro
dw
ayi
Cin
eole
a-P
hel
lan
dre
ne
Eu
caly
ptu
ssa
lmo
no
ph
loia
*C
ym
ene
Pip
erit
on
e
Eu
caly
ptu
ssi
der
oxy
lon
a-P
hel
lan
dre
ne
Cin
eole
Eu
caly
ptu
sso
cia
lis*
Cy
men
eC
ineo
le
Eu
caly
ptu
sst
aig
eria
na
*G
eran
ial
Ger
any
lace
tate
Cin
eole
Lim
on
ene
Eu
caly
ptu
ste
reti
corn
is*
Cy
men
eb-
Pin
ene
Cin
eole
Eu
caly
ptu
sto
dti
an
a*
b-P
inen
eC
ineo
le
Eu
caly
ptu
su
rop
hyl
la*
Cin
eole
Cy
men
e
Eu
caly
ptu
svi
min
ali
s*C
ineo
lea-
Pin
ene
Eu
caly
ptu
sw
an
do
oC
ineo
leC
ym
ene
Eu
gen
iasp
ecio
saL
imo
nen
ea-
Pin
ene
Mel
ale
uca
aca
cio
ides
sub
als
op
hil
aa-
Pin
ene
Cy
men
e
Mel
ale
uca
als
op
hil
aC
ineo
leC
ym
ene
Mel
ale
uca
alt
ern
ifo
lia
Ter
pin
ole
ne
Ter
pin
en-4
-ol
Cin
eole
Mel
ale
uca
arc
an
aa-
Pin
ene
Ter
pin
en-4
-ol
M.
arg
ente
aL
imo
nen
eT
erp
inen
-4-o
lN
ero
lid
ol
Mel
ale
uca
carr
iia-
Pin
ene
Cin
eole
Mel
ale
uca
citr
ole
ns
Ger
ania
lC
itro
nel
lal
Cit
ron
ello
lC
ineo
leP
iper
ito
ne
Met
hy
lcin
nim
ate
706 Phytochem Rev (2014) 13:695–716
123

Ta
ble
3co
nti
nu
ed
Sp
ecie
s(*
den
ote
sfi
rst
rep
ort
of
po
ssib
lech
emo
typ
icv
aria
tio
n)
Mo
no
terp
ene
chem
oty
pes
Ses
qu
iter
pen
e
chem
oty
pes
Ch
1C
h2
Ch
3C
h4
Ch
1C
h2
Ch
3
M.
con
cret
aa-
Pin
ene
Ter
pin
en-4
-ol
Cin
eole
M.
dis
siti
flo
raC
ineo
leT
erp
inen
-4-o
l
Mel
ale
uca
eric
ifo
lia
Lin
alo
ol
Cin
eole
Mel
ale
uca
exu
via
Cin
eole
Ter
pin
en-4
-ol
Mel
ale
uca
ha
lop
hil
aC
ineo
leT
erp
inen
-4-o
l
Mel
ale
uca
ha
ma
taC
ineo
leT
erp
inen
-4-o
l
Mel
ale
uca
leu
cad
end
raC
ineo
leM
eth
yle
ug
eno
lM
eth
yl
iso
eug
eno
l
Mel
ale
uca
lin
ari
ifo
lia
Cin
eole
Ter
pin
en-4
-ol
Ter
pin
ole
ne
Mel
ale
uca
pen
tag
on
av
ar.
La
tifo
lia
Cin
eole
My
rten
ala-
Pin
ene
Mel
ale
uca
plu
mea
Cin
eole
a-P
inen
e
Mel
ale
uca
spic
iger
aL
inal
oo
lC
ineo
le
Mel
ale
uca
sten
ost
ach
yab-
Pin
ene
Cin
eole
Mel
ale
uca
tric
ho
sta
chya
Cin
eole
Ter
pin
ole
ne
Mel
ale
uca
tub
ercu
lata
var
.m
ela
leu
caa
cro
ph
ylla
Cin
eole
Lin
alo
ol
Mel
ale
uca
un
cin
ata
a-P
inen
eC
ineo
leT
erp
inen
-4-o
l
Mel
ale
uca
vin
nu
laC
ineo
lea-
Pin
ene
Mel
ale
uca
viri
difl
ora
a-P
inen
eC
ineo
lec-
Ter
pin
ene
Met
hy
lcin
nim
ate
Ma
rlie
rea
eug
enio
pso
ides
a-P
inen
eT
erp
ino
len
e
Myr
cia
nth
esrh
op
alo
ides
*C
ineo
leG
eran
ial
Lin
alo
ol
Myr
tus
com
mu
nis
*C
ineo
leL
imo
nen
ea-
Pin
ene
Pim
enta
dio
ica
*E
ug
eno
lM
eth
yle
ug
eno
l
Pim
enta
race
mo
sav
arg
rise
a*
Met
hy
lis
oeu
gen
ol
Met
ho
xy
iso
eug
eno
l
Psi
diu
msa
rto
ria
nu
ma-
Pin
ene
Lim
on
ene
Tis
tan
iop
sis
coll
ina
My
rcen
ea-
Pin
ene
Tis
tan
op
sis
lau
rin
ea-
Pin
ene
Lim
on
ene
Wa
terh
ou
sia
flo
rib
un
da
a-P
inen
ea-
Ter
pin
eol
Phytochem Rev (2014) 13:695–716 707
123

The most striking pattern in foliar terpenes across
Myrtaceae is that a-pinene and 1,8-cineole are both
the most common and the most abundant compounds
across the majority of species in this family (Supp
Table 1). In most cases, 1,8-cineole or a-pinene are
the most abundant compounds in that foliage sample.
However, this pattern is not consistent in the clade
containing the tribes Myrteae, Kanieae and Syzygieae,
as well as the tribes Xanthostemoneae, Syncarpieae
and Lindsayomyrteae. In these tribes, 1,8-cineole (a
monoterpene), is primarily replaced by b-caryophyl-
lene (a sesquiterpene) as the most abundant terpene
and in Lindsayomyrteae, Metrosidereae and Back-
housieae, allo-aromadendrene (a sesquiterpene) is the
most abundant compound rather than the monoterpene
a-pinene (Tables 1, 2 and Supp Table 1). However,
we only have foliar terpene data for one species in
each of the latter three tribes, so this conclusion may
be an effect of sample size, and an avenue for future
research is to fill this sampling gap.
Species of Myrtaceae have some of the highest
concentrations of 1,8-cineole in the plant kingdom. De
Vincenzi et al. (2002) surveyed a range of aromatic
and medicinal plants with the focus on 1,8-cineole,
and found that essential oil from E. globulus (Myrt-
aceae) consists for 70–80 % this monoterpene. Other
published data show that some species of Eucalyptus
contain this compound at greater than 90 % of the total
essential oil (e.g. E. polybractea—Brophy and South-
well 2002). Few other plants outside the Myrtaceae
contain such large proportions of 1,8-cineole, however
there are some exceptions such as Cardamon (Elet-
taria cardamomum (Zingiberaceae)—De Vincenzi
et al. 2002) and camphor laurel (Cinnamomum cam-
phora (Lauraceae)—Stubbs et al. 2004). This suggests
that although 1,8-cineole is not unique to Myrtaceae,
high concentrations are found predominantly within
the Myrtaceae and so high foliar 1,8-cineole is a
defining feature of the family.
Mapping the patterns in foliar terpenes to the
phylogeny (Table 2), there are two evolutionary
trajectories that could have occurred (Fig. 2). The
first evolutionary trajectory is that the common
ancestor to all Myrtaceae had high foliar levels of a-
pinene and low foliar levels of 1,8-cineole. Subse-
quently, high concentrations of 1,8-cineole evolved in
the leaves in the common ancestor to Melaleuceae,
Syncarpieae, Lindsayomyrteae, Eucalypteae, Lepto-
spermeae and Chamelaucieae but was subsequentlyTa
ble
3co
nti
nu
ed
Sp
ecie
s(*
den
ote
sfi
rst
rep
ort
of
po
ssib
lech
emo
typ
icv
aria
tio
n)
Ch
1C
h2
Ch
3C
h4
(C)
Au
stro
myr
tus
flo
rib
un
da
Far
nes
ol
Far
nes
yla
ceta
te
Eu
caly
ptu
sn
ova
-an
gli
caA
rom
aden
dre
ne
Ner
oli
do
lG
lob
ulo
lE
ud
esm
ol
Eu
gen
iach
lorp
hyl
laG
lob
ulo
lC
ary
op
hy
llen
eO
xid
eb
-Car
yo
ph
yll
ene
Eu
gen
iad
ysen
teri
cab
-Car
yo
ph
yll
ene
c-C
adin
ene
Eu
gen
iain
volu
cra
tab
-Car
yo
ph
yll
ene
Bic
ycl
og
erm
acre
ne
Lep
tosp
erm
um
gra
nd
ifo
liu
mb
-Car
yo
ph
yll
ene
Vir
idifl
oro
l
Myr
cia
bra
ctea
ta*
Far
nes
ene
Ner
oli
do
lS
pat
hu
len
ol
Myr
cia
fall
ax
Bis
abo
lG
uai
ol
Myr
cia
sylv
ati
caC
alm
enen
eS
pat
hu
len
ol
Sel
ino
l
Psi
diu
mg
uin
een
seb
-Bis
abo
len
eB
isab
ol
Wa
terh
ou
sia
un
ipu
nct
ata
Ger
mac
ren
eD
Bic
ycl
og
erm
acre
ne
Th
ech
emo
typ
esar
eli
sted
inan
arb
itra
ryo
rder
(i.e
.ch
emo
typ
e1
isn
ot
the
do
min
ant
chem
oty
pe
inth
esp
ecie
s,n
or
isit
the
ance
stra
lch
emo
typ
e).
(A)
Sp
ecie
sw
ith
chem
oty
pes
of
bo
thm
on
ote
rpen
esan
dse
squ
iter
pen
es(i
.e.in
vo
lvin
gtw
ob
iosy
nth
etic
pat
hw
ays)
,(B
)sp
ecie
sw
ith
on
lym
on
ote
rpen
ech
emo
typ
es,(C
)sp
ecie
sw
ith
on
lyse
squ
iter
pen
ech
emo
typ
es
708 Phytochem Rev (2014) 13:695–716
123

lost in the Syncarpieae and Lindsayomyrteae lineage.
The second possible evolutionary trajectory is that the
common ancestor to all Myrtaceae had high foliar
levels of 1,8-cineole and a-pinene, then the machinery
leading to high foliar 1,8-cineole was lost several
times in the Xanthostemoneae and the Syncarpieae
and Lindsayomyrteae lineages as well as the clade
containing Myrteae, Tristanieae and Syzygieae. In
each case, the occurrence of high foliar a-pinene is a
trait common in most tribes of Myrtaceae and is
therefore most likely a trait present in the first species
of Myrtaceae. Since a-pinene is common in the leaves
of many species of Myrtaceae (Supp Table 1) and in
many species from closely related families (e.g.
Toudahl et al. 2012; Ogunbinu et al. 2007; Kaur and
Kaur 2010) it is likely that this characteristic was
present in the common ancestor and was lost in a few
species in several tribes of Myrtaceae. Both evolu-
tionary trajectories suggest it was likely that the
capacity to produce high foliar a-pinene was lost three
times, in Lindsayomyrteae, Metrosidereae and Back-
housieae. The major difference between the two
possible evolutionary trajectories is that one involves
a gain-of-function and a loss-of-function and the other
involves three loss-of-function events.
Fahnrich et al. (2011) compiled a review of the
major and minor products from monoterpene syn-
thases from Nicotiana whose major product was
produced by 1,8-cineole synthase. The analysis
showed that the only monoterpene synthases that
produced 1,8-cineole as a minor product were the a-
terpineol synthases of tobacco (Nicotiana langsdorfii
and N. suaveolens) and grape (V. vinifera). These two
monoterpenes are structurally similar and a-terpineol
is produced as an intermediate in 1,8-cineole synthesis
(Keszei et al. 2010), therefore it is possible that an a-
terpineol synthase could gain the ability to produce
1,8-cineole without significantly reducing the activity
of the enzyme (Kampranis et al. 2007). a-Terpineol is
rarely a dominant compound in Myrtaceae, but it is
frequently found in low abundance. Nonetheless, it is
often in the five most abundant compounds when 1,8-
cineole is the most abundant compound (Supp
Table 1). It would be instructive to survey both 1,8-
cineole synthases and a-terpineol synthases in the
leaves of Myrtaceae species and use these in site-
directed mutagenesis studies to obtain an a-terpineol
synthase that produced large amounts of 1,8-cineole.
We predict a simple sequence change will convert an
a-terpineol synthase to a 1,8-cineole synthase, perhaps
in a similar way to the sesquiterpene synthase from
maize (Zea mays) that has two areas in the catalytic
pocket (Kollner et al. 2006). We further hypothesise
that the distribution of these two terpene synthases will
be similar across the family Myrtaceae.
There is a large diversity of terpenes in the leaves of
Myrtaceae where a-pinene and 1,8-cineole are the
dominant compounds. Since many samples contain
Fig. 2 The two possible evolutionary trajectories of Myrtaceae
with respect to high foliar 1,8-cineole concentration. The dotted
lines in the phylogeny represent lineages with the high foliar
1,8-cineole trait, and the black lines represent lineages without
this trait. The crosses represent the predicted loss of high foliar
1,8-cineole, and the star represents gaining this trait. Tribes
whose names are green are those with the high foliar 1,8-cineole
trait
Phytochem Rev (2014) 13:695–716 709
123

high foliar levels of a-pinene, this compound is the
likely ancestral foliar chemotype of all Myrtaceae and
high foliar 1,8-cineole appears to be a defining feature
of the family. The reaction cascade that leads to these
two compounds include the same carbocation inter-
mediate: the a-terpinyl cation (Kampranis et al. 2007),
which suggests only a small change in the amino acid
sequence of one enzyme could allow it to produce
significant quantities of both compounds.
Could the structure of terpene synthase enzymes
constrain the diversity of terpenes found
in Myrtaceae (Fig. 1)?
We found 72 unique terpenes that occurred at high
abundance in the leaves of Myrtaceae (Table 2),
representing all of the carbocation intermediates in
terpene biosynthesis (Keszei et al. 2008). On top of
this, there are 112 functional and expressed terpene
synthases in the E. grandis genome, 80 of which are
expressed to some level in young or mature leaves.
This is the largest terpene synthase gene family
described in a single species (Kulheim et al. 2013).
However, the germacrone-type sesquiterpenes, which
includes elemanes and cadinanes (Iguchi et al. 1969),
are typically rare or missing from the foliar terpene
profile of Myrtaceae (this study, Brophy 2012, pers.
comm.), despite there being a functional and
expressed isoledene synthase that can also be used to
produce cadinanes in significant amounts present in
the genome of E. grandis (Kulheim et al. 2013). This
suggests that terpene synthases impose very few
constraints on the foliar terpene profile.
The presence of a functional enzyme capable of
producing the rare germacrone-type compounds in the
leaves of E. grandis, prompted us to investigate the
evolution of the terpene synthases in Myrtaceae by
examining relevant site-directed mutagenesis studies.
Surprisingly, there are only 10 angiosperm terpene
synthases that have been used in site-directed muta-
genesis studies: 1,8-cineole synthase from Salvia
officinalis (Kampranis et al. 2007); (?)-germacrene
D synthase and a (-)-germacrene D synthase from
Solidago canadensis (Prosser et al. 2006); 5-epi-
sesquithujene synthase and sesquithujene synthase
from Z. mays (Kollner et al. 2004); (E)-a-bergamo-
tene/(E)-b-farnesene synthase from Z. mays (Kollner
et al. 2009); 5-epiaristolochene synthase from
Nicotiana tabacum (Greenhagen et al. 2006); zing-
iberene synthase, b-sesquiphellandrene synthase and
(E)-b-farnesene synthase from Sorghum bicolor (Zhu-
ang et al. 2012). Amino acids contributing to the
internal surface of the catalytic pocket are in approx-
imately the same position in the sequence, relative to
the highly conserved DDXXD motif (Fig. 1), how-
ever, there is a region in the C-terminal domain that
shows variation across the 10 genes. Single amino acid
changes introduced into the terpene synthase lead to
relatively simple changes in the terpene profile such as
changes in stereochemistry or the ratio of products
(Prosser et al. 2004). However, when several amino
acids (in this case up to seven) were changed, the
entire product profile was altered (Kampranis et al.
2007). There is strong evidence that terpene synthases
have diversified within distinct lineages by duplication
and neofunctionalisation resulting in an array of
enzymes across the plant kingdom (Degenhardt et al.
2009; Keeling and Bohlmann 2006). The site-directed
mutagenesis studies show that changes of just a few
amino acids can radically change its production of
particular terpenes in a variety of angiosperms, which
provides a method by which neofunctionalisation of
duplicate terpene synthases can occur. If a mutation
occurs in one copy of the terpene synthase, then it may
not affect the terpene profile since the other copy is
functioning normally. This single mutation could
result in a functionally distinct terpene synthase which
is then exposed to selection pressures. This hypothet-
ical scenario is unlikely to evolve quickly, but the
family Myrtaceae arose at least 66 mya and has been
diversifying since (Biffin et al. 2010).
Most often the amino acid changes induced through
site-directed mutagenesis result in a different ratio of
particular terpenes produced, and in natural systems,
this might lead to different dominant compound.
A Thus, the ability could evolve to produce large
quantities of any compound found in its terpene product
profile through a short series of mutations. Evidence of
this is seen in thea-terpineol synthases of many species,
which are the only characterised terpene synthases, that
are not 1,8-cineole synthases, but produce significant
amounts of 1,8-cineole (Fahnrich et al. 2011).
The limited number of studies available for review
would suggest that enzymatic constraints do not limit
the diversity of terpenes in Myrtaceae, despite the
general absence of germacrone-type compounds in the
leaves. Terpene synthase diversity probably arose
710 Phytochem Rev (2014) 13:695–716
123

through duplication and small, step-wise sequence
changes in ancestral enzymes. At different times these
genetic changes were exposed to different selection
pressures. It is clear that site-directed mutagenesis
studies could be used more widely to tackle evolu-
tionary questions, especially in families that contain
many species with closely related terpene synthases,
such as Eucalypteae and Melaleucaeae. Analysis of
this type of data could be used to better understand
how the diversity in terpene synthases arises and even
to design transgenic plants that could be used to test
the role of different terpenes in mediating ecological
interactions.
What is the role of different species of Myrtaceae
within ecosystems?
Myrteae and Syzygieae are the only two tribes of
Myrtaceae to contain fleshy-fruited species (Biffin
et al. 2010; Thornhill et al. 2012; Wilson et al.
2005) and of the 256 species, which have low foliar
1,8-cineole, the majority (*90 %—Table 1) belong
to the these two tribes. On top of this, these species,
with low foliar 1,8-cineole concentrations, have a
greater diversity of abundant foliar sesquiterpenes
than species with high foliar 1,8-cineole (Table 1).
A recent description of the foliar terpene profile of
two chemically mosaic Eucalyptus trees indicates
that resistance to herbivory is determined by the
concentration of one or a few compounds, whilst
susceptibility to herbivory is determined by the
absence of these compounds (Padovan et al. 2012).
If terpenes are playing a defensive role in Myrta-
ceae, then there are evolutionary forces acting on
these terpenes associated with resistance, and if the
resource allocation to terpene production does not
change, we could expect that the terpene profile of
undefended leaves should contain a greater variety
of compounds than the terpene profile of defended
leaves. Since 1,8-cineole can act as a defensive
compound in Myrtaceae (Butcher et al. 1994;
Lawler et al. 1998) and the foliar terpene profile
of some tribes is more diverse than others, these
data suggest that defence may not be the primary
function of terpenes in species of Myrteae and
Syzygieae. Alternative roles such as attracting
beneficial animals or meditating root-microbe inter-
actions in the soil are possible (Steffen et al. 2012).
Which species should be explored by the essential
oil industry and which species might prove
problematic?
Industrially valuable essential oils are produced from a
number of species of Myrtaceae. 1,8-cineole is sought
from several species of Eucalyptus (e.g. E. polybrac-
tea, E. globulus and E. loxophleba) but also from
Melaleuca cajaputi. A second species of Melaleuca
(Melaleuca alternifolia) produces industrially desir-
able oils rich in terpinen-4-ol. There are two Austra-
lian standards for euclayptus oil. The first standard is
for eucalyptus oil that is 70–75 % 1,8-cineole (AS
2113.1-1998) and states that the terpene profile must
contain 1,8-cineole (70–75 %), limonene (1–15 %)
and a-terpineol (0.5–9 %). Whilst a-pinene, a-phel-
landrene and iso-valeric aldehyde can only be present
in trace amounts. The second standard is for eucalyp-
tus oil that is 80–85 % 1,8-cineole (AS 2113.2-1998)
and states that the terpene profile must contain 1,8-
cineole (80–85 %), limonene (1–15 %) and a-terpin-
eol (0.5–9 %). Whilst a-pinene, a-phellandrene and
iso-valeric aldehyde are only allowed in trace amounts
(\6 % of the oil). Although there are more than 900
species of Eucalyptus, only 20 species have been
exploited for the commercial production of eucalyptus
oil (Boland et al. 1991).
A similar story is true for the tea tree oil. The
Australian standard for tea tree oil (AS 2782-2009; ISO
4730:2004) states that the terpene profile must contain
terpinen-4-ol (30–48 %), c-terpinene (10–28 %), 1,8-
cineole (\15 %) and a-terpinene (5–13 %). Whilst a-
pinene, sabinene, limonene, q-cymene, terpinolene, a-
terpineol, aromadendrene, ledene, d-cadinene, globu-
lol and viridiflorol must be kept to trace amounts
(\8 % of the oil). There are more than 350 species of
Melaleuca, however only four species have been
exploited by the tea tree oil industry (Australian
Standards for tea tree oil—AS 2782-2009).
The major challenges for the essential oil industries
using species from the family Myrtaceae are that
require careful selection of stock to avoid undesirable
chemotypic effects, understanding the degree to which
planting stock is resistant to emerging pathogens (e.g.
Myrtle Rust) and awareness of regulatory issues
around undesirable traits. For example limonene and
its oxidation products are considered undesirable
because of irritant effects on the skin in high
concentrations (Christensson et al. 2009). However,
Phytochem Rev (2014) 13:695–716 711
123

limonene is formed by the same carbocation interme-
diates as is 1,8-cineole and is often the product of
cineole synthases. Understanding the origins and
enzymatic control of limonene formation may indicate
the degree to which these two compounds could be
uncoupled so that 1,8-cineole is produced without
limonene.
The following are a list of species that could be
considered by these two essential oil industries as a
means of diversifying the stock populations. Eucalyptus
oil: Blepharocalyx salicifolius, Callistemon pinifolius,
Callistemon viminalis, Eucalyptus aromaphloia, Euca-
lyptus johnsoniana, Melaleuca armillaris sub. armil-
laris, Melaleuca fulgens sub. corrugata, Melaleuca
hypericifolia, Melaleuca lateralis, Melaleuca paludi-
cola, Melaleuca phoenicea, Myrcianthes cisplatensis,
Myrcianthes pungens, Pimenta obscura, Callistemon
glaucus, Callistemon acuminatus, Callistemon citrinus,
Callistemon recurvus, Eucalyptus angustissima, Euca-
lyptus cinerea, Eucalyptus luehmanniana, Eucalyptus
parvifolia, Eucalyptus risdonii, Eucalyptus robertsonii,
Eucalyptus willisii, Melaleuca acutifolia, Melaleuca
calothamnoides, Melaleuca linophylla, Melaleuca oro-
phila, Melaleuca recurva, Melaleuca sabrina, Melal-
euca stereophloia and Myrcianthes ‘black fruit’. Tea
tree oil: Eucalyptus ovata, Leptospermum polygalifoli-
um Salisb sub. cismontanum, Psidium caudatum (also
known as Calycolpus moritzianus), Melaleuca concre-
ta, Melaleuca ochroma, Uromyrtus sp McPherson
Range, Melaleuca argentea, Melaleuca foliolosa and
Melaleuca dissitiflora. Caution should be taken to
evaluate each species thoroughly before using it as an
industrial source of essential oils.
Future directions
Our database is almost entirely focussed on terpenes
from mature leaves because this is the most common
material studied when considering whether a plant is
suitable for industrial purposes. Terpenes play a
crucial role in protecting leaves from herbivory and
infection (DeGabriel et al. 2009; Macel and Klinkh-
amer 2010; Matsuki et al. 2011), but Myrtaceae must
employ terpenes in important roles in root and wood
tissues since many terpene synthases are highly
expressed in these tissues as they are in leaves
(Kulheim et al. 2013, Rasmann et al. 2012). There is
a distinct gap in our understanding of the role terpenes
play in roots and wood which limits our understanding
of the ecological importance of terpenes to plants. If
we are truly to understand the role of terpenes in
Myrtaceae, this will need to be addressed.
The second significant gap in data we identified was
in the diversity of site-directed mutagenesis studies of
plant terpene synthases. There are only 10 angiosperm
terpene synthases used in site-directed mutagenesis
studies (Greenhagen et al. 2006; Kampranis et al.
2007; Kollner et al. 2009; Kollner et al. 2004; Prosser
et al. 2004; Zhuang et al. 2012), with only a few more
from the gymnosperms (Hyatt and Croteau 2005;
Katoh et al. 2004). These experiments provide insights
into terpene synthase evolution that is unparalleled by
any other method. In Myrtaceae the most useful target
of these approaches would be a detailed examination
of the function of the catalytic pocket, with the focus
on enzymatic evolution.
The final and arguably the most important future
direction is to standardise the method of collecting and
reporting constitutive terpene data in Myrtaceae.
Almost every author cited in this study used a different
method for extracting terpenes and presenting their
data that is independent of the journal requirements.
Most of this revolves around whether data is reported
either as proportions or as concentrations relative to
dry or wet weight. Birks and Kanowski (1993)
reviewed these issues for analysis of conifer resin
and propose standardised statistical approaches for
analysis and reporting proportional composition data.
For a comparison amongst different Myrtaceae, we
believe that terpene data should be presented as
concentration since proportion can be calculated from
concentration (or presented along with concentration)
but concentration cannot be calculated from propor-
tional data. Approximately half of the studies that we
reviewed present concentration data in terms of fresh
weight and the other half dry weight. One of the
problems in presenting dry weight data of terpene rich
leaves is that many drying methods volatilize the
terpenes as well as the water but approaches have been
developed to account for this (Ammon et al. 1985).
Although analyses of leaves for industrial purposes
might favour reporting concentration on a wet weight
basis, most if not all ecological analyses should report
concentration on a dry weight basis and we urge future
studies to standardise on these units or to report both.
There are numerous terpene extraction methods
currently being used, each with different advantages
712 Phytochem Rev (2014) 13:695–716
123

and limitations. The two main methods of terpene
extraction that underlie the data discussed are distil-
lation (steam, hydro or thermo) and organic solvent
extraction (ethanol, pentane, hexane and dichloro-
methane—data not shown). The major disadvantages
of distillation are the loss of thermolabile and highly
volatile compounds. For example, cryptomeridiol and
4-epicryptomeridiol are two eudesmol hydrates that
are not observed after isolation of oil by steam
distillation as they are thermolabile (Cornwell et al.
2000b); and cubenol and epicubenol break down to d-
cadinene, epicubenol and cubenol as a result of the
processes involved in steam distillation (Cornwell
et al. 2000a, b, 2001). Themajor disadvantages of
solvent extraction are inaccurate measures of concen-
trations due to evaporation and the varied success of
extraction of different monoterpene alcohols (e.g.
extraction of a higher amount of linalool compared
with distillation methods, anda lower amount of 1,8-
cineole (Charles and Simon 1990). We believe that
structural re-arrangements are far more detrimental to
a survey of plant terpene diversity than differences in
the proportion of high abundance compounds and thus
solvent extraction methods should be favoured over
distillation methods.
Acknowledgments This research was supported by a grant
from the Australian Research Council to WJF (LP110100184).
We thank our partners in that work (Australian Tea Tree
Industry Association and GR Davis) for their support and
appreciate the advice and comments of many essential oil
chemists.
References
Aharoni A, Giri AP, Verstappen FWA, Bertea CM, Sevenier R,
Sun Z, Jongsma MA, Schwab W, Bouwmeester HJ (2004)
Gain and loss of fruit flavor compounds produced by wild
and cultivated strawberry species. Plant Cell 16:3110–3131
Ammon DG, Barton AFM, Clarke DA, Tjandra J (1985) Rapid
and accurate chemical determination of the water content
of plants containing volatile oils. Analyst 110:917–920
Andrew R, Wallis I, Harwood C, Henson M, Foley W (2007)
Heritable variation in the foliar secondary metabolite sid-
eroxylonal in Eucalyptus confers cross-resistance to her-
bivores. Oecologia 153:891–901
Andrew RL, Wallis IR, Harwood CE, Foley WJ (2010) Genetic
and environmental contributions to variation and popula-
tion divergence in a broad-spectrum foliar defence of
Eucalyptus tricarpa. Ann Bot 105:707–717
Andrew RL, Keszei A, Foley WJ (2013) Intensice sampling
identifies new chemotypes, population divergence and
biosynthetic connections among terpenoids in Eucalyptus
tricarpa. Phytochemistry 94:148–158
Arnold K, Bordoli L, Kopp J, Schwede T (2006) The
SWISS-MODEL workspace: a web-based environment
for protein structure homology modelling. Bioinformat-
ics 22:195–201
Biffin E, Lucas EJ, Craven LA, Ribeiro da Costa I, Harrington
MG, Crisp MD (2010) Evolution of exceptional species
richness among lineages of fleshy-fruited Myrtaceae. Ann
Bot 106:79–93
Birks JS, Kanowski PJ (1993) Analysis of resin compositional
data. Silvae Genet 42:340–350
Bohlmann J, Keeling CI (2008) Terpenoid biomaterials. Plant J
54:656–669
Bohlmann J, Crock J, Jetter R, Croteau R (1998) Terpenoid-
based defenses in conifers: cDNA cloning, characteriza-
tion, and functional expression of wound-inducible (E)-
alpha-bisabolene synthase from grand fir (Abies grandis).
PNAS 95:6756–6761
Boland DJ, Brophy JJ, House APN (1991) Eucalyptus leaf oils.
Use, chemistry, distillation and marketing. Inkata Press,
Sydney, Australia
Bouwmeester HJ, Verstappen FWA, Posthumus MA, Dicke M
(1999) Spider mite-induced (3S)-(E)-nerolidol synthase
activity in cucumber and lima bean. The first dedicated step
in acyclic C11-homoterpene biosynthesis. Plant Physiol
121:173–180
Brophy JJ, Southwell IA (2002) Eucalyptus chemistry. In:
Coppen JJ (ed) Eucalyptus: the genus Eucalyptus. Taylor
and Francis, New York
Brophy JJ, Goldsack RJ, Forster PI (2006) A preliminary
examination of the leaf oils of the genus Xanthostemon
(Myrtaceae) in Australia. J Essent Oil Res 18:222–230
Butcher PA, Doran JC, Slee MU (1994) Intraspecific variation in
the leaf oils of Melaleuca alternifolia (Myrtaceae). Bio-
chem Syst Ecol 22:419–430
Carnegie AJ, Lidbetter JR, Walker J, Horwood MA, Tesoriero
L, Glen M, Priest MJ (2010) Uredo rangelii, a taxon in the
guava rust complex, newly recorded on Myrtaceae in
Australia. Australas Plant Pathol 39:463–466
Carson CF, Hammer KA, Riley TV (2006) Melaleuca alterni-
folia (tea tree) oil: a review of antimicrobial and other
medicinal properties. Clin Microbiol Rev 19:50–62
Charles DJ, Simon JE (1990) Comparison of extraction methods
for the rapid determination of essential oil content and
composition of basil. J Am Soc Hortic Sci 115:458–462
Chen H-C, Sheu M-J, Lin L-Y, Wu C-M (2007) Chemical
composition of the leaf essential oil of Psidium guajava L.
from Taiwan. J Essent Oil Res 19:345–347
Christensson JB, Forsstrom P, Wennberg A-M, Karlberg A-T,
Matura M (2009) Air oxidation increases skin irritation
from fragrance terpenes. Contact Dermatitis 60:32–40
Cornwell CP, Reddy N, Leach DN, Grant WS (2000a) Origin of
(?)-delta-cadinene and the cubenols in the essential oils of
the Myrtaceae. Flavour Fragr J 15:352–361
Cornwell CP, Reddy N, Leach DN, Wyllie SG (2000b)
Hydrolysis of hedycaryol: the origin of the eudesmols in
the Myrtaceae. Flavour Fragr J 15:421–431
Phytochem Rev (2014) 13:695–716 713
123

Cornwell CP, Reddy N, Leach DN, Wyllie SG (2001) Ger-
macradienols in the essential oils of the Myrtaceae. Flavour
Fragr J 16:263–273
Crankshaw DR, Langenheim JH (1981) Variation in terpenes
and phenolics through leaf development in Hymenaea and
its possible significance to herbivory. Biochem Syst Ecol
9:115–124
Croteau R, Davis E, Ringer K, Wildung M (2005) (-)-Menthol
biosynthesis and molecular genetics. Naturwissenschaften
92:562–577
De Vincenzi M, Silano M, De Vincenzi A, Maialetti F, Sca-
zzocchio B (2002) Constituents of aromatic plants: euca-
lyptol. Fitoterapia 73:269–275
DeGabriel J, Moore B, Shipley L, Krockenberger A, Wallis IR,
Johnson C, Foley WJ (2009) Inter-population differences
in the tolerance of a marsupial folivore to plant secondary
metabolites. Oecologia 161:539–548
Degenhardt J, Kollner TG, Gershenzon J (2009) Monoterpene
and sesquiterpene synthases and the origin of terpene
skeletal diversity in plants. Phytochemistry 70:1621–1637
Dudareva N, Andersson S, Orlova I, Gatto N, Reichelt M, Rhodes
D, Boland W, Gershenzon J (2005) The nonmevalonate
pathway supports both monoterpene and sesquiterpene for-
mation in snapdragon flowers. Plant Biol 102:933–938
Fahnrich A, Krause K, Piechulla B (2011) Product variability of
the ‘cineole cassette’ monoterpene synthases of related
Nicotiana species. Mol Plant 4:965–984
Govaerts, R (2008) World checklist of Myrtaceae: Kew Pub
Grattapaglia D, Vaillancourt R, Shepherd M, Thumma B, Foley
W, Kulheim C, Potts B, Myburg A (2012) Progress in
Myrtaceae genetics and genomics: Eucalyptus as the piv-
otal genus. TGG 8:463–508
Greenhagen BT, O’Maille PE, Noel JP, Chappell J (2006)
Identifying and manipulating structural determinates
linking catalytic specificities in terpene synthases. PNAS
103:9826–9831
Grubb PJ, Metcalfe DJ, Grubb EA, Jones GD (1998) Nitrogen-
richness and prtoection of seeds in Australian tropical
rainforest: a test of plant defence theory. Oikos 82:467–482
Guex N, Peitsch M, Schwede T, Diemand A (1995–2011)
DeepView/Swiss-PdbViewer v4.0.4. Swiss Institute of
Bioinformatics
Hall D, Robert J, Keeling C, Domanski D, Lara A, Jancsik S,
Kuzyk M, Hamberger B, Borchers C, Bohlmann J (2011)
An integrated genomic, proteomic and biochemical ana-
lysis of (?)-3-carene biosynthesis in Sitka spruce (Picea
sitchensis) genotypes that are resistant or susceptible to
white pine weevil. Plant J 65:936–948
Homer LE, Leach DN, Lea D, Slade Lee L, Henry RJ, Baver-
stock PR (2000) Natural variation in the essential oil con-
tent of Melaleuca alternifolia Cheel (Myrtaceae). Biochem
Syst Ecol 28:367–382
Horner JD, Gosz JR, Cates RG (1988) The role of carbon-based
plant secondary metabolites in decomposition in terrestrial
ecosystems. Am Nat 132:869–883
Huang M, Sanchez-Moreiras AM, Abel C, Sohrabi R, Lee S,
Gershenzon J, Tholl D (2012) The major volatile organic
compound emitted from Arabidopsis thaliana flowers, the
sesquiterpene (E)-b-caryophyllene, is a defense against a
bacterial pathogen. New Phytol 193:997–1008
Hyatt DC, Croteau R (2005) Mutational analysis of a mono-
terpene synthase reaction: altered catalysis through direc-
ted mutagenesis of (-)-pinene synthase from Abies
grandis. Arch Biochem Biophys 439:222–233
Iguchi M, Nishiyama A, Yamamura S, Hirata Y (1969) Con-
version of elemene-type sesquiterpenes into cadinene-type
compounds and formation of ten-membered germacrone-
type intermediates. Tetrahedron Lett 10:4295–4298
Ireland B, Hibbert D, Goldsack R, Doran J, Brophy J (2002)
Chemical variation in the leaf essential oil of Melaleuca
quinquenervia (Cav.) S.T. Blake. Biochem Syst Ecol
30:457–470
Kampranis SC, Ioannidis D, Purvis A, Mahrez W, Ninga E,
Katerelos NA, Anssour S, Dunwell JM, Degenhardt J,
Makris AM, Goodenough PW, Johnson CB (2007)
Rational conversion of substrate and product specificity in
a Salvia monoterpene synthase: structural insights into the
evolution of terpene synthase function. Plant Cell
19:1994–2005
Kant M, Baldwin I (2007) The ecogenetics and ecogenomics of
plant–herbivore interactions: rapid progress on a slippery
road. Curr Opin Genet Dev 17:519–524
Katoh S, Hyatt D, Croteau R (2004) Altering product outcome in
Abies grandis (-)-limonene synthase and (-)-limonene/
(-)-alpha-pinene synthase by domain swapping and
directed mutagenesis. Arch Biochem Biophys 425:65–76
Kaur R, Kaur H (2010) The antimicrobial activity of essential oil
and plant extracts of Woodfordia fruticosa. Arch Appl Sci
Res 2:302–309
Keeling C, Bohlmann J (2006) Genes, enzymes and chemicals
of terpenoid diversity in the constitutive and induced
defence of conifers against insects and pathogens. New
Phytol 170:657–675
Keszei A, Brubaker CL, Foley WJ (2008) A molecular per-
spective on terpene variation in Australian Myrtaceae. Aust
J Bot 56:197–213
Keszei A, Brubaker CL, Carter R, Kollner T, Degenhardt J,
Foley WJ (2010) Functional and evolutionary relationships
between terpene synthases from Australian Myrtaceae.
Phytochemistry 71:844–852
Kollner TG, Schnee C, Gershenzon J, Degenhardt J (2004) The
variability of sesquiterpenes emitted from two Zea mays
cultivars is controlled by allelic variation of two terpene
synthase genes encoding stereoselective multiple product
enzymes. Plant Cell 16:1115–1131
Kollner TG, O’Maille PE, Gatto N, Boland W, Gershenzon J,
Degenhardt J (2006) Two pockets in the active site of
maize sesquiterpene synthase TPS4 carry out sequential
parts of the reaction scheme resulting in multiple products.
Arch Biochem Biophys 448:83–92
Kollner TG, Gershenzon J, Degenhardt J (2009) Molecular and
biochemical evolution of maize terpene synthase 10, an
enzyme of indirect defense. Phytochemistry 70:1139–1145
Kulheim C, Webb H, Yeoh SH, Wallis I, Moran G, Foley W
(2011) Using the Eucalyptus genome to understand the
evolution of plant secondary metabolites in the Myrtaceae.
BMC Proc 5:O11
Kulheim C, Padovan A, Hefer C, Krause S, Degenhardt J,
Myburg A, Foley WJ (2013) The Eucalyptus terpene syn-
thase gene family. New Phytol (in press)
714 Phytochem Rev (2014) 13:695–716
123

Langenheim JH, Foster CE, McGinley RB (1980) Inhibitory
effects of different quantitative compositions of Hymenaea
leaf resins on a generalist herbivore Spodoptera exigua.
Biochem Syst Ecol 8:385–396
Lawler IR, Foley WJ, Eschler BM, Pass DM, Handasyde K
(1998) Intraspecific variation in Eucalyptus secondary
metabolites determines food intake by folivorous marsu-
pials. Oecologia 116:160–169
Levin DA (1976) The chemical defenses of plants to pathogens
and hrbivores. Annu Rev Ecol Syst 7:121–159
Lichtenthaler HK (1999) The 1-deoxy-D-xylulose-5-phosphate
pathway of isoprenoid biosynthesis in plants. Annu Rev
Plant Physiol Plant Mol Biol 50:47–65
Linhart YB, Thompson JD (1995) Terpene-based selective
herbivory by Helix aspersa (Mollusca) on Thymus vulgaris
(Labiatae). Oecologia 102:126–132
Macel M, Klinkhamer P (2010) Chemotype of Senecio jacobaea
affects damage by pathogens and insect herbivores in the
field. Evol Ecol 24:237–250
Maida M, Carroll A, Coll J (1993) Variability of terpene content
in the soft coral Sinularia flexibilis (Coelenterata: Octo-
corallia), and its ecological implications. J Chem Ecol
19:2285–2296
Martin D, Aubourg S, Schouwey M, Daviet L, Schalk M, Toub
O, Lund S, Bohlmann J (2010) Functional annotation,
genome organization and phylogeny of the grapevine (Vitis
vinifera) terpene synthase gene family based on genome
assembly, FLcDNA cloning, and enzyme assays. BMC
Plant Biol 10:226–248
Matsuki M, Foley WJ, Floyd R (2011) Role of volatile and non-
volatile plant secondary metabolites in host tree selection
by Christmas beetles. J Chem Ecol 37:286–300
McCaskill D, Croteau R (1995) Monoterpene and sesquiterpene
biosynthesis in glandular trichomes of peppermint (Mentha
x piperita) rely exclusively on plastid-derived isopentenyl
diphosphate. Planta 197:49–56
Moore BD, Andrew RL, Kulheim C, Foley WJ (2013) Causes
and consequences of intraspecific chemical diversity. New
Phytol (in press)
Ogunbinu AO, Ogunwande IA, Walker TM, Setzer WN (2007)
Study on the essential oil of Lawsonia inermis (L) Lythra-
ceae. J Essent Oil Bear Plants 10:184–188
Padovan A, Keszei A, Koellner TG, Degenhardt J, Foley WJ
(2010) The molecular basis of host plant selection in
Melaleuca quinquenervia by a successful biological con-
trol agent. Phytochemistry 71:1237–1244
Padovan A, Keszei A, Wallis IR, Foley WJ (2012) Mosaic euca-
lypt trees suggest genetic control at a point that influences
several metabolic pathways. J Chem Ecol 38:914–923
Pichersky E, Gershenzon J (2002) The formation and function of
plant volatiles: perfumes for pollinator attraction and
defense. Curr Opin Plant Biol 5:237–243
Prosser I, Altug IG, Phillips AL, Konig WA, Bouwmeester HJ,
Beale MH (2004) Enantiospecific (?)- and (-)-germac-
rene D synthases, cloned from goldenrod, reveal a func-
tionally active variant of the universal isoprenoid-
biosynthesis aspartate-rich motif. Arch Biochem Biophys
432:136–144
Prosser IM, Adams RJ, Beale MH, Hawkins ND, Phillips AL,
Pickett JA, Field LM (2006) Cloning and functional char-
acterisation of a cis-muuroladiene synthase from black
peppermint (Mentha 9 piperita) and direct evidence for a
chemotype unable to synthesise farnesene. Phytochemistry
67:1564–1571
Rasmann S, De Vos M, Casteel CL, Tian D, Halitschke R, Sun
JY, Agrawal AA, Felton GW, Jander G (2012) Herbivory
in the previous generation primes plants for enhanced
insect resistance. Plant Physiol 158:854–863
Slee AV, Brooker MIH, Duffy SM, West JG (2006) EUCLID
eucalypts of Australia third edition. CSIRO, Canberra
Steffen RB, Antoniolli ZI, Steffen GPK, da Silva RF (2012)
Essential oil of Eucalyptus grandis Hill ex Maiden in
stimulating mycorrhizal sibipiruna seedlings (Ceasalpinia
peltophoroides Benth.). Cienc Florest 22:69–78
Steinbauer MJ (2010) Latitudinal trends in foliar oils of euca-
lypts: environmental correlates and diversity of chryso-
melid leaf-beetles. Austral Ecol 35:204–213
Stubbs BJ, Specht A, Brushett D (2004) The essential oil of
Cinnamomum camphora (L.) Nees and Eberm.—variation
in oil composition throughout the tree in two chemotypes
from eastern Australia. J Essent Oil Res 16:200–205
Suni T, Kulmala M, Hirsikko A, Bergman T, Laakso L, Aalto P,
Leuning R, Cleugh H, Zegelin S, Hughes D, van Gorsel E,
Kitchen M, Vana M, Horrak U, Mirme S, Mirme A, Sev-
anto S, Twining J, Tadros C (2008) Formation and char-
acteristics of ions and charged aerosol particles in a native
Australian eucalypt forest. Atmos Chem Phys 8:129–139
Thornhill AH, Crisp MD (2012) Phylogenetic assessment of
pollen characters in Myrtaceae. Aust Syst Bot 25:171–187
Thornhill AH, Hope GS, Craven LA, Crisp MD (2012) Pollen
morphology of the Myrtaceae. Part 4: tribes Kanieae,
Myrteae and Tristanieae. Aust J Bot 60:260–289
Toudahl AB, Filho SAV, Souza GHB, Morais LD, Santos ODH,
Jager AK (2012) Chemical composition of the essential oil
from Microlicia graveolens growing wild in Minas Gerais.
Rev Bras Farmacogn 22:680–681
Unsicker SB, Kunert G, Gershenzon J (2009) Protective per-
fumes: the role of vegetative volatiles in plant defense
against herbivores. Curr Opin Plant Biol 12:479–485
Van Poecke RMP, Posthumus MA, Dicke M (2001) Herbivore-
induced volatile production by Arabidopsis thaliana leads
to attraction of the parasitoid Cotesia rubecula: chemical,
behavioral, and gene-expression analysis. J Chem Ecol
27:1911–1928
Vickers CE, Gershenzon J, Lerdau MT, Loreto F (2009) A
unified mechanism of action for volatile isoprenoids in
plant abiotic stress. Nat Chem Biol 5:283–291
Wallis IR, Keszei A, Henery ML, Moran GF, Forrester R,
Maintz J, Marsh KJ, Andrew RL, Foley WJ (2011) A
chemical perspective on the evolution of variation in
Eucalyptus globulus. Perspect Plant Ecol Evol Syst
13:305–318
Webb H, Lanfear R, Hamill J, Foley WJ, Kulheim C (2013) The
yield of essential oils in Melaleuca alternifolia (Myrta-
ceae) is regulated through transcript abundance of genes in
the MEP pathway. PLoS One 8:e60631
Wilson PG (2011) Myrtaceae. In: Kubitzki K (ed) The families
and genera of vascular plants, vol 10. Springer, Berlin,
pp 212–271
Wilson P, O’Brien M, Heslewood M, Quinn C (2005) Rela-
tionships within Myrtaceae sensu lato based on a matK
phylogeny. Plant Syst Evol 251:3–19
Phytochem Rev (2014) 13:695–716 715
123

Wise ML, Savage TJ, Katahira E, Croteau R (1998) Monoterpene
synthases from common sage (Salvia officinalis). cDNA
isolation, characterization, and functional expression of (?)-
sabinene synthase, 1,8-cineole synthase, and (?)-bornyl
diphosphate synthase. J Biol Chem 273:14891–14899
World-Health-Organisation (1999) d-Limonene. In Some
chemicals that cause tumours of the kidney or urinary
bladder in rodents and some other substances: summary of
data reported and evaluation. IARC, pp 307–327
Zhuang X, Kollner TG, Zhao N, Li G, Jiang Y, Zhu L, Ma J,
Degenhardt J, Chen F (2012) Dynamic evolution of her-
bivore-induced sesquiterpene biosynthesis in sorghum and
related grass crops. Plant J 69:70–80
716 Phytochem Rev (2014) 13:695–716
123





























